genetic variation in cultivated rheum tanguticumpopulations · genetic variation in cultivated...
TRANSCRIPT

Genetic variation in cultivated Rheum tanguticum populations
Yanping Hu1, Xiaolong Xie2, Li Wang1, Huaigang Zhang1, Jian Yang1 and Yi Li1
1Northwest Institute of Plateau Biology, the Chinese Academy of Sciences, Xining, China.2School of Pharmacy, Henan University of Traditional Chinese Medicine, Zhengzhou, China.
Abstract
To examine whether cultivation reduced genetic variation in the important Chinese medicinal plant Rheumtanguticum, the levels and distribution of genetic variation were investigated using ISSR markers. Fifty-eight R.tanguticum individuals from five cultivated populations were studied. Thirteen primers were used and a total of 320DNA bands were scored. High levels of genetic diversity were detected in cultivated R. tanguticum (PPB = 82.19,H = 0.2498, HB = 0.3231, I = 0.3812) and could be explained by the outcrossing system, as well as long-lived and hu-man-mediated seed exchanges. Analysis of molecular variance (AMOVA) showed that more genetic variation wasfound within populations (76.1%) than among them (23.9%). This was supported by the coefficient of gene differenti-ation (Gst = 0.2742) and Bayesian analysis (�B = 0.1963). The Mantel test revealed no significant correlation betweengenetic and geographic distances among populations (r = 0.1176, p = 0.3686). UPGMA showed that the five culti-vated populations were separated into three clusters, which was in good accordance with the results provided by theBayesian software STRUCTURE (K = 3). A short domestication history and no artificial selection may be an effectiveway of maintaining and conserving the gene pools of wild R. tanguticum.
Keywords: cultivated populations, genetic variation, ISSR, Polygonaceae, Rheum tanguticum Maxim. ex Balf.
Received: November 26, 2013; Accepted: May 12, 2014.
Introduction
Rheum tanguticum Maxim. ex Balf. (Dahuang in Chi-
nese) is an endangered perennial herbaceous plant endemic
to China, with a distribution mainly in Qinghai, Gansu
Provinces and west Tibetan Autonomous Region at alti-
tudes of 2,300-4,200 m above sea level; this plant is found
along forest edges and in valleys (Yang, 1991; Liu, 1997;
Li, 2003; Wang and Ren, 2009). The roots and rhizomes of
this plant officially listed in the Chinese Pharmacopoeia
have been used in traditional medicine for over 2,000 years
to treat various syndromes caused by circulatory problems
(e.g., dysmenorrhoea, hypermenorrhea, hematemesis, lo-
wer abdominal pain, etc), jaundice, diarrhea and constipa-
tion (Yang, 1991; Komatsu et al., 2006; Li et al., 2006;
Chinese Pharmacopoeia Commission, 2010).
As a result of over-harvesting and destruction of its
habitat by humans in recent decades, natural populations of
R. tanguticum have declined dramatically (Hu et al., 2010;
Wang, 2010). This led to R. tanguticum being listed as a
key protected plant of Qinghai Province by the government
in 2009 (The People’s Government of Qinghai Province,
2009). The breeding system of R. tanguticum is outcross-
ing, with pollination probably involving wind and insects
(Chen et al., 2009). Large panicles produce many winged
seeds that are dispersed by wind. The seed set and germina-
tion rates of R. tanguticum are very high (Xie X (2009) PhD
thesis, Graduate University of the Chinese Academy of Sci-
ences, Xining).
Since the 1960s, R. tanguticum has been extensively
cultivated in its original area of production (Qinghai Prov-
ince). The species is propagated predominantly by sexual
reproduction involving seeds. Farmers usually randomly
collect mature seeds directly from wild populations in local
or distant areas, mix them together, and plant them in the
fields. Sometimes, the seeds are exchanged with relatives
or friends, an activity that disperses the germplasm to other
places. To date, most studies of R. tanguticum have focused
on cultivation, tissue culture and analysis of the plant’s
chemical constituents (Zhang, 2004; Che et al., 2006;
VanMen et al., 2012). Although previous reports have pro-
vided preliminary assessments of the genetic diversity of
wild R. tanguticum (Chen et al., 2009; Hu et al., 2010;
Wang et al., 2012b), there has been no report on the genetic
diversity of cultivated populations.
Inter-simple sequence repeats (ISSR) are molecular
markers with primers designed based on repeat motifs
(microsatellites) of eukaryotic genomes that require no
prior knowledge of DNA sequence (Zietkiewicz et al.,
1994). These markers have good stability and large poly-
morphism. In recent years, ISSR has been successfully
Genetics and Molecular Biology, 37, 3, 540-548 (2014)
Copyright © 2014, Sociedade Brasileira de Genética. Printed in Brazil
www.sbg.org.br
Send correspondence to Yi Li. Northwest Institute of Plateau Biol-ogy, the Chinese Academy of Sciences, 23 Xinning Road, 810008,Xining, China. E-mail: [email protected].
Research Article

used to investigate the genetic diversity and relationships at
species, population and cultivar levels in many plants, in-
cluding some medicinal species (Tacuatia et al., 2012;
Chen et al., 2013; Kumchai et al., 2013; Thriveni et al.,
2013; Verma and Rana, 2013). In this study, we used ISSR
markers to: (1) detect the level of genetic diversity and
structure in five cultivated populations and (2) evaluate the
possible impact of cultivation practices on the genetic di-
versity of R. tanguticum. The results obtained should be
useful in developing strategies for efficient management of
the genetic resources of R. tanguticum and for future breed-
ing programs.
Materials and Methods
Plant materials
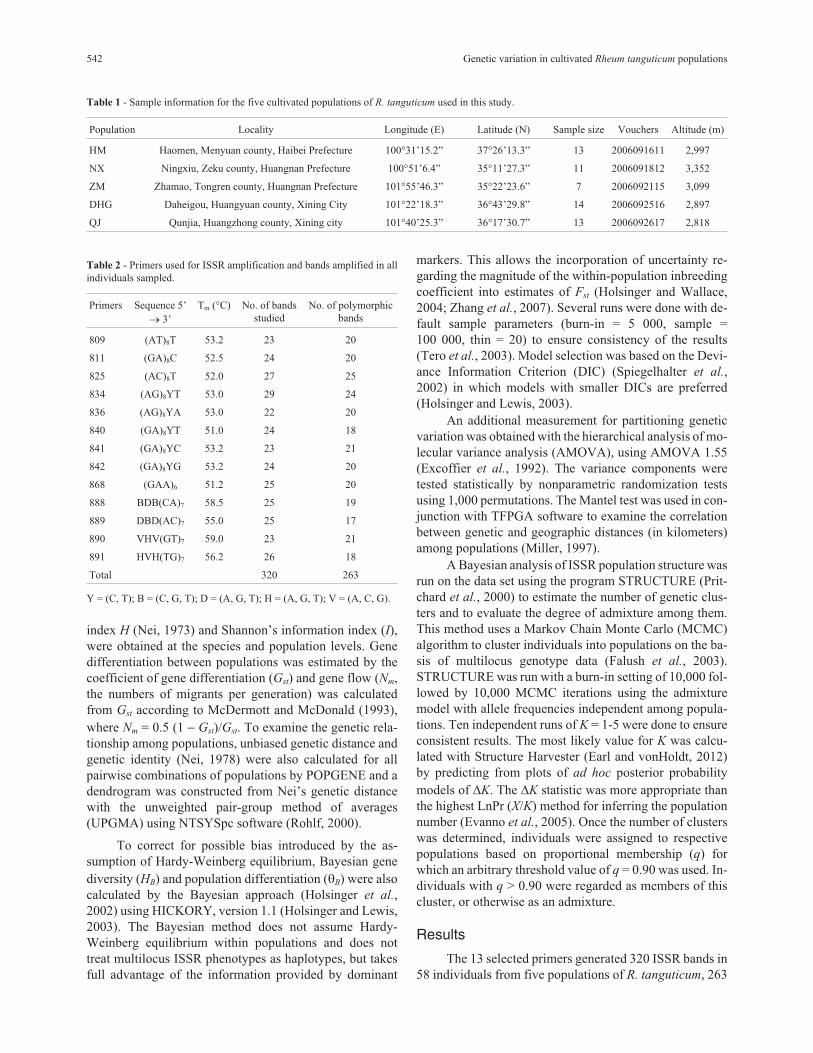
Fifty-eight individuals of R. tanguticum were sam-
pled from five cultivated populations in Qinghai Province,
China (Figure 1). Each population was positioned by GPS,
with the location details listed in Table 1. Young leaf tis-
sues were collected from individual plants located at least
10 m apart and then dried in silica gel. All of the material
collected was identified by Dr. Xuefeng Lu and voucher
specimens were deposited in the Qinghai-Tibetan Plateau
Museum of Biology, Northwest Institute of Plateau Biol-
ogy, Chinese Academy of Sciences.
DNA extraction and ISSR amplification
Genomic DNA was extracted using the modified
cetyltrimethylammonium bromide (CTAB) method de-
scribed by Doyle and Doyle (1987). The DNA concentra-
tion was determined by comparing the sample with known
standards of lambda DNA in 0.8% (w/v) agarose gels. The
isolated genomic DNA was diluted to 30 ng/�L and stored
at -20 °C until ISSR amplification.
One hundred primers from the University of British
Columbia (UBC set no. 9) were initially screened for PCR
amplification and 13 primers that produced clear, reproduc-
ible banding patterns were chosen for final analysis (Ta-
ble 2). PCR amplifications were done in a 20 �L reaction
volume consisting of 30 ng of genomic DNA, 3.0 mM
MgCl2, 0.1 mM dNTP, 10 pmol of primer, 0.75 U of Taq
DNA polymerase (TaKaRa Biotech Co., Ltd.) and 2.0 �L
of 10 PCR buffer. ISSR-PCR amplifications were done in a
PTC-221 thermocycler (MJ Research, Bio-Rad, USA) us-
ing the following program: an initial step of 5 min at 94 °C
followed by 20 s at 94 °C, 60 s at the appropriate annealing
temperature (see Table 2 for details) and 80 s at 72 °C for
38 cycles, with a final extension of 6 min at 72 °C. The neg-
ative control consisted of replacing template DNA with
ddH2O to test for contamination. The amplification prod-
ucts were separated in 1.5% agarose gels stained with
ethidium bromide and photographed with a GDP-8000
System (UVP Inc., USA). Molecular weights were esti-
mated using a 200 bp DNA ladder (TaKaRa Biotech Co.,
Ltd.).
Data analysis
Only unambiguously and reproducibly amplified
ISSR bands were scored as present (1) or absent (0).
Smeared and weak bands were excluded. The resulting bi-
nary data matrix was analyzed using POPGENE version
1.32 (Yeh et al., 1999) to estimate the level of genetic di-
versity under the assumption of Hardy-Weinberg equilib-
rium. Genetic diversity parameters, including the
percentage of polymorphic bands (PPB), the gene diversity
Hu et al. 541
Figure 1 - Locations of the five R. tanguticum populations sampled from Qinghai Province in this study.
index H (Nei, 1973) and Shannon’s information index (I),
were obtained at the species and population levels. Gene
differentiation between populations was estimated by the
coefficient of gene differentiation (Gst) and gene flow (Nm,
the numbers of migrants per generation) was calculated
from Gst according to McDermott and McDonald (1993),
where Nm = 0.5 (1 � Gst)/Gst. To examine the genetic rela-
tionship among populations, unbiased genetic distance and
genetic identity (Nei, 1978) were also calculated for all
pairwise combinations of populations by POPGENE and a
dendrogram was constructed from Nei’s genetic distance
with the unweighted pair-group method of averages
(UPGMA) using NTSYSpc software (Rohlf, 2000).
To correct for possible bias introduced by the as-
sumption of Hardy-Weinberg equilibrium, Bayesian gene
diversity (HB) and population differentiation (�B) were also
calculated by the Bayesian approach (Holsinger et al.,
2002) using HICKORY, version 1.1 (Holsinger and Lewis,
2003). The Bayesian method does not assume Hardy-
Weinberg equilibrium within populations and does not
treat multilocus ISSR phenotypes as haplotypes, but takes
full advantage of the information provided by dominant
markers. This allows the incorporation of uncertainty re-
garding the magnitude of the within-population inbreeding
coefficient into estimates of Fst (Holsinger and Wallace,
2004; Zhang et al., 2007). Several runs were done with de-
fault sample parameters (burn-in = 5 000, sample =
100 000, thin = 20) to ensure consistency of the results
(Tero et al., 2003). Model selection was based on the Devi-
ance Information Criterion (DIC) (Spiegelhalter et al.,
2002) in which models with smaller DICs are preferred
(Holsinger and Lewis, 2003).
An additional measurement for partitioning genetic
variation was obtained with the hierarchical analysis of mo-
lecular variance analysis (AMOVA), using AMOVA 1.55
(Excoffier et al., 1992). The variance components were
tested statistically by nonparametric randomization tests
using 1,000 permutations. The Mantel test was used in con-
junction with TFPGA software to examine the correlation
between genetic and geographic distances (in kilometers)
among populations (Miller, 1997).
A Bayesian analysis of ISSR population structure was
run on the data set using the program STRUCTURE (Prit-
chard et al., 2000) to estimate the number of genetic clus-
ters and to evaluate the degree of admixture among them.
This method uses a Markov Chain Monte Carlo (MCMC)
algorithm to cluster individuals into populations on the ba-
sis of multilocus genotype data (Falush et al., 2003).
STRUCTURE was run with a burn-in setting of 10,000 fol-
lowed by 10,000 MCMC iterations using the admixture
model with allele frequencies independent among popula-
tions. Ten independent runs of K = 1-5 were done to ensure
consistent results. The most likely value for K was calcu-
lated with Structure Harvester (Earl and vonHoldt, 2012)
by predicting from plots of ad hoc posterior probability
models of �K. The �K statistic was more appropriate than
the highest LnPr (X/K) method for inferring the population
number (Evanno et al., 2005). Once the number of clusters
was determined, individuals were assigned to respective
populations based on proportional membership (q) for
which an arbitrary threshold value of q = 0.90 was used. In-
dividuals with q > 0.90 were regarded as members of this
cluster, or otherwise as an admixture.
Results
The 13 selected primers generated 320 ISSR bands in
58 individuals from five populations of R. tanguticum, 263
542 Genetic variation in cultivated Rheum tanguticum populations
Table 1 - Sample information for the five cultivated populations of R. tanguticum used in this study.
Population Locality Longitude (E) Latitude (N) Sample size Vouchers Altitude (m)
HM Haomen, Menyuan county, Haibei Prefecture 100°31’15.2” 37°26’13.3” 13 2006091611 2,997
NX Ningxiu, Zeku county, Huangnan Prefecture 100°51’6.4” 35°11’27.3” 11 2006091812 3,352
ZM Zhamao, Tongren county, Huangnan Prefecture 101°55’46.3” 35°22’23.6” 7 2006092115 3,099
DHG Daheigou, Huangyuan county, Xining City 101°22’18.3” 36°43’29.8” 14 2006092516 2,897
QJ Qunjia, Huangzhong county, Xining city 101°40’25.3” 36°17’30.7” 13 2006092617 2,818
Table 2 - Primers used for ISSR amplification and bands amplified in all
individuals sampled.
Primers Sequence 5’
� 3’
Tm (°C) No. of bands
studied
No. of polymorphic
bands
809 (AT)8T 53.2 23 20
811 (GA)8C 52.5 24 20
825 (AC)8T 52.0 27 25
834 (AG)8YT 53.0 29 24
836 (AG)8YA 53.0 22 20
840 (GA)8YT 51.0 24 18
841 (GA)8YC 53.2 23 21
842 (GA)8YG 53.2 24 20
868 (GAA)6 51.2 25 20
888 BDB(CA)7 58.5 25 19
889 DBD(AC)7 55.0 25 17
890 VHV(GT)7 59.0 23 21
891 HVH(TG)7 56.2 26 18
Total 320 263
Y = (C, T); B = (C, G, T); D = (A, G, T); H = (A, G, T); V = (A, C, G).
(82.2%) of which were polymorphic (Table 3). The bands
ranged in size from 200 bp to 2,800 bp. The total number of
bands varied from 22 (UBC836) to 29 (UBC834), with an
average of 24.6 fragments per primer. The percentage of
polymorphic bands (PPB) ranged from 42.8% in the ZM
population to 51.8% in the DHG population (Table 3). In
terms of the presence of alleles within the 320 alleles de-
tected, there were 57 common alleles in the five popula-
tions but no unique allele was found for any of the five
populations studied.
The measurements of genetic diversity are summa-
rized in Table 3. The value of Nei’s gene diversity (H) was
0.2498 and Shannon’s Information index (I) was 0.3812 at
the species level while at the populations level the corre-
sponding values were 0.1813 and 0.2677, respectively. The
estimates of heterozygosity (HB) in the full model (which
had the lowest DIC, Table 4) based on the Bayesian proce-
dure were generally higher than Nei’s gene diversity (H).
The expected Bayesian heterozygosity (HB) was 0.3231 at
the species level and 0.2712 at the population level. Among
all the populations investigated, the highest and lowest lev-
els of genetic variability occurred in populations DHG
(H = 0.1949, I = 0.2878) and ZM (H = 0.1671, I = 0.2445),
respectively.
The genetic differentiation among populations (Gst),
estimated by Nei’s method, was 0.2742, which indicated
that 27.4% of the total genetic diversity was distributed
among populations, whereas 72.6% occurred within pop-
ulations. Furthermore, the level of gene flow (Nm) was es-
timated to be 1.3236 individuals per generation between
populations, suggesting that gene exchange between pop-
ulations was high. AMOVA analysis further revealed a
similar pattern of genetic differentiation among and
within the populations (Table 5). Of the total variation,
23.9% was attributed to among-population differences, a
value much lower than the within-population proportion
(76.1%). AMOVA also showed that differentiation
among populations and within populations was significant
(p < 0.0010).
The results for �B (analogous to Wright’s Fst) esti-
mated by the Bayesian analysis are shown in Table 4. The
best model, which had the smallest DIC (3876.31), was the
full model, with �B = 0.1963 and f = 0.0886.
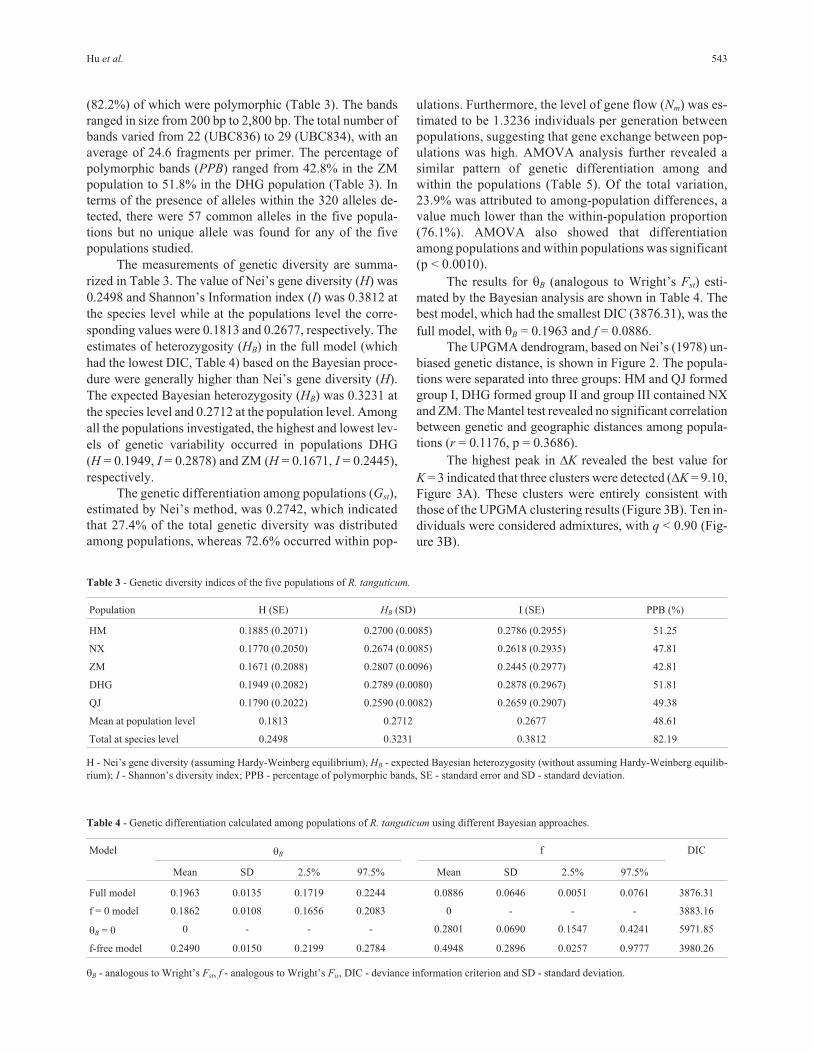
The UPGMA dendrogram, based on Nei’s (1978) un-
biased genetic distance, is shown in Figure 2. The popula-
tions were separated into three groups: HM and QJ formed
group I, DHG formed group II and group III contained NX
and ZM. The Mantel test revealed no significant correlation
between genetic and geographic distances among popula-
tions (r = 0.1176, p = 0.3686).
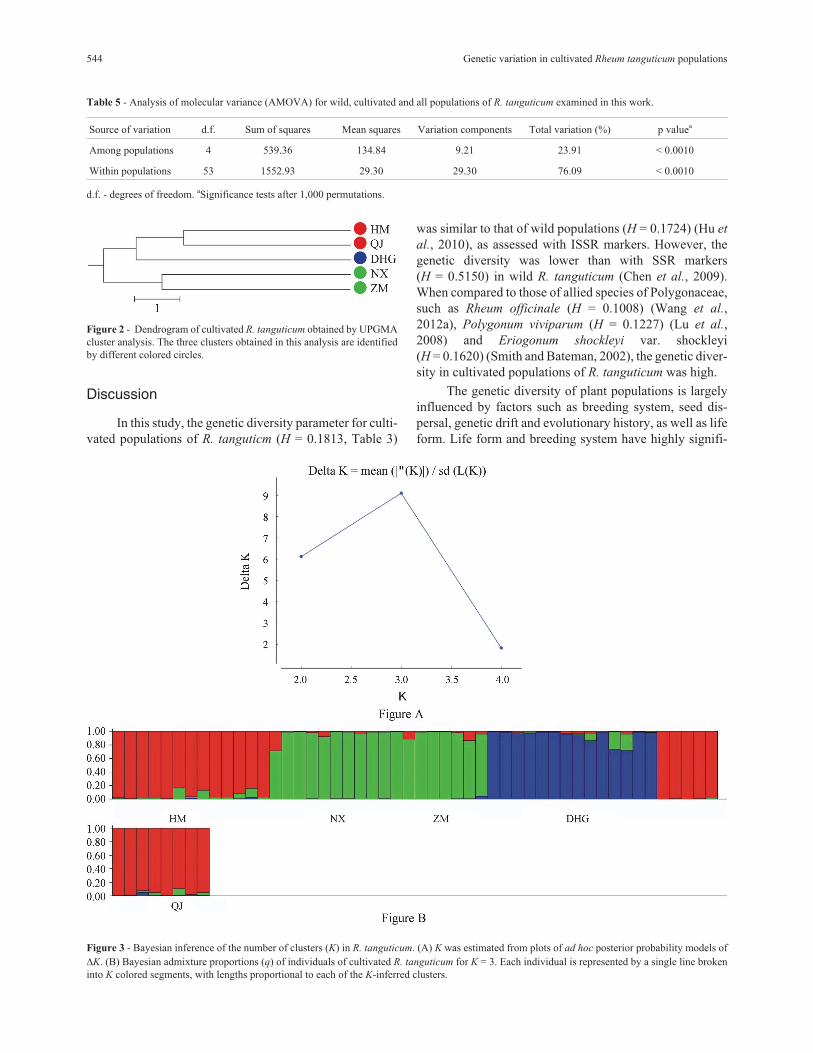
The highest peak in �K revealed the best value for
K = 3 indicated that three clusters were detected (�K = 9.10,
Figure 3A). These clusters were entirely consistent with
those of the UPGMA clustering results (Figure 3B). Ten in-
dividuals were considered admixtures, with q < 0.90 (Fig-
ure 3B).
Hu et al. 543
Table 3 - Genetic diversity indices of the five populations of R. tanguticum.
Population H (SE) HB (SD) I (SE) PPB (%)
HM 0.1885 (0.2071) 0.2700 (0.0085) 0.2786 (0.2955) 51.25
NX 0.1770 (0.2050) 0.2674 (0.0085) 0.2618 (0.2935) 47.81
ZM 0.1671 (0.2088) 0.2807 (0.0096) 0.2445 (0.2977) 42.81
DHG 0.1949 (0.2082) 0.2789 (0.0080) 0.2878 (0.2967) 51.81
QJ 0.1790 (0.2022) 0.2590 (0.0082) 0.2659 (0.2907) 49.38
Mean at population level 0.1813 0.2712 0.2677 48.61
Total at species level 0.2498 0.3231 0.3812 82.19
H - Nei’s gene diversity (assuming Hardy-Weinberg equilibrium), HB - expected Bayesian heterozygosity (without assuming Hardy-Weinberg equilib-
rium); I - Shannon’s diversity index; PPB - percentage of polymorphic bands, SE - standard error and SD - standard deviation.
Table 4 - Genetic differentiation calculated among populations of R. tanguticum using different Bayesian approaches.
Model �B f DIC
Mean SD 2.5% 97.5% Mean SD 2.5% 97.5%
Full model 0.1963 0.0135 0.1719 0.2244 0.0886 0.0646 0.0051 0.0761 3876.31
f = 0 model 0.1862 0.0108 0.1656 0.2083 0 - - - 3883.16
�B = 0 0 - - - 0.2801 0.0690 0.1547 0.4241 5971.85
f-free model 0.2490 0.0150 0.2199 0.2784 0.4948 0.2896 0.0257 0.9777 3980.26
�B - analogous to Wright’s Fst, f - analogous to Wright’s Fis, DIC - deviance information criterion and SD - standard deviation.
Discussion
In this study, the genetic diversity parameter for culti-
vated populations of R. tanguticm (H = 0.1813, Table 3)
was similar to that of wild populations (H = 0.1724) (Hu et
al., 2010), as assessed with ISSR markers. However, the
genetic diversity was lower than with SSR markers
(H = 0.5150) in wild R. tanguticum (Chen et al., 2009).
When compared to those of allied species of Polygonaceae,
such as Rheum officinale (H = 0.1008) (Wang et al.,
2012a), Polygonum viviparum (H = 0.1227) (Lu et al.,
2008) and Eriogonum shockleyi var. shockleyi
(H = 0.1620) (Smith and Bateman, 2002), the genetic diver-
sity in cultivated populations of R. tanguticum was high.
The genetic diversity of plant populations is largely
influenced by factors such as breeding system, seed dis-
persal, genetic drift and evolutionary history, as well as life
form. Life form and breeding system have highly signifi-
544 Genetic variation in cultivated Rheum tanguticum populations
Figure 2 - Dendrogram of cultivated R. tanguticum obtained by UPGMA
cluster analysis. The three clusters obtained in this analysis are identified
by different colored circles.
Table 5 - Analysis of molecular variance (AMOVA) for wild, cultivated and all populations of R. tanguticum examined in this work.
Source of variation d.f. Sum of squares Mean squares Variation components Total variation (%) p valuea
Among populations 4 539.36 134.84 9.21 23.91 < 0.0010
Within populations 53 1552.93 29.30 29.30 76.09 < 0.0010
d.f. - degrees of freedom. aSignificance tests after 1,000 permutations.
Figure 3 - Bayesian inference of the number of clusters (K) in R. tanguticum. (A) K was estimated from plots of ad hoc posterior probability models of
�K. (B) Bayesian admixture proportions (q) of individuals of cultivated R. tanguticum for K = 3. Each individual is represented by a single line broken
into K colored segments, with lengths proportional to each of the K-inferred clusters.

cant influences on genetic diversity. In general, long-lived
and outcrossing species have higher levels of genetic diver-
sity than selfing and clonal plants (Hamrick and Godt,
1996). Bayesian analysis revealed that the inbreeding coef-
ficient (f = 0.0886) in R. tanguticum was small, a finding
that confirmed the outcrossing breeding system of this spe-
cies. Outcrossing and elevated longevity contribute to the
moderately high level of genetic diversity in R. tanguticum.
Notably, the current high level of genetic diversity in culti-
vated R. tanguticum may be markedly influenced by the tra-
ditional, irregular and sparse agricultural practices of this
plant. Traditionally, growers collect and preserve R.
tanguticum seeds randomly without deliberate selection
and mix them together before planting. This practice would
preserve more genetic diversity than in wild plants.
R. tanguticum has been planted in Qinghai Province
since the 1960s. The domestication history is short, there is
no artificial selection and cultivation apparently does not
influence genetic diversity. In cultivated populations, ge-
netic diversity is partly determined by the way genetic ma-
terial is passed from one cultivated generation to the next
(Miller and Schaal, 2006; Yao et al., 2012). For R.
tanguticum, seeds are the main genetic material used to es-
tablish the cultivated population. Cultivated R. tanguticum
may have been established from the seeds of a large number
of wild progenitors. According to local growers, R.
tanguticum are collected randomly and mixed together be-
fore planting. This was confirmed by the proportional
membership (q) calculated with Bayesian analysis. The
value of q in 10 individuals was < 0.90 (Figure 3B), indicat-
ing that the origins of these 10 individuals differed from
that of other individuals in the same population.
The germplasm can occasionally be dispersed to
other places by farmers’ relatives or friends. For exam-
ple, the QJ population was planted by researchers of our
institute. The seeds of this population were collected
from different places in Golog Prefecture and mixed to-
gether. This was confirmed by Bayesian analysis with
STRUCTURE software. Figure 3B shows that there were
five individuals with a proportional membership < 0.90.
The frequent exchange of seeds further improves the
maintenance of genetic diversity (Guo et al., 2007; Yao
et al., 2012). Consequently, traditional agricultural prac-
tice applied to this plant was another important factor
that influenced the abundant genetic diversity of culti-
vated R. tanguticum.
Compared to annual plants, cultivated populations of
some perennial plants may harbor a relatively higher per-
centage of genetic diversity than their wild ancestors
(Otero-Arnaiz et al., 2005; Miller and Schaal, 2006; He et
al., 2009). Our findings were similar to several other
long-lived cultivated plants in which genetic diversity is as
high as in the wild relatives (He et al., 2007; Shi et al.,
2008). Cultivated R. tanguticum had moderate levels of ge-
netic diversity compared with cultivated populations of
other endangered Chinese medicinal plants (Table 6) (Wu
et al., 2006; Guo et al., 2007; Qiu et al., 2009; Song et al.,
2010; Li et al., 2011; Yao et al., 2012), i.e., there was no ev-
idence that a “cultivation bottleneck or founder effect” af-
fected the genetic diversity.
Long-lived and outcrossing species retain most of
their genetic variability within populations. In contrast, an-
nual and selfing species allocate most of their genetic vari-
ability among populations (Hamrick and Godt, 1996;
Nybom, 2004). Our study was consistent with this trend.
AMOVA results revealed that in cultivated R. tanguticum
76.1% of the genetic variance was retained within popula-
tions while 23.9% was among populations (Table 5). In the
long-lived tree Spondias purpurea, the proportion of ge-
netic variation distributed among populations was greater
in cultivated populations than in wild populations, a reflec-
tion of the relative amounts of vegetative propagation (Mil-
ler and Schaal, 2006). The opposite was observed in culti-
vated R. tanguticum. Cultivated populations (Gst = 0.2742,
�st = 0.239) had a lower proportion of their genetic vari-
ability distributed among populations than wild popula-
tions (Gst = 0.3585, �st = 0.290) (Hu et al., 2010). Bayesian
analysis revealed that the inbreeding coefficient
(f = 0.0886) in R. tanguticum was small. Based on the re-
cords of local growers, these cultivated R. tanguticum were
grown from seeds rather than being propagated vegeta-
Hu et al. 545
Table 6 - Comparison of genetic diversity of cultivated R. tanguticum with other important herbs in cultivated populations.
Species Genetic diversity values Markers References
R. tanguticum PPBs = 82.19%; Hs = 0.2498; PPBp = 48.61%; Hp = 0.1813 ISSR Present study
Salvia miltiorrhiza PPBs = 100%; Hs = 0.1951; PPBp = 65.00%; Hp = 0.1591 ISSR Song et al. (2010)
Panax ginseng PPBs = 98.96%; Hs = 0.2886; PPBp = 58.01%; Hp = 0.1890 ISSR Li et al. (2011)
Gastrodia elata PPBs = 61.04%; Hs = 0.1280; PPBp = 35.71%; Hp = 0.1000 ISSR Wu et al. (2006)
Corydalis yanhusuo PPBs = 71.54%; Hs = 0.1630; PPBp = 25.32%; Hp = 0.0940 ISSR Qiu et al. (2009)
Codonopsis pilosula PPBs = 87.23%; Hs = 0.2992; PPBp = 73.43%; Hp = 0.2367 RAPD Guo et al. (2007)
Eucommia ulmoides PPBp = 51.23%; Hp = 0.1572 AFLP Yao et al. (2012)
Hp - Nei’s gene diversity at the population level, Hs - Nei’s gene diversity at the species level, PPBp - percentage of polymorphic loci at the population
level and PPBs - percentage of polymorphic loci at the species level.

tively. Thus, the mating system was outcrossing in culti-
vated populations, the same as in wild populations.
STRUCTURE analysis showed that 10 individuals
were admixtures with q < 0.90 (Figure 3B), indicating that
cultivated R. tangutium had been introduced to cultivation
from the seeds of a large number of wild progenitors. The
planting records revealed that the germplasm had been dis-
persed to other places by the growers, with mixing and ex-
changing of seeds before planting. The frequent exchange
of seeds enhanced the gene flow from wild populations into
cultivated populations. Subsequently, the genetic variation
within populations increased while that among populations
decreased in cultivated populations.
The findings of this study indicate that R. tanguticum
has maintained a relatively high level of genetic diversity in
cultivated populations that may play a crucial role in con-
serving this species in the face of declining wild popula-
tions. A short cultivation history and no artificial selection
do not decrease the genetic diversity of R. tanguticum and
no special cultivar is formed. The primitive agricultural
practices, i.e., random collecting, preserving and planting
of seeds without deliberate selection may be an effective
way of maintaining and conserving the gene pools of wild
plants (Guo et al., 2007; Song et al., 2010). Furthermore,
given that the genetic diversity of the QJ cultivated popula-
tions of R. tanguticum was relatively higher than that of
wild populations at the population level, the current plant-
ing at this site can be seen as a preliminary step for ex situ
conservation. Additionally, other effective approaches,
such as Good Agricultural Practice (GAP), also need be
adopted to meet the market demand and achieve future sus-
tainable use of this medicinal plant.
Acknowledgments
This work was supported by the National Key Tech-
nology R&D Program of China (grant no.
2012BAC08B04), the National Nature Science Foundation
of China (grant no. 31200245) and the West Light Founda-
tion of the Chinese Academy of Sciences for Doctors
(2011).
References
Che G, Li Y, Wang L, Zhou G, Suo Y and Chen G (2006) Sea-
sonal change of anthroquinone content in cultivated Rheum
tanguticum. Acta Bot Boreal-Occident Sin 26:2378-2382.
Chen F, Wang A, Chen K, Wan D and Liu J (2009) Genetic diver-
sity and population structure of the endangered and medi-
cally important Rheum tanguticum (Polygonaceae) revealed
by SSR markers. Biochem Syst Ecol 37:613-621.
Chen DX, Li LY, Zhang X, Wang Y and Zhang Z (2013) Genetic
diversity in wild Dipsacus chinensis populations from China
based on ISSR markers. Genet Mol Res 12:1205-1213.
Chinese Pharmacopeia Commission (2010) Pharmacopoeia of the
People’s Republic of China. vol 1. China Medical Science
Press, Beijing, pp 22-23.
Doyle J and Doyle J (1987) A rapid DNA isolation procedure for
small quantities of fresh leaf tissue. Phytochem Bull 19:11-
15.
Evanno G, Regnaut S and Goudet J (2005) Detecting the number
of clusters of individuals using the software STRUCTURE:
A simulation study. Mol Ecol 14:2611-2620.
Excoffier L, Smouse P and Quattro J (1992) Analysis of molecu-
lar variance inferred from metric distances among DNA
haplotypes: Application to human mitochondrial DNA re-
striction data. Genetics 131:479-491.
Falush D, Stephens M and Pritchard JK (2003) Inference of popu-
lation structure using multilocus genotype data: Linked loci
and correlated allele frequencies. Genetics 164:1567-1587.
Guo HB, Lu BR, Wu QH, Chen JK and Zhou TS (2007) Abundant
genetic diversity in cultivated Codonopsis pilosula popula-
tions revealed by RAPD polymorphisms. Genet Resour
Crop Evol 54:917-924.
Hamrick JL and Godt MJW (1996) Effects of life history traits on
genetic diversity in plant species. Philos Trans R Soc Lond B
Biol Sci 351:1291-1298.
He J, Wang H, Li DZ and Chen SF (2007) Genetic diversity of
Paris polyphylla var. yunnanensis, a traditional Chinese me-
dicinal herb, detected by ISSR markers. Planta Med
73:1316-1321.
He JS, Chen L, Si Y, Huang B, Ban XQ and Wang YW (2009)
Population structure and genetic diversity distribution in
wild and cultivated populations of the traditional Chinese
medicinal plant Magnolia officinalis subsp biloba
(Magnoliaceae). Genetica 135:233-243.
Holsinger KE and Lewis PO (2003) HICKORY: A package for
analysis of population genetic data Ver 1.1. Department of
Ecology and Evolutionary Biology, University of Connecti-
cut, Storrs, USA.
Holsinger KE and Wallace LE (2004) Bayesian approaches for
the analysis of population genetic structure: An example
from Platanthera leucophaea (Orchidaceae). Mol Ecol
13:887-894.
Holsinger KE, Lewis PO and Dey DK (2002) A Bayesian ap-
proach to inferring population structure from dominant
markers. Mol Ecol 11:1157-1164.
Hu Y, Wang L, Xie X, Yang J, Li Y and Zhang H (2010) Genetic
diversity of wild populations of Rheum tanguticum endemic
to China as revealed by ISSR analysis. Biochem Syst Ecol
38:264-274.
Komatsu K, Nagayama Y, Tanaka K, Ling Y, Basnet P and
Meselhy MR (2006) Development of a high performance
liquid chromatographic method for systematic quantitative
analysis of chemical constituents in rhubarb. Chem Pharm
Bull (Tokyo) 54:941-947.
Kumchai J, Wei YC, Lee CY, Chen FC and Chin SW (2013) Pro-
duction of interspecific hybrids between commercial culti-
vars of the eggplant (Solanum melongena L.) and its wild
relative S. torvum. Gen Mol Res 12:755-764.
Li A (2003) Flora of China 5. Science Press, Beijing, 444 pp.
Li M, Li LX, Liu Y and Liu Y (2006) Study survey on rhubarb in
recent years. World Sci Technol-Mod Tradit Chin Med Mat
Med 8:34-39.
Li S, Li J, Yang XL, Cheng Z and Zhang WJ (2011) Genetic diver-
sity and differentiation of cultivated ginseng (Panax ginseng
C. A. Meyer) populations in north-east China revealed by
546 Genetic variation in cultivated Rheum tanguticum populations

inter-simple sequence repeat (SSR) markers. Genet Resour
Crop Evol 58:815-824.
Liu S (1997) Flora of Qinghai, vol 2. Qinghai People’s Publishing
House, Xining, 463 pp.
Lu J, Yang X and Ma R (2008) Genetic diversity of clonal plant
Polygonum viviparum based RAPD in eastern Qinghai-
Tibet Plateau of China. J Northwest Normal University (Nat
Sci) 44:66-72.
McDermott J and McDonald B (1993) Gene flow in plant patho-
systems. Annu Rev Phytopathol 31:353-373.
Miller AJ and Schaal BA (2006) Domestication and the distribu-
tion of genetic variation in wild and cultivated populations
of the Mesoamerican fruit tree Spondias purpurea L.
(Anacardiaceae). Mol Ecol 15:1467-1480.
Nei M (1973) Analysis of gene diversity in subdivided popula-
tions. Proc Natl Acad Sci USA 70:3321-3323.
Nei M (1978) Estimation of average heterozygosity and genetic
distance from a small number of individuals. Genetics
89:583-590.
Nybom H (2004) Comparison of different nuclear DNA markers
for estimating intraspecific genetic diversity in plants. Mol
Ecol 13:1143-1155.
Otero-Arnaiz A, Casas A, Hamrick JL and Cruse-Sanders J
(2005) Genetic variation and evolution of Polaskia chichipe
(Cactaceae) under domestication in the Tehuacan Valley,
central Mexico. Mol Ecol 14:1603-1611.
Pritchard JK, Stephens M and Donnelly P (2000) Inference of
population structure using multilocus genotype data. Genet-
ics 155:945-959.
Qiu YX, Zong M, Yao Y, Chen BL, Zhou XL, Chen ZL and Fu
CX (2009) Genetic variation in wild and cultivated Rhizoma
Corydalis revealed by ISSRs markers. Planta Med 75:94-98.
Rohlf F (2000) NTSYS-pc: Numerical taxonomy and multivariate
analysis system, ver 2.1. New York, NY.
Shi W, Yang CF, Chen JM and Guo YH (2008) Genetic variation
among wild and cultivated populations of the Chinese me-
dicinal plant Coptis chinensis (Ranunculaceae). Plant Biol
10:485-491.
Smith JF and Bateman TA (2002) Genetic differentiation of rare
and common varieties of Eriogonum shockleyi (Polygo-
naceae) in Idaho using ISSR variability. West N Am Nat
62:316-326.
Song Z, Li X, Wang H and Wang J (2010) Genetic diversity and
population structure of Salvia miltiorrhiza Bge in China re-
vealed by ISSR and SRAP. Genetica 138:241-249.
Spiegelhalter DJ, Best NG, Carlin BR and van der Linde A (2002)
Bayesian measures of model complexity and fit. J R Stat Soc
Series B-Stat Methodol 64:583-616.
Tacuatia LO, Eggers L, Kaltchuk-Santos E and Souza-Chies TT
(2012) Population genetic structure of Sisyrinchium
micranthum Cav. (Iridaceae) in Itapua State Park, southern
Brazil. Genet Mol Biol 35:99-105.
Tero N, Aspi J, Siikamäki P, Jäkäläniemi A and Tuomi J (2003)
Genetic structure and gene flow in a metapopulation of an
endangered plant species, Silene tatarica. Mol Ecol
12:2073-2085.
The People’s Government of Qinghai Province (2009) List of key
protected wild plants of Qinghai Province. Qinghai Prata-
culture 18:50-51.
Thriveni H, Sumangala R, Shivaprakash K, Ravikanth G, Vasu-
deva R and Babu HR (2014) Genetic structure and diversity
of Coscinium fenestratum: A critically endangered liana of
Western Ghats, India. Plant Syst Evol 300:403-413.
VanMen C, Jang YS, Zhu HM, Lee JH, Trung TN, Ngoc TM, Kim
YH and Kang JS (2012) Chemical-based species classifica-
tion of rhubarb using simultaneous determination of five
bioactive substances by HPLC and LDA analysis. Phyto-
chem Anal 23:359-364.
Verma S and Rana TS (2013) Genetic relationships among wild
and cultivated accessions of curry leaf plant (Murraya
koenigii (L.) Spreng.), as revealed by DNA fingerprinting
Methods Mol Biotechnol 53:139-149.
Wang XM (2010) Optimization of DNA isolation, ISSR-PCR sys-
tem and primers screening of genuine species of rhubarb, an
important herbal medicine in China. J Med Plants Res
4:904-908.
Wang X and Ren Y (2009) Rheum tanguticum, an endangered me-
dicinal plant endemic to China. J Med Plants Res 3:1195-
1203.
Wang XM, Hou XQ, Zhang YQ, Yang R, Feng SF, Li Y and Ren
Y (2012a) Genetic diversity of the endemic and medicinally
important plant Rheum officinale as revealed by inter-simple
sequence repeat (ISSR) markers. Int J Mol Sci 13:3900-
3915.
Wang X, Yang R, Feng S, Hou X, Zhang Y, Li Y and Ren Y
(2012b) Genetic variation in Rheum palmatum and Rheum
tanguticum (Polygonaceae), two medicinally and endemic
species in China using ISSR markers. PLoS ONE 7:e51667.
Wu HF, Li Z and Huang H (2006) Genetic differentiation among
natural populations of Gastrodia elata (Orchidaceae) in
Hubei and germplasm assessment of the cultivated popula-
tions. Biodiv Sci 14:315-326.
Yang M, Zhang D, Zheng J and Liu J (2001) Pollen morphology
and its systematic and ecological significance in Rheum
(Polygonaceae) from China. Nord J Bot 21:411-418.
Yang YC (1991) Tibetan Medicine. Qinghai People’s Press,
Xining, 670 pp.
Yao X, Deng J and Huang H (2012) Genetic diversity in
Eucommia ulmoides (Eucommiaceae), an endangered tradi-
tional Chinese medicinal plant. Conserv Genet 13:1499-
1507.
Yeh F, Yang R and Boyle T (1999) POPGENE version 1.32.
Microsoft Window-based freeware for population genetic
analysis. Molecular Biology and Biotechnology Center,
University of Alberta, Canada.
Zhang XF (2004) Xining rhubarb. Science Press of Shanxi, Xi’an,
168 pp.
Zhang D, Chen S and Gao Q (2007) Patterns of genetic variation
in Swertia przewalskii, an endangered endemic species of
the Qinghai-Tibet Plateau. Biochem Genet 45:33-50.
Zietkiewicz E, Rafalski A and Labuda D (1994) Genome finger-
printing by simple sequence repeat (SSR)-anchored poly-
merase chain-reaction amplification. Genomics 20:176-183.
Internet Resources
Primer Sets of University of British Columbia (UBC) set no. 9,
http://www.michaelsmith.ubc.ca/ser-
vices/NAPS/Primer_Sets/Primers_Oct2006.pdf (October 5,
2006).
Earl DA and von Holdt BM (2012) STRUCTURE HARVESTER:
A website and program for visualizing STRUCTURE output
Hu et al. 547

and implementing the Evanno method. Conserv Genet Res
4:359-361. http://taylor0.biol-
ogy.ucla.edu/structureHarvester/ (November 1, 2012).
Miller M (1997) Tools for population genetic analyses (TFPGA)
1.3: A Windows program for the analysis of allozyme and
molecular population genetic data. Department of Biologi-
cal Sciences, Northern Arizona University, Arizona, USA.
http://www.marksgeneticsoftware.net/_vti_bin/shtml.exe/tf
pga.htm (March 5, 2009).
Associate Editor: Everaldo Gonçalves de Barros
License information: This is an open-access article distributed under the terms of theCreative Commons Attribution License, which permits unrestricted use, distribution, andreproduction in any medium, provided the original work is properly cited.
548 Genetic variation in cultivated Rheum tanguticum populations