gap junctions and cell polarity: connexin32 and...
TRANSCRIPT

2609Journal of Cell Science 108, 2609-2617 (1995)Printed in Great Britain © The Company of Biologists Limited 1995
Gap junctions and cell polarity: connexin32 and connexin43 expressed in
polarized thyroid epithelial cells assemble into separate gap junctions, which
are located in distinct regions of the lateral plasma membrane domain
Annie Guerrier1, Pierre Fonlupt1, Isabelle Morand1, Rachida Rabilloud1, Christine Audebet1,Vladimir Krutovskikh2, Daniel Gros3, Bernard Rousset1 and Yvonne Munari-Silem1,*1Institut National de la Santé et de la Recherche Médicale U369, Faculté de Médecine Alexis Carrel, Rue Guillaume Paradin,69372, Lyon, Cedex 08, France2International Agency for Research on Cancer, 150 cours Albert Thomas, 69372, Lyon, France3Laboratoire de Génétique et Physiologie du développement, UMR C-9943, Faculté des Sciences de Luminy, 13288, Marseille,France
*Author for correspondence
Epithelial cells of the thyroid gland present an uncommonconnexin expression pattern, they coexpress connexin32and connexin43. In the present work, we have analyzed themembrane distribution of these two connexins todetermine: (i) whether they co-assemble in the same gapjunctions or form separate gap junctions; and (ii) whethertheir location is somehow related to the thyroid cellpolarity. Immunofluorescence analyses of the localizationof the two connexins in thyroid tissue sections revealed thatconnexin32 and connexin43 are located in different regionsof the plasma membrane. We further analyzed the locationof each of the two connexins with regard to that of the tightjunction-associated protein, ZO1. Laser scanning confocalmicroscope observations of connexin32 or connexin43 andZO1 double-immunolabelled thyroid cells, gave evidencefor a separate localization of gap junctions made of each ofthese two connexins. Connexin32 gap junctions appearedas fluorescent spots scattered over the lateral membranedomain, while connexin43 gap junctions formed a meshednetwork superimposable with that of tight junctions in the
subapical region of the cells. Western blot analyses of thedistribution of connexins in thyroid plasma membrane sub-fractions obtained by ultracentrifugation on a sucrosegradient led to the identification of membrane sub-popula-tions enriched in either connexin32 gap junctions orconnexin43 gap junctions. Connexin32 gap junctions andconnexin43 gap junctions were found to differ in theirresistance to solubilization by N-lauroylsarcosine. Increas-ing concentrations of this detergent from 0.12% to 0.42%caused a progressive solubilization of connexin43 whileconnexin32 remained membrane-bound. These datademonstrate that connexin32 and connexin43, co-expressedby polarized thyroid cells, form separate gap junctions withdistinct membrane location, gap junctions made up ofconnexin43 being precisely located within tight junctions.We report here, for the first time, a relationship betweenconnexin distribution and cell polarity.
Key words: gap junction, connexin, thyroid
SUMMARY
INTRODUCTION
In organized tissues, cells exchange directly ions and smallmolecules (Mr<1000) through channels which connect thecytoplasm of adjacent cells (Loewenstein, 1981). Suchchannels are assembled into ubiquitous structures named gapjunctions (GJs). Connexins (Cx), which are the exclusive GJconstitutive proteins known until now, belong to a family ofrelated but distinct integral membrane proteins (Beyer, 1992;Kumar and Gilula, 1992). Cx molecules are assembled in theplasma membrane as hexamers named connexons. Connexonsfrom two adjacent cells interlock in the extracellular space toform an entire channel which is therefore composed of twelveCx molecules (Revel and Karnovski, 1967; Makowski et al.,
1977; Milks et al., 1988). From nucleotide and derived aminoacid sequences of the three first and best known Cx, namelyCx32 (Paul, 1986; Kumar and Gilula, 1986), Cx43 (Beyer etal., 1987) and Cx26 (Zhang and Nicholson, 1989), it has beendeduced that the pore of each channel is probably bordered bythe third transmembrane region of each Cx molecule, whichpresents amphiphilic properties (Milks et al., 1988; Bennett etal., 1991).
In the past few years, new members of the Cx family whichmainly differ by their intracellular domains have been discov-ered (Willecke et al., 1991; Haefliger et al., 1992). It is nowestablished that the expression of Cx is tissue- and cell type-dependent (Traub et al., 1989; Willecke et al., 1991; Haefligeret al., 1992; Hoh et al., 1991; Henneman et al., 1992; Kanter

2610 A. Guerrier and others
et al., 1992), many cell types expressing more than one Cx.The underlying reason for multiple Cx expression is so farunknown; it could likely increase the number of possibilitiesfor a given cell type to regulate cell-to-cell communicationwith adjacent cells (Goodenough and Musil, 1993).
We recently found that thyroid epithelial cells present anunusual Cx expression pattern: they coexpress Cx32 and Cx43(Munari-Silem et al., 1994). Studies on pig thyrocytes inprimary culture led us to show that the expression of Cx32, incontrast to that of Cx43, is dependent upon histiotypic mor-phogenesis, i.e. the reconstitution of follicular structures(Munari-Silem et al., 1994). Therefore, thyroid cell monolay-ers only express Cx43 whereas reconstituted thyroid folliclesco-express Cx32 and Cx43. Thyroid cells cultured either asmonolayers or in the form of follicles, reconstitute functionalGJs (Munari-Silem et al., 1990). GJ-mediated cell-to-cell com-munication between thyrocytes is regulated by thyrotropin,TSH, which acts through the cAMP-protein kinase A pathway(Munari-Silem et al., 1991).
Pursuing our detailed analysis of thyroid GJs, we haveinitiated comparative studies of the spatial distribution andproperties of Cx32 and Cx43, in this tissue. We used anti-Cx32and anti-Cx43 antibodies to: (i) localize each of the two Cx inintact tissue sections by immunofluorescence; and (ii) analyzethe Cx content of thyroid membrane subfractions by westernblot. In the present paper, we show that Cx32 and Cx43assemble into GJ plaques, which are distinguishable both mor-phologically and biochemically. The two sets of GJs appearedto be located in different regions of the lateral plasmamembrane of thyroid cells in a way related to cell polarity.
MATERIALS AND METHODS
TissuesOrgans (thyroid, liver and heart) obtained at the local slaughterhousefrom adult pigs were immediately immersed in ice and processedwithin one to two hours after animal death.
AntibodiesAnti-Cx43 antibodies, named anti-Cx43 (314-322) were produced inrabbit against a peptide corresponding to residues 314 to 322 of therat Cx43. These residues correspond to a sequence of the cytoplasmiccarboxy-terminal region of Cx43. The antibodies, affinity-purified onthe immobilized peptide, specifically recognize the Cx43 protein.They have been characterized by western blot, immunofluorescenceand electron microscope immuno-gold labelling (El Aoumari et al.,1990; Fromaget et al., 1990; Giaume et al., 1991).
We have used two different anti-Cx32 antibodies. Anti-Cx32 anti-bodies, named anti-Cx32 (98-124) were produced in rabbit against asynthetic peptide corresponding to a sequence located in the cyto-plasmic loop at rat Cx32 (residues 98-124). The specificity of theseanti-Cx32 antibodies has been previously documented (Munari-Silemet al., 1994). Anti-Cx32 antibodies named anti-Cx32 (266-282) wereproduced in hen against a peptide corresponding to residues 266-282of the carboxy-terminal region of rat Cx32. The affinity-purified anti-bodies specifically recognize the Cx32 protein by western blot andindirect immunofluorescence (Mazet et al., 1992).
The rat monoclonal anti-ZO1 antibody used was the culture super-natant of the clone R26-4C (Stevenson et al., 1986; Anderson et al.,1988), obtained from the Developmental Studies Hybridoma Bankand maintained by the Department of Pharmacology and MolecularSciences, Johns Hopkins University School Medicine, Baltimore,
MD, and the Department of Biology, University of Iowa City, IA,under contract NO1-HD-6-2915 from the NICHD.
Indirect double immunofluorescence labellingsSmall pieces of pig thyroid tissue were cut immediatly after collec-tion of the organ, embedded in Tissue-Teck OCT compound (MilesInc. Elkart, IN, USA) and frozen into liquid nitrogen. Tissue sections(5 µm thick) prepared at −20°C using a Reichert-Jung cryostat(Vienna, Austria), were mounted on gelatinized slides and stored at−20°C until use. Tissue sections were simultaneously fixed and per-meabilized by incubation in 4% (w/v) paraformaldehyde and 0.05%(v/v) Tween-20 in 10 mM phosphate buffered saline (PBS), for 1 hourat room temperature. After extensive washing in PBS, sections wereincubated in PBS containing 1 mg/ml bovine serum albumin (PBS-BSA) for 30 minutes at room temperature.
For Cx32 and Cx43 double labelling, sections were incubatedovernight at 4°C with the rabbit anti-Cx43 (314-322) (3 µg/ml) andthe hen anti-Cx32 (266-282) (3 µg/ml) in PBS-BSA. After extensivewashing, the antigen-antibody complexes were revealed using a TexasRed-conjugated goat anti-rabbit Ig antibody (Amersham, Aylesbury,UK) and a fluorescein-conjugated goat anti-hen Ig antibody(Kirkegaard and Perry Laboratories, Gaithersburg, USA). The Cx32and Z01 double immunolabelling was carried out using the rat anti-Z01 antibody (final dilution 1/50) mixed with the hen anti-Cx32 (266-282) (6 µg/ml). Antigen-antibody complexes were revealed by incu-bating the sections with Texas Red-labelled sheep anti-rat Igantibodies (Amersham, Aylesbury, UK) mixed with fluorescein-labelled goat anti-hen Ig antibodies (Kirkegaard and Perry Laborato-ries, Gaithersburg, USA). The double immunolabelling Cx43 andZO1, was carried out using the rat anti-ZO1 antibody (final dilution1/50) mixed with the rabbit anti-Cx43 (314-322) (3 µg/ml). Antigen-antibody complexes were revealed by incubating the sections withTexas Red-labelled sheep anti-rat Ig antibodies mixed with fluor-escein-labelled goat anti-rabbit Ig antibodies (Sigma, St Louis, USA).For controls, each primary antibody was replaced by the correspond-ing fraction of the preimmune sera.
Conventional fluorescence microscopyFluorescence images were taken using a SIT camera (Lhesa elec-tronique, Cergy Pontoise, France), installed on an Axiophot micro-scope equipped with the appropriate excitation-emission filters forTexas Red or fluorescein (Zeiss, Oberkochen, Germany). Images werenumerized using an image processing system (Sapphire from Quantel,Montigny-le Bretonneux, France) and stored on a Bernoulli box. Pho-tomicrographs were then prepared using a video printer (UP 5000 Pfrom Sony, Tokyo, Japan).
Laser scanning confocal fluorescence microscopyThe three-dimensional distribution of fluorescent structures wasanalyzed using a laser scanning confocal microscope (LSM fromZeiss) equipped with a PI-Neofluar (40×) oil immersion objective(NA=1.3) (Centre Commun de Quantimétrie, Université ClaudeBernard, Lyon, France). Observations were made on single opticalsections or on serial optical sections taken at 1 µm focus steps in thez-axis of the tissue sections. fluorescein and Texas Red fluorescenceimages from double-labelled samples were collected sequentiallyusing an argon (488 nm) laser beam for excitation and a broad pass(515-545 nm) emission filter for fluorescein and an helium/neon (543nm) laser beam for excitation and a long-pass (590 nm) emission filterfor Texas Red. We verified that there was no mechanically inducedshift during the procedure of sequential image collection. The simul-taneous detection of a given antigen-antibody complex with a fluor-escein-labelled conjugate and a Texas Red-labelled conjugate yieldedsuperimposable fluorescence images. We also verified that there wasno contribution of the fluorescence from one fluorophore to the imageof the other. The image dimensions were 512×512 pixels with 8 bitsresolution.

2611Gap junctions and cell polarity
Images were transferred on a 80486-based IBM compatiblepersonal computer equipped with a Cyclope imaging card (DigitalVision, Chatillon, France). Softwares, performed using turbo Pascal(Borland), were designed for image noise reduction and pseudocolourimage restitution (green for fluorescein and red for Texas Red).Double-labelled images were obtained by pseudocolour image resti-tution from the original black and white images corresponding to eachof the two fluorophores (4 bits resolution for each colour).
Preparation of crude thyroid plasma membrane fractions All tissue fractionation steps were carried out at 0-4°C. The proceduredescribed here was designed for about 250-300 g of pig thyroid tissue.Thyroid glands were minced with a razor blade and placed in 1 mMNaHCO3, pH 8.0, containing 1 mM phenylmethyl sulphonyl fluoride(PMSF) (buffer A). The minced tissue was homogenized in buffer Aby forcing it through a metallic sieve (pore size: 0.3 mm). Thishomogenization procedure led to the disruption of thyroid folliclesand release of intralumenal protein, i.e. thyroglobulin (Alquier et al.,1985). The material passing through the sieve was diluted to 2 litresin buffer A and centrifuged at 700 g for 15 minutes. The supernatantwas discarded. The cell pellet was resuspended in 500 ml of buffer Aand homogenized with a glass/glass Potter homogenizer rotated at 800revs/min with about 10 up-and-down strokes. The homogenate wascentrifuged at 700 g for 15 minutes. The supernatant was kept at 4°C.The pellet was resuspended in 300 ml of buffer A, homogenized againas mentioned above and centrifuged at 700 g for 15 minutes The two700 g supernatants were pooled and centrifuged at 100,000 g for 30minutes. The resulting 100,000 g pellets were resuspended in bufferA. The suspension (75 ml) was mixed with 150 ml of buffer A con-taining 67% (w/v) sucrose (to reach a final sucrose concentration of45%) and distributed in twelve 25 ml centrifugation tubes. The sus-pension was overlayed with 7 ml of buffer A containing 30% (w/v)sucrose. The tubes were then centrifuged at 140,000 g for 90 minutesin a 50.2 Ti Beckman rotor. The material floating at the 30% to 45%sucrose interface was collected, diluted in buffer A and centrifuged at31,300 g for 30 minutes. The final pellet named ‘crude plasmamembrane fraction’ was washed in buffer A and stored at −20°C.
Isolation of GJs from thyroidGJs were isolated from crude thyroid plasma membrane fractionsusing the method of Hertzberg and Gilula (1979). All steps wereperformed in the presence of an additional protease inhibitor,leupeptin (20 µM) (buffer A + leupeptin: buffer B). Leupeptin wasadded in order to inhibit membrane proteases that could be activatedby detergents (Guenet et al., 1982). Briefly, crude thyroid plasmamembranes were washed in buffer B supplemented with 0.1 M NaCland centrifuged at 12,000 g for 15 minutes. The pellets were resus-pended in buffer B and the resulting membrane suspension was mixedwith an equal volume of 0.9% (w/v) N-lauroylsarcosine sodium salt(Sigma, St Louis, USA) in buffer B (final concentration: 0.45%). Thesuspension was sonicated, allowed to stand for 20 minutes at roomtemperature and centrifuged at 31,000 g for 20 minutes. The pelletscontaining the N-lauroylsarcosine-resistant membranes werecollected and washed with 0.1 M Na2CO3, pH 11. After centrifuga-tion at 31,000 g for 20 minutes, the pellet was resuspended in bufferB supplemented with 15% (w/v) sucrose and 1 M urea. This mixturewas overlayed on top of a discontinuous sucrose gradient constitutedby 60%, 45% and 30% sucrose (w/v) in buffer B containing 1 M urea.After centrifugation at 195,000 g for 90 minutes, the GJ-enrichedmaterial banding at the 30% to 45% sucrose interface was collected,washed in buffer B and stored at −20°C.
Isolation of GJs from liver and heartLiver GJs and heart GJs were prepared from 300 g of pig liver or pigheart as described by Hertzberg and Gilula (1979) and Manjunath etal. (1984), respectively.
SDS-polyacrylamide gel electrophoresis and western blotMembrane proteins were fractionated by SDS-PAGE according toLaemmli (1970) using 4.5% acrylamide for the stacking gel and 11%acrylamide in the separating gel. Samples were solubilized in 62.5mM Tris-HCI, pH 6.8, containing 16% (v/v) glycerol, 10% (v/v) β-mercaptoethanol and 4% (w/v) SDS for 30 minutes at room temper-ature. After migration, proteins were either stained with CoomassieBrillant Blue R-250 or transferred onto Immobilon-P membrane(Millipore, Bedford, USA). Electro-transfer was carried out in 25 mMTris, 192 mM glycine, 20% (v/v) methanol, for 50 minutes at 90 V.At the end of the transfer, Immobilon-P membranes were pre-incubated in a saturation solution (20 mM phosphate buffer, pH 7.4,containing 0.15 M NaCl, 2% (w/v) skim-milk and 0.2% (v/v) Tween-20) for 30 minutes at room temperature and then incubated with theanti-Cx antibodies (rabbit anti-Cx32 (98-124): final dilution 1/1,000or purified rabbit anti-Cx43 (314-322): 4 µg/ml) for one hour at roomtemperature. After extensive washing, the blots were incubated witha goat anti-rabbit IgG antibody conjugated to alkaline phosphatase(Sigma, St Louis, USA) for 1 hour at room temperature. Immunecomplexes were revealed according to the established procedure ofHarlow and Lane (1988), using bromochloroindolyl phosphate/nitroblue tetrazolium (Sigma, St Louis, USA) as substrate. Control exper-iments were performed by replacing the first antibody by the corre-sponding fraction of preimmune sera. Standards of known relativemolecular mass (Bio-Rad Laboratories, Richmond, CA) were: phos-phorylase b, 94,400; bovine serum albumin, 66,200; ovalbumin,42,700; carbonic anhydrase, 31,000; soybean trypsin inhibitor, 21,500and lysosyme, 14,400.
Other methodsProtein content of crude membranes and N-lauroylsarcosine-treatedmembranes was determined by the method of Lowry et al. (1951),using bovine serum albumin (Boehringer Mannheim, Germany) asstandard. Particulate samples were pre-treated with 1% sodiumdeoxycholate for 1 hour at room temperature. N-lauroylsarcosine, atconcentration up to 0.45%, did not interfere in the protein assay.
Quantitative analyses of western blots was performed using animage-processing system (Crystal Sapphire from Quantel, Montigny-le-Bretonneux, France).
RESULTS
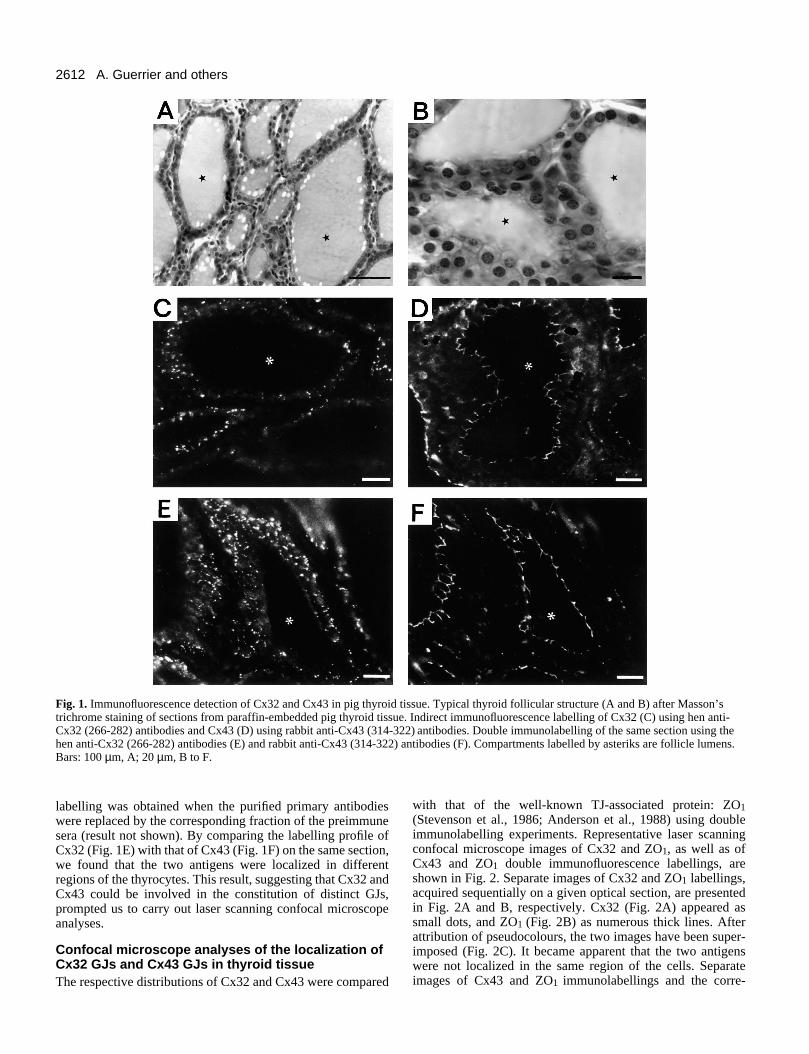
The typical cell organization of the thyroid gland is illustratedin Fig. 1A and B. Single layers of polarized epithelial cellsform spherical structures, the thyroid follicles, and delimitclose compartments, the follicle lumens, which contain thethyroid prohormone, thyroglobulin. Contiguous follicles areseparated by connective tissue. The follicles exhibit variablesize and shape (Fig. 1A). Among the follicles shown at a highermagnification (Fig. 1B), one follicle has been cut close to thepole in such a way that one can see a portion of the thyroidcell epithelium.
Visualization of thyroid GJs by single or doubleimmunofluorescence labelling of CxGJs between thyroid epithelial cells were detected by single(Fig. 1C,D) or double (Fig. 1E,F) immunofluorescencelabelling of Cx on cryosections using either hen anti-Cx32(266-282) or rabbit anti-Cx43 (314-322) or both antibodies.
The anti-Cx32 (266-282) labelled numerous bright spots,scattered within the cell layers forming the follicles (Fig.1C,E). In contrast, anti-Cx43 (314-322) labelled continuousbright lines located close to the follicle lumens (Fig. 1D,F). No
2612 A. Guerrier and others
Fig. 1. Immunofluorescence detection of Cx32 and Cx43 in pig thyroid tissue. Typical thyroid follicular structure (A and B) after Masson’strichrome staining of sections from paraffin-embedded pig thyroid tissue. Indirect immunofluorescence labelling of Cx32 (C) using hen anti-Cx32 (266-282) antibodies and Cx43 (D) using rabbit anti-Cx43 (314-322) antibodies. Double immunolabelling of the same section using thehen anti-Cx32 (266-282) antibodies (E) and rabbit anti-Cx43 (314-322) antibodies (F). Compartments labelled by asteriks are follicle lumens.Bars: 100 µm, A; 20 µm, B to F.
labelling was obtained when the purified primary antibodieswere replaced by the corresponding fraction of the preimmunesera (result not shown). By comparing the labelling profile ofCx32 (Fig. 1E) with that of Cx43 (Fig. 1F) on the same section,we found that the two antigens were localized in differentregions of the thyrocytes. This result, suggesting that Cx32 andCx43 could be involved in the constitution of distinct GJs,prompted us to carry out laser scanning confocal microscopeanalyses.
Confocal microscope analyses of the localization ofCx32 GJs and Cx43 GJs in thyroid tissueThe respective distributions of Cx32 and Cx43 were compared
with that of the well-known TJ-associated protein: ZO1(Stevenson et al., 1986; Anderson et al., 1988) using doubleimmunolabelling experiments. Representative laser scanningconfocal microscope images of Cx32 and ZO1, as well as ofCx43 and ZO1 double immunofluorescence labellings, areshown in Fig. 2. Separate images of Cx32 and ZO1 labellings,acquired sequentially on a given optical section, are presentedin Fig. 2A and B, respectively. Cx32 (Fig. 2A) appeared assmall dots, and ZO1 (Fig. 2B) as numerous thick lines. Afterattribution of pseudocolours, the two images have been super-imposed (Fig. 2C). It became apparent that the two antigenswere not localized in the same region of the cells. Separateimages of Cx43 and ZO1 immunolabellings and the corre-

2613Gap junctions and cell polarity
Fig. 2. Confocal microscope images of pig thyroid tissue double-labelled with anti-Cx (Cx32 or Cx43) and anti-ZO1 antibodies.Cryosections from intact pig thyroid tissue (5 µm thick) were fixed,permeabilized and immunolabelled simultaneously for Cx and ZO1antigens, as described in Materials and Methods. Left panels, Cx32and ZO1 double immunolabelling; right panels, Cx43 and ZO1double immunolabelling. In each case, the image of fluorescein-labelled Cx was first acquired and stored (image A for Cx32; imageD for Cx43). The corresponding images of Texas Red-labelled ZO1were then acquired and stored separately (images B and E). ImagesC and F were obtained by the superimposition of images A and B,and D and E, respectively. Appropriate pseudocolours wereattributed using the computer programme described in Materials andMethods. Cx labelling appears in green and ZO1 labelling in red.Regions of co-localization of Cx and ZO1 appear in orange/yellow.The asterisk indicates the position of the follicle lumen. Bar, 25 µm.
Fig. 3. Confocal microscope serial optical sections of pig thyroidtissue double-labelled for Cx (Cx32 or Cx43) and ZO1. Cryosectionsfrom intact thyroid tissue (5 µm thick) were fixed, permeabilized andimmunolabelled simultaneously with anti-Cx (green fluorescence)and anti-ZO1 (red fluorescence) antibodies as described in Materialsand Methods. (A) Cx32 and ZO1 double labelling; (B) Cx43 andZO1 double labelling. Images a to d correspond to four consecutiveoptical sections, one µm apart. The orange/yellow colour indicates aco-localization of the two fluorophores. Bar, 10 µm.
A B
sponding superimposed image are presented in Fig. 2D,E andF, respectively. Cx43 appeared as rather thick broken lines andZO1 as continuous lines. The appearance of a yellow/orangecolour in Fig. 2F gives evidence of a complete overlapping ofCx43 and ZO1 immunolabellings and indicates, therefore acolocalization of the two antigens.
To determine precisely the degree of co-localization of Cx43and ZO1, we have performed a complementary analysis onserial optical sections at higher magnification. The differentpanels of Fig. 3A show the Cx32 and ZO1 immunolabellingpatterns obtained on 4 consecutive sections, 1 µm apart. Cx32(green spots) and ZO1 (red lines) exhibited a different distrib-
ution. These observations confirmed that Cx32 GJs were notlocated in the vicinity of TJs. A similar approach was used tocompare the tissue distribution of Cx43 with that of ZO1 (Fig.3B). The Cx43 immunolabelling in green was only observedon 3 consecutive optical sections, whereas the ZO1 immuno-labelling in red was clearly present on the 4 consecutive

2614 A. Guerrier and others
Fig. 4. Fractionation of thyroid plasma membranes byultracentrifugation on a sucrose gradient. Identification of sub-fractions enriched in either Cx32 or Cx43. Crude thyroid plasmamembrane fractions were resuspended in buffer B containing 0.22%(w/v) N-lauroylsarcosine. The suspension was sonicated and allowedto stand for 20 minutes at room temperature. Membranes werecollected by centrifugation at 100,000 g for 50 minutes at 4°C,washed in buffer B and finally resuspended in buffer B containing30% (w/v) sucrose and 1 M urea, to reach a final proteinconcentration of 1 mg/ml. After sonication, 1 ml of the membranesuspension was overlaid on the top of a sucrose gradient, prepared inbuffer B containing 1 M urea as follows: 0.5 ml of 60% (w/v)sucrose and 11 ml of a linear 40% to 20% (w/v) sucrose gradient.After centrifugation at 195,000 g for 90 minutes at 4°C, gradientswere collected by fractions of 0.5 ml and the absorbance at 340 nmwas measured. Fractions were then pooled three by three andsupplemented with buffer B to decrease sucrose concentration. Theparticulate material present in each pool was collected bycentrifugation at 100,000 g for 50 minutes at 4°C and assayed forprotein and analyzed for its Cx content by western blot. (A)Membrane distribution along the gradient assessed by absorbancemeasurements at 340 nm (continuous line); sucrose concentrationprofile (broken line). (B) Western blot analyses of the Cx32 andCx43 content of each membrane sub-fraction (pool I to pool VIII).The amount of protein loaded in each lane was proportional to theprotein content of each pool. Cx were immunodetected using rabbitanti-Cx43 (314-322) and rabbit anti-Cx32 (98-124) antibodies. (Aand B) Results of a representative experiment. (C) Quantitativeanalysis of the distribution of Cx32 (black columns) and Cx43 (opencolumns) in thyroid plasma membrane sub-fractions along thegradient. Densitometric measurements on western blots obtainedfrom three membrane fractionation experiments. Columns andvertical bars represent the mean ± s.e.m.
sections (panels a, b, c and d). Interestingly, Cx43 labellingwas more intense in one optical section (section b) than in thethree other sections. In this section, Cx43 and ZO1 labellingsoverlapped in most place as indicated by the yellow/orangecolour. The Cx43 labelling appeared thicker than that of ZO1and extended somewhat beyond that of ZO1. These observa-tions show that Cx43 GJs are rather homogeneously distrib-uted around each cell and segregated within the TJ belt.
Separation of thyroid plasma membrane sub-fractions enriched in either of Cx32 GJs or Cx43 GJs Cx32 and Cx43 distribution among membrane sub-fractionsobtained by ultracentrifugation on a sucrose gradient wasanalyzed by western blot (Fig. 4). Fig. 4A shows the membraneprotein profile and Fig. 4B the distribution of Cx (Cx43 andCx32) along the gradient in a representative experiment. Thebulk of protein remained in the first half of the gradient. Thedistribution of Cx32 and Cx43 along the gradient did notcoincide. Cx43, in its different isoforms, was mainly found inmembrane sub-fractions floating between 15% and 30%sucrose. Cx43 was present in very low amount in the secondhalf of the gradient corresponding to high sucrose concentra-tions. Cx32 peaked in the middle of the gradient around 30%sucrose. These differences in the distribution of Cx32 andCx43 among thyroid plasma membrane sub-fractions werereproducibly observed. Fig. 4C gives the results of the densit-ometric analysis of western blots obtained in three separatemembrane fractionation experiments such as that presented inFig. 4A and B.
Thyroid Cx32 GJs and Cx43 GJs differ in theirresistance to solubilization by N-lauroylsarcosineThe connexin composition of pig thyroid GJs was comparedto that of pig liver GJs and pig heart GJs (Fig. 5). Rabbit anti-Cx32 (98-124) antibodies strongly labeled a 28 kDa antigen inthyroid GJ-enriched fractions (Fig. 5A, lane T). The antibodiesalso reacted with a molecular species in the 50-55 kDa region,which corresponds to Cx32 dimers (Hertzberg and Gilula,1979; Green et al., 1988). A similar, if not identical, profilewas obtained with GJs isolated from liver (Fig. 5A, lane L).Anti-Cx32 antibodies failed to detect any antigen in GJfractions isolated from heart (Fig. 5A, lane H). As expected,the anti-Cx43 antibodies labelled several closely located bandswith an apparent molecular mass of 43-46 kDa (Fig. 5B, laneH) in isolated heart GJs. These bands, corresponding to theknown isoforms of Cx43 (Musil et al., 1990; Musil and Good-enough, 1991) were not found in isolated liver GJs (Fig. 5B,lane L) and only trace amounts of Cx43 antigens were detectedin thyroid GJ-enriched fractions (Fig. 5B, lane T). To under-stand why Cx43 was not present in thyroid Gj-enrichedfractions, we followed the fate of each of the two Cx at thedifferent steps of the isolation procedure. We found that Cx43GJs were lost at the step of treatment of membranes by N-lau-roylsarcosine. This is shown in Fig. 6. Crude thyroidmembrane preparations were submitted to increasing concen-trations of N-lauroylsarcosine and the distribution of Cx32 andCx43 between solubilized and detergent-resistant membraneprotein fractions was analyzed by western blot. Increasing con-centrations of N-lauroylsarcosine from 0.12 to 0.42% very effi-ciently solubilized membrane proteins. At the concentration of0.42% and at a detergent/protein mass ratio of about 2, more

2615Gap junctions and cell polarity
than 97% of membrane protein were found in the detergent-solubilized fraction (Fig. 6A). N-lauroylsarcosine, up to 0.32%,did not extract Cx32 from membranes (Fig. 6B). At the highestconcentration, i.e. 0.42% (the concentration corresponding tothat used in the GJ isolation procedure), more than 90% ofimmunoreactive Cx32 was found in the detergent-resistantmembrane protein fraction. This detergent treatment step ledto a 20- to 30-fold purification of Cx32 GJs. In contrast, theCx43 content of thyroid membranes was already decreased ata concentration of detergent of 0.12% which caused the solu-bilization of only 50-60% of membrane proteins. At this con-centration, N-lauroylsarcosine induced the selective and almostcomplete solubilization of the Cx43 isoform with the lowestapparent molecular mass, described as the non-phosphorylatedform of Cx43 (Musil et al., 1990). N-lauroylsarcosine, from0.12 to 0.42%, caused a progressive solubilization of the Cx43isoform(s) of higher molecular mass. At 0.42%, almost allCx43 was found in the solubilized membrane protein fraction.
DISCUSSION
The present data, based on complementary immunocytologicaland biochemical approaches, unequivocally demonstrate thatCx32 and Cx43 form distinct GJs which are located in differentmembrane regions of the polarized thyroid cells. Cx43 GJs arelocated in the subapical membrane region within the TJ beltand Cx32 GJs are randomly distributed along the lateralmembrane domain of the cells.
Thyroid cells represent, at the moment, the only cell typeknown to co-express Cx32 and Cx43. In other endocrineglands, Cx43 is present in association with variable levels ofCx26 but no Cx32 is found (Meda et al., 1993). We did notdetect any Cx26 in pig thyroid cells. In contrast, exocrineglands express high levels of Cx32 with variable levels of
Fig. 5. Cx content of isolated thyroid, liver and heart GJs. GJ-enriched membrane fractions prepared from pig thyroid (T), pig liver(L) and pig heart (H) were analyzed by western blot. About 10 µgprotein was loaded in each lane. The immunodetection of Cx32 (A)and Cx43 (B) was performed using rabbit anti-Cx32 (98-124) andrabbit anti-Cx43 (314-322) antibodies, respectively. The black arrowindicates the position of Cx32 and the arrowhead the position of thedimer of Cx32. The open arrow indicates the position of the Cx43isoforms. The positions of markers of known relative molecular massare indicated on the side of each panel (×10−3).
Cx26, but no Cx43 (Meda et al., 1993). Thus, the thyroid gland,by coexpressing Cx32 and Cx43, resembles partly otherendocrine glands and partly exocrine glands. It is noteworthythat thyroid cells have an exocrine function by exporting thethyroid prohormone, thyroglobulin, into the follicular lumen,and a typical endocrine function by releasing thyroid hormonesinto the vascular compartment. In the three best-studied celltypes that express several Cx (two or more), cardiomyocytes,hepatocytes and lens fiber cells, a given GJ plaque containsmore than one Cx. In cardiomyocytes, Cx43 and Cx40 on theone hand, and Cx43 and Cx45 on the other, have been foundin the same plaques (Kanter et al., 1992, 1993; Gros et al.,1994). In hepatocytes, Cx32 and Cx26 are present in the sameGJs (Nicholson et al., 1987; Traub et al., 1989). Similarly, inlens fiber cells, Cx46 and Cx50 (also known as MP70) arelocated in the same junctional macula (Paul et al., 1991; Whiteet al., 1992). Thyroid cells, like some other cells, express Cxthat assembles into separate GJs. The Cx43 and Cx26expressed in leptomeningeal cells (Spray et al., 1991) and in
Fig. 6. Effects of increasing concentrations of N-lauroylsarcosine onthe Cx content of thyroid plasma membranes. Crude thyroid plasmamembranes (12 mg protein), prepared as described in Materials andMethods, were suspended in 7 ml of buffer A containing 0.12% (a),0.22% (b), 0.32% (c) or 0.42% (d) N-lauroylsarcosine. After 20minutes at room temperature, the membrane suspensions werecentrifuged at 100,000 g for 30 minutes. (A) Protein content of thepellet (p) and the corresponding supernatant (s) expressed as % oftotal protein. (B) Western blot analysis of Cx32 and Cx43 inuntreated thyroid plasma membrane fraction (control) and in p and sfractions using rabbit anti-Cx32 (98-124) and rabbit anti-Cx43 (314-322) antibodies. In each experimental condition (a, b, c and d), theamount of protein loaded in p and s lanes represents 1/70th of theamount of protein recovered in these fractions. A non-specific bandof unknown origin (found throughout the blot), but often seen, wasobserved on Immobilon-P membranes incubated with Cx43 (314-322) antibodies. The arrows on the right of the figure identify the Cxpolypeptides.

2616 A. Guerrier and others
keratinocytes (Kamibayashi et al., 1993) form separate GJs.One cannot exclude the possibility of the presence of anotherunidentified Cx in these GJs, which may be formed from asingle Cx. It can be inferred from our study that Cx32 and Cx43probably cannot assemble into the same connexon or group ofconnexons. Indeed, being synthesized simultaneously inthyroid cells (Munari-Silem et al., 1994), the two Cx should bein a position to co-assemble (in one connexon) on themembrane of the endoplasmic reticulum, if this molecularevent could take place.
The most important finding of our study is the demonstra-tion of the presence of a definite subset of GJ, Cx43 GJ, withinthe TJ. The existence of GJ within the TJ lattice of variousepithelial tissues, including thyroid tissue, has been known fora long time (Goodenough and Stoeckenius, 1972; Friend andGilula, 1972; Thiele and Reale, 1976). We report here that TJ-associated GJs differ in their Cx composition from the GJslocated elesewhere on the lateral membrane domain of thepolarized thyroid cells. Cx43 GJs are only found within TJ andCx32 GJs are found outside TJ. These observations give thefirst evidence that GJ distribution is related to cell polarity. Onemay consider the possibility of a selective delivery of Cx43connexons to pre-existing TJ or the selection of plasmamembrane Cx43 connexons at the time of TJ assembly. Thepresence of Cx43 GJ within TJ could also result from theexclusion of Cx32 connexons from this specialized membranedomain. Information on the distribution of Cx in other epitheliais now required to determine whether our finding correspondsto a general property of the polarized cell. It is worth notingthat Cx43 has been found in kidney and intestine. Preliminaryobservations (D. Gros) on mouse intestine suggest that Cx43could be present in TJ of the polarized intestinal cell. Thepresence of Cx43 in liver is a matter of debate.
Cx32 GJs and Cx43 GJs, which are distinct entities on thebasis of morphological approaches, are biochemically distin-guishable. First, thyroid plasma membrane sub-fractionsbearing Cx32 GJs exhibit a higher buoyant density than plasmamembrane sub-fractions bearing Cx43 GJs. Second, Cx32 GJshave a higher detergent stability than Cx43 GJs. These datasupport the conclusions drawn from immunofluorescenceanalyses and give evidence for physicochemical differencesbetween the two types of GJs. Our results, showing a differen-tial susceptibility of Cx32 GJs and Cx43 GJs to N-lauroylsar-cosine from the same cells, extend the observations made byGoodenough and Stoeckenius (1972) and Kensler and Goode-nough (1980) on liver GJs and heart GJs. These authors reportedthat heart GJs are less resistant to N-lauroylsarcosine than liverGJs. These differences do not seem to be related to intrinsicstructural properties of the constitutive proteins. Indeed, Cx32and Cx43, which have the same overall structure (Zimmer etal., 1987; Yeager and Gilula, 1992), differ mainly by the lengthof their carboxy-terminal cytoplasmic extension, a differencewhich should be of little importance for protein-lipid or protein-detergent interactions. The differences of buoyant density anddetergent sensitivity could rather result from differences in themolecular environment of the two types of GJs. Variations,even discrete, in the composition of the lipid bilayer within thejunctions or in their immediate environment (Malewicz et al.,1990) could contribute to the differential sedimentationbehaviour and stability to detergent. The marked difference inthe apparent buoyant density of plasma membrane sub-fractions
with either a high Cx43 content or a high Cx32 content suggeststhat the two membrane sub-fractions could differ in theirlipid/protein ratio. Finally, the differential biochemical proper-ties of the two types of GJs might be related to their distinctlocation within the lateral membrane domain of the thyroidcells. It is indeed reasonable to think that the stability of Gjscould be affected by their physical association with TJs, or bythe presence of TJ structures in their near surroundings. Theprotein and lipid composition of the plasma membrane sub-domains forming the TJ belt, although still largely unknown, isexpected to be different from that of the other parts of the baso-lateral domain of the polarized epithelial cells (Furuse et al.,1993; Citi, 1993; Anderson et al., 1993).
What could be the physiological significance of the presenceof two distinct sets of GJs? The only known function of GJchannels is to allow the cell-to-cell transfer of small cytoplas-mic molecules. One might consider that the pores formed byeither of these GJs could exhibit different permeability prop-erties (Steinberg et al., 1994). GJ channels present rapid tran-sitions between open and closed states, and vice versa, inresponse to intracellular signals. Different Cx are expected tohave different sensitivity to these various gating stimuli (Saezet al., 1993). The presence of GJs composed of different Cxcould provide a given cell population with different modes ofcontrol of cell-to-cell coupling.
We thank Dr C. Souchier for her contribution to the laser scanningconfocal microscope analysis. The authors also thank Dr Yamasakifor providing rabbit anti-Cx32 antibodies and Mrs T. Jarry-Guichardfor the purification of rabbit anti-Cx43 antibodies and hen anti-Cx32antibodies. This work was supported by a grant from the Associationpour la Recherche sur le Cancer (grant no. 3070).
REFERENCES
Alquier, C., Guenin, P., Munari-Silem, Y., Audebet, C. and Rousset, B.(1985). Isolation of pig thyroid lysosomes. Biochemical and morphologicalcharacterization. Biochem. J. 232, 529-537.
Anderson, J. M., Stevenson, B. R., Jesaitis, L. A., Goodenough, D. A. andMooseker, M. (1988). Characterization of ZO-1, a protein component of thetight junction from mouse liver and Madin-Darby canine kidney cells. J. CellBiol. 106, 1141-1149.
Anderson, J. M., Balda, M. S. and Fanning, A. S. (1993). The structure andregulation of tight junctions. Curr. Opin. Cell Biol. 5, 772-778.
Bennett, M. V. L., Barrio, L. C., Bargiello, T. A., Spray, D. C., Hertzberg,E. and Saez, J. C. (1991). Gap junctions: new tools, new answers, newquestions. Neuron 6, 305-320.
Beyer, E. C. (1992). Gap junctions. Int. Rev. Cytol. 137C, 1-37. Beyer, E. C., David, L. P. and Goodenough, D. A. (1987). Connexin 43: A
protein from rat heart homologous to a gap junction protein from liver. J. CellBiol. 105, 2621-2629.
Citi, S. (1993). The molecular organization of tight junctions. J. Cell Biol. 121,485-489.
El Aoumari, A., Fromaget, C., Dupont, E., Reggio, H., Durbec, P., Briand,J. P., Böller, K., Kreitman, B. and Gros, D. (1990). Conservation of acytoplasmic carboxy-terminal domain of connexin 43, a gap junctionalprotein, in mammal heart and brain. J. Membrane Biol. 115, 229-240.
Friend, D. S. and Gilula, N. B. (1972). Variations in tight and gap junctions inmammalian tissues. J. Biol. Chem. 53, 758-776.
Fromaget, C., El Aoumari, A., Dupont, E., Briand, J. P. and Gros, D.(1990). Changes in the expression of connexin 43, a cardiac gap junctionalprotein, during mouse heart development. J. Mol. Cell Cardiol. 22, 1245-1258.
Furuse, M., Hirase, T., Itoh, M., Nagafuchi, A., Yonemura, S., Tsukita, S.and Tsukita, S. (1993). Occludin: a novel integral membrane proteinlocalizing at tight junctions. J. Cell Biol. 123, 1777-1788.

2617Gap junctions and cell polarity
Giaume, C., Fromaget, C., El Aoumari, A., Cordier, J., Glowinski, J. andGros, D. (1991). Gap junctions in cultured astrocytes: single-channelcurrents and characterization of channel-forming protein. Neuron 6, 133-143.
Goodenough, D. A. and Stoeckenius, W. (1972). The isolation of mousehepatocyte gap junctions. J. Cell Biol. 54, 646-656.
Goodenough, D. A. and Musil, L. S. (1993). Gap junctions and tissuebusiness: problems and strategies for developing specific functionalreagents. J. Cell Sci. 17, 133-138.
Green, C. R., Harfst, E., Gourdie, R. G. and Severs, N. J. (1988). Analysis ofthe rat liver gap junction protein: clarification of anomalies in its molecularsize. Proc. R. Soc. Lond. B 233, 165-174.
Gros, D., Jarry-Guichard, T., Ten Velde, I., De Maziere, A., Van Kempen,M. J. A., Davoust, J., Briand, J. P., Moorman A. F. M. and Jongsma, H.J. (1994). Restricted distribution of connexin 40, a gap junctional protein, inmammalian heart. Circ. Res. 74, 839-851.
Guenet, L., Leray, G., Godet, J. P., Le Treut, A. and Le Gall, J. Y. (1982).Evidence for intrinsic proteolytic activity in rat liver plasma membranes.Biochem. Biophys. Res. Commun. 108, 486-494.
Haefliger, J. A., Bruzzone, R., Jenkins, N. A., Gilbert, D. J., Copeland, N.G. and Paul, D. L. (1992). Four novel members of the connexin family ofgap junction proteins. Molecular cloning, expression and chromosomemapping. J. Biol. Chem. 267, 2057-2064.
Harlow, E. and Lane, D. (1988). Immunoblotting detection with alkalinephosphatase. In Antibodies, a Laboratory Manual, p. 505, Cold SpringHarbor Laboratory, NY.
Hennemann, H., Schwarz, H. J. and Willecke, K. (1992). Characterization ofgap junction genes expressed in F9 embryonic carcinoma cells: molecularcloning of mouse connexin31 and -45 cDNAs. Eur. J. Cell Biol. 57, 51-58.
Hertzberg, E. L. and Gilula, N. B. (1979). Isolation and characterization ofgap junctions from rat liver. J. Biol. Chem. 254, 2138-2147.
Hoh, J. H., John, S. A. and Revel, J. P. (1991). Molecular cloning andcharacterization of a new member of the gap junction gene family,connexin31. J. Biol. Chem. 266, 6524-6531.
Kamibayashi, Y., Oyamada, M., Oyamada, Y. and Mori, M. (1993).Expression of gap junction proteins connexin26 and 43 is modulated duringdifferenciation of keratinocytes in newborn mouse epidermis. J. Invest.Derm. 101, 773-778.
Kanter, H. L., Saffitz, J. E. and Beyer, E. C. (1992). Cardiac myocytesexpress multiple gap junction proteins. Circ. Res. 70, 438-444.
Kanter H. L., Laing, J. G., Beyer, E. C., Green, K. G. and Saffitz, J. E.(1993). Multiple connexins colocalize in canine ventricular myocyte gapjunctions. Circ. Res. 73, 344-350.
Kensler, R. W. and Goodenough, D. A. (1980). Isolation of mousemyocardial gap junctions. J. Cell Biol. 86, 755-764.
Kumar, N. M. and Gilula, N. B. (1986). Cloning and characterization ofhuman and rat liver cDNAs coding for a gap junction protein. J. Cell Biol.103, 767-776.
Kumar, N. M. and Gilula, N. B. (1992). Molecular biology and genetics of gapjunction channels. Semin. Cell Biol. 3, 3-16.
Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly ofthe head of bacteriophage T4. Nature 227, 680-685.
Loewenstein, W. R. (1981). Junctional intercellular communication: the cell-to-cell membrane channel. Physiol. Rev. 61, 829-913.
Lowry, O. H., Rosebrough, N. J., Farr, A. L. and Randall, R. J. (1951).Protein measurement with the Folin phenol reagent. J. Biol. Chem. 193, 264-275.
Makowski, L., Caspar, D. L. D., Phillips, W. C. and Goodenough, D. A.(1977). Gap junction structures. II. Analysis of the X-ray diffraction data. J.Cell Biol. 74, 629-645.
Malewicz, B., Kumar, N. M., Johnson, R. G. and Baumann, W. J. (1990).Lipids in gap junction assembly and function. Lipids 25, 419-427.
Manjunath, C. K., Goings, G. E. and Page, E. (1984). Cytoplasmic surfaceand intramembrane components of rat heart gap junctional proteins. Am. J.Physiol. 246, H865-H875.
Mazet, J. L., Jarry, T., Gros, D. and Mazet, F. (1992). Voltage dependence ofliver gap-junction channels reconstituted into liposomes and incorporatedinto planar bilayers. Eur. J. Biochem. 210, 249-256.
Meda, P., Pepper, M. S., Traub, O., Willecke, K., Gros, D., Beyer, E. C.,Nicholson, B., Paul, D. and Orci, L. (1993). Differential expression of gapjunction connexins in endocrine and exocrine glands. Endocrinology 133,2371-2378.
Milks, L. C., Kumar, N. M., Houghten, R., Unwin, N. and Gilula, N. B.
(1988). Topology of the 32-kd liver gap junction protein determined by site-directed antibody localizations. EMBO J. 7, 2967-2975.
Munari-Silem, Y., Mesnil, M., Selmi, S., Bernier-Valentin, F., Rabilloud,R. and Rousset, B. (1990). Cell-cell interactions in the process ofdifferentiation of thyroid epithelial cells into follicles: a study bymicroinjection and fluorescence microscopy on in vitro reconstituted thyroidfollicles. J. Cell. Physiol. 145, 414-427.
Munari-Silem, Y., Audebet, C. and Rousset, B. (1991). Hormonal control ofcell to cell communication: regulation by thyrotropin of the gap junction-mediated dye transfer thyroid cells. Endocrinology 128, 3299-3309.
Munari-Silem, Y., Guerrier, A., Fromaget, C., Rabilloud, R., Gros, D. andRousset, B. (1994). Differential control of connexin32 and connexin43expression in thyroid epithelial cells. Evidence for a direct relationshipbetween connexin 32 expression and histiotypic morphogenesis.Endocrinology 135, 724-734.
Musil, L. S., Cunningham, B. A., Edelman, G. M. and Goodenough, D. A.(1990). Differential phosphorylation of the gap junction protein connexin 43in junctional communication-competent and -deficient cell lines. J. Cell Biol.111, 2077-2088.
Musil, L. S. and Goodenough, D. A. (1991). Biochemical analysis of connexin43 intracellular transport, phosphorylation and assembly into gap junctionalplaques. J. Cell Biol. 115, 1357-1374.
Nicholson, B., Dermietzel, R., Teplow, D., Traub, O., Willecke, K. andRevel, J. P. (1987). Two homologous protein components of hepatic gapjunctions. Nature 329, 732-734.
Paul, D. L. (1986).: Molecular cloning of cDNA for rat liver gap junctionprotein. J. Cell Biol. 103, 123-134.
Paul, D. L., Ebihara, L., Takemoto, L. J., Swenson, K. I. and Goodenough,D. A. (1991). Connexin46, a novel lens gap junction protein, inducesvoltage-gated currents in nonjunctional plasma membrane of Xenopusoocytes. J. Cell Biol. 115, 1077-1089.
Revel, J. P. and Karnovsky, M. J. (1967). Hexagonal array of subunits inintercellular junctions of the mouse heart and liver. J. Cell Biol. 33, C7-C12.
Saez, J. C., Berthoud, V. M., Moreno, A. P. and Spray, D. C. (1993). Gapjunctions. Multiplicity of controls in differentiated and undifferentiated cellsand possible functional implications. In Advances in Second Messengers andPhosphoprotein Research, vol. 27 (ed. S. Shenolikar and A. C. Nairn), pp.163-198. Raven Press Ltd, New York.
Spray, D. C., Moreno, A. P., Kessler, J. A. and Dermietzel, R. (1991).Characterization of gap junctions between cultured leptomeningeal cells.Brain Res. 568, 1-14.
Steinberg, T. H., Civitelli, R., Geist, S. T., Robertson, A. J., Hick, E.,Veenstra, R. D., Wang, H. Z., Warlow, P. M., Westphale, E. M., Laing, J.G. and Beyer, E. C. (1994). Connexin43 and Connexin45 form gapjunctions with different molecular permeabilities in osteoblastic cells.EMBO J. 13, 744-750.
Stevenson, B. R., Siliciano, J. D., Mooseker, M. S. and Goodenough, D. A.(1986). Identification of ZO-1: a high molecular weight polypeptideassociated with the tight junction (zonula occludens) in a variety of epithelia.J. Cell Biol. 103, 755-766.
Thiele, J. and Reale, E. (1976). Freeze-fracture study of the junctionalcomplexes of human and rabbit thyroid follicles. Cell Tiss. Res. 168, 133-140.
Traub, O., Look, J., Dermietzel, R., Brümmer, F., Hüsler, D. and Willecke,K. (1989). Comparative characterization of the 21-kD and 26-kD gapjunction proteins in murine liver and cultured hepatocytes. J. Cell Biol. 108,1039-1051.
Willecke, K., Hennemann, H., Dahl, E., Jungbluth, S. and Heinkes, R.(1991). The diversity of connexin genes encoding gap junctional proteins.Eur. J. Cell Biol. 56, 1-7.
White, T. W., Bruzzone, R., Goodenough, D. A. and Paul, D. L. (1992).Mouse Cx50, a functional member of the connexin family of gap junctionproteins, is the lens fiber protein MP70. Mol. Biol. Cell 3, 711-720.
Yeager, M. and Gilula, N. B. (1992). Membrane topology and quaternarystructure of cardiac gap junction ion channels. J. Mol. Biol. 223, 929-948.
Zhang, J. T. and Nicholson, B. J. (1989). Sequence and tissue distribution of asecond protein of hepatic gap junctions, Cx26, as deduced from its cDNA. J.Cell Biol. 109, 3391-3401.
Zimmer, D. B., Green, C. R., Evans, W. H. and Gilula, N. B. (1987).Topological analysis of the major protein in isolated intact rat liver gapjunctions and gap junction-derived single membrane structures. J. Biol.Chem. 262, 7751-7763.
(Received 2 December 1994 - Accepted 29 March 1995)