genetic phenotypic diversity bacillus soil rhizosphereaem.asm.org/content/58/6/1894.full.pdfgenetic...
TRANSCRIPT

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, June 1992, p. 1894-19030099-2240/92/061894-10$02.00/0
Genetic and Phenotypic Diversity of Bacillus polymyxa in Soiland in the Wheat Rhizosphere
PATRICK MAVINGUI,l* GISELE LAGUERRE,2 ODILE BERGE,' AND THIERRY HEULIN'
Equipe d'Ecologie Microbienne de la Rhizosphere, Centre de Pedologie Biologique, Unite Propre6831 du Centre National de la Recherche Scientifique Associe6e a l'Universite Nancy I, B. P. 5,F-54501 Vandoeuvre-les-Nancy Cedex, 1 and Laboratoire de Microbiologie des Sols, Institut
National de la Recherche Agronomique, BV 1540, 21034 Dijon Cedex, 2 France
Received 18 November 1991/Accepted 25 March 1992
Diversity among 130 strains ofBaciUlus polymyxa was studied; the bacteria were isolated by immunotrappingfrom nonrhizosphere soil (32 strains), rhizosphere soil (38 strains), and the rhizoplane (60 strains) of wheatplantlets growing in a growth chamber. The strains were characterized phenotypically by 63 auxanographic(API 50 CHB and API 20B strips) and morphological features, serologically by an enzyme-linked immuno-sorbent assay, and genetically by restriction fragment length polymorphism (RFLP) profiles of total DNA incombination with hybridization patterns obtained with an rRNA gene probe. Cluster analysis of phenotypiccharacters by the unweighted pair group method with averages indicated four groups at a similarity level of93%. Clustering of B. polymyxa strains from the various fractions showed that the strains isolated fromnonrhizosphere soil fell into two groups (I and II), while the third group (III) mainly comprised strains isolatedfrom rhizosphere soil. The last group (IV) included strains isolated exclusively from the rhizoplane. Strainsbelonging to a particular group exhibited a similarity level of 96%. Serological properties revealed a highervariability among strains isolated from nonrhizosphere and rhizosphere soil than among rhizoplane strains.RFLP patterns also revealed a greater genetic diversity among strains isolated from nonrhizosphere andrhizosphere soil and therefore could not be clearly grouped. The RFLP patterns of sorbitol-positive strainsisolated from the rhizoplane were identical. These results indicate that diversity within populations of B.polymyxa isolated from nonrhizosphere and rhizosphere soil is higher than that of B. polymyxa isolated fromthe rhizoplane. It therefore appears that wheat roots may select a specific subpopulation from the soil B.polymyxa population.
It is well established that beneficial interactions occur
between plants and microorganisms living in their rhizo-sphere (29). Beneficial plant-associated microorganisms in-clude symbionts, such as the root-nodulating Rhizobium andactinomycete Frankia spp. and the endomycorrhizal fungi,and nonsymbionts known as rhizobacteria (29). Specificrelationships between plants and associated microorganismsexist and have been well illustrated in the Rhizobium sym-biosis (14).
Beneficial free-living rhizobacteria belonging to the genusBacillus have been isolated from the rhizosphere of Grami-nae spp. (9-11, 39, 40). The presence of a nitrogen-fixingbacterium, Bacillus polymyxa (20), in the rhizosphere ofwheat has also been reported (27, 35). Recently, B. poly-myxa was used as a potential growth-promoting rhizobacte-rium for forage plant species (8, 21). The association be-tween B. polymyxa and plants such as perennial ryegrass(Lolium perene L.) and white clover (Tnfolium repens L.)was found to be specific and probably involves coadaptation(8). The adapted bacterial populations living in a specializedecological niche, such as plant roots, could be generated byphenotypic selection, genomic rearrangements within indi-viduals, or gene transfers between microorganisms, all ofwhich occur in natural microbial populations (18, 26, 46).The potential for and the extent of bacterial adaptation to
the highly heterogeneous and fluctuating environment ofcultivated soil depend on their genetic diversity. Althoughdiversity among free-living soil bacteria has been measured
* Corresponding author.
(49, 50), very little work has compared the phenotypic andgenotypic diversity of the bacteria in the rhizosphere. Theeffect of plant roots on the size and composition of soilbacteria has been well documented (33, 44). It has beenshown that the rhizoplane (RP) and the rhizosphere soil (RS)of many crops contain larger numbers of microorganismsthan the corresponding nonrhizosphere soil (NRS) (41).Preferential stimulation of bacteria in the RP and the rhizo-sphere has also been reported (42). A decrease in thephenotypic diversity of bacterial populations in the RS andthe RP compared with the NRS of rosana western wheat-grass (Agropyron smithii) and fourwing salbush (Atriple-xcanescens) has been reported (32). These studies examinedmicrobial communities, including several bacterial speciesand genera.Very few attempts have been made to study the diversity
of bacterial populations found within and around the rhizo-sphere, possibly because of the lack of suitable techniquesfor isolating a sufficient number of strains belonging to thesame species. In the present work, diversity was evaluatedwithin one species, B. polymyxa. This bacterium has beendescribed as a clearly distinct species by several workers(12, 38) and was suitable for this study.
In this study, we aimed to determine the diversity amongpopulations of B. polymyxa associated with roots of wheatby their phenotypic, serological, and genetic characteristics.Three populations of B. polymyxa isolated by immunotrap-ping from (i) NRS, (ii) RS, and (iii) the RP of spring wheatwere compared. The use of the immunotrapping techniquepreviously described (31) allowed the isolation of strainsrepresenting less than 1% of the culturable microflora. This
1894
Vol. 58, No. 6
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. POLYMYXA FROM SOIL AND RHIZOSPHERE 1895
approach may help in an understanding of the capacity ofplants to select a bacterial population from soil and themechanisms involved in plant-microbe interactions.
MATERIALS AND METHODS
Soil and plants. The soil was a silty eutric cambisol (pH5.5) sampled from a crop field in eastern France (Dieul-ouard). The regular crop rotation in this field over 5 years isrape (Brassica napus L.) followed by wheat or maize. Threesoil samples were collected from fallow sites 3 m apartaround the crop field. The soil was sampled from the top 30cm and passed through a 5-mm-pore-size sieve. Three potswere filled with the soil of each sample. Spring wheatseedlings (Triticiwn aestivum L., cv. Fidel) were grown inpots (11-cm diameter) containing 1.5 kg of soil in a growthchamber with a 16-h photoperiod of 300 microeinsteins m-2s-'. Day and night temperatures were 21 + 1 and 15 + 20C,respectively, with a relative humidity of 85% + 5%. Uncul-tivated pots were also maintained under similar conditions oftemperature and humidity.
Sampling and immunotrapping. Plants were harvested at18 days of growth. The following fractions were considered:(i) NRS, i.e., the uncultivated soil; (ii) RS, i.e., maceratedsamples of roots plus the closely adhering soil; and (iii) theRP, i.e., macerated samples of roots washed free of soil withsterile distilled water by gentle shaking. Pots containing fourplants at the three-leaf stage were pooled and considered anindividual sample. Samples of 2.5 g of roots or 5 g of soilwere macerated with a sterile mortar and pestle in 45 ml ofphosphate buffer (0.72% [wt/vol] NaCl, 0.28% Na2HPO4,0.04% KH2PO4 [pH 7.2]) to obtain the first dilution. Subse-quent 10-fold serial dilutions in phosphate buffer were pre-pared and used for immunotrapping of bacteria as describedby Mavingui et al. (31). Microtiter plates were coated withpartially purified rabbit polyclonal antibodies (PcAbs) (10-3dilution). A 10-3 dilution of soil or root sample was added(100 [l per well). After incubation at 4°C, the plates werewashed twice with phosphate buffer (pH 7.2) and the bacte-ria were desorbed with 0.1 M KCl (pH 5.5) (100 p. per well).Desorbed bacteria were plated on sucrose (4%) agar me-dium. The typical production of exopolysaccharides (levan)by B. polymyxa colonies makes easy their capture. Counts ofbacterial populations were expressed per gram (dry weight)of root or soil (60°C, 72 h).
Bacterial strains and media. Type strains of B. polymyxa(ATCC 842), Bacilllus inacerans (ATCC 8244), and Bacilluscirculans (NCIB 9374) and B. polymnyxa CF43 isolated fromthe rhizosphere of spring wheat in our laboratory were usedas reference strains. Modified Luria-Bertani broth (0.1%[wt/vol] tryptone, 0.5% yeast extract, 0.5% NaCl) was usedas the bacterial growth medium, and nutrient agar (Prolabo)was used to estimate the culturable microflora.
Identification and numerical analysis of phenotypic fea-tures. A total of 63 characters were used to identify anddetermine similarity among the strains of B. polymnyxa. Theproduction of acid metabolites from 49 carbohydrates (API50 CHB) and tests such as the proteolysis of gelatin; activ-ities of nitrate reductase, 3-galactosidase, urease, and tryp-tophanase; H2S formation; and the production of acetoin andcitrate utilization (API 20B) were used (28). The tests weredone in accordance with the recommendations of API-bioMerieux (la Balme les Grottes, Montalieu Vercieu,France). Discs impregnated with dimethyl-p-phenylenedi-amine (API-bioMerieux) were used to test for the presenceof cytochrome oxidase. The presence of catalase was tested
TABLE 1. Origins and designations of B. polyrnyxa strainsisolated by immunotrapping from the NRS, the RS, and theRP of 18-day-old spring wheat (T. aestivumn L., cv. Fidel)
PMD strains from:
Sample NRS RS RP
(n = 32)" (n = 38) (n = 60)
1 51, 52, 57, 66, 100, 101, 102, 112, 113, 200 to 21967, 69, 70, 75, 120, 121, 123, 124,76 127, 134
2 58, 59, 60, 61, 103, 105, 106, 114, 116, 220 to 23977, 78, 79, 80, 117, 118, 119, 122,81, 82 125, 128, 133
3 62, 63, 64, 65, 104, 107, 108, 109, 110, 240 to 25983, 84, 85, 86, 111, 115, 126, 129,87, 90, 91, 92, 130, 131, 132, 135,93 136, 137
"n, number of strains.
by adding a drop of 1.5% H202 to a young colony of B.polymyxa growing on nutrient agar; gas production (02)indicated the presence of the enzyme. The Gram stain testwas carried out in accordance with Cerny's enzymaticmethod (7). Cell shape, motility in a 24-h-old Luria-Bertanibroth culture, and the presence of swelling endospores in2-day-old colonies on nutrient agar were examined by use ofphase-contrast microscopy.Data for phenotypic features were submitted to a cluster
analysis by use of the APILAB computer program. Proxim-ity was measured with Gower's similarity coefficient withnegative matches (17). The clusters were analyzed by use ofthe unweighted pair group method with averages (47).
Serological testing. Species-specific PcAbs were raised inrabbits by hyperimmunization with whole cells of B. poly-myxa CF43 (A. Dorier, Biologie Appliqu6e, Institut Univer-sitaire de Technologie, Lyon, France) and then partiallypurified by precipitation with ammonium sulfate (3.9 M) anddialysis against phosphate-buffered saline (0.80% [wt/vol]NaCl, 0.27% Na2HPO4. 12H20, 0.40% NaH2PO4- 2H20[pH 7.4]). PcAbs were stored at -20°C until used. Theindirect enzyme-linked immunosorbent assay (ELISA) was
conducted as previously described with microtiter plates(Nunc Immuno-plate, Maxisorp F96) (4). A 10-4 dilution ofPcAbs was used. The bacterial concentration was main-tained at 10' to 106' cells per well (100 p.l), since thehomologous reaction (CF43-anti-CF43) was maximum atthis bacterial concentration. The immunoenzymatic reactionwas expressed as the A40- (Dynatech MR 5000 spectropho-tometer). Data for serological properties were log trans-formed to fit the normal distribution by use of the Kolmog-orov-Smirnov one-sample test (22). The variability of thedata was evaluated by use of Bartlett's test and coefficientsof variation (45) on the STATGRAPHICS software com-
puter program (Release 4.2; Uniware STSC, Inc.).Preparation of genomic DNA. Bacterial cultures were
grown overnight at 30°C in 50 ml of Luria-Bertani broth.Aliquots of 25 ml were pelleted at 9,000 x g for 20 min andstored at -20°C until used. The pelleted cells were washedonce in 1 ml of TES buffer (20 mM Tris, 50 mM EDTA, 150mM NaCI [pH 7.9]), centrifuged for 2 min at 9,000 x g, anddissolved in 1.2 ml of TES buffer. The cells were lysed bytreatment with 300 p. of lysozyme (5 mg ml-') (EC 3.2.1.17;Sigma L 6876) and incubation at 37°C for 60 min. Then, 90 p.l
Vol.. 58, 1992
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.1896 MAVINGUI ET AL.
PMD254 PD5PMD247 PMD205PMD252 PMD245PMD215 PMD245PMD227 PMD232PMD234 PMD232PMD253 PMD256PMD240 PMD224PMD223 PMD224PMD208 PMD233PMD239 PMD248PMD241 PMD244PMD243 PMD242PMD231 PMD229PMD228 PD2PMD213 PMD220PMD209 PMD212PMD206 PMD207PM2O PMvD204
PMD200 PMD201PMD249 PMD246PMD249 PMD257PMD202 PMD225PMD210 PMD211PMD23 PMD255PMD236 PMD230PMD226 PMD214PMD251PMDIOPMDIOOPMD132
PMD259 PMD937PMD115 PMD110PMD238 PMD219PMD107 PMD102PMD80 PMD78PMD122 PNM130PMD128 PMD123PMD135 PMD117PMD109 PD0PMD101 PMD111PMD136 PMD129PMD119 PMD112PMD137 PMD124PMD113 PMD105PMD103 PMD77PMD133 PMD125PMD127 PNM1181
PMD134PM8PMD116 PMD85PMD57 PMD114PMD79 PMD814PMD237 PMD812PMD82 PMD631PMD61 PMD60PMD52 PMD218PMD86 PMD628PMD131 PMD65PMD64 PMD91PMD92PMD104 PMD75PMD100 PMD126PMD84 PNM76PMD87 PMD66PMD59 PMD67PMD58 PMDI20PMDS1PMD70PMDOO
Group (%) strains per group
NRS RS RP
IV (0) (0) (100)
i (11.5) (77) (11.5)
]El (68) (23) (9)
I (67) (33) (0)
90 95 100% SIMILARITY
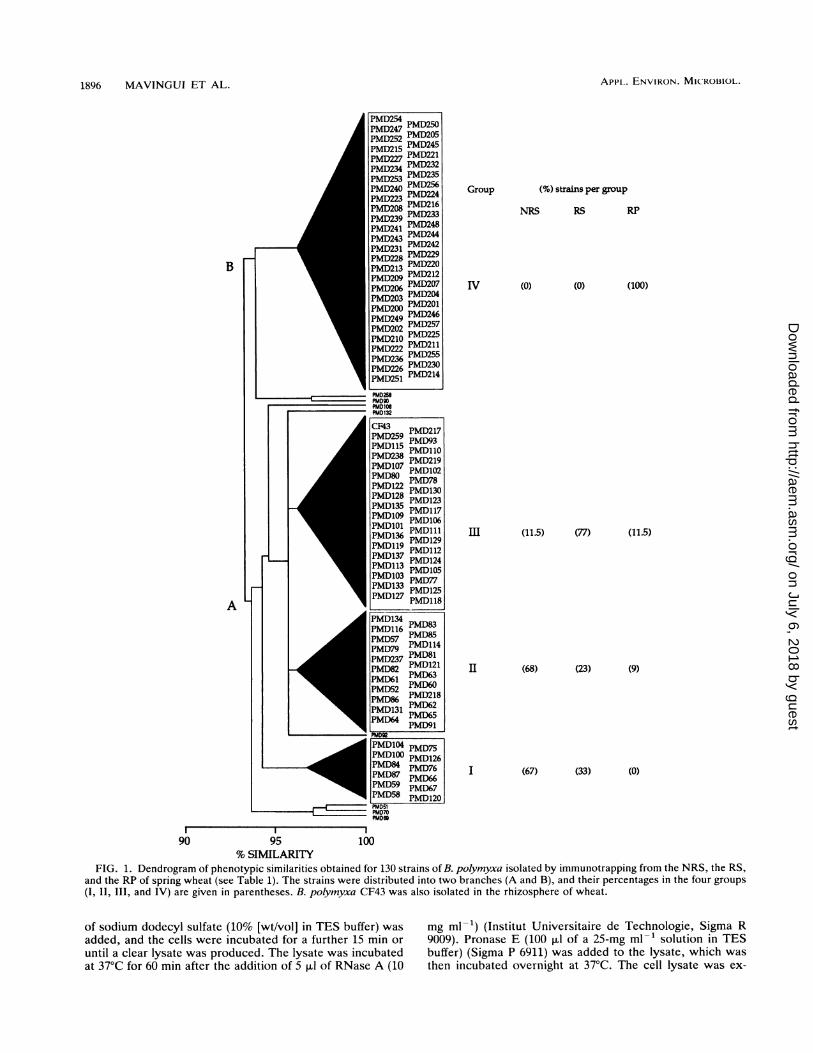
FIG. 1. Dendrogram of phenotypic similarities obtained for 130 strains of B. polymyxa isolated by immunotrapping from the NRS, the RS,and the RP of spring wheat (see Table 1). The strains were distributed into two branches (A and B), and their percentages in the four groups(I, II, III, and IV) are given in parentheses. B. polymyxa CF43 was also isolated in the rhizosphere of wheat.
of sodium dodecyl sulfate (10% [wt/vol] in TES buffer) wasadded, and the cells were incubated for a further 15 min or
until a clear lysate was produced. The lysate was incubatedat 37°C for 60 min after the addition of 5 p.l of RNase A (10
mg ml-1) (Institut Universitaire de Technologie, Sigma R9009). Pronase E (100 p.l of a 25-mg ml-' solution in TESbuffer) (Sigma P 6911) was added to the lysate, which was
then incubated overnight at 37°C. The cell lysate was ex-
B
A
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
B. POLYMYXA FROM SOIL AND RHIZOSPHERE 1897
A4052.5
2.0 -
1.5 -
1.0
0.5 -
0.0 -
55.8 6.0 6.2 6.4 6.6 6.8 7.0 7.2
log N cells per ml
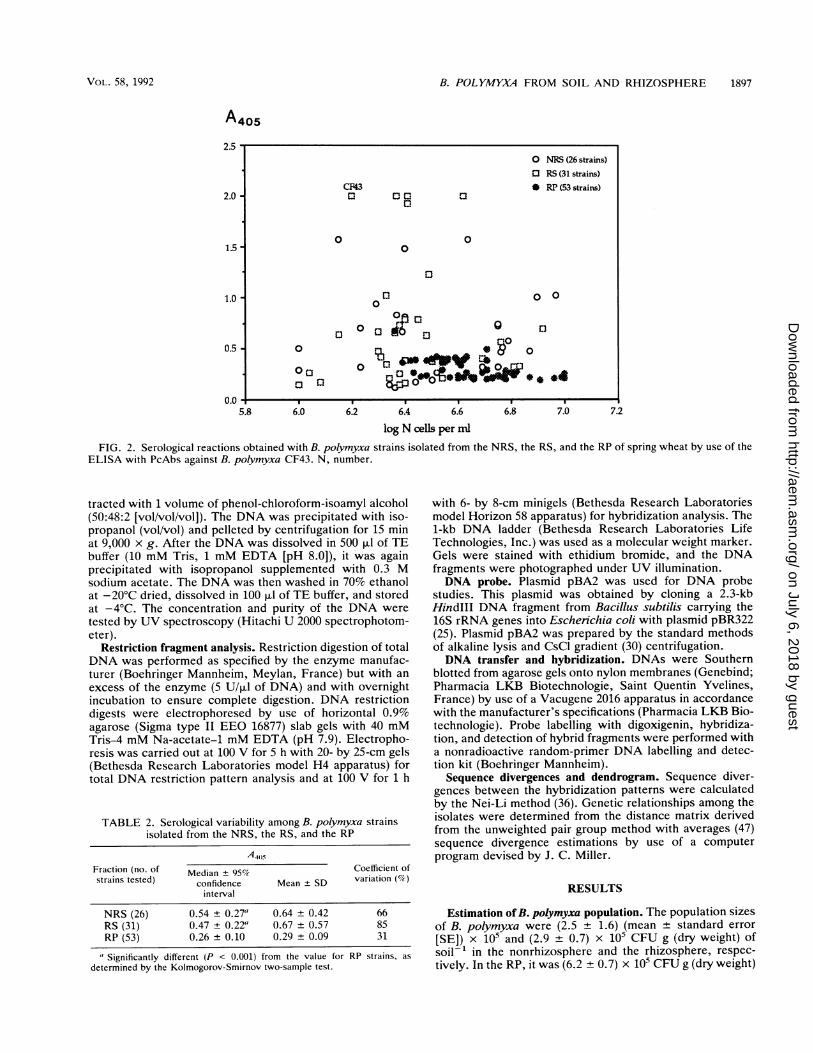
FIG. 2. Serological reactions obtained with B. polymyxa strains isolated from the NRS, the RS, and the RP of spring wheat by use of theELISA with PcAbs against B. polymyxa CF43. N, number.
tracted with 1 volume of phenol-chloroform-isoamyl alcohol(50:48:2 [vol/vol/vol]). The DNA was precipitated with iso-propanol (vol/vol) and pelleted by centrifugation for 15 minat 9,000 x g. After the DNA was dissolved in 500 Vl of TEbuffer (10 mM Tris, 1 mM EDTA [pH 8.0]), it was againprecipitated with isopropanol supplemented with 0.3 Msodium acetate. The DNA was then washed in 70% ethanolat -20°C dried, dissolved in 100 ,ul of TE buffer, and storedat -4°C. The concentration and purity of the DNA weretested by UV spectroscopy (Hitachi U 2000 spectrophotom-eter).
Restriction fragment analysis. Restriction digestion of totalDNA was performed as specified by the enzyme manufac-turer (Boehringer Mannheim, Meylan, France) but with anexcess of the enzyme (5 U/,l of DNA) and with overnightincubation to ensure complete digestion. DNA restrictiondigests were electrophoresed by use of horizontal 0.9%agarose (Sigma type II EEO 16877) slab gels with 40 mMTris-4 mM Na-acetate-1 mM EDTA (pH 7.9). Electropho-resis was carried out at 100 V for 5 h with 20- by 25-cm gels(Bethesda Research Laboratories model H4 apparatus) fortotal DNA restriction pattern analysis and at 100 V for 1 h
TABLE 2. Serological variability among B. polymyxa strainsisolated from the NRS, the RS, and the RP
A405Fraction (no. of Median + 95% Coefficient ofstrains tested) confidence Mean + SD variation (%)
interval
NRS (26) 0.54 ± 0.27a 0.64 ± 0.42 66RS (31) 0.47 ± 0.22a 0.67 ± 0.57 85RP (53) 0.26 + 0.10 0.29 ± 0.09 31
Significantly different (P < t).001) from the value for RP strains, asdetermined by the Kolmogorov-Smirnov two-sample test.
with 6- by 8-cm minigels (Bethesda Research Laboratoriesmodel Horizon 58 apparatus) for hybridization analysis. The1-kb DNA ladder (Bethesda Research Laboratories LifeTechnologies, Inc.) was used as a molecular weight marker.Gels were stained with ethidium bromide, and the DNAfragments were photographed under UV illumination.DNA probe. Plasmid pBA2 was used for DNA probe
studies. This plasmid was obtained by cloning a 2.3-kbHindIII DNA fragment from Bacillus subtilis carrying the16S rRNA genes into Escherichia coli with plasmid pBR322(25). Plasmid pBA2 was prepared by the standard methodsof alkaline lysis and CsCl gradient (30) centrifugation.DNA transfer and hybridization. DNAs were Southern
blotted from agarose gels onto nylon membranes (Genebind;Pharmacia LKB Biotechnologie, Saint Quentin Yvelines,France) by use of a Vacugene 2016 apparatus in accordancewith the manufacturer's specifications (Pharmacia LKB Bio-technologie). Probe labelling with digoxigenin, hybridiza-tion, and detection of hybrid fragments were performed witha nonradioactive random-primer DNA labelling and detec-tion kit (Boehringer Mannheim).
Sequence divergences and dendrogram. Sequence diver-gences between the hybridization patterns were calculatedby the Nei-Li method (36). Genetic relationships among theisolates were determined from the distance matrix derivedfrom the unweighted pair group method with averages (47)sequence divergence estimations by use of a computerprogram devised by J. C. Miller.
RESULTS
Estimation of B. polymyxa population. The population sizesof B. polymyxa were (2.5 + 1.6) (mean + standard error
[SE]) x 105 and (2.9 0.7) x 105 CFU g (dry weight) of
soil-' in the nonrhizosphere and the rhizosphere, respec-tively. In the RP, it was (6.2 + 0.7) x 105 CFU g (dry weight)
O NRS (26 strains)o1 RS (31 strains)
CF43 * RP (53 strains)M C C. Q
0 00
0
0
6
00 pi a
o 0 jD° co O- D°n~~~~~ ° _Evs
VOL. 58, 1992
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
1898 MAVINGUI ET AL.
A al a2 al a6 c2 a3a4 a5 al b 3b4 b5. E .. . .. .... M . .... R\,
lt,
0: X,,,"i: -00
S: *; <Xls 000f:ttt 0 t :'Ds ' DE D o75.:fNF DA'si^y ............................................. -' : 0DE t ' - : ot we r. wis - :iH}* ++0s0v?z=^( Z:d f..h:Z ^w>qa4te tE;ZZ*Xp---w Sso-w- wtwfA00 0 0 00 : 0.; 0 .. ;00 H . f.7 :S S3: ':f
iS fftVUi;) fiCk;0" 000#' XffsCt 's.
000 ,00 05
B a e b2 bi c o f j h a i g
aib1 b5 b2 c4c1c2c3a2 d e f g(1) (2) (3) (4)
9.26.1 -EEt*-tE3.33-..4.1 -
c d
",> to;-,*
..,:2
410-*
0tT"t X~ 'K I
a k I rl n I b2 0 p q r(1) (2) (3) (4)
12.2 -
9.2 -
6.1 -
4.1
2.1 -
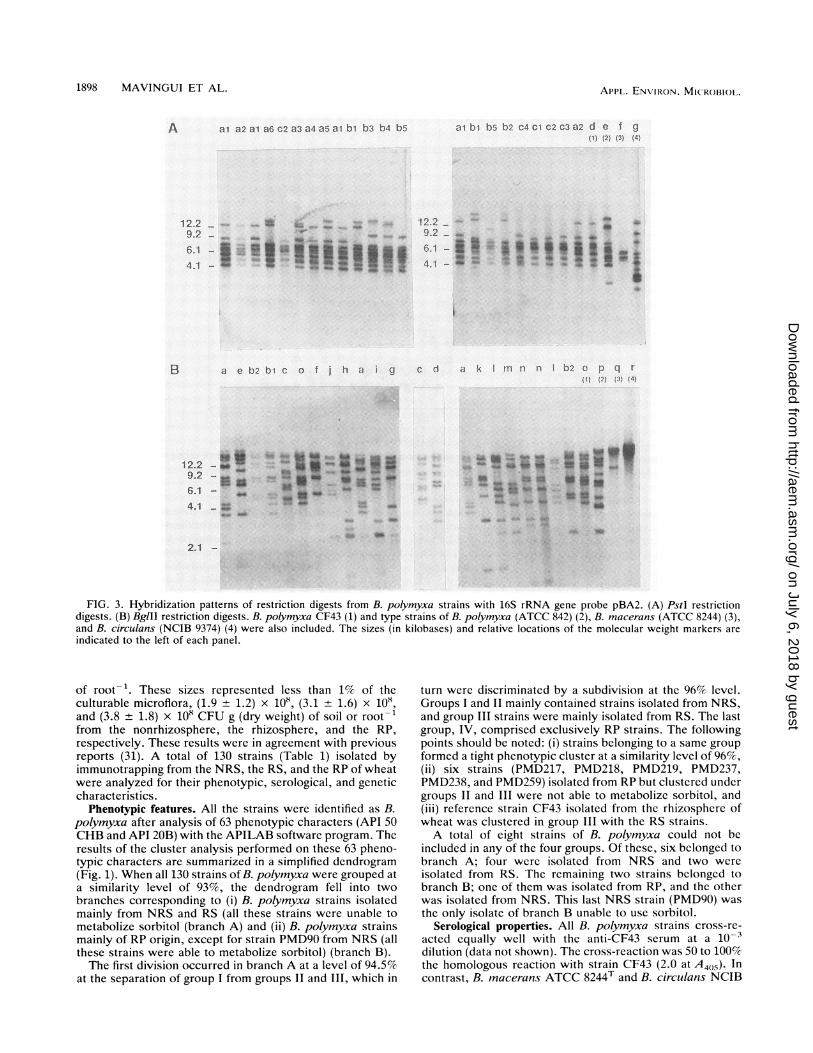
FIG. 3. Hybridization patterns of restriction digests from B. polymyxa strains with 16S rRNA gene probe pBA2. (A) PstI restrictiondigests. (B) BglII restriction digests. B. polymyxa CF43 (1) and type strains of B. polymyxa (ATCC 842) (2), B. macerans (ATCC 8244) (3),and B. circulans (NCIB 9374) (4) were also included. The sizes (in kilobases) and relative locations of the molecular weight markers are
indicated to the left of each panel.
of root-'. These sizes represented less than 1% of theculturable microflora, (1.9 + 1.2) x 108, (3.1 + 1.6) x 108,and (3.8 + 1.8) x 108 CFU g (dry weight) of soil or root-1from the nonrhizosphere, the rhizosphere, and the RP,respectively. These results were in agreement with previousreports (31). A total of 130 strains (Table 1) isolated byimmunotrapping from the NRS, the RS, and the RP of wheatwere analyzed for their phenotypic, serological, and geneticcharacteristics.
Phenotypic features. All the strains were identified as B.polymyxa after analysis of 63 phenotypic characters (API 50CHB and API 20B) with the APILAB software program. Theresults of the cluster analysis performed on these 63 pheno-typic characters are summarized in a simplified dendrogram(Fig. 1). When all 130 strains of B. polymyxa were grouped ata similarity level of 93%, the dendrogram fell into twobranches corresponding to (i) B. polymyxa strains isolatedmainly from NRS and RS (all these strains were unable tometabolize sorbitol (branch A) and (ii) B. polymyxa strainsmainly of RP origin, except for strain PMD90 from NRS (allthese strains were able to metabolize sorbitol) (branch B).The first division occurred in branch A at a level of 94.5%
at the separation of group I from groups II and III, which in
turn were discriminated by a subdivision at the 96% level.Groups I and II mainly contained strains isolated from NRS,and group III strains were mainly isolated from RS. The lastgroup, IV, comprised exclusively RP strains. The followingpoints should be noted: (i) strains belonging to a same groupformed a tight phenotypic cluster at a similarity level of 96%,(ii) six strains (PMD217, PMD218, PMD219, PMD237,PMD238, and PMD259) isolated from RP but clustered undergroups II and III were not able to metabolize sorbitol, and(iii) reference strain CF43 isolated from the rhizosphere ofwheat was clustered in group III with the RS strains.A total of eight strains of B. polymyxa could not be
included in any of the four groups. Of these, six belonged tobranch A; four were isolated from NRS and two wereisolated from RS. The remaining two strains belonged tobranch B; one of them was isolated from RP, and the otherwas isolated from NRS. This last NRS strain (PMD90) wasthe only isolate of branch B unable to use sorbitol.
Serological properties. All B. polymyxa strains cross-re-acted equally well with the anti-CF43 serum at a 10-3dilution (data not shown). The cross-reaction was 50 to 100%the homologous reaction with strain CF43 (2.0 at A405). Incontrast, B. macerans ATCC 8244T and B. circulans NCIB
12.29.26.14.1
Appt-. ENVIRON. MICROBIOL.
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. POLYMYXA FROM SOIL AND RHIZOSPHERE 1899
9374T did not cross-react. To investigate the serologicalvariability among B. polymyxa strains, we used a 10-4dilution of serum. The serological properties measured bythe ELISA revealed heterogeneity among the strains tested(Fig. 2). The variances were not homogeneous within eachfraction, namely, NRS, RS, and RP, as revealed by Bart-lett's test (P < 0.05), indicating the presence of at least two
distinct statistical populations. The nonparametric Kolmog-orov-Smirnov two-sample test (22) revealed that the strainsisolated from NRS and RS with similar medians formed a
distinct serological subgroup different (P < 0.001) from thesubgroup of RP strains (Table 2). In addition, the strainsisolated from the former two fractions (NRS and RS) were
more variable serologically than the RP strains, as indicatedby their higher coefficients of variation (Table 2).RFLP analysis. From an initial sample of 130 B. polymyxa
strains, a subsample of 36 strains (12 each from the NRS,RS, and RP fractions) represented in the phenotypic groups
described in Fig. 1 was used for restriction fragment lengthpolymorphism (RFLP) analysis. The results of the pheno-typic cluster analysis performed on these 36 strains were inagreement with the cluster analysis results obtained when130 strains were used (data not shown). In addition, incor-poration of type strains in the subsample cluster analysisshowed that B. polymyxa ATCC 842T was related to theclusters formed by B. polymyxa strains at a similarity level of94%. In contrast, B. macerans ATCC 8244T and B. circulansATCC 9374T were more distant (<88% similarity).The hybridization patterns of total DNAs from the 36
strains are shown in Fig. 3. B. polymyxa ATCC 842T sharedseveral hybridizing restriction fragments in common withother B. polymyxa strains, but B. macerans ATCC 8244T andB. circulans NCIB 9374T displayed distinctly different pro-
files. The hybridization patterns revealed greater heteroge-neity among the strains isolated from NRS and RS thanamong those isolated from RP (Table 3). Ten and 9 differentprofiles were seen with NRS and RS strains, respectively. Incontrast, with strains isolated from RP, there were onlythree profiles with PstI digests and two profiles with BglIIdigests. Five profiles appeared common to the strains iso-lated from NRS and RS. However, most of the strainsisolated from RP (10 of 12) had one profile (Table 3). Theremaining two RP strains (PMD218 and PMD259) were
unable to metabolize sorbitol and were also differentiatedfrom the other RP strains in phenotypic clustering (Fig. 1). Itshould be noted that two NRS strains, PMD92 and PMD93,with identical hybridization profiles were, however, sepa-
rated phenotypically. Strain PMD63 isolated from NRS andB. polymyxa CF43 showed identical profiles.The global cluster analysis of hybridization patterns with
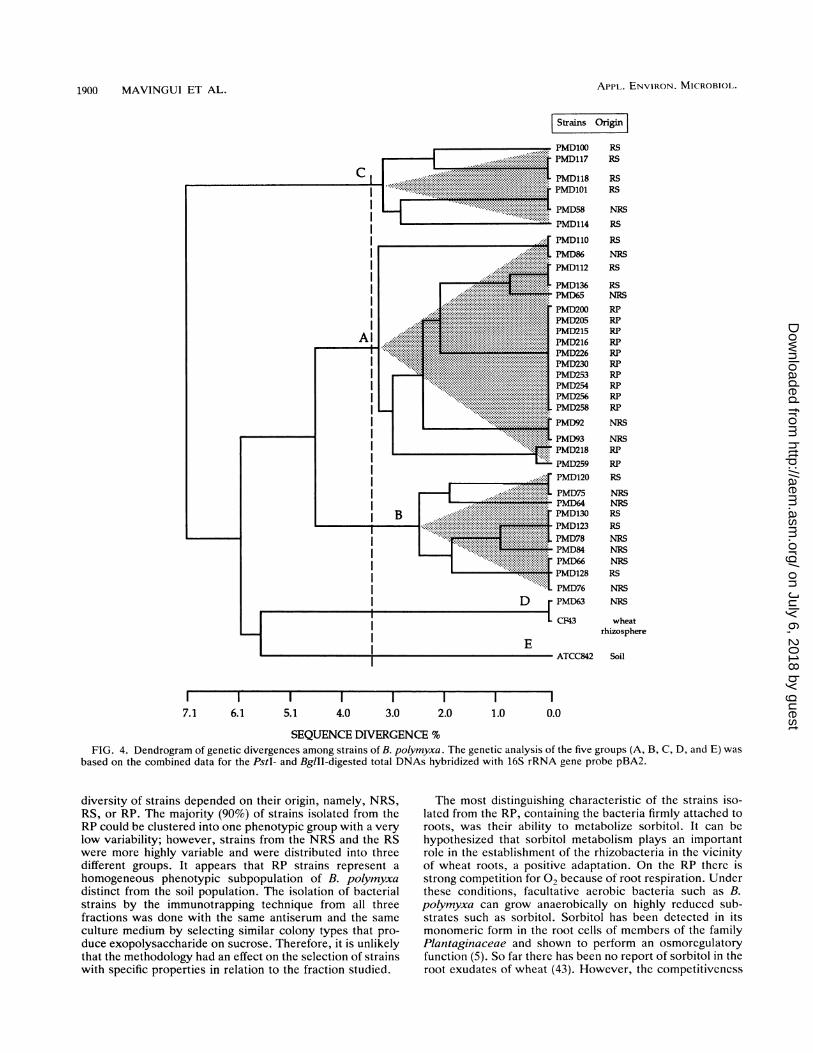
PstI and BglII generated five groups (A, B, C, D, and E) atthe genetic distance level of 3.6% (Fig. 4). The most distantgroup, C, was at the genetic distance level of 7.1%. Se-quence divergence among strains belonging to the same
group did not exceed 3.4%. All the strains isolated from RPwere clustered under group A, whereas the strains originat-ing from NRS and RS were distributed into four (A, B, C,and D) of the five groups. B. polymyxa CF43 belonged togroup D. B. polymyxa ATCC 842T constituted a distinctgroup, E, on its own. B. macerans ATCC 8244T and B.circulans NCIB 9374T were even more distant and thereforewere excluded from the analysis.An examination of total DNA HindlIl restriction patterns
confirmed the RFLP analysis based on hybridization withthe pBA2 probe. All the strains not differentiated by hybrid-ization pattern analysis were also not differentiated by their
TABLE 3. Distribution of RFLP pattern groups (a to p) andsubgroups (1 to 6) of B. polymyxa strains in the samples
pBA2hybridization
Genotypic pattern Strain' Fraction Sample"group' obtained with:
PstI BglII
A a, 200 RP 1a, a 205 RP 1a1 a 215 RP 1a, a 216 RP 1a, a 226 RP 2a1 a 230 RP 2a1 a 253 RP 3a1 a 254 RP 3a1 a 256 RP 3a1 a 258 RP 3a1 b1 65 NRS 3a2 b2 136 RS 3a2 b2 112 RS 1a3 c 259 RP 3a4 c 218 RP 1a. d 86 NRS 3a5 d 110 RS 3a6 e 92 NRS 3a6 e 93 NRS 3
B b1 f 120 RS 1b1 f 75 NRS 1b2 g 64 NRS 3b3 h 130 RS 3b3 h 123 RS 2b3 h 78 NRS 2b4 i 84 NRS 3b5 j 66 NRS 1b5 j 128 RS 2b5 j 76 NRS 1
C c1 k 100 RS 1c2 1 117 RS 2c2 1 118 RS 2C3 m 114 RS 2C4 n 101 RS 1C4 n 58 NRS 2
D d o 63 NRS 3d o CF43d Rhizosphere
E e p 842d Soil
a Derived from a cluster analysis of hybridization patterns with PstI andBglII (see Fig. 4).
h PMD strains, unless otherwise indicated.'*See Table 1.d CF43 is a B. polymyxa strain isolated from the rhizosphere of wheat; 842
is B. polymyxa ATCC 842w.
total DNA restriction patterns (Fig. 5). All the strains foundto be closely related by the hybridization pattern analysis(genetic distance, <1%) also shared identical or very similartotal DNA restriction patterns. On the whole, strainsgrouped by the genotypic cluster analysis showed similar-ities in their total DNA fingerprints.
DISCUSSIONThe cluster analysis performed on phenotypic features of
B. polymyxa strains isolated in this study showed that at thespecies level (i) the strains exhibited a high level of similarity(93%) and their identification was unambiguous and (ii) the
VOL. 58, 1992
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from
APPL. ENVIRON. MICROBIOL.1900 MAVINGUI ET AL.
Strains OriginPMD100 RSPMD117 RS
PMD118 RSPMD101 RS
PMD58 NRS
PMD114 RS
PMD11O RSPMD86 NRSPMD112 RS
PMD136 RSPMD65 NRSPMD200 RPPMD205 RPPMD215 RPPMD216 RPPMD226 RPPMD230 RPPMD253 RPPMD254 RPPMD256 RPPMD258 RP
PMD92 NRS
PMD93 NRSPMD218 RPPMD259 RPPMD120 RS
PMD75 NRSPMD64 NRSPMD130 RSPMD123 RSPMD78 NRSPMD84 NRSPMD66 NRSPMD128 RS
PMD76 NRSPMD63 NRS
CF43 wheatrhizosphere
ATCC842 Soil
I I I I I I 17.1 6.1 5.1 4.0 3.0 2.0 1.0 0.0
SEQUENCE DIVERGENCE %FIG. 4. Dendrogram of genetic divergences among strains of B. polymyxa. The genetic analysis of the five groups (A, B, C, D, and E) was
based on the combined data for the PstI- and BglII-digested total DNAs hybridized with 16S rRNA gene probe pBA2.
diversity of strains depended on their origin, namely, NRS,RS, or RP. The majority (90%) of strains isolated from theRP could be clustered into one phenotypic group with a verylow variability; however, strains from the NRS and the RSwere more highly variable and were distributed into threedifferent groups. It appears that RP strains represent a
homogeneous phenotypic subpopulation of B. polymyxadistinct from the soil population. The isolation of bacterialstrains by the immunotrapping technique from all threefractions was done with the same antiserum and the same
culture medium by selecting similar colony types that pro-duce exopolysaccharide on sucrose. Therefore, it is unlikelythat the methodology had an effect on the selection of strainswith specific properties in relation to the fraction studied.
The most distinguishing characteristic of the strains iso-lated from the RP, containing the bacteria firmly attached toroots, was their ability to metabolize sorbitol. It can behypothesized that sorbitol metabolism plays an importantrole in the establishment of the rhizobacteria in the vicinityof wheat roots, a positive adaptation. On the RP there isstrong competition for 02 because of root respiration. Underthese conditions, facultative aerobic bacteria such as B.polymyxa can grow anaerobically on highly reduced sub-strates such as sorbitol. Sorbitol has been detected in itsmonomeric form in the root cells of members of the familyPlantaginaceae and shown to perform an osmoregulatoryfunction (5). So far there has been no report of sorbitol in theroot exudates of wheat (43). However, the competitiveness
CI
AlI
,. .............
...... .. ...
...- .. ...-..--............
.... ................,.. ..
.....g.:.....-.....s..>
I..t.........
......... ....... .... ....-. ..
. ,-,...........
............
.....ss.....
... ....
......... ............
... ...... .......
...... .. .
...............
.......;s;
...-.-;sSsr.,.,.. ...s,'',,T i,,iiv.T,..:::. .,:.::.:. .
.:''.:S :--::::.:-. :.:.
E-1
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. POLYMYXA FROM SOIL AND RHIZOSPHERE 1901
A B D E C
0*C0 04 Or..v-
OW O" v 0 cv) U Yr T-Nv-WM N N r- - 0(-0 (0
FIG. 5. Agarose gel electrophoresis of HindlIl-digested totalDNAs from B. polymyxa strains. Numbers above lanes indicatestrain designations (see Table 1). A, B, C, D, and E correspond tothe groups indicated in Fig. 4.
of Rhizobium leguminosarum strains for pea nodulation hasbeen correlated with the ability of these strains to catabolizethe amino acid homoserine, found in large quantities in pearoot exudates (24). Monosaccharides such as inositol, glu-cose, glucuronic acid, galactose, galacturonic acid, arabi-nose, and fucose contained in the exudates of several plantshave been also shown to be specifically able to induce thevirulence genes of the plant pathogen Agrobacterium tume-faciens (1), leading to its proliferation in the plant tissue.
The serological variability of B. polymyxa strains was alsohigher among the strains isolated from NRS and RS thanamong RP strains. No cross-reaction was found with B.macerans ATCC 8244T and B. circulans NCIB 9374T, inaccordance with the results obtained with agglutination tests
by Porter et al. (37), although these organisms are relatedtaxonomically to B. polymyxa species (38). However, withB. polymyxa somatic antigens, cross-reactions with certainstrains of B. macerans have been reported (13). Little is
known about the serology of B. polymyxa. Porter et al. (37),using heated vegetative cells as antigens, found that no one
particular antiserum agglutinated all the strains of B. poly-myxa, although some antisera agglutinated 60% of the strains
tested, suggesting the presence of several serogroups. Our
results suggest that the strains of B. polymyxa isolated from
NRS and RS and from wheat roots belong to the same
serogroup. This suggestion could be based on the specificimmunological method used, which allows the isolation of
strains closely related serologically (23). However, the
strains of B. polymyxa isolated from the RP of wheat
represent a serological subgroup distinct from that of strains
isolated from the NRS and the RS. Several studies have
demonstrated that in the case of Bradyrhizobium-soybeansymbiosis, plant genotype can affect serogroup recovery and
the distribution of Bradyrhizobium japonicum in the soil (6,15). Moawad et al. (34) showed that a particular indigenousB. japonicum serogroup was found more frequently than two
other serogroups in nodules of field-grown soybeans. The
presence of a distinct subserogroup of B. polymyxa closelyassociated with wheat roots, i.e., the RP, may indicate
subpopulation selection from an existing heterogeneous res-ident soil population by the host.On the basis of the RFLP patterns, we found that the
strains isolated by immunotrapping are related to B. poly-myxa ATCC 842T. In contrast, B. macerans ATCC 8244Tand B. circulans NCIB 9374T are clearly distant species.These results confirm the findings based on phenotypic andserological characteristics. Nonprobed and probed RFLPpatterns of genes coding for rRNA, used as a taxonomic toolfor microbial classification, have been reported (2, 19, 48).Stahl et al. (48) demonstrated that the separation of Lacto-bacillus strains into two different species by scanned RFLPpatterns of total DNA was in agreement with DNA-DNAhybridization data. Species of the genus Bacillus have alsobeen separated by use of RFLP patterns of hybridizationbands of genes coding for rRNA (16). Our studies showedthat the RFLP patterns of the 10 strains isolated from the RPand able to catabolize sorbitol represented a single genotype.This dominant genotype from the vicinity of wheat roots (theRP) was not found among the 24 soil (nonrhizosphere andrhizosphere) isolates tested. It could be postulated that thisgenotype resulted from a clone. However, this possibilityseems unlikely because three independent RP samples(each sample containing four plants) were used for isolatingB. polymyxa strains (PMD200, PMD205, PMD215, andPMD216 from sample 1; PMD226 and PMD230 from sample2; and PMD253, PMD254, PMD256, and PMD258 fromsample 3) (Table 1). The genotype of RP strains that was notisolated could exist as a minor constituent of the soilmicrobial population. The polymerase chain reactionmethod (3) may be useful if the frequency of this genotype inthe soil is too low for its detection otherwise. However, allthe B. polymyxa strains from the RP were clustered in a
group (A) which also contained some strains from the NRSand the RS, suggesting that they are closely related geneti-cally. In contrast, strains isolated from the NRS and the RSwere clustered in four different groups (A, B, C, and D),indicating that genotypic heterogeneity was higher amongthe strains isolated from the NRS and the RS than amongthose isolated from the RP.
Despite the lack of correlation among the genetic subdivi-sions of subsample strains and the groups obtained byphenotypic features, these methods are sensitive indicatorsof diversity among the strains isolated from the NRS and theRS with respect to the relative homogeneity of the RPstrains. Torsvik et al. (50) have also shown that bothphenotypic and genetic measures provide a good estimate ofsoil bacterial diversity. Using this approach, we were able todifferentiate the subpopulation of B. polymyxa isolated fromthe RP from the NRS and RS subpopulations. It appears theroots of wheat select a specific phenotypic, serological, andgenotypic subpopulation of B. polymyxa, probably from thenatural soil populations. This finding suggests that, over thetime frame of this study, the diversity of the soil bacterialpopulation was lowered following selection by the plantroots. Whether successive interactions between roots andsoil microorganisms would have resulted in the stabilizationof the diversity of the overall soil population needs to beverified.
ACKNOWLEDGMENTS
We are grateful to N. Amarger (INRA, Dijon, France) forproviding the laboratory facilities and to J. P. W. Young (John InnesInstitute, Norwich, England) and P. Hebbar (CNRS, Nancy,France) for critical evaluations.
VOL. 58, 1992
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

1902 MAVINGUI ET AL.
REFERENCES
1. Ankenbauer, R. G., and E. W. Nester. 1990. Sugar-mediatedinduction ofAgrobacterium tumefaciens virulence genes: struc-tural specificity and activities of monosaccharides. J. Bacteriol.172:6442-6446.
2. Armstrong, J. L., N. L. Fowles, and P. T. Rygiewicz. 1989.Restriction fragment length polymorphisms distinguish ectomy-corrhizal fungi. Plant Soil 116:1-7.
3. Bej, A. K., S. C. McCarty, and R. M. Atlas. 1991. Detection ofcoliform bacteria and Escherichia coli by multiplex polymerasechain reaction: comparison with defined substrate and platingmethods for water quality monitoring. Appl. Environ. Micro-biol. 57:2429-2432.
4. Berge, O., T. Heulin, W. Achouak, C. Richard, R. Bally, and J.Balandreau. 1991. Rahnella aquatilis, a nitrogen-fixing entericbacterium associated with the rhizosphere of wheat and maize.Can. J. Microbiol. 37:195-203.
5. Briens, M., and F. Larher. 1983. Sorbitol accumulation inPlantaginaceae; further evidence for a function in stress toler-ance. Z. Pflanzenphysiol. 11OS:447-458.
6. Caldwell, B. E., and E. E. Hartwig. 1970. Serological distribu-tion of soybean root nodule bacteria in soils of south-easternU.S.A. Agron. J. 62:621-622.
7. Cerny, G. 1976. Method for the distinction of gram-negativebacteria from gram-positive bacteria. Eur. J. Appl. Microbiol.53:223-225.
8. Chanway, C. P., F. B. Holl, and R. Turkington. 1988. Genotypiccoadaptation in plant growth promotion of forage species byBacillus polymyxa. Plant Soil 106:281-284.
9. Chanway, C. P., and L. M. Nelson. 1990. Field and laboratorystudies of Triticum aestivum L. inoculated with co-existentgrowth-promoting Bacillus strains. Soil Biol. Biochem. 22:789-795.
10. Chanway, C. P., L. M. Nelson, and F. B. Holl. 1988. Cultivar-specific growth promotion of spring wheat (Tniticum aestivumL.) by coexistent Bacillus species. Can. J. Microbiol. 34:925-929.
11. Clark, F. E. 1949. Soil microorganisms and plant roots. Adv.Agron. 1:242-288.
12. Claus, D., and R. C. W. Berkeley. 1986. Genus Bacillus Cohn1872, 174AL, p. 1105-1139. In P. H. A. Sneath, N. S. Mair,M. E. Sharpe, and J. G. Holt (ed.), Bergey's manual of system-atic bacteriology, vol. 2. The Williams & Wilkins Co., Baltimore.
13. Davies, S. N. 1951. The serology of Bacillus polymyxa. J. Gen.Microbiol. 5:807-816.
14. Djordjevic, M. A., D. W. Gabriel, and B. G. Rolfe. 1987.Rhizobium-the refined parasite of legumes. Annu. Rev. Phy-topathol. 25:145-168.
15. Fuhrmann, J. 1989. Serological distribution of Bradyrhizobiumjaponicum as influenced by soybean cultivar and samplinglocation. Soil Biol. Biochem. 21:1079-1081.
16. Gottllieb, P., and R. Rudner. 1985. Restriction site polymor-phism of ribosomal ribonucleic acid gene sets in members of thegenus Bacillus. Int. J. Syst. Bacteriol. 35:244-252.
17. Gower, J. C. 1971. A general coefficient of similarity and some
of its properties. Biometrics 27:857-874.18. Graham, J. B., and C. A. Istock. 1979. Gene exchange and
natural selection cause Bacillus subtilis to evolve in soil culture.Science 204:637-639.
19. Grimont, F., and P. A. D. Grimont. 1986. Ribosomal ribonucleicacid restriction patterns as potential taxonomic tools. Ann. Inst.Pasteur/Microbiol. 137:165-175.
20. Hino, S., and P. W. Wilson. 1958. Nitrogen fixation by a
facultative Bacillus. J. Bacteriol. 75:403-408.21. Holl, F. B., C. P. Chanway, R. Turkington, and R. A. Radley.
1988. Response of crested wheatgrass (Agropyron cristatum
L.), perennial ryegrass (Lolium perenne L.) and white clover(Trifolium repens L.) to inoculation with Bacillus polymyxa.Soil Biol. Biochem. 20:19-24.
22. Hollander, M., and D. A. Wolfe. 1973. Nonparametric statisticalmethods. John Wiley & Sons, Inc., New York.
23. Hranitzky, K. W., A. D. Larson, D. W. Ragsdale, and R. J.
Siebeling. 1980. Isolation of 10 serovars of Vibrio cholerae fromwater by a serologically specific method. Science 210:1025-1026.
24. Hynes, M. F., and M. P. O'Connell. 1990. Host plant effect oncompetition among strains of Rhizobium leguminosarum. Can.J. Microbiol. 36:864-869.
25. Iglesis, A., P. Ceglowski, and T. A. Trautner. 1983. Plasmidtransformation in Bacillus subtilis. Effects of the insertion ofBacillus subtilis rRNA genes into plasmids. Mol. Gen. Genet.192:149-155.
26. Kennedy, C. 1989. The genetics of nitrogen fixation, p. 107-127.In D. A. Hopwood and K. F. Chater (ed.), Genetics of bacterialdiversity. Academic Press, Inc., New York.
27. Lindberg, T., and U. Granhall. 1984. Isolation and characteri-zation of dinitrogen-fixing bacteria from the rhizosphere oftemperate cereals and forage grasses. Appl. Environ. Microbiol.48:683-689.
28. Logan, N. A., and R. C. W. Berkeley. 1984. Identification ofBacillus strains using the API system. J. Gen. Microbiol.130:1871-1882.
29. Lynch, J. M. 1990. Beneficial interactions between micro-organisms and roots. Biotechnol. Adv. 8:335-346.
30. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
31. Mavingui, P., 0. Berge, and T. Heulin. 1990. Immunotrapping ofBacillus polymyxa in soil and in the rhizosphere of wheat.Symbiosis 9:215-221.
32. Metzger, W. C., D. A. Klein, and E. F. Redente. 1986. Bacterialphysiological diversity in the rhizosphere of range plants inresponse to retorted shale stress. Appl. Environ. Microbiol.52:765-770.
33. Miller, H. J., G. Henken, and J. A. Van Veen. 1989. Variationand composition of bacterial populations in the rhizospheres ofmaize, wheat, and grass cultivars. Can. J. Microbiol. 35:656-660.
34. Moawad, H. A., W. R. Ellis, and E. L. Schmidt. 1984. Rhizo-sphere response as a factor in competition among three sero-groups of indigenous Rhizobium japonicum for nodulationof field-grown soybeans. Appl. Environ. Microbiol. 47:607-612.
35. Neal, J. L., and R. I. Larson. 1976. Acetylene reduction bybacteria isolated from the rhizosphere of wheat. Soil Biol.Biochem. 8:151-155.
36. Nei, M., and W. H. Li. 1979. Mathematical model for studyinggenetic variation in terms of restriction endonucleases. Proc.Natl. Acad. Sci. USA 76:5269-5273.
37. Porter, R., C. S. McCleskey, and M. Levine. 1937. The faculta-tive sporulating bacteria producing gas from lactose. J. Bacte-riol. 33:163-183.
38. Priest, F. G., M. Goodfellow, and C. Todd. 1981. The genusBacillus: a numerical analysis, p. 91-103. In R. C. W. Berkeleyand M. Goodfellow (ed.), The aerobic endospore-forming bac-teria: classification and identification. Academic Press, Inc.,New York.
39. Rennie, R. J., and R. I. Larson. 1979. Dinitrogen fixationassociated with disomic substitution lines of spring wheat. Can.J. Bot. 57:2771-2775.
40. Riviere, J. 1963. Rhizosphere et croissance du ble. Ann. Agron.14:619-653.
41. Rouatt, J. W., and H. Katznelson. 1961. A study of the bacteriaon the root surface and in the rhizosphere soil of crop plants. J.Appl. Bacteriol. 24:164-171.
42. Rouatt, J. W., H. Katznelson, and T. M. B. Payne. 1960.Statistical evaluation of rhizosphere effect. Soil Sci. Soc. Am.Proc. 24:271-273.
43. Rovira, A. D. 1969. Plant root exudates. Bot. Rev. 5:35-57.44. Rovira, A. D., and C. B. Davey. 1974. Biology of the rhizo-
sphere, p. 153-204. In E. W. Carson, Jr. (ed.), The plant rootand its environment. University Press of Virginia, Charlottes-ville.
45. Scherrer, B. 1984. Les compraisons de moyennes, p. 401-463.In G. Morin (ed.), Biostatistique. Chicoutimi, Quebec, Canada.
APPL. ENVIRON. MICROBIOL.
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from

B. POLYMYXA FROM SOIL AND RHIZOSPHERE 1903
46. Silverman, M., M. Martin, and J. Engebrecht. 1989. Regulationof luminescence in marine bacteria, p. 71-86. In D. A. Hopwoodand K. F. Chater (ed.), Genetics of bacterial diversity. Aca-demic Press, Inc., New York.
47. Sneath, P. H. A., and R. R. Sokal. 1973. Numerical taxonomy.W. H. Freeman & Co., San Francisco.
48. Stahl, M., G. Molin, A. Persson, S. Ahrne, and S. Stahl. 1990.Restriction endonuclease patterns and multivariate analysis as a
classification tool for Lactobacillus spp. Int. J. Syst. Bacteriol.40: 189-193.
49. Torsvik, V., J. Goksoyr, and F. L. Daae. 1990. High diversity inDNA of soil bacteria. Appl. Environ. Microbiol. 56:782-787.
50. Torsvik, V., K. Salte, R. Sorheim, and J. Goksoyr. 1990.Comparison of phenotypic diversity and DNA heterogeneity ina population of soil bacteria. Appl. Environ. Microbiol. 56:776-781.
VOL. 58, 1992
on July 6, 2018 by guesthttp://aem
.asm.org/
Dow
nloaded from