geneti čke osnove oplemenjivanja organizama · fenotip 10 18 20 učestalost 0.36 0.48 0.16 • ako...
TRANSCRIPT

genetičke osnove oplemenjivanja organizama

• Oplemenjivanje je proces razmnožavanja, ukrštanja, biljaka i životinja (i mikroorganizama) sa ciljem dobijanja željenih osobina. – tako dobijene jedinke pripadaju linijama, rasama, sortama,
sojevima, varijetetima, kultivarima– njihovim medjusobnim ukrštanjem dobijaju se hibridi
• “konvencionalne” metode oplemenjivanja– veštačka selekcija, inbriding
• manipulacija genima i genomima – stvaranje ploida, genetički modifikovanih organizama
• kombinovanje konvencionalnih metoda i molekularnih tehnologija– mapiranje genoma, Molekularno Asistirana Selekcija

• Fenotipovi mogu biti determinisani – monogeno – poligeno– interakcijom gena i sredine
• Kvalitativne osobine• Kvantitativne osobine
– metričke– numeričke– osobine “sa pragom
ispoljavanja

• Fišerov model kvantitativne genetike• poligeni, poligeni + variranje sredine,
interakcije gena i sredine
kontinualna distribucija verovatnoće opisuje učestalosti javljanja genotipova i fenotipova u populaciji


aditivni efekat više alela daje više fenotipova

Slika 10.4. Fenotipske distribucije za osobinu determinisanu potpunom dominansom (gornji deo slike) ili epistatičkom interakcijom alela (donji deo slike) (prerañeno iz Hamilton, 2009).
• nepotpuna dominansa i epistaze menjaju distribuciju fenotipskih učestalosti u odnosu na aditivni model
• alelske učestalosti ostaju iste • neke fenotipske klase izostaju

• efekat sredine na fenotipsku distribuciju

Interakcija uticajagenotipa i sredine

komponente fenotipske i genetičke varijabilnosti
• Vf = Vg+ Vs
• Vf = Vg + Vs + COV gs + Vgxs

komponente genetičke varijabilnosti
• Vg = Va + Vd+ Vi
• Va aditivna varijansa - varijansa usled prosečne razlike izmedju nosilaca različitih alela – gametski doprinos
• Vd varijansa iz dominanse, koja potiče od toga što heterozigoti nisu uvek intermedijarni u odnosu na homozigote
• Vi interakciona varijansa - rezultat razlika u epistatičkim interakcijama izmedju lokusa

• Ukoliko nema razlike u prosečnom efektu alela na jednom lokusu, na primer A i a, onda nema aditivne varijanse, ali ostaje varijabilnost u fenotipovima izmedju genotipova koja je neaditivna.
• Ako heterozigot nije sasvim intermedijaran izmedju homozigota već ima parcijalne dominanse, cilj je da aditivan efekat uzrokovan zamenom alela odvojimo od varijanse uzrokovane dominansom (Vd)
• Ako više od jednog lokusa utiče na neku osobinu, svaka epistatička interakcija izmedju lokusa će uticati na neaditivnu varijansu (Vi)

kompletna dominantnost jednog alela nad drugim (+d, -d)ako jedan od alela kodira za manje aktivan genski produkt ili za
potpuno neaktivan produkt i ako je jedna kopija alela dovoljna zafiziološko funkcionisanje organizma, nema dominanse
ako je fiziološka aktivnost proporcionalna količini aktivnoggenskog produkta, heterozigotni fenotip je tačno intermedijaran izmedjudva homozigotaparcijalna dominansa (h)
– heterozigoti nisu sasvim na sredini (bliži su jednom odhomozigota.
overdominansa– heterozigoti mogu biti van fenotipskih opsega homozigota

Ako je jedan lokus, dva alela:
(+a) je genotipska vrednost jednog homozigota(-a) je genotipska vrednost drugog homozigota
(d ) je genotipska vrednost heterozigota (podrazumevajući da A1 alel povećava vrednost osobine).
Prose čna vrednost kvantitativne osobine u populaciji

prose čna vrednost fenotipa za celu populaciju množenjem vrednosti svakog genotipa njegovom učestalošću i deljenjem sa ukupnim zbirom jedinki u populaciji.
M = a(p-q)+2dpq = genotipska i fenotipska vrednost za tu osobinu u populaciji
doprinos pojedina čnog lokusa prose čnoj vrednosti populacije može biti kroz homozigote a(p-q) i heterozigote 2dpq.
– ako nema dominanse, d = 0, M = a(1-2q)– ako je potpuna dominansa, d = a, prosečna vrednost M = a(1-2q2).
Tabela 10.1. Učestalosti alela i njihov uticaj na prosečnu vrednost osobine u populaciji
genotip učestalost vrednost učestalost x vrednost
A1 A1 p2 +a p2a A1 A2 2pq d 2pqd A2 A2 q2 -a -d2a
zbir = M = a(p-q)+2dpq
-q2

prosečan efekat alela (α)
• ima populacioni smisao i zavisi od genotipskih vrednosti (a i d) i od učestalosti alela– prosečno odstupanje od srednje vrednosti populacije onih jedinki
koje su dobile taj alel (npr. A1) od jednog roditelja, dok alel od drugog roditelja nasleñuju slučajno iz te populacije
– prosečna vrednost tako nastalih genotipova zavisi od prosečnog efekta datog alela..npr A1..
Tabela 10.2. Prosečan efekat alela (α) u odnosu na prosečne genotipske vrednosti (a i d).
tip gameta
vrednosti i učestalosti genotipova
srednja vrednost
genotipova
srednja vrednost populacije
prosečan efekat alela
(α)
A1A1 A1A2 A2A2 +a d -a
A1 p q pa+qd -[a(p-q)+2dpq] q[a+d(q-p)]
A2 p q -qa+pd -[a(p-q)+2dpq] -q[a+d(q-p)]
-p

• prosečan efekat jednog alela je prosečan fenotip svih jedinki koje ga nose i stoji u zavisnosti od učestalosti genotipova
Primer: ako dva alela, A1 A2, segregiraju na lokusu koji utiče na visinu. U nekoj sredini srednje fenotipske vrednosti (visine) i učestalosti tri genotipa mogu biti:
A1A1 A2A1 A2A2fenotip 10 18 20učestalost 0.36 0.48 0.16
• Ako se fenotipske srednje vrednosti razlikuju - postoji genetička varijansa za tu osobinu u populaciji

• aditivan efekat, aditivna genetička varijansa (Va) je prosečan efekat na fenotip koji potiče od tzv. zamenealela A1 sa alelom A2, tj., prosečna visina jedinki saalelom A2 je veća nego svih jedinki sa alelom A1.
• U Primeru: brojem alela A2 ili A1 u genotipu, množenjemvisinama jedinki (fenotipskom vrednošću) i množenjem učestalošću kojom se javljaju, u odnosu na ukupnu učestalost homo- i heterozigota koji imaju taj alel dobija se prosečan efekat alela A1:= [2(0.36)(10)+1(0.48)(18)]/2(0.36)+1(0.48) = 13.20cm
i prosečan efekat alela A2 = 18.80cm
• prosečna razlika izmedju efekata A2 i A1 = 5.60cm

Prose čan efekat alela i oplemenjiva čka vrednost
• prose čan efekat alela ( α) omogućava da se utvrdi oplemenjivačka vrednost jedinki, koja se odnosi na alele koje jedinke prenosesledećoj generaciji a koji utiču na odreñenu kvantitativnu osobinu
• oplemenjivačka vrednost jedinke može se izraziti kao zbir prosečnih efekata alela koje ona nosi, po lokusu i zbirno za više lokusa.
Tabela 10.3. Oplemenjivačka vrednost jedinki tri genotipa jednog lokusa
genotip oplemenjivačka vrednost A1A1 2α1= 2qα A1A2 α1+ α2= (q-p)α A2A2 2α2= -2pα

ODSTUPANJA USLED DOMINANSE
mogu se izraziti preko datih genotipskih vrednosti a i d, oduzimanjem oplemenjivačkihvrednosti od genotipskih (Tabela 10.3)Primer: genotip A1A1 ima genotipsku vrednost (+a), a populaciona srednja vrednost M
je:M = a(p-q)+2dpqgenotipska vrednost izražena kao odstupanje od M: a-[a(p-q)+2dpq] = a(1-p+q)-2dpq = 2q(a -dp)zamenom (a) prosečnim efektom alela (α) (Tabela 10.4) iz α= a+d(q-p): a= α-d(q-p),)genotipska vrednost je: 2q(α-qd).oduzimanjem oplemenjivačke vrednosti 2qα dobija se odstupanje usled dominanse:-2q2dAnalogno za genotip A1A2 odstupanje usled dominanse je 2pqd, za A2A2 je -2p2d. • sva odstupanja usled dominanse su u funkciji od d: ako nema dominanse d = 0
odstupanja su nula, pa su oplemenjivačke i genotipske vrednosti jednake
Tabela 10.4. Vrednosti genotipova u dvoalelnom sistemu, u odnosu na populacionu srednju vrednost M = a(p-q)+2dpq. Prosečan efekat zamene gena je α = a +d(q-p). genotipovi A1A1 A1A2 A2A2 učestalosti p2 2pq q2 vrednosti a d -a odstupanja od populacionog proseka: genotipska vrednost oplemenjivačka vrednost odstupanje usled dominanse
2q(a-pd) 2q(α-qd) 2qα -2q2d
a(q-p)+d(1-2pq) (q-p)α+2pqd (q-p)α 2pqd
-2p(a+qd) -2p(α +pd) -2pα -2q2d
-2p2d

Ako se razmatra pojedinačni lokus,genotipsku vrednost (G) čine oplemenjivačka vrednost (A), kojaje aditivni deo genotipa i odstupanje usled dominanse (D)
Ako se razmatra više lokusa,Ukupnu genotipsku varijansu (G) čini zbir oplemenjivačkih vrednosti (A) za svaki lokus, odstupanja usled dominanse (D) za svaki lokus i odstupanje usled interakcije medju lokusima (I).
Ako nema dominanse medju alelima genotipska i oplemenjivačka vrednost su jednake
Sve ove vrednosti zavise i od učestalosti alela u populaciji.

Ukupno komponente fenotipske varijabilnosti
• Vf = Va + Vd+ Vi + Vs + COV gs + Vgxs
I sredinska varijansa može poticati od spoljašnje sredine, unutrašnje sredine i materinskog efekta., pa su Vs = Vsu + Vss + Vm (komponente sredinske varijabilnosti)

Interakcije sredinske i geneti čke varijabilnosti• norma reakcije genotipa:
jedan genotip može dati više fenotipova u različitim sredinama

ZA SVAKI GENOTIP JE NEKA SREDINA BOLJA
U SVIM SREDINAMA JEDAN GENOTIP JE BOLJI OD DRUGOG
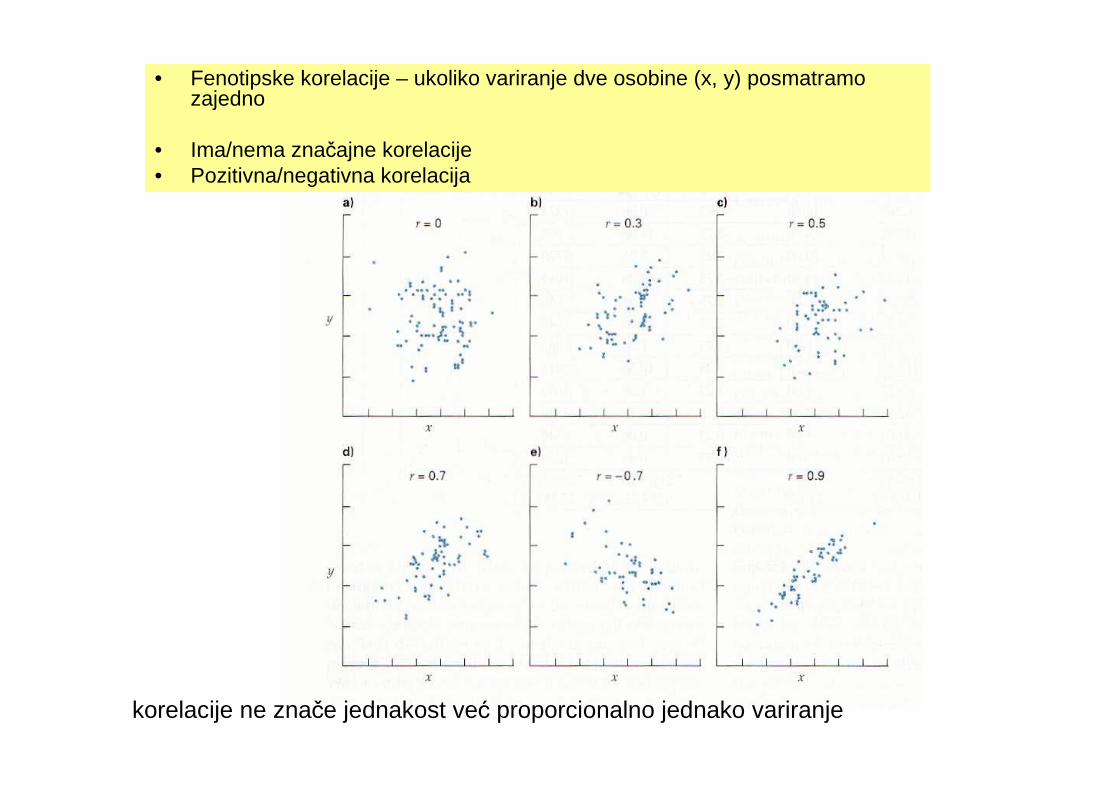
• Fenotipske korelacije – ukoliko variranje dve osobine (x, y) posmatramo zajedno
• Ima/nema značajne korelacije• Pozitivna/negativna korelacija
korelacije ne znače jednakost već proporcionalno jednako variranje

bitno je razlikovati: • gene koji su relevantni za osobinu od genetičkih razlika
relevantnih za razlike u osobini• Ako su geni uključeni u to variranje, onda će u proseku,
biološki srodnici biti sličniji medjusobno nego nesrodne jedinke
• Ta sličnost će se ogledati u pozitivnoj korelaciji izmedju roditelja i potomaka ili izmedju potomaka istih roditelja
• korelacije izmedju srodnika su pokazatelji genetičke varijabilnosti samo ako su sredine srodnika i nesrodnika iste
Fenotipske i geneti čke korelacije medju srodnicima

• Roditelji i potomci imaju polovinu zajedničkih gena, kao i full-sibs, rodjena braća (sestre)
• Njihova genetička korelacija je jednaka verovatnoći da nose isti alel i iznosi 0.5
• Ako su half-sibs, tj. polubraća (polusestre) - isti otac a različite majke onda:
• 0.5 je verovatnoća nasledjivanja identičnog alela od oca, a 0 od majke pa je njihova genetička korelacija prosek toga i iznosi 0.25 (1/4)

• KOLIKO JE VARIJABILNOST NASLEDLJIVA?
• Kako se to može koristiti u oplemenjivanju?

• heritabilnost u širem smislu - doprinos genotipske varijabilnosti (Vg) variranju fenotipova, a nije alelski doprinos varijabilnosti koju nose gameti– ne može se pomoću H2 odrediti genetička
komponenta koja se prenosi kroz generacije putem gameta
• heritabilnosti u užem smislu - uticaj gametske varijabilnosti na Vf koji čini aditivna genetička varijansa (Va), alelski doprinos
HERITABILNOST

• Heritabilnost ne odredjuje koliko je osobina genetička već koliko je njeno variranje genetičko
• Heritabilnost je rezultat interakcije genotipa i sredine; promena uzorka ili promena sredine ne menja način na koji geni determinišu osobinu, ali menjaju udeo Vg ili Vs u Vf te osobine, što utiče na procenu heritabilnosti
• Heritabilnost nije karakteristika jedinke već populacije
• Heritabilnost nije univerzalna za osobinu. Menja se sa promenom sredine
• Visok koeficijent heritabilnosti ne znači da varijabilnost osobine nije pod uticajem sredine

Kako meriti h 2? • Nije nekada moguće identifikovati nijedan lokus koji bi
uticao na kvantitativnu osobinu • Nije uvek moguće meriti genotipove i odrediti genotipske
srednje vrednosti• Samo deo fenotipske varijanse je prenosiv kroz gamete• Mora se odvojiti sredinska od genotipske varijanse
Moguće je – različite homozigotne linije gajiti u istoj sredini pa meriti varijansu
izmedju linija– heteroizigote nastale od ovih linija gajiti u različitim sredinama pa
meriti varijansu unutar linija
• Nekoliko načina za procenu parametara heritabilnosti se uglavnom zasnivaju na fenotipskim sličnostima medju srodnicima

•Regresija osobine jednog roditelja na osobinu potomaka istog pola
h2=2b (kada je regresija na srednju vrednost jednog rodit elja)
Procena koeficijenta heritabilnosti u užem smislu

• Regresija proseka osobine oba roditelja na prosek osobine potomaka
h2=b (kada je regresija na srednju vrednost oba roditelj a)


Tabela 10.10. Koeficijent heritabilnosti izračunat za istu osobinu, na tri načina
metod procene heritabilnost regresija potomaka na roditelje 0,51±0,07
korelacija polusrodnika 0,48±0,11 korelacija potpunih srodnika 0,53±0,07

Interakcije aditivnih vrednosti (Vaa) doprinose kovarijansama svih srodstava
Interakcijske komponente koje uključuju dominansu doprinose kovarijansi samokod FS. Koeficijent kovarijanse za bilo koji tip srodstva je cov = rVa + uVd., odnosno, zbir je korelacija medju
oplemenjivačkim vrednostima i varijanse iz dominanse (Tabela 10.6.).
Tabela 10.7. Koeficijenti komponenti varijanse u kovarijansama kada se uključe dvofaktorijskeinterakcije meñu lokusimaa.
komponente varijanse i koeficijentinjihovog doprinosa
srodstvo Va Vd Vaa Vad Vddpotomstvo-roditelj cov(OP) = 1/2 - 1/4 - -
polusrodnici cov(HS) = 1/4 - 1/16 - -
potpuni srodnici cov(FS) = 1/2 1/4 1/4 1/8 1/16
uopšteno cov = r u r2 ru u2

Fenotipske sli čnosti srodnika – sredinska komponenta
• genetički uzroci nisu jedini uzroci sličnosti meñu srodnicima, jer postoje sredinski uslovi koji utiču da srodnici liče
• članovi porodice koji zive zajedno dele zajedničku sredinu
• ako neki uslovi sredine uzrokuju različitosti meñu nesrodnim jedinkama različitih porodica, oni nisu i uzroci razlika unutar porodice

razlikovati: • gene koji su relevantni za osobinu od genetičkih razlika
relevantnih za razlike u osobini– ako su geni uključeni u to variranje, onda će u proseku, biološki
srodnici biti sličniji medjusobno nego nesrodne jedinke– sličnost će se ogledati u pozitivnoj korelaciji izmedju roditelja i
potomaka ili izmedju potomaka istih roditelja
• ovakve korelacije izmedju srodnika su pokazatelji genetičke varijabilnosti samo ako srodnici ne dele zajedničku sredinu više nego što je nesrodne jedinke dele, odnosno– ako su sredine srodnika i nesrodnika iste, onda je korelacija
medju srodnicima mera genetičke varijabilnosti

Sredinsku varijansu možemo razložiti na komponente zajedničke sredine Vec - eng. common environment -nevezano za srodstvo
• Mnogi činioci Vec variraju (ishrana, klimatski uslovi, antropogeni faktori).
• Vec uzrokuje sličnost unutar grupe i odnosi se na meñugrupnu komponentu varijanse
• Ono što označimo kao Vec, zavisi od grupisanja jedinki kada procenjujemo komponente fenotipske varijabilnosti.
Na primer: ako su grupe potpunih srodnika (FS) onda Vecpredstavlja sredinske uzroke sličnosti meñu FS

Tabela 10.8. Analiza kovarijasne fenotipskih vrednosti u zavisnosti od genetičkih i sredinskih komponenti
srodstvo kovarijansa regresija (b) ili korelacija (t) potomstvo i jedan
roditelj aV2
1
P
a
V
Vb
2
1=
potomstvo i prosek roditelja aV
2
1
P
a
V
Vb =
polusrodnici (HS) aV
4
1
P
a
V
Vt
4
1=
potpuni srodnici (FS) Ecda VVV ++
4
1
2
1
P
Ecda
V
VVVt
++= 4
1
2
1
2
cov
P
OPOPb
σ=

• Ista sredina roditelja i potomaka• Odabiranje fenotipova
• Ako iz nekog uzorka, za roditelje uzmemo fenotipove sa dva ekstrema distribucije:
• 1. F1 ima srednju vrednost bližu odabranim roditeljima –varijabilnost osobine ima aditivnu genetičku komponentu -heritabilnost
• 2. F1 ima srednju vrednosti osobine istu kao ceo uzorak iz kog su uzeti roditelji –varijabilnost osobine nema aditivnu genetičku komponentu
Procena koeficijenta heritabilnosti iz veštačke selekcije

• SD=Xt-Xp
• R=Xp-XF1
• h2=R/S

• kvantifikovanje koeficijenta heritabilnosti: 0 - 1

Odnos selekcionog odgovora R i selekcionog diferencijala S. Svaka ta čka predstavlja par roditelja i njihovo potomstvo, k ao prose čne vrednostiRegresiona linija daje regresiju potomaka na srednj u vrednost roditelja
Početak je prose čna vrednost populacije .

• uspešnost selekcije će zavisiti od prisustva aditivne genetičke varijanse
• u praksi, ako se želi povećanje visine biljke i vrše selektivna ukrštanja, uspešnost selekcije zavisi od R odnosno od Va kao dela h2 .
• što je veća aditivna genetička varijansa, to je viša h2
usled većeg stepena korelacije roditelja i potomaka i biće R veće.
• selekcija će biti potpuno neefikasna ako postoji overdominansa, tj., ako su heterozigoti najbolji za selekciju željene osobine, a oba homozigota lošija.
• tada se ukrštanje vrši medju heterozigotima, a takvim ukrštanjem samo bi se restaurirala početna populaciona varijabilnost.

KOEFICIJENT HERITABILNOSTI ZA RAZLIČITE OSOBINE KOKOŠI

Dva tipa selekcije sa odsecanjem distribucije kontinuirano varirajućih osobina: a) linija označava konstantnu tačku odsecanja. b) tačka odsecanja je proporcionalna. U prvom slučaju intenzitet selekcije opada vremenom kako sve veći deo populacije prelazi tačku odsecanja, dok u drugom ostaje konstantan intenzitet selekcije, ali se tačka odsecanja pomera sa distribucijom.

• Direkciona selekcija• Bidirekciona selekcija

Bitno za planiranje programa veštačke selekcije• kada isti geni utiču na variranje više osobina
(plejotropija) - genetičke korelacije
• pozitivne ili negativne korelacije –može postojati antagonistička plejotropija
• korelisan ogovor na selekciju - je kada se odabiraju osobine koje su genetički korelisane

• ako se utvrdi da je h2 niska, a i H2 takodje niska, znači da je visoka sredinska varijansa u odnosu na genetičku, te se primenjuje promena sredine i Ve snižava.
• ako je h2 veoma niska, a H2 visoka, mora se u oplemenjivanju koristiti neki drugi metod pored selekcije, jer uzrok variranja nije sredinskavarijansa, aditivna je već mala, pa se moraju primenjivati posebne šeme ukrštanja kojima se iskoristi neaditivna varijansa (Vd, Vi).
• Najčešći metod je inbriding – ukrštanje u srodstvu

INBRIDING• Kada je niska Va, niska h2, selekcija ne daje odgovor• Primenjuje se inbriding kao metod dobijanja željenih
genotipova, linija koji daju željene osobine• Najveći broj inbridingovanih linija se dobija
samooplodnjom, ukrštanjem u najvećem stepenu srodstva


Inbridingom se postiže se homozigotizacijagenoma
uvećava se verovatnoća da su aleli identični po poreklu

• Verovatnoća da su dva alela na jednom lokusu kod odreñene jedinke indentična po poreklu – Koeficijent inbridinga (F)
Tabela 6.7. Koeficijent inbridinga (F) u potomstvu različitih srodnika tip parenja F samooplodnja (ili, majka - sin) potpuni srodnici (brat - sestra) ujak - sestričina,tetka - sestrić, dvostruki prvi roñaci prvostepeni roñaci (brat - sestra od stričeva) tetka i polubratanac (sin prvostepenog roñaka) drugostepeni roñaci tetka i sin drugostepenog roñaka trećestepeni roñaci
1/2 1/4 1/8 1/16 1/32 1/64 1/128 1/256


Genotipske učestalosti
Generacije A1A1 A1A2 A2A2 F q
0 1/4 1/2 1/4 0 0,51 3/8 1/4 3/8 1/2 0,52 7/16 1/8 7/16 3/4 0,53 15/32 1/16 15/32 7/8 0,54 31/64 1/32 31/64 15/16 0,5t [1 - (1/2)t + 1]/ 2 (1/2)t [1 - (1/2)t+ 1]/ 2 1 - (1/2)t 0,5
∞ 1/2 0 1/2 1 0,5
Smanjenje heterozigotnosti pri sistematskoj samooplodnji,sa početnim ravnotežnim stanjem u populaciji (p = q = 0,5)

Učestalosti alela u populaciji se ne menjaju pri inbridingu.....
• Ako je učestalost alela A1 u roditeljskoj generaciji = p, a alela A2 = qonda je učestalost homozigota A1A1 jednaka p2 = P a heterozigota A1A2 = 2pq = H
• P’ i H’ su učestalosti genotipova A1A1 i A1A2 u F1 generaciji: • P’ = P + 1/2(1/2)H i H’ = (1/2)H
• U generaciji potomaka učestalost alela A1 je p’ = P’ + (1/2)H’, onda je
p’= P + (1/4)H + (1/4)H= p2 + (1/2)2pq= p2 + pq= P + (2/4)H= p2 + p(1 - p)= p

2t broj predaka svakog diploida, t generacija unazad
U slučajnom uzorku 2N gameta iz genofonda t-1 generacijeverovatnoća da će 2 identična alela da se zajedno nañu je
1/(2N), a da će dva potpuno nesrodna, 1-1/(2N)

Alelske učestalosti u populaciji se ne menjaju bez obzira na F....
• H = 2pq - 2Fpq H/2 = pq - Fpq (H/2)pq = 1 - F F = 1 - (H/2pq)
• p’ = P + H/2= (p2 + Fpq) + (1/2)(2pq - 2Fpq)= p2 + pq = p2 + p(1 - p) (q = 1 - p)= p
P = p2 + FpqH = 2pq - 2FpqQ = q2 + Fpq
F se može izraziti preko učestalosti heterozigota

HD = HO - HOFHOF = HO - HD
F = (HO - HD) / HO
F je u funkciji odnosa dobijene i očekivane (HW) heterozigotnosti
Heterozigotnosti po generaciji pri parenju po principu slučajnosti populaciji ograničene veličine dat je izrazom
Ht = H0 (1 - 1/2N)t ≅ H0 e -t / 2N

Inbriding depresija –- usled homozigotizacije štetnih recesivnih alela- usled asocijativne dominanse –heterozigoti na nekim lokusima su bolji nego oba homozigota
Efekti inbridinga
Fenotipska uniformnost - samo ukolikosu Vg i Vs aditivni i nezavisni.
Genetička uniformnost

HETEROZIS
• Ako se inbredovane linije ukrštaju daju hibride• efekat asocijativne dominanse - daje najbolje heterozigote
• Inbridingovana linija 1: AA BB cc dd EE• Inbridingovana linija 2: aa bb CC DD ee• Hibrid: Aa Bb Cc Dd Ee
• Medjutim, heterozigoti nastali ukrštanjem inbridingovanih, homozigotnih linija mogu biti prosečni, bolji ili lošiji od obe roditeljske linije (na primer po prinosu)
• pozitivan heterozis (hibridna bujnost, vigor)• negativan heterozis (autbriding depresija)
• zato se odabiraju najbolje kombinacije

• Pri ukrštanju jedinki koje potiču iz genetički udaljenih i različitih grupa dolazi do fenomena na nivou fenotipa poznatog pod nazivom heterozis (hibridni vigor, hibridna snaga).
• On se ogleda u tome da hibridno potomstvo (F1 generacija) od roditelja koji pripadaju različitim inbredovanim linijama ili sortama (rasama) pokazuje značajne promene odreñenih fenotipskih osobina (pa i onih koji su bitne za preživljavanje i razmnožavanje jedinki) u odnosu na roditelje.
• Ako F1 generacija prevazilazi u posmatranoj fenotipskoj vrednosti roditelja sa višom vrednošću odgovarajućeg karaktera, govori se o pozitivnom heterozisu, a ako je hibridna generacija "lošija" od "lošijeg" roditelja govori se o negativnom heterozisu.

Odredjivanje genotipa kod kvantitativnih osobina
• mapiranje širom genoma• odredjivanje kandidat gena• kombinacija mapiranja i kandidat gena
Nisu novi pristupi ali primena je nova

mapiranje preko genomske vezanosti
• Genetički markeri pokazuju asocijaciju sa fenotipskim variranjem kroz vezanost, jer su na istom hromozmu blisko locirani, a nisu direktni uzrok variranja fenotipa
• jačina vezanosti – blizina lokusa je obično obrnuto proporcionalna rekombinacionoj učestalosti
• Genetička “rezolucija” je ograničena brojem rekombinacionih dogadjaja u jednoj generaciji uzorka
• markeri identifikuju delove genoma koji utiču na fenotip

• Quantitative Trait Loci (QTL)- pozicije regiona na hromozomu koje odredjuju kvantitativne osobine
• QTL procenjuju broj i relativni značaj gena koji utiču na kvantitativne osobine
• Za odredjivanje QTL potrebni su podaci iz rodoslova(čovek) ili ukrštanja medju linijama (biljke i životinje) o dobijenim rekombinantima
• Treba dovoljno polimorfnih markera rasutih po genomu, da ma koji QT lokus nije udaljen više od 10 cM od marker lokusa

Ukrštaju se 2 linije koje se značajno razlikuju po fenotipu i marker genima
Ako je QTL blisko vezanza marker gen, različitimarker genotipovi i QTLovi će se nasledjivati zajedno,a različiti marker genotipovi u F2će imati različite srednje fenotipove
Ako je marker blizu QTL, dobiće se razlikau srednjim vrednostima fenotipova medjumarker genotipovima
Prosečan fenotipski efekat klase M =5(0,9)+0(0,1) = 4,5Prosečan fenotipski efekat klase m =5(0,1)+0(0,9) = 0,5Razlika izmeñu gameta koji nose M i gameta koji nose m = 4,5-0,5 = 4Razlika izmeñu proseka F2 M/M homozigota i proseka F2 m/m homozigota = 8(8% je razlija izmedju početnih linija, a QTL nosi 10% različitosti jer je RF =0,10


