genetic biochemistry
TRANSCRIPT

.2010 – 2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
18.10 – 1הרצאה –ביוכימיה
3במהלך הסמסטר נעסוק ב. קריאה לנושא ביולוגיה מולקולריתחומר –שקף ראשון במצגת :נושאים עיקריים
(. ר דון"ד)תהליכי שעתוק ושכפול , DNAמבנה , ביולוגיה מולקולארית -
(.ר הס"ד)קואופרטיביות של חלבונים , קיפול חלבונים -
(. ר כהן"ד)של אנזימים ' קינטיקה וכו, פעילות, הפעלה –אנזימולוגיה -
:השאלות בהן עוסקת הביוכימיה
?מהם מבני המולקולות בעלות החשיבות בביולוגיה .1
?וכיצד הן מתרחשות, מהן האינטראקציות המתקיימות בין מולקולות אלו .2
?כיצד מולקולות ביולוגיות נוצרות ומפורקות .3
?וכיצד התא משתמש בה, מיוצרת אנרגיה בתאכיצד –מאזן האנרגיה בתא .4
?מהם המנגנונים המארגנים את המולקולות הביולוגיות שבמבנה התא .5
?מועבר ומבוטא, איך המידע הגנטי מאוחסן .6
:הקדמה לקורס
:י מולקולות ביולוגיות"ישנם שני סוגי קישור המקוימים ע
י קשר "נוצר ע DNA)דות של תת יחי קישור קוולנטיי "הנוצרות ע, מולקולות לינאריות .1כך גם . י קשר קוולנטי בין חומצות אמינו"חלבונים נוצרים ע, קוולנטי בין נוקליאוטידים
(. ושומנים פוליסכרידים
המאפשרים קיום קשרי מימן או קישור בין מולקולרי אחר – קישור לא קוולנטי .2ככל שקשרים אלו חזקים יותר כך המערכת יציבה . קונפורמציות ומבנים תלת ממדיים
הם , םיהרבה יותר חלשים מאשר הקשרים הקוולנטיתמיד אומנם מאחר והם , יותרדוגמא לאינטראקציה לא . גמישים יותר וניתנים לפירוק ולארגון מחדש בקלות רבה יותר
גר בזמן המאפשרים להם להיפתח ולהיס, DNAהיא קשרי המימן שבין גדילי הקוולנטית אשר – מתבטאת בפעילותם של פקטורי השעתוק דוגמא נוספת. תהליכי שכפול ושעתוק
.DNAהם חלבונים המקיימים אינטראקציות לא קוולנטיות עם גדילי ה
בציטופלזמה אז אם –המולקולות הביולוגיות והתהליכים הביולוגיים מתרחשים במדיום מסוים ממולקולות אז במדיום המורכב ואם בממברנה, (ים הידרופילמדיו)מדובר בתמיסה מימית
(. 'כולסטרול וכד, פוספוליפידים)בי ומדיום הידרופ, שומניות
. הם טעון שליליתמורכבת משני גדילים אשר כל אחד מ שתימצא בתוך הגרעין DNAמולקולת למרות המטען הזהה שיש , ולא דוחים זה את זה, ידוע כי הגדילים קשורים זה לזה בנוסף
.לשניהם
תופעת הוא ש (אך יכול להתאים במקרה דומה, DNAשאינו מתאים למולקולת ה) אפשרי הסבר
גורמת לכך שיונים שליליים מתחרים במולקולות השליליות על הקישור למולקולות salting inהזה סברבכדי לדעת אם ה. נצמדות זו לזוכך שהמולקולות השליליות שוקעות ובתוך כך –המים
או למומסים אחרים , עלינו להכיר את היחס שבין המומס לממס הקיים בתמיסה מתאים . שבתמיסה
כך , מחוברות לחלבונים הטעונים חיובית הן, טעונות שלילית למרות שהן – DNAמולקולות ה גביל
. של אנרגיה אדירהמבלי להשקיע כמות DNAר של שני גדילי שמתאפשר חיבו

.2010 – 2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
מדובר . אחד הכוחות הלא קוולנטיים המרכזיים הפועלים במערכות הביולוגיות הוא קשרי המימןוך אטום הוא נשאר בת, אלקטרון מבלי לתרום אותו" תורם)"המימן האלקטרו חיובי בקישור בין
י "ליסוד אלקטרו שלילי המושפע ע( ו החנקןה על החמצן אהמימן אך משפיע בצורה חזק .האלקטרון שבמימן
לא רק עצם (. וחומרים דומים)והם אחראיים לכל תכונות המים , קשרי המימן הם חשובים מאודאם קיימת .עצמהחשיבות רבה בפני תהקישור בעל הכיווניות שלאלא גם , קיום הקשר חשוב
עובדה זו אחראית . חוזק הקשר הוא הגדול ביותר –זווית שטוחה בין שני אטומים שבקשר מימן קשרי מימן בין 4הקפאת המים גורמת להיווצרות –לאנומליה של המים במעבר מנוזל למוצק
כך שהחומר –( דבר אשר גורם למרווחים גדולים בין המולקולות)מולקולות בזוויות שטוחות . קשיח יותר אך גם תופס נפח רב יותר
:נוסף (לא קוולנטי) ריקישור בין מולקול
י זיקה חשמלית בין מטענים "הנוצר ע, קישור המוכר בחלבונים –גשר מלח = גשרי מימן .1מאחר והאלקטרונים לא , לא מדובר בקישור יוני .מנוגדים שבאזורים שונים בחלבון
.מועברים בשלמותם מאטום לאטום
ביות יעדיפו לצמצם מגע מולקולות הידרופו, במדיום מימי – אינטראקציות הידרופוביות .2 . עם מולקולות המים ולכן יקשרו זו לזו
אטומי חמצן –מולקולות המים עוטפות את יוני המלח המומסים –מים משמשים כממס מצוין מים ההיא כי הסיבה לכך . בעוד אטומי המימן מתקרבים ליסוד השלילי, מתקרבים ליסוד החיובי
(. 80)הם בעלי מקדם דיאלקטרי גבוה מאוד
מספר טהור המבטא באיזו מידה החומר מפחית את הזיקה , מקדם דיאלקטרי הוא תכונהאם נכניס נתרן כלורי , כלומר(. כלור ונתרן לדוגמא)מנוגדים החשמלית שבין שני מטענים
הוא בעל מקדם , ל לדוגמאאלכוהו. 80חוזק הקישור בין הכלור לנתרן יקטן פי , לתמיסה מימית . אומנם זו תהיה קטנה בהרבה מאשר זו של המים, לכן הוא בעל יכולת המסה. 24-25דיאלקטרי
אתר הפעיל לכאשר סובסטראט מגיע –תגובות ביוכימיות רבות מתרחשות בתוך תגובה מימית שמדובר או( גובה תתרחשואף הכרחיים בכדי שת) לקישור עשויים לתרוםהמים – של האנזים
רק לאחר קישור יתבצעולכן –( תרשים תחתון)בתגובה הזקוקה לסביבה הידרופובית . שמולקולות המים יידחו מן האזור
:הכללים התרמו דינמיים המאפיינים כל תהליך המתרחש בעולם
לפי חוק זה סך כל (. והמסה)חוק שימוש האנרגיה –החוק הראשון של התרמודינאמיקה .1 . אלא רק תשנה צורתה, אנרגיה לא תיווצר ולא תעלם –האנרגיה בעולם קבוע
ויאבד –חלק מן האנרגיה שנדרשה לפעולה הופך ללא פעיל , בכל פעולה המתבצעת .2אך כן תשתנה זו , כך שלא תשתנה כמות האנרגיה שבעולם)חום לסביבה בתהליך כ
(. המצויה במערכת
A + B C + D + energy
התגובה המתוארת מעלה מתארת מצב בו אנרגיית התוצרים גדולה מן האנרגיה שהייתה
:עשויה להשתחרר באחת משתי צורות( G∆המתוארת כ)כמות אנרגיה זו . למגיבים
.אנטרופיית המערכת, הנותרת מגדילה את אי הסדר במערכתכך שהאנרגיה , כחום .1
כך שרק חלק מכמות החום תגרום לעליית –אנרגיה צימוד לריאקציה נוספת הדורשת .2 . פעיל תגובה לא ספונטנית כלשהיוהשאר י, האנטרופיה

.2010 – 2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
כך שלעולם לא מתקיים , תאבד אנרגיה לסביבה –בכל מעבר מצורת אנרגיה אחת לשנייה . אנרגיה 100%מעבר של
יכלול רק את האנרגיה נטו G∆. נחשב לפי ההפרש בין אנרגיית המגיבים לתוצרים G∆את האנרגיית השפעול היא הסיבה לכך שלא . שהשקענו במערכת אנרגיית השפעולללא , שהשתחררה
.כל תגובה אקסוגנית תתרחש במהירות
תגובה ספונטנית, תגובה המשחררת אנרגיה –תגובה אקסוגנית ,∆G שלילי .
מן התגובות הביוכימיות חלק גדול . תגובה הדורשת השקעת אנרגיה –תגובה אנדוגנית .ולכן יש צורך בתגובות אקסוגניות מצומדות להן הגורמות להן להתרחש, דורשות אנרגיה
עליה להתרחש בקרבה לתגובה , בכדי שתגובה שכזו תוכל לנצל את האנרגיה מן הצימוד .וזמן קצר לאחריה, האקסוגנית
: י המשוואה"ר מבוטא עהדב. היא ספונטנית, לאנטרופיה, הנטייה לחוסר סדר מקסימלי
∆H = ∆𝐺 + 𝑇∆𝑆 . לעולם , האנרגיה הפוטנציאלית של המערכת, שהאנתלפיהמשוואה זו מראהחלק מן האנרגיה תנוצל , המוחלט 0מאחר ובכל טמפרטורה שאינה ה, י מערכת"לא כולה תנוצל ע
. י האנטרופיה"ע
∆G =0 – והתהליך המדובר הוא הפיך, מ"מצב בו המערכת מצויה בש .
כל איבוד של חום מן המערכת יגדיל את אי הסדר .
:מקורות אנרגיה חופשית שבתא
קיים שוויון ריכוזים בין ית הגדול ביותר יתרחש כאשר אנטרופיהה מצב: מפל ריכוזים -י "ע, תרמו דינמיתה ניתן ליצור תגובות מועדפות מבחינכלומר –שני צדי הממברנה
קום בו ריכוזם נמוך למ (אחד מצידי הממברנה) יונים ממקום בו ריכוזם גבוה העברת (. צידה השני)
כן צימודה לתהליך הגורם ול, משחררת אנרגיהתגובה זו : Pו ADPל ATPפירוק מולקולת -
סופי G∆מאזן ) התהליך הכולל יוגדר כספונטני –הקטנת האנטרופיה תאפשר את קיומו ל . הדבר מתואר בשקף. (שלילי
. ת בתור מקור אנרגיהמשמש במקרים רבים בתא כתגובה מצומד ATPפירוק מולקולות :את מולקולה זו ניתן לפרק בכמה דרכים
י כך לקבל "אותם ניתן לשבור וע P-Pישנם שני קשרי .א𝑘𝑗
mol .מכל אחד 31
בודד אשר ממנו נקבל רק C-Pקשר .ב𝑘𝑗
mol 4.
:הביוכימיה של ביטוי גנים
כי בתא ים הםהעיקרי יםהבדלשני ה. אאוקריוטיותא פרוקריוטיניתן לראות תא בתרשים
וכמו כן חסרים בו אברונים רבים , לא מצוי בגרעין אלא מפוזר בציטופלזמה DNAה פרוקריוטידוגמת )או שהם קיימים בצורה שונה ( דוגמת מיטוכונדריות)הקיימים בתאים אאוקריוטים
עצם העובדה שהריבוזומים החיידקיים שונים מן הריבוזומים ההומניים מאפשרת (. ריבוזומים . פגיה בריבוזומים החיידקיים שימוש באנטיביוטיקה המבוססת על
מנגנוני העברת המידע הגנטי ברובם , למרות ההבדליםעובדה המאפשרת לחקור , יימים דמיון רב בשני סוגי התאיםמק
ולבצע , בצורה נוחה מאוד את תהליכים אלו בחיידקים . התאמות לתאים אאוקריוטים לאחר מכן

.2010 – 2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:החומר הגנטי
:DNAהתורשתי הוא הניסויים הוכיחו שהחומר 3
:פעמיים ערכואותו :הניסוי הראשון
, זן לא אלים) Rוזן ( זן אלים) Sזן –נדגמו שתי אוכלוסיות חיידקי דלקת ריאות .א
ועל הטחול שלהם היה , כך שהם מתו, הוזרקו לעכברים Sחיידקים מזן (. מחוספס
לא קרה להם דבר , הוזרקו לעכברים Rכאשר חיידקים מזן . Sניתן למצוא מושבות
כך שלא – הורתחו ולאחר מכן הוזרקו לעכברים Sחיידקי . והטחול שלהם נמצא נקי
והזריקו את , רגילים Rאת החיידקים המורתחים ערבבו עם חיידקי . קרה להם דבר
נמצאו על Sומושבות מסוג , כתוצאה מכך העכברים מתו. תערובת זו לעכברים .הטחול שלהם
, הניסוי הראה שגורם כלשהו מן החיידקים המורתחים עבר לחיידקים הלא אלימיםוידוע שההרתחה הורסת את מאחר. כאשר גורם זה הוא האחראי על אופי החיידק
הוא החומר DNAוי הוכיח שההניס, DNAאומנם לא פוגעת בגדילי ה, החלבונים .החומר התורשתי, שעבר
בו החיידקים הלא פתוגנים החיים , אומנם נוסף שלב, הניסוי הראשון בוצע בשנית .בכאשר הם עורבבו עם מולקולות –עורבבו עם מאקרומולקולות שונות בכל פעם
אומנם . העכברים שרדו, חלבוניות או סוכריות מתוך החיידקים הפתוגנים, שומניות
. העכברים מתו, (רשיםהחלק הרביעי בת) DNAכאשר החיידקים עורבבו עם ה
:ייס'ניסוי הרשי צ: הניסוי השני
:בעזרת שני איזוטופים רדיואקטיביים ים'פאגבקטריוסימנו
.הקיים באופן ודאי ובלעדי בחלבונים( S35)איזוטופ סולפאט .1
. DNAהקיים באופן ודאי ובלעדי ב( P32)איזוטופ פוספט .2
ת א. ים תקפו את החיידקים והתרבו בהם'כך שהפאג ,ים אלו'חשפו תרחיף של חיידקים לפאג :2כך שהוא נחלק ל, לצנטריפוגות הכניסו התרחיף
(.חלקים שאינם קשורים למידע התורשתיה)ובו שאריות הוירוסים , נוזל עליו -
. (ים'ובהם הפאג) משקע ובו החיידקים -
:תוצאות. בכל אחת מן הפרקציות נבדקה נוכחות הפוספט והסולפאט הרדיואקטיביים
.(P32נוכחות גבוהה של )נמצא ברובו במשקע החיידקים DNAה -
. (S35נוכחות גבוהה של ) בנוזל העליוןברובה נמצאה ' המעטפת החלבונית של הפאג -
מגדירה כי החומר שיוחדר לחיידקים הוא , עצם העובדה כי הוירוסים מתרבים בעזרת החיידק
DNAהוכח באופן בטוח כי ה –נוכחות פוספט ולכן מאחר ונמצאה –האחראי על המידע הגנטי . הוא המידע הגנטי
:הניסוי השלישי
נקבל Aאם נדביק עלי טבק בוירוס מסוג . ותוקף עלי טבק( Bו A)קיים בשני זנים TMVהוירוס
והושתל בתוך Bמוירוס מסוג RNAנלקח ה, בניסוי. Bל לגבי וירוס מסוג "וכנ, Aוירוס נוסף מסוג
, אם החלבון אחראי על החומר התורשתי –ולאחר מכן הדביקו את העלים , ולהפך Aמעטפת מסוג
.2010 – 2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הוא האחראי נקבל תוצאה DNAאך אם ה, Aייצר וירוסים מסוג Aוירוס מסוג ) לא יהיה הבדל
. (Aייצר וירוסים מסוג Bוירוס מסוג ) הפוכה
היום ידוע כי וירוסים רבים מסוג : 1הערהRNA הם מסוכנים מאוד(דוגמא לHIV) , . וחלקם ידועים כמסרטנים
הניסוי הוכיח שהחומר התורשתי הוא לא רק : 2הערהDNA , אלא גםRNA .
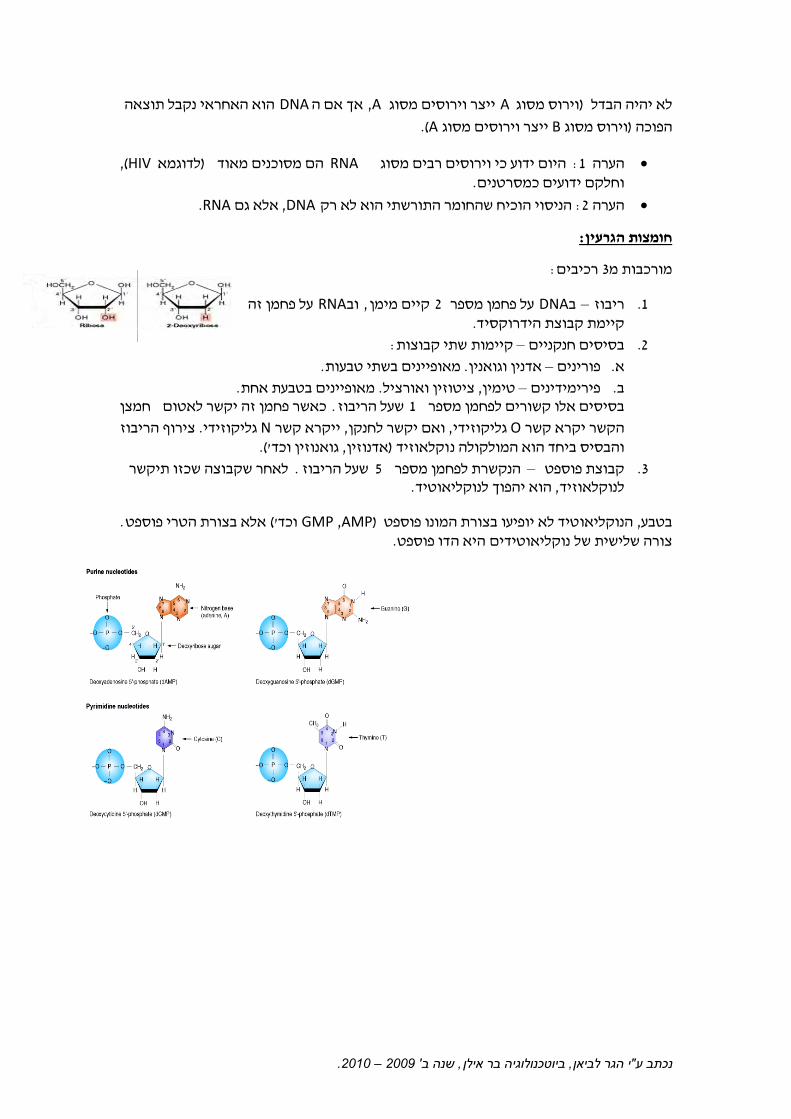
:חומצות הגרעין
:רכיבים 3מורכבות מ
על פחמן זה RNAוב, קיים מימן 2על פחמן מספר DNAב –ריבוז .1 . דהידרוקסיקיימת קבוצת
:שתי קבוצותקיימות –בסיסים חנקניים .2
.מאופיינים בשתי טבעות. אדנין וגואנין –פורינים .א
.מאופיינים בטבעת אחת. ציטוזין ואורציל, טימין –פירימידינים .ב חמצןכאשר פחמן זה יקשר לאטום . שעל הריבוז 1בסיסים אלו קשורים לפחמן מספר
הריבוז צירוף. גליקוזידי Nקרא קשר יי, ואם יקשר לחנקן, גליקוזידי Oהקשר יקרא קשר (. 'גואנוזין וכד, אדנוזין) וקלאוזידוהבסיס ביחד הוא המולקולה נ
לאחר שקבוצה שכזו תיקשר . שעל הריבוז 5הנקשרת לפחמן מספר –קבוצת פוספט .3 . לנוקליאוטידהפוך הוא י, לנוקלאוזיד
. אלא בצורת הטרי פוספט( 'וכד AMP ,GMP)טיד לא יופיעו בצורת המונו פוספט הנוקליאו, בטבע . דו פוספטהיא ה נוקליאוטידיםצורה שלישית של

1 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:20.10 – 2הרצאה –' ביוכימיה א
תחילת ל dנוסיף DNAבצורת ה( AMPאו ATP ,ADP)נוקליאוטיד כאשר נרצה לציין, לשם סימון
ידובר בנוקליאוטידים המרכיבים מולקולות , אם לא נוסיף...(. dATP ,dADP)שם המולקולה
RNA.
:איזומרים מבניים –טאוטומרים
בתמונה . 'טימין וכד, ישנם מספר טאוטומרים לאדנין –הנוקליאוטידים קיימים בצורות שונות :מספר דוגמאות לכךניתן לראות משמאל
. מינו-צורת האמינו וצורת האו, וציטוזין אדנין יםבסיסב שני איזומרים הקיימים .1 .הטאוטומר השכיח יותר הוא האמינו
בה )צורת הקטו –וגואנין תימיןבבסיסים ימים ראות שני איזומרים הקיניתן ל למטה .2
את רק נמצא , RNAוה DNAבכל מבני ה. האנול וצורת( הפחמן והחמצן יוצרים קשר כפול . הקטוצורת
בו קשרים בנה טבעתי מיוחד וגם הפורינים וגם הפירימידנים מקיימים מבולעים אור בצורה יעילה מבנה זה גורם לכך שהנוקליאוטידים . כפולים
כאשר נרצה למדוד את כמות , לכן(. UVתחום ה)ננומטר 260באורך גל של , שבתא( או כמות הנוקליאוטידים בצורתם החופשית)חומצות הגרעין
ניתן לראות בגרף ששיא . ננומטר 260נשתמש בספקטרופוטומטר המכויל ל .הנוקליאוטידים מתרכז באזור זההבליעה של
חומצות הגרעיןהעובדה ש: הערה ,DNA וRNA , קולטות קרינתUV לה מאודבצורה יעי ,
UVבשעות אלו קרינת ה. בקיץהצהריים הסיבה לכך שמסוכן להיחשף לשמש בשעות היאבדרך כלל מערכות . וחומצות הגרעין אשר קולטות אותה עשויות להיפגם, היא חזקה
עשויות לא אך בשעות אלו כמות הקרינה כה גדולה שהן , תיקון מתקנות את נזקי הקרינה, הפגם יגרום למוות של התא –במקרה הטוב .וחלק מן הפגמים לא יתוקנו, לעמוד בעומס
שאופייה הקטלני יגרום לסרטן או , אומנם במקרה רע עשויה להתרחש מוטציה שקטה .למחלה אחרת
:RNAו DNAגדילי ה: פולינוקלאוטידים
:שונים לכל גדיל ישנם שני קצוות –כיום אנו יודעים שגדילים אלו הם בעלי כיווניות
וספט הקשורה לפחמן קבוצת הפ ישנה הקצה בו על הנוקליאוטיד האחרון –' 5קצה .1, לא תהיה קשורה אליו ןגם אם קבוצת הזרח' 5קצה יוגדרקצה זה . שעל הריבוז' 5
במקרה . המסוגל ליצור קישור לקבוצת הפוספט' 5אלא פשוט הריבוז אשר בו פחמן .פנוי' 5ונה קצה זה הוא יכ
.שעל הריבוז פנוי לקישור' 3הקצה בו על הנוקליאוטיד האחרון פחמן –' 3קצה .2הנוקליאוטידים שיתווספו –העובדה כי פחמן זה פנוי היא המגדירה את הכיווניות
. לשרשרת תמיד ייקשרו לקצה זה
לכךסיבה מבנה זה הוא ה .'3ל' 5יד נבנות מכיוון כך שהשרשראות תמהכיווניות מוגדרת לעניין זה השלכות משמעותיות בתהליכי הכפלה –' 3ל' 5נזימים תמיד יפעלו מכיוון האש
.והתחלקות
הגרעין. שני מושגים הנוגעים לאופי ח:
(.קיימות בצורה זו RNAמרבית מולקולות ה) SS = single strand .א

2 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
.DS = double strand .ב
משלימים הישנם אזורים במולקולות מסוג זה , SSהן RNAלא כל מולקולות ה
SSשחלקה RNAדוגמא למולקולת משמאל ישנה .בסיסים והופכים לדו גדיליים
.SDוחלקה
:חיבור נוקליאוטיד לגדיל קיים
] דורש)דינמית תהליך לא מועדף מבחינה תרמוהוא תהליך זהkj
molהגדיל יעדיף –( 25[
התהליך , בכדי לפתור בעיה זו .מאשר להמשיך ולהיבנות (שלילי G∆תהליך בעל )להתפרק המשחררת מספיק אנרגיה בכדי שהתהליך כולו יהיה מועדף , תריאקציה אחרמצומד ל
הריאקציה השנייה היא פירוק נוקליאוזיד טרי פוספט לנוקליאוזיד . מבחינה תרמודינמית
]ובמהלכה משתחררים , מונו פוספטkj
mol אהסופי הו G∆כך ש – 31 [
kj
molשתי ) -6 [
מסיבה זו ניתן לבנות גדיל . (תרמו דינמיתהריאקציות המצומדות מועדפות מבחינה
DNA/RNA רק בעזרת המונומריםATP ,GTP ולא בעזרתAMP ,GMP.
:הערות
במהלך יצירת הקשר של קבוצת הפוספט לפחמן משתחררת מולקולת מים.
י האנזים "נוצר ע, נוקלאוטידים 2הקיים בין , הקשר הפוספודיאסטריDNA פולימראז.
קבוצת הפוספט בחומצות הגרעין מקור האופי החומצי של(PO4אשר טעונה שלילית , (-
של החומצה pKaהפיזיולוגי גבוהה מן ה pHה)מאחר ושחררה את הפרוטון שהיה עליה (. הזרחתית
:ושגיםמספר מ
הוא עדיין לא פולינוקלאוטיד BP 20,25מ קצרנוקליאוטידים הרצף : אוליגונוקלאוטיד - .פריימרים הם סוג של אוליגונוקלאוטיד. אלא אוליגונוקלאוטיד
הגדרה זו . אשר בסופו של הדבר יביא ליצירת חלבון, DNAקטע על פני מולקולת ה: גן -
אשר RNAאלא ישועתק ל –הגן לא יתורגם לחלבון בהם מקרים מעטים נכונה מלבד ל .הוא יהווה כיחידה פעילה בעצמו
אחוז קטן יותר מן )הדבר כולל גם אזורים המקודדים לגנים . שבתא DNAכלל ה: גנום -
אלא ( RNAלא לחלבון ולא ל)וגם רצפים שאינם מקודדים לגורם פונקציונאלי ( הגנום . 'וכד מבנה, בקרה, נצרכים לשם ארגון
כך, ולקת למספר מולקולות אדירותמח DNAכל תכולת ה: הגנום מסודר בכרומוזומים, חלבונים לדוגמא)חומרים רבים אחרים ל בנוסף, ומוזום מיליוני נוקליאוטידיםבכל כרש
, בודד DNAכל כרומוזום נוצר מרצף (. וחלקם מבניים, אשר חלקם קשורים בביטוי גניםכרומוזום גדול יכול להכיל אחוז )כאשר גודל הכרומוזום לא מעיד על כמות הגנים שבו
(.גבוה מאוד של אזורי בקרה ומעט מאוד אזורים המקודדים לגן
מסובבים סביב DNAגדילי ה .הכרומוזוםהמרכיב את DNAהצורת מבנה : נוקלאוזום -החלבון המסובב . (חת חלבוניםמשפ)המורכבים מן ההיסטונים קומפלקסים חלבוניים
ל נוקלאוזום הוא בעצם כ. H1 –סביב הקומפלקס החלבוני הוא גם היסטון DNAאת ה
כמות אדירה של . המלופף סביבה DNAמקטע + של קומפלקס חלבוני יחידה בודדתבזמן . לאחר קיפולים רבים, כרומוזום השלםנוקלאוזומים בשרשרת היא היוצרת את ה
אומנם בזמנים אחרים רמת , (Xצורת ה)יהיה במצבו הדחוס ביותר חלוקה הכרומוזום
. 'שכפול וכד, פנוי לתהליכי שעתוק DNAכך שה, הדחיסה יורדת

3 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
כאשר מקור כל עותק , כל כרומוזום קיים בשני עותקים –הוא גנום דיפלואידי , הגנום האנושילים בין נוקליאוטידים ישנם הבד) הכרומוזומים אינם זהים לחלוטין באופיים. באב או באם
. כל יצור המתרבה רבייה מינית הוא יצור דיפלואידי. וזו הסיבה לכך שישנם שני סטים, (בודדיםכאשר יצור שכזה מתרבה הוא . בעלי סט בודד של כרומוזומים –הם האפלואידים יצורים אחרים
.האפלואידים( לדוגמאספורות )ומתחלק לשני גורמים –מכפיל את הגנום שלו
.XYאו , XXוזוג אחד יהיה , זומים זהיםמתוכם מייצגים כרומו 22 –זוגות 23מכיל קריוטיפ אנושימרבית מקרי . חריגויות במספר הכרומוזומים שבקריוטיפ מכונות אנאאופלואידיה
בשלב ההיריון או )אלא נגמרים במוות מוקדם , האנאופלואידיה אינם מגיעים למצב מתפתח (. לאחר הלידה
מקרה . חוסר של כרומוזום בודד/ בו קיים עודף, מצב לא מאוזן היא בעצםופלואידיה אאנאהבמקום שני 21( טריזומיה)כרומוזומי 3י קיום "הנגרמת ע –מוכר של תופעה זו הוא תסמונת דאון
מקרי בניגוד ל. 13טריזומיה הקיימת בכרומוזום תופעה פחות מוכרת היא . עותקים, מקרים כאלו בכרומוזומי מין הם פחות בעייתיים, 1-22ם כרומוזומיהאנאאופלואידיה של
.חיים מפותחים ומאפשרים
:DNAהמבנה המרחבי של ה
ולאחר שראו זאת ווטסון , מורכב משני גדילים פענחה רוזלין פרנקלין DNAאת העובדה שה :מסקנותיהם. פענחו את המבנה כולוהם , וקריק
. בנוי כשני גדילים המשולבים כסליל DNAה -
(. '5מול ' 3)בעלי כיווניות הפוכה , הגדילים הם אנטי מקבילים -
עובדה )מופנה כלפי חוץ המולקולה , השלד ועליו יוני הפוספט, DNAהאזור הטעון של ה -
. בעוד הבסיסים החנקניים מופנים כלפי פנים( מסיס במים DNAהמסבירה מדוע ה
הצורה החזקה ביותר של –מדובר בקשרי מימן בזווית שטוחה : יליםהקישור שבין הגד -
קשרי 2מקיימים Tו Aוהבסיסים , קשרי מימן 3מקיימים Gו Cהבסיסים .קשרים אלו
ולכן צורת הסליל היא אחידה , T (1.08nm)ל Aזהה למרחק שבין Cל Gהמרחק בין . מימן .הלכל אורכ
–ניתן לראות את הטבעות החנקניות – של הסליל" על"השרטוט מציג מבט בין זוג (.כמעט לחלוטין וכמעט תמיד)אשר נמצאות במצב מישורי
כך שבשביל להשלים סיבוב יש , 36°נוקליאוטידים תמיד תהיה הסטה של בין שני סטים של נוקליאוטידים ( לגובה)מרחק . נוקליאוטידים 10צורך ב
בכדי שאינטראקציות ואן . בעתשל טהמדויק VDVשזהו קוטר , Å 0.34הוא כעל המולקולות להיות במרחק , דר ואלס יתקיימו בצורתן האופטימלית
.DNAוהדבר מתקיים בין גדילי ה –( 0.34) זו מזו ספציפי ביותר
התאמה )יחידות טבעתיות 10ו, ננומטרים 3.4בסיבוב ישנם : 1הערה (. מוחלטת לרדיוס ואן דר ואלס
אם נעביר ציר דמיוני במרכז –קשרי המימן חוצים בדיוק את מרכז הסליל : 2 הערהבבסיס , לדוגמא, ולא יפגע)הסליל הכפול הוא יעבור בצורה ישרה דרך קשרי המימן
(. חנקני
המופיעות , גדולות וקטנות, בכניסות ניתן להבחין – הסתכלות על הסליל מן הצד
ניין זה המשמעות של ע(. minor grooveמול ולצד mojor groove)לסירוגין יכולים להיקשר רק ( אנזימי שעתוק לדוגמא)מתבטאת בכך שחלבונים מסוימים
(. grooveאו רק לסוג כלשהו של , לדוגמא major groovesרק ל)לאזורים מסוימים

4 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
ובזכותם לחלבון היכולת , יםקישור ספציפי של חלבון לרצף מסו אזורים אלו מאפשרים . להיקשר את הסליללאן לזהות
החלק הצבעוני )מראה את ההבדלים שבין החשיפה של הנוקליאוטידים התרשים
. minor grooveלבין חשיפתם באזור ה major grooveבאזור ( במולקולה
כך ששני , וקישור חלבונים אליה יכול לגרום לה להתכופף, היא מולקולה אלסטית DNAה
יכולת זו של החלבונים בעלת . יכולים להתקרב זה לזה DNAאזורים יחסית רחוקים ב
והיא הגורם המאפשר מקרים בהם אזורים , DNAחשיבות למנגנוני תיקון שונים של ה
. משפיעים זה על זה DNAרחוקים ב
נגד כיוון , )+ויכול להיות סליל שמאלי , (עם כיוון השעון, )-יכול להיות סליל ימני DNAסליל האשר אלו מזהים את השינויים המרחביים ולא –של הדבר נוגעת לאנזימים החשיבות(. השעון
לכן הסליל במציאות הוא (.)-אלא רק לצורת הסליל הימני –מסוגלים להיקשר לשתי הצורות .ימני
מאחר וישנם שני – DNAהמבנה הדו גדילי שמצאו ווטסון וקריק היווה פיתרון לצורת הכפלת היכולה , יאורטיבאופן ת, דרך הכפלה זו. כל אחד יכול לשמש לתבנית לגדילים חדשים, גדילים
:האפשרויות 3. כאשר רק אחת קיימת במציאות, צורות 3להיעשות ב
תוצר ההעתקה היא מולקולה דו גדילית –מנגנון קונסרבטיבי .1ושני ,הסליל המקורי שנפתח מתחבר מחדש –חדשה לחלוטין
.הגדילים שסונתזו מתחברים זה לזה ויוצרים סליל נוסף
לאחר ההכפלה תתקבל מולקולה –מנגנון סמי קונסרבטיבי .2התוצר הוא שני . המורכבת מחד גדיל מקורי וחד גדיל חדש
. סיבים דו גדילים זהים
שום גדיל לא נותר כפי שהיה בצורתו –מנגנון דיספרסיבי .3
. ק ומסונתז מחדש גם הואמתפר backboneה –המקורית
:הוכיח כי המנגנון הקיים הוא הסמי קונסרבטיבי – לסון סטאלניסוי מס
( התמקדות רק בגרפים השמאליים)שלושת הגרפים משמאל
:נותשו DNAשל צפיפות מולקולות מראים התפלגות
.מספר מולקולות= Yציר -
. צפיפות= Xציר -
שגודלו בנוכחות DNAהגרף הראשון מראה צפיפות של גדילי
במקום חנקן רגיל , (N15)איזוטופים רדיואקטיביים של חנקן
(N14 .) הגרף השני מראה צפיפות של גדיליDNA שגודלואת צפיפות המולקולות מדדו לפי . בנוכחות חנקן רגיל
/ אשר מפריד את המולקולות כך שהכבדות, גרדיאנט צפיפות . יהיו למעלה והקלות יותר, צפופות יותר יהיו למטה
, הוא הצפוף יותר( הסליל החום) N15ניתן לראות שהסליל ובו
(.14N)גדולים יותר מחנקן רגיל N15האיזוטופים והדבר הגיוני מאחר ו
לאחר מכן העבירו את החיידקים .בלבד( N15)חנקן המכילות DNAהשתמשו במולקולות בניסוי
ירדה בחצי DNAההתוצאה היא שצפיפות . כך שהם שכפלו עצמם בנוכחותו, בלבד N14למצע ובו

5 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הגדילים החדשים יימצאו בין הגדילים , בגרדיאנט צפיפות – ממה שהייתה לפני השכפול
. N14המקוריים לבין גדילים שגודלו בנוכחות
ניתן להבין כי הן , ירדה צפיפות המולקולות –ממצא זה פוסל את האפשרות הקונסרבטיבית
אם היינו מקבלים שתי קבוצות . N15וממולקולות בהן , N14מורכבות בחלקן ממולקולות בהן לא הוכיח זה יש לציים כי ניסוי. אז היה מדובר בצורה הקונסרבטיבית, בעלות צפיפות שונה
. מאחר וגם הצורה הדיספרסיבית אפשרית –באופן מוחלט את הצורה הסמי קונסרבטיבית
:כיח את המנגנון הסמי קונסרבטיביהוהמשך הניסוי אשר
השמתם בתמיסה י"ע DNAה הפרידו את גדילי. הניסוי מתואר בגרפים הימניים בתמונה שלמעלה
את . שוכפלו בנוכחות חנקן קל DNAגדילי ה –הניסוי הקודם מכן חזרו עללאחר . בסיסית מאוד –( של גדילים ולא של סלילים)תוצרי הדור הבא שמו בגרדיאנט צפיפות ומצאו שתי אוכלוסיות
ומצאו כי גם לאחר , חזרו על הניסוי. אחת בעלת צפיפות נמוכה והשנייה בעלת צפיפות גבוהההמתאימה )ות נמוכה אך כעת מירב הגדילים בעלי צפיפ –הכפלה נוספת נותרו שתי אוכלוסיות
ניסוי זה שלל את . בעוד כמות הגדילים בעלי הצפיפות הגדולה לא משתנה, (לחנקן הקל שבמצעהיינו מקבלים קבוצה בודדת בעלת צפיפות בינונית ואם היה מתקיים המנגנון הדיספרסיבי מאחר
התחלנו את ולא את אותן שתי הקבוצות איתן, (אשר יורדת עם כל דור שגדל במצע של חנקן קל) .הניסוי
:מבנים מרחביים שונים
.בלבד B-DNAמבנה מסוג ל אותו הסברנו עד עכשיו נכון DNAתיאור המבנה של סליל וגדילי ה
ניצור מולקולות אומנם אם. B-DNAתמיד יופיע כ DNAה, הסביבה הקיימת בתא, בסביבה מימיתלדוגמא אם נוציא חומצת גרעין מן התא בכדי לבדוק את מבנה באמצעות , בסביבה לא מימית
.נראה כי ישנם מבנים מרחביים נוספים של הסליל הכפול – Xrayדיפרקציית או , קריסטלוגרפיה
A-DNA:
, גם סליל זה הוא ימני. כאשר רמת הלחות נמוכה, נוצר בתנאים יבשים יותר A-DNAהמבנה
כמו , קשרים אלו הם קשרי מימן. Cנקשר ל Gו Tנקשר ל Aוגם הוא מקיים זיווג בסיסים בו
נוצרים על מרכז לא הם. אומנם קשרים אלו הם בעלי אופי שונה, B-DNAהקשרים הקיימים ב .אם נסתכל על הסליל מלמעלה נראה כי מרכזו ריק .מעטפתה, אזור השלדב אלא, הסלילקשרי , נוכל לראות הטיה מאוד גדולה של הבסיסים, אם נסתכל על גדיל זה מן הצד, בנוסף
(. B-DNAבניגוד לזווית כמעט ישרה שיוצר ה)ליל הבסיסים יוצרים זווית חדה ביחס לסהמימן ו
majorיהיה קשה להבחין בין ה, מן הצד A-DNAהדבר גורם לכך שכאשר נסתכל על גדיל ה
groove לminor groove ,מאחר וגודלם כמעט זהה.
מבנהו המרחבי , DNAה נראה כי בניגוד לסליל, בסביבה מימית RNAניצור סליל כפול של אם
המורכבת מגדיל , אם ניצור מולקולה היברידית. ויישאר כזה גם בסביבה לא מימית, A-RNAיהיה
DNA וגדילRNA , גם היא תהיה בעלת מבנהA-DNA .
-Bנמצאות מולקולות מים אשר מייצבות את מבנה ה, בסביבה מימית major grooveבאזורי ה
DNA ,והמבנה , מולקולות המים חסרות, בסביבה לא מימית. ולכן בסביבה שכזו הוא מועדף
ולה זו ישנה כי בריבוז של מולקהיא , הדבר לא מתקיים RNAהסיבה לכך שב. A-DNAמשתנה ל
major grooveהמונעת ממולקולות המים להיכנס לאזור ה, 2על פחמן מספר OHמולקולת
. B-RNAגם בסביבה מימית וגם בסביבה לא מימית לא יתקיים , לכן(. יוצרת הפרעה סטרית) לנוקליאוטידים יש את האפשרות ליצור יותר קשרי מימן מאשר אלו שהם יוצרים בין
.החנקנייםהבסיסים
1 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:25.10 – 3הרצאה –' ביוכימיה א
:מקרים 3עליו למדנו יתקיים באחד מ A-DNAמבנה ה
.מצוי בתנאי לחות נמוכים DNAסליל ה .1
(.לדוגמא tRNAב)יוצר מבנים דו גדיליים RNAכאשר .2
:דוגמאות למקרה זה. RNAומגדיל DNAכאשר נוצר סליל המורכב מגדיל .3
.בעת שעתוק .א
רצף משלים לקטע )ניקח פרוב , DNAאו RNAכאשר אנו רוצים למצוא מולקולת .ב
כך , (שעל הנוקליאוטידים Pαהמסומן באופן רדיואקטיבי דרך , שאנחנו מחפשים
.RNAו DNAכך לעתים נוצר סליל היברידי של . שיתחבר למולקולה שחיפשנו
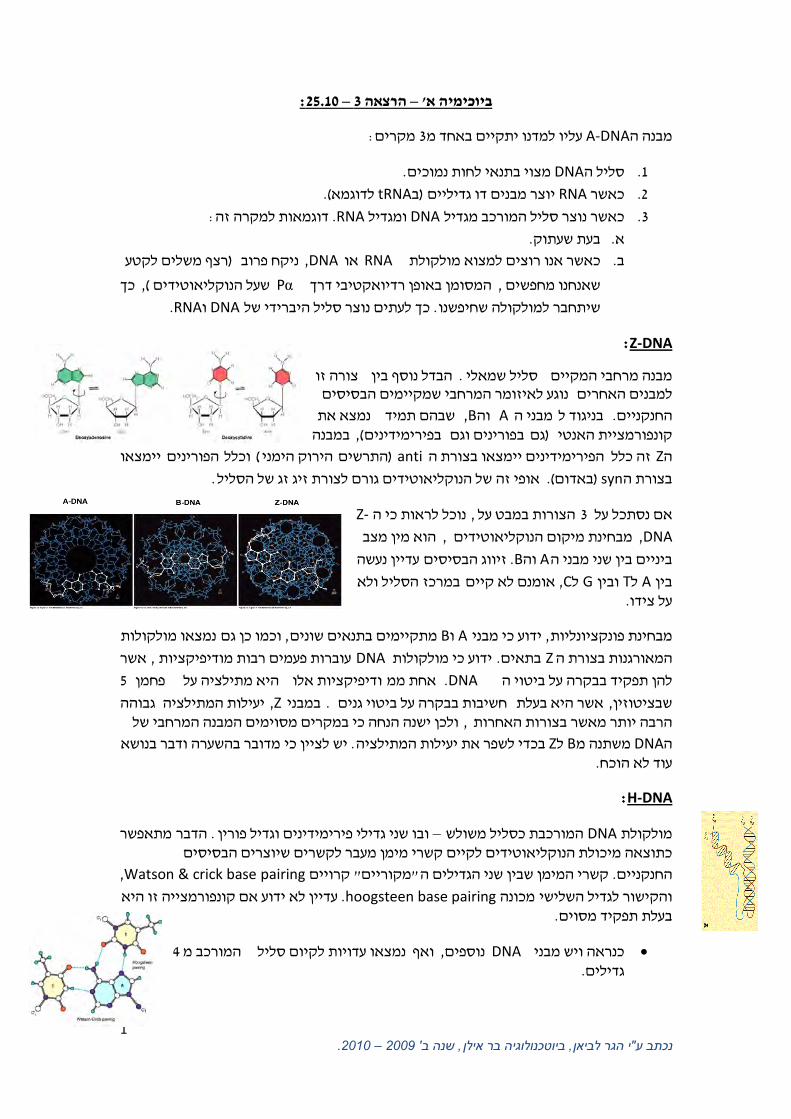
Z-DNA:
צורה זו הבדל נוסף בין . סליל שמאלימבנה מרחבי המקיים נוגע לאיזומר המרחבי שמקיימים הבסיסים למבנים האחרים
נמצא את בהם תמידש, Bוה Aמבני הבניגוד ל. החנקניים במבנה, (פירימידיניםבפורינים וגם ב גם)קונפורמציית האנטי
יימצאו הפורינים לוכל (הירוק הימניהתרשים ) antiהפירימידינים יימצאו בצורת ה זה כלל Zה
.אופי זה של הנוקליאוטידים גורם לצורת זיג זג של הסליל (. באדום) synבצורת ה
-Zנוכל לראות כי ה, הצורות במבט על 3אם נסתכל על
DNA ,הוא מין מצב , מבחינת מיקום הנוקליאוטידים
עדיין נעשה זיווג הבסיסים .Bוה Aביניים בין שני מבני ה
ל ולא הסלי במרכזאומנם לא קיים , Cל Gובין Tל Aבין . על צידו
נמצאו מולקולות וכמו כן גם , מתקיימים בתנאים שונים Bו Aידוע כי מבני , מבחינת פונקציונליות
אשר , עוברות פעמים רבות מודיפיקציות DNAידוע כי מולקולות . בתאים Zהמאורגנות בצורת ה
5פחמן היא מתילציה על דיפיקציות אלו ואחת ממ. DNAלהן תפקיד בבקרה על ביטוי ה
גבוההיעילות המתילציה , Zבמבני . חשיבות בבקרה על ביטוי גנים אשר היא בעלת, שבציטוזיןולכן ישנה הנחה כי במקרים מסוימים המבנה המרחבי של , הרבה יותר מאשר בצורות האחרות
מדובר בהשערה ודבר בנושא יש לציין כי . בכדי לשפר את יעילות המתילציה Zל Bמשתנה מ DNAה .לא הוכחעוד
H-DNA:
מתאפשרהדבר . ובו שני גדילי פירימידינים וגדיל פורין –המורכבת כסליל משולש DNAמולקולת כתוצאה מיכולת הנוקליאוטידים לקיים קשרי מימן מעבר לקשרים שיוצרים הבסיסים
, Watson & crick base pairingקרויים " מקוריים"קשרי המימן שבין שני הגדילים ה. החנקניים
עדיין לא ידוע אם קונפורמצייה זו היא . hoogsteen base pairingמכונה והקישור לגדיל השלישי . בעלת תפקיד מסוים
כנראה ויש מבניDNA 4המורכב מ נמצאו עדויות לקיום סלילואף , נוספים . גדילים

2 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:תלוי רצף DNAמבני
:לדוגמא. אשר קריאה שלו מימין או משמאל תהיה זהה –מדובר ברצף פלינדרומי
5' GCC GAG TAG CTA CTC ATT 3'
3' CGG CTC ATC GAT GAG TAA 5'
כזיים מאפשרים יצירה של צורת הנוקלאוטידים המר 6, מבחינה מעשית
ניתן לראות בתמונה )בשני כיוונים מנוגדים hairpinי יצירת מבנה "ע, צלבחשיבות . פעמים רבות אלו נוצרותצורות צלב כ, וףבתהליכי שחל(. משמאל
DNAאנזימים היודעים לזהות מולקולות מבנים אלו מתבטאת בקיום, במקרים רבים רצפים אלו הם רצפים פולינדרומים. יםיבאזורים מאוד ספציפ ולחתוך אותן
יכול להיווצר מבנה , מאחר ואין חובה לכך)שאם תתרחש , המאפשרים התארגנות מיוחדת במרחב
. האנזים יוכל להתאים עצמו לצורה זו ולפעול על הרצף( stem and loopלדוגמא , אחר
:ארגון החומר התורשתי בתוך התא
הטבלה שבשקף מראה יצורים . DNAולא תמיד הוא יהיה , DSלא כל החומר התורשתי יהיה
דוגמת )לי או שבהם מבנה מעג, (הגנום האנושי כפי מבנה, סליל)לינארי DNAשונים שבהם
.DS ולעתים SS לעתים הואהתורשתי חומר הניתן לראות כי ישנו (. פלסמיד
:מעגלי DNAלגבי
זה יכול להופיע בצורה DNA ,('פאג)או במולקולה קטנה ( חיידק)בין אם מדובר במולקולה גדולה
גורם לצורת או בצורה בה מתח מסוים, בה לא קיים מתח, (relax)המעגלית הפשוטה
מהווה supercoilצורת ה .במצגת ניתן לראות את הרמות השונות משמאל לימין. supercoilה
ככל שהיא מתפתלת –מבחינת נפח שתופסת המולקולה במרחב . DNAרמה נוספת של פיתול של ה . כך היא תופסת נפח קטן יותר בתוך התא, יותר
. כך היא דורשת יותר אנרגיה, (supercoiledיותר )ככל שהמולקולה דורשת פחות מקום , עם זאתכך יש להשקיע יותר אנרגיה ולמצוא יותר דרכים לפתוח אותה , ככל שהיא יותר מפותלת, בנוסף
. כדי שהגנים יהיו נגישים לתהליכי שכפול ושעתוק
Superciol תלוי בכיוון הליפוף, או שליליתובית ויכולה להיות חי, הוא צורה בעלת כיווניות .
–למצב המלופף יש צורך במנגנון שיגרום למתח בסליל relaxedיעבור מהמצב DNAבכדי שסליל מתח נוצר עקב הגדלה או הקטנה של מספר , מבחינה מעשית. ללא מתח הסליל לא יתלפף
:supercoilי סוגי ישנם שנ. relaxedבמצב ה( LKה)הסיבובים שהסליל מקיים בין הגדילים
.תגרום ליצירת מתח שלילי ועקב כך לליפוף בעל כיווניות שלילית – LKהקטנת ה -
. תגרום ליצירת מתח חיובי ועקב כך לליפוף בעל כיווניות חיובית – LKהגדלת ה -
מספר הסיבובים שהסליל יוצר סביב עצמו מוגדר כ: הערהW.

3 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
אם נריץ מולקולות : 2הערהDNA כאשר כל המולקולות בעלות מטען , באלקטרופורזה
נראה כי המולקולות המלופפות נעות מהר יותר מן המולקולות שבמצב , (pH = 7)שלילי
relaxed ,אם ניצור חתך בסליל כך שהמולקולה כבר . מאחר ותופסות פחות מקום במרחב
כל המולקולות כך ש, supercoilו relaxedכבר לא יתקיימו מבני –לא תהיה מעגלית . ירוצו יחד
. איזומרים בעלי טופולוגיה שונה, חת אנזימים היוצרים טופואיזומריםמשפ –טופואיזומראזות
ישנם שני סוגים . supercoilשלבסוף גורם למבנה , חלבונים אלו הם האחראיים על יצירת המתח :עיקריים של טופואיזומראזות
לאחר יצירת . אנזים היודע לחתוך את המולקולה הדו גדילית בגדיל אחד בלבד – 1סוג .1כך שמתקבל מצב בו , הגדיל המשלים יכול לעבור דרך החיתוך ולסגור את החתך, החיתוך
כלומר אם היה מתח . מסתובבים אחד סביב השני הגדיליםהופחת מספר הסיבובים בהם המתח העודף היה גורם , ה במצב אופטימליאם הגדיל כבר הי. ועודף הדבר גרם לשחרור
לרוב הוא אומנם , LKאנזים זה יכול גם להקטין וגם להגדיל את ה. למולקולה להתפתל . מספר הסיבובים הקטנת, משמש לשחרור המתח
כך , י שחרור הקשר הפוספודי אסטרי"אנזים זה משיג את האנרגיה לביצוע התהליך ע
י פתיחת הקשר הוא אוגר "את האנרגיה שהאנזים משיג ע. ATPשאין לו צורך באנרגיית . אצלו ולא משחרר לציטופלזמה
כך שהוא יכול לפתוח את אחד , ישנם מקרים בהם מתרחשת מוטציה באנזים זה
תא רגיל .שאינם מתוקנים DNAשברי כך שנוצרים , אך לא לסגור אותו מחדש, הגדיליםלא יעבור אפופטוזיס כך , סרטני לדוגמא תא, אומנם תא אחר, ימות עקב מקרה כזה
מראז מסוג טיפולים אנטי סרטניים רבים מכוונים כנגד אנזים הטופואיזו .שהשבר יתבטא .תוך ניסיון לעכב את פעילותו, 1
תוך שימוש יודע לבצע חיתוך וסגירה של שני הגדילים – DNA gyraseמכונה – 2סוג .2
הוא מאפשר מעבר מולקולות דרך , DNAלת המלבד ליצירת מתח במולקו. ATPבאנרגיית עניין זה שימושי בזמן המעבר . ולאחר מכן סגירה של הסליל כאילו לא קרה דבר, החתך
צריך לעבור ממצב מפוזר בגרעין אל DNAכאשר ה –מן האינטרפאזה אל שלב החלוקה אנזימים אלו מקלים על הארגון בכך שמאפשרות לסלילים –הכרומוזומים המסודרים
. הסבוכים זה בזה לעבור אחד דרך השני
, LKזאת מאחר וביכולתו להקטין את ה, מולקולהשל האנזים זה יכול ליצור דחיסת יתר .דבר אשר יגרור פיתולים נוספים של המולקולה
:להיות חד גדילית DNAלת הישנן שתי סיבות עיקריות אשר בגללן היינו מצפים ממולקו
אשר היינו מצפים מהם , מדובר בשני גדילים הטעונים שלילית –דחייה אלקטרוסטטית .1 .לדחות זה את זה ולא להיצמד זה לזה
האנטרופיה גדולה יותר כאשר הגדילים חופשיים לעצמם ולא , מבחינה תרמודינמית .2 . כאשר הם צמודים ובעצם מסודרים יותר
י "והדבר מוכח ע –בין הגדילים גורמים לרווח של אנטלפיה ם שקשריה, ייהלמרות הדח, אומנםהגדיל ייצור קשרי מימן עם –אם נפריד בין הגדילים . ת אנרגיה בכדי לפתוח אותםכך שנדרש
ולא מספיק לקבלת , הוא כנראה די נמוךהנובע מקשרי המימן ולכן הרווח של האנטלפיה , המים
. שליליים מספיקים Gערכי
היונים –לא מצויות במים מזוקקים אלא בתמיסה בעלת חוזק יוני גבוה DNAמולקולות הידוע כי
ובכך ממסכים את DNAנקשרים לשלד הטעון שלילית של ה( אשר בחלקם דו ערכיים)החיוביים ככל שהתמיסה , כלומר. 0ומקטינים את הדחייה האלקטרוסטטית עד כדי , המטענים השליליים

4 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הזיקה )כך הקישור בין הגדילים יהיה יעיל יותר , בעלת חוזק יוני גבוה יותר DNAבה מולקולות ה (. ביניהם תהיה חזקה יותר
אחת הסיבות לכך . לים ייפרדוידהג, בסביבה בסיסית DNAיש לציין כי אם נשים את מולקולות הביים וימנע הוא ייקשר ליונים החיו –של התמיסה ( החיובי)היא כי הוא מקטין את החוזק היוני
. מהם להפחית את הדחייה האלקטרוסטטית
בין VDVקשרי מאחר וקיימים, ידי המעלה את האנטלפיהקשרי המימן הם לא הגורם היח, בנוסף :לסיכום(. הטבעות נמצאות במצב האופטימלי והחזק ביותר לקשרים אלוהזכרנו כי )הטבעות
קשרי המימן וקשרי ה –אנטלפיית המערכת מורכבת ממספר גורמיםVDV ( כאשרבעוד , אלו הם הגורמים לאנטלפיה שלילית שני כוחות. (לאחרונים ההשפעה הגדולה יותר
אומנם אינה מספיקה בכדי , פועלת בכיוון אנטלפיה חיוביתהאלקטרוסטטית דחייה ה . יע על האנטלפיה הכללית של התהליךלהשפ
המולקולות שבמערכת קטן מאשר אם מספר מאחר ו, שליליתאנטרופיית המערכת היא .הגדילים היו נפרדים
G∆לפי המשוואה , בכדי שהתגובה תהיה ספונטנית, לכן = ∆𝐻 − 𝑇∆𝑆 ,עליה להתקיים בטמפ 'תתרחש ', כאשר נעלה את הטמפ(. כך שלערך האנטרופיה אין משקל רב לעומת האנטלפיה)נמוכות
. והם ייפרדו זה מזה –דנטורציה של הגדילים
:שהתפרקה לגדילים DNAמעקב אחר כמות ה
ולכן אם , שהתפרק לגדילים מכילה יותר חלקיקים מאשר המצב לפני הפירוק DNAתמיסה ובה תוך שימוש בערכי בליעת גל באורך , הגרעין שבה בעזרת ספקטרופוטומטר. נבדוק את ריכוז ח
260 nm ,נמצא כי הנתונים יהיו גבוהים יותר לאחר הפירוק.
כך שלא קיים מצב בו , קטן מאוד' הדנטורציה מתרחש בטווח טמפ תהליךכמעט תמיד התהליך –רק חצי מן החומר התורשתי יתפרק לשני גדילים
לכל מולקולת . ויתרחש בצורה מהירה מאוד, יכלול את כל המולקולות
DNA אורך : והיא מושפעת מן הגורמים, בה הגדילים נפרדים, היתוך' יש נק
ההיתוך ' ניתן לראות את תלות נ. A-Tל G-Cס מספר זוגות ההמולקולה ויח
כאשר ככל שהקו עולה מעלה כך נדרשת יותר , בגרף G-Cזוגות ה %ב
(.H∆)אנרגיה
בנוגע לחומר הנלמד בה, תשובות במצגת+ ישנן שאלות .

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
27.10 – 4הרצאה –ביוכימיה
:שעתוק ותרגום, שכפול: תהליכי העברת המידע הגנטי – 24פרק
יצירת סליל –שכפולDNA סליל חדש בעזרתDNA קיים.
תהליך בו נוצרים סוגים שונים של מולקולות –שעתוקRNA – tRNA ,mRNA ,rRNA .
שני הגדילים פעילים ומשמשים –שונים DNAשונות ייווצרו לפי גדילי RNAמולקולות .כתבנית
עיבוד מולקולות –תרגוםmRNA הקומפלקס הריבוזומלי סורק את . ויצירת חלבון
לאחר מכן . המקודד להתחלת יצירת החלבון – AUGעד שמגיע לקודון mRNAמולקולת ה
בעוד , המתאימות לפי התאמת זיווג בסיסיםהאמינו . מביאות את ח tRNAמולקולות .בין החומצות האמיניות האנזימים שבריבוזום יוצרים את הקשרים הפפטידיים
הארכת)אמצע ,(initiation) התחלה –חלקים 3נחלקים ל, כל אחד, שלושת תהליכים אלו
–חשיבות העניין בבקרה שנעשית על תהליכים אלו . (termination) וסוף( elongation, הפולימר .להמשיך או להפסיק את סינתזת הפולימר, להתחיל הגורמים שבזכותם ידוע כי יש
:שכפול
ניתן לראות נתונים מספריים הנוגעים לחלוקת בתמונהוהשוואה בין התהליך בתאים אנימליים ובתאים , התא
:דקייםיחי
אוטידים המחושבת אותם צריך כמות הנוקלי -ובתאים ,109היא לשכפל בתאים אנימליים
.3.9X106חיידקיים היא
תהליך שכפול נוקלאוטיד באנימליים אורך -בתאים חיידקיים התהליך . שניות 60-90כ
.שניות 850אורך כ
תוצאה לא הגיונית –חודשים 5תהליך השכפול בתא אנימלי אמור לארוך כאם נחשב נראה כי ההסבר .בתאים חיידקיים שעות 0.67ו ,בתאים אנימליים שעות 8אורך כתהליך מאחר וידוע שה
בכמה , אלא באופן סימולטני, נוקליאוטיד כפול לא נעשה נוקליאוטיד אחרבעובדה כי השנמצא . אתרי שכפול במקביל
Initiation – תחילת התהליך:
oriאזורי אלפיו, בודד בכרומוזום החיידקי oriקיים . הרצפים בם מתחיל השכפול – Oriרצפי ,ישנם מספר אזורים כאלום אנימליבתאים מאחר ו .םאנימליה הכרומוזומים הפרושים על
שיטה )כיוונית יכולה להיות דו –צורת השכפול . מתאפשר שכפול סימולטני מאזורים שונים
ויכולה להיות ( ישנה התקדמות לשני הכיוונים oriמתוך כל נקודת מאחר ו, חסכונית מבחינת זמן :שנערכו בגנום החיידקי את הצורה הקיימת במציאות גילו לפי שני ניסויים .חד כיונית
והשוו את שלו DNAאת הזמן שלוקח לחיידק לשכפל את ה מדדו – בדיקה באופן חישובי .1 (.הנוקליאוטידים וקצב שכפול כל נוקליאוטידלפי מספר )התוצאות לזמן המחושב
כאשר הם משתמשים בחומרים שבו בכדי ליצור חומרים , חיידקים על מצע רגילגידלו .2של אחד פולס על חיידקים אלו הפעילו. הנוקליאוטידים לשם השכפולוביניהם , שונים
(. המחושבלאחר שעבר הזמן )בסוף ההכפלה ואחד נוסף, בתחילת ההכפלה( H3)טריטיום

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
, י סרט צילום"כך שאם נכניס את המולקולה ונבחן אותה ע, הטריטיום יחדור למולקולה . פולטהוא הרדיואקטיבית שגרגרי כסף ששקעו בעקבות הקרינה נוכל לראות
רק האזורים בהם מתחיל , בתחילת ההכפלה ובסופהמאחר ופולס הטריטיום ניתן רק לאורך המולקולה , אם ההכפלה היא חד כיוונית. י גרגרי הכסף"ונגמר השכפול יסומנו ע
ההתחלה )כאשר מן הצד השני האזור יהיה מושחר פחות , נראה אזור מושחר בודדנוכל , אם ההכפלה היא דו כיוונית(. יך מתרחשים באותה הנקודהוהסיום של התהל
ומכך נובע , נמצאו שני אזורים מושחרים .(סוף והתחלה) אזורים מושחריםלהבחין בשני .כי הכרומוזום המעגלי של החיידק עובר הכפלה דו כיוונית
:אנימלי DNAבנוגע לשכפול
גידלו תאים אנימליים על מצע ובו ריכוז נמוך של –לינארי DNAאת הניסוי ערכו בשנית על
תחילת , בתחילת הניסוי. י הטריטיום בצורה חלשה"יהיה מסומן ע DNAכך כל ה. טריטיום
נראה שני –אם ההכפלה היא דו כיוונית . ניתן פולס קצר מאוד של טריטיום DNAהכפלת ה
לה חד כיוונית נראה סימון שחור אם ידובר בהכפ. replication bubbleסימנים שחורים סביב כל .גם בחיידקים וגם בתאים אנימליים השכפול מתרחש באופן דו כיווני, כלומר .בכל תחילת בועה
מלבד לכרומוזום המעגלי קיים , בחלק מן החיידקים. העניין לא נכון לכל חומצת גרעין, זאת עם
אם נבדוק את ההכפלה (. יכפיל עצמו באופן עצמאחוץ כרומוזומלי המ DNAמקטע )פלסמיד גם . מכפיל עצמו בצורה חד כיוונית נמצא כי הוא, של פלסמיד זה
:oriמטרתו הייתה הגדרת תפקיד הניסוי נוסף ש
פלסמיד זה מסוגל לעבור הכללה לתוך , במצבים מסוימים. פלסמיד בחיידק בעל הניסוי נערךאינו )הפלסמיד מאבד את עצמאותו , בשלב זה. כך שהוא הופך להיות חלק ממנו, הכרומוזום
בשלב זה הקטע שמקורו ) והופך לנתון תחת הבקרה המופעלת על הכרומוזום( מתחלק לבד
, אינו מקרי oriתפקוד המתוך כך ניתן ללמוד ש. (אפיזוםבפלסמיד ועבר הכללה לכרומוזום מכונה אומנם , לעתים הוא היה מתחיל באפיזום ולעתים ברצף הכרומוזומלי, היה מקרי מאחר ואם אכן
. אלא רק של הפלסמיד, לעולם לא תשתנה הכיווניות של הכרומוזום. הדבר לא כך
ח בצורה "קפים בעישנם וירוסים התו, לדוגמא –עניין זה לא קיים רק בחיידקים : הערהבדומה )כך שהוא כבר לא עצמאי , בה הגנום שלהם עובר לכרומוזומים האנימליים
(. לפלסמיד
' אם נעלה את הטמפ –בחלק השני של הניסוי השתמשו בחיידק דומה ולו רגישות לטמפרטורה לאחר' נעלה את הטמפ אם, אומנם. יפסיק להתחלק החיידק( 42ל)ר החד' מטמפ מעלות 5בכ
oriלפי ה, ימשיך להתחלק בצורה חד כיוונית החיידק –הפלסמיד של אותו חיידק עבר הכללה ש :מתוך הניסוי ניתן להסיק שתי מסקנות .של האפיזום
ניס אם נכ .הם לא אתרים אקראיים אלא רצפים מוגדרים וקבועים oriניתן ללמוד שה .1חיל ההכפלה של משם תתגם –קיים להת אמור לארצף שכזה לאזור בגנום בו הוא
.DNAה
oriה את מיקומם של רצפים חלבונים שבעזרתם התא ומנגנוני השכפול מזהים מיקיי .2את לא היו חלבונים שיזהו, למרות שהרצף לא נפגע', פמאשר העלנו את הטכ. הנכונים
כנראה שישנם חלבונים אשר חוסמים . האפיזומלי oriנעשה שימוש בכך ש, המקורי Oriה
. החיידק השתמש בו, מעלות 42ומאחר והם לא קיימים ב, האפיזומלי oriאת ה
ישנם מספר רצפי –בתאים אנימליים העניין יותר מורכב : הערהori , ולמרות שידועיודעים מהם המיקומים של . עד היום לא הצליחו לאפיין אותו, שמדובר ברצף מוגדר

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
אומנם מאחר ומדובר בכמות אדירה של בסיסים עדיין קשה , הרצפים על הכרומוזומים . מדי למצוא אותם
:מושגים מספר
הרפליקון הוא כלל , בחיידק. בודד oriשמוכפלת תחת בקרה של DNAיחידת –רפליקון - .הכרומוזום המעגלי
- Ter – terminator sequence .האזור בו השכפול הסתיים.
.DNAהקומפלקס האחראי על הכפלת ה –רפליזום -
. קומפלקס חלבוני הכולל את הפרימאז וההליקאז –פרימוזום -
:עם הכפלת התא DNAסנכרון הכפלת ה
40נראה כי מדובר בערך ב, אם נחשב כמה זמן לוקח לחיידק להכפיל את כל הכרומוזום שלו. דקות 20נראה כי מדובר ב, אם נחשב את הזמן שלוקח לחיידק להכפיל את התא כולו. דקות
שבו כבר מצוי באמצע DNAה, הוא שברגע בו תא מתחיל את חייו כתא עצמאי ההסבר לכךעניין זה לא . כך בעצם החיידק נמצא בשתי הכפלות בו זמנית. הההכפלה לקראת החלוקה הבא
אחד מן השלבים הוא עלייה . במושבת חיידקים ישנם שלבים שונים של גדילה –מתרחש כל הזמן . ורק בשלב זה מתרחשות ההכפלות המקבילות, לוגריתמית של מספר החיידקים
:ניסוי הנוגע לעניין
סומן בחומר פלואורוסנטי terה רצף, בחומר פלואורוסנטי ירוקהה וסומן זו חיידקי Oriרצף בעת חלוקת התא ניתן היה לראות . הליך החלוקה בתאים חייםכך ניתן היה לעקוב אחר ת. אדום
כאשר התא התחיל להתחלק בפעם השנייה ניתן . אחת Terונקודת ( אחת על כל גדיל) oriנקודות 2
כלומר החיידק לא סיים את ההכפלה הראשונה ובכל זאת –ת בודד Ter' ונק ori' נק 4היה לזהות .כבר התחיל את השנייה
Elongation – הארכת השרשרת המסונתזת:
DNA :סוגים 3קיימים בחיידקים כאשר , פולימראז DNA האנזים י"תהליך זה מתרחש ע
א לקשור וה םתפקיד. בלבד' 3ל' 5פולימראז עובדים מכיוון DNAכל אנזימי ה. 3ו 2, 1פולימראז כלומר הנוקליאוטיד המתווסף . )'3נוקליאוטיד לקבוצת ההידרוקסיד הנמצאת על פחמן מספר
(.זרחן עליו קבוצת, שלו' 5י קצה "מחובר ע
פולימראז DNAמאחר וה. מסמל את הגדיל המקורי שבתרשים החוםהגדיל יש ( אלא רק לגדיל המתארך)לא מסוגל לחבר נוקליאוטיד לגדיל התבנית
יש צורך בפריימר , leading strandה, בשביל הגדיל המוביל. צורך בפריימרהגדיל , לעומתו .'3ל' 5לאחריו השרשרת ממשיכה להתארך מן ה, בודד
נו מוכפל מאחר ואי, זקוק למספר פריימרים, the lagging strand, המפגר
okazaki)המכונים מקטעי אוקאזאקי , אלא במקטעים, ברצף אחד ארוך
model .)כל מקטע זקוק לפריימר משלו .DNA הוא האנזים – 3פולימראז הפריימריםבהמשך התהליך . ומייצר את מקטעי האוקאזאקי, המכפיל את הגדיל המוביל
.האוקזאקים ומחוברים למקטעי מושלמי (שעליהם היה פריימר)מקטעים החסרים וה, מוסרים
כפי .אך בכיוון הפוך, שני חלקים הפועלים בצורה מסונכרנת –פועל כדימר 3פולימראז DNAה
מושכת lagging strandהיחידה המשכפלת את הי כך ש"הדבר מתאפשר ע ,נהושניתן לראות בתמ
. leading strandוכך מתקדמת באותו כיוון אליו מתקדמת היחידה המסנתזת את ה, DNAאת ה

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הפרימאז הוא חלק מקומפלקס . את הפריימרים המסונתזים על הגדילים מייצר האנזים פרימאזאנזים שתפקידו לפתוח את הסליל הכפול בכיוון –בו קיים גם ההליקאז ,הפרימוזום ,חלבוני
צאיםמהפרימוזום נקומפלקס ב. בכדי שיהיה ניתן לסנתז את הפריימרים ואת הגדילים, ההכפלה
לאחר שההליקאז פותח את הסליל . SSB – SS DNA binding protein חלבוני –חלבונים נוספים ם אלוחלבוניאת לכן הוא משאיר –על הפרימוזום לדאוג שהגדילים לא יתאחדו חזרה , הכפול
בנוסף הם גם . ומונעים מהם להיצמד ולחזור למבנה הסליל, אשר נקשרים לגדילים שהופרדו
& stem, לדוגמא –להתרחש בתוך אותו הגדיל העשויים )מונעים מהם לייצר מבנים שניוניים
loop .)החלבונים לא מונעים מנוקליאוטידים חופשיים , יש לציין כי במקביל לפעולות אלו .להשלים את הגדיל
ז פותח את הסליל סיבוב אחר עם התקדמותו ההליקא: ות ההליקאזהערה בנוגע לפעילהאנזים , או ייהרסבכדי שהסליל לא ייתקע . ובכך יוצר מתח בסליל, סיבוב
. והוא סוגר את החתך מחדש, המתח משתחרר, פותח את אחד הגדילים 1טופואיזומראז
מרבית הניסויים המדוברים נערכו בחיידקים ורק לאחר קבלת תוצאות נבדקו : 2 הערה . במרבית המקרים נמצאה התאמה קרובה מאוד. גם בתאים אנימליים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
1.11.09 – 5' הרצאה מס –ביוכימיה
מתאר מצב בו הרפליזום מתקדם ( replication bubbleמנגנון ה) עליו למדנומנגנון ההכפלה הוא DNAוה, בו הרפליזום נותר במקומו, אומנם קיים מנגנון נוסף. ומשכפל אותו DNAלאורך ה
ישנם שטוענים כי הוא ו, טוענים שהמנגנון השני הוא העיקריהישנם אף חוקרים . זה שזז ומתקדם ון הנפוץהמנגנ, לפי מרבית הדעות. למנגנון ההכפלהאך באחוז נמוך יחסית ,מתקייםאומנם
.replication bubbleביותר הוא ה
השונים לא מסוגלים להתחיל את תהליך ההכפלה או לסיים DNA polאנזימי , לפי מה שלדנו :הסיבה לכך נעוצה במנגנון פעולת האנזימים. רים באנזימים אחרים לשם כךונעז, אותו
. '3השרשרת הקיימת מסתיימת בקבוצת הידרוקסיד המצוייה על פחמן מתרחשת התקפה , לאחר שהנוקלאוטיד הנכון זוהה להשלמה
יש לציין כי . על הזרחן של הנוקלאוטיד המצטרף OHנוקלאופילית של ה 4מאחר ו, העובדה שהזרחן יחסית חיובי ההתקפה מתרחשת בזכות
אלקטרוני , בהמשך התגובה. חמצנים מושכים ממנו את אלקטרוני הקשרומשתחררת קבוצת ( הקשר ניתק)אל החמצן עוברים P-Oהקשר
ואטומי הזרחן , P𝛼אטום הפוספט הנותר בגדיל מסומן כ. פירופוספט
. P𝛾ו Pβשבקבוצת הפיפרופוספט מסומנים כ
ליצור תנאים הוא DNA pol 3תפקיד האנזים , בתוך מנגנון זהניתן לשים לב כי האנזים משתמש . מתאימים להתקפה הנוקלאופילית
לפחות נוקלאוטיד )כלומר צריך התחלה של גדיל –קיימת OHבקבוצת ההתחלה של . שכן אם לא תהיה התחלה כזו לא יהיה נוקלאופיל, (אחד
יש צורך כי , בנוסף. י האנזים פרימאז"ע, שסונתז קודם לשכפולהגדיל היא בעצם הפריימר
בכדי לפנות מקום ( פירופוספט במקרה זה)יוכלו לעבור לאזור מסוים P𝛽-Oאלקטרוני הקשר ליצור את הקשר )האנזים לא יכול לסיים את התהליך , מסיבה זו(. קבוצה עוזבת)לקשר החדש
(.ן אין קבוצה עוזבת במקרה כזהשכ –בין נוקלאוטידים המצויים על הגדיל
, יש צורך באנזים ליגאז, DNAלאחר שהנוקלאוטידים של הפריימר הוחלפו בנוקלאוטידים מסוג ולכן אין לו צורך , DNA pol 3מנגנון הליגאז שונה ממנגנון ה. אשר יקשר בין המקטעים השונים
. בקבוצה עוזבת שתיקח אליה את האלקטרונים המיותרים
:ק הפריימריםתהליך סילו
DNA pol 1 לשני אזורים פונקציונלים( לא מבחינת רצף אלא מבחינת פעילות)מחולק:
ומורכב , proof readingאזור ה אחראי על ה – C-treminus – Klenow fragmentאזור ה .1 :בעצמו משני חלקים
. DNA polמקיים את פעילות ההאזור ה .אהם אנזימים המסוגלים לחתוך אקסונוקלאזות – exonuclease '3 .ב
אזור זה . DNAלפרק , DNAנוקליאוטידים בודדים מן קצוות ה .'3אזור מנוקלאוטיד בודד מאפשר להסיר DNA pol 1ב
להוריד נוקלאוטידים משני קצוות מסוגל – אקסונוקלאז' 5אזור ה .2 . DNAאחראי על ההכפלה ועל תיקון .השרשרת הקיימת
Nick translation: נוקלאוטיד אחר , התהליך בו הפריימר מסולק ומוחלףהאנזים , DNAבאזור בו ישנו שבר בגדיל ה. DNA pol 1י אנזים ה"ע, נוקלאוטיד
DNA pol 1 ,מסוגל לסלק את הנוקלאוטיד , אקסונוקלאז' 5בעזרת אזור המסוג )ולאחר מכן לחבר נוקלאוטיד אחר , (RNAמסוג )' 5הראשון שבקצה
DNA) ך אנזים זה ממשיך לאורך כל הפריימרכ. במקומו.

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
ויוצר , RNAבעצם מאקטב את הידרוליזת הפריימר העשוי DNA pol 1האנזים , בתהליך זהבנוי מנוקלאוטידים מסוג מקטע הdNMP .
:פעילות הליגאז
יש , לאחר שסולקו הפריימרים DNAצורך בקישור בין מקטעי ה לליגאז. השונים שבונים את הגדיל
החומצה האמינית על האנזים קיימת .NMPלמולקולות ATPהיכולת לקשור מולקולות ישמסוג ) לנוקלאוטיד ATPקבוצה זו קושרת את ה, מתאימים ATPבתנאי . NHקבוצת ועליה, ליזין
NMP )לאטום הזרחן '3החמצן שעל פחמן יוצר קשר בין האנזים, בעת הקישור. המצוי על הגדיל ,כך שההתקפה –העוזבת את האנזים NTPים מועברים לקבוצת הוהאלקטרונים הנותר
. הנוקלאופילית מתאפשרת
Proof reading:
אומנם לעומתו שיעור המוטציות , קצב הוספת הנוקלאוטידים במהלך השכפול הוא מהיר מאודמקיים מנגנון DNA pol 1מכך ניתן להסיק שה. שהיינו מצפים לראות הוא הרבה יותר קטן
כאשר לאחר הוספת כל נוקלאוטיד הוא בודק שאכן , (proof readingמנגנון ה)בדיקה ותיקון מנגנון . ואם לא הוא מחליף אותו, אם כן הוא ממשיך את הגדיל. מדובר בנוקלאוטיד הנכון
:DNA pol 1שינוי קונפורמציה של י "מתבצע ע proof readingה
כאשר במעבר ישנה , עובר דרך האנזים DNAסליל ה, כפי שניתן לראותנכנס נוקליאוטיד DNAאם לגדיל ה(. מוקפת בעיגול)נישה בלתי מנוצלת
הנוקליאוטיד , עקב כך. ישתנה( של הסליל כולו)מבנהו המרחבי , לא נכוןוהשכפול , יסלק אותו exonuclease '3אזור ה, הלא נכון ייפול לנישה זו
. ימשיך
:האנזים הליקאז
לאחר . ליל כך שיהיו שני גדילים נפרדיםסאנזים זה לפתוח את ה תפקידהוא משאיר אחריו חלבונים , או יצרו מבנה חד גדילי שניוני, בכדי שהגדילים לא יסגרו מחדש, מכן
. ATPההליקאז צורך אנרגיית , בעת פעילותו. SS DNA binding proteins –מסוג
:(י גן שונה"כל אחד מקודד ע) שני חלבוניםההליקאז הוא קומפלקס המורכב מ, בחיידקים
חלבון זה מגייס את החלבון השני , לאחר הזיהוי .oriה' חלבון המזהה את נק .1 .שבקומפלקס
. פוליפפטידים המבצעים את פעולת פתיחת הגדיל 6חלבון הבנוי מ –הקסאמר .2
Prossesivity – תכונה המגדירה את יכולת האנזים להתחבר למולקולתDNA מרחק לאורך' כאשר הוא מתחבר לנק –היא גבוהה מאוד , יכולת הפרוססיביות של ההליקאז לדוגמא. מסוים
. מבלי לפול כלל עד לסיום פעולתו lagging strandהוא מתחיל להתקדם על ה oriה
ומספר על leading strandפריימר בודד על ה –מסנתז את הפריימרים – האנזים פרימאז . lagging strandה
י "משחרר את הלחץ שיוצר ההליקאז ע – 1טופואיזומראז מסוג . ותיקון הגדיל שנחתך, ביטול מספר ליפופים, חיתוך אחד הגדילים
הערה :DNA pol 2 – אנזים זה לא תורם לתהליך השכפול . DNAכאשר עיקר פעילותו נוגעת לתיקון , עצמו

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:DNA pol 1,2,3השוואה בין
Holo enzyme :
אין יכולת DNA pol 3לאנזים , כפי שניתן לראות בטבלהלמרות שיש לו צורך בה יותר משני , פרוססיביות גבוהה
DNAעליו לעבור על כל גדיל ה –האנזימים האחרים עובר על לא DNA pol 3ה, אומנם. במהלך ההכפלה
בו – holo enzymeאלא כחלק מקומפלקס ה, הגדיל לבדעצמו מסומן באדום DNA pol 3האנזים . חלבונים רבים
וביניהן )כאשר רוב תכונותיו , (αתת יחידה )תת . מקורן בחלבונים הצמודים אליו( הפרוססיביות
.pol cנוצרת מן הגן αיחידה ה
האזור )אקסונוקלאז ' 3פעילות הוהיא המקנה לאנזים את , α קשורה לתת יחידה – 𝛆תת יחידה . DNA pol 3ומהווה חלק ממנגנון בדיקת הטעויות של האנזים , (proof readingהאחראי על ה
ויכולת בדיקת הטעויות נובעת כולה מתוך , אקסונוקלאז בעצמו' 3אין אזור DNA pol 3לאנזים . תת יחידה זו
אומנם עוד לא ברור , כאשר ידוע שיש לה תפקיד, 𝛉תת יחידה קשורה , לתת היחידה העיקריתכלשהו שנפגע עקב הסרת תת יחידה זו מן הקומפלקס פחו לזהות פנוטיעד היום לא הצלי. מהו
.החיידקי
כך שנוצר דימר האחראי על , תפקידה לחבר בין שני האנזימים –( מסומנת בירוק) 𝛕תת יחידה . השכפול המקביל
. RNAל DNAבין ה עוזרת לקומפלקס להבדיל, (אדפטור)משמשת כמתאם – Xתת יחידה
קשור holo enzymeבעלת מבנה טבעתי שבעזרתו ה –אחראית על הפרוססיביות –𝛃תת יחידה . ואיתה שאר הקומפלקס, הגדיל בעצם לכוד בתוך הטבעת. בצורה בה לא ינתק ממנו DNAל
DNAהחלבון ממנו מורכבת תת יחידה זו בנוי מסלילי אלפא הידרופוביים המופנים כלפי ה .לקומפלקס לעבוד עליו הובכך לא מפריע, יללגד הלא צמוד הטבעת, בצודה זו. הטעון

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
אחראיות על טעינת הטבעת במולקולת – (מוקפות בעיגול, ועוד כמה) 𝛄ו 𝛅 ,𝐗תת היחידות כאשר מולקולת (. אין צורך באנרגייה)מבלי שתתקיים הידרוליזה , ATPתהליך זה דורש . DNAה
ATP שהן פותחות את טבעת המבנה היחידות המטעינות משתנה כך , נקשרת לאנזיםβ , ולאחריש צורך , כאשר הטבעת נפתחת בשנית, יש לציין כי בסוף התהליך. סביב הגדיל סוגרות אותהמכן
(. כלומר יש שימוש באנרגיה)המתפרקת ATPבמולקולת
Termination :
ממול )בצד השני של הרפליקון , בכרומוזום החיידקי הטבעתי. סיום תהליך ההכפלה. שני הרפליזומים מזהים רצפי סיום אותם – אזורי טרמינציה 7 ישנם( oriה' לנק
הוא מסיים את עבודתו אך לא , כאשר אחד הרפליזומים נכנס לאתר הטרמינציה . עד כי הרפליזום מגיע לאתר מן צידו השני DNAמתנתק מן ה
והדבר מהווה סימן , משתחררות βה טבעות , לאחר ששני הרפליזומים הגיעו לאתר הטרמינציהשחרור הגדילים ) DNAשחרור ה. להשתחרר וכי הוא יכול להשתכפל מחדש DNAלכך שעל ה
בצע חתך דו גדילי ובכך אנזים זה מ. 2י טופואיזומראז מסוג "מתאפשר ע( החדשים זה מזה . להתנתק זו מזו DNAלקולות המאפשר למו

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
3.11.09 – 6' הרצאה מס –ביוכימיה
:תהליך ההכפלה בתאים יוקריוטים
הראשונים עיקריים לתהליך 3כאשר רק ה, פולימראז DNAסוגי 5וקריוטים קיימים יבתאים :השכפול
DNA pol 𝛼 – לאחר . אחראי על סנתוז הפריימרמצוי בתוך קומפלקס עם הפרימאז ה
את מקטעים .dNMPהמורכב מ, קצר מאוד, מסנתז קטע נוסף האנזים, נתזוסהפריימר ש
(. אקסונוקלאז' 3באנזים זה לא קיים אזור ) proof reading אלו האנזים מייצר בלי לבצע
סיביות של פרוסבעל ו, lagging strandמצוי בעיקר על ה הוא, בגלל הקישור שלו לפרימאז . נוקלאוטידים 100כ
DNA pol 𝛿 – ם מחליף את האנזיDNA pol 𝛼 ומסיים את נוקליאוטידיםלאחר מספר
, נמוכה פרוססיביותאנזים זה כשלעצמו בעל . (בחיידקים DNA pol 3מקביל ל) הרצף
פרוססיביותוכך מאפשר , βמתפקד בצורה דומה לטבעת ה PCNAנם בעל חלבון מאו .גבוהה
DNA pol 휀 – בעל יכולות דומות לאנזיםDNA pol 𝛿. אם ניצור מוטציה בDNA , כך
, 𝛿לעומת אנזים . 휀תהליך ההכפלה לא ייפגם בגלל האנזים , DNA pol 𝛿שייפגם האנזים
' 3מקיים פעילות . PCNAגבוהה גם ללא החלבון סיביותפרוסאנזים זה בעל .אקסונוקלאז
DNA pol 𝛽 – שגויים המצויים במרכז מקטע נוקליאוטידיםמחליף , קשור בבקרה (. שאינם צמודים לפריימר)האוקזאקי
DNA pol 𝛾 – מעורב בשכפולDNA אשר זה בעל קוד שונה מן ה, מיטוכונדריאליDNA .לא קיים אנזים חיידקי דומה, מאחר ובחיידקים אין מיטוכונדריות. הגנומי
PCNA:
, בנוסף לכך. 2משרשראות פוליפפטידיות ולא 3מלבד לכך שמורכב מ, βחלבון דומה מאוד לטבעת תפקיד . הרצפים שלהן חסרי דמיון, למרות שהתפקוד של שתי תת יחידות אלו כמעט זהה לחלוטין
לא DNA pol 𝛿 אשר ה, (זאקסונוקלא' 3בעל אזור ) proof readingנוסף של חלבון זה הוא ה
ידות בעזרת תת יח DNAגם חלבון זו מוטען על ה, בדומה למודל החיידקי .צמומסוגל לקיים בע .נוספות
RNase H1:
חד DNAחד גדיליות המוצמדות למולקולות RNAאנזים זה מעכל מולקולות , וקריוטיםיבתאים
אנזים זה פועל בצורה . האחרון RNAמלבד לנוקליאוטיד ה, כך הוא מסיר את הפריימר. גדיליות
יוקריוטים לא מתרחש בולכן , (המקבילה החיידקית שלו מבחינת תפקיד) DNA pol 1שונה מן ה
. nick translationתהליך ה
FEN1:
נתז השס DNAעובר על מקטע ה, מסיר את הנוקליאוטיד האחרון. 'endonuclease 5סוג של
,DNA pol 𝛼 מבצע וproof reading . גםשגוי הוא מסיר אותו נוקליאוטידאם הוא מוצא.
DNA pol 𝛿:

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
מסוג נוקלאוטידים הסיר FEN1אם , בנוסף. הפריימראזור שהוסרו מ הנוקליאוטידיםממלא את
DNA אם . הוא ישלים גם אותם, באזור הקצה FEN1 ימלא , הסיר נוקליאוטידים מאמצע הקטע
.DNA pol βאותם האנזים
:השכפול הלינארי טרמינציה של תהליך
הפריימר הפריאמז מסנתז את שלאחר . בעלי חשיבות DNAגם בקצוות הכרומוזומים ישנם רצפי
. DNAואין אנזים המסוגל לסנתז אותו מחדש כ –הפריימר מוסר , (המצוי בקצה הגדיל)האחרון
עם כל, כך בעצם. אלא רק להאריך שרשרת קיימת, לא יודעים לייצר התחלה DNA polאנזימי ה
DNAבעיה זו שייכת רק ליצורים בעלי . בכל כרומוזום', 5של קצה DNAחלוקה נאבד חלק מן ה . היא לא קיימת, בחיידקים לדוגמא –ארי לינ
:באורגניזמים שונים ישנן צורות שונות לפתור את הבעיה
ים קיימים רצפים החוזרים על עצמם 'בבקטריופאג – (Aתרשים ) T7או T4מסוג ים'בקטריופאג
' 3רצפי ה, מכפיל את עצמו וכבר ישנם שני גנומים DNAלאחר שה. בקצוות הכרומוזומים. כך שהאנזימים יכולים להשלים את האזור בו היה פריימר, באופן זמני' 5משלימים את אזורי ה
התהליך חוזר על . והיא מכפילה את עצמה בשנית, כך נוצרת מולקולה המורכבת משני גנומיםאנזים אחר חותך את המולקולה ליחידות של . עצמו עד כי ישנה מולקולה ובה עשרות גנומים
. תוקף' ים רבים מתוך כל חיידק שבקטריופאג'כך שמתקבלים בקטריופאג –דים גנומים בוד
הכפיל עצמו ' לאחר שהבקטריופאג, לדוגמא –לא כל הגנומים נותרים שלמים : הערההסיבה לכך . 4ולא , נקבל רק שני גנומים שלמים,(מקטעים 4ישנה מולקולה ובה )פעמיים
ורק אחד , (שפעם היה בו פריימר)המרכזי רצף היא שכל שני גנומים חולקים ביניהם את ה .מהם יוכל לקבל אותו
כאשר . יםגדילאחד ה על' 5אלו ישנו חלבון הנקשר לקצה ים בוירוס –( Bתרשים ) אדנו וירוסים
עליו הוא ' 5שממול לקצה ה' 3לקצה (המצוי בתוכו) חלבון זה קושר נוקליאוטיד, מוכפל DNAה הוא נסגר לצורה מעגלית –הגדיל השני נותר חד גדילי . כך שהבעיה נפתרת לגבי גדיל זה, נמצא
. מעגלי DNAכך שניתן לשכפל אותו כ, (קצות הגדיל משלימים זה את זה)
Poxvirus ( תרשיםC )– לא מדובר במבנה דמוי . שני הגדילים מקיימים בקצותיהם מבנה טבעתיהשכפול עצמו נעשה בשני . ולא מורכבת מסליל שלם, טבעת היא חד גדיליתמאחר וה –פלסמיד
לווירוס זה (. מהן ההכפלה היא חד כיוונית oriישנן שתי נקודות , כל כיוון לפי כל גדיל)הכיוונים . אחד חדש ואחד ישן –כך שישנם שני גנומים מופרדים , ישנו חלבון היודע לחתוך כל גדיל בנפרד
רק שכעת הוא יהיה , הוא יודע לסגור את הגדיל כפי שהיה בצורתו הראשונית, לאחר שחתך אותם (. מבנה סמי קונסרבטיבי)מחובר לגדיל החדש
וירוסים מסוג זה ידועים . RNAוירוסים אלו בעלי חומר תורשתי מסוג –( Dתרשים ) רטרו וירוס, בעבר היה קשה ליצור את הקישור בין וירוסים אלו. סרטניים בכך שגורמים להתפתחות גידולים
ההקשר נהיה ברור רק לאחר גילוי האנזים . לבין מחלה שבעיקרה נובעת ממוטציות, DNAחסרי
reverse transcriptase ,מ –וירוס ומאפשר שינוי בכיווניות תהליך השעתוק והמצוי בRNA
. DNAל
(. long terminal repeatsמכונים )וכים החוזרים על עצמם ברטרווירוסים ישנם רצפים יחסית אר
הוא משתמש (Tכ ללימפוציט מסוג "בד)נכנס לתא , HIVלדוגמא , RTכאשר וירוס מסוג
הוא משתמש באנזים , לאחר מכן. בתור פריימר( אליו הוא חדר של התא) tRNAבמולקולות ה
reverse transcriptase ומייצר קטעDNA האנזים . קצר שישלים את הפריימרRNase H1 מעכל

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
, המולקולה מתעגלת, בהמשך התהליך. DNAכך שנותר רק ה, long terminal repeatאת מקטע ה
עד כי ישנו DNAמשלים את ה reverse transcriptaseה. DNAכאשר כבר יש עליה התחלה של
כך שאת התהליך השלם ניתן לתאר , DNAעל גנום זה מסונתז גדיל משלים של . גנום ויראלי שלם
מולקולה זה עוברת אינטגרציה . דו גדילית DNAמולקולת >-חד גדילית RNAמולקולת : בקיצור :כך שעשויים להתפתח שני מקרים, לגנום שבתא בצורה אקראית
והאדם יהיה נשא , (מסלול ליזוגני)המולקולה תעבור אינטגרציה לאזור שאינו מתבטא - . של הוירוס
. כך שהוירוס ימשיך להשתכפל, מולקולה תעבור אינטגרציה לאזור שכן מתבטאה -
:אאוקריוטים
רצפים שאינם –קיימים הטלומרים ( '5וקצה ' 3קצה )האאוקריוטים בקצות הכרומוזומיםרצפים אלו . אלפי פעמים םוחוזרים על עצמ, (לא מקודדים לשום גן) אינטגרלים לכרומוזום
ובכך מונעים ממנגנוני תיקון שבתא לשנות םמופיעים בקומפלקס עם חלבונים אשר חוסמים אות . םאו לחתוך אות
החלק עליו היה )חלק מן הטלומר , עם כל חלוקה. הטלומרים אינם מוסיפים דבר לגנום היוקריוטיהטלומר יאבד כולו , חר מספר חלוקותלא. כל שהטלומר מתקצר, נאבד בתהליך השכפול( הפריימר
. כלומר נאבד גן –והחלק הנוסף אותו נאבד בתהליך השכפול כבר יהיה שייך לכרומוזום עצמו – . וימות, התא לא יוכל להתקיים, במצב כזה
סוג של ) י האנזים טלומראז"הטלומרים עצמם נוצרים ע
DNA pol) .אנזים זה מושתק ברוב , כאשר האורגניזם נולדתאי , אומנם נותר פעיל ברקמות האשכים, רקמות שלוה
האנזים חוזר למצב , כאשר תא הופך לסרטני. תאי גזע, הדם . פעיל ממצבו הרדום
. אנושי reverse transcriptaseהטלומראז הוא סוג של
המשלים RNAמדובר בקומפלקס המכיל רצף כך הוא יכול . לנוקליאוטידים האחרונים שבכרומוזום

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
מסוגל להאריך רק את ) בהתאם לרצף הקיים מולו DNAולהאריך את גדיל ה, אליהםלהיקשר
הטלומראז . dNTPההשלמה נעשית עם . (קיים' 3מוסיף נוקליאוטידים לקצה – של הגדיל' 3קצה .יכול להמשיך ולהאריך את הטלומר בעשרות רצפים
קיים הרצף, אדם בבני. הרצף החוזר על עצמו בטלומר הוא ספציפי לכל מין: 1 הערה :
TTAGGG מסיבה זו טלומראז אנושי לא יוכל . מאות פעמים בקצה כל כרומוזום . להאריך טלומרים באורגניזם אחר
את הטלומרים נוהגים לסמן בעזרת נוגדנים פלואורוסנטים הנקשרים לחלבונים : 2הערהמקטעי –י פרובים "דרך נוספת לסימון הטלומריםהיא ע. המצויים בקומפלקס סביבם
DNA (. כך שניתן לזהותם)מסומנים רדיואקטיבית המשלימים לרצף הטלומר
שאינם )תפקידם של הטלומרים הוא לפתור את בעיית קצוות הכרומוזומים : 3הערה, (ושל התא)הם משמשים גם כמדד לגילו של הכרומוזום , עם זאת(. יכולים להשתכפל
. וכמה עוד נותרו לו, לוקות התא עברמעין שעון ביולוגי המאפשר לנו לדעת כמה ח
הטלומרי DNAארגון ה –מבנה הטלומרים הוא מורכב מאוד הטלומרים מאורגנים . הוא שונה מאשר ברצפים המקודדים
–לדוגמא , גדילים בסליל אחד 3-4במבנים מרחביים הכוללים
.hoogstein pairingבשרטוט ניתן לראות טטראגדיל מסוג

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
08.11.09 –7הרצאה –ביוכימיה
:DNAארגון מחדש של ה – 25פרק
:פרק זה כולל את הנושאים
.DNAרמת בקרה מבנית של ה – DNA( מודיפיקציות)חיתוך ושינוי .1 . DNAמנגנוני תיקון נזקי .2רקומבינציה שבין כרומוזומים הומולוגים או בין כרומטידות אחיות במהלך –שחלופים .3
.המיוזהאו מעבר של מקטעים , מספר פעמים לאורך הגנום חזרה של רצף –אימפליפיקציה .4
(. או בין כרומוזומים)למקומות שונים בכרומוזום מגנום , (שהם חלבונים בעצם)ייצור כמות אדירה של נוגדנים , לדוגמא –ארגון מחדש .5
. שכמותו מוגבלת
:מודיפיקציות
:DNAמולקולות ההמודיפיקציה העיקרית שעוברות : מתילציה
:עובר מתילציה DNAישנם שני אתרים עיקריים בהם ה, בתאים פרוקריוטים
- N6-methyl adenine – חנקן עלC 6' על נוקליאוטידים מסוג אדנין . - N4-methyl cytosine – חנקן עלC 4' שעל נוקליאוטידים מסוג ציטוזין .
:בפרוקריוטים חשיבות המתילציה
יזהה את התא , או שאינו מתאים לרצף, אם נוקליאוטיד הוחלף – DNAתיקון בעיות ב .1 .י מתילציה"ע האזור הדרוש תיקון
.oriזיהוי רצפי ה .2חיידק עשוי לייצר אנדונוקלאזות . ים או וירוסים אחרים'מנגנון הגנה מפני בקטריופאג .3
מן DNAאם כך ש, (ולא רק מן הקצה, גם במרכזו DNAאנזימים היודעים לחתוך )בכדי למנוע . התא ייעכל אותו ולא יאפשר לו להתבטא, יחדור אליו 'הפאג
מתרחשת מתילציה על הגנום של החיידק , החיידקי DNAמהאנדונוקלאזות לעכל את ה
. מפני האנדונוקלאזות DNAמתילציה זו מגנה על ה – N4ו N6באתרים אם נצליח למנוע . י מתילציה"מוגברים ע/ גורמי אלימות בחיידקים מסוימים מתאפשרים .4
.הם לא יהיו אלימים, את המתילציה בחיידקים אלו
ורק אם לפני ציטוזין , methyl cytosine-5מתילציה נורמלית תתרחש רק על , בתאים אנימליים
C(cytosine)p(phsphate)G(guanine)' 3: אתר מסוג זה ייראה כך סימון. זה יופיע נוקליאוטיד מסוג גואנין5' .
לא כלCpG זה יהיה באתר שכזה בלבד, אבל אם מתילציה אכן תתרחש, יעבור מתילציה . בדומה לתאים )לציטוזין ' 3מתילציה תתרחש רק בין גואנין המצוי –בתאי צמחים
. יכולים להיות נוקליאוטידים שונים Gל Cאומנם בין ה, (אנימליים אם באתרCpG גם על אותו אתר היא תקוים, כלשהו על אחד הגדילים תהיה מתילציה
, מתוך עובדה זו ניתן להבין כי המתילציה מועברת מדור לדור. אשר על הגדיל המשלים לו . מתא לתא
מן הגנום עובר מתילציה 3-5%כ. קבוצות המתיל המוספות לגדיל לא מפריעות לזיווג הנוקליאוטידים המשלימים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
י האנזים מתיל טרנספראז"המתילציה עצמה מבוצעת ע .
קבוצות המתיל –המתילציה מספקת רמה נוספת של אינפורמציה גנטית – אפיגנטיתתורשה . ולעתים קובעות האם גן יתבטא או האם יושתק לגמרי, השונות משפיעות על רמת הביטוי של הגן
אנזימי המתיל טרנס פראזות מזהים את –המתילציה עצמה מתרחשת לאחר תהליך השכפול . ולפי זאת מוסיפים קבוצות מתיל על הגדיל המשלים ,המתילים שעל הגדיל הקיים
Adomet (S – adenosyl המולקולה ממנה תילקח קבוצת המתיל היאMethionine) .
:אפיגנטיקה –תפקיד המתילציה
, ('המוגלובין עוברי וכד, דוגמת טלומראז)ישנם גנים המבוטאים בתקופה העוברית העוברות ישנם פקטורי שעתוק על הגנים בזמן . לנצח, אשר מושתקים לאחר הלידה
הפרומוטורים ש כך, תבטאהפקטורים אלו מפסיקים ל, לאחר הלידה. המתורגמיםלאחר . י מתיל טרנס פראזות"מתילציה ע ורבשויים לעוע –של גנים אלו חשופים
חלבונים אחרים נקשרים אליהם ויוצרים קומפלקס אשר לא ,קישור המתילים
. הגן הושתק – DNAהמאפשר כלל שעתוק של
על הכרומטין , באופן רגיל. אנזימים המסירים קבוצות אצטיל –דה אצטילאזות
קבוצות אלו מרווחות . ישנן קבוצות אצטיל( DNAקומפלקסי ההיסטונים המצויים במבנה ה)
, הדה אצטילאזות בעצם גורמות להסרה של קבוצות אלו. וכך מאפשרות לו לעבור שעתוק DNAה
.ולמניעת השעתוק שלו DNAת הובכך לדחיס
פרומוטרים שעברו מתילציה מסיבית לא . CpG islandsמכונים י מתילציה"שהושתקו עאזורים .הגן לא יתבטא לעולם, אם לא יהיה מנגנון שיעשה דה מתילציה. יאפשרו שעתוק של הגן כלל
לגנים שלא יש צורך במוטציה אשר תאפשר, בכדי לפתוח את קומפלקס המתילים והחלבונים (. טלומראז בתאים סרטניים, לדוגמא)להתבטא בתאים מסוימים , אמורים להתבטא
נוכל לייצר , באפיגנטיקה המתילציה תפקידבכדי לאמת את
, שאינם יכולים לעבור מודיפיקציה מסוג זה DNAגדילי . מתבטאים( שלא מתבטאים בתא תקין)ולבדוק האם הגנים
שעברו CpGבתרשים ניתן לראות סליל ועליו שני אתרי כך שנוצר , משמאל ניתן לראות הכפלה נורמלית. מתילציה
ניתן , מימין. גדיל חדש ועליו קבוצת המתיל באותו אתר לראות הכפלה שהתרחשה בנוכחות סוג מיוחד של ציטוזין
(azadeoxycytidine) , בו יש אטום חנקן במקום פחמןבהכפלה . שלא יכולה להתרחש מתילציה כך', 5בעמדה
כך לאחר . גם בגדיל המשלים לגדיל זה לא יהיה מתיל, הבאה( אפיגנטיקה)רמת הבקרה של המתילציה , כמה חלוקות
. וכאשר בדקו ראו כי הגן שהיה מושתק כעת מתבטא, בוטלה
מחלה בה תת היחידות המרכיבות את –טלסמיה
כך שההמוגלובין מורכב , לא מיוצרות βההמוגלובין מסוג
המוגלובין שכזה הוא (. βושתי αבמקום שתי ) αיחידות 4מ . ומוביל חמצן בצורה מוגבלת, מוגבל
אשר הוא דומה , γהתאים מייצרים גלובין , בשלב העוברי
כך שעד ללידה אין -αהרבה יותר מאשר גלובין βלגלובין

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
כך , י מתילציה"האזור המקודד לגלובין גמא מושתק ע, בעת הלידה. פגיעה משמעותית בעוברניסו לגרום לכך שבעזרת מתן , לכן. שההמוגלובין מתחיל לתפקד בצורה לא תקינה
azadeoxycytidine תבוטל השתקת גלוביןγ .כך שחלבונים אחרים כמובן ויש לשמור על :ביטול השתקה של גן בודדדוגמא ל. או גם הםהאמורים להיות מושתקים לא יבוט
את תאים אלו .ולבודד מספר תאים בהם אנו רוצים לבצע את השינוי, תאי מח עצם ניתן לשאוב
אם נצליח . תתבטל בתאים אלו כך שהמתילציה, azacitidineמספר חלוקות בנוכחות נגדל למשךשבו אנו רוצים , מדובר באזור ספציפי)להנדס תא כזה ולהחזירו לאזור ממנו שאבנו את מח העצם
. יהיה מדובר במרפא לטלסמיה –( לטפל
Genetic imprinting:
אחד מן האם ואחד מן , שני אללים, צאצא מקבל שני עותקים של אותו הגן, בכל יצור דיפלואידי. כאשר רק הראשון יתבטא בתאים, אלל דומיננטי ואלל רצסיבי קיימיםמצב הקלאסי ב. האב
מאחר והדבר , האלל שיתבטא לאו דווקא יהיה הדומיננטי ,genetic imprintingבמצב של , אומנם . האם הגיע מן האב או מן האם –תלוי במקור שלו
מתואר מצב בו ישנו עובר , לדוגמא. הגורם לתופעה זו היא המעורבות של מתילציה בביטוי הגנים, כתוצאה מכך. מהי מושתק בחלקויבעוד הגנום הא, עכברי שבו הגנום האבהי נקי ממתילציה
ללא קשר לדומיננטיות –( מתוך הגנים המושתקים)הצאצאים יבטאו רק את האללים האבהיים . או רצסיביות במצב הבלתי מושתק
גן שעבר הטבעה גנטית= י מתיל "גן מושתק ע . Imprinting אם הגן –אומנם הדבר תלוי באופי הגן –יכול להיות אימהי או אבהי
הוא יהיה כזה בכל עכבר , אימהי Imprintingהמקודד לצבע הפרווה בעכבר מקיים .שהוא
עכברים בעלי אלל –דוגמא במצגת יש שאלת: הערהA ומיננטי במצב ד)לאוזניים כחולות
לזוג עכברים ישנם שני צאצאים (. רצסיבי במצב הרגיל)לאוזניים ירוקות aואלל ( הרגילההסבר לכך הוא . כאשר כל אחד מן הצאצאים בעל אוזניים בצבע שונה, הטרוזיגטיים לגן
. וכל צאצא קיבל מן האם את האלל הקובע, שהגן עבר אימפרינטינג ה עובר לא כל גן שעבר מתילצי: 2הערהimprinting .
:תאור ניסוי
אם . שעות הצלחת תתמלא כולה בחיידקים 24תוך כ, בצלחת פטרי Kמזן Ecoliאם נזרע חיידקי כל . במצע החיידקים( חורים)שעות נוספות נראה פלאקים 24לאחר , ים לצלחת'נוסיף בקטריופאג
בחור יש רק –כך שהם מתו , שהדביקו את החיידקים באזור החור ים'פלאק נוצר מבקטריופאג . ים'פאג
לא , Bמזן Ecoliים אלו ונזרע אותם על צלחת פטרי שגדלים בה חיידקי 'אם נעביר חלק מפאגאם . אנו עשויים לבסוף למצוא פלאק בודד, ואם נחזור על התהליך פעמים רבות, נראה פלאקים
נקבל , Bמזן Ecoliונזרע אותם על צלחת פטרי חדשה ובה חיידקי ים 'מתוך פלאק זה נוציא פאג . פלאקים רבים
לא – Kמזן Ecoliונעביר אותו לצלחת בה חיידקי , Bמזן Ecoliשגדל על מצע ' אם ניקח פאג . או שנקבל פלאק בודד לאחר מספר ניסיונות, נקבל פלאקים
:הסבר תוצאות הניסוי

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
. של וירוסים החודרים לחיידק DNAאנזימים המעכלים רצפי –בכל חיידק ישנם אנדונוקלאזות ובכך , החיידק גורם למתילציה על הגנום שלו –בכדי שהנוקלאזות לא ייעכלו את הגנום החיידקי
Kהעברנו את הוירוסים מזן , כך כאשר(. שיש עליו מתילים DNAהנוקלאזות לא ייעכלו )מגן עליו הם לא –( אשר יודע להגן על עצמו מפני וירוסים אלו) Bלזן ( םאשר לא יודע להגן על עצמו מפניה)
באחד החיידקים כנראה , לאחר שניסינו פעמים רבות, אומנם. הצליחו להתרבות וליצור פלאקים
לא ' של הפאג DNAה, כך. 'של הפאג DNAכך שהוספו מתילים גם ל –השתבש מנגנון המתילציה
כאשר העברנו וירוס (. מוגן DNAאשר גם לצאצאיו יש כ)והוא המשיך להתרבות בחיידקים , עוכל
הם לא יכלו להגן – Bלמצע ובו חיידקים מזן ( שלו כבר יש מתילים DNAכך שעל ה)מפלאק זה . על עצמם כי הוירוס כבר היה מוגן מפני האנדונוקלאזות
אם נעביר את , (על רצפים שונים בגנום) Kובזן Bמאחר והמתילציה מתרחשת בצורה שונה בזן
של DNAיידע לעכל את ה Kזן – Kלצלחת ובה הזן ( שלו עבר מתילציה DNAשה)הוירוס המוגן
. ישתבש Kאלא אם כן התהליך יתרחש שוב ומנגנון המתילציה של אחד מחיידקי –' הפאג
:אנזימי רסטריקציה
באתרים DNAהחותכים את רצף ה ,הם אנזימי רסטריקציה BamH1 ,EcoRIהאנזימים (. דוגמא בתרשים)כל אנזים שכזה בעל אתרי חיתוך הספציפיים לו –מסוימים
, בעזרתם ניתן לחתוך גנום באתרים מסוימים –אנזימים מסוג זה הם הבסיס להנדסה הגנטית
במידה )הרצוי DNAולבודד את מקטע ה, להריץ את המקטעים שמתקבלים באלקטרו פורזהולשלב גן זה בגנום אנושי , אם נצליח לבודד גן רצוי מתוך גנום חיידקי(. וידוע גודלו מראש
ניצור גנום אנושי מהונדס , (להוסיף גן זה לרצף הומני ולחבר את שני הרצפים בעזרת ליגאז) . ומשופר
:דוגמאות לשימוש באנזימי רסטריקציה למטרה רפואית
חזירים וכד, נהגו לקחת אינסולין מסוסים, נסולין לחולי סכרתבכדי לייצר אי, בעבר' .הוא , ח אלו לא דומה לחלוטין לאינסולין האנושי"מאחר והאינסולין המיוצר בבע, אומנם
הפיקו את , בכדי לפתור בעיה זו. גרם לתופעות לוואי ולאלרגיות שונות בקרב המטופליםפלסמיד זה . וסיפו אותו לתוך פלסמידוה, הגן לייצור אינסולין מתוך הגנום האנושי
. שאינו גורם לתופעות לוואי או לאלרגיות –אשר מייצר אינסולין אנושי , הוחדר לחיידק שיטה בה נהגו )לא משתמשים בנגיף מוחלש –בצורה דומה מייצרים חיסון כנגד צהבת
._____________________אלא ב, (המכילה סיכון מסוים, בעבר

1
DNAשיעור השלמה בנושא מנגנוני תיקון ב –ביוכימיה
חמור בהרבה מנזק DNAאומנם הנזק שיגרם למולקולת –כל מולקולות התא חשופות לנזקים
DNAוניתן לייצר אותן מן ה, כל המולקולות הן זמניות DNAמלבד . שייגרם לכל מולקולה אחרת
היא הבסיס לכל המולקולות ולכל החלבונים שייוצרו בתא DNAמולקולת ה. אם יוצרו באופן שגוי
של תא DNAבמקרה הטוב מוטציה ב. ולכן שגיאה בודדת עשויה לגרום לנזק רב אשר הוא קבוע – .אומנם במקרה הגרוע יותר הנזק יוכל לעבור לדור הבא, מותתגרום לו ל
:בשני מקורות DNAנזקי ותמקור
בגלל כמות , למרות שישנם מנגנוני תיקון. ההכפלה בזמן DNA polטעויות של אנזימי ה .1עדיין נשמרות ,והמהירות הגבוהה של התהליך, הנוקליאוטידים האדירה המוכפלת
.טעויות
.וירוסים, חומרים כימים, חשיפה לקרינה –מקורות חיצוניים .2
היא נראה כי שכיחות השגיאה E.coliנוצרים באחר קצב הנוקליאוטידים השגויים ה אם נעקובעשויות אומנם לאורך זמן –מדובר בדיוק רב . נוקליאוטידים נכונים 10,000בנוקליאוטיד אחד ל
רואים כי הוא ,(שלא תוקן)הנותר בפועל כאשר בודקים את אחוז הטעויות. להצטבר טעויות רבות . הרבה יותר קטן
:ניתן לחלק לשני סוגים DNAאת נזקי ה
.נזק המתרחש בגדיל בודד .1
2. DS breaks- הדורשים מנגנוני תיקון מיוחדים, שברים דו גדיליים .
:מספר משפחות של מנגנוני תיקון שגיאות בגדיל בודד
1. Direct repair – התרחשה מודיפיקציה שגויה על אחד הנוקליאוטידים מתקן מצב בו.
, פולימראז DNAלדוגמא אם קבוצת מתיל מצויה על הגדיל ובכך משבשת את פעולת ה .ון תיקון זה פשוט יסיר את קבוצת המתילמנגנ
2. NER (nucleotide excision repair )– כל –במקרה בו לא ניתן לתקן את הגורם להפרעהונוקליאוטידים חדשים משלימים אותו בעזרת הגדיל , הרצף המופרע מוצא מן הגנום
. המשלים לו
3. BER (base excision repair )– התא ישתמש , אחדנוקליאוטיד אם ישנה שגיאה בבדומה , כ"בד .מוצא ומוחלף בנוקליאוטיד תקיןוקליאוטיד השגוי נבו ה, זהבמנגנון
ע במקט לא מדובר .אוטידים גם אם רק אחד שגוייוחלפו מספר נוקלי ,לשיטה הקודמת
.(NERבניגוד ל) אלא במספר נוקליאוטידים בודדים, שלם
הוא יחדור למולקולת , יל אחד תקיןסלכאשר ישנו רק –מנגנון המבוסס על רקומבינציה .4
אומנם מגבלתו הוא כי הוא , צויןמדובר במנגנון מ. השגויה וישמש כתבנית לתיקון DNAה
לא עבר DNAבמצב בו ה –יוכל להתרחש רק במצב בו קיימות שתי מולקולות דו גדיליות שני ילים ולא בין תתרחש רק בין שני סל)לא תוכל להתרחש רקומבינציה –שכפול
ולא, Sאו בשלב ה, G2מנגנון זה יתקיים רק לאחר שלב המסיבה זו (. גדילים משלימים
.G1בשלב
5. Miss match repair – במקרה בו המנגנון זה יתקייםDNA פולימראז הכניסלדוגמא אם היה )נוקליאוטיד שגוי כך שמתקיימת אי התאמה בין הגדילים המשלימים
(.AGובמקומו נוצר , ATצריך להיות זוג

2
:המנגנוניםפירוט 0
Direct repair:
שרמתה הולכת – UVהשמש חושפת את תאי הגוף לקרינת קרינה זו בעלת עוצמה יחסית . וגדלה עם הירידה ברמת האוזון
השיבוש . DNAב ושונים רבים והיא גורמת לשיבושים, בוההגעל פני גידל של פירימידין דימריםהנפוץ ביותר הוא יצירת
, בו שני פירימידינים, ניתן לראות בתרשים גדיל בודד. אחד
נוסף קשר קוולנטי יוצרים, UVאשר בעקבות החשיפה לקרינת –קישו זה יכול להיעשות בשתי צורות . אלא גם דרך הבסיסים החנקניים לא רק דרך הסוכר –
של 4של פירימידין אחד לפחמן 6בין פחמן והשנייה יוצרת קשר ,אחת יוצרת מבנה ציקלובוטאן
. (Cכ יהיה "שבד) שניהפירימידין ה
כאשר אנזימי ,כך. helix distortion – הדימרים גורמים לשיבוש בארגון המרחבי של הסלילפול או כוייתקעו בעת כל ש, לעבוד יוכלוהם לא או השעתוק צריכים לעבור על הגדילההכפלה
התא –ולא יכול לשעתק חלק מחלבוני התא , הכפיל עצמוכתוצאה מכך התא לא יכול ל. שעתוק .ימות
בצורה יחסית טובה עם יתן לראות כי חיידק זה יכול להתמודדנ, E.coliמתוך מחקר שנעשה בהוא –( מעל סף מסוים דימרים אלו כן יהיו לטלים) כמות מסוימת של דימרים מסוג ציקלובוטאן
ם התא החיידקי יתקשה ת הצורה השנייה של הדימריאומנם אם נוצר. מסוגל לתקן ולסלק אותםלא –מצד אחד השגיאה הגנטית לא תועבר לדור הבא –הדבר יכול להיות חיובי או שלילי . יותר
. שני החיידק מת מצד. יתבטא שינוי פנוטיפי
:מנגנוני התיקון השונים שנועדו להתמודד עם הדימרים באופן ישיר
1. DNA photolyase (photoreactivating enzime):
יודע להשתמש באנרגיה של האור הנראה על מנת לפתוח את הפוטוליאז נזיםאמנגנון זה יתקיים רק בפרוקריוטים או ביצורים יוקריוטים . מבנה הציקלובוטאן
. להמיר אנרגיית אור לאנרגיה כימית המסוגלים, (צפרדעים לדוגמא) נחותים
ולא)נוצר דימר מסוג ציקלובוטאן לאתרים בהם האנזים נקשר באופן ספציפיאותו האנזים מבצע ,הוא לא תלוי אור הקישור תהליך. (דימרים מן הסוג השניל
הקישור יש לו צורך באנרגיה על מנת לשבור את הקשר רחלא, אומנם, גם בחושך רצפטורים לאור שבתא מעבירים את האנרגיה שקלטו. בין שני הפירימידינים
בהמשך הוא ישתמש באנרגיה זו לפתיחת . ותה לאנרגיה כימיתהממיר א לאנזים. ובכך לביטול הדימר ולחזרה למצב התקין, הקשר
.מנגנון זה לא קיים ביונקים .הנוקליאוטידים בהסרתצורך אין – המנגנון ישיר כפי שניתן לראות
:מנגנון שני .2
גובה של אנזימיםעשויה לגרום לת או למולקולות מסוימות חשיפת התא לגירויים חיצונייםמודיפיקציה זו של .נוקליאוטידיםהעל ( הוספת קבוצות אלקיליות מסוג)המבצעים מודיפיקציות
, נפוצה למקרה כזה דוגמאמשמאל ניתן לראות .קישור קבוצות אלקיליות יכולה לגרום לנזק רב. (6 חמצןעל )לגואנין קבוצת מתיל להוספת סוג זה תגרוםמהתא לריאגנט חשיפת בו
שתי לכך ו ,לאחר המודיפיקציה לא יזהו את הבסיס DNAאנזימים הפועלים על ה, במקרים רביםאו לחלוקה , לדור הבא מוטציהאו יעביר את ה, עקב המוטציה התא ימות – תוצאות אפשריות

3
להיות מוטציות מסוג חלקן יכולות, א תוצאה של הצטברות מוטציות רבותסרטן הו, כידוע .הבאה
DNAהדי שבכ, ים חושפים את החולה לחומרים כאלובמקרים רב, בטיפול בסרטן. אלקילציהי "ע יפתח מוטציות נוספות שיגרמו למות התאים הסרטניים, את הריאגנטים הפגום יקלוט
.(DNAפול השכ, חלוקהעת ההמוטציה מתבטאת רק ב)הפגיעה בחלוקה שלהם
–מאחר והחלוקה בתאם סרטניים מתרחשת בתדירות גבוהה הרבה יותר מתאים אחרים . לאלקילציה תפגע יותר בתאים הסרטניים מאשר בתאים אחרים החשיפה לריאגנטים הגורמים
נוטים גם הם –תאי מעי , תאי עור, תאי מין, תאי דם כי –טיפול זה מקיים בעיה של ספציפיות . ולכן יפגעו מטיפול מסוג זה, להתחלק בתדירות גבוהה
:פעילות המנגנון
בתא שייחשף תתרחש הכפלה –פולימראז DNAלא תמנע את פעילות ה קבוצת האלקיל
כך שהוא יכניס בשכיחות גבוהה DNA polקבוצה זו תבלבל את ה, אומנם. לריאגנט
עשוי לגרום לשינוי טוטאלי הדבר . GCבמקום ATבדור הבא נקבל , כך. Cבמקום Tמאוד
הוא יכול למנוע – ORFהגם אם הנוקליאוטיד הוא לא חלק מן . של החלבון שנוצר ךלגרום להם להיקשר אל הגן כשאין בכאו –מפקטורי שעתוק שונים להיקשר אל הגן
. ובכך לפגוע בבקרת השעתוק, צורך
:עם מוטציה מסוג זה התא התמודדות
היודע להסיר את O6 alkyl guanine transferase אנזיםהקיים , גם בתאי האדם, בכל התאים
הקבוצה האנזים נקשר לנוקליאוטיד בעל. גואניןב O6מחמצן , או כל קבוצה אלקילית, המתילר ציסטאין שבאתר הפעיל של וקושר אותה לשיי, מנתק את הקבוצה האלקילית, האלקילית
ולא –האנזים סיים את תפקידו , לאחר מכן. DNAמן ה הקבוצה האלקילית הוסרהשכך , האנזיםובל לכו הוא יו –אין שום מנגנון המסיר את הקבוצה האלקילית מן האנזים –יוכל לתפקד שוב
O6 alkyl guanineיצירת אנזים במאקטאנזים זו , לפחות בחיידקים, לפני הפירוק. לפירוק
transferase (. ג זהסומאקטב שעתוק של מולקולות נוספות מ) חדש
התא לא הסיר את הנוקליאוטידים אלא –גם במקרה זה נעשה תיקון ישיר . פתר את הבעיה של הנוקליאוטידים השגויים
:NERמערכת
אוד דומה שלו בתאים מ אומנם מצאו צורה, בחיידקים נחקר NERגנון נהממדובר במנגנון תיקון . בתאי האדם גםוצורה דומה ומעט מורכבת יותר , יוקריוטים
. בו המקטע הפגום מוסר מן הגנום ומוחלף, לא ישיר
מרכז מהווים שלושה , גנים מרכזיים הפועלים במנגנון זה 5בחיידקים ישנם
DNAסורק את מולקולת ה uvrBו uvrAהקומפלקס . uvrA, uvrB, uvrC –המנגנון . ילית גדולה או דימר של פירימידיןה אלקקבוצ –בחיפוש אחר מוטציה כלשהי
DNAמולקולת ה, ATPותוך פירוק , את השגיאה הוא נעצר מזהה כאשר הקומפלקסלאחר היווצרות שינוי זה . שינוי במבנה המרחבי של המולקולה –תיצור כיפוף
אנזים העוזב את הקומפלקס ובמקומו נכנס uvrAהאנזים , (בעזרת הקומפלקס)
uvrC . מסוגל לחתוך את ה –פעילות של נוקלאז החדש בעלהקומפלקסDNA .נוקליאוטידים 3-4בחיידקים הקומפלקס יחתוך את המקטע הבעייתי במרחק של כ
(. נוקליאוטידים 13של ככ הסרה"סה)' 5נוקליאוטידים בכיוון 8-9ו', 3מכיוון

4
ישתחררו קשרי המימן המחברים בין הקטע , (בעל פעילות של הליקאז) uvrDבנוכחות האנזים
DNAבעזרת . uvrBבתהליך זה משתתף גם האנזים . סרוהקטע השגוי יו –החתוך לקטע המשלים . וייסגר מאוחר יותר בעזרת ליגאז, פולימראז הקטע יושלם
:שני הבדליםל מלבד, מנגנון זה באדם מאוד דומה
שיצר את לא אותו קומפלקס –הוא קומפלקס אחר המתפקד כנוקלאזהקומפלקס - . הקישור והחיתוך
20הוא כ downstreamרק מקטע ה)נוקליאוטידים 13סר יותר ארוך מהמקטע המו - (. נוקליאוטידים
הסיבה לרגישות . UVרגישות גבוהה מאוד לקרינת מאופיינת במשפחת מחלות הגורמות ללבקנות
. חסר uvrאחד אנזימי –הגבוהה היא מחסור באחד או יותר מן המרכיבים של המנגנון שתואר גם ביום שמש רגיל מתרחשים כל –יא מאוד גבוהה החולים לסרטן העור הרגישות , כתוצאה מכך
.שלא מתוקנים DNAהזמן נזקי
פעיל בצורה NERמנגנון ה, לדוגמא ההטרוכרומטיןאזור , נזק מתרחש באזור לא פעילאם ה: הערהבעיקר מאחר והגישה –אנזימי התיקון יעילים יותר היכן שנדרשים , כלומר. הרבה פחות יעילה
כאשר, ייפתח ההטרוכרומטין נזקים כאלו יבלטו במקרה בו. כה מאודנמו להטרוכרומטיןשלהם .יהיה צורך לתרגם אותו
:BERמנגנון
עלולה להתרחש , לכן. (DNAמסוג U נוקליאוטיד) dUMP בתא מיוצרים גם נוקליאוטידים מסוג
ולכן נדמה –ים ידים אלו מאוד דומשני נוקליאוט. Tבמקום Aמול Uיוכנס בעקבותיהטעות
ובהכפלה הבאה – שגיאהפולימראז לא יזהו RNAוה DNAה –בעיה המוטציה לא גורמת לכאילו
. Tבמקום Uבתא קיימים שני מנגנונים המונעים כניסת למרות זאת. Aיחזור להיות Uה
א יוכל להגיע ל Uכך נוקליאוטיד . ופירופוספט dUMPל dUTPהיודע לפרק אנזים – מנגנון ראשון .מנגנון זה מונע את המוטציה .לשרשרת הגנום
:פעילות המנגנון. Tומכניס Uהמרחיק את ה BERמנגנון ה –מנגנון שני
הבסיס החנקני הקשר הגליקוזידי שבין את חותך uracyl DNA N glycosylaseהאנזים .1 . מופרד מן השלדהבסיס –לריבוז
apyrimidinic, אנזים שני, (פירימידין-א)לאחר שהושג ריבוז חסר בסיס חנקני .2
endonuclease ,לאפירימידין' 5פירימידין וחותך את הקשר הגליקוזידי -מזהה את הא ,
. DNAחתך ב, nickכך שנוצר
פעילות מסוג ) 1פולימראז DNAי ה"המבוצע ע, nick tranlstionבחיידקים קיים מנגנון ה .3ולהכניס ,פירימידין-את הא, נוקליאוטיד להסירבעזרת פעילות זו האנזים יודע (. '5
וישלים , נוספיםנוקליאוטידים מספר יסיר נזיםאכ ה"בד. במקומו נוקליאוטיד חדשוהליגאז יסגור , (סיביות נמוכהבעל פרוס)כן הוא ייפול מן הגדיל לאחר מ. ותם בחזרהא
. את הרווח
? לו המפריעכ לשנות מוטציה שלא "מדוע התא מתאמץ כ
–במקום נוקליאוטיד אחר מופיע Uאומנם לעתים ה. באמת אין בעיה – Tנכנס במקום Uאם ה
עובר דה אמינציה Cכאשר (. מחליפה קבוצת אמיןמולקולת מים ה)שעבר דה אמינציה Cלדוגמא

5
המנגנון לא התפתח יק כימכך ניתן להס. שינוי ברצף, מדובר במוטציה רצינית – Uלהוא הופך
, Uהמנגנון לא יודע מה מקור הו מאחר. Cמפני החלפת אלא בכדי להגן Tבכדי להגן מפני החלפת
. Tיתקן גם החלפות של הוא ,רק כי עליו להחליף אותו אלא
:סילוק דימרים של תימין – BERדוגמא נוספת לשימוש של התא ב
אומנם – דימרים של פירימידין לא קיים בחולי הלבקנותהמנגנון הרגיל המטפל ב
מנגנון זה חולים אלו לא בזכות. שנוצרים Tה דימרימ עושה זאת בחלק BERמנגנון תיאור מנגנון .שורדים ומגיעים לגיל בגרותאלא , שנה בגיללוקים בסרטן עור
:Tהדימר הרחקת
בין הסוכר לבסיס החנקני ש הגליקוזידי מנתק את הקשר N glycosylaseה .1
(T) – ה נותק אחד ממרכיביT.
יחתוך apyrimidinic endonuclease (deoxyribophosphodiesterase )ה .2כך מתקבל נוקליאוטיד ועליו . שבגדיל עצמוהפוספודיאסטרי את הקשר
סים חנקניים המנותק מן השרשרתשני בסי
. המנגנון ימשיך כפי שתואר קודם .3
:עקת חמצון
לא עקב מוטגן )במהלך המטבוליזם הרגיל של התא . בעיה מאוד קשה בתאים
–reactive oxygen species (ROS )ים מיוצר( קרינה או חומר כימי, כלשהות לגרום לחמצון ועשויו, מולקולות מסוג זה הן פעילות מאוד. מולקולות שהן רדיקליים חופשיים
חומרים שהוא להגן על עצמו מפני התא כך על. מודיפיקציה מאוד מוטגנית =של הנוקליאוטידים
י"ע בתנאי חמצון שתיווצר, oxoguanine 8היא הביותר מוטגניות אחת המולקולות ה. צריימ, בעת שכפול היא כיהסיבה לכך שחומר זה מאוד מוטגני . 8אטום חמצן לעמדה קישור
. Cבמקום Aנוקליאוטיד זה עשוי לקשור
.מולקולה זו נוצרת בתדירות גבוההו מאחר בני אדםעד לחיידקים וזו מאוד נפוצה מבעיה
:התמודדות התא עם נזק חמצוני מסוג זה
:תא נעזר בהם בכדי לפתור את הבעיהגנים שה 3ישנם Ecoliבחיידקי
בכך הוא –ופירופוספט dGMPל oxodGTP 8הגורם לפירוק , mutT, תוצר הגן הראשון .1פעילות גן זה בעצם נועד למנוע את. זה לתא ןקליאוטיד מחומצמונע הכנסה של נו
.המוטגן
. mutYאו mutMיפעל אחד מן האנזימים , DNAבמצב בו מולקולה זו כבר נכנסה לתוך ה .2מנגנון הראשון מזהה את המולקולה ומסלק אותה בעזרת
BER . בכך הוא נותן לתא הזדמנות להכניס נוקליאוטיד . גואנין תקין לרצף
oxo guanine 8מסלק את הנוקליאוטיד שמול ה mutYה .3
מדובר . Cויאפשר לתא להכניס , Aכלומר יסלק את ה –
ולכן – Aאו Cבהכפלה הבאה יוכל להיכנס –בפתרון זמני גם אנזים זה . על האנזים יהיה לפעול כל הכפלה מחדש
אומנם לא על הנוקליאוטיד – BERמשתמש במנגנון .הבעייתי
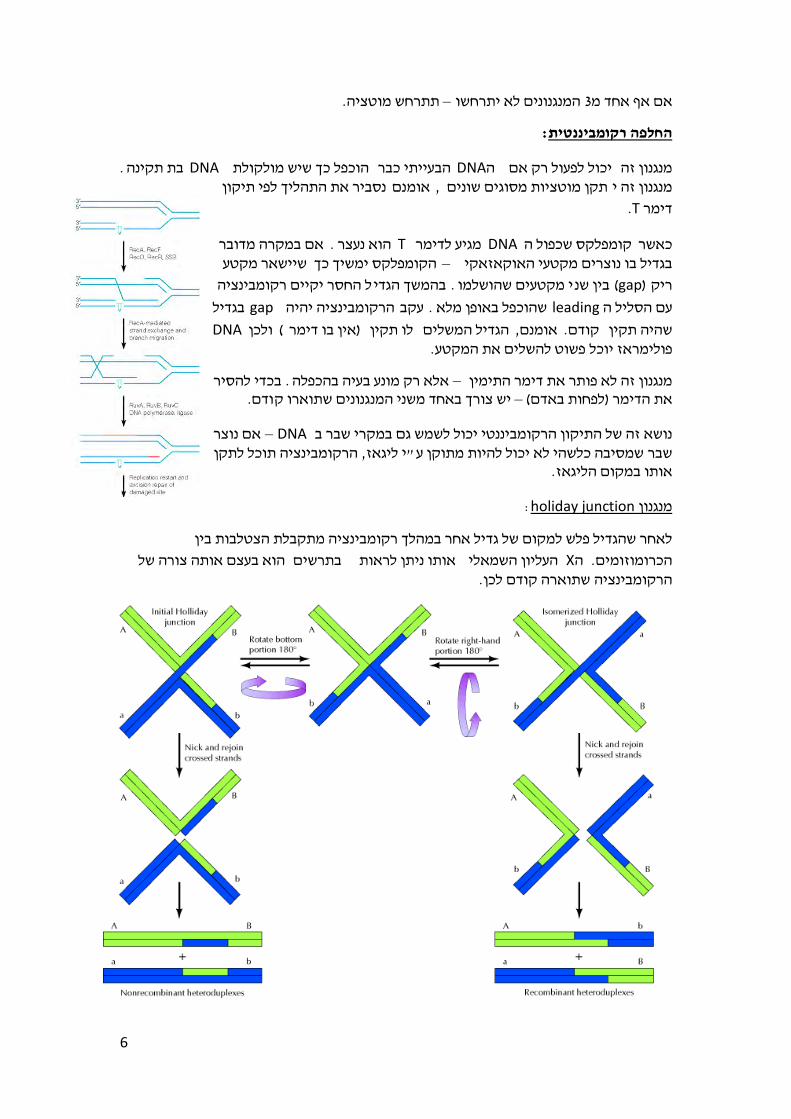
6
. תתרחש מוטציה –המנגנונים לא יתרחשו 3אם אף אחד מ
:החלפה רקומביננטית
. בת תקינה DNA הוכפל כך שיש מולקולתהבעייתי כבר DNAהיכול לפעול רק אם מנגנון זה נסביר את התהליך לפי תיקון נםאומ, תקן מוטציות מסוגים שוניםמנגנון זה י
. Tדימר
אם במקרה מדובר . הוא נעצר Tמגיע לדימר DNAקומפלקס שכפול הכאשר הקומפלקס ימשיך כך שיישאר מקטע –בגדיל בו נוצרים מקטעי האוקאזאקי
ל החסר יקיים רקומבינציה בהמשך הגדי .י מקטעים שהושלמובין שנ( gap)ריק
גדיל ב gap הרקומבינציה יהיה עקב . שהוכפל באופן מלא leadingעם הסליל ה
DNAולכן (אין בו דימר) לו תקין הגדיל המשלים, אומנם. קודםשהיה תקין . פשוט להשלים את המקטעפולימראז יוכל
בכדי להסיר . אלא רק מונע בעיה בהכפלה –מנגנון זה לא פותר את דימר התימין .ודםגנונים שתוארו קיש צורך באחד משני המנ – (לפחות באדם)הדימר את
אם נוצר – DNAנושא זה של התיקון הרקומביננטי יכול לשמש גם במקרי שבר בהרקומבינציה תוכל לתקן , י ליגאז"שמסיבה כלשהי לא יכול להיות מתוקן עשבר
. אותו במקום הליגאז
:holiday junctionמנגנון
לאחר שהגדיל פלש למקום של גדיל אחר במהלך רקומבינציה מתקבלת הצטלבות בין
הוא בעצם אותה צורה של בתרשיםאותו ניתן לראות העליון השמאלי Xה. הכרומוזומים. שתוארה קודם לכןהרקומבינציה

7
השמאלי Xחלק התחתון של השל ה 1800שנוצר אחרי סיבוב , אופקי מבנה פתוח= המרכזי Xהלהיות פתיחה צריכה , מאחר והאנזימים לא יודעים לחתוך את הצורה המלופפת(. ההתחלתי)
אם יתרחש חיתוך אנכי ולא . נקבל את הפתיחה השמאליתזי המרכ Xבמצב של ה. לכיוון כלשהוגדיל אחד –רקומבינציה חלקית במקרה הראשון התרחשה . אופקי נקבל את הפתיחה הימנית
שום גדיל לא –הצורה השנייה גורמת לרקומבינציה מלאה , עם זאת. נותר כמו הגדיל המקורי . זהה לגדילי המקור
:Miss match repairמנגנון
התרחש זיווג לא DNAבמולקולת ה במקרים בהם. מנגנון זה קיים גם בתאים יוקריוטים עילאיים
אומנם לעתים , BERשרות להשתמש במנגנון הנכון בין נוקליאוטידים יש אפ . מנגנון זה יוכל לפעול רק מיד לאחר השכפול .יש צורך במנגנון שונה
להבחין מי מבין הנוקליאוטידים המקיימים לקיים את המנגנון יש בכדי
miss match אנזימים ממשפחת הי "שתעשה עפעילות – הוא השגויmut –
mutS, mutH, mutL. שלבי המנגנון :
. miss match אחר ומחפש DNAרץ על ה, mutSדימר של שני .1
בעקבותיו, DNAשל ה נוצר כיפוף ATPהידרוליזת במקביל ל .2 .האחריםים מתווספים שני האנזימ
כאשר אזור –לשני כיוונים DNAמושך את ה הקומפלקס שנוצר .3
הקומפלקס ממשיך ומושך את הגדילים זה . במרכז missmatchה
או upstreamמשנה אם לא , GATCמזה עד כי הוא פוגש ברצף
downstream .
אומנם האדנין השני –כך על האדנין הישן יש מתילציה . תהליך זה מתרחש מיד לאחר השכפול. תנכנס בטעו מהם הוא הנכון ומילזהות מי ניתן ו –עדיין לא הספיק לעבור מתילציה ( החדש)
. (אדום)בו האדנין לא עבר מתילציה הוא יחתוך את המקטע GATCכאשר הקומפלקס מגיע לרצף
.nickפולימראז ישלים אותו מחדש וליגאז יסגור את ה DNA, מאחר והגדיל המקורי תקין
וא המקורי הקומפלקס לא ידע איזה מן הגדילים ה –אם המנגנון לא יתרחש מיד לאחר ההכפלה . ועשוי לגרום להוספת מוטציה לגנום –
DS breaks:
שברים דו גדיליים . נגעו לנזקים שהתרחשו בגדיל בודדכל המנגנונים עליהם דיברנו עד עכשיו
(DSB )מקרים מסוכנים הם .בים לחמורים מאודייננת ונחשעשויים להתרחש עקב קרינה מ
לחבר את יםעשוי נוני התיקון ינסו לתקן את השבר הםאשר מנגכ בהם, DSBמקרים של ריבוי : בתא יש שני מנגנונים שמטרתם לתקן נזק מסוג זה .הקצוות הלא נכונים זה לזה
1. NHEJ – nonhomologous end joining – שברכאשר התא מזהה . המנגנון המיידי ,אם נפלו –בעייתי זה הוא פתרון. צוות והם מחוברים מחדש באופן מיידינבחרים שני ק
את הם ילכו לאיבוד מאחר ומנגנון זה לא יזהה –נוקליאוטידים מקצוות השברים
בודדת DNAמנגנון זה הוא המנגנון היחידי שיתרחש בתא בשלב בו יש מולקולת . חסרונם
(. G1שלב ה)
2. Homologous recombination – מנגנון זה יתקיים במצבG2 , מוכפל הגנום כברבו .בכדי לדעת אילו נוקליאוטידים צריכים להיות התא משתמש במולקולה השלמהבמנגנון
. אשר יהיה מועדף כאשר אפשרי, מדובר במנגנון מאוד מדויק. קצוותב

8
את הכתוב על , 25לקרוא את פרקtransposable elements – שלא הספקנו נושא . ללמוד

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
10.11.09 –8' הרצאה מס –ביוכימיה
Transcription– תהליך השעתוק :
: מספר הערות
mRNA– מן ה 3-5%כמותו כRNA שבתא. rRNA– מרבית הRNA שבתא. מלבד לm, r, t , ישנן מולקולות מסוגsmall RNA ולהן חשיבות רבה .
: הבדלים עיקריים בין שכפול לשעתוק
.RNAבעוד בשעתוק תמיד ישועתק גדיל בודד של , בשכפול משתכפלים שני גדילים - .בעוד שכפול הגנום מבוצע על כולו, רק חלק מהגנום משועתק, זמן נתונה' בנק -גבוה פי כמה וכמה מאשר שיעור המוטציות RNAשיעור המוטציות במהלך יצירת ה -
היא DNAחר ומוטציה במא, מאחורי עובדה זו היגיון רב. DNAבמהלך שכפול ה
וגם אם לא תהיה , היא מולקולה זמנית RNAה –RNAמשמעותית הרבה יותר מאשר ב
לעומת . התקינה DNAתמיד יהיה ניתן להיפטר ממנה ולייצר חדשה מתבנית ה, תקינה . שגיאה בשכפול לא תמיד ניתן יהיה לתקן, זאת
: מושגים חשובים
הגן המשמש . ולשני לא תהיה משמעות, רק אחד הגדילים ישמש כתבנית, לגבי גן מסוים
. antisense strandאו ה noncoding strandכתבנית מכונה הזהה ברצפו לרצף ה, הגדיל שאינו משועתקRNA מכונה , שנקבל בסוף התהליךcoding
strand או הsense strand. .RNAר לנוקליאוטידים מסוג מסוגל להיקש antisenseרק גדיל ה -הדבר משתנה –ולהפך senseישמש כ antisenseהגדיל אשר שימש כ, לגבי גנים אחרים -
. לגבי גנים שונים- ORF– אזור זה יימצא בין קודון ההתחלה . האזור המקודד לחלבון(ATG ) לבין קודון
. הסיום .השעתוק מבקר את תחילת תהליך, ATGל' 5אזור בקרה המצוי –פרומוטור -- 5' UTR– נעצר ב, מקטע משועתק שאינו מתורגםAUG .בעל תפקיד בקרתי. - 3' UTR– בעל תפקיד . מתחיל לאחר קודון הסיום, מקטע משועתק שאינו מתורגם
. בקרתי- RBS – ribosome binding site– 5נקשר ל' UTR ,באזור הסמוך לAUG – באזור זה
.התרגוםמתחיל - uORF 5' – upstream open reading frame– לעתים ניתן יהיה למצואORF קצרה
מקטעים אלו גם הם יכולים להיות קשורים (. לעומת מאות בגן רגיל BP 25כ)מאוד לעתים אזור (. שאינם ברורים כיום)וייתכן ויש להם תפקידים נוספים , בבקרה או יציבות
ין לא ברור לגמרי מהו התפקיד השלם של עדי –ולעתים לא , זה יתורגם כחלבון נפרד . המקטעuORF הקודונים המקודדים לו לא , אומנם. קיים בתוך מסגרת הקריאה של הגן
כך שרק , נוקלאוטידים 2,1אלא לאחר , מתחילים מספר שלשות לאחר הקודון התחילי
הנוסף של AUGנוכל לזהות את ה( לא לפי שלשות)אם נתחיל קריאה לא רגילה של הגן . מקטע זה
. initiation, elongation, termination –שלבים 3גם תהליך השעתוק מורכב מ, כמו השכפול

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
Initiation– תחילת השעתוק :
כ באזור "בד, (פקטורי שעתוק) DNAי חלבונים הנקשרים ל"הבקרה על תחילת התהליך נעשית ע. קישור זה הוא המבקר את רמת הביטוי של הגן. הפרומוטור
סוג של פקטור שעתוק המדכא ביטוי של –רסור רפDNA ( ישנם פקטורים חיוביים (.ושליליים
λ 𝑐𝑠𝑜, λ 𝑟𝑒𝑝𝑟𝑒𝑠𝑠𝑜𝑟- רפרסורים שונים .
: תהליך השעתוק בחיידקים
: ניסו שנערך בנושא מנגנון פירוק של לקטוז בחיידקים
, במצבים בהם אין גלוקוז. כמקור פחמן ואנרגיה חיידקים משתמשים בגלוקוז, במצב נורמליחיידקים , (גלקטוזמורכב מגלוקוז ו, די סכריד)דוגמת לקטוז , אומנם יש מקורות סוכר אחרים
האנזים המפרק לקטוז לשתי . גלקטוזי פירוקו ליחידות הגלוקוז וה"מסוגלים להשתמש בו ע
𝛽יחידות אלו מכונה − 𝑔𝑎𝑙𝑎𝑐𝑡𝑜𝑠𝑖𝑑𝑎𝑠𝑒 .
. נמוכה מאוד האנזים של הביטוי רמת כי נראה, גבוהה גלוקוז רמת בנוכחות חיידקים נגדל אם, לקטוז במצע יימצא זאת ובמקום, מאוד נמוכה תהיה בסביבתם הגלוקוז רמת אם, זאת לעומת
𝛽 ה רמת − 𝑔𝑎𝑙𝑎𝑐𝑡𝑜𝑠𝑖𝑑𝑎𝑠𝑒 רמת מאוד קצר זמן תוך, גלוקוז ףנוסי שוב אם. אחת בבת עולה . בשנית מאוד נמוכה לרמה תרד האנזים
להגיב חייבת זו ותבנית, האנזים לביטוי המשמשת תבנית להיות חייבת כי הבינו, זה ניסוי מתוך להיווצר המסוגלת מתווכת מולקולה ישנה כי הבינו, בנוסף. הסביבה לתנאי רגישה מאוד בצורה
. במהירות ולהתפרק
: שונים מוטנטים נמצאו, הניסוי בהמשך
𝛽 של גבוהה רמה מקיים גלוקוז בנוכחות גם אשר מוטנט נמצא - − 𝑔𝑎𝑙𝑎𝑐𝑡𝑜𝑠𝑖𝑑𝑎𝑠𝑒 . . נמוכה הייתה האנזים רמת גלוקוז של נמוכה ברמה גם כאשר –הפוך מוטנט גם נמצא -
רגיש מאוד RNAמתוך כך הם הסיקו כי ישנו סוג של . רמת הבקרה על החלבון נפגמה, כלומר
(mRNA) ,כאשר מתרחשת בקרה על , מולקולה דינמית והמתפרקת ונוצרת לפי תנאי הסביבה . מולקולה זו
: lac-operonמודל ה הניסויהמודל הראשון שנוצר עקב
.פוליפפטידית אחתמקטע המקודד לשרשרת –ציסטרון
מולקולת סוג של . המקודד לחלבונים שונים בעלי תפקיד מטבולי בודד DNAאזור ב –אופרון RNA לדוגמא גנים )גנים השייכים לנתיב מסוים , במקרים רבים בפרוקריוטים. פוליציסטרונית
3מתעתיק זה ייווצרו . בודד mRNAתעתיק –מצויים יחד באופרון אחד ( הקשורים בעיבוד לקטוזאופרון הוא תופעה הקימת .אומנם הגנים מצויים תחת בקרת שעתוק משותפת, חלבונים שונים
.בפרוקריוטים בלבד
Lac-operon– ב, לפי הבנת החוקרים. אופרון המכיל גנים המטפלים בלקטוזlac operon 3ישנם : גנים
𝛽 הגן ליצירת האנזים .א − 𝑔𝑎𝑙𝑎𝑐𝑡𝑜𝑠𝑖𝑑𝑎𝑠𝑒 . . גן המאפשר הכנסת לקטוז לתא –פרמאז .ב

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
ולתא אין צורך , (גלקטוז)המיותר שנכנס לתא גן שתפקידו לסלק את החומר –אצטילאז .ג . בו
( רפרסור)השערת החוקרים הייתה כי ישנו חלבון מבקר . גנים אלו מצויים תחת אותה הבקרה 3
הגנים לא , וכי כאשר הרפרסור קשור לאזור זה, (operator)נים הג 3הנקשר לאזור המצוי לפני
.בקרת הרפרסור היא בקרה שלילית. RNAPיכולים לעבור שעתוק מאחר והחלבון חוסם את ה
ייכנסו בתחילה מולקולות הלקטוז, כאשר נכניס למערכת לקטוז ונדאג לרמת גלוקוז נמוכההמולקולות שייכנסו ייקשרו לרפרסור וייגרמו לו לשנות קונפורמציה כך . לתאים בצורה לא יעילה
ובה ( האופרון)בודדת mRNAצר מולקולת יי RNA polכך האנזים . operatorשהוא עוזב את ה .גניםה שלושת
RNA pol – RNA polymerase:
גם . DNAשעל גדיל ה' 3לקצה rNMPהוא מחבר . DNA polאנזים זה עובד בצורה דומה ל
.מלבד לעובדה שאין לו צורך בפריימר, מבחינת מנגנון DNA polהריאקציות דומות מאוד ל
. RNA – mRNA, rRNA, tRNAהיוצר בעצמו את כל סוגי ה, RNA polבחיידקים יש סוג בודד :שני ניסויים הוכיחו עניין זה
.RNAוהיא גרמה לכך שלא נוצר שום סוג של , RNA polבאנזים יצרו מוטציה בודדת .1
–אנטיביוטיקה זו מונעת את תהליך השעתוק – rifampicin מסוג אנטיביוטיקהשימוש ב .2
. RNAהוספתה למצע תגרום לכך שלא נקבל שום סוג של
:RNAPסוגי 3ביוקריוטים ישנם
1. RNAP 1 – אחראי על יצירת ה, מבצע את רוב פעילות האנזיםrRNA . 2. RNAP 2 – אחראי עלmRNA. 3. RNAP 3 – אחראי על יצירתsmall RNA ועלtRNA ( סוג שלsmall RNA .)
וגידלו תאים יוקריוטים בנוכחות רעל מעכב , כאשר ניסו לקיים ניסוי דומה לזה שביצעו בחיידקים
α)שעתוק − 𝑎𝑚𝑎𝑛𝑖𝑡𝑖𝑛) ,ראו כי הרעל לא גרם לעיכוב יצירת סוגי הRNA במידה שווה :
- RNAP 1 – לא עוכב גם בריכוזים גבוהים , עמיד לרעל( לא נפגמה יצירתrRNA .) - RNAP 2 – י הרעל גם בריכוזים נמוכים"עוכב ע . - RNAP 3 – י הרעל רק בריכוזים גבוהים"עוכב ע .
.השונים באנזימים שונים RNAמקור סוגי ה, כלומר
:נוספות מולקולות מעכבות שעתוק
1. cordycepin – קיים מימן ' 2מלבד לעובדה שעל פחמן , מולקולה דומה מאוד לנוקלאוטיד
השרשרת לא תוכל , לאחר שמולקולה כזו תתווסף לשרשרת, עקב כך. OHבמקום קבוצת .להמשיך להתארך
.Dאקטינומיצין .2
Bacterial RNAP:
:סוגים 5מ, שרשראות פוליפפטידיות 6אנזים זה מורכבת מ

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
יחידה𝛂 ( עותקים 2המופיעה בהשרשרת) , יחידה𝛃 , יחידה𝛃′- 3 יחידות אלו
על הוספת , RNAהן אחראיות על יצירת ה – core enzymeמשמשות כ . הנוקליאוטידים
. זיהוי הפרומוטר, בעלת תפקיד בנוגע לאזור בו מתחיל השעתוק -𝛉תת יחידה -ולכן , (ולא באופן שוטף)ישנם גנים אשר צריכים לעבור שעתוק רק במקרים מיוחדים
. שונה θמתקיים מערך במצבים כאלו . להם יש פרומוטרים מיוחדים
עבודת , ωללא RNA polאם נבנה . לא ברור מהו התפקיד של יחידה זו - 𝛚תת יחידה - . האנזים לא תיפגע בשום צורה
RNA pol מתקדם לאורך הDNA ,עד כי יחידת ה θבשלב זה החלבון נעצר . מזהה את הפרומוטר (. הרצף שזוהה כפרומוטר)אוטידים ינוקל 50י קישור לרצף של כ"ע
כך . יהיה לו סימן ומספר, כאשר נגדיר מיקום נוקליאוטיד מסוים על גבי הרצף: הערה . 1-והנוקליאוטיד לפניו יהיה , 1+הנוקליאוטיד הראשון המשועתק יהיה , לדוגמא
:תיאור התהליך
DNAמאות אנזימים כאלו סורקים את ה)וסורק אותה DNAמתקדם על גבי RNAPה .1
מזהה אותו כך שהאנזים θיחידת –מגיע לאזור הפרומוטר כאשר האנזים (. ביחידת זמן (. רצף זה הוא הפרומוטר)נוקליאוטידים 50י קישורו לרצף של כ"כולו נעצר ע
.DNAיוצר קומפלקס סגור סביב ה RNAPה .2
מפרידות בין שני ′βו α ,βהיחידות –יוצר קומפלקס פתוח RNAPה, Mg+2בנוכחות יוני .3
. קומפלקס זה הרבה יותר יציב מאשר הסגור. DNAגדילי ה4. RNAP באנזים זה ישנם שני אתרים. מתחיל לייבא נוקליאוטידים:
. purinשברוב המכריע של המקרים יהיה , אתר המוקדש לנוקליאוטיד הראשון .א
נוקליאוטיד . Gאו Aיהיה ( הנוקלאוטיד הראשון שישועתק) 1+כלומר הנוקליאוטיד . רזה הוא סוג של פריימ
. יוסיף את שאר הנוקליאוטידים RNAPלאחר קישור נוקליאוטיד זה ה .בשנוסף הוא בעל פרוססיביות מאוד ( נוקליאוטידים 10כ)המקטע הראשון , אומנם
כך . בעצם ממשיך להתקדם על המולקולה מבלי לייצר כלום RNAPה –נמוכה
10)יב עד כי ייווצר מקטע יחסית ארוך ויצ, נוספים RNAPימשיכו להגיע
. תתחיל להיווצר mRNAומולקולת ה( נוקליאוטידים יישארו מבלי שחלקם ייפלו
רק אז –עוזבת את הקומפלקס θהמקטע היציב נוצר לאחר שתת יחידה . הפרוססיביות שלו עולה
Elongation– הארכת השרשרת:
כאשר –במהלך השעתוק נוצר אזור המתואר כבועת שעתוק 20מדובר בכ. לאזור זה נוספים הנוקליאוטידים החדשים
כאשר בין , (אזור חד גדילי) DNAנוקליאוטידים פנויים של
חשיבות . RNAמצויים במצב דו גדילי עם מולקולת 8-9העניין היא בכך שסיום תהליך השעתוק תלוי ברמת החוזק
מספר . DNAנוקליאוטידים אלו לגדיל ה 8-9ור של של הקיש
מוסיף RNAPי כך שכל פעם שה"נוקליאוטידים אלו נשמר ע .הוא סוגר אחד מן הקצה השני, נוקליאוטיד

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
RNAP יכול להיעצר ממספר סיבות:
רצף ארוך של נוקליאוטידים מסוגGC . פתיחת )במקרה כזה האנזים עשוי לעצור ולחכות
ולאחר , (בגלל מספר קשרי המימן ATקשה הרבה יותר מאשר קשרי GCקשרים מסוג . לעתים האנזים אף יחזור אחורה. זמן מה להמשיך
בעיה בDNA ,mismatch או שבר בDNA לדוגמא.
DNAהחלק שהוא כבר סנתז ומצוי לפניו ייפול מן גדיל ה, יחזור אחורה RNAPבכל מקרה בו הובכך עצם )פעולה זו . וך מקטע זה וינסה להתקדם הלאההאנזים יחת, לכן. ויפריע לו להמשיך
. DNAמהווה סוג של סיגנל להבאת מנגנון תיקון ה( RNAPעצירת ה
:Consensus sequence
כאשר תת יחידה טטא . המשותף לגנים רבים, נוקליאוטידים 6קיים רצף של כ -35ו, -10באזורים
פקטור זה (. מטרה להתחיל את השעתוקב)ייעצר RNAP, מזהה רצף הדומה מאוד לקונצנזוסהדבר תלוי באחוז הזהות של רצף הקונצנזוס –מסוגל לגרום לרמות יעילות שונות של שעתוק הגן
. שבגן לרצף הכללי
:חשיבות כל נוקליאוטיד באזור הקונצנזוס ליעילות אתר הפרומוטר
זיהוי הרצף ובדקו עד כמה יעילות , שינו נוקליאוטיד בודד בכל פעם, בניסויירידה מתחת לקו האפס מראה , כפי שניתן לראות בגרף. ירדה או עלתהכנראה –אם קיימת גם עליה וגם ירידה באותו אזור . ולהפך, ירידה ביעילות
:נמצא כי, בנוסף. ויש תלות בנוקליאוטיד המחליף
.מסומנים בצהוב –הנוקליאוטידים השמורים ביותר -
.מסומנים בסגול –מן המקרים 60-65%הנוקליאוטידים הקיימים ב -
. מסומנים בכחול –מן המקרים 40-60%הנוקליאוטידים הקיימים ב -
יתכן . יש לציין כי רצף הקונצנזוס יהיה זה הקיים במרבית המקרים במציאות . אומנם לרוב אין צורך ברצף היעיל ביותר –ורצף זה לא יהיה היעיל ביותר
Termination – סיום התהליך:
:בפרוקריוטים יש שני מנגנוני טרמינציה
יופיעו שני אזורים : אתר זיהוי הטרמינציה נקבע לפי רצף –המנגנון השכיח יותר .1
משלימים זה את זה בתוך GCרצפי ה. Uולאחריהם אתר עשיר ב, GCעשירים ב
, השני GCמגיע לאתר RNAPכאשר ה. stem & loopכך שנוצר מבנה , RNAגדיל הכך שהרצף שכבר שועתק מתחיל ליצור את מבנה שניוני , את דרכו הוא מאט מאוד
. זהיוצר מבנה סטרי הגורם ללחץ , Uבנוסף לזיקה החלשה של רצף ה, זה שניונימבנה
:שתי הערות. DNAמגדיל ה mRNAהמאפשר ניתוק של מולקולות ה
כנראה ומלבד לשני אלמנטים אלו ישנם חלבונים נוספים התורמים .התהליךלסיום
RNAP הוא יכול להמשיך להישאר עליו –לא בהכרח עוזב את הגדיל . ולשעתק גן נוסף
, ρ שרשראות פוליפפטידיות מסוג 6המורכב מ -ρבמנגנון זה משתתף החלבון .2
, הנוצרת mRNAרצף ספציפי שעל מולקולת ההקסאמר בעל זיקה להומוהיוצרות

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
ρ שה לאחר. המקיים פעילות הדומה לאנזים הליקאז − 𝑝𝑟𝑜𝑡𝑒𝑖𝑛 ייקשר ל ρ −
𝑏𝑖𝑛𝑑𝑖𝑛𝑔 𝑠𝑖𝑡𝑒 ,תוך שימוש בATP גדיל )הוא מנתק אזורים דו גדילייםRNA וגדיל
DNA )כך שהmRNA ררתחמש.
:בקרת התהליך –רגולציית השעתוק
–גנים 3ישנם lac-operonידוע כי ב. י חיידקים"בחזרה לניסוי בנוגע לניצול גלוקוז וגלקטוז ע
𝛽 − 𝑔𝑎𝑙𝑎𝑐𝑡𝑜𝑠𝑖𝑑𝑎𝑠𝑒 ,פרמאז ואצטילאז .
הכולל )והפרומוטר , operatorה, האזור המקודד לרפרסור –במערכת הבקרה קיימים , בנוסף
.אם הרפרסור לא נמצא עליו RNAPאליו נקשר ה( operatorבתוכו את ה
בניסוי שתואר נבדקה רק הבקרה השלילית . היא גם חיובית וגם שלילית lac-operonהבקרה על ה . כיצד עוצרים את ביטוי האופרון אם אין צורך בחלבונים שהוא מקודד אליהם, כלומר –
:בקרה חיובית
האנזים אדנולאט ציקאז הופך , (רמות גלוקוז נמוכות בסביבה) האופרון כאשר יש צורך בחלבוני
מולקולה זו קרויה . בה הפוספט יוצר מבנה טבעתי AMPמולקולת – cAMPל ATPמולקולת
כך שחלבון , (רצפטור למולקולה המתעגלת) CRPהיא נקשרת ל וכאשר היא נוצרת, שליח שניונישינוי הקונפורמציה גורם לחלבון להיקשר לאזור הפרומוטר בצורה בה . זה משנה קונפורמציה
RNAP וישועתק כך שהגנים –יכול להיקשר לפרומוטור גם כן .
הCRP הוא סוג של גורם בקרה חיובי .
:lac operonטבלת סיכום של נושא ה
1 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
15.11.09 – 9הרצאה –ביוכימיה
קונספט זה נוצר . אותה ניתן להפעיל אותה מבחוץמערכת מושתקת –מערכת אינדיוסיבילית ניתן להחדיר את . המופעלת עליו( שליליתהחיובית וה)הבקרה ו, לאחר גילוי מערכת האופרון
-lacוהפרומוטור של הכאשר לגנים אלו מצורף הגן לרפרסור , הגנים שאנו רוצים לבטא לחיידקoperon .גן בנוכחות לקטוז ה –שונים לחלוטין יימצאו תחת בקרה שאנו מכירים חלבונים, כךכיום מוכרות מערכות . ובכך נוכל לשלוט בביטוי שלו –שאנו מעוניינים לבטא יתבטא לחלבון
(.lacoperonמלבד ל)אינדיוסיביליות רבות אחרות
הוא יכול להשתמש ,(חומצה אמינית חיונית) ך בטריפטופןכאשר לחיידק צור – trpאופרון עליו לייצר אותו –אם חומצה זו לא מצויה במצע . בחומצות המצויות במצע הגידול
המצויים כולם ישנם גנים רבים trpבמסלול הייצור של , כפי שניתן לראות בתרשים. בעצמו . על אופרון בודד
במצב הרגיל של התא רפרסור זה מצוי בתא באופן –לא פעיל ( צהוב)ניתן לראות רפרסור ומאפשר שעתוק של גנים ( לא על הפרומוטור)חופשי
כאשר יש כמות גדולה , אומנם. האחראיים על יצור טריפטופן, היא נקשרת לרפרסור ומאקטבת אותו –של חומצה זו בתא
בקרה )וייעכב את ייצור החומצה ייקשר לפרומוטור כך שהוא . (שלילית
ישנה בקרה ברמת –בנוסף לבקרת הרפרסור , ןפכאשר קיימת כמות גדולה של טריפטו. האינטיניואציה
(מצוי בתרשים מטה) של גן זה מקבל מבנה שניוני DNAהמבנה . UUUורצפי ( GCמסוג )שנוצר עקב אזורים משלימים
ר זה הוא מגיע לאזו RNA polכאשר –זה מזכיר את המבנה המאותת לטרמינציה של שעתוק . לגנים אלו mRNAלא יהיה –לא משעתק את הגן –מפסיק את השעתוק
Atinuated mRNA – הרצף הגורם למבנה השניוני .
לגדיל תהיה צורת –המבנה השניוני יהיה אחר –כאשר יש צורך בטריפטופןstem & loop בניגוד למספר מבנים כאלו בנוכחות רמות גבוהות של )בודדת
.והוא כן ישועתק ויתורגם –( טריפטופן
אם נייצר את מולקולת – antisense RNAי "שבתא נעשית ע RNAרמת בקרה נוספת על כמות הלמבנה זה . יווצר דו גדיליהן ייקשרו זו לזו כך ש – mRNAהמשלימה לרצף ה, האנטיסנס RNAה
.כן שהגן ששועתק לא יתורגם –לא יכולים להיקשר ריבוזומים
ישנם מקרים מעטים מאוד בהם החיידק משתמש בשיטה זו –ובלת בתוך התא שיטה זו לא מקאם נרצה לסמן . (in vitro) שיטה זו נהוגה הרבה במעבדות, לעומת זאת. (in vivo) בעצמו
נייצר רצף רדיואקטיבי מסוג –ואם כן אז איפה –כך שנדע אם היא קיימת , RNAמולקולת antisense ( איזוטופיבו הזרחן רצף) אם ה. ונדאג לביטויו בתאmRNA אכן קיים הם ייצמדו–
המסומן נמצא נראה כתם שחור בגלל ההתפרקות mRNAובמקום בו ה, ל'נריץ אותם בג . הרדיואקטיבית
:rRNAאופרון ה
. RNAהמורכבות מחלבונים ומיחידות –קטנה וגדולה –הריבוזום מורכב משתי תת יחידות :ורכב ממ( 70S)הריבוזום הפרוקריוטי
1. 30S – ממולקולת המורכבת היחידה הקטנהקומפלקסRNA קטנה(16S + )21 חלבונים. 2. 50S – מורכבת משתי מולקולות ההיחידה הגדולה קומפלקסRNA (23S, 5S) ,31ומ
.חלבונים

2 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
. מצויות על אותו אופרוןrRNA (16,23,5 )מולקולות ה 3נוצרת מולקולת – mRNAלא נוצרת דרך rRNAמולקולת ה
RNA (שהיא בעצם האופרון )האנזים ,לאחר יצירתה. ארוכהRNAse III ( נוקלאז החותך חומצות גרעין מסוגRNA) ,חותך
של rRNAמבנים שניוניים אלו הם יחידות ה .step & loopרצפים המצויים במבני ה אתמתחיל בחיתוך המולקולה תוך כדי RNAse IIIה (.tRNAחלק ממולקולת ה)+ הריבוזומים
אם ניצור מוטציה באנזים . ה השלםבבתא כמעט ולא ניתן למצוא מולקולה זו במצכך ש, יצירתהמסיבה זו אנזים זה מהווה בקרה על יצירת . נראה הצטברות של מולקולה זו בתא, RNAse IIIה . שבתא rRNAה
לחיידק יש צורך בכמותrRNA ל כל הזמן לתרגם חלבוניםבכדי שיוכ –גבוהה מאוד .
מדובר רק בחלק . tRNAהראשונית מצויות מולקולות RNAבמולקולת ה, כפי שניתן לראותכך . קא מצויות בתוך אופרוניםוולאו דו ,מפוזרות בגנום החיידקכאשר השאר , ממולקולות אלו
:התהליך .ריםמספר אנזימים אחי "אלא ע, RNAse 3י ה"עיבוד מולקולות אלו לא נעשה ע, או כך
9במרחק של כהארוכה RNAאת מולקולת הבשלב הראשון סוג של אנדונוקלאז יחתוך .1 . stem & loopנוקליאוטידים מן ה
2. RNAse D ,עד , 7כך –אחד אחד , מן הקצהנוקליאוטידים חותך ,סוג של אקסונוקלאז . ני נוקליאוטידיםשכי נשארים רק
tRNAחותך את החלק השני של ה RNAse Pהאנזים .3 .כך שהיא חתוכה משני הצדדים ('5קצה )
4. RNAse D ממשיך ואוכל את שני הנוקליאוטידים 9שבמקור היו בו )הרצף שנשאר בקצה גדיל זה . שנשארו
רצף זה הוא (. '3אזור ה) CCA הוא( נוקליאוטידים עודפיםאכל חלק RNAseאם מסיבה מסוימת ה –רצף חיוני
יחזור ויסנתז _____ אנזים ה –מנוקליאוטידים אלו . אותם מחדש מאחר והם עקרוניים מאוד
עוברת מודיפיקציות שונות שבודדה tRNAמולקולת ה .5 :RNAאשר הן ספציפיות ל
אורציל הופכים לפסואידו נוקליאוטידים מסוג .א .יורידין
._____________________ –ריבותימידין .ב .________________________ –ין דיהידרויוריד .ג
כך , (אנזים)וממרכיב חלבוני RNAמורכב ממולקולת Pהאנזים ריבונוקלאז – RNAse Pבנוגע ליש צורך בכל ( in vivo)מסתבר כי בתוך התא . RNAמכיל בעצמו RNAשהאנזים החותך
, in vitroאומנם אם נחסיר את החלבון במצב , (RNAגם החלבון וגם ה)הקומפלקס האנזימטי . בלבד RNAי מולקולת ה"הפעילות האנזימטית תתבצע ע
מולקולותRNA בעלות פעילות אנזימטית מכונות ריבוזים.
היא מולקולה בעלת זמן mRNAה – בפרוקריוטים mRNAדרך נוספת לבקרת – RNAית ידגרדצ –משועתקות ומפורקות בתא כל הזמן mRNAלות מולקו. דקות 2-3כ, חיים קצר מאודמחצית
המגיבות במהירות רבה לתנאי , עובדה זו היא ההופכת מולקולות אלו לאדפטיביות ביותר . הסביבה המשתנים
:שעתוק בתאים יוקריוטים
:הגנום היוקריוטי

3 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
לעומתם . נוקליאוטידים 106המורכב מכרומוזום אחד ובו , לחיידקים יש גנום יחסית קטן. מיליארד נוקליאוטידים 10בין מיליארד ל –בבני אדם . ליון נוקליאוטידיםילשמרים ישנם כמ
נראה כי בבני אדם –אם נסרוק את מספר הגנים הצפוי . גנום גדול יותר מאשר לאדם –לאפונה , למרות שלא נראה כי מדובר במספר גדול מאוד. גנים 30,000יש כ
גנים 30,000יכולה לייצר מ, י תהליכים שונים"ע, יתהמערכת התא .חלבונים שונים 150,000אלו כ
הגנומים של ניתן לראות כי ישנה וריאביליות מאוד גדולה בין קא נוגע למספר וכאשר מקורה לאו דו –האורגניזמים השונים
ההסבר לכך מגיע מן (. כפי ניתן לראות בתרשים ובטבלה)הגנים שלא ,DNAמרבית ה( ביורקיוטים בעיקר) – noncoding DNAת גדולה של העובדה שיש כמו :זה DNAסוגים של 3שנם י. עובר שעתוק כלל
1. satellite DNA – רצפים קצרים שחוזרים על עצמם מאוד ואלפי פעמים אחד אחרי השניכתוצאה –י הכפלות לאורך האבולוציה "רצפים אלו נוצרו ע(. לא בצורה מפוזרת, ביחד)
, (או את כולו) DNAאם נקרין מקטע . ויראלי שחדר לגנום DNAמטעות או מהכנסה של הם כולם , ואם נריץ אותם בגרדיאנט צפיפות, ( BP 300כ)נקבל מקטעים שאוכם זהה
נשתמש , DNAלשאר ה sateliteבכדי להפריד בין מקטעי ה. יימצאו באותה הפרקציהאם ניצור את הגרדיאנט בתנאי – Tו Aדים מסוג בעובדה כי הם עשירים בנוקליאוטי
והם יימצאו , יפרדו לשני גדילים כך שצפיפותם קטנה sateliteמקטעי ה, חום מסוימים . במקום גבוה יותר בגרדיאנט
לדוגמא )ההנחה היא כי יש להם תפקיד מבני –לא ברור מהו התפקיד של אזורים אלו וחשיבותו נובעת מכך שסיבי , ים תעתיקיםמן הצנטרומר לא נוצר(. במבנה הצנטרומר
satelliteגם הטלומרים הם סוג של . הכישור נקשרים אליו במהלך החלוקה המיטוטיתDNA .
2. Alu elements – אחד אחר , נוקליאוטידים אשר לא מופיעים ביחד 300מקטעים של כ Alu1שמם נובע מכך שאנזים הרסטריקציה . אלא משובצים במקומות רבים בגנום, השני
ולא ברור , מן הגנום 10%אזורים אלו מהווים כ. חותך רצף המצוי במרבית אזורים אלועוד תופעה . מאוד מאוד דומיםרצפים אלו תמיד יופיעו בין שני רצפים . מהו תפקידם
מכרומוזום )המשויכת למקטעים אלו היא כי הם מדלגים ממקום למקום בגנום אלמנטים . human transposoneההנחה היא כי מקטעים אלו הם חלק מ(. לכרומוזום
.אלו קיימים יותר בפרימטים שאינם בני אדםאם (. לא קיימים כלל בתאים פרוקריוטים)אופייניים לתאים יוקריוטים –אינטרונים .3
נראה כי –שקודד לו DNAששועתק ונשווה אותו לרצף ה mRNAנבדוק את רצף ה, premRNAמקטעים אלו מוצאים ממולקולת ה, splicingבתהליכי . חסרים מקטעים
3לראות גן ובו במצגת ניתן . נקשרים זה לזה מחדש( אקסונים)והקטעים שנותרו אינטרונים יכולים להיות קצרים או . אינטרונים 2אקסונים ו
כך או כך הם יוצאים מן המולקולה כך שהם לא –גדולים מאוד . noncoding RNAכובעצם ניתן להגדיר אותם , יתורגמו לחלבון
– alternative splicingחשיבות אינטרונים אלו מתקיימת בעת . premRNAלבונים שונים מאותו מקטע תהליך המאפשר יצירת ח
mRNAוהאדום הוא ה premRNAכחול הוא ה –בתרשים . שיתורגם
גם , לדוגמא. ישנם גנים המופיעים במספר רב של עותקים –סיבה נוספת לגודל הגנוםלכן –בתאים פרוקריוטים וגם בתאים יוקריוטים יש צורך בכמות אדירה של ריבוזומים
כ "רצפים אלו בד. יש צורך בעותקים רבים של הגן המקודד לחלבונים הבונים אברון זהההתרחבות הנראית בתמונה סביב הגדיל . tandem repeatsמופע המכונה – יופיעו ביחד
. הנוצרת RNAהיא מולקולת ההריבוזומלי ישנם חלבונים נוספים שיש צורך בכמות RNAבנוסף לגנים המקודדים ל
. היסטונים לדוגמא –דולה שלהם ג
:שעתוק יוקריוטי

4 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
כך שלעתים התרגום ) בהם השעתוק והתרגום מתרחשים באותו אזור –לעומת הפרוקריוטים בעוד , מתרחש בגרעין בתא –כל תהליך השעתוק ביוקריוטים , (מתחיל עוד לפני שנגמר השעתוק
. התרגום מתרחש בציטופלזמה
שם נוצרות יחידות –תפקיד הגרעינון נוגע ליצירת הריבוזומים . בתוך הגרעין קיים הגרעינוןנוצרים בתוך ( הריבוזומליםיחד עם החלבונים )השלמים הקומפלקסים גם , כמו כן. rRNAה
.הגרעינון
( דחוס, כהה)בה ניתן לזהות אזורי הטרכרומטין –גרעין התא בתמונה ניתן לראות אתכל החומר הגנטי (. פחות דחוס כך שמאפשר מעבר קרני אור, בהיר)אאוכרומטין ואזורי
עניין זה מתבטא ברמת . אומנם קיימות רמות שונות של דחיסות –מאוד מאוד דחוס אזורים לא פעילים , לעומתם. פתוחים המשועתקים צריכים להיות יותראזורים –השעתוק
.מבחינת שעתוק יופיעו כהטרוכרומטין
עובר רמת דחיסה ראשונית סביב DNAה. DNAמרק ומטין לא בנויהכראשר הם מייצרים מבנה דחוס עוד ( מבנה הכולל קומפלקס היסטונים)נוקלאוזומים
בשלב חלוקת התא המבנה נהיה עוד יותר דחוס כך . כך עד לקבלת הכרומטין –יותר נוספים בוניםוחל, היסטונים, DNAקומפלקס הכרומטין כולל . שמתקבל כרומוזום
. nonhistone proteinsהמכונים (התורמים למבנה)
DNAהעדות הראשונה לקיום הנוקלאוזומים הייתה כי כאשר הפיקו מולקולת לדוגמא מיקרוקוקל אנדונוקלאז החותך כל )וחשפו אותה לנוקלאזות לא ספציפיות
DNA המייצגים נתקבלו בנדים , פורזהלאחר הרצה באלקטרו(. שנמצא בדרכודים לא אוטייניתן לראות כי ההבדל של הנוקל. אוטידיםינוקל 200הבדלים של כ
ככל שגודל המולקולה גדול יותר כך –לינארי אלא אקפוננציאלית ___________________________________________________.
, אזותמוגנים מפני הנוקל 145מתוכם – נוליאוטידים 200ישנם רצפים של כ, כלומר 145הרצפי . (שנחתכוהאזורים )מקשרים שאינם מוגנים נוקליאוטידים 55וביניהם
. מוגנים בעצם מאחר והחלבונים מונעים לא מאפשרים גישה של הנוקלאזות לרצף .החלבונים הוגדר כנוקלאוזום+ המבנה כולו של רצף כזה
כל ניתן לדעת כי –אנדונוקלאז לא ספציפי מאחר והשתמשנו ב . הופרדו הנוקליאוטידים הלא מוגנים
י שימוש "שנים לאחר מכן ניתן היה לראות את מבנה השרשרת הנוקלאוזומית ע .במיקרוסקופ אלקטרונים
לינקרDNA – הDNA הלא מלופף .
. חלבונים שכל אחד מהם מופיע פעמיים 4ליבת הנוקלאוזום היא קומפלקס אוקטאמרי המורכב מ :ההיסטוניםחלבונים אלו הם
- H2A - H2B - H3 - H4
את הקישור של ליבת הנוקלאוזום ותפקידו ליצור, הוא לא חלק מליבת הנוקלאוזום H1ההיסטון . בתמונה ניתן לראות היסטון זה בצהוב. סביב הליבה DNAי ליפוף ה"ע, DNAעם ה
ן וליזיןהם חלבונים מאוד בסיסיים ומכילים כמות גדולה מאוד של ארגני( H1כולל )ההיסטונים בטבלה ניתן . שיירי הליזין במיוחד בעלי תפקיד חשוב מאוד במבנה זה. (חומצות אמינו בסיסיות)
. 1/5לראות כי אחוז הליזין והארגנין שבהיסטונים נע בין רבע ל

5 . 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הבסיסיות שלההיסטונים היא
המאפשרת קישור יעיל גם . זה pHטעון שלילית ב DNAבעוד ה, pH 7מאחר וטעונים חיובית ב, DNAשלהם ל
.המצויים בנוקלאוזום הם בעלי אופי בסיסי nonhistoneחלבונים מסוג

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
17.11.09 – 10הרצאה –ביוכימיה
:הערות להרצאה קודמת
באופן חלקי בגלל מקטעי , הגנום היוקריוטי גדול בהרבה מהגנום הפרוקריוטיDNA שלא
בעלי . נוקליאוטידים 300אורכם כ – Alu elementsמקטעי , לדוגמא. מקודדים לגנים (.טרנספוזום יורקיוטיסוג של )לים לדלג ממקום למקום בגנום גמסו. תפקיד לא ברור
סוג של מקטע אינטרונים הם גםnoncdoing .לעתים אינטרון של גן אחד יקודד , אומנם
. noncodingולכן לא מדטבר במקטע שהוא לגמרי , לגן אחר
זומים מתפרקים הכרומטין מתארגן מחדש כך שהנוקלאובעזרת חלבונים שונים , במהלך השכפול
נוקליאוטידים ___ כ. יכול לעבור ולשכפל את הגדיל DNA polהבצורה זו (. לפני מזלג השכפול) . הנוקלאוזומים מתארגנים מחדש ונוצר מבנה הכרומטין, לאחר שהמזלג עבר
:(tRNAו mRNA ,rRNAמלבד ל) שבתא RNAמגוון מולקולות
1. Small nuclear RNA – snRNA – ספלייסינג )בעלות מספר תפקידים , ות גרעיניותמולקול . (לדוגמא
2. Small nucleolar RNA (snoRNA )– למולקולות אלו . מולקולות המצויות בגרעינון
, המקודד לריבוזומים מצוי שם DNAומאחר ורוב ה, תפקיד חשוב מאוד בארגון הריבוזום .גם מולקולות אלו יהיו באזור זה
3. scaRNA – בעיבוד מעורבותsnoRNA .
4. miRNA (mir???) – שאלה מסמנים את מספר מולקולת הסימני הmiRNA . בעלותתהליך התרגום של מולקולת מבקרות את –חשיבות רבה בבקרה על ביטוי גנים
. mRNAישנם אזורי קישור ל בו, שאינו מתורגם, UTR' 3לרוב הן ייקשרו לקצה . mRNAה
.רום לביטוי גן או לדיכוי שלוהן עשויות לג mRNAכאשר מולקולות אלו ייקשרו לעשויות –מולקולות אלו מהוות פקטור בהתפתחות של גידול סרטני לאחרונה גילו כי
מקרה כזה הוא דוגמא לכך שלעתים גם אם לא . לגרום לביטוי גן שלא אמור להיתרגם .עשויה לגרום לפגם בחלבון עת עליובעיה בבקרה המבוצ, התרחשה שום מוטציה בגן
5. Small interference RNA (siRNA )– תמולקולות קצרות המתלבשות על מולקולו
כאשר רוצים (. פירוק)התאי לדגרדציה RNAתפקידן לדחוף את ה. שבתא mRNAהניתן , (לדוגמא בכדי לבדוק כיצד התא מתפקד בלי החלבון)במעבדה להשתי ביטוי של גן
. להשתמש במולקולות אלו
:וקריוטיםבי RNA polאנזימי ה
- RNAP 1 – מרבית מולקולות סינתזתי על אאחר rRNA (28S,18S,5.8S.)
- RNAP 2 – אחראי על סינתזתmRNA , עלsnoRNA ,miRNA וחלק מן הsnRNA.
- RNAP 3 – י על האאחרtRNA, מולקולת rRNA הקטנה (5S) ,חלק מן הsnRNA ומן
. small RNAה
RNAP 1:
תופעה המכונה ) קיימים מספר עותקים שלו בגנום. מדובר באנזים הפשוט ביותר מבין השלושה
tandem repeats) .ומוטורים שונים לכל לא קיימים פרrRNA ( רצף הפרומוטור לקטעים אלו
RNA pol 1. ולכן אין צורך בפקטורי שעתוק רבים, (ולא בתוך הגנום –משתנה רק בין מין למין . שי יזהה את רצף הפרומוטור האנושי בלבדהאנו

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
חלקן, סיביותפרוס מקנות לאנזיםחלקן –תת יחידות 13מכיל כ של אנזים זה holo enzymeה . 'בזיהוי הפרומוטר וכד בעלות תפקיד
:rRNAתהליך שעתוק ה
מקטע זה הוא הדומה ביותר לאופרון הקיים בתאים אנימלים המקטע אשר רואים בתרשים חוזר(. מההוא לא אופרון אך דו)
. על עצמו פעמים רבות בגנום
מקודדים למקטעי ישנם אזורים ה ניתן לראות כי במקטע
ישועתק בתחילה הפרקורסור. השונים שבריבוזום rRNAה
, (שונים rRNAכ יהפוך לשלושה מקטעי "שאח 45Sמקטע )מודיפיקציות . מודיפיקציות שונות בגרעיןולאחר מכן יעבור
:אלו כוללות
ווספת במתילציה זו מת. שת רק באזור הפרקורסורמתילציה מאוד ספציפית המתרח .1
תהליך זה מכונה . שעל הריבוז 2' פחמן מסשעל OHקבוצת הקבוצת מתיל ל
2Omethylation . עוברים מתילציה בצורה זו באתרים כולם, נוקלאוטידים 200על פני מולקולה זו כ
. ספציפיים מאוד
.באתרים ספציפים שעל המולקולה, פסואידו יורידיןיורידין ל תהפיכ – איזומרציה .2
היחידות –המתילציה והפסואידויורידינציה מתרחש חיתוך באתרים ספציפיים לאחר .3
18S ,5.8S ,28וS 5יחד עם ה, לאחר מכן. מופרדות זו מזוS י ה"המיוצרת עRNA pol 3
, 40S –תת יחידה קטנה )נוצרות שתי תת היחידות המרכיבות את קומפלקס הריבוזום
(. 60S –תת יחידה גדולה
או את היורידין שצריך לעבור , התא יודע לזהות את האתרים אליהם צריך להיקשר המתיל
קומפלקס . snoRNP חלבוןה ובוקומפלקס המצויות ב, snoRNAבעזרת מולקולות ה, מודיפיקציה
כל אחד יודע ,snoRNP לישנם סוגים שונים ש)זה נקשר לאתרים שצריכים לעבור מודיפיקציה כך שהתא יודע לבצע את המודיפיקציה באתרים אלו , (לפסואידויורידינציהלסמן למתילציה או
. המסמנות אתרי חיתוך במולקולת הפרקורסור snoRNAישנן מולקולות . בלבד
RNA pol 3:
. ועוד snoRNA ,tRNAחלק מה, 5S rRNA –הקטנות RNAמייצר את מולקולות התת 14המורכב מ( אחת טידיתלא שרשרת פוליפפ)גם אנזים זה הוא קומפלקס
. לא מכיל מגוון גדול של פקטורי שעתוקו, יחידות
כל אחד מכיל , tRNAישנם כשני פקטורי שעתוק מרכזיים הקשורים בשעתוק : מספר שרשראות פוליפפטידיות
- TFIIIC – לזהות בכדי ,בשני אתרים שונים, הראשון שנקשר למולקולה
לאתר תחילת TFIIIBלאחר מכן נקשר .את האתר הדורש שעתוק
. RNA pol 3השעתוק ומגייס את האנזים
- TFIIIB – נקשר למולקולה לאחר קישורTFIIIC ,לאתר תחילת השעתוק ,
. tRNAהמשעתק את ה RNA pol 3ובכך מגייס את האנזים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
מזהה אתרים המצויים בתוך האזור ה ,TFIIIA, נדרש פקטור שעתוק נוסף 5S rRNAלצורך שעתוק
.לפי המנגנון שתואר קודם TFIIICו TFIIIBלאחריו יפעלו פקטורי השעתוק .המשועתק
שני תהליכים אלו לא אופייניים למולקולותmRNA – כ פקטורי השעתוק לא "בהן בד (. המקודד)ייקשרו לאזור הרצף המשועתק
כאשר הrRNA 5יחידת ה, משועתק בעודףS נקשרת לפקטור השעתוקTFIIIA , כך שנמנע
. לא ישועתק עד שכמותו בתא תקטן שוב rRNAהו, השלב הראשון
טורי שעתוק עיקריים בפק 3ישנם –לסיכוםholoenzyme של הRNA pol 3 , המקיימים .תהליך יחסית פשוט
RNA pol 2:
כל חלבוני התא כך שניתן להגיד כי , שבתא mRNAאחראי על ייצור כל הה, מורכב יותר אנזים
וחלק מן snoRNA ,miRNAבנוסף הוא אחראי על סנתוז . RNA pol 2שייצר mRNAבמקורם
.snRNAה
לא כל )או בתאים מסוימים , (לא באופן שוטף)ישנם חלבונים אשר אותם הגוף צריך בזמן מסוים
יהיו משועתקות לפי mRNAמולקולות ה, RNAבניגוד לשאר מולקולות ה, לכן(. הרקמותנעשית לכן . ויוצר בקרה על השעתוק הגן המופיע אחריו, תנאי שעתוק הגןהמגדיר את , הפרומוטר
:שני מנגנוני שעתוקהפרדה בין
.המוחל על כל גן בגנום, (bazal transcription machinery)מנגנון השעתוק הבסיסי .1
הגן ואיפה ימת RNA pol 2ל היודעים לסמן ,נוספים שעתוק הכולל פקטורי שעתוקמנגנון .2 . (לא ואיפה ומתי)צריך להיות משועתק
הקיימים במנגנון ( שלא משתנים)י שעתוק בסיסיים ורקיימים אותם פקט (ללא יוצאי דופן) לכל גן .ישנם פקטורי שעתוק שוניםלעתים לגנים שונים , מעבר לכך. (הבזלי) השעתוק הבסיסי
Housekeeping genes – ר כך שלא דורשים פרומוט, התאיםבכל , ים המשועתקים כל הזמןגננמצא כי בפרומוטר של גנים . מאחר ואין באמת הוראות למתי ואיפה הגן צריך להתבטא, מיוחד
מנגנון שעתוק . RNA pol 2י "והוא אלמנט המוכר ע, GC boxהמכונה , GCעשיר באלו קיים רצף (. -35ואזור -10אזור )דומה מאוד לזה המוכר בחיידקים זה
גנים המבוטאים באופן סלקטיבי ב, housekeeping genesלעומת ה
רצף עשיר ) TATA boxנמצא את ה( זמנים מסוימים וכד, תאים מסוימים)
לאתר תחילת השעתוק upstreamבסיסים 25-30במרחק של כ (Aוב Tב
הוא הסיגנל הראשוני המגדיר את האזור TATA boxה. )+1אתר ) . כפרומוטור
א אשר הו, TBP (TATA binding protein)חלבון נקשר ה TATA boxאל ה
פקטור השעתוק הוא TBPה. TFIIDחלק מקומפלקס גדול יותר המכונה
מכיל פקטורי שעתוק נוספים TFIIDהקומפלקס . הראשון הנכנס למערכת
(other TATA binding proteins) ,חלבוני , וחלבונים נוספיםTAF –
TATA associated proteins ,לרצפי ה הנקשריםTATA . לאחר קישור
. הפרומוטר מזוהה TAFחלבוני ה

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הקישור של הTAF לDNA אשר הוא חשוב מאוד בכך שעוזר , יוצר כיפוף של הגדיל . לפקטורי השעתוק הבאים לזהות את האזור ולהיקשר אליו
ובכך מייצב את הקומפלקס , נקשר לאתר גם הוא TFIIA פקטור השעתוק ,הפרומוטר זיהוילאחר
(DNAמבלי להיקשר ל) RNApol 2נקשר ל TFIIF פקטור השעתוק .TFIIBלאחריו נקשר . הקיים
חשובה מאוד מאחר ומונעת TFIIFהפעילותו של . TFIIBי זיהוי ה"ומכוון אותו לאזור הפרומוטור ע
.להתחבר לאתרים אחרים RNA pol 2מהאנזים
GTF =general transcription factor =הכולל את כל פקטורי , מנגנון השעתוק הבסיסי
.?TFIIהשעתוק
:הוא כפול TFIIHתפקיד ה. TFIIH הפקטור הוא להפעיל את TFIIEתפקידו שלפקטור השעתוק
noncodingלקרוא את ה RNA pol 2שר לפאומ DNAפותח את ה –משמש כהליקאז .1
strand .
בלעדיו –זרחון זה קריטי . RNA pol 2לי של הטרמינ Cב הרחון הזנאחראי על ז .2
בגלל . DNAויישאר תקוע על מולקולת ה, התחיל את השעתוקהקומפלקס לא יוכל ל
.הוא קינאז TFIIHפעילות זו הפקטור
ניתקים מן המולקולה ?TFIIכל חלבוני ה, מתחיל את תהליך השעתוק RNA pol 2ברגע בו ה
נופל ממנו יחד עם לא RNA pol 2הזרחן שנקשר ל. חדש TATAומתחילים את התהליך באתר
מולקולות נוספות החשובות RNA pol 2מאחר ומאוחר יותר יקשור ל, פקטורי השעתוק .לספלייסינג ולתהליכים נוספים, לפוליאדנילציה
, TATA box ,GC box – המגדירים את הפרומוטר ניתן לראות את האזורים השונים בתרשים
CAAT box ,octamer motif .100ה כ בין"בד, יעו באתר שלפני השעתוקאלו תמיד יופ מוטיבים
. לאתר תחילת השעתוק upstreamנוקלאוטידים המצויים ה
מרבית הגנים לא יכילוTATA box ,אלא רצפים אחרים .
גם במקרים בהם אין את רצף הTATA, אלא כל אזור אחר
(GC כברירת מחדל) הTBP ייקשר ויתחיל את תהליך
אומנם גם , מקד את חלבון זהל TATAהתפקיד . השעתוק
. פשוט באופן פחות ספציפי, יפעל TBPה GCבאזור
:RNA pol 2מבנה ההערות בנוגע ל
מכיל אזור –( צורת האגרוף) DNA polמקיים מבנה דומה מאוד ל -אזור יציאה בנוסף בעל פפטיד טבעתי הגורם לפרוס , כניסת רצף
. סיביות מאוד גבוהה
- Rudder – מקטע המפריד בין מולקולת הDNA למולקולת הRNA דו )דוש לפעילותו יוני מגנזיום ( אדום)אנזים זה . הנוצרת (.ערכיים
- α חומר המעכב כמעט רק את –אמניטיןRNA pol 2 . חומר זהמונע את כניסת הנוקליאוטידים לתוך ,אנזיםנקשר ל
. ובכך מונע את הארכת השרשרת, holoenzymeה

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:ספציפית לשעתוקבקרה
:לאחד משני אזורים ייקשרו( לא בזליים)ספציפיים פקטורי שעתוק
תחילת לאתר upstreamבסיסים 1000ועד לכ 100מ –אזור הפרומוטור המרוחק - .השעתוק
אתרים , enhansoresעשויים להיקשר לפקטורים – אזור הפרומוטור המרוחק עוד יותר -ובכל זאת ,ליאוטידים מאתר תחילת השעתוקנוק מאות אלפי/אלפיהמצויים במרחק
כך שלמרות , אלא מקופלת, היא לא מולקולה ישרה DNAה. את תהליך השעתוק מבקריםבצורה המקופלת הם יהיו צמודים , שבין שני מקטעים ישנם מאות אלפי נוקליאוטידים
– '5או ' 3מיקום ביחס לולכן אין חשיבות ל, הקיפול יכול להתקיים בכל כיוון. זה לזהחייבים להיות במגע ישיר אומנם , בכל אחד משני הכיוונים להימצאיכולים האנהנסורים
.גן עצמו לאחר הקיפולעם ה
:כ יש שני אזורים"בדהספציפיים השעתוק ילפקטור
1. DNA binding domain – כ ייקשר לרצף דרך ה"חלבון בדאזור זה בminor והmajor
groove .
2. Activation domain – אינטראקציה בין אזור זה לבין חלבוני הbazal transcription
machinery תשפיע על גיוס החלבונים לאתר .
בשיטות של( בין האזורים)ניתן להפריד ביניהם –שני אזורים אלו מכיל כל פקטור שעתוק מאחר ו ________________________________________.אנזימי רסטריקציה)הנדסה גנטית
.(הקלטה)______________________
(:DNAבהתאם למנגנון הקישור שלהם ל)פקטורי השעתוק נחלקים למספר משפחות
1. Zn fingers – נה החלבון מב. צורת אצבע ובמרכזה אטום אבץ בעליחלבוניםי שיירי ציסטאין והיסטידין המצויים במרחקים זהים מן האטום "מתאפשר ע
יכולות להשתלב אשר בליטות בשילוב עם האבץ נוצרות .המקשר ביניהם
. ברצפים ספציפיים DNAכך שהחלבון נקשר למבנה ה, DNAשל ה groovesב
שתי שרשראות בכל דימר –חלבונים המופיעים כדימרים – לאוצין זיפר .2חומצות 6כ כל "בד) שיירי לאוצין במרחקים שווים אלו מאלו בעלותפוליפפטידיות
כל , groovesכך שבאמצעות מבנה ה, 'רץ' רי הלאוצין יוצרים מבנה ריץייש. (אמינומבנה חלבוני זה יכול להיות הטרודימרי . שלוחת פוליפפטידית יוצרת קישור לגרוב
צירופים שונים של (. שתי שרשראות זהות)או הומודימרי ( שתי שרשראות שונות) .השרשראות השונות יזהו גנים שונים
3. Helix loop – יחידות חלבונים הבנויים מα ים לינארים טעהליקס ומק(loop )לסירוגין .
הליקס הן αיחידות ה. מבנים כאלו עשויים להופיע בצורת דימרים ועשויים שלא
. DNAהיוצרות את הקישור ל
4. Helix-turn-helix – מקיימים מבנה מאוד דומה לחלבוני הhelix loop , אומנם .ות מקטע היוצר פנייה בשרשרת החלבוניתבמקום מקטע לינארי מקשר בעל
יוכלו DNA polוה RNA polוגם אנזימי ה, בכדי שכל פקטורי השעתוק עליהם דיברנו
–מאורגן במבנה דחוס DNAידוע כי ה. לגדילצריכה להיות גישה פיזית , DNAלהיקשר לרצף המבנה מדובר ב, נםמאו. לא מאפשר קישור קל קיפוליםו ההבנוי מנוקלאוזומים מבנה הכרומטין
.או פשוט לרווח את המרחק ביניהם, נוקלאוזומיםלפרק ניתן להזיז או –דינמי
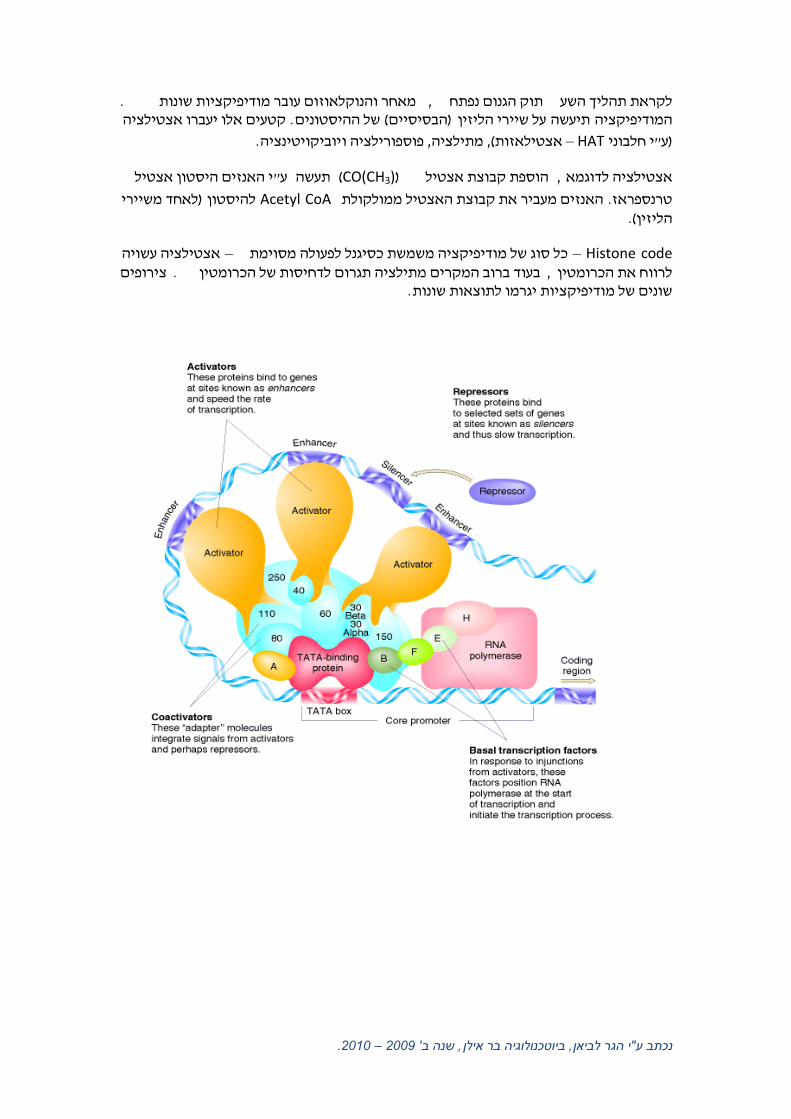
. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
. מאחר והנוקלאוזום עובר מודיפיקציות שונות, תוק הגנום נפתחלקראת תהליך השע קטעים אלו יעברו אצטילציה. של ההיסטונים( הבסיסיים)על שיירי הליזין תיעשההמודיפיקציה
. יוביקויטינציהפוספורילציה ו, מתילציה, (אצטילאזות – HATי חלבוני "ע)
י האנזים היסטון אצטיל "ע תעשה (CO(CH3))הוספת קבוצת אצטיל , אצטילציה לדוגמא
לאחד משיירי )להיסטון Acetyl CoAהאנזים מעביר את קבוצת האצטיל ממולקולת . טרנספראז (.הליזין
Histone code – אצטילציה עשויה –כל סוג של מודיפיקציה משמשת כסיגנל לפעולה מסוימתצירופים .בעוד ברוב המקרים מתילציה תגרום לדחיסות של הכרומטין, לרווח את הכרומטין
.ודיפיקציות יגרמו לתוצאות שונותשונים של מ

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
22.11 – 11הרצאה –ביוכימיה
:הערות לשיעור קודם
י "כל מולקולה המשועתקת עRNA pol 3 י מנגנון השעתוק הבסיסי "משועתקת ע(bazal
transcription machinery) , הדורש אזורGC box . מולקולות מסוימות יכילו בשעתוק
. TATA boxהדורשים אזור , נוספים( פקטורי שעתוק)אנזימים
ההבדלה ביןhousekeeping gene י "ע מקומות ספציפיים נעשית/ המשועתק בזמניםלגן
אליהם קשורים . רחוקים מן הגן המדובר, DNAם על פני היאתרים המצוי –אנהנסורים
. והם אחראים על הזמן והמקום בו הגן ישועתק, bazalפקטורי שעתוק שאינם חלק מה
חלבונים המעכבים גיוס פקטורי שעתוק מסוג ) לאנהנסורים יכולים להיקשר רפרסורים
bazal )או פקטורים המאקטבים שעתוק .
Chromatin remodeling
– histone codeמתאפשרת בזכות ה GCאו ל TATAל( מכל סוג)גישה של פקטורי שעתוק ון גורמות לארגה שונות העוברים מודיפיקציות( וארגנין( בעיקר)ליזין )להיסטונים ישנם שיירים
.מחדש של ההיסטונים
של מודיפיקציותאלא על ( יסטאיןצשיירי ) DNAת על והמתרחש לא מדובר במודיפיקציותכ תגרום לפתיחה של "מתילציה מסוג זה בד, לדוגמא(. של החלבונים, ההיסטונים עצמם
.DNAהכרומטין כך שמתאפשרת גישה של פקטורי השעתוק ל
הוא בעצם צירופים שונים של מודיפיקציות באתרים מסוימים בכרומטין histone codeהלהתרווח , ים להתפרקינוקלאוזומים עשו –המסמנים לתא מהו הארגון שצריך להיות בכרומטין
אזור שלם בכרומטין עובר –מדובר בצירופים מורכבים מאוד . צריך להתרחש שינוי כלשהו –' וכד . כל צירוף גורם א יודעים למהת כאשר כיום עוד למודיפיקציו
:של הכרומטין remodelingתהליך ה
היסטון לגיוס ובכך גורם DNAב חשוף נקשר לאתר GFPשעתוק הפקטור ה להיפתחות ראשונית של מבנהגורמת אצטילציה אנזימים אלו מבצעים .אצטילאזות
ולאחר , באתרים שונים היסטון קינאזות גורמות לפוספורילציהלאחריהם . הכרומטין
כך קומפלקס זה מגיע . chromatin remodeling complexמגויס הקומפלקס מכן
מבנה שונה לחלוטין של יוצרוממשיך עד כי ,histone codeלאחר שנקבע כבר ה
כך שנותר אזור ( sliding)שינויים אלו כוללים תזוזה של הנוקלאוזומים . הכרומטין
. bazal transcription machineryפנוי ל
יש לזכור כי ישנם פקטורי שעתוק האחראיים על ארגון מחדש .
יש הבחנה בין הremodeling לבין המודיפיקציות (modifying).
. חלבונים שונים שאינם חלק מן הקומפלקס יגויסו גם כן, remodelingלאחר השאזור השעתוק ם אותו כךמשנים את מבנה הנוקלאוזום או מסלקיפרונים 'צ, לדוגמא (. הקומפלקספעילות בעיקר לאחר )תרשים מתאר את סדר הגעת החלבונים ה .מתרווח

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
:לאחר שעתוק mRNAתהליכי עיבוד
:(עוד בגרעין) ים בשעתוק יוקריוטיקיימהמתם עיקריים תהליכי 3ישנם
(.'3קצה )פוליאדנליציה .1
2. Capping – (.'5קצה )מתיל גואנין 7קישור
3. Splicing – בו גנים מסוימים ,ביצורים יוקריוטים עילאיים בלבדתהליך המתרחשלדוגמא יהיו יותר גנים כאלו לעומת בבני אדם) .של חלבונים יותר גדולהופכים למספר
(.שמרים
:פוליאדנילציה
של ' 3המגנה על קצה ה, AMPנוקלאוטידים מסוג 200ישנה תוספת של כ, לאחר סיום השעתוק
מגבירים את אשר גם , חלבונים נוספים רצף זה קושר, בנוסף. מפני עיכול mRNAמולקולת הלדוגמא )של מולקולות בקרתיות אחרות ', 3קישור נוסף לקצה ה מתווכיםם וג, יציבות המולקולה
miRNA המעכבות שעתוק.)
אומנם באופן שונה , תלוית רצף גם טרמינציה ביוקריוטים – טרמינציית השעתוק ביוקריוטים
אלא, להפסיק את השעתוק RNAלא מאותת ל DNAביוקריוטים הרצף על ה. מאשר בפרוקריוטים . לחלבונים אחרים
AAUAAA – רצף קונצנזוס(צא בשכיחות הגבוהה ביותר בקצה היימmRNA )
כך , צריך להיגמר mRNAלאזור בו ה upstreamנוקלאוטידים 20יימצא כ
מקודד לסיום גם הוא , Uוב Gהעשיר ב, רצף נוסף. שיתקיים שם חיתוךשני רצפים אלו . נוקלאוטידים מרצף הפוליאדנילציה 50מצוי כו ,השעתוק
מן RNA pol 2הסיום לא מתרחש בנפילה פשוטה של ה. הווים סיגנל לסיוםמטורים האחראיים על שני פק קשורים של אנזים זה אל הקצה המזורחן –הגדיל
תק את שני רצפי שע RNA pol 2לאחר שה .CstFו CPSF –הפוליאדנילציה
נקשרים ו RNA pol 2משתחררים מן ה, פקטורים אלו מזהים אותם, הטרמינציה
נוקלאוטידים 20כ)בנקודה זו RNAמתווכים חיתוך של הבכך הם . עצמו RNAל
downstearm השתחררו מן הלאחר שהם (. לרצף הפוליאדנילציהRAN מבנהופקטורים , בנוסף. השעתוק מופסקכך ש, וכך גם הזיקה שלו לגדיל, משתנה המרחבי
שתפקידו , PAP (poly adenilation polymerase)האנזים מתווכים קישור של אלו
הוא –עושה זאת RNA pol 2לא ה – (poly Aלהוסיף רצף )ליצור את הפוליאדנילציה . סיים את תפקידו לאחר שנחתך מן הגדיל
mRNAהמבקרים את יציבות ה poly Aחלבונים נוספים נקשרים לרצף ה, בהמשך .ועשויים להשתתף בתהליכי בקרת התרגום
Capping:
של מולקולת ' 5קצה ל( מעט שונה מן הגואנין הרגיל)מתיל גואנין 7קישור נוקליאוטיד מסוג
הקישור . '5ל' 5פחמן בין אלא '3דרך פחמן לא –ור הוא קישור הפוך הקיש. נוצרתה mRNAה תימצאלא כ "ישנה קבוצת מתיל שבדעל הגואנין ' 7כאשר באתר , נעשה בין שתי קבוצות הפוספט
מודיפיקציה .'5מפני אקסונוקלאזות מסוג ' 5הקישור של מולקולה זו מגן על הקצה . בגואנין רגיל .בכל התאים היוקריוטים/ מסוג זה תתרחש

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
בהם מתרחש קישור של קבוצות מתיל ( לא יוקריוטים, טפילים)ישנם יצורים . את הקצה ולהגן עליו לסמן שמטרתו' 5נוספות לקצה ה
י חלבונים "מתווכת ע (פוליאדנילציהוגם ה) CAPהוספת קבוצת ה, כפי שתואר
אין ,כלומר .תוך כדי תהליך השעתוק, RNA pol 2טרמינלי של ה Cהקשורים לזנב ה
. יתרחש cappingצורך לסיים את השעתוק בכדי שה
Splicing:
.המקטעים הבונים את החלבון –אקסונים . mRNAמסולקים מן ה –אינטרונים
ואקסונים תמיד טרונים תמיד יהיו אינטרוניםהאינ, קבוע מתבצע באופןישנם גנים בהם תהליך
בגן אחד הם אינטרונים – alternative splicing ישנם גנים המקיימים, לעומתם. יהיו אקסונים
. DNAחלבונים שונים מאותו רצף כך שמתקבלים –אחר של גןאקסונים
Cם על הזנב הימעורבים חלבונים המצויוגם במהלכו , תוך כדי שעתוקתהליך זה מתבצע גם
. גופיף המכונה ספליסוזום, י קומפלקס חלבוני"מבוצע עתהליך זה . RNA pol 2טרמינלי של ה
את הפעילות אשר הן המבצעות, snRNAמולקולות כיל מקומפלקס זה , מלבד לחלבונים . הקטליטית
:רצפים הקריטיים לתהליך 3ישנם
1. 5'SS – 5' splice site – סוף אזור המחבר בין. (צהוב) האינטרוןלסוף ( כחול)האקסון הראשון
כך – AGGUהרצף באזור זה כמעט תמיד יימצאשני הנוקליאוטידים האחרונים באקסון יהיו ש
AG ,הראשונים באינטרון יהיו והשנייםGU.
2. Branching point – חלק מן האינטרון.
3. 3' splice site – 3'SS – גם בו יימצא הרצף , אתר החיתוך השניAGGU.
ובמקביל , U1 –המרכזית snRNAנקשרת מולקולת ה SS'5לאזור ה
, מיד לאחר מכן. brancing pointאחרת נקשרת ל snRNA (U2) מולקולת
,כך שנוצר קיפול U1נקשרות ל U6ו U4 –נוספות snRNAשתי מולקולות
loop (טבעת אותה ניתן לראות בתרשים ) המכונהlariat . הקיפול גורם
, שברצף החיתוך השני Aאל ה (השייך לאינטרון) GUלקישור ה
.lariatוהאינטרון משתחרר כ
פעולתם של : הערהU4 וU6 האת גורמת להם להחליףU1 ובכך
.לקצה השני SS'5את הקצה ה להעביר
מולקולת , במקבילU5 ת את האקסונים בקצותיהם מחדש קושר
mRNAאת קיום מולקולת ה ומאפשרת, לכרי רצף בודדישתחרר רק לאחר חיבור האקסונים זה האינטרון .החדשה
. לזה
כלשהו לא פנוי splice siteספלייסינג אלטרנטיבי יתרחש כאשר אזור הספליסוזום לא , יגרום למיסוך של אזור זה חלבון כלשהו אם –לקישור

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
תהליך זה . הרצף שהיה אמור להיות אינטרון יהפוך לאקסון ויתורגם לחלבון עקב כך. יזהה אותו .קיים בכל הגניםלא
:הערות לגבי תהליכי העיבוד
כל מולקולות הsmall RNA (U למיניהן )נקשרות לאתרים השונים על הmRNA י "ע
.SSהן מכילות נוקליאוטידים משלימים לאתרי ה –השלמת בסיסים
מדובר בפעולה הנחשבת כמתרחשת , גורמת לסיום השעתוק פוליאדנילציהלמרות שה . תוך כדי שעתוק
של ה' 3לזנב ה ,כפי שניתן לראותRNA pol 2 ,האזור הC י "טרמינלי שעבר פוספורילציה ע
אזור זה באנזים בעל –חשיבות גדולה בכל תהליכי העיבוד , TFIIHפקטור השעתוק
.splicingוגם ב cappingגם ב, ם בפוליאדנילציהגתפקיד
RNA editing:
וההערכה היא כי , תהליכי העיבוד שתוארו 3בשנים האחרונות נתגלה תהליך נוסף ללאחר היווצרות , ובמהלכו, התהליך מתרחש ברקמות מסוימות .חשיבותו גדולה מאוד
יאוטיד בצורה בה הוא יהפוך לנוקל, נוקליאוטידתבוצע מודיפיקציה ל, mRNAמולקולת ה
Cמולקולת ( סרת קבוצה אמינית והחלפתה בחמצןה)דה אמינציה י תהליך "ע, לדוגמא. שני
.לא מדובר בהחלפת נוקליאוטיד אלא בשינוי מולקולה קיימת. Uתשתנה ותהפוך ל
אשר )ין אינוז – Iל ,י דה אמינציה"ע, Aהיא הפיכת RNAסוג עוד יותר נפוץ של עריכת
עריכה .Gכ יזהוהאנזימים שבתא אותו, (בצורה טבעית RNAלא קיים כנוקליאוטיד ב
בעת תרגום אנו עשויים לקבל חומצה אמינית – RNAזו גורמת לשינויים בקריאת ה
שיופיע במרכז המולקולה ולא stop codonאו אף , DNAאחרת מזו שציפינו לה לפי ה . בסופה
רק ברקמה בה , חלבון הספציפי לרקמה מסוימת הגוף יכול לגרום לסינתזת, בעזרת תהליך זהנקבל חלבון שלם , כלשהו לא מתרחש ברקמת הכבד תהליך עריכה אם ,לדוגמא. הוא נדרש
מאותו גן את נקבל בעקבותיו, stop codonתהליך עריכה יגרום לקבלת מעיבעוד בתא , ותפקודיי "תהליך עריכה יכול להפוך חלבון מקוצר לחלבון ארוך יותר ע –לחילופין . מקוצרהחלבון ה
. stop codonביטול
תהליך הediting מתרחש גם הוא בגרעין.
תהליך זה מעורב בהיווצרות תאים סרטניים מאחר ומעורב בהיווצרות שברים בDNA.
מודיפיקציות שונות המתרחשות –סיכום ההבדלים בין תאים פרוקריוטים ליוקריוטים
יצא mRNAרק לאחר מודיפיקציות אלו ה. ביורקיוטים לא יתרחשו בפרוקריוטים, בפרוקריוטים השעתוק מתרחש בציטופלזמה, לעומת זאת. לציטופלזמה ויתורגם לחלבון
.מבלי שיתרחשו מודיפיקציות שונות –ולכן התרגום מתחיל עוד לפני שהשעתוק נגמר

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
mRNA export – מולקולת ה, לאחר כל המודיפיקציותmRNA זו תצא מן הגרעין אל . הציטופלזמה ותעבור תרגום
mRNA degradation:
שעות או , ותהן יציבות ובעלות זמן חיים שיכול להימשך מספר דק mRNAמולקולות , באופן כללי
. הוא תהליך בעל חשיבות בקרתית ואחראי על רמת הביטוי של כל גן RNAתהליך פירוק ה. ימים
. כ יהיו יציבים יותר מאשר גנים אחרים"בד Housekeeping genes, לדוגמא
עוד לפני שיגיע mRNAתהליכי העיבוד השונים הם המונעים ממנגנוני הפירוק להרוס את ה
cap מולקולה חסרת –ה בי אזורים שונים ש"מוגדרת ע mRNAיציבותה של מולקולת . לריבוזום
. יציבות של מולקולה עשויות להשפיע על, אם קיימות, uORF. אקסונוקלאזות' 5חשופה לפעילות
poly Aרצף ,כ"בד. חלבונים המפקחים על יציבות המולקולהשוי לקשור לעצמו ע UTR '3קצה המגיע לגודל עד כי ( בהתחלה לאט)מתקצר עם הזמן , ולההמצוי בקצה המולק
– חלבונים מייצבים שהיו עליו ייפלו, זה בגודל(. נוקלאוטידים 30של כ)קריטי
א לא תהיה כך שהי mRNAאקסונוקלאזות יעכלו את מולקולת ה' 3וכעת כך', 5מגנים גם על קצה ה' 3חלבונים אלו שהיו על קצה ה, בנוסף .שמישה
פירוק . וכחות אקסונוקלאזותשני הקצוות פגיעים בנ poly Aשללא ה . '5ל' 3כ יתרחש מ"המולקולה בד
RNA interference – מולקולות של נוסף תהליך עיכולRNA ,עיכול . התלוי ברצפים ספציפייםורך בפירוק דחוף של מולקולת כאשר יש צ ייעשה רקו, המולקולה לא מתרחש בקצוותמסוג זה
RNA המולקולות המבצעות תהליך זה הן ה. כלשהיsiRNA – small interference RNA –
. אחרות RNAמולקולות יציבות המתערבות ב RNAמולקולות
גבירו את כמות העותקים של ה ולכן, רצו לקבל צבע חזק יותר בפטוניה –גילוי תופעת ההתערבות ולא חזק , אומנם בחלק מן המקרים נתקבל דווקא צבע בהיר יותר .גנים האחראיים על הצבע
הגורמים , antisense רצפייטוי י ב"בבקרה שהפעיל הצמח עבתחילה חשבו שאולי מדובר . יותר
.עברו השתקההמקודדות לצבע י RNAשחלק ממולקולות ה לכך
כי כאשר מכניסים מקטעי הראה C eleganceב רךנעמקביל ש מחקר. קרהה לא המסתבר כי זה
DS RNA פירוק של הההשתקה נובעת מ. הגן מושתק, של גן מסויםmRNA כלומר –התאי ,
של (השתקהובכך ל)פירוק גורמות ל, דו גדיליות( נוקלאוטידים 20)קצרות RNAמולקולות
. mRNAמולקולות
:dsRNAישנן שתי משפחות של מולקולות
- siRNA – מולקולות בעלות רצף זהה לחלוטין למולקולת הmRNA מפרקות . המושתקת
.mRNAאת מולקולת ה
- miRNA – מולקולות .אך לא זהה, מולקולות בעלות רצף דומה למולקולה המושתקת
.אך לא מפרקות אותה, mRNAאלו מבקרות ומעכבות מולקולות
, (pre RNAמולקולת ) גדולה RNAאלו מיוצרות בתא בתור מולקולת מולקולות
ם שני כבר בהכך ש, רצפים משלימיםמכילה זו מולקולה. RNA pol 2י "ע
המקטעים המשלימים מזהה את DROSHAהאנזים .מקטעים דו גדיליים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
70כ כל מבנה אורכו – stem & loops שמתקבלים מבניכך וחותך אותה pre RNAמולקולת השב . אוטידיםינוקל
י קומפלקס חלבוני "שם היא מזוהה ע, מוצאת לציטופלזמה DROSHAי ה"שנחתכה ע מולקולה
דו גדילי RNAמקטעי – (נוקלאוטידים 20כ)יותר החותך אותה למקטעים קצרים( dicer)אחר
בו ,RISC (RNA induces silencing complex) –מועברים לקומפלקס נוסף מקטעים אלו . קצרים
, (י קישור אליהן"ע) RNAמזהה את מולקולות ה RISCלאחר ההפרדה ה. עוברים הפרדה הם . כך שהן כעת חלק מהקופלקס –תוכו ומעביר אותן ל
:יםיים להתרחש שני דברשוע, mRNAמולקולת במקרה בו הקומפלקס יכניס לתוכו
מדובר) mRNAה יפרק אתהקומפלקס , קישור מתבצע תוך התאמה מלאהאם ה -
.(siRNAב
(. miRNA)בו בלבד עכי אלא mRNAה את קהקומפלקס לא יפר, תאמה לא מלאהאם הה -
קומפלקס הRISC פועל כדימר העובר במקביל על שני הגדילים .

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
24.11 – 12הרצאה –ביוכימיה
5318963 –טלפון מעבדה רמי דון.
:Translation –תהליך התרגום
, יוצא מן הגרעין ומגיע אל הריבוזום mRNAלאחר שה, תהליך התרגום מתבצע בציטופלזמה
mRNAנקשרת ליחידה הקטנה התת ,בתחילת התהליך. קטנה וגדולה :יחידותתת המורכב משתי
תתחבר תת , לאחר שיימצא הקודון התחילי. ראשון AUGבחיפוש אחר את המולקולה וסורקת
נכנסות, ת עם חומצה אמינית המתאימה להכל אח, tRNA מולקולות לאחר מכן. היחידה הגדולה
היא משתחררת אז, stop codonהשרשרת הפפטידית מתארכת עד כי הריבוזום מגיע ל. לריבוזום
:שתי הערות. mRNAוהריבוזום מתנתק מן ה
mRNA כמו )' 3ל' 5י הריבוזום מ"תמיד ייקרא ע (.שני התהליכים הקודמים
החלבון המיוצר תמיד מיוצר מן הקצה הN
כך החומצה האמינית . טרמינלי Cטרמינלי לקצה ה
Nתמיד תהיה בקצה ה, מתיונין, הראשונה .(לפחות לפני שלבי עיבוד שונים) טרמינלי
Initiation:
. uORFלעתים גם ו, פרומוטר ,מתיל גואנין 7 בו מצוי, UTR' 5צף יש מקטע המורכב מר ORFלפני ה
' 3ישנו ה stop codonלאחר ה. stop codonותסתיים ב AUGתתחיל תמיד ב mRNAשעל ה ORFה
UTR. רצף פולי אזור זה יכילביוקריוטיםA ,יו קשורים חלבונים שונים ואלmiRNA המבקרים . את התרגום
:בנוגע לקוד הגנטיהערות
הקודון התחילי ,AUG ,החומצה האמינית , בפרוקריוטים. מקודד בפרוקריוטים למתיונין
.AUGי "מקודדת ע וגם היא, היא פורמיל מתיונין הראשונה המוכנסת לרצף
ישנם מקרים בהם הריבוזום יתחיל את התרגום לפי קודונים אחרים :GUG ,AUU ,
UUG .יוצא . לה אלטרנטיביים וקיימים בעיקר בפרוקריוטיםכונים קודוני התחאלו מ
אשר התרגום שלו יכול להתחיל , הקיים בתאים יוקריוטים PIN2חלבון דופן הוא ה
. CUGויכול להתחיל מ, AUGמ
יכולה להיות לאוצין, לדוגמא. י יותר מקודון אחד"מרבית חומצות האמינו מקודדות עבעוד האות , שתי האותיות הראשונות זהותבהם –קודונים שונים 4י "מקודדת ע
החומצה האמינית סרין מקודדת . degenerate code: מצב זה מכונה. משתנההשלישית האחרות משתנים 2אומנם ב, מוחלפת האות השלישיתהראשונים 4ין הב –קודונים 6י "ע
קודונים 4מרבית חומצות האמינו כוללות רק . נוקליאוטידים אחרים גם . תאו פחו
. יוקריוטים, פרוקריוטים, קיים בוירוסים –הקוד הגנטי שמור מאוד באבולוציה עניין זה מבטא את הלחץ –יום התא כל שינוי גורם למוטציה הלא מאפשרת ק
–יוצא דופן מכלל זה הוא הקודון התחילי . שמפעיל הטבע שמטרתו שימור קוד זה . ו גם הן לפורמיל מתיוניןישנן מעט וריאציות של הקודון התחילי אשר יקודד

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
עשוי , לחומצה כלשהי דנוספים בהם קודון המקוד םמעטיישנם מקרים , start codonבדומה ל
mRNAקודד להמ, אליהמיטוכונדרי DNAקיימים ב אלו ייםשינומרבית . תאחר חומצהלקודד ל
AUAהקודון , לדוגמא. הקוד הגנטי עשוי להיות מעט שונה בה, ם בתוך המיטוכונדריהורגתהמ .גם בחיידקים ישנו מעט שוני. אומנם במיטוכונדריה מקודד למתיונין, כ לאיזולאוצין"מקודד בד
:תרגום בפרוקריוטים
:תחילת התהליך
עליו למקם את עצמו בצורה נכונה כך שידע לזהות את , mRNAלפני שהריבוזום סורק את ה
וכך –חלבונים שונים 3מקודדת לבודדת mRNAמולקולת , באופרון לדוגמא, לפעמים. AUGהוכן , את תחילת הגן השני, את סיומו, לת הגן הראשוןמנגנון התרגום צריך לדעת לזהות תחי
. הלאה
Gהעשיר בSD (shine delgarno sequence )בחיידקים קיים רצף – זיהוי אתר תחילת התרגום
המשלימה RNA (16 s rRNA )בריבוזום החיידקי קיימת מולקולת .המאותת לתחילת התרגום, Uו
הבא הוא הקודון AUGבעצם מסמן לריבוזום כי ה SDה. כך שהוא מזהה את האתר, SDלרצף הלאחר זיהוי הרצף הריבוזום מתארגן לקראת .ולא סתם קודון המקודד למתיונין, התחילי . התרגום
ולכן עליו , האחראי על תרגום כל הגנים, מצוי בתוך הריבוזום הוא –שמור ביותר SDרצף הכן יהיה זהה בכל אומנם –רצף זה לא קיים בצורה זהה בכל הפרוקריוטים . להופיע בכל פרומוטר
.מין חיידקי בודד הפרטים של
lacZהגן בתרשים ניתן לראות את , לדוגמא
. הקיים בו SDאת רצף הו ,lac-operonמצוי בה
ORFשני , lac operonתן לראות כי בבמצגת ניקודון , לכל אחד מהם יש קודון תחילי –נוספים
.SDורצף , עצר
:tRNAמבנה
' 3קצה . לת מבנה דו ממדי בעל צורת תלתןבע, מולקולה חד גדיליתהיא tRNAה
(. אחרון A) CCAסתיים בנוקליאוטידים של כל מולקולה כזו י
בעוד , מסמלים את הנוקליאוטידים השמורים tRNAהחלקים האפורים שב
tRNAאחת ל tRNAהאזורים התכולים הם האזורים המשתנים בין מולקולת
רק או (Y)כלשהו פירימידיןרק ישנם אתרים במולקולה העשויים להיות . אחרת
בנוסף .וישנם שיכולים להיות מורכבים מכל נוקליאוטיד שהוא, (R)כלשהו פוריןהרצף הכתום מסמל את האנטי . (דוגמת פסוידו יוירידין) ישנם אתרים העוברים מודיפיקציה
של ' 3הקשורה לקצה ה)ולפיו מוספת החומצה האמינית הנכונה mRNAהאזור הנקשר ל –קודון
(. tRNAה
ניתן לראות משמאל בתרשיםtRNA בה מקטע, של לאוצין variable loop, השונה מאוד
. שאר המולקולה זהה, זה loopמלבד ל. של ליזין tRNAמן המקטע הקיים ב
ניתן לראות תמונה תלת ממדית של מימיןtRNA.

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
: המודיפיקציות המתרחשות על מולקולה זו
.פסואידו יורידין -
. זה RNAאומנם הוא כן קיים ב, RNAנוקליאוטיד אותו אנו לא מכירים ב –ריבותימידין - . הוא לא דאוקסי, הוא תימידיןלמרות שנוקליאוטיד זה
.tRNAב Dכמסומן –דהידרויורידין -
יאלחומצה שה tRNAה מולקולת כנראה ומשמעות המודיפיקציות היא בזיקה הספציפית שביןבאופן מפורט יותר . ולהכרת הקודון המתאים לה, ריבוזוםב ליכולת ההשתלבות שלה, תקושר
.המודיפיקציות עוד לא ברורתפקיד
באופן , נעשית כמו כל הכרה דו גדילית mRNAלקודון שעל ה tRNAההכרה בין האנטיקודון שב
. tRNAשל ה '3יהיה מול ה mRNAשל ה' 5קצה ה – אנטי מקביל
Wobble pairing – אין צורך , למרות שישנן חומצות אמינית המקודדות לפי מספר קודונים
, tRNAמבחינת ה)הנוקליאוטיד השלישי שבקודון . אחת לכל קודון, שונות tRNAבמולקולות
& Watsonשהיא לא מסוג , מסוגל ליצור קשרי מימן בצורה גמישה( '5מדובר בקודון שבעמדה
crick .טבלת קוד גנטי קטנה המתאימה לנוקליאוטיד השלישי:
ולקודון , AAAיודע להיקשר לקודון , UUUהאנטיקודון , לדוגמא
AAG .הבסיס אנטי קודון : דוגמא נוספתI (אינוזין ) יוכל ' 5שבעמדה
. בקודון' 3שבעמדה Aאו U,Cלזהות את
מספר מולקולות הtRNA (. קודונים 61חומצות אמיניות ו 20לעומת ) 21כ "בד –שבתא
ב קבוצת הפורמיל מוסרת בשל. ועליו פורמיל מתיונין tRNAלריבוזום נכנס , AUGלאחר זיהוי הכך נוצר מצב בו לא כל חלבון חיידקי . ולעתים תוסר גם המתיונין איתה, (תמיד) מאוחר יותר
החומצה האמינית , התהליךבזמן , עם זאת. יתחיל במתיונין( וקריוטיםיוהתופעה נכונה גם ל)
.יקודדו למתיונין רגילה AUGשאר קודוני ה. הראשונה תמיד תהיה מתיונין
:בחומצה אמינית tRNAטעינת ה
Cדרך הקצה ה)בשלב הראשון נוצר קשר בין החומצה האמינית .1תוך , אל האדנין( הקבוצה האלפא קרבוקסילית, טרמינלי
. שחרור קבוצת פירופוספט
AminoAcyl tRNAי האנזים"תוצר הביניים מזוהה ע .2
synthetase ,של ה' 3הקושר את חומצה זו לקצה הtRNA
(. ACCל)
על ) OHשתי קבוצות מכיל tRNAולקולת השל מ' 3הקצה ה
AminoAcylהאנזים . אותן ניתן לראות בתרשים, (2ו 3פחמן
tRNA synthetase קושר את החומצה האמינית והאדנין לאחת
ההבדל )כך שיכולים להתקבל שני תוצרים – OHמקבוצות ה (. ביניהם חסר משמעות
אותו י"ושניהם מבוצעים ע, של החומצה האמינית שני השלבים שתוארו מהווים אקטיבציה
.AminoAcyl tRNA synthetase ה, אנזים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
. לליזין 2, אחד לכל חומצה אמינית, AminoAcyl tRNA synthetaseאנזימי 21ישנם E.coliב
איןtRNA קבוצת הפורמיל תיקשר למתיונין רק לאחר –מיוחד לפורמיל מתיונין
. tRNAשהמתיונין תיקשר ל
לקשור את tRNAלאילו מולקולות בעזרתם האנזים יודע , השונות tRNAישנם הבדלים בין ה
הוא הקובע את הקישור לחומצה tRNAהאנטיקודון שעל ה, בחלק מן המקרים. אמיניתהחומצה הישנם אתרים נוספים על פני המולקולה אשר הם ,במקרים אחרים. האמינית
האתרים )כנושאת חומצה אמינית ספציפית לה אחראיים על הזיהוי של המולקו
עשויה לגרום למוטציה tRNAכך בעצם מוטציה אשר תתרחש ב. (אדוםבהמסומנים . אפילו אם הגן תקין לחלוטין –בחלבון
בהן אזור האנטיקודון לא קשור כלל לקישור tRNAניתן לראות כי ישנן מולקולות ולקולה עדיין תקשור את גם אם נשנה אותו לחלוטין המ –החומצה האמינית
זה גם תגרום למוטציה מוטציה מסוג . רהחומצה האמינית אותה היא אמורה לקשו . תכניס חומצה אמינית מסוימת לפי קודון אשר לא מתאים להמאחר ו, בחלבון
:ת סופרסורמוטציו
אומנם ביחד יבטלו זו את זו , הן בעצם שתי מוטציות אשר בנפרד יגרמו לכשל מוטציות סופרסור
הגורמת , מוטציה הקיימת בגן היא Nonsense mutation, לדוגמא. ויאפשרו חלבון תקין
, אומנם אם המוטציה גרמה לשינוי בנוקליאוטיד השלישי שבקודון. stop codonלהיווצרות
קודון העצר לא יוכר כקודון עצר אלא ש לגרום לכךעשויה tRNAב מוטציה נוספת שתתרחש . כקודון המקודד לחומצה אמינית
:ריבוזומים בפרוקריוטים
:מורכבים משתי תת יחידות
תת יחידה זו היא . חלבונים נוספים 21ומ rRNA 16S מורכבת מ: 30S, תת יחידה קטנה .1
. SDהמזהה את רצף ה
.חלבונים נוספים 31ו rRNA 23S, rRNA 5Sמורכבת מ :50S, תת יחידה גדולה .2
כמו כן המבנה המרחבי של , המבנה המרחבי של כל המולקולות המרכיבות את הריבוזום פוענח .המבנה מצוי במצגת. הריבוזום כולו
Initiation:
:פקטורי אתחול התרגום
מתנתקת מתת אשר עקב כך, תת יחידה הקטנההנקשרים ל (IF1, IF3)פקטורי אתחול ישנם שני
ייקשר IF2פקטור האתחול , SDלאחר עצירה באתר ה. mRNAנקשרת לו היחידה הגדולה
, mRNAנקשרת לת היחידה הגדולה במצב זה ת. הנושאת מתיונין tRNAלקומפלקס ואיתו
. מתנתקים מן הקומפלקס IFושלושת ה
מאחר והIF2 הפורמיל מתיונין תופיע רק ב, מצוי רק בתחילת התרגוםtRNA ולא , הראשון . בשאר הקודונים המקודדים למתיונין
:elongation –תהליך התרגום

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
לאחר. Aהחדש המצטרף תמיד ייכנס לאתר tRNAה. tRNA :E,P,Aעמדות בהן יהיו 3בריבוזום
אל החומצה החדשה המצויה באתר מועברת Pאתר השרשרת הפוליפפטידית המצויה ב, כניסתו
A . כך האתרP נותר רק עםtRNA ,כך –הרצף מתקדם שלשה . ללא שרשרת וללא חומצה אמינית
אתר ) Eיהיה כעת באתר Pשהיה באתר והקודון, Pמצוי כעת באתר Aשהקודון שהיה באתר
(. exitה
תוך ) Aהוא המביא את החומצת האמינו הנכונה לעמדה GTPהקשור ל TUפקטור האלנוגציה
האנזים הקיימת יוצר צה האמינית החדשה לשרשרתבין החומ את הקשר הפפטידי(. GTPפירוק
peptidyltransferase .
ומקבלת אותה בעזרת פקטור, תנועת הריבוזום לעבר השלשה הבאה צורכת אנרגיה
. Gהאלונגציה
תהליך טעינתTU מחדש דורש גם הואGTP י פקטור האלונגציה "ונעשה עTs .
:termination –תהליך הסיום
.כך שהיא נותרת פנויה, לעמדה זו המתאים tRNAאין , Aמצוי בעמדה stop codonכאשר ה
Releasing factors 1 לאתר ייקשרו( כל אחד מזהה קודונים מסוג אחר) 2אוA .ייקשר לאחריהם
RF3 , בהידרוליזת אנרגיהתוך צריכתGTP .פירוק הGTP יגרום לtRNA שבעמדהP ולאחריו , לעזוב . דיתגם השרשרת הפוליפפטי
פקטורי אתחול 3קיימים –סיכום מנגנון התרגום בפרוקריוטים(IF) ,3 פקטורי
elongation (Ts, TU, G )3ו releasing factors .ישנם , על אלו למדנו בהרצאה קודמת .נוספים
:תהליך התרגום

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
29.11 – 13הרצאה –ביוכימיה
Initiation:
כאשר לפני התרגום, (70S)פרוקריוטי שלם מצוי בתא כיחידה אחת ריבוזום
IF3כנראה ו) IF3ו IF1י פקטורי האתחול "תבוצע ע 30Sול 50Sהפרדתו לולאחר ,לתת היחידה הקטנהפקטורים אלו ייקשרו . (אחראי על ההפרדה
GTPהטעון במולקולת , IF2נקשר הפקטורים אל. mRNAשב SDלאזור ה מכן
(. fMET)הנושא פורמיל מתיונין tRNAוב
נוצרת זיקה טובה לקישור תת mRNAקשורים יחד ל IFה 3בו –במצב זה
נפרדים תוך פירוק IFהשלושת לאחר הקישור. היחידה הגדולה לקומפלקס
GTP (וקבלת אנרגיה ) יחידה תת , תת יחידה גדולה –ונותר הקומפלקס
, P מצוי בעמדה tRNAבמצב זה ה. fMETהנושאת tRNAו mRNA, קטנה
. הבא tRNAפנויה ל Aועמדה
Elongation:
ובחומצת GTPעצמו במולקולת טוען TUהפקטור . (EF) גם בתהליך זה ישנם שלושה פקטורים
הוא תהליך זה )יבוזום הפנויה שבר Aלעמדת ה tRNAא את הולאחר מכן מבי, אמינו ספציפית
(. GTP (פירוק)הדורש הידרוליזת
peptidyle transferase את הקישור בין חומצה אמינית זו לשרשרת הקיימת מבצע האנזים
קטליטית המאפשרות RNAמולקולות הפעילות הקטליטית של האנזים מקורה ב. המצוי בריבוזוםהשרשרת הקיימת של פעילות המולקולות נעשית כך ש. יצירת קשר פפטידי בין חומצות האמינו
התהליך ) טרמינלית Cהנוספת לקצה ה, Aחומצות האמינו מועברת לחומצה האמינית שבעמדה . (מוצג בתרשים
עמדה , בהמשך. tRNAמצויה מולקולת Pמצויה השרשרת הפפטידית ובעמדה Aבמצב זה בעמדה
A אלונגציה נוסף בעזרת פקטור מתפנה– G , אשר גם הוא טעון במולקולתGTP. האנרגיה בעזרת
של ' 3של הריבוזום שלשה אחת בכיוון ת התקדמותאפשרתמ GTPהידרוליזת ה הנפלטת בתהליך
mRNA .במצב זה הtRNA הועבר לעמדהE כך שיכול להשתחרר מן הריבוזום.
מפקטור האלונגציה GDPהוא לשחרר את מולקולת ה, Tsפקטור , הפקטור השלישיהתפקיד של
TU ,במולקולתמחדש אותו לטעון ו GTP כך ה. חדשהTU יוכל להביא חומצה אמינית חדשה אל . הריבוזום
Termination:
במקרה בו – stop codonגיע לכאשר הריבוזום מ. RF1, RF2, RF3 –פקטורים 3גם בתהליך זה
(לא משנה איזה) RF2או RF1פקטור , tRNAהתהליך תקין ולא קיימת מוטציה באנטי קודון של ה
קישור זה יגרום לניתוק השרשרת הפוליפפטידית . וייקשרו בסמוך לו stop codonאת אתר ה יזהו
, לאחר מכן .ללא חומצה tRNAוה( 2או 1) RFה רק ותרויובריבוזום שכך ,Pשבעמדה tRNAמן ה
מתנתק מן tRNAכך שה, Aסמוך לאתר בנקשר לריבוזום , GTPגם הוא טעון ב, RF3 הפקטור .יותרכיום ידוע כי מדובר בתהליך מורכב .הריבוזום

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
RF3 פקטור זה קושר . הוא מין פקטור שחזור המאפשר את שחזור התהליךGTP , ובכך מאפשר
ולהשתחרר מן Eהאחרון יוכל להגיע לעמדה tRNAכך שה', 3תזוזה אחרונה של הריבוזום בכיוון
ייקשרו לתת 3ו IF 1כאשר , היחידות אחת מן השנייהמאוחר יותר מתפרקות גם תת . הריבוזום .היחידה הקטנה והתהליך יתחיל מחדש
חומצות האמינו ' אם נגדיר את מס)פפטיד שלם י הפוספט בהם השתמשנו בכדי לסנתזקשר כימות
(:Nשבפפטיד כ
יש צורך – aminoacyl tRNA synthetaseי "ע ATPהחומצה האמינית מאוקטבת בעזרת -
.N GTPבכ
.N GTP – הפירופוספט לאחר מכןפירוק קשר -
.חלבוןהבודדת לכל GTP דורשת מולקולת IF2פעילות ה -
.N GTPיש צורך בכ. GTPקושר TUפקטור האלונגציה , לגבי כל חומצת אמינו נפרדת -
. N GTPגם דורש לפעילותו Gפקטור האלונגציה -
. בודדת לכל החלבון GTPדורשת מולקולת RF3פעילות ה -
. קשרי פוספט לחלבון 4N +2 (≈ 4N)יש צורך ב –כלומר
Poly Ribosome – כאשר הריבוזום נקשר למולקולת . ביוקריוטיםחיידקים וגם ב מבנה הקיים גם
ונוצרים , בו זמנית מצויים מספר ריבוזומים על המולקולה –הוא לא מצוי שם לבד , mRNAה
. poly ribosomeמצב המכונה – מספר פפטידים
ולהגיע לאתר בו הוא , החלבון צריך לעבור קיפול מתאים, השרשרת הפוליפפטידית נוצרתש לאחרהחלבון יופרש אומנם לעתים ,מצוי בתוך התא עצמו( בו הוא פועל)לעתים אתר זה . צריך לפעול
או , (התא המרווח שבין דופן התא לבין ממברנת)בחיידקים למרווח הפריפלזמי – אל מחוץ לתאורם שיעביר יש צורך בג –הממברנה דרךאם החלבון מצוי בתוך התא ועליו לצאת . מחוץ לחיידק
י "ע)חלבונים ספציפיים המזהים את החלבונים שצריכים להיות מופרשים לשם כך ישנם. אותוומוליכים אותם לתעלות ספציפיות המפרישות את החלבון , (רצף סיגנל הזהה בכל חלבונים אלו
י "ייחתך ע ,שסימן אותו ככזה שצריך לצאת לרוב הרצף, לאחר שהחלבון יצא מן התא. מן התעלה .פרוטאזות
:אנטיביוטיקה
ת כולן מונעות א, סוגים שונים של אנטיביוטיקה 5דוגמאות ל. זיהום חיידקי בלבדלנגד תרופת :ריבוזוםי פגיעה ב"יצירת החלבונים החיידקיים ע
זו נקשרת לאתר בריבוזום אליו צריך להיכנס האמינו אציל מולקולה –טטראציקלין .1
tRNA ( אתרA .)א ל)להתארך הרצף הפפטידי לא יוכל, כאשר היא נכנסת לאתר, כךמאחר ומבנה הריבוזום החיידקי שונה . לבוןיצירת הח ותימנע, (יתווספו חומצות אמינו
תרופה זו יעילה . יםלא תפגע בתאים יוקריוט המולקולה, ממבנה הריבוזום היוקריוטי . כנגד מגוון רחב מאוד של חיידקים
לפורמיל IF2קישור פקטור האתחול בעיקר את מונעת מולקולה ה –סטרפטומיצין .2
לא fMETמאחר ו. חלבונים לא יוכלו להתחיל להיווצר בחיידק והוא ימותכך ש, מתיונין .היא לא תפגע בתאים כאלו, קיים ביוקריוטים
כיום . פוגעת במערכת השמיעה –יקה זו היא כי היא אודיוטוקסית הבעיה עם אנטיביוטיתרונה הוא כי היא . המודעות לנזק זה גרמה לכך שהשימוש בה נעשה בצורה מחושבת
.יעילה מאוד כנגד זיהומים

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
–ומונעת את התקדמות הריבוזום לשלשה הבאה 23S rRNAנקשרת ל –אריתרומיצין .3
chloramphenicolאנטיביוטיקה גם ה. Gהאלונגציה מונעת את השלב בו משתתף פקטור .(מונעת אלונגציה) פועלת בצורה דומה
.תרופה הפוגעת בתאים יוקריוטים ופרוקריוטים –פורומיצין .4
מעבדה ברבות משמשת. תרופה הפוגעת בתאים יוקריוטים בלבד –ציקלוהקסאמיד .5 (. חלבון שכבר תורגםלבטל תרגום חלבונים בכדי לזהות רמת יציבות של כשיש צורך
, מלבד לממברנה(. ולא במנגנון התרגום)פוגעת במנגנון יצירת הדופן של החיידק – פניציליןמבנה המורכב משרשרות סוכריות המחוברות –ישנה דופן הבנויה מפפטידוגליקן לחיידקים
. י פפטידים קצרים"ביניהן ע
אחד ההבדלים המרכזיים בין חיידקים גרם חיוביים דופן זו היא (תרשים תחתון) בחיידקים גרם שליליים –לגרם שליליים
לעומת חיידקים גרם חיוביים , הפפטידוגליקן הוא דק מאודלכן חיידקים גרם חיוביים . בהם הדופן עבה מאוד( תרשים עליון)
.פגיעים יותר בנוכחות פניצילין
לק עליו לפרק את הדופן וליצור אותה בכל פעם בה החיידק מתחדק לא יוכל לבנות את הדופן החיי, בנוכחות פניצילין. מחדש
. מחדש וימות
הבעיה בפניצילין היא כי בחיידקים מסוימים ישנו פלסמיד . כך גן זה מקנה עמידות כנגד הפניצילין. המכיל גן שהתוצר החלבוני שלו יודע לפרק את הפניצילין
הסיכוי . הסיכוי להתפתחות זן עמיד גדל, באנטיביוטיקה בצורה תכופה יותר ככל שאדם ישתמשבו כמות גדולה של )ח "כי חולה יפתח בעצמו זן כזה נמוך בהרבה מהסיכוי כי חולה המטופל בביה
.(אנטיביוטיקה
אדם המקבל טיפול לכן –נוטות לפגוע בחיידקי הקיבה האנטיביוטיות שהוזכרו (. 'יוגורט וכד)יומלץ לאכול מאכלים בהם חיידקים טבעיים , טיקהאינטנסיבי של אנטיביו
:תרגום ביוקריוטים
. מדובר במנגנון מאוד דומה המערב כמות גדולה יותר של פקטורים
לדיסוציאציה וגורם( 60S)לתת היחידה הגדולה נקשר( מסמלת יוקריוטי E) eIF6פקטור האתחול
אשר כרגע מופרדת נקשרים פקטורי אתחול ( 40S)ה לתת היחידה הקטנ. בין שתי תת היחידות
אומנם ישנם , אותם צריך לזכור) IF4Eו IF4G, IF4Aהמורכב מן הפקטורים , IF4F: ביניהם, רבים
פקטור זה . mRNAשל ה '5ובכך מעביר את כל הקומפלקס לקצה , CAPנקשר ל IF4Eהפקטור (.עוד
תהליך . הראשון AUGעד כי הוא מוצא את ה mRNAרץ על מולקולת הו '5קצה מתיישב על
. ייחודי ליוקריוטים mRNAה סריקת
ביטוי מוגבר שלו גורם להתפתחות של גידול סרטני – IF4Eהאתחול חשיבות נוספת של פקטור . לא ברור עוד כיצד, (פקטור שעתוק זה הוא אונקוגן)
, לאחר מכן. הנה הקטהוא מגייס אליו את תת היחיד', 5לאחר שהקומפלקס נקשר לקצה ה
נקשר ( אך רגילה) MET-tRNAול GTPגם הוא קשור ל, וקריוטיהפר IF2הפקטור המקביל ל
בתהליך זה גם הפקטורים . הראשון AUGעד כי הוא מגיע ל mRNAאשר סורק את ה, לקומפלקס
IF3 וIF4C עוזרים בקישור הMET-tRNA.

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
הפקטור .גם היא mRNAתת היחידה הגדולה נקשרת ל, הראשון AUGכאשר הקומפלקס מגיע ל
IF5 כך . מסלק את פקטורי השעתוק שהיו בקומפלקס ומאפשר את קישור תת היחידה הגדולההריבוזום השלם מצוי על המולקולה ויכול להתחיל את תהליך התרגום בצורה מאוד דומה למנגנון
. הפרוקריוטי
:וקריוטים והיוקריוטיםטורים הפרהשוואה בין הפקטבלת
. AUGומחפשים mRNAקומפלקס היחידה הקטנה וכל פקטורי האתחול רצים על גבי ה: הערה
. לא נקבל חלבון כלל –שלא יהיה במסגרת הקריאה –מוקדם AUGאם מוטציה תגרום ליצירת
טרמינלי Nקצה הנקבל תוספת קצרה ל –המצוי במסגרת הקריאה AUGאם מדובר ב, אומנם . חלבון שרצינו לקבלב
uORF – עולה השאלה איך , מאחר ולמדנו כי מנגנון התחלת התרגום הוא מנגנון של סריקה
ישנם (. mRNAולא נופל מן ה)הריבוזום יודע כי קיים חלבון נוסף לאחר המקטע החלבוני הראשון :מספר מודלים שהוצעו
ORFייפול מיד ויספיק לזהות את ההריבוזום לא , אם מדובר במרחק קטן בין השניין -תזהה את , ייתכן ותת היחידה הקטנה תישאר, קאם הריבוזום יספיק להתפר. השני
.הנוסף והתהליך יאותחל בשנית AUGה
אם כן נרצה שהחלבון יעבור .תרגום החלבוןאת מטרתו למנוע –הוא בקרתי uORFה -
. mRNAנוספת על הרמת בקרה – stem & loopיהיה במצב של uORFה –תרגום
:י אותם ריבוזומים"המתבצעים ע, בציטופלזמה ישנם שני סוגי תרגום, (בלבד)ריוטים קבתאים יו
1. Free ribosomes – ת וריבוזומים חופשיים בציטופלזמה הפוגשים במולקולmRNA .חלבונים אשר פועלים בציטופלזמה יתורגמו בצורה זו .תורנדומלי

. 2010 –2009' שנה ב, ביוטכנולוגיה בר אילן, י הגר לביאן"נכתב ע
2. mRNA צמודות לER – לאזורי הrough ER ,ריבוזומים אלו יתרגמו. עליה ריבוזומים , תעלותולשמש כ)חלבונים אשר צריכים לצאת מן התא או להיות מעוגנים בממברנה
ומאפשרת את תהליכי , י'היא רשת ממברנות האחראית על ייצור הגולג ERה(. רצפטוריםאשר יישלחו , עיותבו, הוא יוכנס לוזיקולות, ר באזור זהווצכאשר חלבון יי. האקסציטוזה
. לממברנה לאחר יצירתן
Nבקצה ה signal peptideמכיל רצף , ואמור להישלח אל מחוץ לציטופלזמה, ERחלבון המיוצר ב
ומאפשרים את כניסת מזהים את רצפים אלו SRPחלבוני . המשמש כסיגנל, טרמינלי של החלבון
כשכבר לא יהיה בו , (ERב) יעבור חיתוך signal peptideמאוחר יותר רצף ה .ERהחלבונים לתוך ה . צורך
הנקשר אליו , SRPי ה"חלבונים אלו יזוהו ע –בנוגע לחלבונים שתפקידם להיות מעוגנים בממברנה
וישנם כאלו שצריכים ERישנם רצפים בחלבון שצריכים להיות בתוך ה. ERופותח לו תעלות באו , מגדירים כי יש להכניס את המקטע הבאכך על החלבון ישנם רצפים ה. להיות מחוץ לה
ונותר , הוא לא נכנס לתעלה, מתורגם( כתום) stop transfer seqכאשר ה. להוציא אותו .בציטופלזמה
ואזורים ( מחוץ לממברנה)כאשר בודקים חלבון מסוג זה ניתן לזהות אזורים הידרופיליים או שמעוגן , מדובר בחלבון טרנסממברנלי י כך לשער האם"וע, (יהיו בתוך הממברנה)הידרופוביים
. 'וכד, באופן חלקי לממברנה

1 . 2010 –2009, בר אילן' ביוטכנולוגיה שנה ב, י הגר לביאן"נכתב ע
1.12 – 14הרצאה –ביוכימיה
צריכת –בכדי לקיים את התופעה יש צורך בסימני חיים . החיים הם תופעה המקיימת את עצמהר את הבעיה התהליכים אותם מלמדת וחוקרת הביוכימיה נועדו לפתו. יכולת רבייה ועוד, נרגיהא
. של קיום החיים
לא מדובר . מתחלפים כל הזמן שבו תאיםהולכן , הגוף נשחק. תאים הם לא אינסופייםוה הגוף מדובר בשחיקה של החומר. לאחר תקופה כולם נהרסים –בניין , גם אבן –בתופעה ביולוגית
300האקלים קרוב ל –הסביבה החומרית היא סביבה תרמית . שנובעת מן הסביבה בה אנו חייםים כולם יובילו להרס התהליכים השונ –חום הוא תנועה אקראית של חלקיקים . מעלות קלווין
. אי הסדר רק יתגבר עם הזמן – שינוי מבפנים המוביל לאנטרופיה בכיוון בודד –מבנים מסודרים
היא התפתחות עוברית , לדוגמא. תופעת החיים היא תופעה היוצרת מבנים ומקיימת סדר, אומנם, תופעת החיים. שנים 15תאים לאחר כ 1018הופך ל, המתחיל בתא אחד, גניזםהאורתהליך בו
, ולכן יש צורך בתהליכים רבים ובתופעות שונות, רהתופעה הטבעית מנוגדת לגמרי לעולם החומ . הסותרת את קיומם אנטרופית בסביבה מסודרים יאפשרו קיום חיים שכולם
:כים הכלליים ביותר המאפשרים חייםהתהלי
האנטרופיה היא ביטוי בו אנו . אנטרופיה –כל גוף עובר שינוי מבפנים הגורם להרס שלו ישנם תהליכים המייחדים את , מאחר והחיים הם תופעה לא טבעית. לתיאור הרס זה משתמשים
הכימיה של התא שונה מהכימיה של . האורגניזם בכך שמאפשרים לו לבצע היפוך תהליכיםן ריאקציות של הריאקציות הביוכימיות ה –בתא לא נשברים או נוצרים קשרים –המבחנה
:העקרונות .החלפת קשרים
גניזם רהאו זהבעזרת מנגנון . תופעה הכללית ביותרה –הכימיה של צימוד תהליכים .1התהליכים , ייצור אנטרופיה, תהליכי פירוק טבעי –מצמד את התהליכים הטבעיים
לנו לשם כך יש . תהליכי בנייה –לתהלים ההפוכים לטבע –הספונטניים של שינוי מבפנים , הן החלבונים –מכונות צימוד אלו . על פעולתן האורגניזם פנוי –" מכונות צימוד"צורך ב
תנועה תרמית , י חום"נים מופעלים עהחלבו. החומר הראשון הנחוץ לקיום האורגניזם .צורך בהחלפתםהגורמת לשחיקתם כך שיש
.החלפת קשרים במקום בנייה ופירוק –עקרון שני .2
ממולקולה –י כך שכל המבנים "עולם החיים מתקיים ע. החלפה, ורמחז –עקרון שלישי .3עצם העובדה שהאדם חי רק . כולם מתחלפים –עד לאטמוספרה , אורגניזםדרך ה, בודדתישנן מולקולות הקיימות בגוף רק למשך מספר . שנים היא לא תקלה אלא כוונה 100
, ורגניזם צעיר ולא שחוקכל הזמן קיים אכך . יום כל תא דם מוחלף 120תוך . שניות . ובזכות כך החיים מתרחשים ומקיימים את עצמם, מולקולות צעירות נטולות תקלות
החיים הם –הביולוגיה עברה אופטימיזציה לא לטובת הפרט אלא לטובת המין המסקנה היא כי כדי אלא רק ב, לעד מסיבה זו הפרט לא חיי. נועדו לקיום המין אלא, לא פרטיים לגבי כל אדם
מבחינת הביולוגיה האדם סיים , לאחר הולדת הצאצאים. להוליד צאצאים – לבצע את תפקידו . ואין בו עוד צורך, את תפקידו
. תהליכי בנייה עצמיתבו( הגנום)יש צורך במנגנון כימי , בכדי לקיים את תהליכי הבנייה
כי אנו לא משתנים מתארתהקביעות . עקרון הקביעות וההשתנות: עקרון משני וחשוב .4גם הקביעות היא אינה . י הגנום"הנקבעת ע מוכוונתבצורה אקראית אלא בצורה
.והתרחשה האבולוציה, עקב מוטציות נגרמות מחלות –אבסולוטית
:התנהגות המולקולות
חיבור , השינויים הבסיסיים ביותר ברמה המולקולארית, הצעדים האלמנטאריים של כל תהליךהכל זז באופן שוטף –לפי עולם החומר , נעשים באופן אקראי –של שני אטומים אחד לשני

2 . 2010 –2009, בר אילן' ביוטכנולוגיה שנה ב, י הגר לביאן"נכתב ע
העיקרון הבסיסי ביותר המאפשר ? בנייה והתפתחות, כיצד נוצרים סדר, אם כך. ובצורה אקראיתהשמת –קבלת תהליך מכוון לפי עקרונות אקראיים –סדר במקום בו אין סדר ויצירת , בנייה
היא תפעל רק בכיוון בו אינה –כאשר ישנן מגבלות על תנועה אקראית . מגבלות על המערכת . מוגבלת
( מפלי חום חסרת)התא הוא מערכת איזותרמית . לותפקיד הביוכימיה הוא ליצור מגבלות אקת בשאלה מהם מפלים יוכימיה עוסהב. היוצרת מפלים כימים –( חץמפלי ל חסרת)ברית ואיזו
כיצד הכוחות בתוך התא מכוונים את . פוטנציאל אלקטרוכימימפלי אנרגיה חופשית ו, אלו . ולא מפלי חום או לחץ, התהליכים על בסיס מפלי ריכוזים
באמצעות , עצמיתישנו זיכרון כימי ומכניזם של בנייה , בכדי לשמר את הקביעות, ברמת המיןאומנם יוצרות כיווניות בגלל , מכונות המונעות לפי תנועה אקראית של חום, ריאקציות של העברה
שני עקרונות אלו הם המבדילים בין . ועוד שני עקרונות חשובים נוספים( חלבונים)המבנה שלהן .בלבדאלא את המצב השני , האורגניזם לא מקיים שיווי משקל. שיווי משקל למצב עמיד
בה , למערכת המצויה במקסימום אנטרופיה, כימית, דינמיתהגדרה תרמו –משקל שיווילא משנה לאיזה שינוי נגרום –מדובר במצב יציב . ההסתברות לשינוי היא קטנה מאוד
הקביעות של הפרמטרים המאקרוסקופיים . הקיום הממוצע של המצב יישאר אותו דברכאשר אנו מצויים –לפי המכניזם . שיווי המשקלהיא הגורם המאפשר לנו למדוד את
אין עוד יכולת שינוי , לא ניתן יותר לגרום לאי סדר –במצב של מקסימום אי סדר ולכן לא , שינוי, התפתחות, אומנם האורגניזם דורש גידול. ולכן המערכת יציבה, מבפנים
.יכול להתקיים במצב של שיווי משקלקביעות ניתן להשיג . יש לנו צורך בקביעות, דבר מסוכן למרות ששיווי משקל יכול להיות
. י השקעת אנרגיה"ע, במצב שהוא לא מקסימום אנטרופיה, במצב לא יציבאם ננסה להשיג . המתקיימת לאורך זמן מטה סטביליות היא מצב של יציבות מוגבלת
.לאורך זמןלא נוכל לשמר אותו , שיווי משקל מקומי, יצירת מינימל מקומי י"מצב זה עמ הרצוי "מ המקומי לש"זאת מאחר והפרעות קטנות יכולות להסיט את המצב מהש
קיום מצב מטה סטבילי לאורך זמן הוא קיום שלא בנוי .כל הזמן( מקסימום אנטרופיה) . י כוחות מנוגדים מאוזנים אשר ישמרו על מצב יציב"על מקסימום אנטרופיה אלא ע
המאקרוסקופיים של המערכת הם יציבים ואינם מצב בו הפרמטרים –מצב עמיד, הכוח יכול להיות חשמלי. י השקעת כוח מתמדת"אומנם יציבותם נקבעת ע, משתנים
בכדי לקיים את , לכן האדם צריך להשקיע אנרגיה באופן שוטף. מכני או כל אחד אחרורגניזם כאשר הא. מצויה במצב עמיד, או הביוספרה, אורגניזם, לכן כל תא. המצב העמיד
. מת הוא מגיע למצב של שיווי משקל
בכדי לשמור את המצב העמיד במצב יציב נדרשים זרמים של חומר ואנרגיה –הומיאוסטזיס מדובר בזרמים מנוגדים המאזנים זה את זה ומאפשרים את תהליכי . במבנים והמכונות שבגוף
מהווים , ים לתוצאה הפוכהשני חומרים הגורמ –האינסולין והגלוקגן , לדוגמא. שימור הגוף . כוחות מנוגדים המווסתים זה את זה
:כיצד מבוצעים התהליכים הביוכימיים
. עליהן מבוסס כל עולם החי, המכונות של החייםהן ( פולימרים)מאקרומולקולות חלבונים , (ארבעה בסיסים)רעין חומצות ג, המאקרומולקולות העיקריות של עולם החי הן סוכרים
תת יחידות אשר 60ישנן כ, כלומר(. מתילנים של חומצות שומניות)ושומנים ( ת אמינוחומצו 20)נובע כולו , גיוון עצום, הגיוון של עולם החי. הן אחראיות על בניית המאקרומולקולות
.הסדר שלהן בשרשרת הפולימר, מקומבינציות שונות של תת יחידות אלו
דוגמת , להבדיל מפולימרים כימיים)גיות התכונות החשובות של המאקרומולקולות הביולו (:פלסטיק

3 . 2010 –2009, בר אילן' ביוטכנולוגיה שנה ב, י הגר לביאן"נכתב ע
ולכן מאפשרים , (לא עשויים ממונומר אחד)פולימריים הפולימריים הביוכימיים הם הטרו .1 .הסדר של המונומרים בתוך הפולימר הוא התכונה החשובה הראשונה. סידורים שונים
יתרחשו ,ייםפולימרבניגוד להומו, קיפול והמבנים המרחביים של ההטרופולימריםה .2קרומולקולות יש למ. ה הנבנית על פי ספציפיותיכולת הבחנ –ים בהעדפה לכיוון מסו
, בגוף ישנם נוגדנים. אומנם רק אחד הוא המועדף ביותר –אינסוף מבנים אפשריים או נקשרים לחומר ספציפי , המקיימים פעילות ספציפית חלבונים אחריםו, תעלות יוניות
ית הבנייה עקרון זה הוא מהווה את הפתרון לבעי. מועדף שלהםבזכות המבנה ה – בלבד . קרומולקולותי המ"הנעשית ע, העצמית
ינן לבין מספר חומצות אמינו שא( פוליפפטיד)ההבדל העיקרי בין שרשרת חומצות אמינו וגדלה ההסתברות , הוא כי קיימת בה כמות אי סדר קטנה הרבה יותר, מחוברות זו לזו
השמנו מגבלה על החומצות האמיניות ולכן יותר סביר –צוי הספציפי לקיום המבנה הר, הקישוריות מקטינה את מספר אפשרויות הסידור. כי נקבל אותן בצורה בה הן רצויות
ובכך את פער האנטרופיה בין המצב –מקטינה את האנטרופיה של המצב ההתחלתי –ברת נמדדים במיליונים סדרי הגודל של ההבדלים באנטרופיה המדו. הסופי להתחלתי
. ההסתברות קטנה פי מיליון לפחות
תכונת חלבונים המאפיינת את –המונומרים בשרשרת מאפשר לנו לשמר מידע סידור .3. נגרום לאיבוד המידע, אם ניצור הידרוליזה לחומצות אלו. תפקידן של חומצות הגרעין
לא היה ידוע מי האחראי על נשיאת , למרות שידעו זמן רב על קיום חומצות הגרעין, בעבר . המידע הגנטי
מאחר ומוכתב לפי , יהיה דומה, גם אם שונות, מבנה אוסף מולקולות, בעזרת הקישוריות .4. לכל מונומרים מסוג מסוים יש מבנה חלקי משותף. זהה בין מונומר למונומר קישוריות
כך הדבר . הקשר הפפטידי זהה בין כל חלבונים בכל האורגניזמים –גם בין פולימרים הדבר – בכדי לבנות חלבונים בגוף הטורף, ן הצומחמ, מאפשר שימוש בחלבונים מן החי
.את קיום שרשרת המזון, מאפשר את המחזור
ליצור פולמרים אף ניתן. שלישוני, שניוני, מבנה ראשוני –קיום היררכיה של מבנים .5 . מבנה רביעוני – היוצרים אינטראקציות עם פולימרים אחרים
לא , בנויה על אינטראקציות חלשות ,או בתוכן, ולקולותקרומהאינטראקציה בין המכל .6ת ביחס לקישור כולן חלשואומנם , ר או חלשות יותרישנן חזקות יות. קוולנטיותאומנם כל קישור ( בין המונומרים)בכל מקרומולקולה קיים קישור קוולנטי . הקוולנטי
. נוסף יהיה חלש מכךאלא ביחס לכך שהן –ההגדרה של הקשרים כחלשים לא נעשית ביחס לקישור הקוולנטי
יש לנו היכולת . לשבור קשרים אלוולכן יש לנו היכולת , בתנועה אקראית, תלויות בחוםהקשרים , כל הריאקציות הפיכות –בין רצפטור לליגאנד , להפריד בין נוגדן לאנטיגן
בפיקו . אשר היא תלויה בחום, מהירות המולקולותבוהכל תלוי –שנוצרים נשברים כאשר מספיק . מולקולה בודדתמאקרוכאלף מולקולות מתנגשות ב, שנייה בודדת
האינטראקציות . ל עליה עולה על הכוח של התאחיזההכוח המופע, מולקולות מתנגשות (. מולקולות מים, מולקולות התמיסה)חלשות ביחס לכוח הממוצע המופעל על מולקולה
0.6ים היא האנרגיה הממוצעת של תנועת המ, יםבה אנו חי, קלווין 310בטמפרטורה של או יותר 10-100פי אינטראקציה יציבה צריכה להיות גדולה, כלומר. קילו קלוריות למול
היא מקור היחיד , אנרגיית המים, ההתנגשויות האקראיות. מאשר אנרגיה זו של המיםסיבה זו היא הגורמת לכך שיש לנו צורך . לכל פעולה ותהליך ביולוגיים( טיןלחלו)
.בטמפרטורת גוף קבועה
:אינטראקציות חלשות
, כוח שנוצר באמצעות שני גופים –טראקציה היא פעולה הדדית ינא. מתקיימות לאורך טווח קצרמסיבה זו אנו (. שני דיפולים, בדומה לשני מטענים חשמליים)זה על זה כוח אשר הם מפעילים
מאחר , זמן הקיום של אינטראקציות אלו הוא קצר .ולא על קשר, מדברים על אינטראקציהלפי עקרונות . מן להתנגשויות מולקולות מיםוהמצב המטה סטבילי בו הן מצויות נתון כל הז
כאשר באופן כללי , ללית משתנה וגורמת לכמה אפשרויותעוצמת ההתנגשויות הכ, הסטטיסטיקה

4 . 2010 –2009, בר אילן' ביוטכנולוגיה שנה ב, י הגר לביאן"נכתב ע
י היחס בין הכוח שהיא מקיימת לכוח שמפעילות עליה "זמן החיים של אינטראקציה נקבע ע .מולקולות המים המתנגשות
שלא , קילו קלוריות 0.6בניגוד להתנגשות של , רום לניתוקקילו קלוריות למול תג 22התנגשות של . ההסתברות היא כי ככל שההתנגשות תהיה חזקה יותר כך היא תהיה נדירה יותר. תגרום לדבר
אומנם אם . היא תופר פעמים רבות, קילוקלוריות למול 1.5אם מדובר באינטראקציה שגודלה של כל מצב , אורך החיים של האינטראקציה .היא תופר כפעם בדקה, קילו קלוריות 10גודלה
להתגבר על , באנרגית ההתנגשות הנדרשת לבטלה סטייה מן הממוצעי גודל "נקבע ע, מטה סטבילי (. שבין המולקולות)כוח המשיכה
אם האינטראקציות בין . תלויה ביכולת האינטראקציות להתפרק בזמן, יכולת האורגניזם לזוזאורגניזם לא היה מסוגל לרוץ ברגע בו הבחין –ת יותר משהן עכשיו מולקולות השריר היו חזקו
.בסכנה
:סיכום
קיים תאנו מסוגלים לה. הן המקרומולקולות, לבעיית קיום החיים, ברמה המולקולארית, הפתרון . בזכות מספר קטן מאוד של רכיבים היוצרים אינסוף מבנים אפשריים
אין שניים בעלי , מיליארד בני אדם בעלי גנום דומה 6תופעה המאפשרת כי מתוך –פולימורפיזם (. מלבד תאומים זהים)גנום זהה לחלוטין
שהיא , הן המאפשרות את תכונת הבנייה עצמית, כל התכונות שתיארנו הנוגעות למקרומולקולות . שחייו נוגדים את עקרון הטבע ,פותרת את בעיית האורגניזם
.המצב העמיד מתקיים בכל מערכות החיים .המצב העמיד של הפרט –הומיאוסטזיס
שכבה שומנית, הגורם המאפשר לתא תכונה זו היא הממברנה. החיים הבסיסיתיחידת –התא המפלים . ומאפשרת קיום מפלים כימיים, היוצרת הפרדה, ננומטר 4יה כישעוב ,חסרת חוזק מכני
ילות הביוכימית של הפע. הכימיים הם המאפשרים את פעילות הגורמים והחלבונים שבתאמאפשרת פוטנציאל המאפשר יצירת כיווניות מתוך תנועה , הממברנה היא כי היא מהווה חיץ
הפונקציה של הממברנה היא לא פונקציה . מפל זה הוא המניע את כל התהליכים. אקראית –לדוגמא . מפלי הריכוזים ומפלי המתחים הם הגורם לכל העבודה המתבצעת בתא. מכנית
, והזרמתם דרך המכונה, י יצירת מפל פרוטונים"נעשית בקרום המיטוכונדריון ע ATPזת הביוסינת . בכיוון המפל, דרך הקרום
יגרום –חור בודד בתא . הם המאפשרים כל פעילות –מפלי הריכוזים , מפלי הפוטנציאל הכימי . אנטיביוטיות רבותתרופות פועלות , לפי עקרון זה. למותו רק בגלל סיבה זו