genetical studies on the skeleton of the mouse xviii. three genes for syndactylism
TRANSCRIPT

T r I 0ENErlCAL STUDIES ON THE SKELETON OF TKE MOUSE
XVII I TH]REE GENES FO~ $7~ N I)AC,TX LISle{
BY HANS GRONIgBERG
3le~Hcd l~sec~,~elz Co~dt, (}ro~2) fo~ Ez2~er4't~se~zt~d R~sec~rd~. 4~. I~tmr'ite?.. Diseases, U~ivers@y Cdbge, Londo~*
(With Thirty TextJigm'es)
(Received 29 J~oae 1955)
INTJ~OD~OTYO2q
!t'.hi~ paper deals will1 three different genes which ealtse syndaetylism of the. ha.nds ~nd f~e< either alone oi' in combination with o~hm" anomalies. Two of tl~ese genes have not. be~n desoriloed t~efore: The third gene, shsker with syndaetylism, has been b ie f fy de- 'scribed by Rs discoverer (Hertwig, 1942), who has also sttldied the structure of its la%y- fN~h (Hertwig, i951); the effects of the g~ne on the skeleton have, however, not yet Been ~Ves~igat.ed. This paper will give a description of the osseo~:s and cartilaginous skeletons of!r thsee rout.ants based on aEzaz'in and methyl.ene-bkle clearance preparations and afew papain macerations- Ohondrifioation Of the skdeton of hands and feet first becomes aeteetable in methylene blue preparations in 14oday mouse embryos. This has been taken ~s..:~h~ starting point for alI three mutants, while earlier stages will be reserved for later ~apers As wilt appear below, each. of the three mutants hascertain complicating featm'es;
~[iese also remain to be studied.
~YNDACTYLISM
Ge.r~etics. Syndaety]ism was discovered in 1951 as a, s]?ontaneotls mutation in the A/Fa �9 ' ~ " collected by Dr D. S. lealconer are given in inbred strain in Edinbm'gh. Segreeatmn data a~.appendix io this paper. Animals of the strain (derived from linkage experiments and lienee genetically heterogeneoas) were kindly hs,nded over to me in October 19.58. ~[ost ofehe work reported here was done on a i m a l s bred from the original stock, but a small :9!refers to the inbred s~rain CBA/@r was also carried, out. Segregation data collected in .{his.laboratory (Table 1) rote in general agreemen.t wRh. those of lealeoner and eox~firm his conclusion that the condition is dne to a. si~gle recessive gene :['o:t' which we propose the �9 symbol 's.m. '. In all sea,'e~atil~,g ~enerations, there is a considerable d.efmieney ofreoessives aS.hompared wi.gh the expectadoa, As sm./sm animals are invariably affected on all four feat, it is very mdikely that the defmiency is due to the oeeurrenoe of >omnal overlaps. Data on embryos to be given below show no deleieney of syndaetyls. The deficiency is ~hus obyiously due to eel.retire dhnination at or soon after birth. There are in :fact always inany stillborn and often mutila.ted bodies m segregating m.ating~, m~d the mothers presumably eat some a:ffec~ed young before the littm:s are recorded- The.~ e is also a, gre~tly ii~ereased mortali ty of s.yndaetyls in the first :few days after bh'dL }Iowever, the animals ~vhieh live to be weaned generally survive and breed. The fertilRy of syndactyl males
8 Oenet. 54

114 Geneticc~Z s'htdies o'~ ~he sk, eleton of the ,mou~e
seems to be :fairly normal, but that of syndaetyl females of~e~ leaves much to be desirpd:.: The stock is thus best maintained baekcross :fashion, ma~,ing syndaetyl males to the~?: heterozygous sisters.
The reduced[ viability of s~/~'-m is iaflue~cecl by the gene~io backgroun& It was rnu~li lower in the F~ generation re,~tdting from an outoross to th.e inbred strain CBA/Gr than {if the original sbo ok (uos. 9 a:~ d 3, Table 1 }, tl:l.e difference being highly signiw (X"= 18'3~:. ~ = 1 ; P=2 x 10 -~ approx.). The causes fbr the increased morfialJty of .s.,m,/s'~n a5 bir6~ and soon after have not yet been investigaOed.
Table I, ,%9'regc~io~ of syq~dacty~is'r~ ets ve.eo'~des at 5irth o* soo'~ rCte*" Pheno~y~e of
Genotype of parent offspl'ing
No. c~ 9 marinas + #~ Total t * -ml~ + / + :] 18 0 18 2 + / . ~ -'/s~,~ (a) ,~ 215 § 259 B +/s)~ +/s in (~) 5 231 ]2 2~3 4 +/ .s~ ~m#m 9 144 75 219 5 ~/s,,~z +/sin 8 140 62 202
Via?Jiligy
0-61 0.t6 0.52 0.4,1;
Note, The intsrcross rantings no. 2 were made within the o r i ~ i s~oek from which most o~' the baokoro~s rantings (nee. ,I s ad 5} are ak~o derived. The Jn~ereross mat, ings no. 3 are ~n _,V a generation following an outcrops. ~o the inbred strain OBA/Gr (no, I). T h e ' viabil i ty ' given in She I~st column h~s been calculated from the segrega~ don ratios; as it includes some stillborn bu t classifiable ymmg, it represents a m a x i m u m rather . than ~ ~0ruo vahi~..
Table 9. _Relationship of tail anomagies to s~/ndactylism. A&dts Normal Sb~adaetyl
Tail T~fl Tail Tail Viability _No. Type of mating normal affeo~ecl normal afl%cted Total of sr~
I +# f~ x + # m (~) 150 4 t l I1 I76 0-4,3 +/sm x + / s~ (5) 195 2 7 0 204 0-11
3 +/3r~ x s~,~/sm (~) 86 3 9 t2 110 0-Z3 4 + / s~ x sin~am (b] 73 0 16 5 94 0.28
~%:ote. Nee. 1 znd 2 correspond to nos. 2, and 3 in T~ble I. The baekzrosses nos. 3 and ~ differ in ~h~t the pareii~l
~m/~'~ animals in no, 3 had %nil anom~ whil~ tho~e in no. ~ had norma.] ta.ils; in both cases reciproc~l b~ok~
crosses have been pooled.
Synd~ot, yls often have tsi]. anomalies which are always in the dJs~al half of the ~ i l ~.nd may vary from a eompIe~e twis~ go one or two marked tail 1/nks and finally d o w n g defects which are b~rely de~eetable. The data of Table 2 show 5hat 28 out of 71 adair; syndaotyls, or nearly '40 %, had tail anomalies marked enough to be visible in the living:. animal ~oreover, there is a suggestion 0hat the incidence of ~ai[ anomalies in s,m/sm mice:: is %flueneed by the generic baokgronnd; +,he baokcross data, nee. 3 and 4,, Table 2, differ.:. in tJaa~ in ~he former the syn/sm parents had abnormal tails, while in the latter the sm/s/(9i parents had normal tails; in the offspring of the former group, tail anomalies oeourredm i2 out of 21 sm/s~ animals, but oMy i~a 5 tag of 21 i~?. that of ~he lather; the differenCg~i jns$ significan$. (X 2 = 4.842; ~ = I; P - 0.028).
The situation is complicated by the fact that a similar kind of anomaly .is sometime.s fotmd in normal liSter-mates of syndaetyls, there being 9 among 513 normal adultsi. :0} t-75% . The usual ~ues~ion 5hus immedia~ely arises as to wb.edzer the tail a n o m g ~ among the normats ~re heterozygous mamfestations of sm (% which ease they should 0~ ~-. be :found in. +/sin), or whether sin~sin Am-ply enhances t;he manifestation of ~t~ e n g ~ w}fieh otherwise does no~ cleon show up among the normals (in. which case the affected:

I c i n g s G r ~ J ~ J ~ t ~ 1 15
~o;rm~ls migh~ be eibher +/s.m or § ). Other and less l ikely possibifi~ies are the
e.jsgence of g gone for ta i l anomal ies l inked t,o sin, or the exis tence of bwo c]Jfferent ~aiI
~oma/ies in t he s tock, one pemdiar to s,n@~n while the ocher is not . These quest ions can
~e~e be raised, b a t no t answered. The last co lumn in Table 2 gives the v i ab i l i t y of s~n/sm as deduced i i 'om the segregat ion
~f~i0s ~mong a d u l t surv ivors ; i t is much lower t han the cor responding va laes deduced
fkom:the segregat ion ra t ios a~ b i r th (Table 1). The difference be tween the two is a rough
~e~sure of the select, i r e m o r t a l i t y af ter b i r t h ment ioned pre~dot~s[y~
Table 3. Segregatio~ of Syndactylis~n in glutens of eenbryos
Type of mating ,.. TobaI
No. O ~ O No. of li~t~r~ 4- s~n omhryos ~'[e~n litter size 1 +/sm +~sin 4 26 5 31 7.75
s~)m + / s ~ * t2 &9 56 105 8-75 3 ~m/s~ 7~ normM 18 -52 90 14~ 7-89 4 sin~sin 7~ normal~ 7 ~0 0 70 10.00 5 sm/.sm sin~sin 2 0 10 i0 5"00
"Lhvo o f thes~ ht,~s z~me s m~*,,ings of +l~m c~d • ~m#m 9% One o f these li~,f~er~ came f rom ~ ran.ring o f -F2 normal c~ • s ~ / , ~ ~.
Segregation data from li~ters of 12-]fi-day~old embryos ~re gi~ef~, in TM31e 3. The olassi- fi0~tion of sm/s~ is qMte easy f rom the 13-day s tage onwards, b u t is somet imes a l i t t le
�9 ~ u l t in 12-day etnb~:yos; while i t is betievect l~hat the l a t t e r h a v e been cor~:eogly
01/Ssified, the poss ib i l i ty of a smal l quota of errors cannot be excluded. The unseleeted - ~ .~and b~ckcross t i t ter~ (nee. ] and 2, Table 3) are fair ~ p r o x i m a ~ i o n s to :3 " 1 and 1 " ] ra t ios
respectively which s t radd]e the expec ted vak~es. 18 out of 25 F~ normal s (nee. 3 and ~, i.Tgbh 3) p r o v e d to be + / s ~ and 7 § / § ; this is in fMr agreement wi th t he expee6ed 2 : 1
rmglo. However , t he 18 backoross l i t te rs of no..-3, Table 3, show a h igh ly signif icant excess
o~.Sm/sm over ~iorma] embryos (X ~ = 9-6~1 ; ~ = 1 ; P = 0'0018 approx . ) ; bhe d%ree of
selee~io~ inhe ren t in ~hese da ta is so s l ight t h a t i t canno~ account for the excess of sTndac~yis I t may , of course, be due go an aociden~ of Sarm}p]hl.g. An excess of a m u t a n t
~Ype is a v e r y rare even t and ~o~ ~eadily explicable. As more e m b r y o m c maberial is going f:~.5; eollec~ed, i t seems bus3 a~ this j unc tu re ~o awai~ whether the phe nome non will r epea t
it~etf in a la rger sample , or whether i t wi]I prove ~o have bee~ due to a run of luck.
Anatomg, The descr ip t ion of/)he aclulb a n a t o m y is based on a l izar in c learance prepara,- ~iO~s of ~6 sm/s~n mice ranging in ~ge f rom I6 be 286 days , Iror ~I of gh.em n.ormal l i t ter-
mates are ava i l ab le for comparison. I n every case, all :four :[:'eet~ ~vere a.ffecged. I)igibs m a y b~oined b y sof~ ~i.~sues only,* or ~hey m a y be jo ined b y bone. In. Tab le 4, digi ts inc luded
Ia normM forefee~ and hindfee% twin ses~moid~ ~,re presen~ on @e vole,r aapee# of ca, oh megae~rpa.l and on ~he plantar asl~oe~ of e~eh mehat~zsa'[~ in each ea.~e near its distal end. $2nd~etylo~s digits ~*igh o.sseous fusion 6[phalanges usually have only ~hree sesamoids between them, the odd one in the midclle b~mg common ~o both ~f'them though often composite (e.g. Figs. 2 ~nd 3). ~fhe s~ae is often ffound where s.vnd~etylo~as digi~s ~re 9~herwL~e only held ~;ogeghe," by so{'~, titmice (e-g. digit 2 ~nd i'~ composite oeighbo~w in Fig. 2~.); in such cases flit eddii>iom~l atga~hmen~ by means of the common se~az~loid ha, s bee~ ignored.
SOme an@ors h~'e useff ~he word 'zygodsetyli~m' for e~ses where the adjaee~.~g digits are joined by soft ..~ue~ only, 'sy~d~etyllsm" being reserved for o,~seoua fusione. A.~ g~d~ p~per w.ill give extensive evidan~e ~h~g {'~:ese ~brms ~ure only the ext, remes of ~ continuous distrib~gio.u, ~ d ~s a,II Our three genes produce ~he whole .ta:~ge of phenomena, we have refrMJned from uMng ~ terminology w'h[eh ere~,ges a gulf where ~o~ae ill f~ct exist,s. ~ll types of iimion between ~djaeenI~ digigs have thus be.:~n referred to a.s syndaotylimn, no m~gter whether they .a.re held ~ogegher by soft tissues only or whet.bur there is g Inore in~imt~te fusion by cartilage or bone.
8-2

116 Genetical studies on the skdeton of the mouse
in bracke&s are fused by bone (though l~ot necessarily along their whole length) ; otherwise they are fused by soft tissues only. The syndactylism, of the forefeet is usually by means of soft tissu.es only (r out o:[' 52 feet), while in the hind feet osseous fusions were reg~darly presenL It is obvious that the primary Nsio~ takes place between digits 3 and ~, as thes~ are syndaetylmls in every ease (for a single exception see below, Fig. 2b). Digit 2 is al.~0 ot'~en included; however, the prilnacy o:f the 3 + ~ f'nsio~t is obvious in m~ny oases (e.g: it~ Figs. 1 e, ha) by the fact that digits .3 and 4 are fused by bone while digit 2 is attached ~o them by soft tissues only. No~ rarefy, all three middle digits are fused by bone (Fig. 1 b), in the living animal, such %ee look rather like those of a bird. Digit 5 is never affected.
Table 4. Types oJ: fl~sio'n.~' encou'ntered i~ twenty-six ~d,uh syndactyb D i g i t involved Fores l-lhadfee~
3, '4. 26 - - {&-*) - -
2, 3, ~t 18 - -
(2, 3, =') 3 r e
i , 2 (~, ~) -- a
( i , 2) (& ,~) -- i
(I, 2,3,4,) - - 1 Total 52 52
t-Iowever, digit 1 is sometimes involved in the hindfeet; ff so, osseous fusions betwee~ digits, t and 2 are confined ~o the ~erminaI phalanges ; the general impression is that such involvemen~s of digit 1 are secondary or~ly. In ~h.e forefeet, digit 1 cannot be involved, as the rudimentary pollex of the mmlse is in any ease completely syndacbylous wi~h the rest of the hand.
Where there is an osseous fusion between neigJabouring dgits, ~his involves as a rule th~ phalanges only; the metacarpals and metatarsals remain separate throughout thei~ length (see, however, below for exceptions). Usually, all three phalangas are fused,
Among 37 hincl feet with osseous fusion between digit s .3 and 4b only, aE three phalange~ were Nsed ,33 times ; twice %sion was confitmd to phalanges I and2~ the terminal phalange~ remaining separate; and ~wiee R~sion was by means of phalanges 1 and 3; in the lat~er., gJo.e fusion of the terminal phalanges was probably a secondary event. The remaining i5 hinclfeet had osseous fusions involving all three middle digits; in each case, the fusi0n. between digits 3 and 4 was complete, but the attachment of digit 2 to its composite neighbour happened as follows:
By ph~Iange~ 1, 2, 3 9 times By phalanges 1, 2 0nee By phalanges 2, 3 3 times By phalanx 2 Once By phalanx 3 Once
On the referees the situation is similar, excep~ that osseous fusions are uncommon (8 6 ~ of 52 fee~). Five times digits 3 gnd 4 only were affected (fl:~.sio~ by all three phalanges 3 times; by phalanges 1 and 2 once; and by phalanx 3 once); one nqo~se had fusi0n~ involving all ~hree n/ddle digits on both forefeet; in each ease, the %sion betweell digits arld ~ was by means of the termin.i phalanges only; *,he fusions between cligits 3 and.~" involved all three phalanges on one side and phalanges I and 3 oI~ ehe other. Finaii.y, there is the foot illustrated in Fig. 2b which is completely exceptional, in that the prima~Y: fusion is between digits 2 and{ 3 rather than between digits 3 and 4.
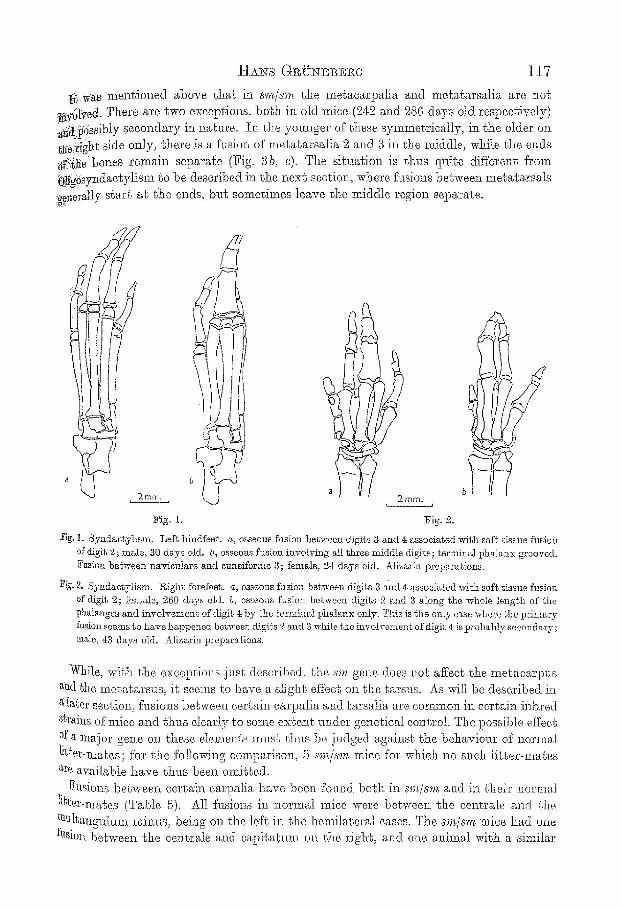
H ~ s G g ~ N E ~ c ~ 117
!~...vCas msntioued above that in sr the metacarpalia a~d metatarsalia are not ~rb[Ved. There are two exceptions, both in old mice (242 and[ 286 days old respectively) .~41)0ssibly secondary iz~ nature. In the yo~mger of these symmetrically, in the older on ~0.right side only, there is a fusion of metatarsalia 2 and 3 in the middle, while the ends ~f~:~he bones remain separate (Fig. 3 b, e). The situation is thus sit, ire different from 0!i~oSyz~dactylism ~o be described in the next section, where fusions between metatarsals ge~m.ally start at the ends, bl~g sometimes leave the n~ddle region separate.
2mm+ t - - I
Fig. 1, ]Fig. 59.
b
/
Eig. 1. 8yndaetyliem. Left hindfeet, a, osseous fusion between digits 3 and 4 associated vdl,h soft tissue fusion of digit 2; male, ao days old. b, osseous fusion involving all ~hree middle digits; terminal phalanx grooved. ] ~ i o n 5etween na,rienlare and e=neiforme 3 ; fmnMe, 24 days old. Alizarin preparations.
iP~g. 2. Syndastytism. R,igh~ forefeet, a, osseous fusion between digits 3 and 4 associated with soft tissue fusion of digit 2; t~male, 260 days old. b, osseous fusion between digits 2 and 3 a b n g the whoIe lengbh of the t~halanges and involvement of digit. 4 by the termh'lM phalanx only. This is the only case where t, be pr.lnt~ry Naion seems to have happened between digits 2 and $ while the involvement of digit 4 is probably secondary.: m~te, 43 days old. A'[iz~rin pre]?a.rat,%ns.
While, with the exceptions just described, the s~~, gent does not affect the metacarpus and the metatarsus, it seems to have a slight etfeet on the tarsus. As will be d.sseribed in ~ia~er section, fusions betweeu certain carpalia and tarsalia are common in certain i~]bred ah'ains of mice and thus clearly to some extent m~.der gsnebical toni)tel. The ]?ossible eft'cog of a major gone on these elemeiihs must thus be judged against the behaviour of normal litter-mates; %r the following comparison, 5 s.m/~',z mice %r which no such ]it, tot-mates are available have thus been omJtt~ed.
Fusions between certain oarpalia have been found both in srr~./s~)'~ and in their normal litter-mates (Table 5). All %sions in normal mice were between ~he centra].e and the tadeangulum n~in~l.s, being on the left in the hemilal~eral cases. The sm/s','~ mice had one Nsion between the centrals and eaphtatrtm on l)he right, and. one animM wi~h a similar
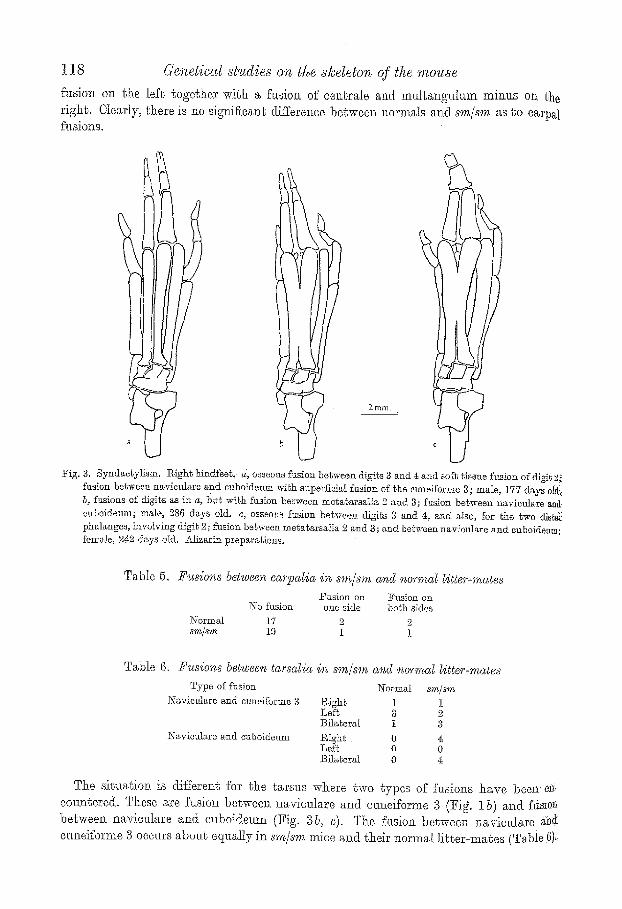
118 Genetical studies on the s]cdeIvn of the mouse
flmion on ~b.e lef~ together wi~h a fusio~ of cenl:r~le and mul~angultun minus on ~he righ~. Clearly, t he r e is no sig~figeane difference b e t w e e n normals and s m / s m as Co earp~l fttsions.
[ 2ram. �9
Fig. 3. Syndae~ylism. iZight l~dfeeg, d, osseous fusion bel~ween di~ts B and 4 ~nd soft iissue fusion ofdlgif2~ fusion betwee~ naviculare and euboideum wi~h superticia[ fusion of the etmsfforme 3 ; male, 177 days oldi~ b, fusions of digits a~ i~ a, t)ut with ft~ior~ between me~arsatia ~ and 3; fusion between navieuIare and cuboideum; male, 286 ds,ys old. c, osseot~ fusion between digits 3 and 4, and also, for the l~wo dls~g[ phalanges, involving digi~ ~; fusion between megatarsalia 2 and 3; and beSween navionlare and euboideam; female, 2r days old. _41izarin preparations.
Tab le 5. 2~'~sions bet~veen earpa~ia in s,r~@m and uofmaZ gitter-mates Fasion on Fusion on
No fusion one side bo~h sides Normal 17 2 2 sm/~m 19 I I
Tab le 6. Yusions between tarsa~ia iqz xm/s~n aqzd noqw~a~ litter'-mates TyVo of fusion
Ns~viculare and ouneiibrme 3
Naviculare and euboideum
~N-ormat s.m/sm P~igh~ I I Loft; 3 ]3ilaterM I 3
Night. 0 4: Lei% 0 0 ]~ila~eral 0 4
The situation is different for the tarsus where ~wo types of • have been:eu-
coun t e r ed . ThEse a re fus ion between, na .viculare a n d cune i fo rms 3 (Fi.~. 1 b) a n d f~mn
bel ,ween n ~ v i e a l a r e and eubo ide t tm (Fig. 35, o). T h e fus ion b e [ w e e n n a v i c t d a r e and~
c u n e i f o r m s 5 occurs a b o u t e q u a l l y in sm/sm mice a.nd t.heir n o r m a l l i t t e r - m a t e s (Table.6)-.

0;t'.glis other hand, the fl~sion between ~avioulare and enboidemn seems to be confined t,e r syndactyls.* tror a sig:aifieaaace test, ~he data may be az'ranged izt a 3 x 9 table as
t0~to~S~"m" : Normal ~m/sm
No ~hsiota 1.6 7 Nav ieu ta re + euneiforme 3 5 6 Navicu]are + e~boidenm 0 S
e/hence X "~= 11.612; ",,=2; P=0 '003 approx. I t thus appears that the sm gone does not mffusncs the ft~sions between navieu].are and cuneiforms 3, but is responsible for fusions between naviculare and euboideum. However, as only 8 oat of 21 s'rn/s~)~ mice were ~hus ~ffe~ted, i~ is probable tha t this is a remote effect of the gone.
I~ was mentioned tha t many sm/s~n sy~daetyls have various tail anomalies. 1 t of gh.e ~6 ~dults studied in alffzarin preparations had completely normal tails. The remaining ifi~}'animals included a few with tail anomalies too slight to be de~ectable in the living mo~se. The region affected, is exchsively the distal half of the tail, with a strong preference [0~%he third quarter (caudal vertebrae 16-21), while tlie last quarter is agMn less affected ~8~:vertebrae affected in region 1.6-21, 1.3 in region 22-27, with a single ease onvertebra 14). ~ffsoted vertebrae may be ordy slightly shortened; often they are vdry small and then us~ally more or Iess wedge-shaped. I t is common for a normal vertebra to be flanked by .two abnormal ones such as one would expect to find in the case of a tail ~wist. Embryo- !~gieal Nets to be reported in a later paper seem to indicate that the osseous tail anomalies ~e preceded by azaomalies of %all shape prior to the chondriflcation of caudal vertebrae.
No other effects of s~. or~ the skeleton have been discovered.
L l m m . b
)
]~ig. 4. Synd~,cLylis,~l. Loft, hindf'eeg of Id -day-o ld embryos, ~, n o r J ~ l ; b, ~m/sm ([it%sr-ma~ea). l~{e~hy!ene-blne prepara t ions .
~mbryoi~yy. s~n/~'m, el?abryos can be distinglfisl~ed from their normal litts~-ma.tes by rife shape of their feel, froze the ].2oday stage onwards (earlier stages ].,ave not yet been examined); these early manifestations will be descril~ed on a later occasion. In 14-day .~mbryos (Fig. 6), Carsa]ia and metatarsalia ha;~e slatted to ehondrify, and the basal ~halanges of the t)aree middle di.gGs have made their appearance. In the. sm/8~ embryo, the otttline of the footpl~te has four projections for the digits rather tlxa~a the nor~aal five, ~nd .metatarsalia 3 and 4, instead of diverging as in the m~o~'ma,l embryo, converge and have ~,. co]nr~on basal phalaI~x. Strictly spsaki~a, t]~ere is t:hu,s no 'fusion' between adjasen.t ear ~ilaginous pha] a, nges as a tempos)be phalanx is pros ent from the very b eginning. ~s ~he ns and ulti:mately ehe termhml phalanges ehondrify, they ~:tsaall.y oon:~il~.ne to
* ~INere were %we more b.ilaeeral eases and oz~e oJa Lhe righ~ among ~t~e five s m ) m mice which had to be
~&cl~ded here a,s no no rma l liI',ter~mages a,re ava i l ab le for comparison.
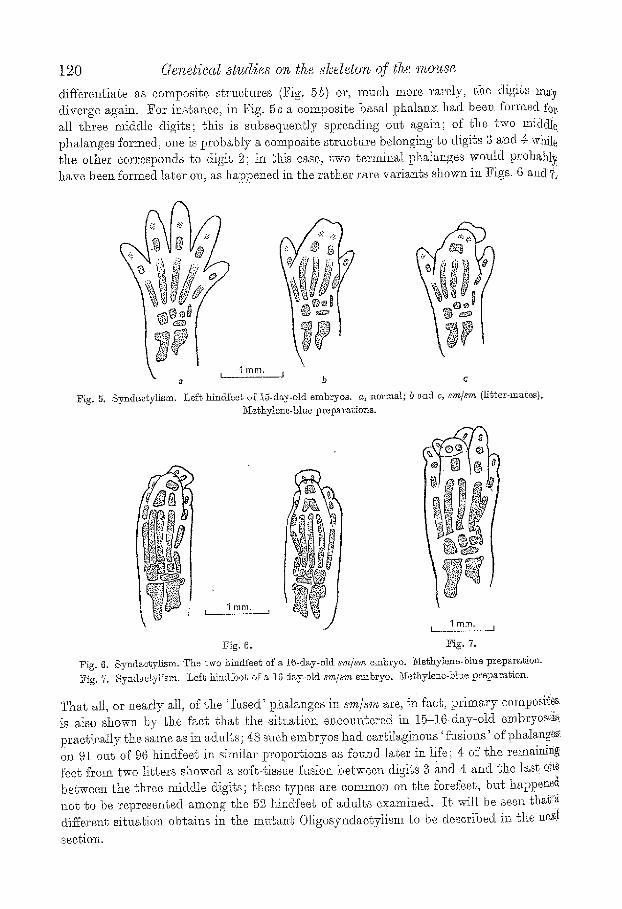
120 r s~udies on the slcdetoro of the mouse
differentiate as composite structures (Fig. 5b) or, much more rarely, ~he digits m~y diverge again. For instance, in Fig. 5 c a composite basal phalanx had been formed fez' all three middle digits; this is snbseqr~ently spreading out again; of the two m.iddIe phalanges formed, one is probably a composite structure belongilag to digits 3 a~ad 4: wNle the other corresponds to digit 2; in this ease, two terminal phalanges woald probabl~. have been fbrmed later on, as happened itl the rather rare variants show~ i~x Figs. 6 and?.
\ .1 mm. a
~ig. 5. Syndactylism.
l ] b c
Lef~ hindfeeg of 15-d~y-old embryos, a, norton.l; b and c, s.m/sv~ (Hb~er-maees)~ ~Ie~hylene-blue preparagions,
I royal. L 1
�9 1 mm. r 1
Fig. 6. Fig. 7.
Fig. 6. Synd~e~ylism. The two hindfee~ of a 16-d~y-old s,m/~m embryo. ~iethylen~-blue pr~t~ara~ion. Fig. 7, Synd~etyH~m. Lef~ hindfbot of a 10wday.old sm/~'m embryo. Nethylene-bh~e preparation.
That all, or ilearly all, of the 'Nsed ' phalanges in sm/s~'~ are, in fact, primary composites is also shown by the fact that th.e siLuatiol~ encountered in 15-16day-old embrycs..::is practically the same as in adults; 48 such embryos had cartiIaginons ' fusions' of pha]anges. on 91 out of 96 hindfeet in similar proportions as fouz~d later in life; 4 of the remain]N feet from two litters showed a soft-tissue fusion between digits 3 ~nd 4 and the last one between the three middle digits; these types are common on the forefeet, but happene~ aot go be represented among the 52 liindfeet of adzdts examined. I t will be seen t h a ~ differene sittmtion obtains in the mu~ar~ Oligosyndaetyiism to be described in the nex~
section.

H.a ,xs @X~~-NSB~R@, t 2 1
While the phalanges are thas composi te f rom the s%art, this does not apply to the t~rsalia which originate separa te ly f rom each other (Figs. ~-6). Cartilaginous fusions between navioulare al~d euboideum are found in newborn s#n/sm; t hey are thus real lesions between e lements which were originally separate. The same ( thoagh p robab ly between bones} is clearly the ease %r the rare %sions between metatars~lJa encountered
in old sm/s~n mice. OLzc4osY~cmYzrs~
@e~,eties. This dominan t mu tan t , which I owe to the kindness of Dr W. L. ~ussel l (Biology Division, Oak Pddge Nat ional Labora to ry , O~k g idge , Tmmessee, U.S.A.), ~rose
r~diatiou exper iments . The fa ther of the first m u t a n t animal was an av, male f rom a 0ross between the inbred strahas 101 and O g H s a d had been i r rad ia ted with a dose of 600 r. of X~rays before being ma ted to an unt rea ted female of a mul t ip le recessive strain; ~he gone thus p r o b a b l y arose as ~ r e s Q t of the t rea tment . Two affected males which were Nm" or five generat ions removed f rom the origi~aal mutant female reached me in October 1958 through the good o~ices of Dr D. S. Falconer (Edinburgh). T h e y were outcrossed to
T~hle 7. Seg~'egati~n of Ob;gosy,~dac@ism a,s recorded at birth or soo~ @e,r No. Type of math~g No. of m~thngs Oa + Total
1 Os/+ x + / + 8 69 52 1'21 2 Os/+ x +/-4- 6 61. 64 12.5 g Os/+ x Os/+ 13 233 10t 334:
Note. No. 1 is the outcrops of the original male~ to CBA females; no. 2 are braker'oar-type mabiugs in ~a~r generations.
Table 8, SW~'egetion of O~igosy~zdae~yZism in ~itters of embryo~
Type of mating e - -
No. s 1 Os/+ + / +
+ / + 0.#+
- - x No. of Solid Dead litters Os + moles embryos Total ~le~n litter size
19 7~ 8.5 9 4 t72 9"05 7 80 24: 6 - - 60 8"57
26 104 109 I~5 d 232 8"92 3 Os/+ Os/+ 14 44 28 20 - - 92 6.57
females of the inbred straha @BA/@.r, and the whole of our mater ia l iv derived Kern t ha t outcross. As will be desmibed below, tJQe anatomical s i tuat ion ranges f rom soft4issue Nsion. be tween ad jacent digits (syndactyl ism) down to complete absence of a &igit (oIigodaeWlism). The gone m a y thus be calt~d 'Otigosynd.acbylisns' with syxubol 'Os'. The condition is easily classitiable a t bir th, a r d the affected animals are of about norms1 sdabilRy and ferti lRy. Whereas be, choreas- type mat ings give a reasonable fit to a t : I ratio (i30 : 116),/7~ mat ings do not :fit a ,3 : 1 rat io a.t all well (XZ=4,'S90 ; ~ = 1 ; ]?=0"027); on the other hand, they are iu good agreement with a 2 : it ra t io (X e = 1.4.39 ; oa = 1 ; P = 0-23). This sugges% t h a t the Os/Os homozygote m a y be inviable.
The same s i tua t ion is again encountered ha litters of embryos (Table 8)~ T ) e baokeross- type mat ings give a close approach to a 1 : 1 ratio (10~f : 109), bu t the F 2 ratio differs significantly s a 3 : 1 ratio (X2--.7.4:07 ; ~=1. ; P =0-0065); it is in good agreemen~ with a 2 : 1 rati.o (X ~ = 1.000 ; n = 1 ; P - 0'32). Combining the da ta of Tables 7 and 8, the back- e, ross rat io is 234: : 225, the if2 ratio 277 : 129. Fur the r evidence for the de~e].~ i9~, utero o:f
.the Os/Os homozygo te cornes f::om th.e fact that there are significantly more 'solid moles ' m Os/+ x()a/-~ tBan in Ca /+ x +[ -F ~nd + / + x O s / + m.atit~gs (X2-15-46; n = I ; P=0-000,085). 'Sol id moles ' are small p lacentae whose embryos have been completely

].29 Genetical sa~dies on lhe slce~e~on of ~he mm~se
absorbed and thus represent zygotes whxch have been eliminated aftez' implantation. A small quota of such cases is neazqy al~ays to be fou~d, bLI~ ies signiflc~n% i.uc~eu~e ill OS/+ X OS/+ rantings is strong evidence tha~ ~he excess of solid moles inck~des the Os/O~ homozygotes, or at least par t of them. ~4~ctually, having regard to the background per- centage of embryonic death, the incidence i.a the 7?~-type rantings is less than the expcs~ed~ 25 %; perhaps some of ~he Os/Os homozygotes die before implantation, in which ease their absence would only be noted m a reduced litter: size as is indeed probably the case (see last column of Table 8). As more ma.terial will have to be collected for the aetna! ideneiScaeion cf the O~/Os homczygote, the ma~ter need. nee be discussed here ihrther
Table 9. Ano'magies of tlz~ d@its' i'~ twenty-xix c~du~t Oi~9osyndac@s. sta~ed, syndac~y~.ism i.~.~vd'l,es d~gits 2 a,r~d S onL~
No. Type of anomaly Yorefeeg 2[tindfeeL I Synd~a~ylism by soft tissues only 3,].. 18" 2 Osseous syndaetyliam by phalanx i 7 9 3 Os~eo'ns syndaety]ism by phalan~ 1 and 2 g 5 ,1, Osseous ~y~.d~ctyHsm by phalanx 1, 2 ~nd 3 3 I0 5 Osseous s2ndae~ytism by phalanx I, 2 and 3 and by ."lj ~ 2
megae~rpa-ls or mel,~tarsals 6 Oligodaetylism 2 6 7 Oligod~cbyHsm with polyclaeVti~m 0 2
Total 52 52
* This includes 3 t~et m which digi~ ~i was also superficially webbed. ? In one ease the end phalange~ (3] were separate.
U~.~Zess otT~e~r'w.i~e
i~a~omy. Materi~h 26 alizarin clearance preparations of 08/+ mice ranging in age from I6 to 968 days together with. 90 normal litterrrnates. Whereas the effects of s~n/~ are: almost confined to the region of the phalanges, all parts of hands and feet are affected in Ox/+ mice, and hence the anatomical situation is considerably more complex.
_~AI four feet are regularly affected. As in sm/sm, the ibrefeet are involved to a lesser extent than the h in~ee t , but the difference is much less marked; thus there are more.. osseous fusions on the forefeet, while on the hindfeet soft-tissue Nsions are much coin- moner. With few exceptions, syndactyli.~m is confined to digits 2 and g (disregarding �9 fusions between metaoarpaHa and metatarsalia ~ and 5 which will be described later):
a general summary is given in. Table 9. Some of the anomalies ennmerated in Table 9 are shown in l~igs. 8-10. Thus synda~-
~ylism by soft tissues only is shown in Figs. 8 b an& 10 b: osseous involvement of ~he basg~ phalan~ in Fig. 80, in .vehement of bas~l, and middle phalanges in ~ig. 9 ~, arid of all three phalanges in Fig. 8 d- In Figs. 8 e and 95 ~he fusion of all phalanges is accompanied by ~: fusion of the corresponding metatarsals which are, however, still separate in the middle: Finally, in the foot shown in Fig. 8]] a state of oligodaetylism has been reached in which nothing indicates any more tha t the element next to the hallux is a composite of digits, ~
and $~i f , indeed, i t is. Events happen in a similar fashion on the forefeet, bu~ the more extreme reductions
are Iess commonly encountered. Two stages only are shown in Fig. 10: soft~Lissne fusion of digits 2 and 3, and osseous %sion of the same digits involving th.e basal phalanges. Th~ general cm~rse of events as deduced from adult specimens is thus as follows. I f the fasio~ does net remail~, confined, t;o the soft tissues, osseous fueiokt starts with the basal phalangeS' amd ehenee proceeds in a diseal direction until all phalanges are %sod tc each o~her bY'

HA~S Q~ON~BE~e 123
bone, Only ai'ber this prooess has reached ssturr do the metacarpals or mebatarsals become involved, and complete eoalesoenoe may ultimately lead ~o a st~te of o]igodaetylism ~ which digits 9.9 and 3 are represented by a single element whioh does no~ betray ~xtez'- nal!y that it is composite. We shall see latex', on examining embryos, that this deduction ~ probably in need of amendments.
r ' 2 mm:.--r
d
lqg, 8. Oligosynib;o|,yi~sm.' High~ hind~eet, a., :normal c!, :33 d~ys old; b, Os/4- o 23 d~k3~s okl; ~, Osj + -9, 33 days old; d, O~/+ 9, 36 d~ys old; e, Os/+ ~, 33 d~y,s old; j[. O~'/ F ~, 33 d~ys old.. a, c, c a n d f ~re libeor-mates, b ~rl(] g sisters from different, lit.~ers, Expla.l~al,ion h~ the ~ext. Alizarin prepar~io.as.
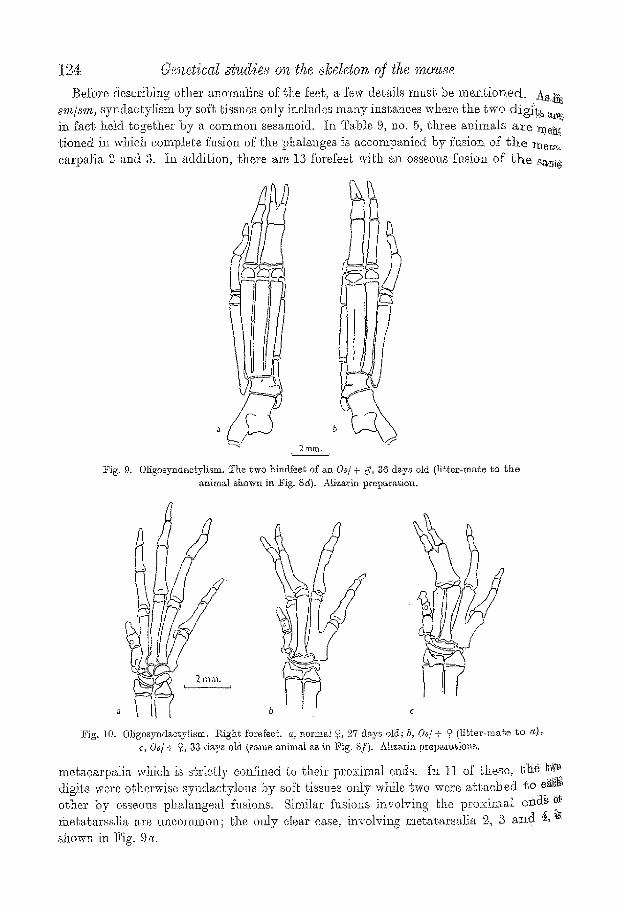
t24- 6.'/snetical ~tud~es on the ~keleton qf t, he mouse
Before describing~ othei' anomalies of the feeh a few del, Mls mush be m.en~,ioned. AS:~; ..... s'm/sm, syndzc~ylism by soft tissues ozfly inelndes many instances where ~.he two d ig i~ a~.i, i~ fact held together by a common sesamoid, he Ta])ls 9, no. 5, three animMs a~e :me~ tioned in which complete fklsion of the phalanges is accomt~anied by fusion of t h e meT,~ ca~'pali,~ 2 and 3. In addition, there are 13 forefeel~ w:ith an osseous fusion of t h e sa~s
!} 2mrn.
-~ig. 9. Oligosyndae~ylism, ~he ~wo hindfeet of a~ 0~ /+ c~, 36 days old (lil,ter-ma~e ~o t h e ~nimM shown in Fig. Sg). Alizarin preparation.
�9 \ \
~ ITIm.
Fig. t0, 01igosyndactv]ism. l~ighg forefeet, a, normM o 27 days old; b, Os/+ o li'e~er-ma~e 1,o eQi:,. c, 0 s / + 9, 33 ds~ya old (s~,me animM ,~,e in :Pig. 8f). Aiizarin p r e p ~ t i o ~ s .
metacar:pMia which is s~ricdy coni3.ned ~o their proximal ends. I:l 11 of Lhese, t l ie ~o digits were o~hsrwise syndacty.lous by so~e tissues only while two were a gtac}oed go Si~ii, other by osseoas pi?alangeal %sio~zs. Similar Nsions involving Ge proximal ei/aS ~:~ me~agarsglia are uncommon; the mJy dear case, involving metatarsMia 2, 3 a~d ~ :is. shown in Fig,. 9a.

125
?{hereaS ~he invoh:eme~t of digit~ 2 al~d 3 seems to be' primary', nearly all animals show ~sseet~s f~sions involving the bases of metaoarpalia 4 and 5 ~nd simil.arly of me~atarsalia 4, ~ud!5. Ia~ the forefeet, this always leads to a very marked abduction of digit 5, while i~ ~e hindfeet the angle is often much sm~ller. Exampl~ on the hindfeeg are shown in ;~!g. 8b@ and on the :forefeet in Fig. 10b and c, There were only two animals which were ;ex0et)~ional in this respect. One of the two males originally imported and used for the ia.t~&cross had no saeh fusions on any of its four feet (the other male was not made into a. t~eparation) ; and the animal shown in Fig. 9 Iaeked fuions on the hindfeet, bat no~ on ,flie forefeet. The probable reason for these %sions will be discussed later.
~?~ble 9, no. 7, is an item called' oligodaetylism with polydaetylism'. This somewhat 2~adoxical designation h~s been chosen for l~wo ~dNt, hindi'eet aud several others fonnd ~i~0~g the embryos which have five toes and thus, at first glance, might be th.ought to be
b
I t r im. ! J
~[~. 11. Oligos~d~etylimn- ~lindfeet of two I6 d~y-old embryos ~dSh ~ com~h~tion of otigodacty~ism and rolydaotylism. ~lethylene-blue prep~r~ions~ Expl~na.tion in the text.
n~itker oligodactylous nor polydsotylous. In sc.taal fact, as shown in Fig. 1 I, both pro-
eesses seem to be at work at the same time. Both feet shown have three ordinary toes
:with 3 phalanges each; and it"ore t.heh' position on Lhe feet it is clear that these are digits 3, r 5. Next to blhese t]]ere are two elements whi.oh are much shorter and syndaotylous by soft tissues ; both of them have two phalanges only and, in this respect: they must be classified as h~llaces. ! t is ~rue that in the specimen of Fig. ]la,, the lateral of gb.e Vwin ~[ements artie~dates with the cuneiforme 2 and in this respect betray,~ that it is really �9 a modified digit 2. Iioweve:r~ in the foot shown in. Fig. l ib , t:he a.rtieulatiou of the e0rrespo~ding digit is mainly with the cuneiforme 1 (though a, small process still ex6ends ~0Wards euneiforme 2). In the most extreme cases there is ~ single bloated metatarsale 1 with two digits like a. :fork. It th[ts appear~ that ma~ex~ial u,~uatly desgiued go form digit 2 l~as bee~: shi:ged, towards digi~ ] ; the material for the hallux is thereby increased wighthe �9 rein t that a doable ha,lht~: is being formed. The foot i.s thus a]?]?aren gly oligodaetyl.ous :for @[gig 2, hug po]yda.cgylous for cligit 1, the net result being a pe.neadactylous foot. Now it i~ >robffble that a shi:fg of mater:is] from the region of digit 2 towards the hallax might not .lekd to a double b1~e to a s.ingls hallnx. In that ease, ol]~odaetylism~ . . of' digit 2 would

!26 Ger~e~ic~l sa~dies o~ J~e sfcdeton of ~]~e ,mo~se
result, but the digit next to the hallux w e n d not be a composite of digits 2 a~d 3 (as discussed on the basis of the adN.t specimens), but would represent digit 3 only. Further. evidence in the same dii'eetion will be presen%d later on.
We mention here a fusion between me~atarsalia 1 and 2 which has been seen twice) bilaterally and four times on the le~ side. in such o~ses, the metatarsMe 1 tends to be very short and 5s intimately attached to its neighbo~r either entirely or along most of its length.
The gone for Oligosyndactyiism regularly produces extensive fusions i:a carpus and tarsus. The 20 normM litter~mgtes available for comparison showed, a similar ~baci~- ~ o u n d effect' of carpM fusions as those of the sm/~ stock, there being altogether sevei~ hemilateral Nsi0ns, as follows: capitatnm and centrals 3 times (right); multangulu N minus and centrals 4 times (1 right, 3 left). Unlike the other grou.p of normals, there wen no tarsal %siena at all.
The 26 adt~lt Os/§ mice showed the following carpal Nsions:
Feet mul~angulnm mians--cen~rMe--oapitagum 1 multangulnm minua--centrMe--capi.tatum--hamgtum 28
centrale--capi~attm~--ham~t um 21 centrale--capitatum 2
The m~t[tangLdam maitre was never involved in fusions. The two more extreme types of fusion are shown in Fig. I0 b and 6. In the proximal row of carpalia, most animals ha& a fission between the scapholunatum and the triquetrnm, with ~he pisiforme often attached to the latter (l~ig. i0@ In preparations of this kind and without dissection, it is some-. times difficult to be quite certain whether in a given case the bones are completely f~tsed or only near that state (as in Fig. 10 5). For that reason, no detailed figures will be give~ for the fusions in the p~oximal row of carpalia.
The fusions in tire tarsus are perhaps even more massive. They always include a solid fusion between talus and calcaneus to form a taloca]caneus (Figs. 8 6@] 9 ~, 5, and 11 a, $). ~& second striking anomaly is the fact that the eaIeaneus invariably lacks the processus trochlearis, a very prominent flange of bone on its antero-la~eral side and thus not far: behind the proximal end of the metatarsals 5.
The talocaleaneus is never fused with any of the dis~al tarsalia; nor is the tibiaie mediale ever involved in any fusions. _42I other tarsalia are liable to get included though to a variable extent. Before describing these anomalies in detail, we have to discuss tti~ absence of one of the tarsalia which is invariably found in Os/+ mice. The normal arrangement of the adadt tarsalia is showil in ]fig. 8a and that in 15~day normal embryos in Fig. 13e. Comparing the latter normal animal with i~s O,s/+ litter-mates on the same figm:e, it win be seen that the group consisting of navictilare, ctmeiforme 1 and cuneiforme~ is about the same in normal and abnormal animals. I-towever, while the normal animal! has two additional elements, the cuboideum and the o~meiforme 3, there is only a sing!~ element in the Os/+. In later stages (Fig. l l e) it can be seen tha~ this single element: a,rtic~lates witb. the calcaneus at ~he back and with the metatarsalia 4~ and 5 in front, i.e. it Behaves like the euboidm~m; however, it also articNates with the metatarsals ,3, amili~' this respect it behaves like the cuneiforms 3. I t is thus certain that this element repre- sents both the cuboid.e~m and the o~meiforme 3; in fact, aafieipating evidence to be: presented la%r, it is a primary composite and not derived from a secondary fusion. F0~

[!~ldiff a better name, we shall refer to this element as ' the conlposite' il~ the following
�9 .The tarsal fusions enoonntered in 0~/+ mice are very varied. That between the na.viou~
i~re'and the ' composite' is always present (~'igs. 8, 9); it may be the mxly fusion in this :i.~;~gi0n, as in Fig. 8b, bu t this is rather unusual. Nore eo:mmonly, the cuneiforms 9~ is also N~htcled; ff so, the artic~tlation between cuneiforms 2 and navioulare is usual].y preserved .i~ig~i 8d, e, and 9 ~, b) though of co urse non-functional; ig is the articular surface which ii[:~i~bsf, rarely obliterated (though this ~,lso sometimes happens, such as in ~ig. 8f). ~ i form_ia 1 and ~9 may remain separate from each other and the rest of the tarsalia '.i~: 8b); sometimes they fuse with each other, but not with any other element (~ig. 8@ '~e~s are other variants which eamaot all be described in detail. Apart from the tibiale :~di~le which is always a separate entity, She caneiforme 1 maintains its independence more often than an}, of the other tarsalJa. '~naddition to these fusions between garsalia, there are certain fusions involving tarsatia
'~i~ mebatarsatia. Fusions between cuneiforms 1 and metatarsals 1 were observed three '~i~ies (two left, one right); and the he len between cuneiforms 2 and metatarsals ~ is }~her more common; it was found in 7 animals (three right, one left, ~hree bilateral) ; in ;such cases, the cuneiforms 2 often does not fuse with any of its other neighbonrs.
The redlmtion of the tarsus leads to secondary consequences. The navioulare is narrower )a~ ig no longer has a separate ermeiforme 3 t~ articulate with. The 'composite' is much '.narrower than the sum of its components when separate; hence, while it usually artietdates :.i;~ith metal,ar6alia .3 and 4= in embryonic life {?iga. l l , 14 ~), later on it often retreats from me[atarsa.le ~ more or less compJ.e~ely (Ieig. 8 b-f) ; raetatarsale 5 thus probably moves into a position of abduction largely for lack of suppoa% from the rear. in the cases where the ~eta~rsale 5 has remained separate (Fig. 9), fihe mechanical si~auati.on J.s more i'avom'able. i~:seems, however, that part of' the blame a%aohes to a reduction in the normal locking ::mechanism by which a process of metatarsals 3 fits into a corresponding baeision in :~m~[atarsale ~, and a sim.ilar process of metatarsals 4[ into a~a incision in metatarsals 5; it �9 ;s~ms ~hag these locking devices are all m~eh reduced in Os/+ mice (compare the normal iff.:l~ig. 8 with the five abnormal mice, and with Fig. 9).
..The 'composite ' is also shorter than its neighbonrs (navieulare and cuneiforms 2.). ~ ~ consequence, the long axes of the calcaneus and of the metatarsalia are no longer -~a~allel to each other as in the normal na0z~se, 5ut include an angle of about 150 ~ to 160% ;T~.e.hindfoot thus points outward. ~o that extent (pea valgus).
In the forefeet of Os/+ mice, the ulnar end of the h~matum articulates with. meta- ~a~pr r but does not react metacarpals 5 at all, a.s it does in the normal (Fig. 10). Zetaearpale 5 thus gets into ~,, state of abduction in which it becomes .fixed. The fusions l~'Ween me~acar]?alJa and meta,tarsalia 4 a�9 5 are thus clearly secondary to the reduc- highs in carpus and tarsus respectively.
No o~.her skeletal abnorma.tities have been discovered in Or/+ mice. ~'mbryd, oy 9. Os/-P embryos can be disting~fished from their no:r:m.al litte.r-mates by the
~hal)e of their foot plates ag th.e ~ge of t2 days, as wilI be described on a, later occasion. I~ l{-day embryos (ICig. ]2), o}~ondrific~tion as seen in meghylened)lue preparations has ~g~rtecl in the tarsus, the three middle metatarsalia (and very slightly in mete.tarsals 5) and in the basal phs.langes of digies 2 ,1-. In Os/+ embryos, the only detectable anom.a,ly ~Y be Chat the projections of digits 2 and. 3 at the edge of' the foot p l a t are closer
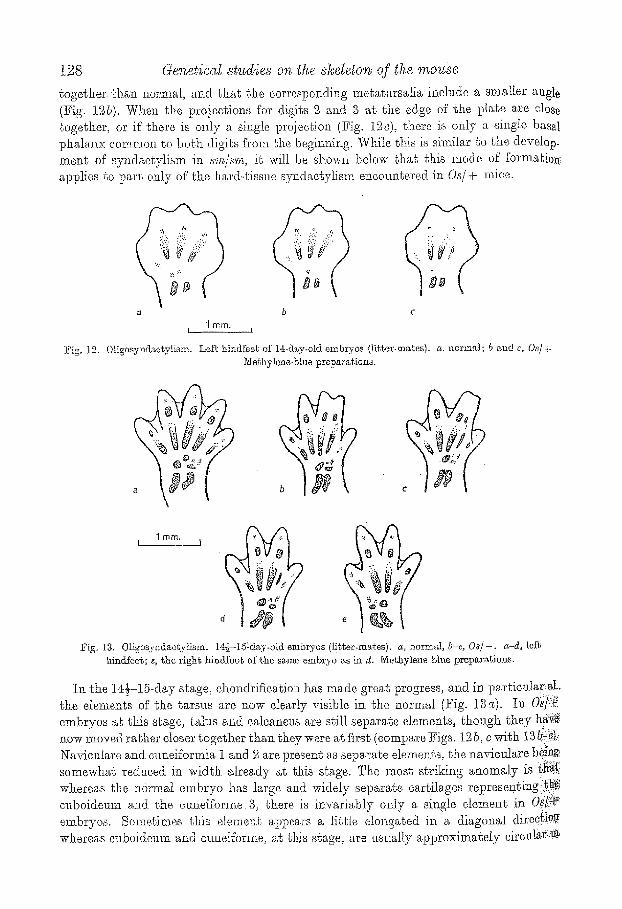
togethm" d~an normal, and tha~ the corresponding metatarsalia include a smaller angle (Fig. ].St). When the pro}e~t, ions fro" digits 2, .~md S at, l, he edge of the pla~e are dose togeSker, or if ehere is only a single projection (Fig. 19~)= there is only a singfe basal phalanx common to bedr digits from d~e begin%t~g, While J~is is sim.ilar ~o the develop_ ment of syndaetylism in a~/~'m, i.~ wilt be shown below ~hat this mode of f'orm.atiot~ applies to part only of ~l~e hard-tissue syndaetylism eneoungered in Os/+ mice.
a b c
J f i l m . I 1
"~ig. 12. Os Left hindfes~ of 14-rl~y-old embryos (li~t.er-ma~es), a, normM; b and c, Oa/+ 3'I~hylene~'atua preparations.
d .r
)
S m i T h t
? ?
~ig. 13. Oligosyndaetytism, I4.{-I5"day-ot4 embryos (litter-me&e@ G normal, h-e, Os/+. a-d-, lef~ hind_feet; e, the righ~ hincktbo~ of the same embKro ~s h~ d. ~ie~hyl~ne blue preparations,
In the 14�89 stage, chml&Ltioatio~ has made great progress, and in par~iotdar aL the elements of th~ tarsus are now clearly visible in the normal (Fig. 13a). In 0~/:::%' embryos at tkis stage, talus and oaIoanez~s are still separate dements, though tkey hgg~ Jlosr moved ~'abher closer ~ogeth.er ~ha~ they were ~t first (compare Figs. lg b, o with 13 h~}:.~ Naviculare and. eundformia I and 2 are present as separate elements, the navientare hdn~ somewhat reduoed in width, already at this s~,age. The most striking anomaly is Gh{[ whereas the normal embryo has large and widely separa% cartilages represenging:i}~{ ouboideum a.nd the cu~.eiform.e 3, there is invariably only a si,lgle element in Oai!~ ~ embryos. Sometimes this eleme~.t appeaes a little elot~gated in a diagonal dired[i0g whereas euboideum and cunei%rme, at this stage, are usually approximately oirort]ar ~

H ~ m s G ~ O ~ s ~ e 129
.~.~li~e~ Later (]~ig. 11 ~) it can be seen that this element artienlates with mstatarsalia ~gi~"in front and with the ealcane~s in the rear; i~ is ~hus clearly the ecl~ivalent of both ~t~boidenm and enneiforme 3 and appears as a single centre of ehondrifieation ti'om the ~gi~, While this is thus a case of primary composite, aN the other f~sions in the tarsus of N:['- : mice arc secondary. For instance, talus and cgleaneus have always coalesced to "~'6~ the talo-ealcaneus at the 16-day stage (t~igs. 11, l i e ) . The fusions between other ,eI~,me~gs are less regular', t~'or instance, c~meiforme S and the ' ~omposit.e' are separate in i~ig.. I1(~, superfeially fused in ]~ig. t~e and intimately joined ~ogether in Fig. 11 D. In at[ :~hree cases, navicnlare and the ~ composite' are still separate, b~t will coalesce at a later
s~ge of gestation.
b
\ ~ ram. t _ _ J
I~ig. 1:~, OIigosynda~tylism- Lef~ hi~dfoo~ ~nd forefoo~ of ~ 16-day-old embryo. Nfethylene-bhe preparation.
�9 in the carpus, some of the elements are so smaU and close together that, in methylene- .15!us preparations, it is difficult to decide whether all entities which are later ioined ~0gether have originally been presen~ separately; it is hoped the6 sectioned material to be l?ressnted later will be more eri~iea.J. In 6he meantime, it cars be seen in 16-day-old ~mbryos (Fig. 1t5) that fusions in ~he carpus are about as far advanced as they will ever ii~',.there being a fusion between maltangul.um reinas, eengrale, eapitatum and h~matnm, aiid simile.fly between scapholun~%um, triqnetrum and pisff'orme. The fusion between in~ga~arpalja & and 5 is always present at this stage and sometimes a lib~le earlier; ~ these elements are quits sepm'age from each other, as are the metatarsalia 4
X~ ~ in the hindfest. Secondary fctsions in Os/+ embryos are indeed by no means confined to carpus and
~ars~ts. 91.us is seen on comparing ~he relative frequency of soft-tissue and hard-tissue ~sYridaetylism in. embryos and adults (Table 10, in which all oligodaetylous feet have been ~ Botb in the forefee;~ and in the hindfeet, many more d~gits are fused, by bone in !h@. adult ghana, are %sod by ~a,~tilage in 15-.16-day old. embryos.. The diffe~:ence is highly Signitican~ in each case (X~.)=7"6~3 and X~1)=1876 respectively). Clearly, some of the ftx,sions observed later in life nmst have taken place after the 15-16-d~y s~age; i,e. they :faUst be secondary fcl.sions; indeed, it is clea~' that there axe more secondary fusions bh~n ~timary composites- Several su~h c~ses ]aa,~e been recorded as ' ? abou~ !;o fuse' in Ca/+,
Genoa. S4

130 Ge~,eticcg studies o.~z ~t~e s/cege~os of a/~,e ~,oz~se
but not in s~Ws~r~ embryos, before the exisbence of secondary phMangeM fusions was suspected. A case in progress involving metacarpalia 2 and 3 is shown in Fig. l'~b; one. cannot, of com'se, be certain how cOral?lute this particular fusion would have become l~ter on.
Whereas all, or nearly all, cases of osseous %sion in sm/s"o'~ are thus primary, bids applies to some only of the fusions of Os/+ mice. Rather more than one-half of those uncompleted in adults are secondary fusions between elements laid down as separate entities, tn the tarsus, the fusion between cuboideam and cundforme 3 which resnlts in the ~'compesite ~ is primary, all the others are secondary.
Table 10. bzcide~zes of s@4isst~e sy~dacty~is~n ~'~d of ha.rd-t.iss~e (cc~'rtigc~ge c~'~d bo'~�9 fl~sion i'~ 15-] 6 day-old Os/ + enzbr:qos c~,~d Os/ + cd,~lts
~oretegs Hindlegs
"Embryos Adults Igmbryos Adubs
Soft-tissue syndactyl ism 116 3~ 88 19 Kard-t,issue syndacgylism 19 16 38 26
']?otal 135 50 lOG 48
Fig. 18 illustrates a mode of d~velopment of oggodactylism in emb~'yos width i8 different from that deduced from a&dt material. The four abnormal limbs b-s can be regarded as steps in this process. In b, digits 2 and 3 are of about equal calibre, tn c, digit 2 is much thimier than digit 3, but still r6,presented both by a metatarsMe and the basal phManx.* in d, digit 2 is reduced to a mere rudiment of its metatarsale, and in s: there is no truce of digit 2 at all. In ~his a~d other cases, oligodactylism is brought aboa~ not by afusion between digits 2 gad 3 (which are of roughly equal calibre, as in Figs. 8s and 9&), but by a gradual reduction of digit 2 nntil it vanishes altogether. It is not certa}]z what happens to the rudiments of digit 2 as seen in Fig. 13c and d. In a 16-day-d~ embryo (Fig. 1~) , a roughly triangular piece of cartilage which slightly proje0ts between cuneiforme 1 and metatars~le I is obviously a rudiment of metats,rsMe 2. Ne, traces recog~izabIe as rudiments of tJaa~ element have been observed in adult sl~ecime~, Tl~is may be due to an accident of samJing. Perhaps some of the thinner rudiments' lil}~ that in Fig. lad are either :absorbed or secondarily fuse with their bigger ndghboux anS. are later no longer identifiable.
�9 I aminclined ~o suggest that there are three ways in which oligodactylism can be broug]i{. aboutin O~/+ ns these form a continuous sequence. I t looks as if the material for dial{ 2.1 prior to chondrificgtios, is either shifted in its e~tirety towards digit 8 ; if so, the two may- :fuse as about equal partners and so form a composite digit, Or alternatively, {he mate~N {'or digit 2 may be shifted in its entirety Cowards digit 1 ; if so, the material is too much fQ~ a single tiallux, and the remdt is what we have eMled oligodactylism with polydactylissi~, But if the material for digit 2 is .dispersed and partly goes one way and partly the otifer; remnants faay be left behind b. the middle fl'om which a rudimentary digit 2 may g %rmed. Whatever the explanation in any particular instance, tt is clear that the clitorises v l i sm in Os/+ nice involves a digit in the middle (digit 2) and not a digit f,_.om ~I!a margi= of the foot plate, as in Hertwig's oligodaetytism (Freye, 1954).
" A general eendeney :for digi~ 2 ~o be reduced as compared with digit ;3 is quite common in O,s[ ~" emb<Y.9 s~ see, for htsl~ance, Pig. lab.

H ~ x s ga, f : > ~ o 13I
ffe~aetic*. This recessive gent (syn,bol s?/) was obtained by IKertwig (19t-2) in X~ray irradiation experiments. The father of the f_rst known heterozygo%e had been treated ~ith m~ X-ray dose of I000 r. ; so ~he mntagion was probably in.ch~od ra~her %hart spon- ~aneous. As sy/sy mice never live to breed, all segregation data are derived from Fg~type raa~ings; the first few litters obtained by tIer~wig gave a ratio of 2? : 7. Larger data wiU. be:presmzted below. A number of +/sy heterozygotes were kindly handed over fie me by Fret. Yaula Hertwig in Nay i946. The original stock was later outcrossed to a derivative 0f~he GF~' strain which wa~ not highly inbred, and at a still tabor date, an ontcros~ of that, ~toek was made to the @BA/@r inbred strain. The data are snmmarized in Table ] 1. In all tIiree groups of rantings there is a significant deficiency of sy/sy a~fimals which is probably due to sdeetive early post-natal elimination as shown by segregation data among embryos.
Table I1. ,%9regat,ion of dufi~er with ,y~clactygsm as dassified c~t birah or soa~ @or. Mat iws uJith 20 young or over
No. o f r a n t i n g s + ~y T o t a l Perc~mi,~ge sy
Orig inM s t u c k 8 220 58 278 9,0.9 Ou~zroa~ ~o GFF 28 i279 $q~6 162~5 21'3 Ca%cross to CBA/Gr 16 705 118 823 1~.3 To~l 52 Z204. 522 2726 18.0
Table 12 Seq,,'ega~.i6oa. of .~haher with syndaz@is,m.. Litter's of e~bryos L i t t e r s wi~& ,~y
g
L{~ter size No . of l i t t e r s 1 2 3 4 + sy To~M ~y e x p e c t e d 6 1 7 2 ,0 t07 7 i I -- --
S 6 1 - - 4 1 31 17 48 13.3344 9 5 2. 2 - - I 35 10 45 12,1905
10 2 ---- i ~ -- 1.5 5 20 &2.980 ii ;2 . . . . . 2 14 8 22 .5,7420
T o t a l 16 . . . . 101 41 142 38-,5816
The. percentage of sy/sy was similar in the original stock and Jbllowing the outm'oss GFF: but it fell considerably after the outeross to OBA (X z : 17.08 ; n = 2; P - 0"0002 approx.). Wla% many ~y/sy alfimals li~od for ~ or 4 weeks (and sometin-ms considerably longer) in ifie original s to& and aGer the enforces to OF]Y, the gene behaved almost a.% a lethal at bit@ following th.e oatoross to CBA, nearly all ani~als beh:~g either stillborn (a, nd hence often mopped np before being recorded) or dying within 24 hr. or so after birth. As will
be described below, differences in anatomieM strnetm'e were also associated with $hese dhangea in genetic background. Unfm'tuna%ely, it was not reMizod soon enough ~hat, t, he opportunity for the study of post-natal development wmdd be so limited. As a conse- q~ence, %he 1lumber of alizarin clearanoes asd papMn preparations secured was very smM1, and most of the following description ,rill have be be based os embryos and new~ Born young.
Segregations in ligtecs of embryos aged 13-15 days are summarized iu Table 1Z. allowing fbr the selection in these da.ta (~o lit)or being included nnless it contains a,t least One ~9/sy embryo), the number of st~st embryos found is in excellent agreement wfl~h tha~; expected for a ~ : 1 ratio (4t and 38-6 respectively).
9-2

132 aeneliccd studies on the a]~etegon of ~he mouse
A'nc~tomy c~nd deve~ofo'menL In this mutant, abnormalities are found not only in the feet, but also in many other parts of th~ skeleton. We shah here first discuss the foot ~nomalies which clearly form a t~nit; the remainder of the syndrome will be described afterwards; it may no~ be very closely connected with what happens in the feet at a~r early stage in development.
The gene sy aft'cots forefeet and hindfeet; however, unlike sin/sin and Os/+, the forefee~ are oi~en completely normal, alad even in the hindfee~, which are more strongly affected; normal overlapping is not very rare. Out of 29 sy/sy mice of the original stock, I3 were affected on all fbur feet, 6 on three feet, 9 on two feet, and 1 on one fbot onl.y; hence a~ occasional sy/~y mouse with completely normal feet may be expected. The involvement: of the various digits is shown in Table 13. As in s@s'm and in Os/+, ~he three middle digits are involved (neither digit 1 nor digit 5 being ever incNded in syndaotylism),
Table 13. I~tvo~ve~nent of the various digits in the o~'iyi~al stock, of shak,er with syndas@isrn. .Living animals
Table 14.
Digits hivolved Forefee~ Hindfeet Tots[
aN'one "26 1 27 2 gnd a 23 42 65 3 ~nd 4 6 3 9 2, 3 a.nd 4 3 12 15 ~2og~l 58 58 I16
s w~th 3yndac@ism. Invdve~nent of the ~arious digits (eargigaginous fudom' o~tg). ~]f ethylene-bhte pref~arc~tiona
Digits involved Forefee t i Hindfeeg 's None 51 21 72 2 ~ncl 3 0 14 14 3 and 4= 9 9.4 8a 2, 3 and[ i 0 1 1 TotM ~0 60 120
5~ezvborn c~nimals fi'o~ gfs outcross (ezcept 5 from CBA outoross),
However, the situation differs strikingly from ~,hat in sm/~m (in which there is either involvement of digits 3 and 4, or of ~, 3 and 4, but never of 2 and 3 alone) and from that in Ca/-:-(in which there is usually involvement of digits 2 ancl 3, and ra rdy of 2, 3 and 4, but never of 3 and 4 alone). While sin~sin and Os/+ thus each have one single sharply defined primary type of syndaetylism (3 + 4 and 2 + 3 respectively), sy/sy has the choio~ between the two, doing either one thing or the other. In the original stock, fusion between digits 2 and 3 was much commoner ~ha~ that between digits 3 and 4. t:fowever, this' depends very much on the genegical background in which the gone finds itself. This is: shown by the data of Table I4 which is base& on methylene&lue preparations of newborn sy/~y young derived from the G!~I~ outcross, with the exception of five from ~ke CBA out- cross. As in this kind of preparation, soft-tissue syndactylism of newborn ~nimds is not always classifiable with confidence, oartiiaginoas fusions between phalanges only have been tabulated; the data are thus not directly comparable with those of Table 13 which includes both types of syndaetylism. However, it is obvimls that on this genetic back ~ ground, synd~ctylism between digits 3 ~nd 4 is much commoner than syndac@ism between digits 2 and 3.
As in sin/am ~nd in Os/*, abner.real sy/sy embryos can be disting~dshed from their normM litter-mates at the age of 12 days. The stages which san be followed in methylene-

133
blue preparations are fairly similar to ~hose in the other mutants. Figs. 15-1.7 tb.as requ r :e ii~tle explanation. Bard-tissue fusions are confined to the region of the phalanges, ~.~s.in. s,m/sm, the metaoarpalJa and metatarsalia being never involved. Nest commonly, ~Jl ~hree phalanges are %sed, or the middle and the terminal phalanx only. Fusion of lj~al (and middle) phalanges t'oJJowed by separate terminal phalanges so common in ,Os/+ and not very.rare in sm/,~m is very exceptional in sy/sg. Carhlaginous %sions between l~Mlanges are present from the very beginning, and there is no evidence for the occurrence 6{seeondary fusions in these elements. All earpalia and tarsali~ are laid down as separate
b 1ram.
~ ,,~ ~:.:,7.:..
Fig. 16. Shaker with syndaotyli~m, l~ight hindfee~ os 13~-day-old embryo~, a, norms!, b-d, sy#g (litter-mates). Methylene-blue preparations. ~ote. The low-gw~de anomaly in b is confirmed by the fact ~hat the same embryo has a high-grade anomaly on one of its forefeet.
roll%,
b
I
C d
Fig. 16. Shaker wit.h syndaotytism. Right hindibet of 14~-de,[} -old embryas (or perhaps a little cider} c4 norm~d; /~-d~ <q/,~/, a-c are litter m~tes; d • from another litter. ~:[ethylel~bllle ~)reparatiofl,%

134 Genetical studies on, the slceleton of the mouze
C b
1 rnm d r
Fig. 17. Shaker wigh syndaegylism. I:[indfeeg of 15.}--][6-day-old erabryes, a, normM; b-f, ay/~g, e and d are fiwo hindfee~ of one embryo ~vhieh is a ligger-m~te ~o a ,~nd b; e and f from ~ ~lifferen~ 12~ter. ~le~hy~ene- blue l~re;para~ions.
elements, t{owever, there are secondary fusions both in carpus and tarsus. Examples are shown in Fig. 1.8. Five alizarin preparations only are available for examinalion, fern: of these fl'om the original stock, one from the CBA outcrosa, i~usions are present ie every carpus and. every tarsus, as follows:
~'eet
eentra le~capi~atum--ham~um 7 eapi~a~um--hama~um 3
navietd~re--ouboidm~m~cue.efforme 3--cuneiforms 2 7 euboideura--eunei~orme 3--etmeiforme 2 3
The nonLflmctional artic~datiot~ between navicalare and cuneiforms 2 tends to persis~ (Fig. I8b), as in 0~/+ mice. Al~taough no normal litter-mates to ~]~.e above animals are available .for comparison, the fusions observed are so extensive that l,hey can ha~dly be drte to the genetic background as such; they are probably effects of the sy gone itself.~
* N~e added 28 June [955. Since ~he ~bove was written, fo~r ~ddition~ a,~iz,~rin prepar~f2ona of I5-18-d~y~ old sy/sy mice have become available for study. These show Nsions similar to ~hose described a.bove on a[! six~een fee~. As in t, he new anim~ls ~he ~y gone is on aa entirely different gene~ie background, ~here camno~ be. a.ny dmzb~ ~ha.~ the earp~l and t~rsa.1 f~xsiona are due ~o ~he -sy gone it.~elf.
Hs>-s G~iPN.~B J:gG. 135
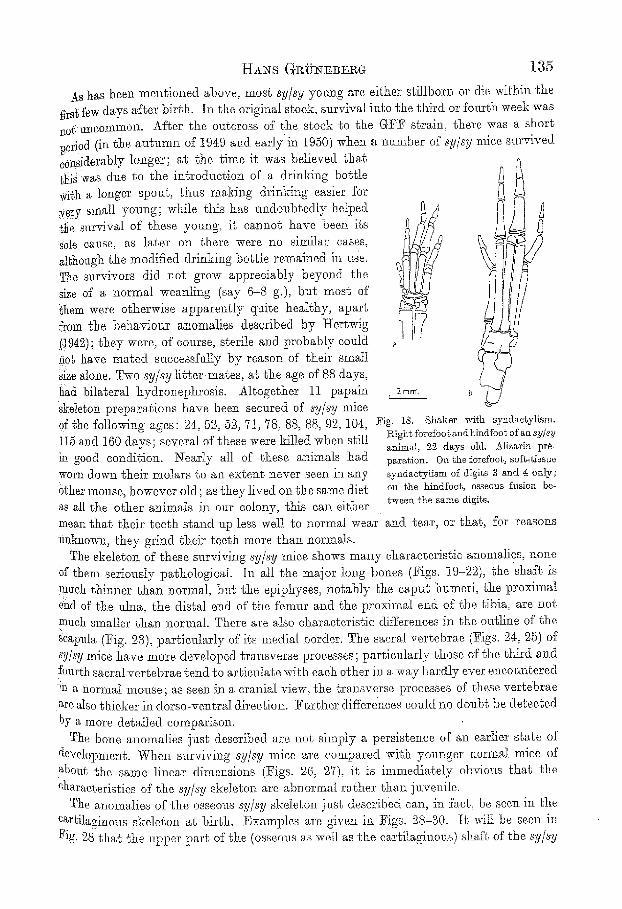
As has been mentioned above, most sy/sy young are either stillborn or die within the .f/rs~ few days ~fter birth. In the original stock, survival into ~he third or fourth week was ::&, tmeommon. After the ougeross of the stock {o the G1?17 strain, there was a short period (in the autumn of 19~9 aild early in I950) when a number of sy/sy mice survived sonsiderably longer; at the time it was believed that {NS w~s dne to the in~roduction of a drinking bottle with a longer spou% thus ranking drinki::g easier for *e~Y small yonng; while this has m:doubtedly helpecl ~tte survival of these young, it cannot have been its .ble cause, as later on there were no similar cases, aN:ough t,he modified drinking bottle remained in use. The survivors did not grow appreciably beyond the siZe of a normal weaning (say 6 8 g.), but most of' ~hem were otherwise apparently quite healthy, apart from the behaviom" anomalies described, by I-Iertwig {!942); they were, of oot:rse, sterile a::d probably couid i~0b have mated suecessf:tIty by reason of t}leir small :SiZe alone. Two sy/sy litte>mates, at the age of 88 days, llad bilateral hydronephrosis. Altogether 11 papain Skeleton preparations have been secured of sy/sy mice Of the following ages" 24:, 55, 52, 71, 78, 88, 88, 9'?,, 104, 1I~ mid 160 days; several of these were killed when still !: good condition. Nearly all of these animals had worn down. their molars to an extent never seen in any 6thor mouse, however old; as they lived Ola the same diet as all the other animals in our colony, this ca:: either
a
a t o m ~ b
Fig. 18. Shaker with syndae~ylism. Right forefoogandt:indfoot of an ~y/sy animal, 2.2 days old. Aliza.rin pre- paration. On the forefoot, softdissue s3mdaotykisrn of digits 3 and g only; on ~Jae hLuds osseous fusion be- tween: the same digits.
mean that their teeth stand up less we].l to normal wear and tear, or that, for reasons mlki:ov::, they grind their teeth more than normals.
The skeleton of these surviving sy/sg mice shows mai:y oharacterisi, io anomalies, none Of then: serionsly pattmlogical. In all the major long bones (Figs. 19-22), the shaft is much thinner than normal, bu~; the epiphyses, notably the caput humeri, ~he ]?roximM ~nd of the ulna, the distal, end of the femur and ~he prommM end of the tibia, are not much smaller than normal. There are also characteristic d_ifferencBs in the outEne of the Scapula (Fig. 23), particularly of its medial })order. The sacral vertebrae (Figs. 24, %) of *s*..I/sy trice have more developed transverse processes ; particularly ~hose of the third a:ld. fottrth sacral vertebrae tend to arUienlate with each other in a way harNy ever encountered ~;. m a normM ram:st; as seen in a cranial view, the transverse processes of these vertebrae are also thicker in dorso-ventra] direction:. Farther differences could no doubt be detected :by a more detailed comparison.
The bone anonialies just described are not simply a persistence of an earlier state of developm.ent. When survivilag s?//sy mice are compared with younger normal mice of about ~he saree linear dimensions (Figs. 26, 27), ie. is immediately obvious that the eharacte:ristics of the sy/sg skeleton are a]ol?ormal rather that], iavenile.
The anomalies of the osseous sy/sy skelet, oii just dese:r:ibed can, in J!act, be seen in the cartilaginous skeleton a~ bir~h. Examples are given in Figs. 28-80. Ie will be seen in 1Pig. 28 that the upper part of the (osseous as well as the cartila.ginou.s) shaft of the sy/sy

136 Genetica~ studies on the d~deton of the mouse
tibia is ~him~er, bu~ ~hat the ~ra~sverse diameter of the ca~?ztL tibiae .is abot~t:l~or~jal. Similarly, ~he charac~ex'istio flat~ening of the anterior part of bhe media.t border of ~[ie scapula (Fig. 23 b) is detectable in the ea~:til~ginous s~l;e at birth (Fig. 29 b). And ~he characteristic ~ransverse processes of ~he sacral ver~ebrae of sy/sy survivors are also pre- formed in cartilage (Fig, 30 b), As usual, i~ is ~hus possible to exonerate the osseous skeleton
i
b a*
<
1
t
. . ff l~
:Fig, 19. :Fig, 20.
:Fig, 19. Shaker with syndac~ylism. I~ight humerus of a sy/sy O0 {b, b') a~d of a normal brother, ][04 days old. Papain preparations.
Fig, 20. Shaker with synd~c~ylism. ]o~na of the same animals as in :Fig. 19. Papain preparations.
a a" b e
I - . 3
?
Fig. 21, :Pig, 22.
Fig, 21. 8hakor with syndaotylism. Lei% f e m ~ of ~he aame <~nim~ls as ia :Figs. t9 and 20. Papa,in prepai, ations,
:Fig. 22. Shaker with syndao~ytisra. I~ighb ~ibi~ ~md fibula. Same animals as in Figs. 19-21. P~pain preparations.

HANS GI~NEB:ERG 137
Fig. 23.
) Fig. 24.
b
2mm.
Fig; 23. Shaker with syndactyHsm. Eight scapula of the same animals as in Figs. 19-22. Papain preparations.
~! ' 2~. Shaker wi~sh syndactytism. Dorsal view of the s~cral vertebrae of the same animals as in Figs. I9-23. P~pain preparations.
~2mm. d'~ 3
m m 8 ' " E ,
Fig. 26.
(
b'
Fig. 25. Fig, '27.
~[g. 25. ~haker wi~h syndae~ylism, Cranial views )f the sacral vertebrae sllown in Fig, 24. Papa.in :[3rel)~ra~ions, a~, normal; a~-d ", ~y/sy.
I% 26. Shaker wi~h syndactylism. :[~igh~ h~,merus 0zR.houb its :proximal epi}2hysi,~ ) of ~ sS.sy ~, 78 d~ys old (5, b') ,~nd oi' ~ 35-day-old normal n'muao from '~he CBA/Gr ,~train (a, a'}. Papa.in t)reparM.ion,~.
~g. 27. Shaker with syndao~y?dsm. Lsf~ femur (without its dis~a} epiphysis) of an sy/sy ~, 78 days old (b, 5"}, and of a 35-day-old normal mouse ;[rom the CBA/C4r s~rain (a, a'}; s~rae animals ~s in Fig. 26. Papain prcpara~ion.~.
138 Genetica~ studies on the skeleton of the m ~ s e
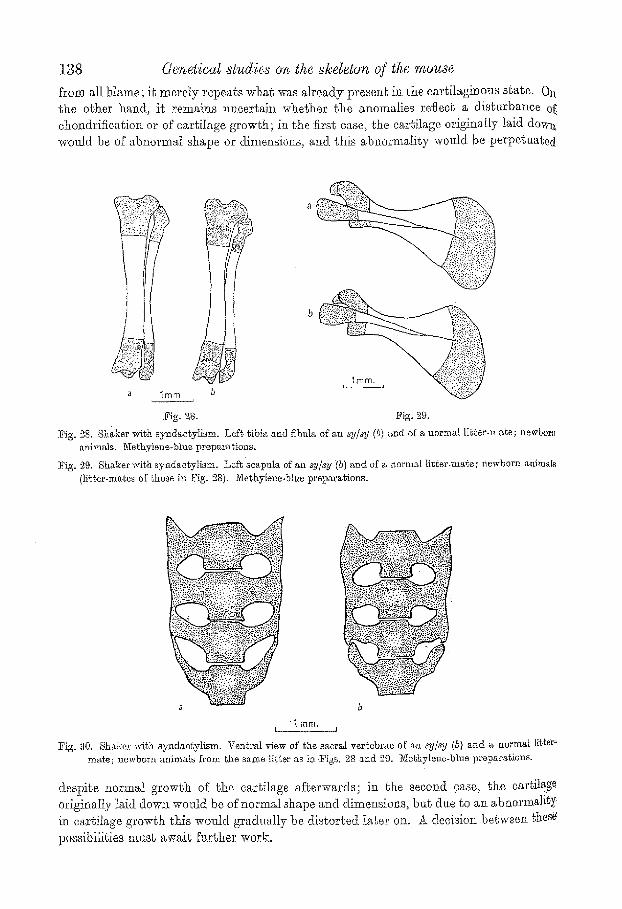
from all blame; it merely repeats what was alreacly presea~ in the cartilaginous s~ate. 0tt the other hand, it remai~s ~meertai~t whether the anomalies reflecb a disturbance of ehondrifieation or of ear,liege growth; in the firs1; ease, the cartilage origina,ll.y laid dowa weald be of abnormal shape or dimensions, arid. ~his abnormality would be perpGttatedi
a
a 1ram. h
l~ig. 28. ~ig. 29,
t?ig. 28. Sha.ker with sy~dactylism. Lef~ ~ibia and ~bnla of an sy/xy (b) and of a normal li~ger-ma~e; newbor~ animals. Ne~hyl~ne-blue preparations.
]rig. ~,9. Shaker wi~h syndaegylism. Lefbscapula ef an sy#y '(b) and of a norma[ Hg~er-'mafie; newborn animals {li~t.er-ma.~es of those in Fig. 28). Methylene-blue preparations.
., j , ,::r : : t'L:
b
"1ram. L 1
Fig. 30, Shaker with syndae~ylism. Ventral view of the sacral vertebrae of ~ zy#y (5} and a normal Iitter~ mat, e; newborn animals from the same ].i[ter a~ in Pigs. 28 a, nct 29, 3'Iel~hyI~ne,blue prep~'~raf2on~.
despite normal growth of the cartilage afLerwards; in the second 9ase, the cartilage originaIly laid dow~ wend be of normal shape and dimensions, ba~ due go an abnormality i:a car~ilage growg~h this wonld gradually be distorted later oJ.a. ~ decision betwee~ ~hese po,ssibili~ies nmsb ~w~ig Nrther work.

HA~s G ~ t ~ s m ~ r 139
CARPAL ~bID TAP~S_4_ZL FUSIONS IN INBI~EI) STi~XINS
~,~aoeem'rence of carpal and ~arsal fusions in normal litter-mates of sm/~m and Os/+ mice ~::been mentioned above. Similar variants have also been fmmd in inbred sSrains of ~co;; :As the inbred strains differ widely from each ot]~er in this respect, ~bese %siena N~is~(eiearly be under genetic control, at leas~, go some extent. In the carpus (Table 15), ~ie~:between multangulam minns and centrde ia Vpical of both 057BL/Gr* ~md of ~ B / c , bug has not been encountered at all in small samples of CBA/Gr and A/Gr. In :~ii;j~ generation involving k./Gr and BALB/c-~, ~here were 16 animals with this type of Nsion and 8 withoat; this is no~, far from a 3 : t ratio, bug in view of all oar other ex- Cel{chess of interstrain differences, t would be greatly surprises if things were ultimately i0itnd to be so simpte, k second kind of fusion involving the capgsatum and een~rMe ~r in all { animals of a particular CBA/Gr litter, bm, not in S animals from Shree gher litters (two ramjets) of the same strain While this might conceivably indicate a gea~lfic differenl,iadon within She strain, I am more inclined to interpret it as a case of <hmehing' within a litter, i.e. as a case of intra-litter correlation due to the r pre- ~igal or post-harM enviromnenL A similar ease for which this is pro.:bable will be men~
~Jm:ed presently.
~a.ble 15, Incidence of ca~pM fl~sions in fou~" i**.bred st~ain~, a~ F~ gener~,tio~ be~wee~. inb~'cd strains, a~zd i~. a~o mixed stoek~
5'iult~ngulum minus and eenl .rMe Capitat.a'm and cenh'Me
50. ~o f,s~o~ ~,. ~. :~. +Zs ~,. L. :< +L. ~ot~:. 1 A/Or 1 ~ - . . . . 12 2 CB_g/Or 8 - - 2 1 1 i2 3 C57BL/Gr -- __ ] 7 - - r - - 17" �9 f BALB/e -- 1 -- 35 . . . . 36 5 (A/Gr x BAL;g/c) F: 8 1 ~ $ . . . . 24 6 "F from sm stock 17 - - 2 :2 . . . . 21 7 + from Os stock 13 1 3 - - 3 . . . . . 20
* This includes one litter of 5 from buckwheat 'feeding experiments.
The tarsal fl1.sio].I8 encountered in the same material were, with one exception, all of .~h~ s~me Mnd, i.e. fus{ons between the navioul~re and the eunMforme 3 (T~ble ],6). This ,}[pe of fusion was almost universal ~n r 57 BL/Gr,:I but absent from 0BA/Gr and BALB/o. :rhvo of the three affected animals in A/Gr represent, the second litter of a pair of mice ~hose first litter (four animats) showed no %siena; {his suggests a real difference be{ween %Ire two litters ra{b.er than an accident of sampling ( P = 0-067 by Fisher's exact, method);
* The s'~me observation has been made independently by Dr ]?au/]~'. ForsghoeFol (Dotroi% g{iehigan, U,S.A.) his sabstrMtl of O 57 ZBL/I.O which also, like our own C 57 BL/Gr, has fusions between nav~eularo and euneiForme 3
m.t.he tarsus. The genetieal basis for these auomMios must have been fixed before the American and BrYtisb m~bti~ea wore ~eparated from each other and seems to have remMued stable ~ e r since. The ]3finish ~ubsl.rain was brough'~ to this eoanbry by J, ]3. S. }IMdane in the autumn of I932, and ~.he l~st common ancestors of g57BL/10 and our own subsh'Mn may have Iived a few years further back. t am grateffd to Dr Forsthoefel
"~br NloWing me to refer s hi~ unpublished data. "~ /~x animals have, unFor tama~eIy, nod been examined; a.s so often, the preparations had been made for qtdte
different purposes. In Ogs particular ease, the 13ALB/e ~toek segregated for the gene tbr tMl-ki~ks (tk) which ~ad ~ust. ari,~e.n i.u its, and t~he same gene Nee segregated in ~,lxe ou~.m'oss I~L; as l, he invidenee of fusions ~ppeara
8b be I, he same in normal and tkflk mice, ~hese two classes have been pooled. Dz Paal F, Fors thoefel informs me that in old C 57 BL/I.0 mice, the znneiForme 2 is Mac otdten involved, which
eviden~,|y indieat.es a ,~econd~ry fusion in t.he o~soous M,a,~e. No such case ~a.~ found in S otd Cd~_BL/Gr mice.

t 4 0 GeneticaZ sa~dies on ~he J~demn of the ~nouse
if so, there is b~tra-litter correlation due to the common environment; in any ease, bh~: occurrence of animals with and without fusions in the same sibship of an inbrecl sgrM~. indicates that, within ghe strain, ~he difference is Fcobably environmental ra~her flIat~ gene~ie.
Fusions between carpals and/or ~arsals as manife,~tations of major skeletal genes have: been reported by several a~tthors (Hovelaeqtle, 1920, in his 'souris lax~es'; Bean, 19291 in 'rnyeleneephalic' blobs; Carter, 1351, in henfimelia tibiae; Orh.neberg, 1953, in coti:,. genital hydrocephalus; Freye, 195t, in olJgoclactylism; Green, 1955, in luxoid; Trus[ov%. 1956, in fidget; and ~his paper). On the other hand, similar f~sions in otherwise normal! mice seem until recently ~o b.sve escaped notice.* Apar~ from the observations o{:
Table 16. I~vidence of t~fad fl~sio~vs in jbu.r inb~'ed st~'Mns, c~9~ Y~ 9e.;~e'ra, tion beaoeen .inb~'ed strains, and i~ two mixed stocfcs
N~vieulare a.nd ,'.une]forme B f
~o. No fusion 1%. L, P.. + L. Tot.M
1 &/Qr 9 1 ] 1 12 0BA/Gr 12 __ _ -- 12 O 57 BL/Qr -- t I 15 !~
~i IBALB/e 36" -- -- -- 36 5 (A/Or x BALB/o) F~ 23 - - --- 1 2,J, 6 + from s~g s~oek 16 I 3 1 21 7 + from Os s~ook 20 - - -- ~ 2.0
* One animal with an incomplete fusion between mmeiformia 2 ~md ~.
Dr Forsthoefel mentioned above, Truslove (1956) found in 95 otherwise normal (i.e. noni polydaetylons) indfeet of ormal litter-mates or dget. 6 ..sen of nsio navie~lare and cnneiforme 3, and I0 instances of fusion between cuneifor~-rda i and% The latter type of fl~sion has not been observed ~t all in the data presentBd in this pape~:i (except, of course, in mutant animals like Os/+ or sy/sy). I t is thus quite likeIy tha0 int~:. ,:, extensive studie~ may bring to light sdditiona~ Nsion pa~terns. Whereas in norms,[ mi~: fusions usually seem to involve two adjacent elements only, in some of the mutants (sa~. as Os/+ and sy/sy) the anomalies tend to be much more extensive.
Similar %sions between carpals and t~rsMs have been described in man. For instane.~) Testut (1911) remarks 'C'esg ainsi que l'on rencontre parfois ia soudure du ca!ean~il~ aveo le scapho~de, de' l'astragMe aveo le calean~t~m, des trois oungJformes entre et{x0n aveo le scapho~de, etc.', and Bryce (19 I5) states that " the tarsal bones may be fused. Wi)!i their ~eigJaboltrs: thns the talus may be trailed to the ealcanens; the tshts to the n~@# ealar; the eMcaneus to the navioular; ~he navicular ~o the cub oid; the ealeaneas Lo ~l~e ouboid; the second to the.third cuneiform. The metatarsals Ii and III may be unit'S&' with l,heir respective cuneiform bones, and the metatarsal I and I t with one anoth&::~ their bases'. Fusions in the carpus seem to be much rarer in man, but Bryce meuti0~ that ' e.g. the kmate and trique/~ral bones are occasionally seen. coalesced'.
* Bean (I929) mentions that some ether~ise norn:M feet showed wrious Mnds of ft~sions. Unforbtm~@!Y~ ~.his auhhor has fMled go idengif'~ by n~me ~he elemeng~ she is writing ~boub, and t,b e supposedly normal foreleg!" and hlnd_feeg described by her differ so sgrH~mgly h'orn ~he norm~t gn~gomy of the mouse tha t t.he authot~ mtJ~ have investigated very peculiar mJ.ee, or else her technique .mus~ he.re been s~dly inadeq~a,l,e.

t tANS GMJ~xge t 41
])~:SCUSSION
$li~:fhree mutants described in ~his paper can be disting~dshed from their normal li~ter- Sa~es:it the 12-day stage of embryonic life by the shape of their limb buds. The limb ~Ms6f the mouse do not yet contain any cartilage at that stage. In %hess mutants the ~li~e~.of hands and feet is thus not determined by the a.ngtomy of the skeleton, bu t the ~a~omY of the skeleton is determined by %he shape of hangs and feet. The situation is ~liss i~ke that in polydactylism (Ohang, 1939), in hemim.elia tibiae (luxate, Carter, 1954) gd:ia oligodac~ylism (Freye, 195~), fl'om which similar conclusions have been drawn, oz' iouid:have bee~ drawn. The next step in the analysis will be ~o discover the causes of the :~@a: anomalies in the limb buds, and, to connect the e~rly effects with the later ones tep:~t~ed here. This, i~ is hoped, will be done i.u a l~tex .paper.
.~:e pathological situs%ion in is.tel stages of development reported here consists mainly SffliSions between adjacent elements of the skeleton. Of these, three different types can ~~(~stinguished thongh there are pzobably intergrades between them.
([) Pris'~a~'y f~sions. These happen in the mesenchyme stage and give rise to a single ~ r e of ohondrification where normalXy there are two. I t is Pr9bable that this in fact .~Chdes two different mechanisms (formation of a single condensation of mesenchyme :i~s~ead of two; and coalescence of two such oentres prior to chondrification). !~xamples ~eghe fused phalanges in s~r@m, sy/sy and, partly, in Os/+ ; and tJae ~ composite' eorre- sppnding to cuboidenm and cuneiforms 3 in Or /+ . }i2) Secondary fusions. These happen in. the ea.rtilage stage and result in the eon-
cr~scence of two cartilaginous elements of separate origin. E.xamples are ~he formation 0~.the talo-caleane~s, and the coalescence of other tarsals and carpals in 0~/+ ; ~he fusion [begween metacarpMia 4~ and 5 and meta~arsalia ~ and 5 in the same mutant, and many
~liers {i) Territory fi~sions. These happen by the coalescence of neighbouring bones either
a~ross the periosteum, or across a fully established artieNar st~rfaee. An example 0f the b~mer is probably the occasional fusion between me~a~arsalia found in some old s~/sm ig~e, while the latter mechanism is probably represented by the fusion of~eaneif'orme 2 ~'ff'S neighbours which oscars in many old O5qBL/10 mice, but is not yet present in giihnals 3-6 weeks old.
These three groups, though asefal for a first classification, are probably not sharply disei~ct For k~stance, the coalescence between two condensations of mesenehyme can hgppen jus t about the t ime of chondriflcation; so She dual origin, of a structm'e may be ~igibh :for a fleeting moxnent in. cartilage ~hongh very soon MteJ.'wards its composite ~a~Ure is no ].onger de~eotable. This is probably 1;h.e case for the naviculare and cuneiibrme 2 l~::congenital hydrocephalus (Grtflteberg, 1953). Siksdla~:ly, a tertiary fusion may l)e �9 [~i/itated if an articulation bet:ween: say, two adjacent tarsals is not quite complete, ])n~ -~ainewhere traversed by a ca.rtitaginm~s bridge.
M1 three types of fusion can octm: in norntal development. They are thus not in t}leln- [email protected] pathological though they become so by happening in the wrong places. Primary b.siens, in the case of the mouse, musS; be assumed Jbr tt~e primitive carpal i a 4. a,ud5 which, ~o judge from methyl.erie-blue preparations, are fl'om ~he first represented by a s~ngle. �9 ~ainatum ; and for the primkbive t~rsMia 4 and 5 which similarly give rise t,o a cu:boideam

t4=2 (i~en~tical studies on the a]celtton of the mouse
which appears as a sk~gle element from the start; the 'composite' present in Os/~ apparently differs from normal, development only by the fac~ tha~ the primitive tarsale a) instead of becoming an independent cuneiforme 3, is also included in. this primary fusion; A secondary fusion in normal :mouse development occurs in tibia and fibula which al-~ formed as widely separate cartilaginous e].emen.ts, but la~er fuse i.n theb distal thirds. I am not aware o[' a case of ter t iary fhsion which is a normal f'eatm-e for the mouse;: fusions between membrane bones (like tha~ betwem~ the two frmxtalia in :man) are possibly= somewhat different. More strictly comparable is the fusion between radius and ulna it~ Xenop~*s hLevis (D. R. Newth, personal comm.tmication) which happens a r e s these.. elements are ossified, and there are donLtless many other examples. The causes for these tMee types of fusion both in ontogeny and in phylogeny are not fu]Iy understood. It g hoped, tha t a more detailed sttldy of these phenomena whm'e they happen anomalously may help in understanding {hub causes in ~ormal development. The same, of com'sg;. applies to cases where normal fusions fail to happen in a mutan% as, for instance, in tM thyroid cartilage in congenital hydrocephM.as (G.riineberg, 1953).
Some of the fusions which are pathological in the mouse are normal features in th~ anatomy of other organisms. s instance, a fusion between t ab s and ealcaneus very si.milar to that found in Os/+ is a normal feature in Sphe.~wdor~, gara'n~s and other lacertilians. In the three-toed sloth B~'a@~s there is a %sion of tarsalia 1, 2 and 3 with the corresponding metatarsalia (Braus, 1906) such as happens for the first and more commonly for the second digit in Os/+. Other parallels will no donbt be obvious ~ readers better versed in comparative anatomy. The type of oligodaegy]ism which occ~s in some Os/+ mice, i.e. elimination of a ray in the middle, is quite ckifferent from the way in which digits have been lost in the course of phylogeny where this has generally started. either on the pre-axia] o} mi the post-axial border of the limb. The only exceptio~ seems: to occur in Cetaeea where, in Balaenopts~'us ~uscu~us, a reduction of the third finger. happens during embryonic life such that later on that ~uger is missing from the middle of the row (Xiikental, 1890; quoted from Brans, 1906).* Syndactylism Rself is, of course= a laormal feature in many wading birds and certain mammals, sash as ~he kangar0~. (Maeropus) where the second and third digits of the hindlimbs are much reduced aM
syndaetylous. Inherited ~yzadactylism as an anomaly within a species has been described in the.
domes{is fowl, in swine and i~ man. In the domestic fowl (Davenport, 1909), syndactyiig~ is by soft tissues only and usually coDtined to the interval between digits 8 and 4; this~:: as Davenport points out, is the intervaI filled m those wading birds in which two toes: oNy are connected by a web ; less commonly, the digits 2 a~d 3 were also webbed. Tl~e gene~ieal sR.uaion has not been completely anaIysed. In swine (Detlefsms & @armiehagli. 1921.), syndactylism has become a breed character in the ' runic-footed' hogs; {he gent.lot 'mule-foot ' is dolmnant over that for cloven feet; all four feet are affected, but unlike thei eonditimas described in this paper; the forefeet tend to show more complete syndactyliSm. than {he hnidfeet. As in sy/sy, the syndaetylism always starts from the tips of the toes but tends to become less complete in a proximal direction. The literature on syndactylism
* The reduction of the second digi~ on hands and feet of certain lori-l[kc lemurs (Nyc~ieebidae) such as:: .Perodic~isus 2o~o, Arc~scsbus scdabare'~uis and rela~ed species is much less complete, moreover, i~ happens as an adaptation t.o tm arboreal habit~t~ attd goes t.ogebher wRh an extreme oppositioa of pollex and halliix: respectively while in Os/+ the material of ~he foot plates tends towards consent.ration in ~he middte.

~.~ali,%aS recently been summarized by Julia Bell (1953), who distinguishes four major
~es~. as: follows : " :~y~ei:A,, in which webbing of the second and third toes occurs with commonly no
.~-~:o~yiy in the hands. "~ ~Tpe.:A.2, in which d~e third and fourth fingers are webbed; these cases are frequently .~s~ig~ed with reduplication o:f the fourth finger--or parts of the fourth finger--inchded
Ni~::web. ~.T~e:B 1 in which the fourth and fifth toes are webbed; these oases are not infrequendy
.N~0~i;~ted with a supernumerary little toe which is included in the web. T~e C refers to cases in which webbing of three or more digits in hands or feet, or in
a~l:.e~i:emities , has occurred.' :t~'actdition, there are a number of pedigrees of ~ mixed ~ypes '; usually, the members of
sTedigree, or most of them, conform to one of these types, but some genes have extremely "v~iabie effects, go far as I can see, aone of omr bh!"ee mouse mute,ribs is closely similar to ,i'~y:'0f the human types so far described. This is perhaps not ran.prising. Presumably -:~ff;re are many genes in either species which may lead to syndae~ylism., and the 'homo- :Iggcus' .ones may not have been observed yet. Moreover, 'homologous' genes may g0&ce different phenotypio effects on ~he vastly different genetic backgrounds of man
~cl::mouse. ,~ UMMAI%Y
1. The recessive gone for syndactylism (s.~) has redueec~ j a b i l i t y ~t birth and soon ~ft.er, bug nearly ~ormal viability and fertility later on. All four feet are regularly 'gffeefied. 8y~dactyli.~m invobes primarily digits 3 and 4~ while digit 2 is also not rarely :f~volved. Fusions are usually osseous in the hindfeet, but often by soft tissues only in the fo~feet. Phalanges only are involved, usually all three of them, wh_ile metacarpals and ra~atarsals are not affected. The ~mviculare and cuboideum tend to be fused. The fusions ~ety~een phalanges are primary, those between tarsalia secondary. Anomalies in the distal
~ilf.0f the tail are present in many s,m,/s~ mice. " 9.... The semi~dominant gone for Oligos?mdaotylism (O~) is lethal in hom ozygous oondi~ ~ioni:.but of no~:~a.l viability and fertility in heterozygotes. All rotor %et are regularly affe~%d. Syndactylism is usually confined i,o digits 2 and 3, but digit l is occasionally .~1~o involved. Soft-tissue fusions are more common o~., the forefeet, hard-tissue fusions on the hind feet. Fusions seem ~o sta.rt at the basM ph.al~m~ and thence spread first ~t~f~Ily; after all phalanges are :Nsed, the process spreads in a proximal dbection iu- voMng metacar]?Ms and metatarsals. Fusion may be so complete as to lead to a four- toecI foot in which there is e~l~.er a eompoNte digit representing rays 2 and $, or in which di~t 2 has completely disappeared; sometimes the material for digit 2 seems go be shif'bed Cowards digit 1 which may tl~eaa be duplicated. There are always ex~.ensive Nsions in carpus and tarsus; these inc]:ade a primary Nsion between euboideum and cuueiforme .3, all others being seconda?:y. Fusions between some phalanges are also primary, but tb.e majority of these and between metacarpals and metatarsals are secondary; these include t?ar~ieklarly fusions between the proximal cads of metaearpalia and metatarsalia 4 and 5 5~hieh are secondary to the reductions in earl?us and ta~rsus respectively. ~3. The recessive gone for elks.her with syndactylism (sy) is semi-lethal, in that really
:aaimals die soon after bir~].~, sad none reach sexna.t maturity. Most hind:feet are s flee.ted, ht~t normal overlapping in the forefeet is common. Anoma.liea involve either digbs 2

144 Gene t i ca l s tud ie~ o u the eke le ton o f the m o u s e
and 3, or digits 3 and t, or all three of ~hem; the relative frequency of these Nsions i~ .partly controlled by the genetic background. As i~ sm/sm, fusions are confined to the ph~Ianges and apparently all primary; metacarpals and metatarsals are not affected, bug there are fairly extensive secondary fusions in carpus and ~arsus. In additiou to the anomalies on the feet, ~here is a gmaeral involvement of tt~e skeleton; the Iong bones have ehi~ sh~f~s, but epiI)hyses of nearly normal size; ~here are ~lso shape anomalies of seapala and of the saorM vertebrae. These anomalies are preformed in c~rtiLq.ge and o~n be ,~een in the newborn animal. Whether they are due to a disturbance of ehondrifioation or el:
cartilage growth is unknown. 4. Fusions between certain earpalia and/or tarsalia have been encomagered both in the
inbred strains A/Gr, @BA/Gr, C57BL/Gr and BALB/c and in heterogeneous stocks o~ mi~e. Thus BALB/c regularly kas ~ fttsion between multanguIum minus and centrale; 057 BL has the same Nsion associated with fusion beeween aavie ulare and mmeiforme a.~ .wtu]e other strains either lack these filsioas completely or have small percentages. These il:sions, or their absence, are thus :ruder geneticaI control.
I am deeply indebted eo Dr D. S. Falconer (Eclinbnrgh), to Nfiss Josephine S. gowe~ and Dr W. L. t;htssell (Oak t~idge, Tennessee, U.S.A.) gild to Prof. Peals Herbwig (Halle/ SaMe, Germany) who turned their genes sm, Os and sy over go me for study, and withou~ whose generosAy ~his work would not have been possible. ~'or technical assisgarxee in various ways I am iedeb~ed to lViiss "Johanaa 8to[ze (now i~'Irs J. Deol). This investigation has been s~tppor~ed by a grant fi'om the ltookefeller I~ounda{iom which is gratefully
acknowledged.
t~EFEP~ENCES
kgE~'% A. N. (1929). A morphological ~n~tysis of the foot ~bnormalities occurring ha the deseend~nts e~ X-rayed mice. Amgr. J. A~tat. 43, P~21-46.
BELL, J'~rxa (1953). 0~syndae~yly ~rtd i~s ~ssoei~ion with polyd~etyly. Tree,try ofH~man ~rnherita, nce:
5,' p~'t 2, pp. 33-50. B~AVS, H. (1906). Die ~ntwicklmag der Form der ~x~remi~i~t~n and de~ Ex~remi~emn~kelett~. In O~k~,r
Ker~dg's Ha~dbu~s der vergldchendsn und ez~oerimsntegen Entwic]~ungslehr~ dsr W~rbeftiere, 3,: p~rt 2, pp. 167-338.
BRYce, T. H. (I915). ~ax @~aln's El~.~gen~s of Anal~mny, t].~h ed. 4, p~r~ 1, pp. 157 end 199. London: Longm~ns, Green and Co.
@~s~E~, T. C, (1951). The genetics of ltLx~te mice. I. ~lorphologic~l abnormalities of h~erozygo~es ~nd i homozygotes. J. gznet. 51}, 277-99.
@ A ~ , T. C. (1954). The genetics of l.ux~te mice. IV. Embryology. Y. Gene& 52, 1-35, C~r~, Tso-KA~ (1939), The development of polydactylism in s, special str~h~ of 2/s mv,.sctd,~s~ [email protected]'
~,rat. Iglsl Bull. 14: 119-32. DAv-~a'oB~, C. B. (1909). Yz~her~b~nee of r m domestic fowl. 2~zbK Uar'~e2. [nstn, no. 121,
pp. i00. D]~TL]~]vSE~, J. A. & Ca_e~izct~L, W. J. (1921). InheA~ance of s:~md~e~ylism, bl~ck, ~nd dilu~io~ in
s~h~e. ]. Agrgm Rss. 20, 595-604. F~.xs, ~, (I954}. Anatomlsche trod ent~viddungsgesehieh~Iiehe Untersuchu~gen. ~m Skele~6 normaie~
m~d oligodacty]er }fguse. Wise. ~..g'Iarli~>Luther-Usdv. 3 (195a/r ~,Xg~h.-na%urw. lgeihe, IS[4, 801-2r
ll~El~k'l, 13/[. O. (195,5), Luxoid ~ new hereditary leg and foe~ abnormality in the house mouse..I. IIere~,,.
46, 91-9.

I~ANS GP~iTIg]3B ERG 145
~ r m ~ I ~ , I-L (1953). Cde~etica.l s~ndiea on the skeleton of ~he mouse. VII. Oongeni~[ hydrocephMus-
.t..(teqtet- 5t, 327-58. ~,~W~e, F. (1942}. irene 3iuta.@onen uud Koppelungsgruppen bei der Hausm~us. Z. i'~d'alal. Abstam~n.-
,~, gervrbLebre, 80, 220-46- I ~ , ~ G , p . (i951):. Entwietdungsgesckichtliehe Untersuohu_ngen fiber Bewegnngsstbrungenbei M~iusen-
Verb. A n a l GeM. (49. Vers. in ]:feidelberg, I6.-I9. April 1951), pp. 97-107. tteVg~xoq~=, A. (1920). _hz~a~omie et morphog~nte d'une anomalie hfr6dtit~ire des membres abdominaux
(Absence eongdnitale du tibia>. Bug. bier. Suppl. 3, 1-156. .~s~w, L. (19tl}. Trc~igd g'A~u3tomie fq,,r 6 ~ ed. 1, 381. paris: Oct~va Dora e~, fiis, T~Vs~ow, G. N. (1956). The ana.tomy ~nd develOpmEnt of the fidget mouse. J . genel. (in the Press),
APPENDIX
BY D. S. FALcoxs~
[nstitute of A~imal ~enetics, Ed,i~bu,rg/~ ~VMaetylism (sin) arose in Edinb~u'gh by spontaneous mutation in the A/Fa in-bred ~rain. It w~s found by Nr J. H. Isaaoson who also worked out the genetics. 0utcrosses were made to several non-inbred stocks and both backeross and intercross matings were m~de. The segregations observed are summarized in the gable. They show syndactylism ~:be recessive, bug there is a marked deficiency of homozygotes in the segregation pr e- g~ies at the time of classification, which was normally made within t clay of birth: i~iertality of syndaetyls after olassifiea~ion was high and the deficiency at birth is most prdbabty due to reduced viability.
Summary of sm seyregations GenotjTe of p~rent PhenoIcrpe of progeny
.~ ~ snl. + Total ' Viability
am~am sin~am. 14 0 14 - - sml~m -F / + ~ 0 92 92. - -
+ I + s.ml**~,. J ~m/am +/am ~6 144 220 0-83 +/~'m sm/s~n 54 78 129 0.72 + ~ors +/am 220 1278 1498 0.52
Tests for linkage were made by means of the linkagedesting stocks described by Carter & Falconer (1951), bub no linkage was found- On account of bhe high mortaI~ty of syn- da0tyls most of the tests have wide limits of error. The following are the marker genes ~gainst Which sm was ~ested.. The figu.re given with e~ch marker is tile closest admissible ilinkage, obtained by deducting twice the ,standard error from the observed reeomJ)jnation fi'equency, c~43, b 43, o 33,,fao, f~ r 39, ~ 26, Zz 36. N 32, p 21, Re 33, ru 28, s 56, 8d. 45, se 39, IV 47, v 30, 7r 45, wa-2 26, Mi a'/~ 48, sex 41. The ,significant excess Of re- combinants with s is not very convilldng as there was a significant de~cieney of as ~egregants as well as of ~m~m.
P~EFERENCE
0 ~ g ~ . T. C, & _FALtering, D. S. (1951)- S~oeks for de,ceding linkage in tLe mouse, aud ~t~e theory of
{;heir design. J . Gcnet. 50, 307-23.
10 @enet.. 54-