general histology (97-2003 version)
TRANSCRIPT

GABRIELA MUŢIU
CIURSAŞ ADINA NICOLETA
GENERAL HISTOLOGY
EDITURA UNIVERSITĂŢII DIN ORADEA
2010

Table of Contents
Chapters Page Number
1. Epithelia and Glands 2-20
2. Connective Tissue 21-36
3. Cartilage 37-45
4. Bone 46-56
5. Muscle 57-70
6. Vascular System 71-83
7. Blood 84-98
8. Lymphoid Tissues 99-113
9. Nervous Tissue 114-128
1

EPITHELIA
Epithelia are tissues consisting of closely apposed cells without intervening intercellular substances. Epithelia are avascular, but all epithelia "grow" on an underlying layer of vascular connective tissue. The connective tissue and the epithelium are separated by a basement membrane. Epithelium covers all free surfaces of the body. Epithelium also lines the large internal body cavities, where it is termed mesothelium. Furthermore, the internal surfaces of blood and lymph vessels are lined by epithelium, here called endothelium.
Epithelia are classified on the basis of the number of cell layers and the shape of the cells in the surface layer.
If there is only one layer of cells in the epithelium, it is designated simple. If there are two or more layers of cells, it is termed stratified. Cells in the surface layer are, as a rule, described according to their height as
squamous (scale- or plate-like), cuboidal or columnar.
Simple Epithelia
Simple squamous epithelium
This type is composed of a single layer of flattened, scale- or plate-like cells. It is quite common in the body. The large body cavities and heart, blood vessels and lymph vessels are typically lined by a simple squamous epithelium. The nuclei of the epithelial cells are often flattened or ovoid, i.e. egg-shaped, and they are located close to the centre of the cells.
Simple cuboidal epithelium
2

Cells appear cuboidal in sections perpendicular to the surface of the epithelium. Viewed from the surface of the epithelium they look rather like small polygons. Simple cuboidal epithelium occurs in small excretory ducts of many glands, the follicles of the thyroid gland, the tubules of the kidney and on the surface of the ovaries.
Simple columnar epithelium
The cells forming a simple columnar epithelium are taller than they are wide. The nuclei of cells within the epithelium are usually located at the same height within the cells - often close to the base of the cells. An example is the simple columnar epithelium which lines the internal surface of the gastrointestinal tract (GIT) from the cardia of the stomach to the rectum.
Identifying Epithelia
The outlines of individual epithelial cells are not always visible, and it may be difficult to identify the shape of the cells.
It is often helpful to look at the shape, location and spacing of the nuclei in the epithelium, which together will allow a very good guess at the shape of the cells forming the epithelium.
How many cell layers seem to be visible in a section depends very much on the angle between the plane of the section plane the surface of the epithelium.
Oblique sections of epithelium will be visible in almost all slides of organs in which epithelium lines a surface with a very irregular profile. A single surface is usually not lined by several types of epithelia.
The number of epithelial cell layers will usually be the smallest number of layers visible anywhere along the surface lined by the epithelium.
3

Suitable Slidessimple squamous epithelium: any section containing blood vessels, sections of organs which include an outer lining (or serosa) of the organs, or sections of kidney (parietal blades of Bowman's capsules) - H&E, trichrome simple cuboidal epithelium: sections of ovaries (epithelium lining the surface), thyroid gland (follicles), kidney (tubules) or large glands (e.g. parotid gland) with well preserved small ductssimple columnar epithelium: sections of the small intestine (duodenum, jejunum or ileum), uterus (uterine glands), liver (large bile ducts) or gall bladder - H&E, trichrome
Sublingual Gland, Human, H&EBlood vessels are probably present in all sections you will ever see. With very few exceptions, they are lined by a simple squamous epithelium. The individual epithelial cells are extremely flattened and form a much larger part of the surface than individual cells in cuboidal or columnar epithelia. The nuclei of the squamous epithelial cells are also flattened and often stain darkly. Not every epithelial cell nucleus will be included in the plane of the section, and if the vessel is very small (e.g. a capillary), there may not be any visible nuclei in the epithelial lining.Capillaries and other small vessels are easily deformed during tissue processing, and the epithelium of larger vessels may be damaged or look corrugated. It may therefore take a little more patience than you expect to find a "good" simple squamous epithelium.Draw a small vessel with its epithelial lining, label the features visible in your drawing and include a suitable scale.
4

Duodenum, Rat, H&E and Ileum, Human - H&EThe small intestines are lined by a simple columnar epithelium. Most of the epithelial cells (enterocytes) are involved in the absorption of components of the digested food in the lumen of the intestines. Complex folds of the intestinal lining increase the surface area available for absorption. The plane of the section will therefore often pass at an oblique angle through the epithelium. The epithelium may look stratified where this happens. Scan along the epithelium until you find a spot where it is cut perpendicular to its surface, i.e. where it looks like a simple columnar epithelium. Mucus producing goblet cells are a second cell type of this epithelium. Mucus stains only weakly or not at all in H&E stained sections. Round, light "hollows" in the epithelium represent the apical cytoplasm of the goblet cells, which is filled with mucin-containing secretory vesicles.Microvilli extend from the apical surface of epithelial cells into the intestinal lumen. They increase surface area by a factor of ~20 and thereby facilitate absorption. Together, the microvilli are visible as a light red band along the apical limit of the epithelium, i.e. the side of the epithelium facing the lumen of the intestine. This band is call the brush border.Draw and label the epithelium. Include goblet cells in your drawing.
5

Stratified Epithelia
Stratified squamous epithelium
Stratified squamous epithelia vary in thickness depending on the number of cell layers present. The deepest cells, which are in contact with the basement membrane, are cuboidal or columnar in shape. This layer is usually named the basal cell layer, and the cells are called basal cells. Basal cells are mitotically active and replace the cells of the epithelium which are lost by "wear and tear". The basal cell layer is followed by layers of cells with polyhedral outlines. Close to the surface of the epithelium, cells become more flattened. At the surface of the epithelium, cells appear like flat scales - similar to the epithelial cells of simple squamous epithelia.Remember that it is the shape of the cell which form the surface of the epithelium which gives the name to the epithelium.
Stratified cuboidal and columnar epithelia
are not common. A two-layered cuboidal epithelium is, for example, seen in the ducts of the sweat glands. Stratified columnar epithelia are found in the excretory ducts of the mammary gland and the main excretory duct of the large salivary glands.
Suitable Slidesstratified squamous epithelium: sections of the oesophagus, tongue or vagina - H&E, van Gieson, trichrome. stratified cuboidal epithelium: skin (excretory ducts of sweat glands) - H&Estratified columnar epithelium: sections of the parotid gland or mammary gland - H&E
6

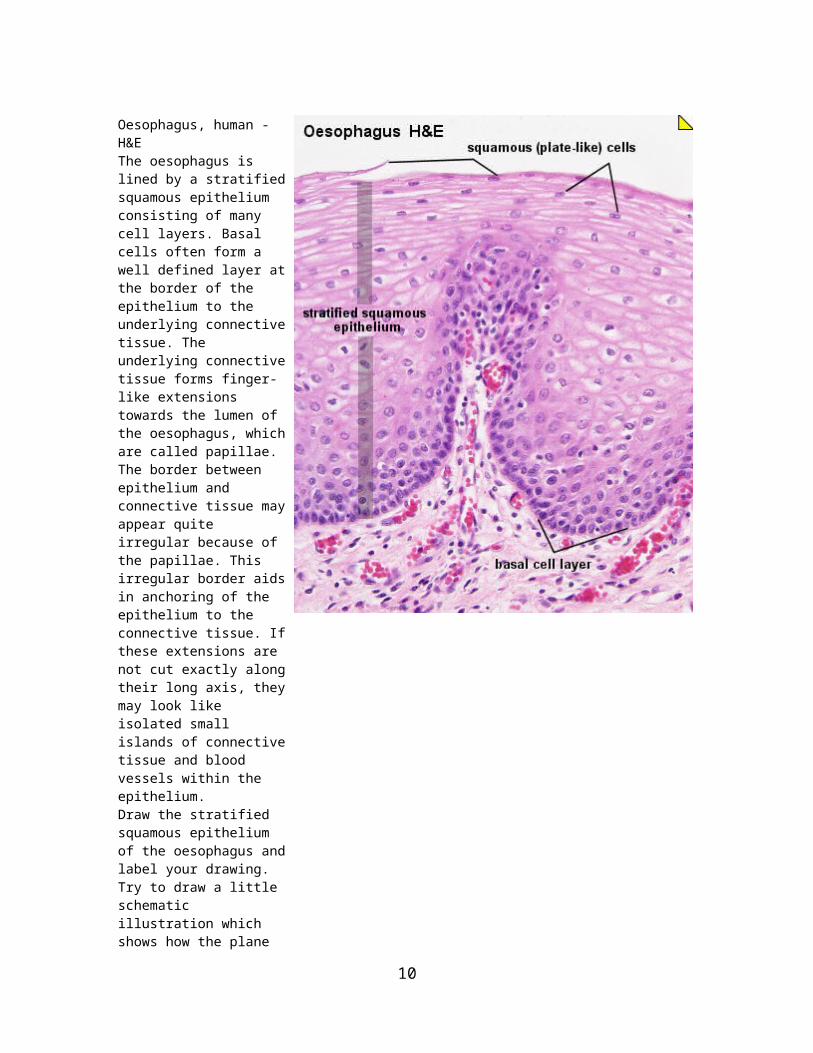
Oesophagus, human - H&EThe oesophagus is lined by a stratified squamous epithelium consisting of many cell layers. Basal cells often form a well defined layer at the border of the epithelium to the underlying connective tissue. The underlying connective tissue forms finger-like extensions towards the lumen of the oesophagus, which are called papillae. The border between epithelium and connective tissue may appear quite irregular because of the papillae. This irregular border aids in anchoring of the epithelium to the connective tissue. If these extensions are not cut exactly along their long axis, they may look like isolated small islands of connective tissue and blood vessels within the epithelium.Draw the stratified squamous epithelium of the oesophagus and label your drawing. Try to draw a little schematic illustration which shows how the plane of section would effect the appearance of the connective tissue extensions.
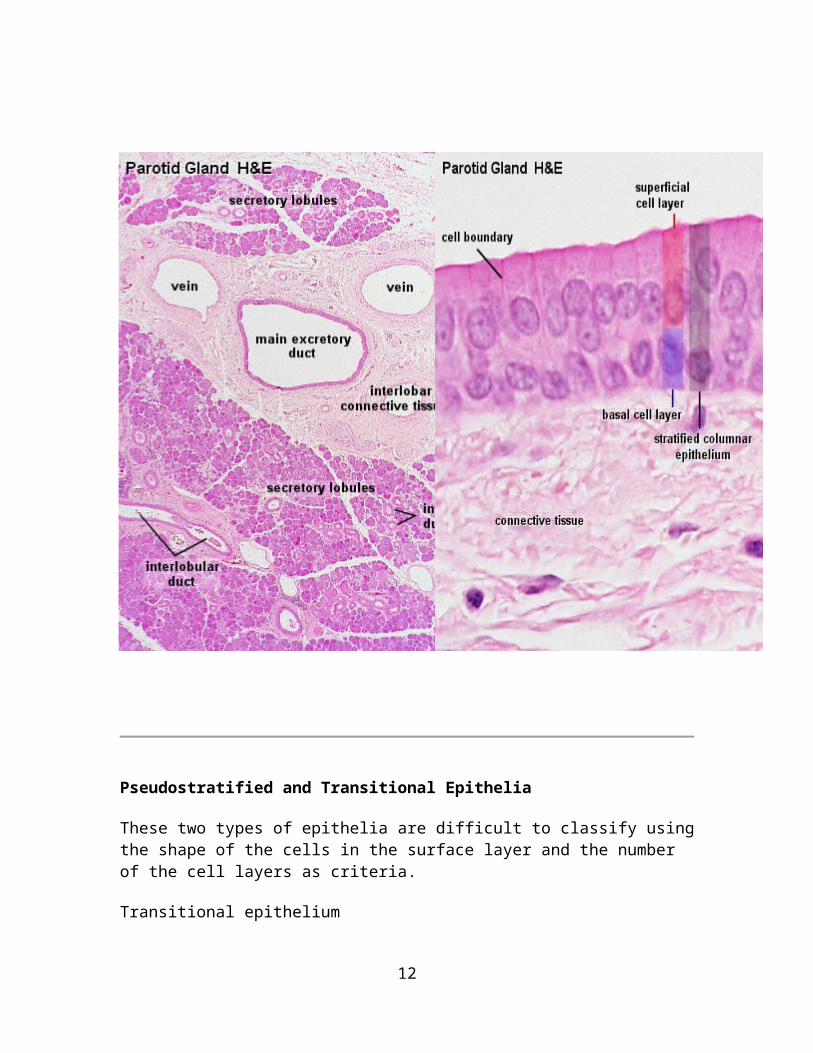
Parotid Gland, Human - H&EStratified columnar epithelia are found in the largest excretory ducts of some glands. The parotid gland, a large salivary gland, is one of them. Several epithelial types are found in the duct system of the parotid. The smallest ducts, which are embedded in the secretory tissue (intralobular ducts), are lined by cuboidal or columnar epithelia. Small ducts, which are embedded in connective tissue located between areas of secretory tissue (interlobular ducts), are lined by columnar or pseudostratified epithelia. These ducts finally coalesce to form the main excretory duct of the parotid which is lined by a stratified columnar epithelium.Draw the stratified columnar epithelium seen in the largest ducts and label your drawing.
7

Pseudostratified and Transitional Epithelia
These two types of epithelia are difficult to classify using the shape of the cells in the surface layer and the number of the cell layers as criteria.
Transitional epithelium
Transitional epithelium is found exclusively in the excretory urinary passages (the renal calyces and pelvis, the ureter, the urinary bladder, and part of the urethra).
8

The shape of the cells in the surface layer of a transitional epithelium varies with the degree of distension of the organs whose lumen is lined by this type of epithelium. In the 'relaxed' state of the epithelium, it seems to be formed by many cell layers. The most basal cells have a cuboidal or columnar shape. There are several layers of polyhedral cells, and, finally, a layer of superficial cells, which have a convex, dome-shaped luminal surface. In the distended state of the epithelium only one or two layers of cuboidal cells are followed by a superficial layer of large, low cuboidal or squamous cells. In the distended state the epithelium will resemble a stratified squamous epithelium.
Pseudostratified columnar epithelium
All cells of this type of epithelium are in contact with the basement membrane, but not all of them reach the surface of the epithelium. Nuclei of the epithelial cells are typically located in the widest part of the cell. Consequently, the nuclei of cells which do or do not reach the surface of the epithelium are often located at different heights within the epithelium and give the epithelium a stratified appearance. The epithelium will look stratified but it is not - hence its name "pseudostratified". Pseudostratified columnar epithelia are found in the excretory ducts of many glands.
Suitable Slidestransitional epithelium: sections of ureter or bladder - H&Epseudostratified epithelium: sections of the trachea - H&E
9
Bladder, Monkey - H&EAt a first glance a transitional epithelium looks like a stratified cuboidal epithelium. Several rows of nuclei appear to be topped by a layer of dome-shaped cells which bulge into the lumen of the ureter. The shape of the surface cells and the number of rows change if the bladder is distended. The number of rows decreases. This decrease should tell us that many of the nuclei located in different layers of the epithelium belong to cells which are all in contact with the basement membrane. With distension, the shape of the cells in the surface layer will become squamous.Draw the epithelium and label the features you can see. Add a simple schematic drawing of how you expect the epithelium to look like if the ureter is distended.
It has not yet been resolved if all the epithelial cells are in contact with the basement membrane. Some texts consider transitional epithelium as a specialised stratified epithelium while others group it with pseudostratified epithelia. Maybe it is best to also consider it 'transitional' in this regard.
10

Trachea, Human - H&EAt least two, sometimes three rows of nuclei are seen in the pseudostratified columnar epithelium lining the trachea. The nuclei belong to cells which are all in contact with the basement membrane. The epithelial lining of the trachea is also one of the few examples of a basement membrane clearly visible in H&E stained sections. Epithelial cells can be ciliated or they can be goblet cells (unicellular exocrine glands). Basal cell regenerate other cell types of the epithelium. Capillaries and small vessels are visible in the connective tissue beneath the epithelium.A ciliated pseudostratified columnar epithelium with goblet cells is a characteristic feature of parts of the respiratory system, where it is call respiratory epithelium. It contains several cell types in addition to ciliated, goblet and basal cells.Draw the epithelium at high magnification and label your drawing.
Special Cytological Features of Epithelia
Basement membrane or basal lamina Epithelia are separated from the underlying connective tissue by an extracellular supporting layer called the basement membrane. The basement membrane is composed of two sublayers. The basal lamina (about 80 nm thick) consists of fine protein filaments embedded in an amorphous matrix. Membrane proteins of the epithelial cells are anchored in the basal lamina, which is also produced by the epithelial cells. The major components of the basal lamina are two glycoproteins - laminin and (usually type IV) collagen. The reticular lamina consists of reticular fibres embedded in ground substance. The fibres of the reticular lamina connect the basal lamina with the underlying conective tissue. The components of the reticular lamina are synthesised by cells of the connective tissue underlying the epithelium.In addition to its function as support of the epithelium, the basal lamina acts as a selectively permeable filter between epithelium and connective tissue.
11
Unless special stains are used, the basement membrane is rarely visible using light microscopy. You can read more about reticular fibres and ground substance on the Connective Tissues page.
Specialisations of the apical surfaceMicrovilli and stereocilia are finger- or thread-shaped extensions of the epithelial cells. Their main function is to increase the surface area of epithelial cells. They are typically found in epithelia active in absorption. Microvilli contain actin filaments, which are in contact with the terminal web of the cell. The only difference between microvilli and stereocilia is their length. Microvilli are much shorter than stereocilia. Stereocilia are, despite their name ("cilia"), not actively moving structures.
Using light microscopy, stereocilia are difficult to discern from cilia.
Specialisations of the lateral and basal surfacesConnective tissue is responsible for the structural integrity of most organs. As mentioned above, it is absent from epithelia. Instead, tissue integrity as well as the barrier function of epithelia is taken care of by extensive cell-to-cell contacts between epithelial cells. These functions are mediated by several specialisations in the lateral and basal parts of the cell membranes of the epithelial cells.
Desmosomes
are specialisations of the lateral cell membranes which mediate cell adhesion. Proteins inserted into the cell membrane of the adjacent cells form a protein-'zipper' linking the cells. Fibers of the cytoskeleton attach to the cytoplasmic side of the desmosome to stabilise the area of contact. Hemi-desmosomes mediate the attachment of the epithelial cells to the basement membrane.A group of glycoproteins (cadherins) inserted into the opposing plasma membranes mediate cell-to-cell adhesion at desmosomes and also at the adhesion zones or patches mentioned below. Integrins, another group of proteins, allow the cell to attach to the matrix proteins of the basal lamina.
Intermediate junctions (zonula adherens)
are structurally not as well-characterised as desmosomes. An intermediate junction typically appears as a close and consistent apposition (15-20 nanometers) of the cell membranes near the apical cell surface. Intermediate junctions surround the entire cell. Again, fibres of the cytoskeleton insert into the cytoplasmic side of this membrane specialisation. Patches of adhesion resemble intermediate junctions structurally, but form more localized, patch- or strip-like contacts between neighbouring cells. They are found scattered over the lateral surfaces of the epithelial cell.
12

The above mentioned membrane specialisations mediate cell-adhesion but are less well suited to support one of the essential functions of epithelia - the isolation of the interior of the body from the outside world. A tight junction (zonula occludens) between epithelial cells mediates this aspect of epithelial function.
Proteins inserted into the cell membranes of adjacent cells 'stitch' the membranes of the cells together and provide an effective barrier to the diffusion of substances from the outside of the epithelium (called luminal side if the epithelium covers the surface of a tubular structure). Several "rows of stitches" may be found. Their number depends on the demand to reduce diffusion across the epithelium. Each of these rows reduces diffusion by about a factor 10 of what it was 'before'.
13

GLANDS
are cells or aggregations of cells whose function is secretion.
Exocrine glands release the secretory product via a system of ducts that opens upon one of the surfaces of the body which are in contact with the external world (skin, gastrointestinal tract etc.).
Endocrine glands release their secretory product (typically hormones) into the spaces between the secretory cells (extracellular space) from which it enters the bloodstream.
Both endocrine and exocrine glands are developmentally derived from epithelia, which form a down-growth into the underlying connective tissue. The cells forming this down-growth then develop the special characteristics of the mature gland. Exocrine glands maintain the connection with the surface epithelium, whereas the connection is lost by endocrine glands.
Classification of Exocrine Glands
Exocrine glands may be classified according to cell number, and/or the shape and branching pattern of their secretory portions and ducts.
Unicellular Glands
consist of a single secretory cell. In mammals the only example of unicellular exocrine glands are goblet cells, which occur in the epithelium of many mucous membranes. Goblet cells secrete the glycoprotein mucin, which by the uptake of water is converted into a slimy substance, mucus.
14

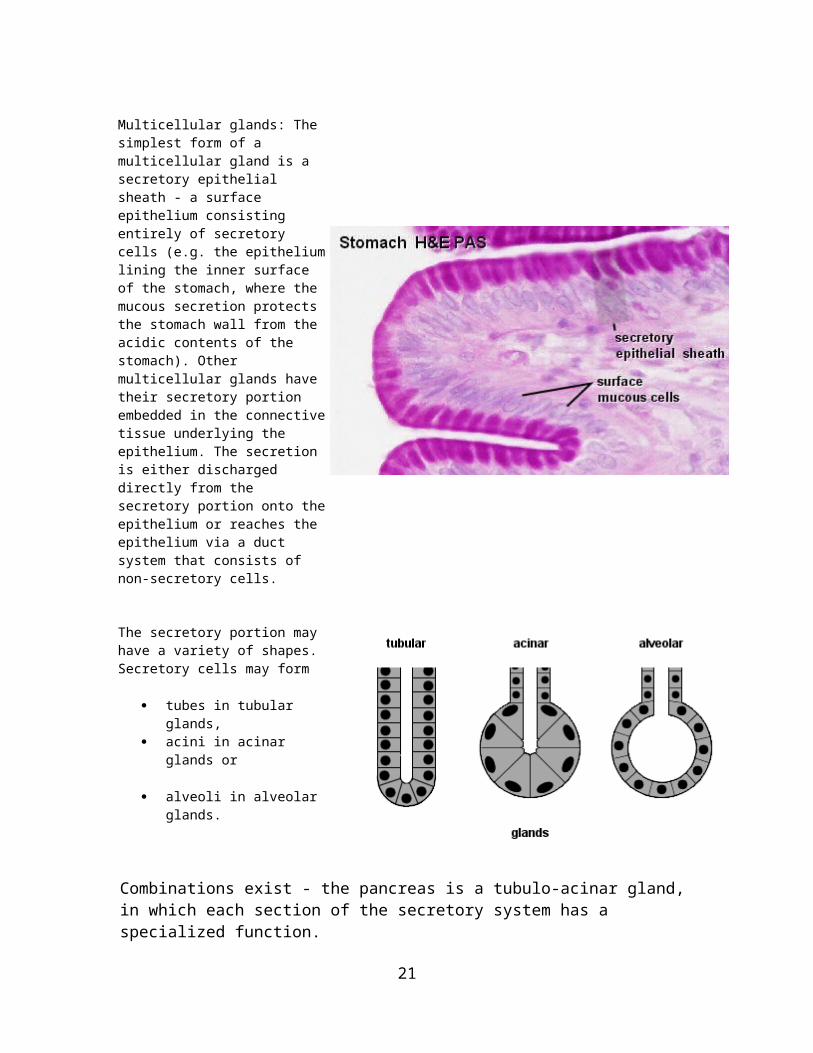
Multicellular glands: The simplest form of a multicellular gland is a secretory epithelial sheath - a surface epithelium consisting entirely of secretory cells (e.g. the epithelium lining the inner surface of the stomach, where the mucous secretion protects the stomach wall from the acidic contents of the stomach). Other multicellular glands have their secretory portion embedded in the connective tissue underlying the epithelium. The secretion is either discharged directly from the secretory portion onto the epithelium or reaches the epithelium via a duct system that consists of non-secretory cells.
The secretory portion may have a variety of shapes. Secretory cells may form
tubes in tubular glands, acini in acinar glands or
alveoli in alveolar glands.
Combinations exist - the pancreas is a tubulo-acinar gland, in which each section of the secretory system has a specialized function.The precursors of digestive enzymes are produced by the acinar cells. Tubular cells secrete the alkaline bicarbonate solution which eventually neutralizes the acidic contents of the stomach that are released into the duodenum.
Multicellular glands with an unbranched excretory duct are called simple. We talk about a compound gland when the excretory duct is branched. Finally, the part of the gland consisting of secretory cells is branched in a (surprise!) branched gland.
The classification scheme may appear somewhat elaborate - but there are many exocrine glands around. All of them can be identified and described by this scheme, and some ideas about their function can be derived from this description.
15

Suitable Slidesunicellular exocrine glands (goblet cells): sections of intestines (duodenum, jejunum, ileum or colon) or trachea - H&Esecretory epithelial sheath: stomach - H&E straight tubular glands: sections of stomach (principal glands) or colon (intestinal glands) - van Gieson, H&Ecoiled tubular glands: sections of skin (sweat glands) - see lab section on the Integumentary System page.
Colon, Human - van GiesonStraight tubular glands extend from the surface of the colon into the underlying connective tissue. Although they are present throughout the intestines they are largest in the colon and, because of the smooth inner surface of the colon, they often show in good longitudinal or transverse sections. The lumen of the glands is narrow and surrounded by secretory cells of several types, which include goblet cells. The connective tissue beneath the epithelium and surrounding the glands in the colon contains more cells than the connective tissue beneath other epithelia that were considered on this page. This is a characteristic feature of the epithelia in the digestive system. Glands cut at slightly oblique angles will connect to the lumen outside of the plane of the section.If possible, draw both longitudinally and transversely sections intestinal glands. Include part of the surrounding connective tissue and surface epithelium.
16

Secretory Mechanisms
The secretory cells can release their secretory products by one of three mechanisms.
Merocrine secretioncorresponds to the process of exocytosis. Vesicles open onto the surface of the cell, and the secretory product is discharged from the cell without any further loss of cell substance.
Apocrine secretiondesignates a mechanism in which part of the apical cytoplasm of the cells is lost together with the secretory product. The continuity of the plasma membrane is restored by the fusion of the broken edges of the membrane, and the cell is able to accumulate the secretory product anew. This mechanism is used by apocrine sweat glands, the mammary glands and the prostate.
Holocrinesecretion designates the breakdown and discharge of the entire secretory cell. It is only seen in the sebaceous glands of the skin.
There are two additional mechanisms by which secretory cells can release their products. Lipid soluble substances may diffuse out of the secretory cell (e.g. steroid hormone-producing endocrine cells). Transporters (membrane proteins) may actively move the secretory product across the plasma membrane (e.g. the acid producing parietal cells of the gastric glands). These secretory mechanisms may not involve any light microscopically visible specialisations of the cell.
Histological Structure of Large Exocrine Glands
The relationship between the secretory tissue (parenchyma) of glands and the supporting connective tissue is similar in most larger glands. Externally the entire gland is surrounded by a layer of dense connective tissue, the capsule. Connective tissue sheets (septa) extend from the capsule into the secretory tissue and subdivide the gland into a
17

number of lobes. Thinner connective tissue septa subdivide the lobes into a number of lobules. Reticular connective tissue (hardly visible in H&E stained sections) surrounds and supports the secretory units of the glands (alveoli, acini etc.) and the initial parts of the excretory ducts if present.
Blood and lymph vessels as well as nerves penetrate the capsule and form a delicate network between the secretory units and the initial parts of the duct system.
The main excretory duct conveys the secretory product to one of the external surfaces of the body. Other parts of the duct system are named according to their relation to the lobes and lobules of the gland.
Lobar ducts are are large branches of the main duct which extend to the lobes of the gland. They may be called
Interlobar ducts if they are found in the connective tissue surrounding the lobes. Interlobar ducts branch and give rise to
Interlobular ducts, which are found in the connective tissue surrounding the individual lobules of the gland. Branches of the interlobular ducts enter the lobules and are now called
Intralobular ducts. The terminal branches of the duct system, which connect intralobular ducts with the secretory units of the gland, are called
Intercalated ducts.
The appearance of the different portions of the duct system is quite variable from gland to gland and may allow the identification of the gland. Quite often, the appearance of parts of the duct system also permits some deductions about their functions.
Note that lobes and lobules are defined by their relationship to each other. Many small lobules may form one large lobe. Neither size nor the spatial relationship between different parts of the tissue can be unequivocally determined in a single, two-dimensional section of the tissue. Lobes and interlobar ducts may therefore be difficult to distinguish from lobules and interlobular ducts.
Suitable Slidesalveolar gland: lactating mammary gland H&E - see lab section on the Female Reproductive System page.serous and mucous acinar glands: sections of parotid gland, sublingual gland or tongue (lingual salivary glands) - H&E
18

Parotid Gland, Human - H&EFind an area of secretory tissue at low magnification, and scan over this area at high magnification. Within the lobules and between the acini of the parotid you can find two types of ducts. Since they are both located within the lobules they are both intralobular ducts. Striated ducts are lined by a simple tall columnar epithelium. Intercalated ducts are lined by a simple cuboidal epithelium and connect individual acini to the striated ducts.Try to capture the features of the acini, intercalated and striated ducts in one compound drawing which shows how they connect to each other. Label your drawing.
Parotid Gland, Human and Sublingual Gland, Human - H&EMany secretory cells and the secretory structures formed by them belong to one of two morphologically distinct forms: serous or mucous. Serous secretions have a low viscosity, i.e. they are rather "watery". Mucous secretions have a high viscosity, i.e. they are rather "slimy". The apical cytoplasm of the cells forming serous acini is usually well-stained. Secretory vesicles are visible in the apical cytoplasm in well-preserve tissue. The nuclei are round or slightly ovoid and located in the basal cytoplasm of the cells. The bluish color of the basal cytoplasm reflects the presence of large amounts of rough endoplasmatic reticulum.The contents of the secretory vesicles in the apical cytoplasm of cells forming mucous acini are only weakly stained. These empty-looking vesicles give the apical cytoplasm of mucus-producing cells a distinct "foamy" or "frothy" appearance. The nuclei of mucous cells appear darker and smaller than the nuclei of serous cells. They also seem to be "pressed" against the basal limit of the cells and may look flattened with an angular ("edgy") outline. Glands containing mucous acini (e.g. the sublingual glands) are called mucous glands. Glands containing serous acini (e.g. the parotid glands) are called serous glands. If both types of acini are present the gland is muco-serous.Identify and draw serous and mucous acini at high magnification. Label your drawing. Make sure that the features which characterise serous and mucous acini are visible in your drawing - if necessary use a little artistic freedom.
19

20
CONNECTIVE TISSUE
Connective tissue consists of cells separated by varying amounts of extracellular substance. In connective tissues cells typically account for only a small fraction of the tissue volume. The extracellular substance consists of fibres which are embedded in ground substance containing tissue fluid. Fibres in connective tissue can be divided into three types: collagen fibres, reticular fibres and elastic fibres.
Extracellular Substance
Collagen fibres
Collagen fibres are the dominant fibre type in most connective tissues. The primary function of collagen fibres is to add strength to the connective tissue.
The thickness of the fibres varies from ~ 1 to 10 µm. Longitudinal striations may be visible in thicker fibres. These striations reveal that the fibres are composed of thinner collagen fibrils (0.2 to 0.5 µm in diameter). Each of these fibrils is composed of microfibrils, which are only visible using electron microscopy.
Microfibrils are assemblies of tropocollagen, which, in turn, is an spiral-like assembly of three collagen molecules (triple helix). The organisation of the tropocollagen within the microfibrils is highly regular. A small gap (60 nm wide) is found between the subsequent tropocollagens which form the microfibrils. Staining solutions used in electron microscopy tend to fill in these gaps, and the alignment of the gaps gives the microfibrils a cross-striated appearance (with 68 nm intervals) in EM images.
Coarse collagen fibres are formed by type I tropocollagen.
There are many different tropocollagen types around (currently named type I to XXI). These types differ in their content of the amino acids hydroxyproline and hydroxylysine. They also differ in the amount of carbohydrates attached to the collagen molecules. The different types of tropocollagen give the fibres the structural and functional features which are appropriate for the organ in which the fibres are found. Types I, II and III are the major fibre-forming tropocollagens. Tropocollagen type IV is an important structural component of the basal lamina.
A tensile force of several hundred kg/cm2 is necessary to tear human collagen fibres. The fibres stretch by only 15-20%.
Reticular fibres
21

Reticular fibres are very delicate and form fine networks instead of thick bundles. They are usually not visible in histological sections but can be demonstrated by using special stains. For example, in silver stained sections reticular fibres look like fine, black threads - coarse collagen fibres appear reddish brown in the same type of preparation.
Because of their different staining characteristics, reticular fibres were initially thought to be completely different from collagen fibres. Cross-striations with the same periodicity as in coarse collagen fibres are however visible using electron microscopy. We now know that reticular fibres consist of collagen - although the main type of tropocollagen found in reticular fibres, type III, is different from that of the coarse collagen fibres.
Reticular fibres give support to individual cells, for example, in muscle and adipose tissue.
Suitable Slidessections of liver, spleen or lymph nodes - reticulin
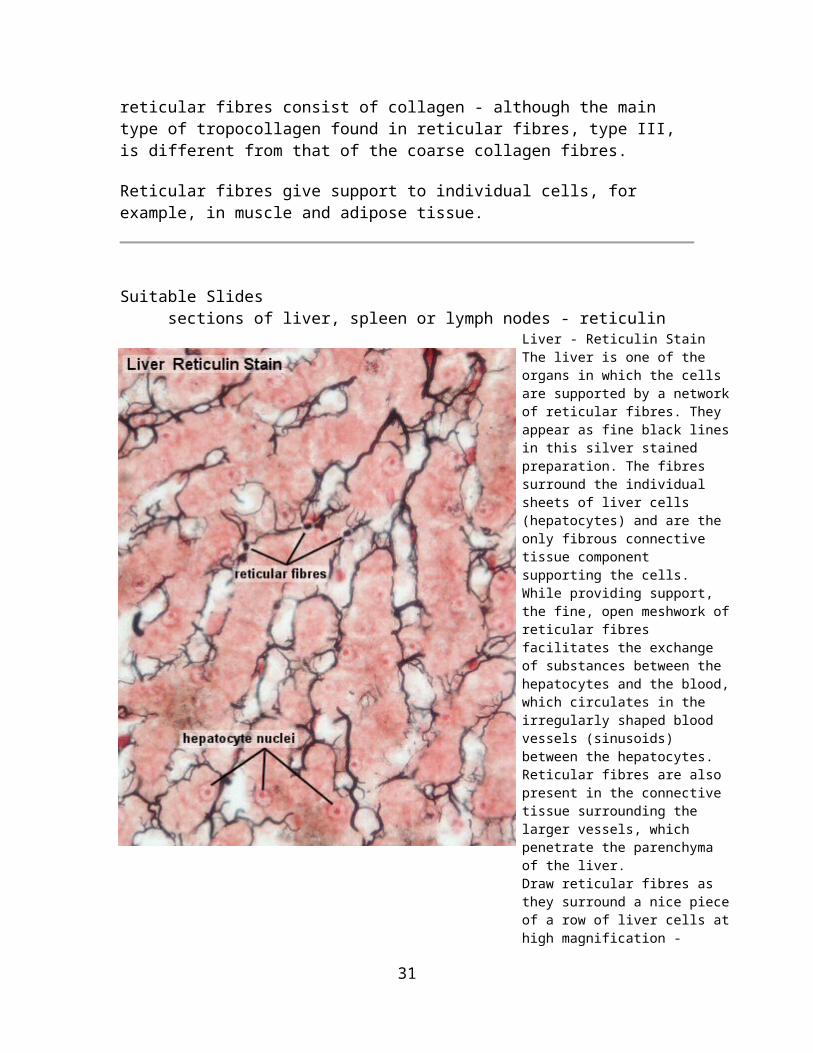
Liver - Reticulin Stain The liver is one of the organs in which the cells are supported by a network of reticular fibres. They appear as fine black lines in this silver stained preparation. The fibres surround the individual sheets of liver cells (hepatocytes) and are the only fibrous connective tissue component supporting the cells. While providing support, the fine, open meshwork of reticular fibres facilitates the exchange of substances between the hepatocytes and the blood, which circulates in the irregularly shaped blood vessels (sinusoids) between the hepatocytes. Reticular fibres are also present in the connective tissue surrounding the larger vessels, which penetrate the parenchyma of the liver.Draw reticular fibres as they surround a nice piece of a row of liver cells at high magnification - include a suitable scale and label your drawing.
Blood will not be visible in some types of preparations and the sinusoids appear empty.
22

Elastic fibers
Elastic fibres are coloured in fresh tissues - they are light yellow - but this colouration is only visible if large amounts of elastic fibres are present in the tissue, for example, in the elastic ligaments of the vertebral column. Special stains are necessary to show elastic fibres in tissue sections. Resorcin fuchsin is one of these stains, which gives the elastic fibres a dark violet colour.
Light microscopy does not reveal any substructure in the elastic fibres. Electron microscopy shows that elastic fibres consist of individual microfibrils, which are embedded in an amorphous matrix. The matrix accounts for about 90% of the fibre and is composed of the protein elastin. Neither the elastin nor the microfibrils are collagens.
Elastic fibres can be stretched to about 150% of their original length. They resume their original length if the tensile forces applied to the elastic fibres are relaxed.
Elastin is a somewhat odd protein in that its amino acid sequence does not determine a specific three-dimensional structure of the molecule. Instead, elastin remains unfolded as a "random coil". Elastin molecules are cross-linked to each other by desmosin and isodesmosin links, which are only found between elastin molecules. Tensile forces straighten the cross-linked mesh of elastin coils.
Suitable Slidessections of blood vessels or skin - elastin
Skin, human - elastin & van Gieson or Artery, human - elastin & eosinLike reticular fibres, elastic fibres require special stains to be visualized. Typically elastic fibres will appear as fine, dark violet and gently undulating fibres in the tissue. Elastic fibres can form membranes - not unlike the collagen membrane in the basal lamina of epithelia. This is the case at some levels in the walls of blood vessels.Collagen and elastic fibres intermingle in the dermis, i.e. the connective tissue beneath the epithelium of the skin. Immediately beneath the epithelium both fibre types are relatively fine - they appear much thicker in the deeper parts of the dermis. At least the internal elastic lamina should be visible in the smaller arteries which course through the dermis.A combination with a second stain is necessary to visualize other tissue components. Colours visible in the sections will depend on the method used in combination with the elastin stain. Eosin gives an even pink or red colour to many tissue components. Nuclei of cells remain unstained without the inclusion of the haematoxylin in the staining solutions.Identify the artery and the vein in the section. Their walls contain large amounts of elastic fibres. Blood vessels: draw a small section of the wall of a vessel, preferably an artery, at high magnification. Identify elastic laminae, fine and coarse elastic fibres in your drawing.Skin: draw a small section of the dermis - preferablyof a part of the dermis where both the very fine and the coarse fibres are visible.
23

Ground substance
Ground substance is found in all cavities and clefts between the fibres and cells of connective tissues. Water, salts and other low molecular substances are contained within the ground substance, but its main structural constituent are proteoglycans.Ground substance is soluble in most of the solvents used to prepare histological sections and therefore not visible in ordinary sections.
Proteoglycans are responsible for the highly viscous character of the ground substance. Proteoglycans consist of proteins (~5%) and polysaccharide chains (~95%), which are covalently linked to each other. The polysaccharide chains belong to one of the five types of glycosaminoglycans, which form the bulk of the polysaccharides in the ground substance.
24

Hyaluronan (or hyaluronic acid) is the dominant glycosaminoglycan in connective tissues. The molecular weight (MW) of hyaluronic acid is very high (~ MW 1,000,000 ). With a length of about 2.5 µm hyaluronan is very large. Hyaluronan serves as a "backbone" for the assembly of other glycosaminoglycans in connective and skeletal tissue, which results in even larger molecule complexes (MW 30,000,000 - 200,000,000).Hyaluronan is also a major component of the synovial fluid, which fills joint cavities, and the vitreous body of the eye.
The remaining four major glycosaminoglycans are chondroitin sulfate, dermatan sulfate, keratan sulfate and heparan sulfate. These glycosaminoglycans attach via core- and link-proteins to a backbone formed by the hyaluronic acid. The coiled arrangement of the hyaluronan and other attached glucosaminoglycans fills a roughly spherical space with a diameter of ~0.5 µm. This space is called a domain. Neighbouring domains overlap and form a more or less continuous three-dimensional molecular sieve in the interstitial spaces of the connective tissues.
The large polyanionic carbohydrates of the glycosaminoglycans bind large amounts of water and cations. The bound water in the domains forms a medium for the diffusion of substances of low molecular weight such as gases, ions and small molecules, which can take the shortest route, for example, from capillaries to connective tissue cells. Large molecules are excluded from the domains and have to find their way through the spaces between domains.
The restricted motility of larger molecules in the extracellular space inhibits the spread of microorganisms through the extracellular space. A typical bacterium ( 0.5 x 1 µm) is essentially immobilised in the meshwork formed by the domains. The pathogenicity of a bacterium is indeed to some extent determined by its ability to find its way through the mesh, and some of the more invasive types produce the enzyme hyaluronidase, which depolymerises hyaluronic acid.
The components of the ground substance, collagen, elastic and reticular fibres are synthesised by cells of the connective tissues, the fibrocytes.
Connective Tissue Cells
Connective tissue cells are usually divided into two groups based on their ability to move within the connective tissue. Fibrocytes (or fibroblasts) and fat cells are fixed cells. Macrophages, monocytes, lymphocytes, plasma cells, eosinophils and mast cells are wandering cells.
Fibrocytes
Fibrocytes are the most common cell type in connective tissues. They are the "true" connective tissue cells. Usually only their oval, sometimes flattened nuclei are visible in LM sections. The cytoplasm of a resting (i.e. inactive) fibrocyte does not contain many
25

organelles. This situation changes if the fibrocytes are stimulated, for example, by damage to the surrounding tissue. In this case the fibrocyte is transformed into a fibroblast, which contains large amounts of the organelles which are necessary for the synthesis and excretion of proteins needed to repair the tissue damage. Fibrocytes do not usually leave the connective tissue. They are, however, able to perform amoeboid movement.
The terms fibrocyte and fibroblast refer here to the inactive and active cells respectively - at times you will see the two terms used as synonyms without regard for the state of activity of the cell.
Reticular cells
Reticular cells are usually larger than an average fibrocyte. They are the "fibrocytes" of reticular connective tissue and form a network of reticular fibres, for example, in the lymphoid organs. Their nuclei are typically large and lightly stained (H&E) and the cytoplasm may be visible amongst the cells which are housed within the network of reticular fibres.
Adipocytes
Fat cells or adipocytes are fixed cells in loose connective tissue. Their main function is (what surprise!) the storage of lipids. If "well fed" the cytoplasm only forms a very narrow rim around a large central lipid droplet. The flattened nucleus may be found in a slightly thickened part of this cytoplasmic rim - if it is present in the section, which may not be the case since the diameter of an adipocyte (up to 100 µm) is considerable larger than the thickness of typical histological sections. A "starving" adipocyte may contain multiple small lipid droplets and gradually comes to resemble a fibrocyte.
Lipid storage/mobilisation is under nervous (sympathetic) and hormonal (insulin) control. Adipocytes also have an endocrine function - they secrete the protein leptin which provides brain centers which regulate appetite with feedback about the bodies fat reserves.Leptin deficiency in experimental animals results in obesity.
Adipocytes are very long-lived cells. Their number is determined by the number of preadipocytes (or lipoblast) generated during foetal and early postnatal development.
26

Macrophages
Macrophages arise from precursor cells called monocytes. Monocytes originate in the bone marrow from where they are released into the blood stream. They are actively mobile and leave the blood stream to enter connective tissues, where they differentiate into macrophages. Macrophages change their appearance depending on the demand for phagocytotic activity. Resting macrophages may be as numerous as fibrocytes.Resting macrophages are difficult to distinguish from fibrocytes in H&E stained sections.
Suitable Slidessections of liver or lymph nodes - carbon injected animal - trichrome or H&E sections of lung - H&EIn the lung, macrophages "patrol" the respiratory surfaces and ingest airborne particles which settle there. They "retire" to the connective tissue of the lung were they can be identified by the accumulations of fine particles in their cytoplasm. sections containing irregular connective connective tissues - Alcian blue & van GiesonIn Alcian blue & van Gieson stained sections macrophages may appear as intensely green, roundish cells with a dark nucleus.
27

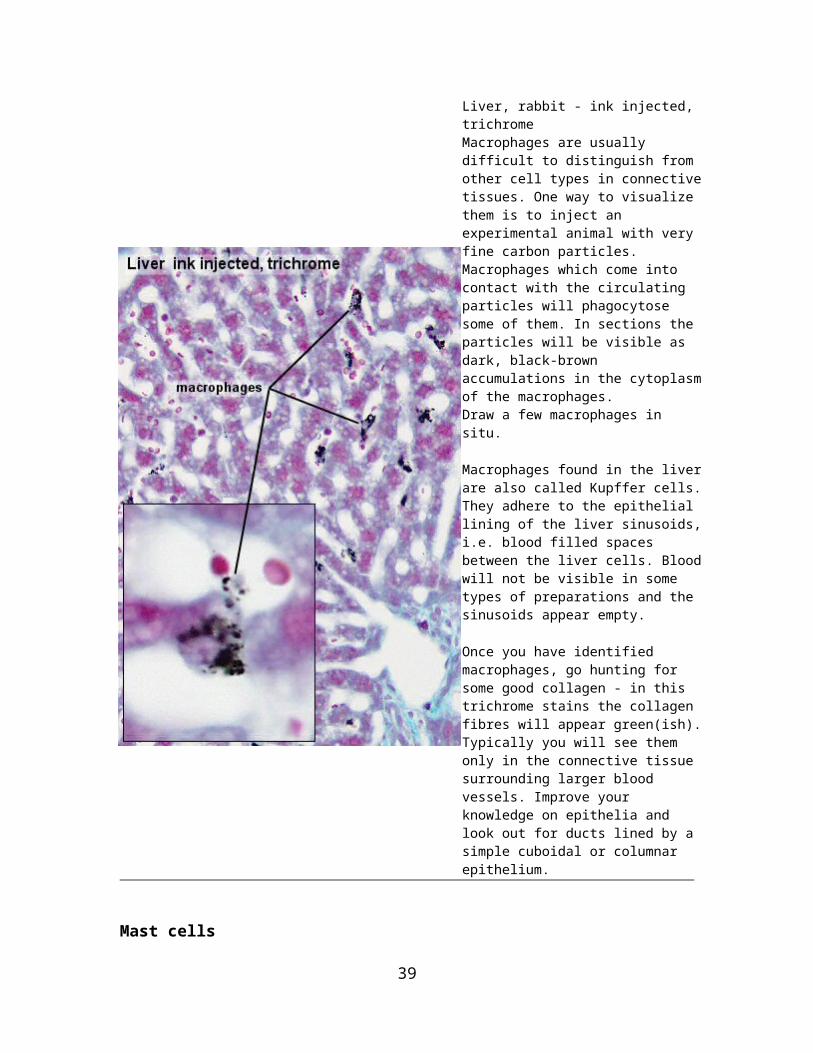
Liver, rabbit - ink injected, trichromeMacrophages are usually difficult to distinguish from other cell types in connective tissues. One way to visualize them is to inject an experimental animal with very fine carbon particles. Macrophages which come into contact with the circulating particles will phagocytose some of them. In sections the particles will be visible as dark, black-brown accumulations in the cytoplasm of the macrophages.Draw a few macrophages in situ.
Macrophages found in the liver are also called Kupffer cells. They adhere to the epithelial lining of the liver sinusoids, i.e. blood filled spaces between the liver cells. Blood will not be visible in some types of preparations and the sinusoids appear empty.
Once you have identified macrophages, go hunting for some good collagen - in this trichrome stains the collagen fibres will appear green(ish). Typically you will see them only in the connective tissue surrounding larger blood vessels. Improve your knowledge on epithelia and look out for ducts lined by a simple cuboidal or columnar epithelium.
Mast cells
Mast cells are - like macrophages, lymphocytes and eosinophils - in demand when something goes wrong in the connective tissue. Quite a few of them are present in healthy connective tissue as they stand on guard and monitor the local situation. The cytoplasm of mast cells is filled by numerous large vesicles. Mast cells discharge the contents of these vesicles if they come in contact with antigens, for example, proteins on the surface of an invading bacterium or, in allergic reactions, in response to antigens found, for example, on the surface of pollen grains.
The most prominent substances contained in the vesicles are heparin and histamine. They increase blood flow in close by vessels and the permeability of the vessel walls to plasma constituents and other white blood cells. By facilitating access to the area, mast cells facilitate an immune response to the antigen which triggered the release histamine and heparin.
28

Other connective tissue cells
Lymphocytes and plasma cells
Lymphocytes are usually small cells (6 - 8 µm). Their nuclei are round and stain very dark. The cytoplasm forms a narrow rim around the nucleus and may be difficult to see. There are many of them in the connective tissue underlying the epithelia of the gastrointestinal tract but usually much fewer in other connective tissues. Again, this situation may change - in this case with immunological reactions. Some lymphocytes may differentiate into plasma cells. Plasma cells are lymphocytes which produce antibodies. To accommodate the necessary organelles for this function the size of the cytoplasm increases dramatically and the cells become basophilic. Plasma cells can occasionally be spotted in the loose connective tissue present in sections.Like eosinophilic cells and monocytes, lymphocytes are white blood cells.
Eosinophilic cells
Eosinophilic cells are typically rounded or oval, large cells, which contain large amounts of bright red granules in their cytoplasm. They originate, like the monocytes, in the bone marrow. They enter connective tissues early in inflammatory reactions, where they phagocytose antigen-antibody complexes. Their numbers in healthy connective tissue vary with location, but a few of them can usually be found.
Mesenchymal cells
During development, mesenchymal cells give rise to other cell types of the connective tissue. A small number of them may persist into adulthood. Mesenchymal cells are smaller than fibrocytes and difficult to detect in histological sections. They may regenerate blood vessels or smooth muscle which have been lost as a consequence of tissue damage.
Suitable Slidessections of tongue, skin, mesentery or other sections containing epithelia and / or loose connective tissue - toluidine blue, cresyl violet
29

Mast Cells, Tongue - toluidine blue andMesentry, Rat - cresyl violet Mast cells are relatively frequent in the connective tissue benath the epithelium and between the muscle fibres of the tongue. In most connective tissue cells and the muscle fibres only the nucleus is stained by the toluidine blue. The cytoplasm of the mast cells is however filled with dark, blue / violet grains which represent their secretory vesicles. At low magnification mast cells stand out as large, dark dots among smaller and lighter stained nuclei and among the very weakly stained remaining connective tissue components.Draw a few mast cells in situ and label both the mast cells and some of the surrounding tissue components.
Connective Tissue Types
Loose connective tissue and dense connective tissues
These two tissues are distinguished according to the relative amounts of fibres they contain. Dense connective tissues are completely dominated by fibres. They are subdivided according to the spatial arrangement of the fibres in the tissue.
In dense irregular connective tissue the fibres do not show a clear orientation within the tissue but instead form a densely woven three-dimensional network. A good example is the dermis of the skin.
We talk about regular dense connective tissue if the fibres run parallel to each other. Good examples of regular dense connective tissue are tendons, ligaments and the fasciae and aponeuroses of muscles.
30
Loose connective tissue is relatively cell rich, soft and compliant. It is also rich in vessels and nerves. It is best understood as a kind of generalised connective tissue in which all connective tissue cell types may occur. Loose connective tissue may occur in some special variants: mucous connective tissue, reticular connective tissue and adipose tissue.
Suitable Slidessections of tendons or ligaments - van Gieson, H&E
Muscle-Tendon Junction, rat - van GiesonIn van Gieson stained preparations collagen stains dark red while other tissue components appear in varying shades of grey (nuclei) and yellow (cytoplasm). Areas of dense regular connective tissue are usually easy to identify in these preparations. Coarse collagen fibres are aligned with each other with only very narrow opens spaces between them. Like in most other connective tissues, there will be only a few cells between the fibres. Their cytoplasm is difficult to identify but the nuclei can be seen scattered among the collagen fibres. Nuclei are often elongated, and their long axis runs parallel to the course of the collagen fibresSketch part of the regular dense connective tissue.
31

Suitable Slidessections of skin or non-lactating mammary gland - H&E, van Gieson, trichrome
Non-lactating Breast - H&EDense irregular connective tissue forms the dermis of the skin, i.e. the layer of connective tissue immediately below the epithelium lining the surface of the skin. Beneath the skin forming the mammae (nipples), dense connective tissue areas are very extensive. This tissue surrounds the resting mammary gland. Both the high density of collagen fibres and the their irregular distribution are easily visible. Again, only a very small fraction of the tissue is taken up by cells. Like in van Gieson stained preparations, their cytoplasm is often not visible in H&E stained sections. Dark spots scattered between the collagen fibres represent the nuclei of the cells.Draw part of the connective tissue including some fibrocytes.
The excretory ducts of the mammary glands are called lactiferous ducts. They are lined by a quite nice stratified columnar epithelium. If you are working with a section of non-lactating breast look for the lactiferous ducts in the connective tissue.
Reticular connective tissue
Reticular connective tissue consists of reticular cells and the network of reticular fibres formed by them. Most connective tissues contain reticular fibres, but only in reticular connective tissue are they the dominant fibre type. In a number of tissues and organs, reticular connective tissue forms the structural framework in which the cells of the organ are suspended. The open meshwork of fine fibres is particularly useful in tissues and organs in which diffusion and / or cell movements are functionally important, for example, in the liver, lymph nodes and the spleen.
32

Adipose tissue
Adipose tissue is essentially loose connective tissue containing large numbers of adipocytes. There are two types of adipose tissue, which derive their names from the colour of the tissue (white or brown) and the number of lipid droplets found in the adipocytes.
Adipocytes of white, unilocular adipose tissue contain one large lipid droplet. Adipocytes of brown, multilocular adipose tissue contain many lipid droplets.
White adipose tissue does not only function in the storage of lipids. For example, in the palms of the hands, on the plantar surface (sole) of the feet and in the gluteal region (buttocks) it has a structural, cushioning function. In these regions, accumulations of adipocytes are surrounded by strong connective tissue fibres. Also, the distribution of white adipose tissue is different in males and females and is part of the secondary sexual characteristics. The storage and mobilisation of lipids does require quite some metabolic activity of the tissue. Consequently, adipose tissue has a rich supply of capillaries.
Brown adipose tissue occurs mainly during development and may account for 2 - 5 % of the body weight in a newborn. In adult individuals most of the brown fat has further differentiated into white fat. Adipocytes in brown fat contain plenty of mitochondria. A very rich capillary supply and the cytochromes found in the mitochondria give the tissue its characteristic colour. A protein (UCP-1 or thermogenin) found in these mitochondria decouples the oxidation of fatty acids from the generation of ATP. Instead, these cells generate heat.
The location of the brown fat reflects its heat-generating function. It is located in the axilla (armpits), between the shoulder blades, in the region of the neck and along large blood vessels. The heat generated by the brown fat warms the blood which supplies nearby organs or which re-enters the trunk from the limbs.
Suitable Slideswhite adipose tissue: sections of skin - H&E, trichrome, van GiesonSection are rarely prepared to show just white adipose tissue. The hypodermis, i.e. lightest and loosest appearing layer, of skin sections will typically contain large areas of adipose tissue. Other good candidates are bone sections which contain yellow bone marrow or sections of lymph nodes which are often embedded in adipose tissue. Small spots of adipose tissue should be present in many other sections. brown adipose tissue: sections of brown adipose tissue or kidney - trichrome, H&E Brown adipose tissue is often located in the connective tissue close to the renal hilus or in the renal sinus of sections which contain the entire kidney of small laboratory animals. Look for an indentation in the outline of the kidney, which corresponds to its hilus.
33

Thick Skin - H&EIn well-preserved thin sections, the adipocytes form a mosaic of rounded or slightly angular white tiles, which correspond to the locations of the lipid droplets, separated from each other by darker seams, which correspond to the cytoplasm of the adipocytes and the sparse connective tissue components between them. Because of the large size of the adipocytes you will only rarely see it "typical" signet ring-like appearance of the cells.Although the tissue my be a little distorted, thicker sections give a good three dimensional feel of the adipose tissue. The cytoplasmic rims of the adipocytes form thin veils which enclose the open spaces which were occupied by the lipid droplets. Oval adipocyte nuclei are often located close to the corners at which the adipocytes meet.Draw two or three adipocytes at high magnification and a survey image which illustrates the appearance of adipose tissue at low magnification.
34

Kidney - trichromeIn the renal sinus, islands of brown adipose tissue are often surrounded by white adipose tissue, which emphasises the different appearances of the two tissue types. In brown adipose tissue, the nuclei of adipocytes are round and located more or less centrally in a cytoplasm which, after the extraction of lipids during tissue preparation, looks very frothy. Cell borders can be difficult to identify. Capillaries are very frequent.Sketch the appearance of the brown adipose tissue. Contrast the characteristic features of white and brown adipose if both types are present side by side.
Mesenchymal connective tissue
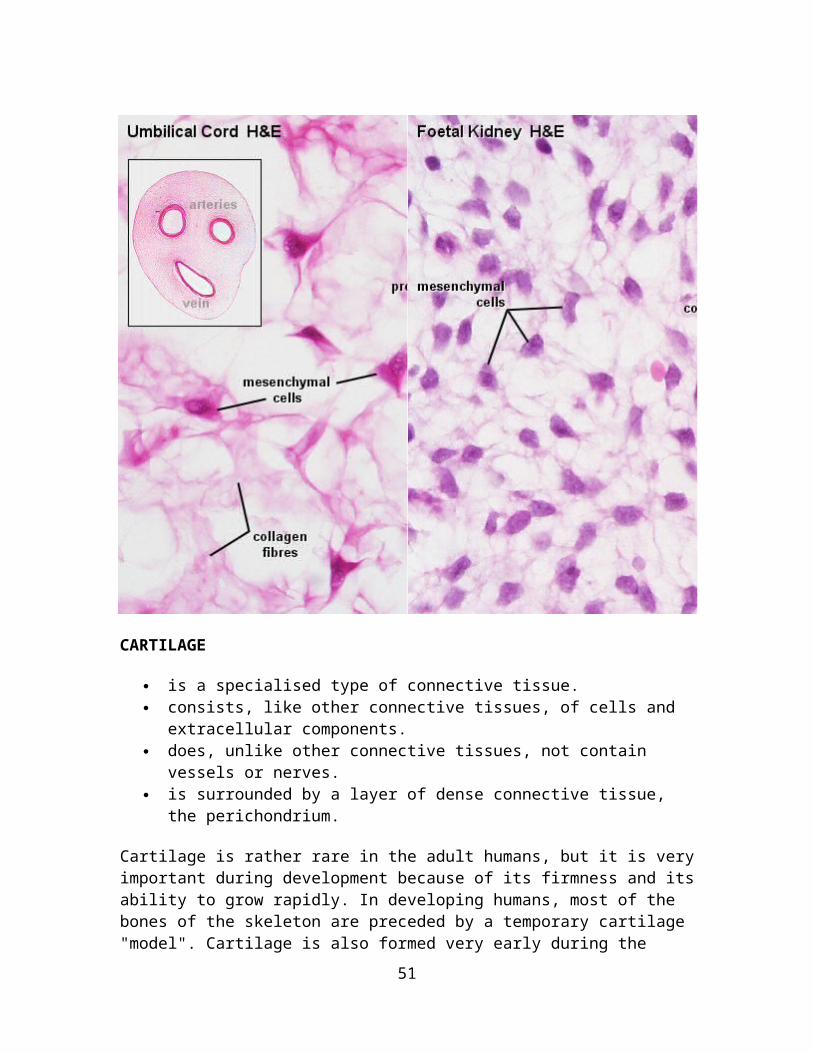
Mesenchyme forms the undifferentiated "filling" of the early embryo. It consists of mesenchymal cells, which interconnect by slender cell processes. Mesenchymal cells have stem cell properties, i.e. they are able give rise to other cell and tissues types. The wide extracellular space between the mesenchymal cells is occupied by ground substance, which can be stained with dyes that also stain mucin - hence the alternative name of this tissue type: mucoid connective tissue. Collagen or reticular fibres may not be visible at all or form a loose network between the cells. With fetal development, mesenchyme forms the connective tissue between and within the developing tissues and organs. Mucoid connective tissue also forms a compliant cushion around the vessels of the umbilical cord, where it is also called Wharton's jelly.
In adult humans, mesenchymal connective tissue is only found in the dental pulp.
35

Suitable Slidessections of umbilical cord, tooth (pulp), or sections of embryonic and early foetal development - H&E, Azan or Alcian blue & van Giesonsection usable for "intramembranous ossification" during foetal development will contain areas of mucoid connective tissue around the developing bone.
Umbilical Cord, Human - H&E andFoetal Kidney, Human - H&E Within the umbilical cord you will be able to identify three large vessels and their walls. Mucoid connective tissue fills the space between the vessels and the simple squamous epithelium lining the surface of the umbilical cord. Note the very fine appearance of the collagen fibres and the lack of apparent specialisations in this type of connective tissue.The number of cells and appearance of the collagen fibres vary depending on the precise location of the tissue. In some locations, mucoid connective tissues will contain a large number of cells and only a few, very delicate collagen fibres. Examples are dental pulp and the mucoid connective tissue which is found between the developing tubuli and glomeruli of the foetal kidney.A small drawing should be sufficient to capture the appearance of the tissue.
36

CARTILAGE
is a specialised type of connective tissue. consists, like other connective tissues, of cells and extracellular components. does, unlike other connective tissues, not contain vessels or nerves. is surrounded by a layer of dense connective tissue, the perichondrium.
Cartilage is rather rare in the adult humans, but it is very important during development because of its firmness and its ability to grow rapidly. In developing humans, most of the bones of the skeleton are preceded by a temporary cartilage "model". Cartilage is also formed very early during the repair of bone fractures.
Types of Cartilage
Hyaline Cartilage
Hyaline cartilage develops, like other types of connective tissue, from mesenchymal cells. From about the fifth foetal week precursor cells become rounded and form densely packed cellular masses, chondrification centres. The cartilage-forming cells, chondroblasts, begin to secrete the components of the extracellular matrix of cartilage. The extracellular matrix consists of, ground substance (hyaluronan, chondroitin sulfates and keratan sulfate) and tropocollagen, which polymerises extracellularly into fine collagen fibres.
Tropocollagen type II is the dominant form in collagen fibres of almost all types of cartilage.
As the amount of matrix increases the chondroblasts become separated from each other and are, from this time on, located isolated in small cavities within the matrix, the lacunae. Concurrently the cells differentiate into mature cartilage cells, chondrocytes.
Growth occurs by two mechanisms
Interstitial growth - Chondroblasts within the existing cartilage divide and form small groups of cells, isogenous groups, which produce matrix to become separated from each other by a thin partition of matrix. Interstitial growth occurs mainly in immature cartilage.
37

Appositional growth - Mesenchymal cells surrounding the cartilage in the deep part of the perichondrium (or the chondrogenic layer) differentiate into chondroblasts. Appositional growth occurs also in mature cartilage.
Like all protein-producing cells, chondroblasts contain plenty of rough endoplasmatic reticulum while they produce matrix. The amount of rough endoplasmatic reticulum decreases as the chondroblasts mature into chondrocytes. Chondrocytes fill out the lacunae in the living cartilage.
The matrix appears structureless because the collagen fibres are too fine to be resolved by light microscopy (~20nm), and because they have about the same refractive index as the ground substance. Collagen accounts for ~ 40% of the dry weight of the matrix.
The matrix near the isogenous groups of chondrocytes contains larger amounts and different types of glycosaminoglycans than the matrix further away from the isogenous groups. This part of the matrix is also termed territorial matrix or capsule. In H&E stained sections the territorial matrix is more basophilic, i.e. it stains darker. The remainder of the matrix is called the interterritorial matrix. Fresh cartilage contains about 75% water which forms a gel with the components of the ground substance. Cartilage is nourished by diffusion of gases and nutrients through this gel.
Suitable Slides
sections of the trachea or larynx - H&E, van Gieson
Trachea, cat, H&E and Trachea, cat, van GiesonBoth stains are equally well suited to look at the organisation of hyaline cartilage. The van Gieson method stains collagen red. The cartilage appears as a wide red zone underneath the epithelium and loose connective tissue, which line the lumen of the trachea. The staining may appear a little lighter close to the lacunae. This lighter stained zone defines the territorial matrix surrounding the lacunae and chondrocytes. Colour intensities appear reversed in the H&E stained section. The two compartments of the matrix are usually better defined than in van Gieson stained sections. The interterritorial matrix appears very light; the territorial matrix is somewhat darker. Groups of chondrocytes surrounded by these lighter (van Gieson) or darker (H&E) staining zones belong to the same isogenous group. A layer of dense connective tissue surrounding the cartilage and blending with it is the perichondrium.The isogenous groups may form small "squares" (e.g. four chondrocytes, separated by thin cartilage membranes, in a 2x2 arrangement) or short columns (e.g. four chondrocytes in a 1x4 arrangement).Draw a small section of the cartilage and identify in your drawing territorial matrix, interterritorial matrix, isogenous groups, and chondrocytes. Think about how the spatial arrangement of chondrocytes in the isogenous group may reflect patterns of cell divisions.
38
Elastic Cartilage
occurs in the epiglottic cartilage, the corniculate and cuneiform cartilage of the larynx, the cartilage of the external ear and the auditory tube.
corresponds histologically to hyaline cartilage, but, in addition, elastic cartilage contains a dense network of delicately branched elastic fibres.
39

Suitable Slides Sections of the epiglottis - elastin
Epiglottis, human, elastinPreparations of the epiglottis are usually dominated by the cartilage surrounded by varying amounts of connective tissue and epithelia. The appearance of the cartilage (in this preparation a blue-green colour) will depend on the method used to show tissue components other than elastic fibres. Although the matrix appears blue-green, the typical organisation of cartilage is readily visible. Within the green matrix you can see the fine elastic fibres which give this cartilage its elastic properties. The elastic fibres may form dense masses in which individual fibres are difficult to distinguish. The staining of these masses of fibres may appear more reddish than dark-violet.A change of the colour of the stain in intensely stained tissue areas is called "metachromatic staining".Draw and label a small section of elastic cartilage.
40

Fibrous Cartilage
is a form of connective tissue transitional between dense connective tissue and hyaline cartilage. Chondrocytes may lie singly or in pairs, but most often they form short rows between dense bundles of collagen fibres. In contrast to other cartilage types, collagen type I is dominant in fibrous cartilage.
is typically found in relation to joints (forming intra-articular lips, discs and menisci) and is the main component of the intervertebral discs.
merges imperceptibly into the neighbouring tissues, typically tendons or articular hyaline cartilage. It is difficult to define the perichondrium because of the fibrous appearance of the cartilage and the gradual transition to surrounding tissue types.
Suitable Slides
sections of intervertebral discs or articular discs - H&E, van Gieson
Fibrous Cartilage, Intervertebral Disc, sheep, H&E and Articular Disc, rabbit, H&EThe fibrous cartilage forming the intervertebral discs varies in appearance from the center of the disc (the nucleus pulposus) the the periphery of the disc (the annulus fibrosus). Centrally, the fibrous matrix is very loose. The jelly-like consistency of the central part allows the intervertebral discs to function as a shock absorber. Towards the periphery, the fibrous matrix is organised into layers. It is often visible that the fibres of different layers are oriented at angles to each other - similar to the orientation of the thread in radial tires. Chondrocytes are very flattened in the periphery and may be difficult to find.Midway between periphery and center of the intervertebral disc, chondrocytes are scattered singly or in small isogenous groups in the dense fibrous matrix of the cartilage. If you take a close look at the cells you will see that their appearance actually resembles that of chondrocytes in other types of cartilage - their characteristic appearance distinguishes fibrous cartilage preparations from connective tissues. The very regular arrangement of the fibres in the articular disc may initially let you guess at dense regular connective tissue. Isogenous groups of chondrocytes again identify the tissue as fibrous cartilage.Draw a small section of the fibrous cartilage, including (if possible) a group of chondrocytes.
41

Articular Cartilage
is a specialised form of hyaline cartilage. transforms the articulating ends of the bones into lubricated, wear-proof, slightly
compressible surfaces, which exhibit very little friction. is not surrounded by a perichondrium and is partly vascularised. is, depending on the arrangement of chondrocytes and collagenous fibres, divided
into several zones:
Tangential layer
Chondrocytes are rather small and flattened parallel to the surface. The most superficial part (lamina splendens) is devoid of cells. Collagen fibres in the matrix of the tangential layer are very fine. They run parallel to the surface of the cartilage.
42

Similar to the collagen fibres of the skin, the general orientation of collagen fibres in articular cartilage is determined by tensile and compressive forces at the articulating surfaces.
Transitional zone
The chondrocytes are slightly larger, are round and occur both singly and in isogenous groups. Collagen fibres take an oblique course through the matrix of the transitional zone.
Radial zone
Fairly large chondrocytes form radial columns, i.e. the stacks of cells are oriented perpendicular to the articulating surface. The course of the collagen fibres follows the orientation of the chondrocyte columns.
Calcified cartilage layer
It rests on the underlying cortex of the bone. The matrix of the calcified cartilage layer stains slightly darker (H&E) than the matrix of the other layers.
The main source of nourishment for articular cartilage is the synovial fluid, which fills the joint cavity. Additional small amounts of nutrients are derived from blood vessels that course through the calcified cartilage close to the bone.Living chondrocytes have been found in small pieces of cartilage floating in the joint cavity after damage to the articular cartilage.
Osteoarthritis, the slow progressive degeneration of articular cartilage, is the most common joint disease. It may be caused by persistent and abnormally high loads on the joint surfaces, which initially result in the loss of proteoglycans and chondrocytes from the articulating surface of the cartilage. Subsequently, the cartilage may crack (fibrillate), erode and expose the underlying bone.
Suitable Slides
Sections of large joints - H&ELayers are difficult to identify in the articular cartilage of small joints.
Articular Cartilage, bovine, H&EThe layers of articular cartilage are easiest to identify in large joints. Note the changing orientations of the lacunae and isogenous groups at different depth in the cartilage. The changing orientations of chondrocytes and isogenous groups reflect the orientations of the collagen fibres in the matrix. The fibres are not visible in the slide. The darker hue of the cartilage close to the bone is caused by the calcification of the cartilage.Draw the articular cartilage at low magnification. Indicate in your drawing the preferred orientations of lacunae and isogenous groups and the expected orientation of collagen fibres.
43

Degeneration and Regeneration of Cartilage
Due to the fairly poor access of nutrients to the chondrocytes they may atrophy in deep parts of thick cartilage. Water content decreases and small cavities arise in the matrix, which often leads to the calcification of the cartilage. This further compromises nutrition. The chondrocytes may eventually die, and the cartilage is gradually transformed into bone.
Chondrogenic activity of the perichondrium is limited to the period of active growth before adulthood. Although chondrocytes are able to produce matrix components throughout life, their production can not keep pace with the repair requirements after acute damage to hyaline or articular cartilage. If these cartilages are injured after the period of active growth, the defects are usually filled by connective tissue or fibrous cartilage. The extracellular matrix of these "repair tissues" is only poorly integrated with the matrix of the damaged cartilage.
44

Fortunately, cartilage is rather well suited for transplantation - the metabolism of the chondrocytes is rather slow, the antigenic power of cartilage is low, and it is difficult, if not impossible, for antibodies or cells of the immune system to diffuse through the matrix into the cartilage.
45

BONE
Bone is the main component of the skeleton in the adult human. Like cartilage, bone is a specialized form of dense connective tissue. Bone gives the skeleton the necessary rigidity to function as attachment and lever for muscles and supports the body against gravity.
Two types of bone can be distinguished macroscopically:
Trabecular bone (also called cancellous or spongy bone) consists of delicate bars and sheets of bone, trabeculae, which branch and intersect to form a sponge like network. The ends of long bones (or epiphyses) consist mainly of trabecular bone.
Compact bone does not have any spaces or hollows in the bone matrix that are visible to the eye. Compact bone forms the thick-walled tube of the shaft (or diaphysis) of long bones, which surrounds the marrow cavity (or medullary cavity). A thin layer of compact bone also covers the epiphyses of long bones.
Bone is, again like cartilage, surrounded by a layer of dense connective tissue, the periosteum. A thin layer of cell-rich connective tissue, the endosteum, lines the surface of the bone facing the marrow cavity. Both the periosteum and the endosteum possess osteogenic potency. Following injury, cells in these layers may differentiate into osteoblasts (bone forming cells) which become involved in the repair of damage to the bone.
46

Histological Organization of Bone
Compact Bone
Compact bone consists almost entirely of extracellular substance, the matrix. Osteoblasts deposit the matrix in the form of thin sheets which are called lamellae. Lamellae are microscopical structures. Collagen fibres within each lamella run parallel to each other. Collagen fibres which belong to adjacent lamellae run at oblique angles to each other. Fibre density seems lower at the border between adjacent lamellae, which gives rise to the lamellar appearance of the tissue. Bone which is composed by lamellae when viewed under the microscope is also called lamellar bone.
In the process of the deposition of the matrix, osteoblasts become encased in small hollows within the matrix, the lacunae. Unlike chondrocytes, osteocytes have several thin processes, which extend from the lacunae into small channels within the bone matrix , the canaliculi. Canaliculi arising from one lacuna may anastomose with those of other lacunae and, eventually, with larger, vessel-containing canals within the bone. Canaliculi provide the means for the osteocytes to communicate with each other and to exchange substances by diffusion.
In mature compact bone most of the individual lamellae form concentric rings around larger longitudinal canals (approx. 50 µm in diameter) within the bone tissue. These canals are called Haversian canals. Haversian canals typically run parallel to the surface and along the long axis of the bone. The canals and the surrounding lamellae (8-15) are called a Haversian system or an osteon. A Haversian canal generally contains one or two capillaries and nerve fibres.
Irregular areas of interstitial lamellae, which apparently do not belong to any Haversian system, are found in between the Haversian systems. Immediately beneath the periosteum and endosteum a few lamella are found which run parallel to the inner and outer surfaces of the bone. They are the circumferential lamellae and endosteal lamellae.
A second system of canals, called Volkmann's canals, penetrates the bone more or less perpendicular to its surface. These canals establish connections of the Haversian canals with the inner and outer surfaces of the bone. Vessels in Volkmann's canals communicate with vessels in the Haversian canals on the one hand and vessels in the endosteum on the other. A few communications also exist with vessels in the periosteum.
Trabecular Bone
The matrix of trabecular bone is also deposited in the form of lamellae. In mature bones, trabecular bone will also be lamellar bone. However, lamellae in trabecular bone do not form Haversian systems. Lamellae of trabecular bone are deposited on preexisting trabeculae depending on the local demands on bone rigidity.
47

Osteocytes, lacunae and canaliculi in trabecular bone resemble those in compact bone.
Note the distinction between macroscopic (visible to the eye) and microscopic (only visible under the microscope) appearance when the bone is named. Lamellar bone forms both trabecular bone and compact bone, which are the two macroscopically recognizable bone forms.
Suitable Slides sections of compact bone (usually part of the diaphysis of a long bone) - ground (unstained), Schmorl stained or H&E
Compact bone, human - Schmorl stainLamellae which run parallel to the surface of the bone are visible both on the outer, convex surface of the bone (circumferential lamellae) and on the inner, concave surface of the bone facing the marrow cavity (endosteal lamellae). The surface formed by the endosteal lamellae is often more irregular than the surface formed by the circumferential lamellae. The space between these two sets of lamellae is filled by Haversian systems and interstitial lamellae. Only few of the Haversian systems are "textbook" circular. Osteocyte lacunae are visible between the lamellae. Canaliculi become visible at high magnification (illustrated in the ground section below).You should be able to see, draw and identify Haversian systems, interstitial and circumferential lamellae and/or endosteal lamellae.
48

Compact bone, human - ground (unstained)The osteocyte lacunae are the main feature of the ground section. They are visible as elongated black spots in the bone matrix. Canaliculi, radiate from the lacunae into the surrounding bone matrix. Some lamellae are visible in the ground section. There is actually no distinct border between most lamellae, but our brain can use the elongated osteocyte lacunae and their orientation to "reconstruct" the lamellae. Volkman's canals connect to a few of the Haversian canals.
49

Suitable Slides sections of part of a vertebra or an epiphysis of a long bone - H&E, van GiesonSections prepared to show articular cartilage will often also contain trabecular bone in the epiphysis below the articular cartilage.
Articular cartilage, bovine - H&EThin sheets and bars of bone, trabeculae, are visible at low magnification. Although they may appear as individual pieces in sections, they form an interconnected meshwork in the living bone. The spaces between the trabeculae, the marrow cavity of the epiphysis, is filled by either red bone marrow or yellow bone marrow. At high magnification, elongated osteocyte lacunae, which in well preserved tissue still contain osteocytes, are visible in the matrix. If the H&E stain also turned out well, it should be visible that the matrix of the trabecular bone is formed by lamellae. Canaliculi are present but hard to identify in most H&E stained sections. Haversian systems are not present in the trabeculae.In trabecular bone obtained from young individuals, in which the bone is still growing, small areas of calcified cartilage are occasionally seen in the bone trabeculae. They are remnants of the cartilage scaffold on which bone matrix was deposited during the formation of the trabeculae. With the reorganisation of bone such areas will eventually be lost.
50
Bone Matrix and Bone Cells
Bone Matrix
Bone matrix consists of collagen fibres (about 90% of the organic substance) and ground substance. Collagen type I is the dominant collagen form in bone. The hardness of the matrix is due to its content of inorganic salts (hydroxyapatite; about 75% of the dry weight of bone), which become deposited between collagen fibres.
Calcification begins a few days after the deposition of organic bone substance (or osteoid) by the osteoblasts. Osteoblasts are capable of producing high local concentration of calcium phosphate in the extracellular space, which precipitates on the collagen molecules. About 75% of the hydroxyapatite is deposited in the first few days of the process, but complete calcification may take several months.
Bone Cells
Osteoprogenitor cells (or stem cells of bone)
are located in the periosteum and endosteum. They are very difficult to distinguish from the surrounding connective tissue cells. They differentiate into
Osteoblasts (or bone forming cells).
Osteoblasts may form a low columnar "epitheloid layer" at sites of bone deposition. They contain plenty of rough endoplasmatic reticulum (collagen synthesis) and a large Golgi apparatus. As they become trapped in the forming bone they differentiate into
Osteocytes.
Osteocytes contain less endoplasmatic reticulum and are somewhat smaller than osteoblasts.
Osteoclasts
are very large (up to 100 µm), multi-nucleated (about 5-10 visible in a histological section, but up to 50 in the actual cell) bone-resorbing cells. They arise by the fusion of monocytes (macrophage precursors in the blood) or macrophages. Osteoclasts attach themselves to the bone matrix and form a tight seal at the rim of the attachment site. The cell membrane opposite the matrix has deep invaginations forming a ruffled border. Osteoclasts empty
51
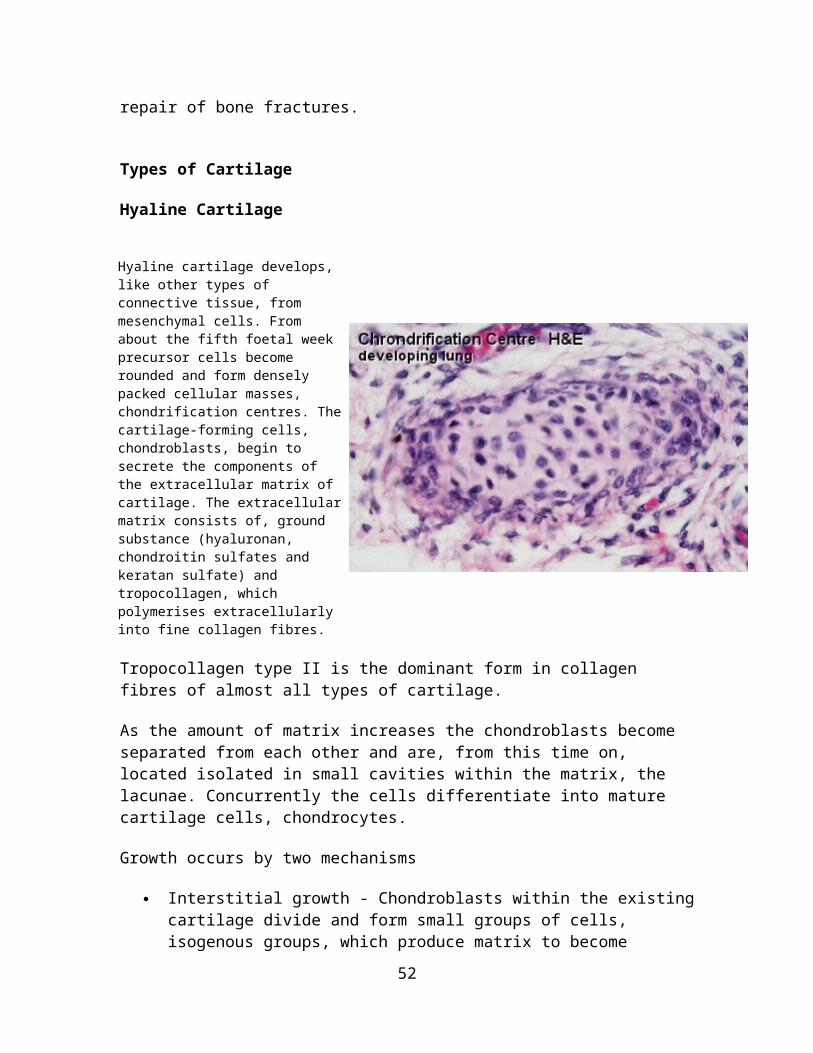
the contents of lysosomes into the extracellular space between the ruffled border and the bone matrix. The released enzymes break down the collagen fibres of the matrix. Osteoclasts are stimulated by parathyroid hormone (produced by the parathyroid gland) and inhibited by calcitonin (produced by specialised cells of the thyroid gland). Osteoclasts are often seen within the indentations of the bone matrix that are formed by their activity (resorption bays or Howship's lacunae).
Formation of Bone
Bones are formed by two mechanisms: intramembranous ossification (bones of the skull, part of the mandible and clavicle) or endochondral ossification.
Intramembranous Ossification
Intramembranous ossification occurs within a membranous, condensed plate of mesenchymal cells. At the initial site of ossification (ossification centre) mesenchymal cells (osteoprogenitor cells) differentiate into osteoblasts. The osteoblasts begin to deposit the organic bone matrix, the osteoid. The matrix separates osteoblasts, which, from now on, are located in lacunae within the matrix. The collagen fibres of the osteoid form a woven network without a preferred orientation, and lamellae are not present at this stage. Because of the lack of a preferred orientation of the collagen fibres in the matrix, this type of bone is also called woven bone. The osteoid calcifies leading to the formation of primitive trabecular bone.
Further deposition and calcification of osteoid at sites where compact bone is needed leads to the formation of primitive compact bone.
Note the distinction between macroscopic and microscopic appearance when the bone is named. We again have the two macroscopically different forms of bone - trabecular bone and compact bone - but their early developmental ("primitive") forms consist of woven bone.
Through subsequent reorganisation the primitive compact and trabecular bone is converted into mature compact and trabecular bone. During reorganisation and growth, woven bone will, in time, be replaced by lamellar bone.
Intramembranous ossification does not require the existence of a cartilage bone model.
52

Suitable Slides sections of the developing mandible (or some other bones of the skull) or clavicle - H&E, van GiesonSections prepared to show endochondral ossification (see below) may be an alternative if no specifically prepared slides of intramembranous ossification are available. The periosteal collar, i.e. the manchette of bone forming around the diaphysis of the cartilage model of the bone, is formed by a mechanism similar to intramembranous ossification and results in the deposition of woven bone.
Mandible, intramembranous ossification - H&EThe developing bone will in sections usually be associated with a number of other tissues which develop in close association with it. In case of the mandible, there can be developing teeth, the tongue, skin and salivary glands.The first job - best done at low magnification - is to find the developing bone. It should look like a coarse meshwork (trabecular bone) of pink tissue surrounding patches of much lighter or unstained tissue. Lamellae are not visible (woven bone) and the lacunae are larger than lacunae in mature bone. Ossification centres appear as areas of a gradual transition from connective tissue to bone. Light, pinkish bone matrix is found between the osteoblasts.Depending on the state of development of the bone, it is occasionally possible to find bone trabeculae which are lined by a layer of osteoblasts. These osteblasts are depositing the first lamellae on the already existing trabeculae. The trabeculae will therefore have a core of woven bone, which is surrounded by lamellar bone. Compare the shapes, sizes and frequencies of lacunae in lamellar and woven bone if both types of bone are present.
53

Endochondral Ossification
Most bones are formed by the transformation of cartilage "bone models", a process called endochondral ossification. A periosteal bud invades the cartilage model and allows osteoprogenitor cells to enter the cartilage. At these sites, the cartilage is in a state of hypertrophy (very large lacunae and chondrocytes) and partial calcification, which eventually leads to the death of the chondrocytes.Invading osteoprogenitor cells mature into osteoblasts, which use the framework of calcified cartilage to deposit new bone. The bone deposited onto the cartilage scaffold is lamellar bone. The initial site of bone deposition is called a primary ossification centre. Secondary ossification centres occur in the future epiphyses of the bone.A thin sheet of bone, the periosteal collar, is deposited around the shaft of the cartilage model. The periosteal collar consists of woven bone.
Close to the zone of ossification, the cartilage can usually be divided into a number of distinct zones :
1. Reserve cartilage, furthest away from the zone of ossification, looks like immature hyaline cartilage.
54

2. A zone of chondrocyte proliferation contains longitudinal columns of mitotically active chondrocytes, which grow in size in
3. the zone of cartilage maturation and hypertrophy. 4. A zone of cartilage calcification forms the border between cartilage and the zone
of bone deposition.
Primary and secondary ossification centres do not merge before adulthood. Between the diaphysis and the epiphyses a thin sheet of cartilage, the epiphyseal plate, is maintained until adulthood. By continuing cartilage production, the epiphyseal plate provides the basis for rapid growth in the length of the bone. Cartilage production gradually ceases in the epiphyseal plate as maturity is approached. The epiphyseal plate is finally removed by the continued production of bone from the diaphyseal side.
Bone formation and bone resorption go hand in hand during the growth of bone. This first deposited trabecular bone is removed as the zone of ossification moves in the direction of the future epiphyses. This process creates the marrow cavity of the bones. Simultaneously, bone is removed from the endosteal surface and deposited on the periosteal surface of the compact bone which forms the diaphysis. This results in a growth of the diameter of the bone.
Suitable Slides sections of bones during the early stages of their development - H&E, van Gieson
Foetal vertebral column, human - H&EHold the section against a light background. The cartilage models will stain very light, and the outlines of the skeletal structures they will form can often be identified. If an ossification centre is present, it will appear as a darker area within the cartilage model. The zonation of the cartilage should be visible in all ossification centres. How much bone is present depends on how far ossification has proceeded. The newly formed bone trabeculae will often consist of lamellar bone with a more or less extensive core of darkly staining calcified cartilage. The lamellar organisation of the bone may not be visible.How the ossification centre exactly will look also depends on the plane of the section in relation to the ossification centre.
55

Suitable Slides sections of the epiphyseal disc of growing bones - H&E
56

Growing bone, rabbit - H&EThe cartilage model has almost entirely been transformed into bone. The only remaining cartilage is found in the epiphyseal disk. Zones of cartilage proliferation, hypertrophy and calcification are visible at high magnification, but only on one side of the epiphyseal disk - towards the diaphysis, which increases in length as the cartilage generated by the epiphyseal disc is transformed into bone.Osteoclasts may be found on the newly formed trabeculae or associated with parts of the cartilage scaffold.Draw the region of the epiphyseal disks. Identify in your drawing the epiphyseal disk and the bone of the epiphysis and diaphysis. Indicate the direction of cartilage proliferation in the epiphyseal disk and the direction of bone growth.
Reorganisation and Restoration of Bone
Changes in the size and shape of bones during the period of growth imply some bone reorganisation. Osteoblast and osteoclast constantly deposit and remove bone to adjust its properties to growth-related demands on size and/or changes of tensile and compressive forces.Although the reorganisation of bone may not result in macroscopically visible changes of bone structure, it continues throughout life to mend damage to bone (e.g. microfractures) and to counteract the wear and tear occurring in bone. Osteoclasts and osteoblasts remain the key players in this process. Osteoclasts "drill" more or less circular tunnels within existing bone matrix. Osteoblasts deposit new lamellae of bone matrix on the walls of these tunnels resulting in the formation of a new Haversian system within the matrix of compact bone. Parts of older Haversian systems, which may remain between the new ones, represent the interstitial lamellae in mature bone. Capillaries and nerves sprout into new Haversian canals. Restorative activity continues in aged humans (about 2% of the Haversian systems seen in an 84 year old individual contained lamellae that had been formed within 2 weeks prior to death!). However, the Haversian systems tend to be smaller in older individuals and the canals are larger because of slower bone deposition. If these age-related changes in the appearance of the Haversian systems are pronounced they are termed osteopenia or senile osteoporosis. The reduced strength of bone affected by osteoporosis will increase the likelihood of fractures.
57

MUSCLE
Motion, as a reaction of multicellular organisms to changes in the internal and external environment, is mediated by muscle cells.
The basis for motion mediated by muscle cells is the conversion of chemical energy (ATP) into mechanical energy by the contractile apparatus of muscle cells. The proteins actin and myosin are part of the contractile apparatus. The interaction of these two proteins mediates the contraction of muscle cells. Actin and myosin filaments, each composed of many action and myosin molecules, form myofibrils arranged parallel to the direction of cellular contraction.
A further specialisation of muscle cells is an excitable cell membrane which propagates the stimuli which initiate cellular contraction.
Three structurally and functionally distinct types of muscle are found in vertebrates:
1. smooth muscle, 2. skeletal muscle and 3. cardiac muscle.
Smooth Muscle
Smooth muscle consists of spindle shaped cells of variable size.The largest smooth muscle cells occur in the uterus during pregnancy (12x600 µm). The smallest are found around small arterioles (1x10 µm).
Smooth muscle cells contain one centrally placed nucleus.The chromatin is finely granular and the nucleus contains 2-5 nucleoli.
The innervation of smooth muscle is provided by the autonomic nervous system. Smooth muscle makes up the visceral or involuntary muscle.
Structure of smooth muscle
In the cytoplasm, we find longitudinally oriented bundles of the myofilaments actin and myosin. Actin filaments insert into attachment plaques located on the cytoplasmic surface of the plasma membrane. From here, they extend into the cytoplasm and interact with myosin filaments. The myosin filaments interact with a second set of actin filaments which insert into intracytoplasmatic dense bodies. From these dense bodies further actin filaments extend to interact with yet another set of myosin filaments. This sequence is repeated until the last actin filaments of the bundle again insert into attachment plaques.
In principle, this organisation of bundles of myofilaments, or myofibrils, into repeating units corresponds to that in other muscle types. The repeating units of different myofibrils are however not aligned with each other, and myofibrils do not run exactly longitudinally
58

or parallel to each other through the smooth muscle cells. Striations, which reflect the alignment of myofibrils in other muscle types, are therefore not visible in smooth muscle.
Smooth endoplasmatic reticulum is found close to the cytoplasmatic surface of the plasma membrane. Most of the other organelles tend to accumulate in the cytoplasmic regions around the poles of the nucleus. The plasma membrane, cytoplasm and endoplasmatic reticulum of muscle cells are often referred to as sarcolemma, sarcoplasm, and sarcoplasmatic reticulum.
During contraction, the tensile force generated by individual smooth muscle cells is conveyed to the surrounding connective tissue by the sheath of reticular fibres. These fibres are part of a basal lamina which surrounds muscle cells of all muscle types. Smooth muscle cells can remain in a state of contraction for long periods. Contraction is usually slow and may take minutes to develop.
Origin of smooth muscle
Smooth muscle cells arise from undifferentiated mesenchymal cells. These cells differentiate first into mitotically active cells, myoblasts, which contain a few myofilaments. Myoblasts give rise to the cells which will differentiate into mature smooth muscle cells.
Types of smooth muscle
Two broad types of smooth muscle can be distinguished on the basis of the type of stimulus which results in contraction and the specificity with which individual smooth muscle cells react to the stimulus:
1. The multiunit type represents functionally independent smooth muscle cells which are often innervated by a single nerve terminal and which never contract spontaneously (e.g. smooth muscle in the walls of blood vessels).
2. The visceral type represents bundles of smooth muscle cells connected by GAP junctions, which contract spontaneously if stretched beyond a certain limit (e.g. smooth muscle in the walls of the intestines).
59

Suitable Slides Sections of the intestines (duodenum, jejunum, ileum or colon) - H&E
Jejunum, baboon - H&EThe outer part of the tube forming the intestines consists of two layers of smooth muscle - one circular layer and one longitudinal layer. If you look at the tissue close to the border between the two layers of smooth muscle, you will be able to see both longitudinally sectioned smooth muscle cells and transversely sectioned smooth muscle cells. The smooth muscle cells are much longer than their nuclei. Transversely sectioned smooth muscle cells may not have their nuclei in the plane of the section.Occasionally you will find small nerves between the two muscle layers, and, if you are lucky and/or patient, you will also see some very large nuclei in this region. These nuclei belong to peripheral nerve cells (ganglion cells of the myenteric plexus), which regulate the contraction of the muscle around the gastrointestinal tract.Draw a small area which contains both longitudinally sectioned and transversely sectioned smooth muscle at high magnification.
60

The only tissues which perhaps could be confused with smooth muscle are dense regular connective tissues and peripheral nerves. Both the number of nuclei and their shapes clearly distinguish smooth muscle from dense regular connective tissues. Nuclei are much more frequent and larger in smooth muscle, and they are very elongated if cut longitudinally. Peripheral nerves will be surrounded by a capsule of cells and connective tissue - the perineurium. The thickness of longitudinally cut nerve fibres is constant while smooth muscle cells are spindle shaped. Also, axon and nodes of Ranvier should be visible in peripheral nerves
Skeletal Muscle
Skeletal muscle consists of very long tubular cells, which are also called muscle fibres.The average length of skeletal muscle cells in humans is about 3 cm (sartorius muscle up to 30 cm, stapedius muscle only about 1 mm). Their diameters vary from 10 to 100 µm.
Skeletal muscle fibres contain many peripherally placed nuclei.Up to several hundred rather small nuclei with 1 or 2 nucleoli are located just beneath the plasma membrane.
Skeletal muscle fibres show in many preparations characteristic cross-striations. It is therefore also called striated muscle.
Skeletal muscle is innervated by the somatic nervous system. Skeletal muscle makes up the voluntary muscle.
Structure of skeletal muscle
Muscle fibres in skeletal muscle occur in bundles, fascicles, which make up the muscle. The muscle is surrounded by a layer of connective tissue, the epimysium, which is continuous with the muscle fascia. Connective tissue from the epimysium extends into the muscle to surround individual fascicles (perimysium). A delicate network of loose connective tissue composed of fine collagenous and reticular fibres (endomysium) is found between the muscle fibres of a fascicle. Finally, each muscle fibre is surrounded by a basement membrane.
Origin of skeletal muscle
The myoblasts of all skeletal muscle fibres originate from the paraxial mesoderm. Myoblasts undergo frequent divisions and coalesce with the formation of a multinucleated, syncytial muscle fibre or myotube. The nuclei of the myotube are still located centrally in the muscle fibre. In the course of the synthesis of the myofilaments/myofibrils, the nuclei are gradually displaced to the periphery of the cell.
61

Satellite cells are small cells which are closely apposed to muscle fibres within the basal lamina which surrounds the muscle fibre. Their nuclei are slightly darker than those of the muscle fibre. Satellite cells are believed to represent persistent myoblasts. They may regenerate muscle fibres in case of damage.
Suitable Slides
Sections of skeletal muscle, tongue or upper esophagus - H&E Tongue, Skeletal Muscle, human - H&ESkeletal muscle in the tongue is arranged in bundles which typically run at right angles to each other. Both longitudinally and transversely cut skeletal muscle fibres are present. In both section planes you can see that the nuclei are located in the periphery of the muscle fibre. Myofibrils may be visible as very fine dots in some of the transversely muscle fibres. Striations formed by the A- and I-Bands of the sarcomeres are visible in longitudinally cut fibres. Z-lines and H-bands can be identified in well-preserved tissue.Details of the sarcomeres stand out more clearly if you close the iris diaphragm of the microscope. Remember to open the diaphragm after you have seen the striations clearly !In the connective tissue between the muscle fibres, the endomysium, numerous capillaries supply the muscle with oxygen and nutrients.Draw a small section of longitudinal and transversely cut skeletal muscle at high magnification.
The muscle surrounding the upper one-third of the oesophagus is skeletal muscle. Smooth muscle surrounds its lower one-third. In section of the middle of the esophagus it is usually possible to identify both muscle types and their appearances can be compared.
62

The Contractile Apparatus of Skeletal Muscle
The spatial relation between the filaments that make up the myofibrils within skeletal muscle fibres is highly regular. This regular organisation of the myofilaments gives rise to the cross-striation, which characterises skeletal and cardiac muscle. Sets of individual "stria" correspond to the smallest contractile units of skeletal muscle, the sarcomeres. Rows of sarcomeres form the myofibrils (), which extend throughout the length of the skeletal muscle fibre.
Depending on the distribution and interconnection of myofilaments a number of "bands" and "lines" can be distinguished in the sarcomeres:
63

I-band - actin filaments,A-band - myosin filaments which may overlap with actin filaments,H-band - zone of myosin filaments only (no overlap with actin filaments) within the A-band,Z-line - zone of apposition of actin filaments belonging to two neighbouring sarcomeres (mediated by a protein called alpha-actinin),M-line - band of connections between myosin filaments (mediated by proteins, e.g. myomesin, M-protein).
The average length of a sarcomere is about 2.5 µm (contracted ~1.5 µm, stretched ~3 µm).
The protein titin extends from the Z-line to the M-line. It is attached to the Z-line and the myosin filaments. Titin has an elastic part which is located between the Z-line and the border between the I- and A-bands. Titin contributes to keeping the filaments of the contractile apparatus in alignment and to the passive stretch resistance of muscle fibres.Other cytoskeletal proteins interconnect the Z-lines of neighbouring myofibrils. Because of this connection, the A- and I-bands of neighbouring myofibrils lie side-by-side in the muscle fibre. These cytoskeletal proteins also connect the Z-lines of the peripheral myofibrils to the sarcolemma.
Muscle-Tendon Junction
At the muscle-tendon junction, the end of a muscle fibre forms deep invaginations, which increase its surface area. The basement membrane of the muscle fibre extends into these invagination and, so do the collagen fibrils of the tendons. The actin filaments of the last sarcomeres extend into cytoplasmic specialisations associated with zonula adherens-like membrane specialisations. Instead of interconnecting two cells, the cell membrane is here anchored to the basement membrane of the muscle cell. The basement membrane is, in turn, connected to the collagen fibrils of the tendons.
64

Excitation and Contraction of Skeletal Muscle
The area of contact between the end of a motor nerve and a skeletal muscle cell is called the motor end plate. Small branches of the motor nerve form contacts (boutons) with the muscle cell in a roughly eliptical area. The excitatory transmitter at the motor end plate is acetylcholine. The space between the boutons and the muscle fibre is called primary synaptic cleft. Numerous infoldings of the sarcolemma in the area of the motor end plate form secondary synaptic clefts. Motor end plates typically concentrate in a narrow zone close to the middle of the belly of a muscle. The excitable sarcolemma of skeletal muscle cells will allow the stimulus to spread, from this zone, over the entire muscle cell.
The spread of excitation over the sarcolemma is mediated by voltage-gated ion channels.
Invaginations of the sarcolemma form the T-tubule system which "leads" the excitation into the muscle fibre, close to the border between the A- and I-bands of the myofibrils. Here, the T-tubules are in close apposition with cisternae formed by the sarcoplasmatic reticulum. This association is called a triad. The narrow gap between the T-tubule and the cisternae of the sarcoplasmatic reticulum is spanned by proteins which mediate the excitation-contraction coupling.
Proteins in the sarcolemma which forms the wall of the T-tubule (dihydropyridine (DHP) receptors) change conformation, i.e. they change their shape, in response to the excitation travelling over the sarcolemma. These proteins are in touch with calcium channels (ryanodine receptors) which are embedded in the membrane of the cisternae of the sarcoplasmatic reticulum. The change in the shape of the proteins belonging to the T-tubule opens the calcium channels of the sarcoplasmatic reticulum. Calcium can now move from stores in the sarcoplasmatic reticulum into the cytoplasm surrounding the myofilaments.
Sites of interaction between actin and myosin are in resting muscle cells "hidden" by tropomyosin. Tropomyosin is kept in place by a complex of proteins collectively called troponin. The binding of calcium to troponin-C induces a conformational change in the troponin-tropomyosin complex which permits the interaction between myosin and actin and, as a consequence of this interaction, contraction.
ATP-dependent calcium pumps in the membrane of the sarcoplasmatic reticulum typically restore the concentration of Ca to resting levels within 30 milliseconds after the activation of the muscle fibre.
65

Types of Skeletal Muscle
Skeletal muscle cells respond to stimulation with a brief maximal contraction - they are of the twitch type. Individual muscles fibres cannot maintain their contraction over longer periods. The sustained contraction of a muscle depends on the "averaged" activity of often many muscles fibres, which individually only contract for a brief period of time.The force generated by the muscle fibre does depend on its state of contraction at the time of excitation. Excitation frequency and the mechanical summation of the force generated is one way to graduate the force generated by the entire muscle. Another way is the regulation of the number of muscle fibres which contract in the muscle. Additional motor units, i.e. groups of muscle fibres innervated by one motor neurone and its branches, are recruited if their force is required. The functional properties of the muscle can be "fine-tuned" further to the tasks the muscle performs by blending functionally different types of muscle fibres:
Type I fibres (red fibres)
Red muscles contain predominantly (but not exclusively) red muscle cells. Red muscle fibres are comparatively thin and contain large amounts of myoglobin and mitochondria. Red fibres contain an isoform of myosin with low ATPase activity, i.e. the speed with which myosin is able to use up ATP. Contraction is therefore slow. Red muscles are used when sustained production of force is necessary, e.g. in the control of posture.
Type II fibres
White muscle cells, which are predominantly found in white muscles, are thicker and contain less myoglobin. ATPase activity of the myosin isoform in white fibres is high, and contraction is fast. Type IIA fibres (red) contain many mitochondria and are available for both sustained activity and short-lasting, intense contractions. Type IIB/IIX fibres (white) contain only few mitochondria. They are recruited in the case of rapid accelerations and short lasting maximal contraction. Type IIB/IIX fibres rely on anaerobic glycolysis to generate the ATP needed for contraction.
Skeletal muscle fibres do not contract spontaneously. Skeletal muscle fibres are not interconnected via GAP junctions but depend on nervous stimulation for contraction. All muscle fibres of a motor unit are of the same type.
66

Fibre type is determined by the pattern of stimulation of the fibre, which, in turn, is determined by the type of neuron which innervates the muscle. If the stimulation pattern is changed experimentally, fibre type will change accordingly. This is of some clinical / pathological importance. Nerve fibres have the capacity to form new branches, i.e. to "sprout", and to re-innervate muscle fibres, which may have lost their innervation as a consequence of an acute lesion to the nerve or a neurodegenerative disorder. The type of the muscle fibre will change if the type of stimulation provided by the sprouting nerve fibre does not match with the type of muscle. The process of reinnervation and type adjustment may result in fibre type grouping within the muscle, i.e. large areas of the muscle are populated by muscle fibres of one type.
Muscle Spindles
Muscle spindles are sensory specialization of the muscular tissue. A number of small specialised intrafusal muscle fibres (nuclear bag fibres and nuclear chain fibres) are surrounded by a capsule of connective tissue. The intrafusal fibres are innervated by efferent motor nerve fibres. Afferent sensory nerve fibres surround the intrafusal muscle fibres.If the muscle is stretched, the muscle fibres in the muscle spindle are stretched, sensory nerves are stimulated, and a change in contraction of the muscle is perceived. Different types of intrafusal fibres and nerve endings allow the perception of position, velocity and acceleration of the contraction of the muscle.The contraction of the intrafusal fibres, after stimulation by the efferent nerve fibres, may counteract or magnify the changes imposed on the muscle spindle by the surrounding muscle. The intrafusal fibres and the efferent nerves can in this way set the sensitivity for the sensory nerve ending in the muscle spindle.
67

Cardiac Muscle
Cardiac muscle, the myocardium, consists of muscle cells, cardiomyocytes, with one centrally placed nucleus.Nuclei are oval, rather pale and located centrally in the muscle cell which is 10 - 15 µm wide.
Cardiac muscle cells exhibit cross-striations. Cardiac muscle cells excitation is mediated by rythmically active modified
cardiac muscle cells. Cardiac muscle is innervated by the autonomic nervous system, which adjusts the
force generated by the muscle cells and the frequency of the heart beat. Cardiac muscle is for these reasons also called involuntary striated muscle.
Structure of cardiac muscle
The ultrastructure of the contractile apparatus and the mechanism of contraction largely correspond to that seen in skeletal muscle cells. Although equal in ultrastructure to skeletal muscle, the cross-striations in cardiac muscle are less distinct, in part because rows of mitochondria and many lipid and glycogen droplets are found between myofibrils.
In contrast to skeletal muscle cells, cardiac muscle cells often branch at acute angles and are connected to each other by specialisations of the cell membrane in the region of the intercalated discs. Intercalated discs invariably occur at the ends of cardiac muscle cells in a region corresponding to the Z-line of the myofibrils (the last Z-line of the myofibril within the cell is "replaced" by the intercalated disk of the cell membrane). In the longitudinal part of the cell membrane, between the "steps" typically formed by the intercalated disk, we find extensive GAP junctions.
T-tubules are typically wider than in skeletal muscle, but there is only one T-tubule set for each sarcomere, which is located close to the Z-line. The associated sarcoplasmatic reticulum is organised somewhat simpler than in skeletal muscle. It does not form continuous cisternae but instead an irregular tubular network around the sarcomere with only small isolated dilations in association with the T-tubules.
Cardiac muscle does not contain cells equivalent to the satellite cells of skeletal muscle. Therefore cardiac muscle cannot regenerate.
68

Suitable Slides Sections of cardiac muscle - Alizarin Blue, Whipf's polychrome, iron haematoxylin, H&E
Cardiac Muscle, human - H&EUse a low magnification to find a part of the tissue in which the cardiac muscle cells are cut longitudinally. At high magnification you should see striations and the large nuclei of the cardiac muscle cells. If you follow the course of individual cardiac muscle cells you will note fine, darker lines which seem to cross the fibres. These are the intercalated discs which connect the individual muscle cells mechanically and permit the conduction of electrical impulses between the cells.
A light streak of cytoplasm is often visible extending from the poles of the nucleus. This part of the cytoplasm does not contain myofibrils, and it appears very light in transversely cut cardiac muscle cells. Myofibrils are often visible in transversely cut cells. Their visible separation reflects the large numbers of mitochondria located between them. Also, the large number capillaries reflect the need of a good blood supply to the constantly active muscle cells.
Draw longitudinally cut cardiac muscle cells which show all the features mentioned. Label the features in your drawing, and include an suitable scale.
Excitation in cardiac muscle
A number of specialised structures, which are composed of modified cardiac muscle cells, ensure that the contraction of the atria and ventricles takes place in the order that is most appropriate to the pumping function of the heart. The excitation of the myocardium originates from the sinuatrial node, which is located in the wall of the right atrium lateral to the opening of the superior vena cava into the atrium. The sinuatrial node initiates the contraction of atrial myocardium. Excitation also reaches the atrioventricular node at the base of the interatrial septum. The myocardium of the atria and ventricles are separated
69

from each other by a zone of connective tissue, the fibrous body of the heart. The fibrous body prevents the spread of excitation from atrial muscle cells to those of the ventricles.
A system of modified cardiac muscle cells, Purkinje fibres, has developed, which conduct stimuli faster than ordinary cardiac muscle cells (2-3 m/s vs. 0.6 m/s). A bundle of Purkinje fibres extends from the atrioventricular node, pierces the fibrous body, divides into left and right bundles, and travels, beneath the endocardium, towards the tip (apex) of the heart. Branches of the bundle contact ordinary cardiac muscle cells by way of specialisations similar to intercalated discs. Purkinje fibres contain large amounts of glycogen but fewer myofibrils than ordinary cardiac muscle cells. Myofibrils are mainly located in the periphery of the cell. Purkinje fibres are also thicker than ordinary cardiac muscle cells.
Modified muscle cells in nodal tissue (nodal muscle cells or P cells; P ~ pacemaker or pale-staining) of the heart exert the pacemaker function that drives the Purkinje cells. The rhythm generated by the nodal muscle cells can be modified by the autonomic nervous system, which innervates the nodal tissue and accelerates (sympathetic) or decelerates (parasympathetic) heart rate.
70

Suitable Slides Sections of cardiac muscle (interventricular septum) - Whipf's polychrome, iron haematoxylin, H&E
Purkinje Fibre, sheep - Whipf's polychromeCardiac muscle cells in this preparation have a red-violet appearance. Much of the connective tissue looks light blue, striations of cardiac muscle cells are visible. Intercalated discs may be more difficult to find, but nuclei stand out very clearly. Bundles of Purkinje fibres are present in areas of connective tissue between areas of "normal" cardiac muscle tissue and beneath the endocardium. Purkinje fibres appear as a chain of light blue profiles with a red rim. Browse through the tissue at low magnification and change to high magnification when you suspect the presence of Purkinje fibres. The red rim is formed by the contractile filaments. They are displaced to the periphery of the cells and can sometimes be used to define the outline of individual cells. The nuclei are large, but the cells are even larger and you will not see a nucleus in each cell.
71

VASCULAR SYSTEM
The cardiovascular system is concerned with the transport of blood and lymph through the body. It may be divided into four major components: the heart, the macrocirculation, the microcirculation and the lymph vascular system.
Essentially, the macrocirculation comprises all vessels, both arteries and veins, that would be visible to the eye. The vessels of the macrocirculation supply and drain a network of fine vessels interposed between them, the capillaries. This network is also called the capillary bed. Water and other components of the blood plasma which exude
72

from the blood vessels form the interstitial fluid, which is returned to the circulation by the lymph vascular system.
General Structure of Blood Vessels
You have already seen blood vessels of various sizes and types in preparations available in other lab sessions, and you should be aware that the histological appearances of vessels of different sizes (arterioles vs. arteries) and different types (arteries vs. veins) are different from each other. These differences are the result of quantitative variations of a common structural pattern that can be seen in all blood vessels with the exception of capillaries, i.e. the division of the walls of the blood vessels into three layers or tunics.
The tunica intima
delimits the vessel wall towards the lumen of the vessel and comprises its endothelial lining (typically simple, squamous) and associated connective tissue. Beneath the connective tissue, we find the internal elastic lamina, which delimits the tunica intima from
the tunica media.
The tunica media is formed by a layer of circumferential smooth muscle and variable amounts of connective tissue. A second layer of elastic fibers, the external elastic lamina, is located beneath the smooth muscle. It delimits the tunica media from
the tunica adventitia,
which consist mainly of connective tissue fibres. The tunica adventitia blends with the connective tissue surrounding the vessel. The definition of the outer limit of the tunica adventitia is therefore somewhat arbitrary.
Variations of Vessel Wall Structure
Arteries
All arterial vessels originate with either the pulmonary trunk (from the right ventricle) or the aorta (from the left ventricle). Specialisations of the walls of arteries relate mainly to two factors: the pressure pulses generated during contractions of the heart (systole) and
73

the regulation of blood supply to the target tissues of the arteries. The tunica media is the main site of histological specialisations in the walls of arteries.
Vessels close to the heart (aorta, pulmonary trunk and the larger arteries that originate from them) are
Elastic arteries
The tunica intima of elastic arteries is thicker than in other arteries. A layer of loose connective tissue beneath the endothelium (subendothelial connective tissue) allows the tunica intima to move independently from other layers as the elastic arteries distend with the increase in systolic blood pressure. Distension of the walls is facilitated by concentric fenestrated lamellae of elastic fibres in a thick tunica media. In adult humans, about 50 elastic lamellae are found in the tunica media of the aorta. The energy stored in the elastic fibres of the tunica media allows elastic arteries to function as a "pressure reservoir" which forwards blood during ventricular relaxation (diastole). Smooth muscle cells and collagen fibres are present between the layers of elastic fibres. Both fibre types are produced by the smooth muscle cells. Each elastic lamella forms together with interlamellar fibres and cells a lamellar unit. The external elastic lamina is difficult to discern from other layers of elastic fibres in the tunica media. The tunica adventitia appears thinner than the tunica media and contains collagen fibres and the cell types typically present in connective tissue.
The walls of these large arteries are so thick that their peripheral parts cannot derive enough oxygen and nutrients from the blood of the vessel that they form. Larger vessels are therefore accompanied by smaller blood vessels which supply the tunica adventitia and, in the largest vessels, the outer part of the tunica media of the vessel wall. The vessels are called vasa vasorum. In macroscopic preparations vasa vasorum are visible as fine dark lines on the surface of the larger arteries.
Suitable Slides sections of the aorta - H&E, elastin
Aorta, human - H&E , elastin & van GiesonThe thin endothelial lining of the aorta corresponds to that of other vessels. The flattened cells are easily damaged during preparation and it may be difficult to identify the endothelium. The subendothelial layer of connective tissue is characterised by a lower density of cells, i.e. fewer nuclei, a fibrous appearance of the tissue and the absence of well-defined elastic layers. Because the lamellae of elastic fibers diffract light differently from the remaining tissues they should also be visible in H&E stained sections. Elastic lamellae become visible in the tunica media. The majority of cells in the tunica media are smooth muscle cells.
74

Smooth muscle cells and collagen fibres are found between the layers of elastic fibres. If you scan the periphery of the aorta you may find small blood vessels, the vasa vasorum, in the tunica adventitia and penetrating into the outer part of the tunica media.
The diameter of individual arteries decreases as we follow them further into the periphery. However, their total diameter increases, which leads to a fall in blood pressure. Also, the properties of the elastic arteries have to some extent evened out differences in diastolic and systolic blood pressure. The amount of elastic fibres in the tunica media decreases with these physiological changes. We now find a type of arteries which are termed
Muscular arteries
The tunica intima is thinner than in elastic arteries. Subendothelial connective tissue other than the internal elastic lamina is often difficult to discern. The internal elastic lamina forms a well defined layer. The tunica media is dominated by numerous concentric layers of smooth muscle cells. Fine elastic fibres and and a few collagen fibres are also present. The external elastic lamina can be clearly distinguished although it may be incomplete in places. The thickness and appearance of the tunica adventitia is variable.
75

The basic structure of the walls of arteries does not change much as we come to the next type of arterial vessels. Size is used to differentiate them from muscular arteries.
Arterioles
are arterial vessels with a diameter below 0.1 - 0.5 mm (different values in different textbooks). Endothelial cells are smaller than in larger arteries, and the nucleus and surrounding cytoplasm may 'bulge' slightly into the lumen of the arteriole. The endothelium still rests on a internal elastic lamina, which may be incomplete and which is not always well-defined in histological sections. The tunica media consists of 1-3 concentric layers of smooth muscle cells. It is difficult to identify an external elastic lamina or to distinguish the tunica adventitia from the connective tissue surrounding the vessel.The smooth muscle of arterioles and, to some extent, the smooth muscle of small muscular arteries regulate the blood flow to their target tissues. Arterioles receive both sympathetic and parasympathetic innervation. The final branching of the arterioles finally gives rise to the capillary network (microcirculation).
Suitable Slides
76

sections of arteries - H&E or elastin (in combination with other stains)Sections of small muscular arteries and arterioles are present in many sections, and the basic features of their structure are usually visible - even in smaller arteries. Large muscular arteries often have their "own section" in teaching collections.
Artery - H&E and elastin & eosinIdentifying muscular arteries in sections is rather straight forward. There are two easily recognizable features which distinguish these arteries from veins. If two vessels have a similarly sized lumen, the walls of arteries will be much thicker and more compact than the wall of veins. At high magnification, the internal elastic lamina forms a pink streak immediately below the endothelial cell lining in arteries and even arterioles, while it is difficult to identify in veins.The layer of subendothelial connective tissue is very thin, and the endothelium seems to rest on the internal elastic lamina. Smooth muscle cell nuclei are frequent in the tunica media. The external elastic lamina stains similar to the internal elastic lamina, but it is thicker and appears fibrous instead of forming a continuous band. Collagen fibres and a few connective tissue cell nuclei are visible in the tunica adventitia.If you close the iris diaphragm of the microscope, the elastic layers will stand out very clearly, but remember to open the diaphragm once you have seen them.
77

In addition to the inner and outer elastic laminae, elastin stains will show fine elastic fibres in the tunica media and coarse elastic fibres between the collagen fibres of the tunica adventitia. The appearance of other structures will depend on the stain used together with the elastin stain. Eosin, the E in H&E, gives a pink colour to both collagen fibres and the cytoplasm of cells. Nuclei are not stained if the H is omitted from the H&E.Draw either one large composite image containing the three tunics and the cellular and fibrous elements which form the tunics.Alternatively, you can draw a low power overview and supplement it with high magnification illustrations of the individual tunics. Focus on an H&E stained section.
Capillaries
The sum of the diameters of all capillaries is significantly larger than that of the aorta (by about three orders of magnitude), which results in decreases in blood pressure and flow rate. Also, capillaries are very small vessels. Their diameter ranges from 4-15 µm. The wall of a segment of capillary may be formed by a single endothelial cell. This results in a very large surface to volume ratio. The low rate of blood flow and large surface area facilitate the functions of capillaries in
providing nutrients and oxygen to the surrounding tissue, in the absorption of nutrients, waste products and carbon dioxide, and in the excretion of waste products from the body.
These functions are also facilitated by a very simple organisation of the wall of capillaries. Only the tunica intima is present, which typically only consists of the endothelium, its basal lamina and an incomplete layer of cells surrounding the capillary, the pericytes. Pericytes have contractile properties and can regulate blood flow in capillaries. In the course of vascular remodelling and repair, they can also differentiate into endothelial and smooth muscle cells.
78

Three types of capillaries can be distinguished based on features of ethe endothelium.
Continuous capillaries
are formed by "continuous" endothelial cells and basal lamina. The endothelial cell and the basal lamina do not form openings, which would allow substances to pass the capillary wall without passing through both the endothelial cell and the basal lamina. Both endothelial cells and the basal lamina can act as selective filters in continuous capillaries.
Fenestrated capillaries
The endothelial cell body forms small openings called fenestrations, which
79

allow components of the blood and interstitial fluid to bypass the endothelial cells on their way to or from the tissue surrounding the capillary. The fenestrations may represent or arise from pinocytotic vesicles which open onto both the luminal and basal surfaces of the cell. The extent of the fenestration may depend on the physiological state of the surrounding tissue, i.e. fenestration may increase or decrease as a function of the need to absorb or secrete. The endothelial cells are surrounded by a continuous basal lamina, which can act as a selective filter.
Discontinuous capillaries
are formed by fenestrated endothelial cells, which may not even form a complete layer of cells. The basal lamina is also incomplete. Discontinuous capillaries form large irregularly shaped vessels, sinusoids or sinusoid capillaries. They are found where a very free exchange of substances or even cells between bloodstream and organ is advantageous (e.g. in the liver, spleen, and red bone marrow).
Suitable Slides Sections of any well preserved tissue - H&E, Whipf's polychromecardiac and skeletal muscle, glands or the papillary layer of the skin contain dense capillary beds.
80

Cardiac Muscle, sheep - Whipf's polychromeLarge numbers of capillaries are present in almost all tissues. At least a few dozen cross sections are present in every sqr. mm of section of poorly vascularised tissues. There may be thousands in highly vascularised tissues. However, a "good" capillary is not that easy to find because of their small size and because the capillary walls are very thin, which often leads to the collapse of the capillary during tissue preparation.Cardiac muscle is highly vascularised. Each muscle cell is surrounded by one or more capillaries. The capillaries roughly follow the course of the muscle cells. To find capillaries in transverse and longitudinal sections it is easiest to first find areas in which the muscle cells have been cut in these planes. Only one or two red blood cells fit side by side in the capillary. A single endothelial cell forms the wall around the entire circumference of a segment of the capillary. Endothelial cell nuclei are therefore not always visible, and some red blood cells are only surrounded by a fine line representing the capillary wall.Identify and draw a few capillaries. Include some of the surrounding tissue features - maybe a cardiac muscle cell, a venule or arteriole - as a scale.
Veins
The walls of veins are thinner than the walls of arteries, while their diameter is larger. In contrast to arteries, the layering in the wall of veins is not very distinct. The tunica intima is very thin. Only the largest veins contain an appreciable amount of subendothelial connective tissue. Internal and external elastic laminae are absent or very thin. The tunica media appears thinner than the tunica adventitia, and the two layers tend to blend into each other. The appearance of the wall of veins also depends on their location. The walls of veins in the lower parts of the body are typically thicker than those of the upper parts of the body, and the walls of veins which are embedded in tissues that may provide some structural support are thinner than the walls of unsupported veins.
Venous vessels originate from the capillary network which coalesce into the smallest venous vessels the
Venules.
81

They are larger than capillaries. Small venules are surrounded by pericytes. A few smooth muscle cells may surround larger venules. The venules merge to form
Small to medium-sized veins
which contain bands of smooth muscle in the tunica media. The tunica adventitia is well developed. In some veins (e.g. the veins of the pampiniform plexus in the spermatic cord) the tunica adventitia contains longitudinally oriented bundles of smooth muscle.Aside from most veins in the head and neck, small to medium-sized veins are also characterised by the presence of valves. The valves are formed by loose, pocket-shaped folds of the tunica intima, which extend into the lumen of the vein. The opening of the pocket will point into the direction of blood flow towards the heart. One to three (usually two) pockets form the valve. Blood flowing towards heart will pass the pockets. If the flow reverses, blood will fill the pockets which will occlude the lumen of the vein and prevent the return of blood into the part of the vein preceding the valve. The ability of the valves to prevent backflow depends to some extent on the state of contraction (tone) of the smooth muscle in the wall of the vein.
The largest veins of the abdomen and thorax
do contain some subendothelial connective tissue in the tunica intima, but both it and the tunica media are still comparatively thin. Collagen and elastic fibres are present in the tunica media. The tunica adventitia is very wide, and it usually contains bundles of longitudinal smooth muscle. The transition from the tunica adventitia to the surrounding connective tissue is gradual. Valves are absent.Vasa vasorum are more frequent in the walls of large veins than in that of the corresponding arteries - probably because of the lower oxygen tension in the blood contained within them.
Suitable Slides
sections of veins - H&E, van Gieson or elastin (in combination with other stains)Like arteries, veins and venules are present in many sections. The basic features of their structure are however more difficult to identify because of the thin walls of veins - in particular in small veins. It is best to resort to sections of large veins, which together with large muscular arteries often have their "own section" in teaching collections.
82

Vein, human - H&EThe tunica intima is very narrow and the internal elastic lamina is difficult to identify - even in elastin stained sections. A few elastic fibres below the endothelium form only a very thin and incomplete internal elastic lamina. Smooth muscle is present in the tunica media, but it is organised less regular than in the artery. The tunica media is, again as compared to the artery, very thin and there is no sharp border between the tunica media and the tunica adventitia. The tunica adventitia of the largest veins contains coarse collagen fibres, elastic fibres and longitudinal bundles of smooth muscle. Small and medium sized veins will not contain smooth muscle in the tunica adventitia.Draw part of the wall of the vein, label the tunics and indicate the presence of smooth muscle, fibres and their types.
Suitable Slides
sections containing small to medium sized veins - H&E, van Gieson or trichromeUnless a specifically prepared slide is available, I would recommend looking at skin slides, in which I have found quite a few nice valves in the veins located at the border between dermis and hypodermis.
83

Vein Valve - H&E Unless the section has been specifically prepared to illustrate valves, you will have to search for a while - probably through several sections. If a vein with valves is present in the section, it should be easy to identify. Valves are only found in small to medium-sized veins. You should see one or two bands of tissue in the lumen of the vein. Each band is formed by two apposing layers of tunica intima. The bands may share their origin from the inner aspect of the wall of the vein or they may have separate origins. Folding of the tissue bands forming the valves is variable.Draw the vein and valve and label the tunics which you can identify. Try to indicate the position of the section in the schematic drawing of the valve.You can observe the function of vein valves. Put your finger firmly on one of the veins which are visible on the back of your hand. Use another finger to stroke over the vein in the direction of the wrist. The vein will empty and not backfill (closed valves!) until you release the first finger.
Additional Specialisations of Vessels
Small arteries and veins often form anastomosing networks, which provides routes for alternative blood supply and drainage if one of the vessels should become occluded because of pathological or normal physiological circumstances. Some arteries are however the only supply of blood to their target tissues. These arteries are call end arteries. Tissues which are supplied by end arteries die if the arteries become occluded.
The segments of the kidney and the heads of the gastrocnemius muscle are examples of tissues supplied by end arteries.
Arteries and veins may also form arteriovenous shunts, which can shunt the blood flow that otherwise would enter the capillary network between the vessels. These shunts usually contain specialisations of the smooth muscle in the region of the shunt. Arteriovenous shunts are frequently seen in the blood supply of distal parts of the limbs and the nose (thermoregulation) and in the blood supply of endocrine organs.
84
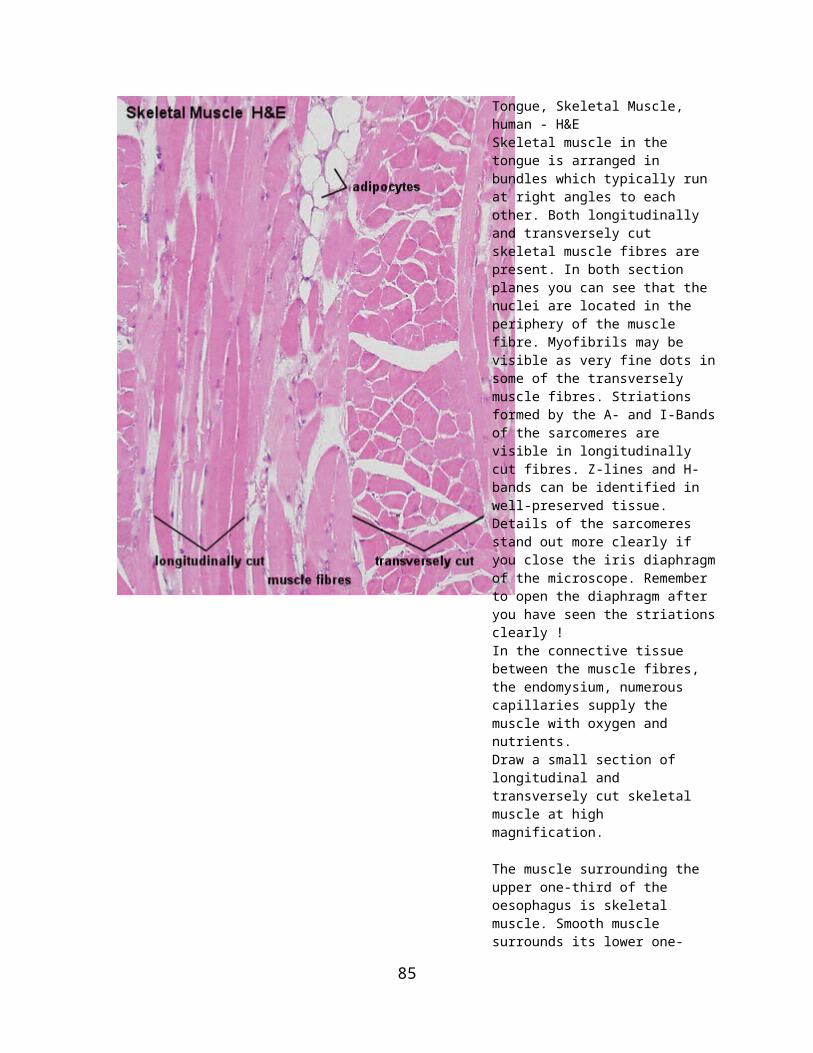
Lymphatic Vessels
Parts of the blood plasma will exude from the blood vessels into the surrounding tissues because of transport across the endothelium or because of blood pressure and the fenestration of some capillaries (this process is partly counteracted by the higher osmotic pressure of the blood). The fluid entering tissues from capillaries adds to the interstitial fluid normally found in the tissue. The surplus of liquid needs to be returned to the circulation. Lymph vessels are dedicated to this unidirectional flow of liquid, the lymph. Three types of lymph vessels can be distinguished based on their size and morphology.
Lymph capillaries
are somewhat larger than blood capillaries and very irregularly shaped. They begin as blind-ending tubes in connective tissue. The basal lamina is almost completely absent and the endothelial cells do not form tight junctions, which facilitates the entry of liquids into the lymph capillary. Temporary openings in the endothelial lining of the lymph capillaries also allow the entry of larger particles into the lymph capillaries (lipid droplets, which are absorbed from the lumen of the gut do not enter blood capillaries, but enter the circulation via lymph vessels which are found in the villi of the ileum and jejunum). Lymph capillaries merge to form
Lymph collecting vessels
which are larger and form valves but otherwise appear similar to lymph capillaries. The lymph is moved by the compression of the lymph vessels by surrounding tissues. The direction of lymph flow is determined by the valves. Lymph vessels empty intermittently into lymph nodes from which the lymph continues in efferent lymph vessels.
Only very little lymph is returned from the limbs if they are immobilized, which illustrates the importance of muscular action in lymph transport. This is also the reason for immobilizing limbs that are either infected or that have been bitten by venomous Australians. The effect can also be observed after long intercontinental flight when you may feel that your shoes and socks are just about one number too small. Finally, impeded lymph drainage is one of the problems associated with surgery which requires the removal of lymph nodes and which thereby interrupts the lymph collecting vessels.
Eventually the lymph collecting vessels merge to form
85

Lymph ducts
which contain one or two layers of smooth muscle cells in their wall (some textbooks call this layer the tunica media of lymph vessels). They also form valves. The walls of the lymph ducts are less flexible in the region of the attachment of the valves to the wall of the duct, which may give a beaded appearance to the lymph ducts. Peristaltic contractions of the smooth muscle contribute to the movement of lymph towards the heart in addition to the compression of the ducts by surrounding tissues.
The largest lymph duct of the body, the thoracic duct, drains lymph from the lower half and upper left quadrant of the body and empties the lymph into the circulation by merging with the vascular system close to the junction of the left internal jugular and subclavian veins. That it is the largest lymph duct does not mean that it is a large vessel when compared to the large arteries and veins. It actually is not much larger (about 5mm in diameter) than one of the superficial forearm veins.
Suitable Slides
Sections of small intestine - H&E Jejunum, baboon - H&EAt low magnification you will see villi extending into the lumen of the jejunum. Find villi which are cut longitudinally and change to medium magnification. In some of the villi you will see fairly large open spaces, which are surrounded by a layer of flattened endothelial cells. They should not contain any red blood cells - if they do you are looking at a capillary, which also should be somewhat smaller. These openings represent the blind end of lymph capillaries which originate in the villi.Their name, lacteals, is derived from the milky appearance of the lymph. This appearance is caused by suspended lipid droplets which enter these lymph capillaries.
Draw a villus containing a lymph capillary. Focus on the features of the lymph capillary and the surrounding connective tissue. Include, if possible, a blood capillary in your drawing.
The jejunum slide is also good for revision. You should be able to find columnar epithelium, goblet cells, smooth muscle, small nerves, a few ganglion cells and, of course, lots of loose connective tissue and blood vessels.
86

BLOOD
Blood is sometimes considered to be a fluid connective tissue because of the mesenchymal origin of its cells and a low ratio of cells to liquid intercellular substance, the blood plasma. In human adults about 5 liter of blood contribute 7-8 % to the body weight of the individual. The contribution of red blood cells (erythrocytes) to the total volume of the blood (haematocrit) is about 43%.
Erythrocytes are the dominant (99%) but not the only type of cells in the blood. We also find leukocytes and, in addition, blood platelets. Erythrocytes, leukocytes and blood platelets are also being referred to as the formed elements of the blood. Erythrocytes and blood platelets perform their functions exclusively in the blood stream. In contrast, leukocytes reside only temporarily in the blood. Leukocytes can leave the blood stream through the walls of capillaries and venules and enter either connective or lymphoid tissues.
Erythrocytes
Erythrocytes do not contain a nucleus. They do contain haemoglobin, which fills almost the entire cytoplasm. Erythrocytes are unable to move actively, but they are remarkably elastic and can withstand deformation. They are typically biconcave disks although their shape is influenced by osmotic forces. The average diameter of the disk is ~7 µm. Since erythrocytes can be found in the vast majority of histological sections - in small numbers even in perfused tissues - they will often allow us to estimate the size of other structures or cells. Mature erythrocytes do not contain organelles, and their cytoplasm looks fairly homogenous - even in the EM.
At high magnification some granularity may be visible in EM images. The granular appearance is caused by haemoglobin molecules. Foetal erythrocytes (up to the 4th month of gestation) are larger than "adult" erythrocytes, and they are nucleated. The later feature they share with erythrocytes of other animal classes (e.g. amphibia and birds).
Functions
Erythrocytes function in the transport of oxygen. Haemoglobin, the oxygen binding protein in erythrocytes, contributes about 30% of the weight of an erythrocyte. The lifespan of an erythrocyte in the bloodstream is 100-120 days.
About 5×1011 erythrocytes are formed/destroyed each day.
87

Suitable Slides blood smear - Leishman, Wright's, Giemsa or May-Grünwald-Giemsa stains
Where to look for cells in a blood smearThe density of cells varies across the smear. Cells will be "heaped and piled" close to the point were the drop of blood was placed on the slide. White blood cells appear shrunken, and some types are difficult to distinguish from each other. There are fewer cells close to the tip of the smear. In this region, white blood cells are sometimes damaged and erythrocytes may be deformed. The best area to look at is between these two regions. Where it is located exactly and how wide it is will depend on the smear, but the middle of the smear is a good starting point.
Blood Smear, human - Leishman stain In a suitable region of the blood smear, the erythrocytes are rarely forming clumps or rows. Instead, they are more or less evenly spaced and occasionally forming goups of maybe 2 or 3 cells. Due to the biconcave shape of the erythrocytes, their center will look lighter than their periphery. Depending on the way in which the blood smear was prepared, blood platelets may be found singly, in small groups or large clumps.How does the shape of the erythrocyte facilitate its function? How would you expect an erythrocyte to look like if it is in an extracellular fluid of very low or very high osmotic pressure?Identify and draw a few erythrocytes.
It is a good idea to do one large composite drawing for all types of blood cells.
88

Leukocytes
Leukocytes can be further subdivided into granular leukocytes, i.e. neutrophils, basophils and eosiniphils, and non-granular leukocytes, i.e. monocytes and lymphocytes.
In healthy individuals the relative numbers of circulating leukocyte types are quite stable. A differential leukocyte count would typically produce the following cell frequencies (numbers in parentheses are the range of normal frequencies reported in different texts):
~ 60% neutrophils (50% - 70%) ~ 3% eosinophils (>0% - 5%) ~ 0.5% basophils (>0% - 2%) ~ 5% monocytes (1% - 9%) ~ 30% lymphocytes (20% - 40%)
Changes in their relative numbers indicate that something abnormal is happening in the organism. A larger than usual number of neutrophils (neutrophilia) would indicate e.g. an acute or chronic infection. The number of basophils and eosinophils may increase (eosinophilia or basophilia) as a consequence of e.g. allergic disorders.
Granular Leukocytes
Granular leukocytes are all approximately the same size - ~ 12-15 µm in diameter. Their nuclei form lobes, and nucleoli cannot be seen. The number of nuclear lobes varies according to cell type. All granulocytes are motile.
The term granulocytes refers to the presence of granules in the cytoplasm of these cells. The granules correspond to secretory vesicles and lysosomes. Specific granules are the granules which are only found in one particular type of granulocytes.
Neutrophil granulocytes (or neutrophils)
have a very characteristic nucleus. It is divided into 3-5 lobes which are connected by thin strands of chromatin. The number of lobes increases with cell age. Up to 7 lobes can be found in very old neutrophils (hypersegmented cells).
Neutrophils (like all other granulocytes, monocytes and lymphocytes) contain all the organelles that make up a typical cell. In addition to the usual complement of organelles, they also contain two types of granules. Primary granules (or A
89

granules) contain lysosomal enzymes and are likely to be primary lysosomes, although they are larger (0.4 µm) than the "ordinary" primary lysosome. Secondary granules (or B granules), the specific granules of the neutrophils, contain enzymes with strong bactericidal actions. The specific granules of neutrophils stain only weakly if they are at all visible - they are "neutral", hence the term neutrophil.
Functions
Neutrophils play a central role in inflammatory processes. Large numbers invade sites of infection in response to factors (e.g. cytokines) released by cells which reside at an infection site. Neutrophils are the first wave of cells invading infection sires. Receptors in their plama membrane allow them to recognise foreign bodies, e.g. bacteria, and tissue debris, which they begin to phagocytose and destroy. The phagocytotic activity of neurophils is further stimulated if invading microorganisms are "tagged" with antibodies (or opsonised). Neutrophils cannot replenish their store of granules. The cells die once their supply of granules has been exhausted. Dead neutrophils and tissue debris are the major components of pus. Their lifespan is only about one week.
Lost neutrophils are quickly replenished from a reserve population in the bone marrow. Because they are younger, their nuclei have fewer lobes than the "average" neutrophil. A high proportion of neutrophils, with few nuclear lobes indicates a recent surge in their release from the bone marrow.
Suitable Slides
90

Blood Smear, human - Leishman stainNeutrophil granulocytes are easy to find. They are the most frequent type of white blood cells, and the complex shape of their nucleus identifies them unequivocally. In darkly stained smears it is possible to see some faintly purple, very small granules in the cytoplasm. These granules represent the primary, lysosom-like granules of neutrophils. Have a close look at the nucleus of a number of neutrophils, and make a qualified guess at the gender of the individual, which donated blood for the slides.Identify and draw one or two neutrophil granulocytes.
How the neutrophils and other leukocytes exactly will look like depends somewhat on how the stain turned out. In some batches (labeled B2 in the images) nuclei are dark and crisp, and the cytoplasm is well demarcated. In other batches (labeled B1 in the images) nuclei and cytoplasm are lighter and their boundaries are less well defined. While the morphology appears clearer in the darker stained smears, it will usually be more difficult to identify eosinophils and basophils.
Eosinophil granulocytes (or eosinophils)
Their nucleus usually has only two lobes. Almost all of the cytoplasm appears filled with the specific granules of the eosinophils. As the term "eosinophil" indicates, these granules are not neutral but stain red or pink when eosin or a similar dye is used in the staining process. Aside from the usual complement of organelles eosinophils contain some large rounded vesicles (up to 1 µm in diameter) in their cytoplasm. These granules correspond to the eosinophilic grains that we see in the light microscope. These specific granules contain, in addition to enzymes that otherwise are found in lysosomes, an electron-dense, proteinaceous crystal. This crystal is composed of major basic protein (MBP).
Functions
The presence of antibody-antigen complexes stimulates the immune system. Eosinophils phagocytose these complexes, and this may prevent the immune system from "overreacting". Their granules also contain the enzymes histaminase and arylsufatase. These enzymes break down histamine and leukotrienes, which again may dampen the effects of their release by basophils or mast cells. MBP, which can also function as a cytotoxin, and its release by eosinophils may be involved in the response of the body against parasitic infections, which are accompanied by an increase in the number of eosinophils.
91

Suitable Slides
Blood Smear, human - Leishman stainEosinophils and basophils are the only cell types present in normal blood which initially may be difficult to distinguish - in particular in darker smears. If you see them side by side in your drawing, the difference between them should become apparent. Chances are 6:1 that the you find an eosinophil before you find a basophil. The two lobes of the nucleus of eosinophils are usually well-defined and of about equal size. The nucleus is embedded in a cytoplasm crowded with granules, which seem to form a solid mass in the cell. The 2-3 nuclear lobes of basophils are not as well defined as those of eosinophils, granules are not as numerous as in eosinophils, and pretty much all of them can be identified "as individuals" rather than the dense mass they form in eosinophils.Note that eosinophils and basophils are much easier to distinguish in B1. In B2, the difference in the staining of their of granules is not as pronounced, and the nuclei do not stand out as clear as in B1.Identify, draw and label an eosinophil and a basophil.
92
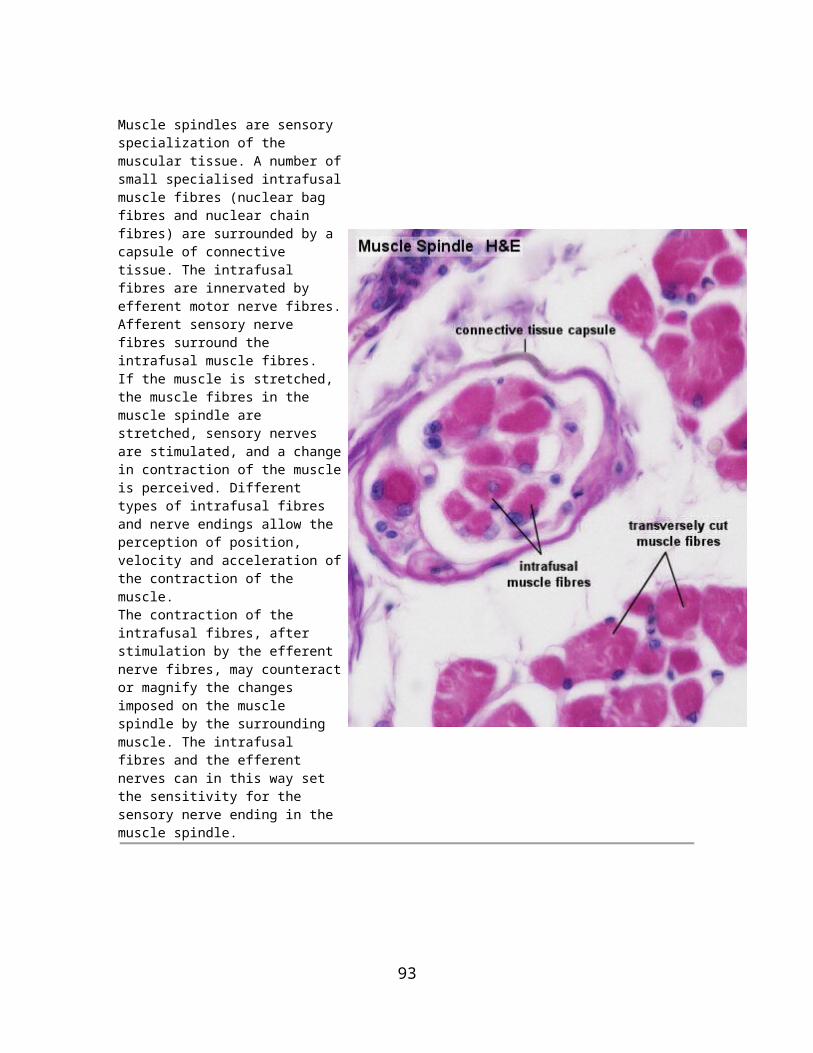
Basophil granulocytes (or basophils)
Basophilic granulocytes have a 2 or 3 lobed nucleus. The lobes are usually not as well defined as in neutrophilic granulocytes and the nucleus may appear S-shaped. The specific granules of basophils are stained deeply bluish or reddish-violet. In some smears, their colour corresponds closely to the colour of the nucleus which sometimes is difficult to distinguish amongst or behind the granules. The granules are not as numerous as those in eosinophils. The specific granules of basophils (~ 0.5 µm in diameter) appear quite dark in EM pictures. They contain heparin, histamine, lysosomal enzymes and leukotrienes. The later correspond to the slow-reacting substance of anaphylaxis (SRS-A).
93

Functions
Heparin and histamine are vasoactive substances. They dilate the blood vessels, make vessel walls more permeable and prevent blood coagulation. As a consequence, they facilitate the access of other lymphocytes and of plasma-borne substances of importance for the immune response (e.g. antibodies) to, e.g., a site of infection. The release of the contents of the granules of basophils is receptor-mediated. Antibodies produced by plasma cells (activated B-lymphocytes) bind to Fc-receptors on the plasma membrane of basophils. If these antibodies come into contact with their antigens, they induce the release of the contents of all of the basophil granules.
Non-granular leukocytes
Monocytes
These cells can be slightly larger than granulocytes (~ 12-18 µm in diameter). Their cytoplasm stains usually somewhat stronger than that of granulocytes, but it does not contain any structures which would be visible in the light microscope using most traditional stains (a few very fine bluish gains may be visible in some monocytes). The "textbook" monocyte has a C-shaped nucleus. Monocytes contain granules (visible in the EM) which in appearance and content correspond to the primary granules of neutrophils, i.e. the granules correspond to lysosomes.
Functions
Once monocytes enter the connective tissue they differentiate into macrophages. At sites of infection macrophages are the dominant cell type after the death of the invading neutrophils. The phagocytose microorganisms, tissue debris and the dead neutrophils. Monocytes also give rise to osteoclasts, which are able to dissolve bone. Osteoclasts are of importance in bone remodelling.
Suitable Slides
Blood Smear, human - Leishman stainMonocytes and lymphocytes definitely look much prettier in darker stained smears (B2) than in lighter ones (B1) - mainly because of a clearer distinction between cytoplasm and nucleus. The cell is very likely to be a lymphocyte if the nucleus is round and surrounded by a narrow rim of cytoplasm (B1 and left in B2). Large lymphocytes with a wider rim of cytoplasm do occur (right in B2), but not often. The C-shaped nucleus of a textbook monocyte may not be easy to find. The nuclei will vary from a peanut- to a "fat" S-shape in smears. Whatever the shape of the nucleus, it is usually not lobed and not round. The nucleus is bound, at least on its concave side, by a wide rim of non-granular cytoplasm. Note also the light area of monocyte cytoplasm, which is often visible close to the concave surface of the nucleus. A large Golgi apparatus is
94

located in the area. The Golgi apparatus does not stain as well as the remainder of the cytoplasm and leaves a light "impression" - the phenomenon is also called a "negative image".Identify and draw lymphocytes and a monocyte.
Lymphocytes
These cells are very variable in size. The smallest lymphocytes may be smaller than erythrocytes (down to ~ 5 µm in diameter) while the largest may reach the size of large granulocytes (up to 15 µm in diameter). How much cytoplasm is discernible depends very much on the size of the lymphocyte. In small ones, which are the majority of lymphocytes in the blood, the nucleus may appear to fill the entire cell. Large lymphocytes have a wider rim of cytoplasm which surrounds the nucleus. Both the nucleus and the cytoplasm stain blue (and darker than in most other cell types in the blood). The typical lymphocyte only contains the
95
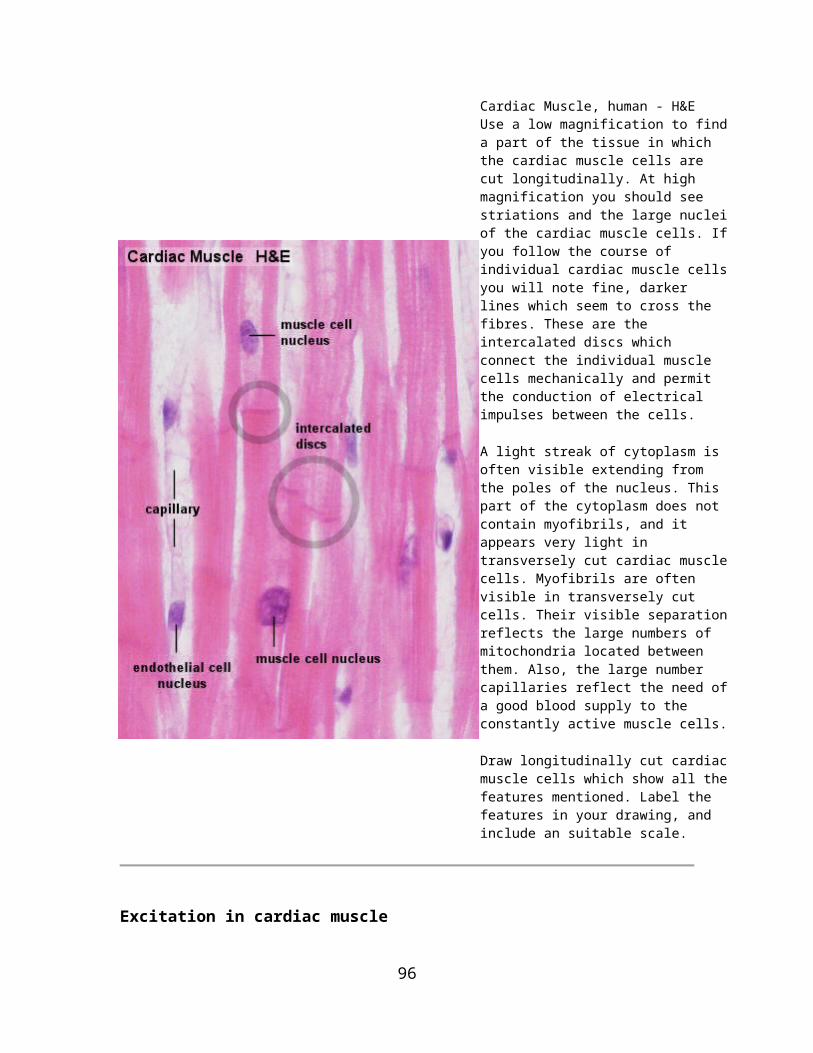
usual complement of cellular organelles. The appearance of lymphocytes may change drastically when they are activated (see below).
Functions
Most lymphocytes in the blood stream belong to either the group of B-lymphocytes (~5%) or the group of T-lymphocytes (~90%). Unless they become activated, the two groups cannot easily be distinguished using routine light or electron microscopy.Upon exposure to antigens by antigen-presenting cells (e.g. macrophages) and T-helper cells (one special group of T-lymphocytes) B-lymphocytes differentiate into antibody producing plasma cells. The amount of cytoplasm increases and RER fills a large portion of the cytoplama of plama cells. T-lymphocytes represent the "cellular arm" of the immune response (cytotoxic T cells) and may attack foreign cells, cancer cells and cells infected by, e.g., a virus.
T-lymphocytes and B-lymphocytes form the vast majority of lymphocytes in the blood stream, but they do not add up to 100%, and they usually are small lymphocytes. The much less frequent medium-sized or large lymphocytes may represent e.g.
natural killer (Nk-) cells which belong to the group of large granular lymphocytes, or
haemopoietic stem cells of which a very few will be circulating in the blood stream.
Blood Platelets (or thrombocytes)
Blood platelets do not contain a nucleus. Unlike erythrocytes, which also lack a nucleus, the blood platelets of mammals have never been nucleated cells. Instead, blood platelets are fragments of the cytoplasm of very large thrombocyte precursor cells, the megakaryocytes. Like other cells involved in the formation in blood cells, megakaryocytes are found in the bone marrow.
Platelets are about 3 µm long but appear somewhat smaller in the microscope. This is because their cytoplasm is divided into two zones: and outer hyalomere,
96

which hardly stains, and an inner granulomere, which contains bluish staining granules. These granules are usually not individually visible with the highest magnification on your microscope, and the granulomere appears more or less homogeneously blue. In addition to different types of vesicles (i.e. the granules), mitochondria, ribosomes, lysosomes and a little ER are present in the thrombocyte granulomere. Different types of vesicles contain either serotonin (electron-dense delta granules; few) or compounds important for blood coagulation (alpha granules; they also contain platelet-derived growth factor (PDGF) which may play a role in the repair of damaged tissue). The hyalomere contains cytoskeletal fibres, which include actin and myosin.
Functions
Platelets assist in haemostasis, the arrest of bleeding. Serotonin is a potent vasoconstrictor. The release of serotonin from thrombocytes, which adhere to the walls of a damaged vessels, is sufficient to close even small arteries. Platelets, which come into contact with collagenous fibers in the walls of the vessel (which are not usually exposed to the blood stream), swell, become "sticky" and activate other platelets to undergo the same transformation. This cascade of events results in the formation of a platelet plug (or platelet thrombus). Finally, activating substances are released from the damaged vessel walls and from the platelets. These substances mediate the conversion of the plasma protein prothrombin into thrombin. Thrombin catalyzes the conversion of fibrinogen into fibrin, which polymerizes into fibrils and forms a fibrous net in the arising blood clot. Platelets captured in the fibrin net contract leading to clot retraction, which further assists in haemostasis.
Blood coagulation is a fairly complex process, which involves a large number of other proteins and messenger substances. Deficiencies in any one of them, either inherited or acquired, will lead to an impairment of haemostasis.
Suitable Slides blood platelets (thrombocytes)megakaryoblasts and megakaryocytes
97

Blood Smear, human - Leishman stainIn lightly stained smears (B1), blood platelets will appear like light blue, fairly ill-defined specks between the other blood cells. In darker smears (B2), you will sometimes be able to see that the blue specks are formed by an accumulation of small bluish grains, the granules of the blood platelets.Identify and include a platelet in one of your other drawings.
Red Bone Marrow, rabbit - H&EThe marrow cavity of this bone is filled with red bone marrow. H&E is not the method of choice for looking at haemopoietic cells, but a few of the numerous named types or broader groups can actually be recognized.Precursors of platelets are the haemopoietic cells easiest to find in red bone marrow. The very dark and large megakaryoblast and the even larger but light megakaryocytes are clearly visible even at low magnifications. Note that adipocytes are present also in the red bone marrow.Identify and draw a megakaryocyte and megakaryoblast.
98
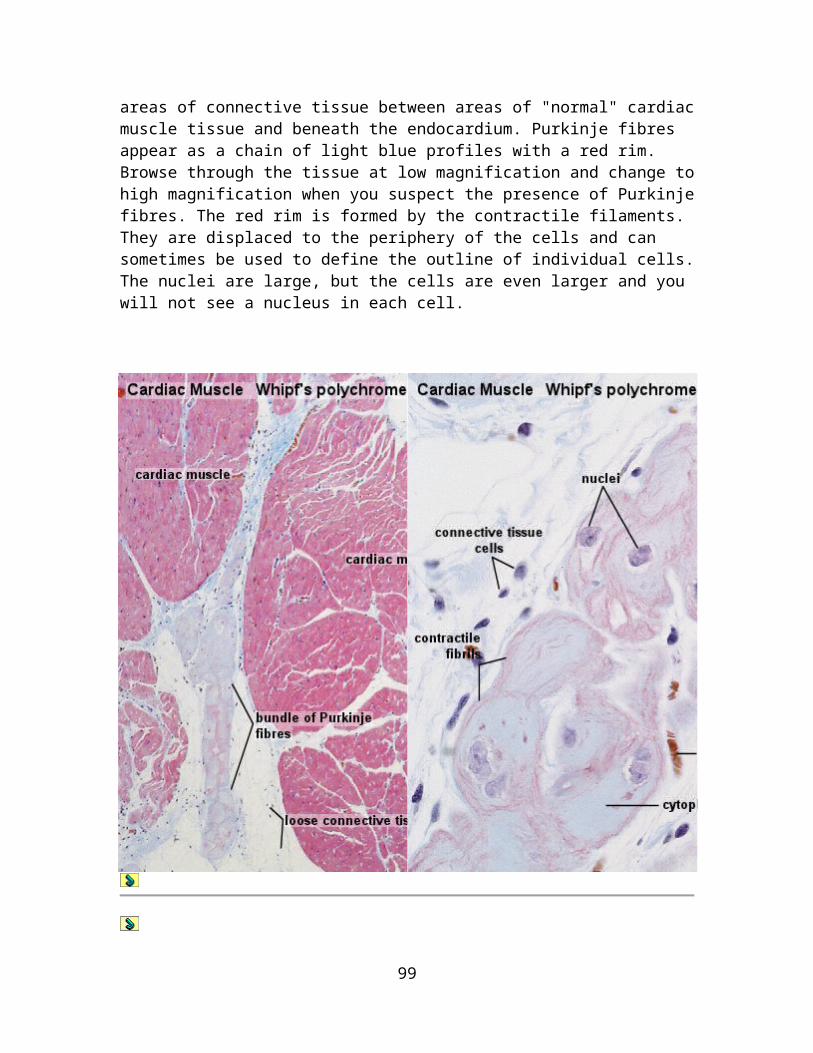
Haemopoiesis
During foetal development, the formation of blood cells (haemopoiesis) commences in wall of the yolk sac. After the second month of foetal development, the liver, and, slightly later, the spleen, become the dominant sites of haemopoiesis. From the 6th month, and dominating from the 7th month onwards, the formation of blood cells occurs in bone marrow, which is the major site of formation of blood cells in normal adult humans.
Yellow bone marrow, which harbours mainly adipocytes, dominates in the hollow of the diaphysis of adult long bones. Haemopoiesis occurs in red bone marrow, which is typically found between the trabeculae of spongy bone in the epiphysis of adult long bones. Both age and demands on haemopoiesis may effect the relative amounts of red and yellow bone marrow. Haemopoietic cells surround the vascular sinusoids and are supported by reticular connective tissue. In addition to the endothelial cells of the
99

sinusoids and the reticulocytes of the connective tissue, macrophages are frequent in red bone marrow.
Haemopoietic Cells
The basis of haemopoiesis is a small population of self-replicating stem cells, which ultimately can generate all types of blood cells. Their progeny may develop into either lymphocytic stem cells or pluripotent haemal stem cells (colony-forming unit - stem cell - CFU-S). The latter type gives rise to stem cells which can form the major groups of blood cells other than lymphocytes. Depending on their progeny it is possible to differentiate
burst-forming unit of the erythroid line (BFU-E), colony-forming unit - granulocytes and macrophages (CFU-G/M), and colony-forming unit - megakaryocytes (CFU-Mk).
Erythrocytes The first identifiable stage of erythropoiesis is the proerythroblast - a large, slightly basophilic cell, which contains a large, lightly stained nulceus. Proerythroblasts proliferate to generate a sequence of cells which show a gradual decrease in size and condensation of their chromatin. They are named after changes in the staining characteristic of their cytoplasm (basophilic erythroblast, polychromatophilic and orthochromic normoblasts). The nucleus is finally extruded from the normoblast. The cell enters circulation as a reticulocyte, which still contains some organelles. Reticulocytes remain for a few days in either the bone marrow or the spleen to mature to erythrocytes.In some blood smears reticulocytes may be recognisable because of a very slight basophilic staining - either homogeneous or in the form of a basophilic stippling.
Granulocytes Myeloblast appear light-microscopically similar to proerythroblast. They proliferate to generate promyelocytes. Promyelocytes begin to accumulate nonspecific granules, but they are still able to divide. The maturation of their progeny, the myelocytes, is characterised by the accumulation of specific granules and changes in nuclear morphology. Metamyelocytes have a C-shaped nucleus.
Blood Platelets (Thrombocytes) are, as mentioned above, fragments of the cytoplasm of megakaryocytes. Megakaryocytes are very large cells (up to 160 µm in diameter), which contain very large, highly lobulated, polyploid nuclei. Megakaryocytes are in turn the product of the differentiation of basophilic megakaryoblasts.
100
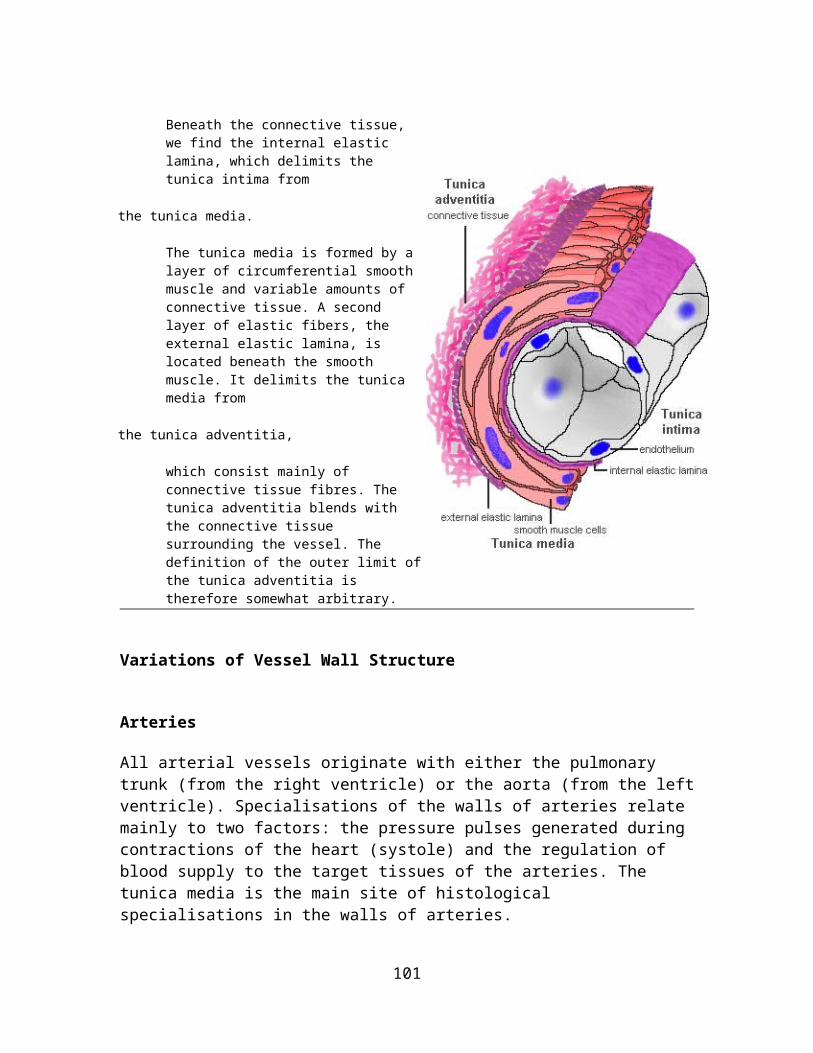
Precursors of blood cells which are usually only found in the bone marrow can be found in peripheral blood in a variety of pathological conditions.If a Rh-negative mother has been immunised by erythrocytes of a Rh-positive foetus, a condition called Erythroblastosis fetalis may develop during subsequent pregnancies. It would show itself in the foetus or newborn by the presence of erythrocyte precursors in peripheral blood - although other, more severe symptoms should be obvious. Chronic myeloid leukemia is another condition - in this case showing itself by the presence of all types of granulocyte precursors in peripheral blood.
The nomenclature employed for haemopoietic cells (but not the number of stages recognized) is somewhat variable across texts. Note also that these cell types refer to stages of development along a morphologically more or less continuous spectrum.
Suitable Slides
sections of red bone marrow - H&E or a bone marrow smear - Leishman, Wright's, Giemsa or May-Grünwald-Giemsa stains
Red Bone Marrow, rabbit - H&EMost of the haemopoietic cells visible will be of the erythroblastic line. The only cell type of this line which is easy to distinguish in H&E stained sections are normoblasts.A very condensed nucleus is seen in late (orthochromic) normoblast. Granulocyte and erythrocyte precursors will mostly intermingle, but may be distinguished by nuclear morphology and/or size. A bent nucleus is found in metamyelocytes - this shape is very pronounced in the last, immature form of neutrophils, which are also called stab or band cells. If the cell (1) is large, with a distinct "clearing" in the otherwise pink cytoplasm and (2) has an ovoid or slightly indented nucleus, it is likely to be a myelocyte. Cells with large light nuclei and almost unstained cytoplasm are either reticulocytes or macrophages.
101

Macrophages are frequently associated with normoblasts, and together these cells form erythroblastic islands. The name for macrophages in these islands, nurse cells, may tell you a bit about their function in addition to the scavenging of the expelled nuclei.Identify normoblasts, myelocytes and metamyelocytes and include them in your drawing of the megakaryocyte/blast.
102

LYMPHOID TISSUES
Lymphoid (or lymphatic) tissues, which mainly consist of dense accumulations of lymphocytes, are widely distributed in the body. Lymphoid tissues are typically located at sites that provide a possible route of entry of pathogens and/or sites that are liable to infections. Epithelia delimit all other tissues from the "outside world", and it is not surprising that lymphoid tissues are often found near them. Such lymphoid tissues are grouped together as epithelium-associated lymphoid tissues. Depending on their precise location these lymphoid tissues may be referred to as e.g. mucosa-associated lymphoid tissue (MALT) or bronchus-associated lymphoid tissue (BALT). The tonsils or Peyer's patches are examples of mucosa-associated lymphoid tissues. Lymphoid tissues represent the sites of proliferation and differentiation of lymphocytes.
Lymphoid organs may be defined as anatomical "entities" which consists chiefly of lymphoid tissues. The thymus is a primary lymphoid organ in that it supplies other lymphoid organs and tissues with T-lymphocytes. Inserted into the blood and lymph vascular system, the spleen and lymph nodes (secondary lymphoid organs) monitor the internal environment of the body.
Thymus
The thymus is situated in the upper parts of the thorax, behind the sternum and the upper four costal cartilages, in the anterior and superior mediastina. The size of the thymus changes in the course of life. It weighs about 10-15 g at birth and reaches its top weight (about 30-40 g) at puberty. After puberty a progressive involution (see below) occurs, which leaves a middle-aged person with a thymus weighing about 10 g. The thymus consists of a right and left lobe which are joined by connective tissue.
The thymus is enclosed by a thin connective tissue capsule from which numerous septa extend into the thymus subdividing the two lobes into numerous lobules (about 0.5 -2 mm in diameter). Blood vessels enter and leave the thymus via the connective tissue septa. Each lobulus is divided into a darker peripheral zone, the cortex, and a lighter, central zone, the medulla. Medullary tissue is continuous from lobule to lobule throughout each lobe.
Reticular cells and macrophages are present in addition to the lymphocytes, which are the dominant cell type within the lobules.
Reticular cells
are quite abundant. Their cytoplasm is eosinophilic, and their large, ovoid and light nuclei may contain one or two nucleoli. The cells are branched, and their slender processes are connected with the processes of other reticular cells to form
103
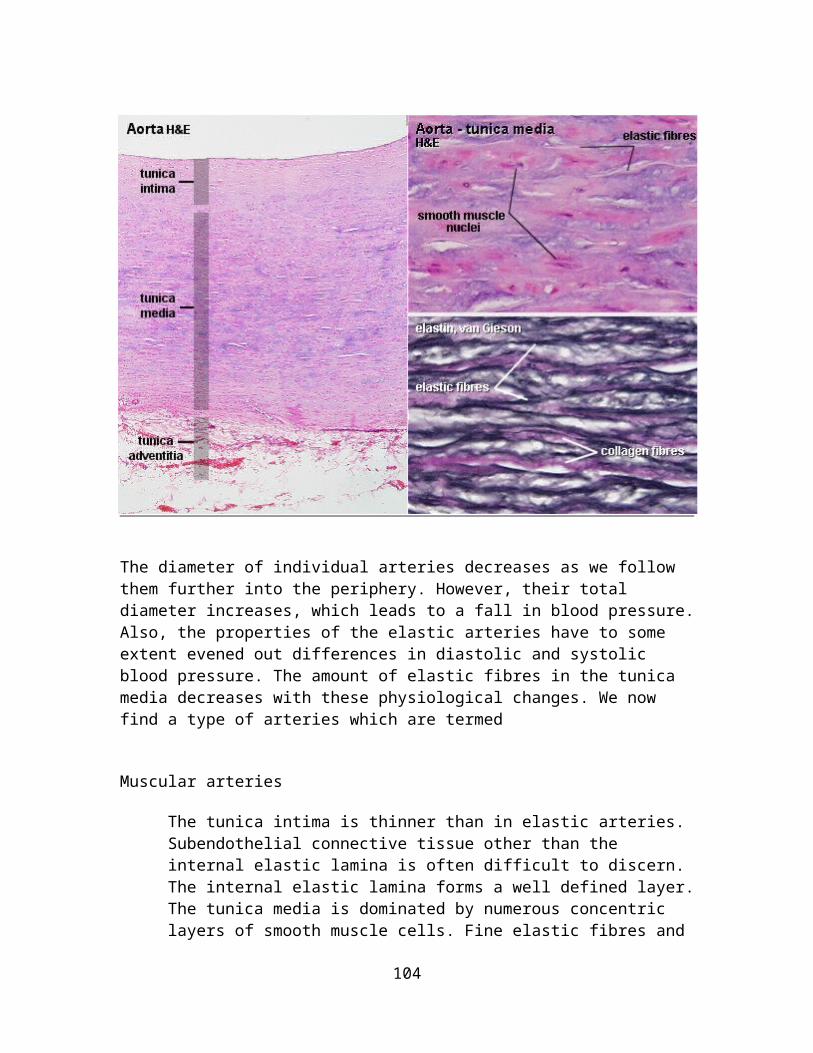
a cellular reticulum (or cellular network). This cellular network (reticular fibres are scant in the thymus) provides support for other cells of the thymus.
Reticular cells sheathe the cortical capillaries; they form an epitheloid layer which delimits the cortical tissue from the connective tissue and secrete substances (hormones and other factors) important for thymic function. Thereby they create and maintain the microenvironment necessary for the development of T-lymphocytes in the cortex. Their functions thus go beyond those of "typical" reticular cells and, to reflect this, they are also referred to as thymic epitheliocytes.
Macrophages
occur in both cortex and medulla. They are difficult to distinguish from the reticular cells in H&E stained sections.
Lymphocytes
are present in both cortex and medulla, but are more numerous (denser) in the cortex. Their sizes are variable (5 - 15 µm) in the cortex but generally small in the medulla. The vast majority of them will be developing T-lymphocytes. They are also called thymic lymphocytes or thymocytes.
Thymus, foetal human - H&EIdentify the connective tissue capsule and septa, a lobule and its cortex and medulla at low magnification. Identify lymphocytes and thymic corpuscles. They look pretty much like a sliced (very, very small) onion. Take a close look at the medulla and try to find some cells which contain large and light nuclei. They will be either macrophages or reticular cells.Sketch part of the thymus at low magnification. Identify medulla and cortex. Draw a segment of a lobule at high magnification and identify lymphocytes and nuclei of reticular cells/macrophages and, if possible a Hassall corpuscle.
104

Function of the Thymus
The thymus is necessary for the development of the recirculating pool of small, long-lived (in humans many years) lymphocytes, the T-lymphocytes. These cells are mainly responsible for the cell-mediated part of an immune response. Stem cells invade the cortical regions of the thymus, where they divide to form lymphocytes. Only a small fraction (estimates range from 10-30%) of the cells generated in the cortex leave the thymus. They migrate via the medulla into the blood stream to populate the T-lymphocyte areas of other lymphoid tissues and organs. Cells which do not express the necessary receptors to recognize antigens presented to them or which react incorrectly towards "self-antigens" die and are removed by cortical macrophages.Since the function of the thymus is to produce lymphocytes for the other lymphoid tissues it is a primary lymphoid organ.
Involution of the thymus
After puberty much of the parenchyma of the thymus, in particular cortical lymphoid tissue, is replaced by adipose tissue. The process, which is called involution, initially proceeds rapidly but slows down in adulthood. Involution is under the control of steroid
105
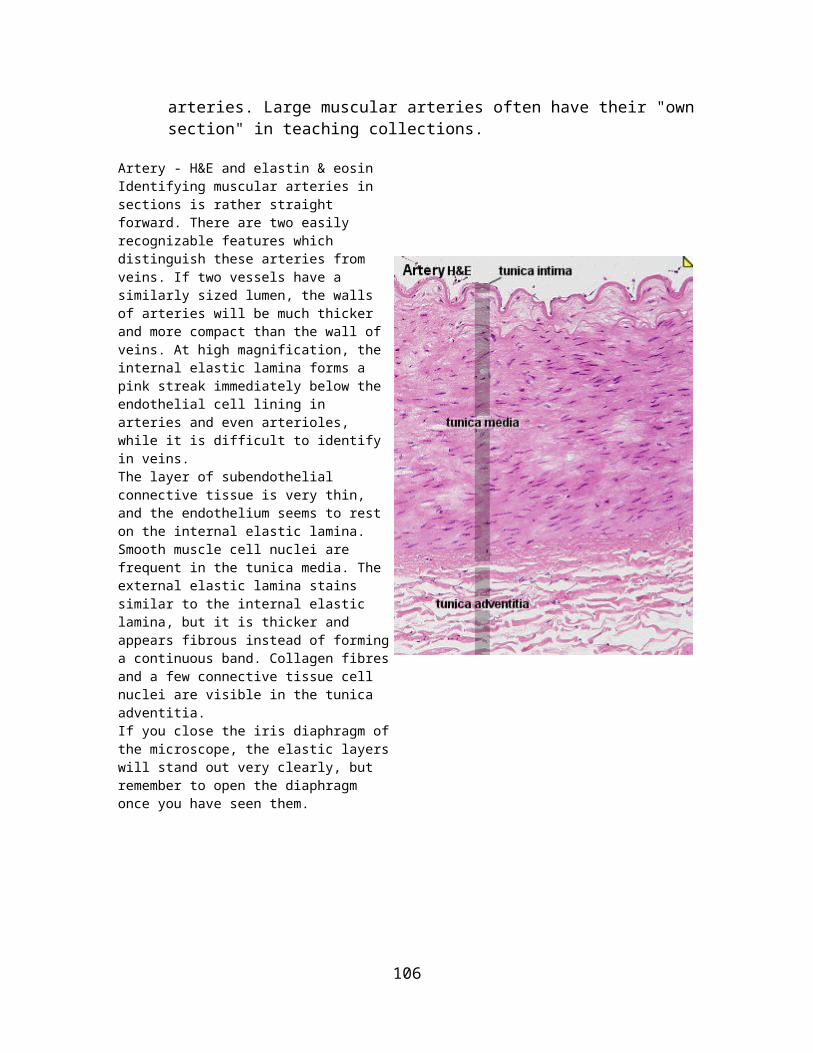
hormones (both sexual hormones and stress hormones). Although most pronounced in the thymus, involution is a common feature of all lymphoid tissues.
Another age-related phenomenon is the increase in size of the thymic (or Hassall's) corpuscles. Thymic corpuscles are rounded eosinophilic structures, which consist of concentrically arranged, flattened cells. Thymic corpuscles are likely to be formed by reticular cells. Similar structures occur also in the tonsils. The size of these structures varies from 20 µm to more than 100 µm in diameter. Thymic corpuscles may calcify, and their core may "dissolve" leading to the formation of a cyst.
Thymus, adult human - H&EDraw a part of the tissue, which illustrates the presence of adipose tissue, and the decrease in the amount of cortical and medullary thymic tissue. Include thymic corpuscles in your drawing if you can find them.
Lymph Nodes
Lymph nodes are small, flattened, oval or bean shaped organs, which are situated in the course of the collecting lymph vessels. Their size is variable (from a few mm to more than 2 cm). The capsule and trabeculae of lymph nodes are formed by connective tissue. Afferent lymph vessels penetrate the capsule and empty into the subcapsular space. The lymph continues thereafter through cortical and medullary sinuses towards the efferent lymph vessels, which emerge from the hilus of the lymph node. The walls of the sinuses can be traversed freely by all components of the lymph, which allows lymphocytes to
106

enter/leave the lymphoid tissue (as part of their constant circulation) or to get in contact with antigens/antigen-presenting cells that may arrive with the lymph.
In lymph nodes we find B- and T-lymphocytes, macrophages and reticular cells.
Reticular cells
(and reticular fibres) form a delicate network between the capsule and trabeculae. Only their large and light nuclei are easily visible in the microscope. The cytoplasm of reticular cells is only weakly eosinophilic. Lymphocytes and macrophages are housed in the network of reticular cells and the reticular fibres formed by them. The processes of reticular cells and reticular fibres extend into and criss-cross within the sinuses.
Lymphocytes
which are located in the outer cortex of the lymph node are likely to represent B-lymphocytes. They are organised into spherical masses - lymphoid nodules or follicles. Sites within the cortex at which B-lymphocytes have been stimulated to proliferate (by contact with an antigen) appear lighter than the surrounding tissue and allow you to identify the centres of lymphoid nodules. The lighter stained parts of the nodules are called germinal centres. Mature B-lymphocytes (plasma cells) are located in cord-like extensions of the lymphoid tissue into the medulla, the medullary cords. T-lymphocytes are located in the more diffuse tissue between the nodules and in the paracortex, i.e. the deep part of the cortex.
Macrophages
are found scattered within the lymphoid tissue. In many preparations they are difficult to distinguish from the reticular cells, but if an H&E stain turns out nice, macrophages can be distinguished from the reticular cells in the sinus system of the lymph node.
Blood Vessels
Blood vessels enter the lymph nodes through the hilus and travel initially in the connective tissue trabeculae that extend from the hilus into the parenchyma of the lymph node. They continue in the medullary cords towards the cortex and give off capillaries to the surrounding tissue as they do so. High-endothelial venules (or postcapillary venules) in the deep cortex have a characteristic low cuboidal epithelium - quite unlike the squamous epithelium that we usually would expect to see. Lymphocytes, which reach the
107

lymph node via the blood stream, may migrate through this epithelium as part of their recirculation. Larger venules accompany the arteriolar branches as they leave the lymph nodes.
Lymph node, rabbit - H&EThe subcapsular and cortical sinus system was hardly (if at all) visible in the slides I looked at. Identify the connective tissue capsule and trabeculae, cortex and medulla of the lymph node, lymph nodules with germinal centres, medullary cords and postcapillary venules.
108

Lymph node - H&E, carbon injectedThis slide illustrates the distribution of macrophages in lymph nodes - take a quick look at them. Note that most of them are located in the paracortex and medulla.
109

Spleen
The spleen is, like the lymph nodes, a discriminatory filter. Unlike the lymph nodes, the spleen is inserted into the blood stream. The spleen clears the blood of aged blood cells and foreign particles and is the site of immune reactions to blood-borne antigens. The spleen is not essential to life in adult individuals. Other organs can take over its functions if the spleen is removed.
The spleen is surrounded by a capsule of dense connective tissue from which branched trabecula extend into the parenchyma of the spleen (sounds familiar). The parenchyma of the spleen is termed the pulp of the spleen. Most of the pulp of a fresh, unfixed spleen is a soft, dark red mass, the red pulp. It consists of large, irregular, thin-walled blood vessels, the splenic sinusoids, interposed between sheets and strands of reticular connective tissue, the splenic cords (of Billroth). Within the red pulp small, oval or rounded greyish white areas, the white pulp, is formed by lymphoid tissue.
Branches of the splenic artery divides into trabecular arteries, which enter the white pulp, where they are called central arteries. Branches of the central artery almost all divide into smaller vessels in the marginal zone, i.e. the border between the red and white pulp. Fine branches of the central artery - penicillar arteries (cuboidal epithelium) - branch again to form arterial capillaries, which, as they exit the white pulp, are surrounded by a sheath of phagocytotic cells and reticular fibres. They are now called sheathed arteries. From here, the blood enters the red pulp. Sheathed arteries may empty the blood which they carry directly into the splenic sinusoids (closed circulation, about 90% in cats) or into the reticular connective tissue of the splenic cords (open circulation). Macrophages are, in addition to reticular cell, the main resident cell population of the splenic cords.
Blood cells which are emptied into the splenic cords re-enter the blood vessels through the endothelium of the sinusoids. The endothelial cells are elongated (in cross section they may appear cuboidal) and oriented along the long axis of the sinusoids. The endothelium of the sinusoids has no junctional complexes and its basement membrane is incomplete (forming narrow circular bands around the endothelial cells with large intervening fenestrations). Macrophages ingest aged erythrocytes, platelets and other particulate matter as they pass through the splenic cords.
The composition of the plasma membrane of erythrocytes changes as the cell ages. It is thought that these changes eventually expose erythrocyte senescence antigens, which bind blood-borne antibodies and thus tag the erythrocyte for removal by macrophages. Erythrocyte removal is also one function of the resident macrophages of the liver, although splenic macrophages take care of most of the job.
The sinusoids continue into the veins of the pulp, which empty into thin-walled trabecular veins, which eventually coalesce to form the splenic vein.
110

The white pulp surrounds the central arteries as a periarterial lymphoid sheath (PALS). Lymphocytes of the PALS are likely to be T-lymphocytes. In addition, we see macrophages and plasma cells in the PALS. Lymph nodules, formed by B-lymphocytes, are present along the course of the central arteries. The central arteries are typically located in the periphery of the nodule.
Spleen, human - H&EFind a place close to the capsule where you can identify trabeculae, white pulp (possibly a nodule with a peripherally placed central artery), and red pulp. Good penicillar and sheathed arteries are very hard to identify - do not despair if you (or the demonstrator) cannot find them. As usual, it is easiest to identify macrophages by accumulations of particulate matter in their cytoplasm, which often will represent disintegrating erythrocytes.
111

Spleen, cat - reticulinThis slide illustrates the distribution of reticular fibres in the spleen. They often appear coarser in the red pulp, where they have a distinct, stranded organisation. The reticular fibres of the white pulp appear somewhat finer and, at times, they are arranged as concentric rings. The peripheral localisation of the central arteries in nodules is quite distinct. Occasionally you may see small rings of reticular fibres in (or close to) the periphery of the white pulp. These rings are likely to represent the reticular fibres surrounding sheathed arteries.
Mucosa-Associated Lymphoid Tissue
The mucosal lining of the alimentary canal and airways is in many ways specialised to facilitate the exchange of substances between the external environment and the body. Unfortunately, these specialisation do not just apply e.g. to components of the digested food but also pathogens. This is combined with excellent living conditions for bacteria in parts of the alimentary canal - in particular the ileum and the colon. Lymphoid tissue located beneath the mucosal epithelia, mucosa-associated lymphoid tissue (MALT), protects the body against pathogens that may enter the body via the mucosa. The importance of this task is reflected in the mass of the MALT, which corresponds to the combined mass of the other lymphoid organs and tissues.
112

The task that the immune cells of the MALT have to accomplish is different from that of other parts of the immune system. We do need a defense against pathogens, but it would not be a good idea to mount an immune response against components of the food. Immune cell activation therefore differs between the MALT and other lymphoid tissues.This difference is mediated by different receptors expressed by immune cells of the MALT and by different substances which they release upon contact with an antigen. Because of their specific functions, immune cells of the MALT do not mingle with other immune cells. Epithelial cells of the vessels supplying the MALT express specific receptor which are recognized by MALT immune cells and allow their homing to the MALT during recirculation. Lastly, MALT plasma cells produce a secretable form of antibodies, immunoglobulin type A dimers, which can be taken up by epithelial cells and then released onto the epithelial surface.Specialisation of MALT immune cells occur at the molecular level. In routine histological preparations, immune cells of the MALT look pretty much like immune cells of other lymphoid tissues.
Often MALT consists of small accumulations of lymphoid cells or one to a few lymph follicles beneath the epithelium and possibly extending into the submucosa. The tonsils and Peyer's patches are large accumulations of lymphoid tissue with associated specialisations of the epithelium.
Tonsils
The tonsils are accumulations of lymphoid tissue surrounding the openings of the digestive and respiratory tracts. The tonsils and smaller accumulations of lymphoid tissue, which may be found between them, are also called Waldeyer's ring.
Depending on their localisation we distinguish between
palatine tonsils (THE tonsils), which are located in the lateral wall of the oropharynx and covered by a stratified squamous epithelium,
113

lingual tonsils which are situated in the lamina propria at the root of the tongue and also covered by a stratified squamous epithelium, and
pharyngeal tonsils (also called nasopharyngeal tonsils or adenoids) which are located in the upper posterior part of the throat (nasopharynx) and covered by a pseudostratified ciliated epithelium with goblet cells.
The tonsils do not have afferent lymph vessels. Efferent lymph vessels are present. Exposure to antigens relies on the contact of antigens with cells of the immune system across the epithelium which covers the tonsils. The epithelium of the palatine and lingual tonsils forms deep crypts into the lymphoid tissue, and the resulting increase of the surface area is one way to facilitate the contact of antigens with the immune cells. In addition, the epithelium may specialise in places to form an open meshwork of cells with an incomplete basal lamina (a reticulated epithelium) which allows the infiltration of the epithelium by lymphocytes and macrophages.Tonsillar lymphoid nodules consist mainly of B-lymphocytes. Other areas are occupied by T-lymphocytes, activated B-lymphocytes and other cells of the immune system.
The tonsils share some histological features with lymph nodes:
1. cells in the tonsils are supported by a fine network of reticular fibres and 2. high-endothelial (postcapillary ~) venules function in the "homing" of circulating
lymphocytes - this is actually a shared feature of all lymphoid tissues and organs.
The palatine tonsils are surrounded by a thick hemicapsule of connective tissue, which delimits them from the pharyngeal muscle and facilitates their removal in tonsillitis.
114
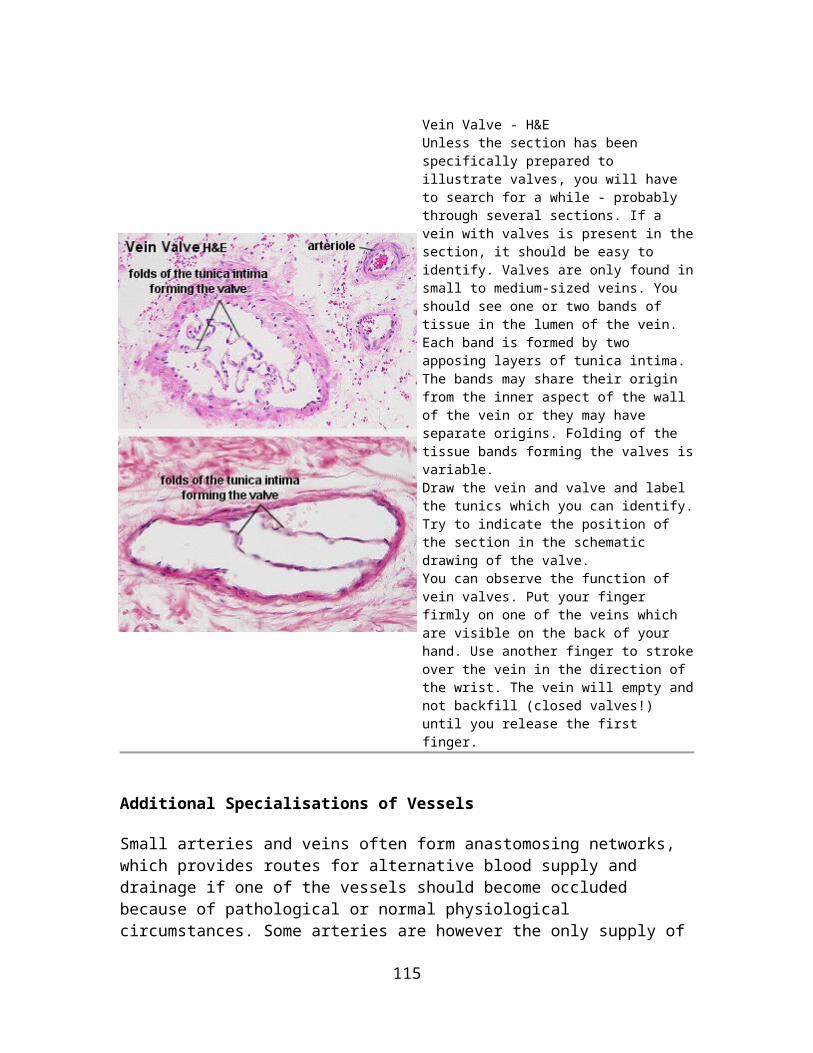
Tonsil, human - H&EIdentify the surface epithelium covering the lymphoid tissue of the tonsils, lymph nodules and tonsillar crypts. Have a look at the epithelium both inside and outside the crypts. Outside the crypts, the epithelium will typically look like ordinary stratified squamous epithelium. Inside the crypts, where cells of the immune system often invade the epithelium, it will be difficult to find the boundary between epithelium and lymphoid tissue. Tissue preservation is not that great, but with a little bit of patience you should be able to find high-endothelial venules in the lymphoid tissue.Sketch the organisation of the tissue at low magnification.
Gut-Associated Lymphoid Tissue - GALT
115

Small accumulations of lymphocytes or solitary lymph follicles are found scattered in beneath the epithelium throughout the gastrointestinal tract. However, the most prominent accumulations occur in the ileum and appendix in the form of Peyer's patches. In the ileum, they form dome-shaped protrusions into the lumen. Beneath the epithelial lining of the domes, Peyer's patches extend from the lamina propria to the submucosa. Within Peyer's patches, lymph follicles with germinal centers are typically located deep in the submucosa.
The epithelium in contact with the lymphoid tissue is specialised to facilitate the contact of antigens with cells of the immune system. The epithelium appears columnar and contains cells with deeply invaginated basal surfaces - microfold cells or M-cells. Immune system cells can enter these invaginations (intraepithelial pockets) where they are exposed to materials which have been endocytosed by the epithelial cells and then released into the invaginations. Goblet cells are rare or absent in the epithelium which covers the domes.
Suitable Slides slides of appendix, ileum or Peyer's patches - H&E Slides of the appendix and ileum may be useful is there is no specially prepared slide of Peyer's patches. Extensive areas of lymphoid tissue are alway present in the appendix, but domes and specialised sections of epithelium may not. If you have a slide of the ileum hold it against the light and see if parts of the wall look darker than the rest. These parts are likely to contain lymphoid tissue and may show domes and/or specialised sections of epithelium.
116

Peyer's Patch, Ileum - H&E If you hold the sections against a light surface the areas which are occupied by the lymphoid tissue should be readily visible - in H&E stained sections they appear darker than the remaining tissue. Look at these areas under the microscope. Identify the domes and their epithelial covering. The epithelium will be lower than the epithelium covering the villi and goblet cells are rare or absent. See if you can find intraepithelial pockets containing immune cells. Even if no clear pockets are present, some lymphocytes should appear scattered "over" the epithelium. In reality they will be located in the intraepithelial pockets. High-endothelial venules should be present in the lymphoid tissue beneath the domes but may be difficult to find. Next go further towards the submucosa and see if you can find lymph follicles.Draw the epithelium covering the domes at high magnification. Include a scetch of the normal intestinal epithelium for comparison. Scetch the appearance of Peyer's patches at low magnification.
117

NERVOUS TISSUE
The nervous system consists of all nervous tissue in the body. It is divided anatomically into the central nervous system and the peripheral nervous system.
Central Nervous System (CNS)
The CNS consists of the brain (encephalon), which is enclosed in the skull, and the spinal cord, which is contained within the vertebral canal. Nervous tissue of the CNS does not contain connective tissue other than that in the three meninges (dura mater, arachnoid membrane and pia mater) and in the walls of large blood vessels. Collagenous fibers or fibrocytes/blasts are consequently not observed, which is quite unlike other tissues. Because of the absence of connective tissue, fresh CNS tissue has a very soft, somewhat jelly-like consistency. The two major classes of cells that make up the nervous tissue are nerve cells, neurones, and supporting cells, glia.
118

Neurons
The vast majority of neurones is generated before birth. Persisting stem cells give rise to a small number of new neurones throughout the lifetime of mammals, including humans. The permanent addition of neurones may be important for the maintenance and plasticity of some parts of the CNS, but it is insufficient to replace neurones that die because of disease or acute damage to the CNS. Neurones should last a lifetime. Mature neurones are not mitotically active, i.e. they do not divide.
Neurones are generally large cells. Neural activity and its control require the expression of many genes, which is reflected in the large and light nuclei of most neurones. The keys to the understanding of the function of a neurone lies in (1) the shape of the neurone and, in particular, its processes, (2) the chemicals the neurone uses to communicate with other neurones (neurotransmitters) and (3) the ways in which the neurone may react to the neurotransmitters released by other neurones.
The shape of the neurone and its processes
Neurones have long processes, which extend from the part of the cell body around the nucleus, the perikaryon or soma. The processes can be divided into two functionally and morphologically different groups, dendrites and axons.Dendrites are part of the receptive surface of the neurone. As a rule, neurones have one to several primary dendrites, which emerge from the perikaryon. Primary dendrites may divide into secondary, tertiary etc. dendrites. Dendrites can be smooth, or they can be studded with small, mushroom-shaped appendages, which are called spines.
Each neurone has as a rule one axon, and never more than one axon which emerges from the perikaryon or close to the trunks of one of the primary dendrites. The point of origin of the axon from the perikaryon is the axon hillock. The axon may, like the dendrites, branch as it travels through the nervous tissue to its destination(s). The axon is the "transmitting" process of the neurone.
119

The axon forms small, bulb-shaped swellings called boutons at the ends (terminal boutons) or along the course (boutons en passant) of its branches. Synapses are morphologically specialised contacts between a bouton formed by one neurone, the presynaptic neurone, and the cell surface of another neurone, the postsynaptic neurone. Synaptic vesicles contain the neurotransmitters. Synaptic vesicles typically accumulate close to the site of contact between the bouton and the postsynaptic neurone. The release of the neurotransmitter from the synaptic vesicles into the synaptic cleft, i.e. the space between the bouton and the postsynaptic neurone, mediates the transfer of information from the pre- to the postsynaptic neurone. Both the release of the synaptic vesiscles and the mediation of the response to the transmitter require membrane-associated specialisations - the pre- and postsynaptic densities.
The shape and orientation of the dendritic tree of the neurone determines the amount and type of information that may reach the neurone. The course of its axon determines to which neurones this information may be passed on. The location of the neurone within the CNS determines to which major system the neurone belongs.
There are several hundred functionally different areas, i.e. groups of neurones, in the CNS. Based on their location, the shape of their dendritic tree and the course of their axon, several thousand types of neurons can be distinguished in the CNS. Transmitters
Neurotransmitters either excite or inhibit the postsynaptic neurone. The most prominent excitatory transmitter in the CNS is L-glutamate. The most prominent inhibitory transmitter in the CNS is GABA (gamma-amino butyric acid). Other "main" neurotransmitters are e.g. dopamine, serotonin, acetylcholine, noradrenaline and glycine. Each neurone uses only one of the main transmitters, and this transmitter is used at all synaptic boutons that originate from the neurone.One or more of the "minor" transmitters (there are several dozens of them - such as cholecystokinin, endogenous opioids, somatostatin, substance P) may be used together with a main transmitter.
120

The molecular machinery that is needed to mediate the events occurring at excitatory synapses differs from that at inhibitory synapses. Differences in the morphological appearances of the synapses accompany the functional differences. The pre-and postsynaptic densities are typically of equal width, or symmetric, at inhibitory synapses. The postsynaptic density is thicker than the presynaptic density at asymmetric synapses, which are typically excitatory.
Receptors
Usually there exists a multitude of receptors which are all sensitive to one particular neurotransmitter. Different receptors have different response properties, i.e. they allow the flux of different ions over the plasma membrane of the neurone or they may address different second messenger systems in the postsynaptic neurones. The precise reaction of the neurone to the various neurotransmitters released onto its plasma membrane at the synapses is determined by the types of receptors expressed by the neurone.
Suitable Slides
sections of spinal cord - H&E, luxol fast blue/cresyl violet (LFB/CV), toluidine blue, Giemsa
Thoracic Spinal Cord, sheep - LFB/CVMost neurones have a light, large nucleus with a distinct nucleolus. The cytoplasm of many neurones contains fairly large amounts of rough endoplasmatic reticulum, which may aggregate within the cytoplasm of the neurone to form Nissl-bodies. Nissl-bodies are prominent in motor neurones located in the ventral horn of the grey matter of the spinal cord. The neurites are difficult to identify in most types of stained sections. Only the most proximal segments of the primary dendrites are seen clearly. The size of the perikaryon depends on the level of activity of the neurone and the length of the processes which the neurone has to support. An usable range for the size of the perikaryon would be 15 - 50 µm, although much smaller and much larger neuronal perikarya exist.Draw the spinal cord at low magnification and indicate the distribution of grey matter and white matter. Find a nice group of neurones in the grey matter and draw them at a high magnification. Finally, have a look at the white matter and identify the nuclei of glial cells. You will find similar nuclei also in the grey matter.
121
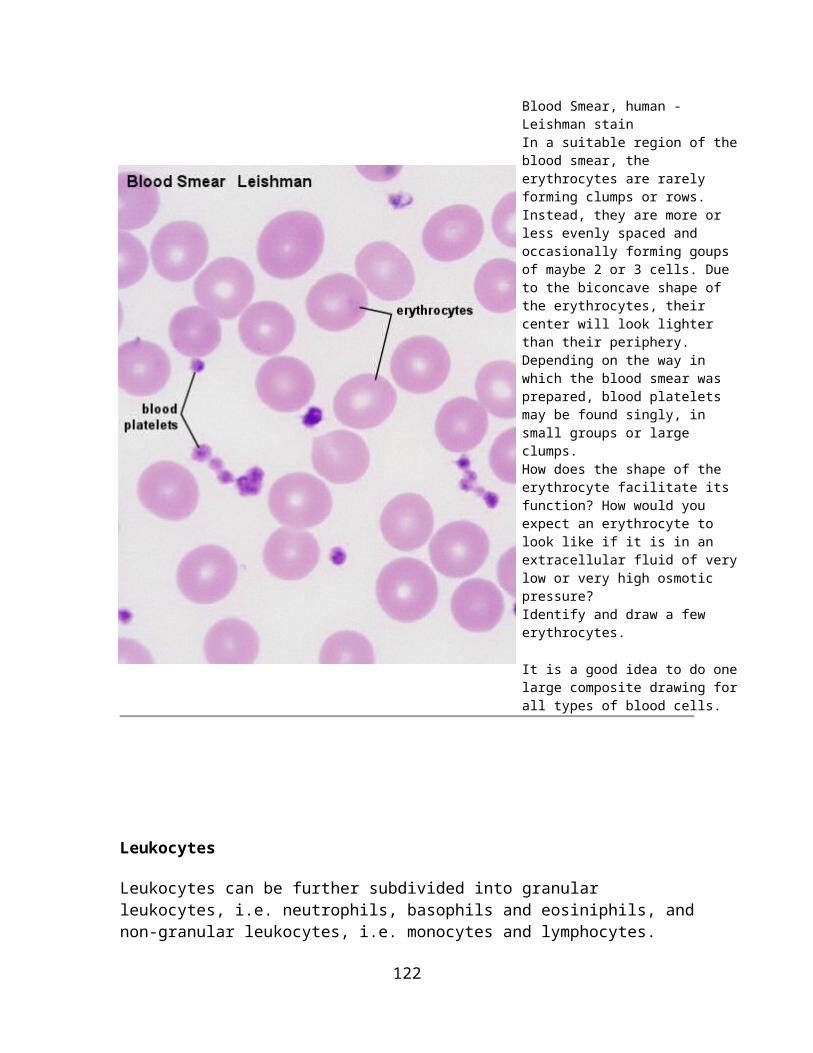
Thoracic spinal Cord - H&E, silver stainThese slides show the same major features as the LFB/CV stained sections. Try to identify neurones (primary dendrites, Nissl-bodies) and glial cell nuclei in the H&E stained section. Differentiate between grey and white matter. The LFB stain showed the myelin sheath nicely. In the H&E stained section we instead can see large, cross-sectioned axons in the white matter. The feltwork of nerve fibres, neuronal and glial cell processes is also called neuropil.Part of the cytoskeleton of neurones is (like the reticular connective tissue fibers) argyrophilic, i.e. they "love" silver and can be stained by silver stains. Aside from the neurones and their processes, fine fibrils are visible in the neuropil. Many of the fibrils represent axons travelling in the grey and white matter of the spinal cord.
122

123

Glial Cells
CNS tissue contains several types of non-neuronal, supporting cells, neuroglia.
Astrocytes (or astroglia) are star-shaped cells. Their processes are often in contact with a blood vessel (perivascular foot processes). Astrocytes provide mechanical and metabolic support to the neurones of the CNS. They participate in the maintenance of the composition of the extracellular fluid. Although not themselves directly involved in the process of communication between neurones, they may be involved in the removal of transmitters from synapses and the metabolism of transmitters. Astrocytes are also the scar-forming cells of the CNS.
Oligodendrocytes (or oligoglia) have fewer and shorter processes. Oligodendrocytes form myelin sheath (see below) around axons in the CNS and are the functional homologue of peripheral Schwann cells. Oligodendrocytes may, in contrast to Schwann cells in the periphery, form parts of the myelin sheath around several axons.
Microglia are small cells with complex shapes. Microglia are, in contrast to neurones and the other types of glial cells, of mesodermal origin. They are derived from the cell line which also gives rise to monocytes, i.e. macrophage precursors which circulate in the blood stream. In the case of tissue damage, microglia can proliferate and differentiate into phagocytotic cells.
124

The ventricles of the brain and the central canal of the spinal cord are lined with ependymal cells. The cells are often cilated and form a simple cuboidal or low columnar epithelium. The lack of tight junctions between ependymal cells allows a free exchange between cerebrospinal fluid and nervous tissue.Ependymal cells can specialise into tanycytes, which are rarely ciliated and have long basal processes. Tanycytes form the ventricular lining over the few CNS regions in which the blood-brain barrier is incomplete. They do form tight junctions and control the exchange of substances between these regions and surrounding nervous tissue or cerebrospinal fluid.
Many glial cells do express neurotransmitter receptors. Neuronal activity may regulate glial function by a spillover of transmitter from synaptic sites, which are typically surrounded by fine processes of glial cells. Occasionally, neurones also make synapse-like contacts with glia cells. Glial cells may also communicate with each other via GAP junctions.
Suitable Slides sections of the forebrain - toluidine blue, Giemsa, luxol fast blue/cresyl violet
125

Forebrain, Cortex, mouse - Giemsa and Forebrain, Hippocampus, mouse - GiemsaMost glial cells are much smaller than neurones. Their nuclei are generally much smaller than neuronal nuclei, and they rarely contain an easily visible nucleolus. Other aspects of their morphology are variable. The glial cytoplasm is, if visible at all, very weakly stained. Different types of glial cells cannot be easily distinguished by their appearance in this type of preparation. Most of the small nuclei located in the white matter of the CNS, where they may form short rows, are likely to represent oligodendrocytes.Browse through the sections at low or medium magnification and try to get a feeling for the structural diversity visible in the section available to you - parts of the section that look different from others are very likely to have different functions.Find a spot that appears interesting (or least boring) to you and sketch its structure at low magnification. Choose a spot for high magnification, and draw some of the visible neurones and glial cells. Note the difference in the size and number of glial cells and neurones.
Peripheral Nervous System (PNS)
126

The PNS comprises all nervous tissue outside the brain and spinal cord. It consists of groups of neurones (ganglion cells), called ganglia, feltworks of nerve fibres, called plexuses, and bundles of parallel nerve fibres that form the nerves and nerve roots. Nerve fibres, which originate from neurones within the CNS and pass out of the CNS in cranial and spinal nerves, are called efferent or motor fibers. Nerve fibres which originate from nerve cells outside the CNS but enter the CNS by way of the cranial or spinal nerves are called afferent or sensory nerve fibres.
The principal neurotransmitters in the PNS are acetylcholine and noradrenalin.
Peripheral Nerves
Afferent, sensory fibres enter the spinal cord via the dorsal roots, while efferent, motor fibres leave the spinal cord via the ventral roots. Dorsal and ventral roots merge to form the spinal nerves, which consequently contain both sensory and motor fibres. As the spinal nerves travel into the periphery they split into branches and the exact composition of the nerve in terms of motor and sensory fibres is, of course, determined by the structures the nerve will innervate.
One nerve fibre consists of an axon and its nerve sheath. Each axon in the peripheral nervous system is surrounded by a sheath of Schwann cells. An individual Schwann cell may surround the axon for several hundred micrometers, and it may, in the case of unmyelinated nerve fibers, surround up to 30 separate axons. The axons are housed within infoldings of the Schwann cell cytoplasm and cell membrane, the mesaxon.
In the case of myelinated nerve fibres, Schwann cells form a sheath around one axon and surround this axon with several double layers (up to hundreds) of cell membrane. The myelin sheath formed by the Schwann cell insulates the axon, improves its ability to conduct and, thus, provides the basis for the fast saltatory transmission of impulses. Each Schwann cell forms a myelin segment, in which the cell nucleus is located approximately in the middle of the segment. The node of Ranvier is the place along the course of the axon where two myelin segments abut.
127

Fibre types in peripheral nerves:
Type A fibres (myelinated) are 4 - 20 µm in diameter and conduct impulses at high velocities (15 - 120 m per second). Examples: motor fibers, which innervate skeletal muscles, and sensory fibres.
Type B fibres (myelinated) are 1 - 4 µm in diameter and conduct impulses with a velocity of 3 - 14 m per second. Example: preganglionic autonomic fibres.
Type C fibres (unmyelinated) are 0.2 - 1 µm thick and conduct impulses at velocities ranging from 0.2 to 2 m per second. Examples: autonomic and sensory fibres.
Peripheral nerves contain a considerable amount of connective tissue. The entire nerve is surrounded by a thick layer of dense connective tissue, the epineurium. Nerve fibres are frequently grouped into distinct bundles, fascicles, within the nerve. The layer of connective tissue surrounding the individual bundles is called perineurium. The perineurium is formed by several layers of flattened cells, which maintain the appropriate microenvironment for the nerve fibres surrounded by them. The space between individual nerve fibres is filled by loose connective tissue, the endoneurium.
Fibrocytes, macrophages and mast cells are present in the endoneurium.Nerves are richly supplied by intraneural blood vessels, which form numerous anastomoses. Arteries pass into the epineurium, form arteriolar networks beneath the perineurium and give off capillaries to the endoneurium.
128

Suitable Slides sections of peripheral nerve - H&E, osmium or plucked preparations of peripheral nerve - osmium
Peripheral Nerve, cat - osmiumWhich structures can be recognized in peripheral nerves depends on the stain that has been used in the preparation. Osmium gives a black color to lipids. In osmium stained preparations it is possible to observe the myelin sheath surrounding the axon. A good impression of the different sizes of the nerve fibres may be obtained. The axon is usually not well preserved. It may only form a little dark spot somewhere within the dark ring which represents the myelin sheath. Lipid droplets in fat cells, which can be found in the connective tissue around nerves, stand out as large (much larger than the nerve fibres), round, homogeneously stained areas.Draw the nerve at low magnification (you may include some of the stained lipid droplets) and a small section of it at high magnification.
Peripheral Nerve, rat - H&EIn longitudinal H&E stained sections it is possible to identify the axon running in its myelin sheath, nodes of Ranvier and Schwann cell nuclei. Components of the connective tissue elements, which accompany the nerve, should be visible and identifiable in both longitudinal and transverse sections. H&E stained and transversely cut preparations give a good picture of the axon in the middle of a ring-like structure (sometimes fussy), which represents the remains of the myelin sheath. Due to their small size and the lack of a myelin sheath, type C fibres are very difficult to detect in either osmium or H&E stains.
129

Draw part of the longitudinally and transversely sectioned nerve at high magnification. Include Schwann cell nuclei, myelin sheath, axons and, if possible, nodes of Ranvier.
Ganglia
Ganglia are aggregations of nerve cells (ganglion cells) outside the CNS. Cranial nerve and dorsal root ganglia are surrounded by a connective tissue capsule, which is continuous with the dorsal root epi- and perineurium. Individual ganglion cells are surrounded by a layer of flattened satellite cells. Neurones in cranial nerve and dorsal root ganglia are pseudounipolar. They have a T-shaped process. The arms of the T represent branches of the neurite connecting the ganglion cell with the CNS (central branch) and the periphery (peripheral branch). Both branches function as one actively conducting axon, which transmits information from the periphery to the CNS. The stem is connected to the perikaryon of the ganglion cell and is the only process originating
130

from it. Ganglion cells in dorsal root ganglia do not receive synapses. Their function is the trophic support of their neurites.
Early in development two processes emerge from the perikaryon of dorsal root ganglion cells, which merge in the course of development. These ganglion cells are therefore also called pseudounipolar neurones. Two processes emerge from the perikaryon of bipolar neurones. The majority of CNS neurones are multipolar, i.e. more than two processes (but only one axon) emerge from their perikaryon.
Autonomic ganglia do contain synapses, and the ganglion cells within them do have dendrites. They receive synapses from the first neurone of the two-neurone chain, which characterises most of the efferent connections of the autonomic nervous system. The second neurone is the ganglion cell itself. Some autonomic ganglia are embedded within the walls of the organs which they innervate (intramural ganglia - e.g. GIT and bladder).
131

Suitable Slides sections of dorsal root ganglia and autonomic ganglia - H&E, toluidine blue, Giemsa, luxol fast blue/cresyl/violet (LFB/CV)
Dorsal Root Ganglion, cat - H&E and Autonomic Ganglion - H&EGanglion cells will typically be several times larger than other cells in the ganglia. The perikaryon is very large and surrounds a large and light nucleus. Only the cells immediately surrounding the ganglion cells as one flattened layer are satellite cells. With a lot of luck you may see the process of a ganglion cell as it passes out of the capsule of satellite cells. Ganglion cells are of course in contact with other parts of the nervous system and with the peripheral tissues which they innervate. Consequently, nerve fibers will be visible close to or within the ganglion. Lipofuscin, a brownish pigment, accumulates with age not only in ganglion cells, but also in several other cell types of the body, e.g., cardiac muscle cells or endocrine cells.Sketch the appearance of the spinal ganglion section at low magnification. Draw a small section of the spinal ganglion and peripheral ganglion at high magnification. Label ganglion cells, satellite cells and, if visible, nerve fibres and connective tissue elements.
132

Bibliography
1. Junqueira LC; Carneiro J.; Kelly RO: Basic Histology, 11 th edition; LANGE, 2005.
2. Dudek RW: High-Yield Histology, 2nd edition; Lippincott Williams & Wilkins, 2000.
3. Kuehnel W: Color Atlas of Cytology, Histology and Microscopic Anatomy; Thieme, 2003.
4. Lodish H; Baltimore D; Berk A; Zipurski SL; Matsudaira P; Darnell J: Molecular Cell Biology; Scientific American Books Inc., 1995.
5. Ross HM and Reith JE: Histology a text and atlas, Harper International Edition, 1985.
Fitzpatrick TB; Eisen AZ; WOLFF K; Freedberg IM; Austen KF: Dermatology in general medicine, Vol. I., Vol. II; Mc Graw Hill, 1993.
133