furin is a direct transcription target of notch1 notch1 auto-activation
TRANSCRIPT

Furin is a direct transcription target of Notch1
1
Notch1 auto-activation via transcriptional regulation of Furin, which sustains Notch1 signaling by 1
processing Notch1-activating proteases ADAM10 and MT1-MMP* 2
3
Hong Qiu, Xiaoying Tang, Jun Ma, Khvaramze Shaverdashvili, Keman Zhang, Barbara Bedogni 4
5
Department of Biochemistry, Case Western Reserve University School of Medicine, Cleveland, OH 44106 6
7
*Running title: furin is a direct transcription target of Notch1 8
9
To whom correspondence should be addressed: Barbara Bedogni, Dept. Of Biochemistry, Case Western 10
Reserve University School of Medicine, 10900 Euclid Ave, Cleveland, OH 44106, USA; Tel: 216-368-7602; 11
Fax: 216-368-3419; email: [email protected] 12
13
Keywords: Notch1, MT1-MMP, Furin, ADAM10 14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
MCB Accepted Manuscript Posted Online 17 August 2015Mol. Cell. Biol. doi:10.1128/MCB.00116-15Copyright © 2015, American Society for Microbiology. All Rights Reserved.
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
2
ABSTRACT 31
Notch1 is an evolutionarily conserved transmembrane receptor involved in melanoma growth. Notch1 is 32
first cleaved by Furin in the Golgi to produce the biologically active heterodimer. Following ligand binding, 33
Notch1 is cleaved at the cell membrane by proteases such as ADAM10 and 17 and MT1-MMP, the latter of 34
which we recently identified as a novel protease involved in Notch1 processing. The final cleavage is 35
γ-secretase-dependent and releases the active intracellular domain (NIC). We now demonstrate that Notch1 36
directly regulates Furin expression. Aside from activating Notch1, Furin cleaves and activates several 37
proteases including MT1-MMP, ADAM10 and 17. By chromatin immunoprecipitation and reporter assay, 38
we demonstrate that Notch1 binds at position -1236 of the Furin promoter and drives Furin expression. The 39
Notch1-dependent enhancement in Furin increases the activity of MT1-MMP and ADAM10 but not 40
ADAM17, as demonstrated by shRNA knockdown of Furin. And promotes the cleavage of Notch1 itself. 41
These data highlight a novel positive feedback loop whereby Notch1-dependent Furin expression can induce 42
Notch1 signaling by increasing Notch1 processing and by potentiating the activity of the proteases 43
responsible of its activation. This leads to Notch1 signal amplification that can promote melanoma tumor 44
growth and progression as demonstrated by the inhibition of cell migration and invasion upon furin 45
inhibition downstream of Notch1. Disruption of such feedback signaling might represent an avenue to treat 46
melanoma. 47
48
INTRODUCTION 49
Melanoma is the deadliest form of skin cancer, causing approximately fifty thousand deaths a year, 50
despite new promising therapeutic treatments (1-3). It is imperative to understand melanoma biology in 51
order to find novel therapeutic targets. 52
Notch proteins are transmembrane receptors of approximately 300 kD. In humans, there are four Notch 53
receptors, Notch1-4, which share the same basic structure: 1) an extracellular domain containing 54
EGF-repeats (epidermal growth factor-like repeat domain), an LNR domain (LIN12-Notch-repeat domain) 55
and a HD domain (heterodimerization domain); and 2) an intracellular domain containing a RAM domain 56
(RBP-JK-associated module, an ANK domain (ankyrin repeat), a TAD domain (transactivation) and a PEST 57
domain (proline-glutamate-serine-threonine) (4). The full length Notch is first cleaved by Furin in the trans 58
Golgi. Such cleavage is necessary to properly position Notch1 at the plasma membrane (5, 6). Once at the 59
membrane and following ligand binding, a second cleavage occurs that is canonically driven by ADAM 60
proteases (ADAM10 and 17) (7-10). This produces an unstable fragment that is immediately cleaved by 61
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
3
γ-secretase to release the intracellular domain (NIC) (11). NIC translocates into the nucleus, and binds with 62
transcription factors CSL (C-promoter binding factor-1) and MAML (Mastermind-like) to regulate the 63
transcription of multiple downstream targets, including HES family members, c-Myc, and Cyclin D3 64
(12-20). 65
The Notch pathway is an evolutionarily conserved signaling cascade that plays essential roles in 66
embryogenesis and in cell renewal in the adult by participating in the maintenance of stem cell pluripotency 67
in a variety of tissues (21-23). Previous studies have shown that Notch1 plays essential roles in melanocyte 68
stem and precursor cell homeostasis (24-27). Furthermore, Notch1 is highly expressed in melanomas. 69
Overexpression of active Notch1 (NIC1) transforms primary human melanocytes in vitro and confers 70
metastatic properties to primary melanoma cells (28-31). Notch1 was also shown to be required for 71
melanoma cell growth and survival (31). However, no Notch1 activating mutations have been observed in 72
melanoma so far, suggesting other mechanisms are responsible for the high activity of Notch1 in melanoma 73
cells. 74
One potential mechanism is an increase in processing. We recently demonstrated that MT1-MMP, a 75
membrane-tethered zinc dependent matrix metalloproteinase, acts as a protease involved in the second 76
cleavage of Notch1 (32). MT1-MMP is very abundant in melanomas and associates with disease progression 77
and poor patient outcome (32, 33). MT1-MMP is synthesized as a pre-pro-enzyme of 64kDa that is also 78
cleaved by Furin prior to going to the plasma membrane as an active 55kDa enzyme (34). MT1-MMP is one 79
of the most important MMPs that promote cell migration and invasion in cancer. In melanoma, we 80
demonstrated that MT1-MMP mediates melanoma growth and metastasis via various mechanisms including 81
activation of Notch1 (32). 82
ADAM10 and ADAM17 belong to the ADAM (A disintegrin and metalloproteinase) family of zinc 83
dependent metalloproteinases and are considered the canonical proteases involved in the cleavage of Notch1 84
at the S2 site (7). Structurally similar to MT1-MMP, they also are activated in the Golgi by Furin, to then 85
arrive at the membrane as active enzymes (34). The dysregulated expression of ADAM proteins has been 86
reported in multiple tumors. They are implicated as positive regulators in tumor cell proliferation, 87
angiogenesis and metastasis (35). Importantly, both ADAM10 and ADAM17 have been shown to be 88
overexpressed in melanoma (36-39). 89
Furin cleaves many substrates, including MT1-MMP, Notch1, ADAM10, and ADAM17 (6, 34, 40-42). 90
Furin is a calcium-dependent serine protease that belongs to the proprotein convertases (PCs) family. The 91
expression levels of Furin vary from different cell types and cell differentiation degree (43-47). Furin has 92
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
4
been reported to regulate tumor growth and malignant tumor phenotypes (48-50). Importantly, Furin 93
expression has been shown to coordinate with that of its substrates, such as TGF, BMP and insulin like 94
growth factor (51-53). 95
Our present data establish that Furin is regulated by its own substrate Notch1. In turn, Notch1 induced 96
Furin further activates MT1-MMP and ADAM10, which can then enhance the cleavage of Notch1. Hence, 97
we have identified a novel positive feedback loop whereby Notch1 signaling self-activates by regulating not 98
only its own processing but that of its regulatory proteases. This novel feedback signaling might account for 99
the high activity of Notch1 in melanoma cells. 100
101
MATERIALS AND METHODS 102
Cell lines - Primary and metastatic melanoma cells were in part purchased from ATCC (American Type 103
Culture Collection, Manassas, VA) or were gifts from Dr. Marianne Broome Powell (Stanford University) 104
(54). The use of the cells was approved by the Case Cancer Institutional Review Board (IRB). The cell lines 105
used in this study are: V2387, WM115, WM266-4 and SKMEL2. These are human melanoma cells derived 106
from primary melanoma (WM115), metastases to the skin (WM266-4 and SKMEL2) and lymph nodes 107
(V2387). All cells were maintained in DMEM (Dulbecco’s modified Eagle’s medium) supplemented with 108
10% FCS (fetal calf serum), 1% glutamine, and 1% penicillin-streptomycin. 109
shRNAs and Expression Plasmids - shRNAs against human Notch1 (TRCN0000003359) and Furin 110
(TRCN000075238 and TRCN000075239) were purchased from Sigma . The Notch1-NIC lentiviral 111
expression vector was constructed by inserting the cDNA sequence corresponding to human Notch1-NIC 112
(base pairs 5278-7668 of full length human Notch1) into the pLM-CMV-Ha-puro-PL3 lentiviral plasmid 113
(55), between XbaI-XhoI. The lentiviral iDuet101a-DN-MAML plasmid (dominant negative MAML) was 114
kindly provided by Dr. Eli Bar (Case Western Reserve University). Viral particles were produced in 293-FT 115
cells using the X-tremegene 9 reagent (Roche, Mannheim, Germany) according to manufacturer's 116
instructions. The packaging plasmids used were pMD2.G and psPAX2 that were purchased from Addgene 117
(Cambridge, MA). 118
Western Blot Analysis - Total protein for all assays was extracted with urea lysis buffer (9 M urea; 75 mM 119
Tris-HCl, pH 7.5, and 100 mM 2-mercaptoethanol), and 20μg/sample was separated by 8–10% SDS-PAGE 120
and transferred onto PVDF membranes. Membranes were probed with the following antibodies: anti-Notch1 121
(C20, Santa Cruz Biotechnology, Santa Cruz, CA); anti Notch1mN1A (full length and cleaved – Novus 122
Biologicals LLC, Littleton , CO); anti-Notch1NIC (cleaved Val-1744 - Cell Signaling Technologies, 123
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
5
Beverly, MA); anti-MT1-MMP (clone LEM-2/15.8, Millipore, Billerica, MA); anti-ADAM10 (Abcam, 124
Cambridge, MA); anti-TACE (tumor necrosis factor- α- converting enzyme, ADAM17) (eBioscience, San 125
Diego, CA) and anti-Furin (Abcam, Cambridge, MA). Bands were detected using Super Signal detection 126
reagent (Thermo Scientific). Loading was normalized with anti-β-actin (Santa Cruz Biotechnology). 127
Densitometric quantifications (by ImageJ) of band intensity were normalized to band intensity of the 128
respective β-actin. 129
Real-time PCR Analysis - cDNA was synthesized from total RNA using SuperScript first-strand synthesis 130
system for RT-PCR (Invitrogen), then used for PCR amplification with SYBR Green PCR master mix 131
(Roche). The following primer sets were used to amplify specific target genes: human ADAM10 forward: 132
5’-CAAAGTCTGAGAAGTGTCGGG-3’; reverse: 5’-CTGCACATTGCCCATTAATG-3’; human 133
ADAM17 forward: 5’-ACCTGAAGAGCTTGTTCATCGAG-3’; reverse: 134
5’-CCATGAAGTGTTCCGATAGATGTC-3’; human GAPDH forward: 135
5’-CGCTCTCTGCTCCTCCTGTT-3’; reverse: 5’-CCATGGTGTCTGAGCGATGT-3’; human MT1-MMP 136
forward: 5’-CTCCCTCGGCTCGGCCCAAA-3’; reverse: 5’-CGCCTCATGGCCTTCATGGTGTCT-3’; 137
human Furin forward: 5’-GAAGCAGCAGCGGCCAGGAT-3’; reverse: 138
5’-CGAAGATCTGGCCCAGGTTGAGG-3’; human PC5 forward: 139
5’-TGCCAGGGACCAACCCAGGA-3’; reverse: 5’-TGCTCTCGGCCATAGTTGTCTGC-3’; human PC7 140
forward: 5’-CACCAGCACGGTTTCGGCCT-3’; reverse: 5’-AGCCGTTGGGATCCGAGTCCAT-3’. 141
Relative quantification of mRNA expression levels was normalized by GAPDH. 142
Chromatin immunoprecipitation assay - Chromatin immunoprecipitation (ChIP) was performed using ChIP 143
assay kits (Upstate Biotechnology) following the manufacturer's recommendations. The following primer 144
sequences were used: Furin promoter at B1 (-3556bp): forward: 145
5’-ATCAGGAGGGTCACCTATGGTGC-3’; reverse: 5’-CAGAGGTGCTGGGATTACAGGC-3’; Furin 146
promoter at B2 (-1236bp): forward: 5’-CTCAGAGCTTAGTTCCCAGCAGAC-3’; reverse: 147
5’-GATCTCCAGGTATTGCCAAATGTC-3’; Furin promoter at B3 (+960bp): forward: 5’- 148
ATGCAGATTGAAGAGGCAAGCTG-3’; reverse: 5’- GCAATACTTGCTGCTTCAGGTGC-3’; HES1 149
promoter: forward: 5’-CGTGTCTCCTCCTCCCATTG-3’; reverse: 150
5’-CCAGGACCAAGGAGAGAGGT-3’. The antibody employed for the immunoprecipitation was 151
previously described (56). 152
Luciferase Assays - The Hes1-reporter plasmid was a kind gift from Dr. Ryoichiro Kageyama (Kyoto 153
University, Kyoto, Japan) (57). The human Furin promoter-luciferase constructs pGL2-Basic, pGL2-SacI, 154
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
6
was generously provided by Dr. Claire M. Dubois (University of Sherbrooke, Canada) (43). The mutations 155
within the pGL2-SacI reporter were generated by PCR using the following primers: 156
5’-cctgtgacgtcacagctcctcattctgcgacagtgg-3’ and 5’-gaatgaggagctgtgacgtcacaggatggtggttag-3’ which replaces 157
the sequence 5’-TTCCCAC-3’ with 5’-TCACAGC-3’. V2387, WM115 or SKMEL2 cells (5 ×104/well, in 158
24-well plates) were transfected using X-tremegene HP reagent (Roche Applied Science) as per the 159
manufacturer’s instructions. After 48 h, cells were lysed in 100 μl of lysis buffer (Promega, Madison, WI). 160
A Renilla luciferase reporter plasmid driven by a CMV promoter was co-transfected with the reporter 161
constructs at a 1:20 ratio to assess transfection efficiency. Activities of firefly and Renilla were assessed by 162
the Dual-Luciferase assay system (Promega), and light production was measured for 10 s in a Monolight 163
2010 luminometer (Molecular Devices, Sunnyvale, CA). 164
MT1-MMP and ADAM10 activity assay - Membrane proteins were extracted from 106 cells by three cycles 165
of freeze-thaw in dry ice/ethanol/37°C baths. The lysates were sonicated for 3 s, and the membranes were 166
pelleted by centrifugation (30 min, 13 000 g, 4°C) and resuspended in PBS. Equal protein amounts per 167
sample were incubated with a fluorogenic MT1-MMP substrate (Mca-PLGL-Dap (Dnp)-AR-NH2) or 168
5-FAM /QXLTM 520 ADAM10 substrate provided by the manufacturer (Anaspec, Fremont, CA, USA). 169
Fluorescence intensity was measured at Ex/Em = 490 + 20 nm/520 + 20 nm using a SpectraMax M2 Elisa 170
reader (Molecular Devices, Sunnyvale, CA, USA). 171
Notch ligand stimulation assay - Notch signaling was induced by plating cells (32,000/cm2) on dishes 172
displaying immobilized FC- or FC-JAGGED1 ligand anchored to protein-A. Plasmids expressing FC- and 173
FC-JAGGED1 (58, 59) were kindly provided by Dr. Aaron Proweller (Case Western Reserve University, 174
Cleveland). 175
Migration and invasion assays – SKMel2 cells were plated in a confluent monolayer in duplicate. A scratch 176
using a pipette tip was produced and the detached cells were gently washed away with PBS. Plates were 177
incubated under a time-lapse microscope for 24 hours. Pictures were taken every hour for the duration of the 178
experiment. Frames were aligned and distance between one of the edges of the wound in the first frame 179
(considered as time 0) to the migration front was calculated for the time points indicated. For the trans-well 180
invasion assay, 25 x104 cells suspension was added to an 8μ pore insert either uncoated (control) or coated 181
with growth factor reduced matrigel (BD Biosciences, MA). 5% FBS containing DMEM media was added 182
to the lower chamber (24 well plate) as chemo-attractant. After a 24 hours incubation cells that migrated 183
through the control inserts or matrigel and collected onto the bottom membrane, were fixed with 4% 184
Formaldehyde and stained with Coomassie blue. Each treatment was done in triplicate. Membranes were 185
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
7
incubated in 10% acetic acid to extract the Coomassie stain and color intensity was quantified at 295nm. % 186
invasion is calculated as follows: (mean reads of matrigel/mean reads of insert)x100. 187
188
RESULTS 189
Notch1 modulates MT1-MMP activity - Our previous data showed that MT1-MMP is involved in the 190
cleavage of Notch1, and that it is a critical modulator of melanoma metastasis and melanoma cell growth (32, 191
33, 60). Thus, we decided to explore the mechanisms of MT1-MMP regulation. Interestingly, we observed 192
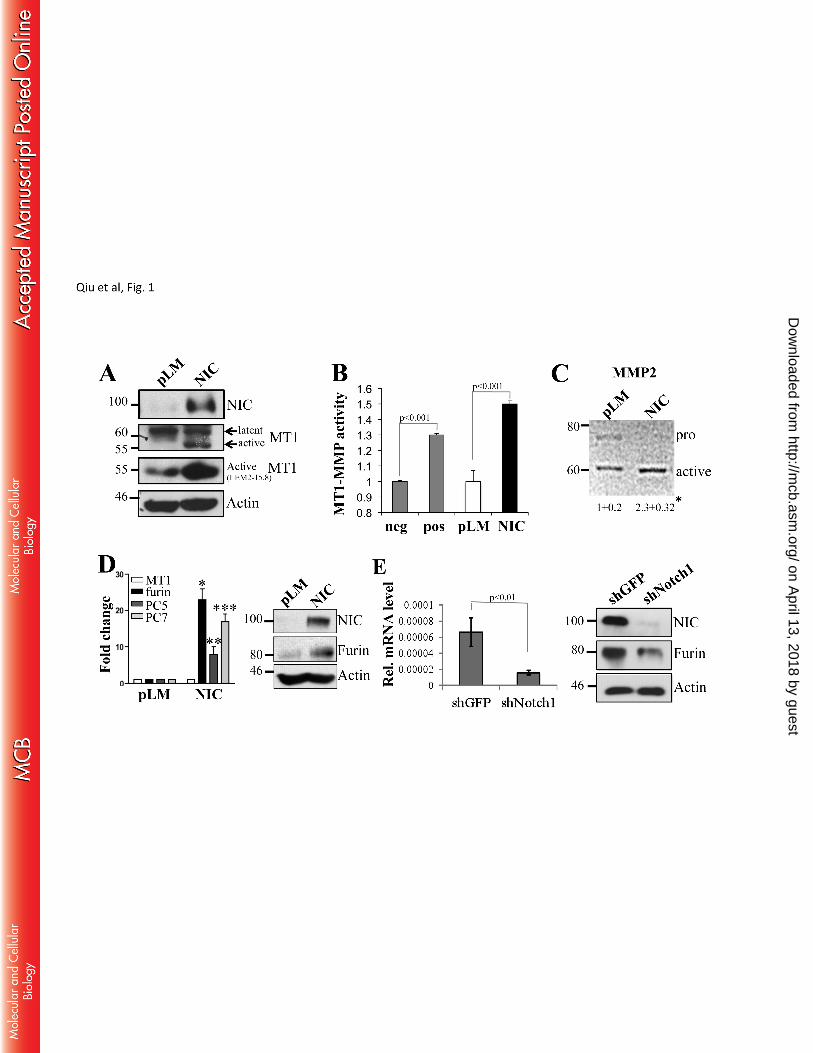
that Notch1 promotes MT1-MMP activity. Expression of an active Notch1 (NIC) in melanoma cells 193
increases the 55 kDa fraction of MT1-MMP (Fig 1A), the catalytically active enzyme, as assessed by two 194
different antibodies, one of which recognizes the active MT1-MMP peptide specifically. This protein 195
fraction is indeed the active protein as it leads to an increase in MT1-MMP activity as measured by an in 196
vitro activity assay (Fig 1B) (61, 62). Furthermore, expression of active Notch1 leads to an increase in 197
MMP2 processing (Fig. 1C), a known target of MT1-MMP (63). Given that MT1-MMP activation requires 198
the cleavage of the inhibitory propeptide by Furin or Furin-like convertases, and given that expression of 199
Notch1 did not change the levels of MT1-MMP transcript in the cells (Fig. 1D), we investigated if Notch1 200
could regulate the expression of the convertases. Indeed, we found that Notch1-NIC increased both mRNA 201
and protein levels of Furin (Fig 1D, E). The transcripts of the Furin-like convertases PC5 and PC7, also 202
involved in MT1-MMP activation, were increased as well (Fig 1D). Conversely, inhibition of Notch1 by a 203
specific shRNA, resulted in a decrease of Furin mRNA and protein. In a parallel experiment, cleavage of 204
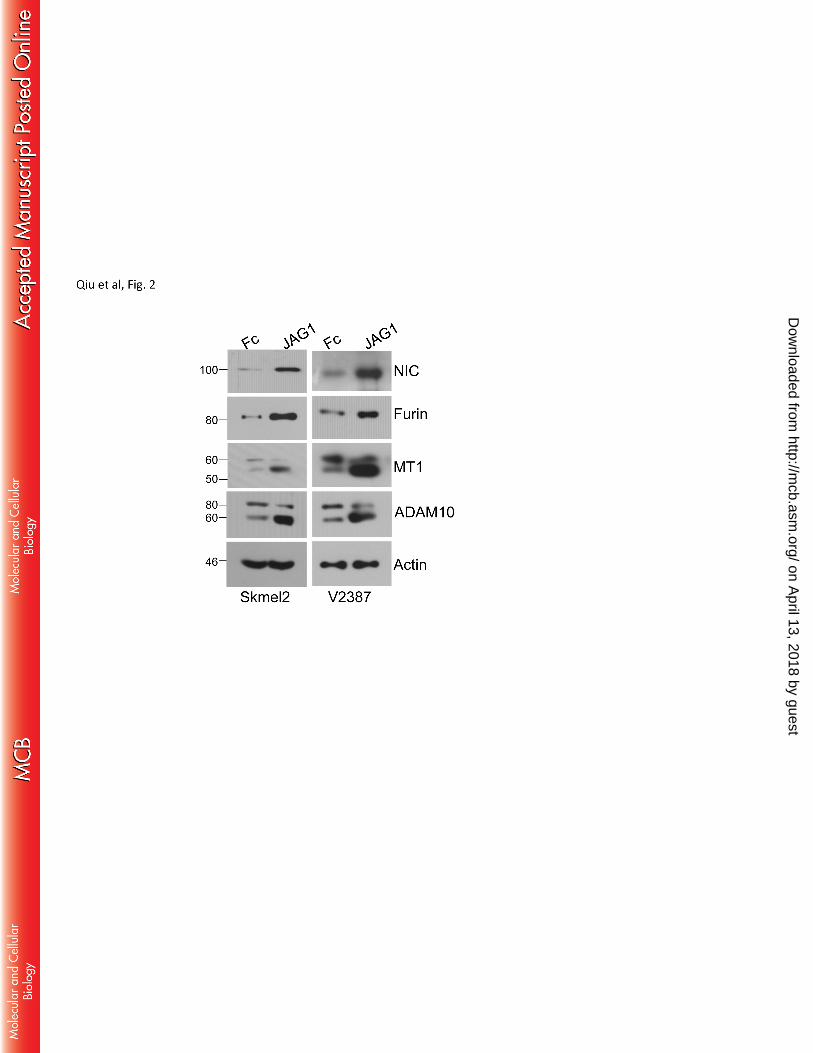
endogenous Notch1 by stimulation with recombinant JAGGED1 ligand, led to an increase in Furin and a 205
correspondent increase in MT1-MMP processing (Fig. 2), further suggesting Notch1 activates MT1-MMP 206
likely via furin expression. 207
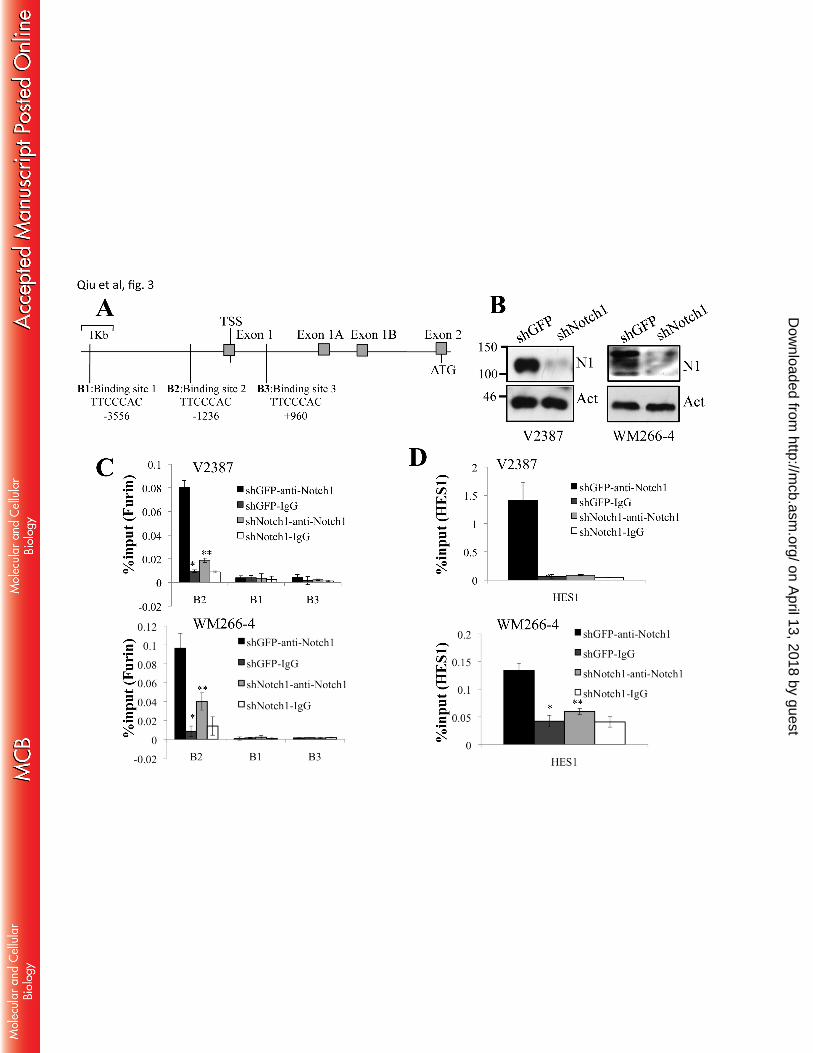
Notch1 affects Furin expression through binding to the Furin promoter - Based on the previous data, we 208
hypothesized that Notch1 affects the activity of its protease MT1-MMP by increasing the levels of Furin 209
available to cleave the inhibitory propeptide. We therefore sought to determine whether Notch1 might 210
regulate Furin by directly driving its expression. Active Notch1, although not capable of directly binding to 211
DNA sequences, functions in a transcription complex together with CBF-1 and Master Mind-Like (MAML). 212
Sequence analysis of the Furin promoter through Motif Search (http://www.genome.jp/tools/motif/) revealed 213
three putative CBF1/Notch-binding sequences (TTCCCAC) within a 5 kb sequence encompassing 4 kb 214
upstream and 1 kb downstream the Furin transcription start site: one at position -3556bp, one at -1236bp, 215
and one at +960bp (Fig. 3A). The sites were then tested for Notch1 occupancy in V2387 and WM266-4 cells 216
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
8
expressing either a control shRNA (shGFP) or shNotch1 (Fig. 3B), by chromatin immunoprecipitation 217
(ChIP) assay. We observed enrichment at the B2 site (position -1236) in shGFP cells that was inhibited in 218
shNotch1 expressing cells, but not at positions -3556 and +960 (Fig 3C). Binding of Notch1 to the HES1 219
promoter was performed as a positive control, which also showed enrichment in shGFP cells compared to 220
shNotch1 cells (Fig 3D). 221
To further confirm that the B2 site is indeed where Notch1 binds and drives transcription of Furin, we 222
performed a series of luciferase reporter assays (fig. 4A). In V2387 cells, the reporter SacI that contains the 223
WT B2 binding site, showed a 10 fold induction of luciferase activity in cells expressing the active form of 224
Notch1 (NIC) compared to control (pLM) expressing cells (Fig 4B). The HES1 reporter construct was used 225
as a positive control. Mutation of the B2 binding site form TTCCCAC to TCACAGC, significantly reduced 226
the induction of luciferase activity in NIC expressing cells, compared to the wild type (WT) in three 227
different melanoma cell lines (Fig 4C). The upper panel shows the expression levels of Notch1-NIC in each 228
line tested. These data identify Furin as a novel Notch1 direct target. 229
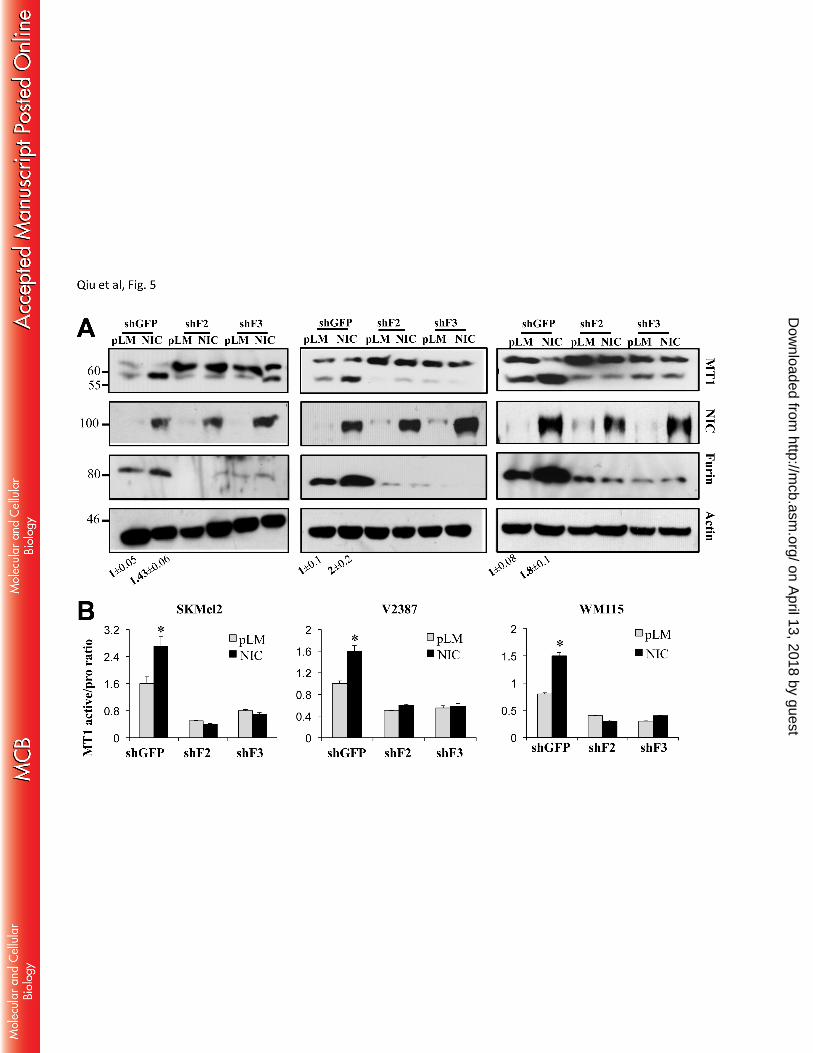
Notch1 affects MT1-MMP activity through Furin - Once we established that Notch1 modulates Furin 230
expression, and given that MT1-MMP activity is dependent on the cleavage of the inhibitory propeptide by 231
Furin, we next evaluated whether Notch1 plays a role in the conversion of MT1-MMP from inactive (64Kd) 232
to active (55Kd) through Furin. We tested whether inhibition of Furin by specific shRNA sequences would 233
impair MT1-MMP activation downstream of Notch1. Since Furin also processes Notch1, we used cell lines 234
expressing an exogenous, active Notch1-NIC that is independent of Furin cleavage. Western blot analysis 235
showed that the ratio of active MT1-MMP increases in NIC expressing cells, but decreases in Furin knock 236
down cells (Fig 5A, 4B). Importantly, inhibition of Furin expression abolishes the ability of active Notch1 to 237
increase the active MT1-MMP fraction (Fig 5A, 4B), indicating that Notch1 affects MT1-MMP activity 238
through Furin. 239
Notch1 affects ADAM10 activity - ADAM10 and ADAM17 are generally accepted as proteases responsible 240
for Notch1 cleavage following ligand binding. Similar to MT1-MMP, ADAM 10 and 17 are also activated 241
by Furin prior to reaching the plasma membrane as fully active enzymes (7, 34, 40). Previous data from our 242
group showed that melanoma tumors and cells do express these proteases, although not at the same levels as 243
MT1-MMP (32). We therefore wanted to determine whether ADAM10 and 17 were also affected by Notch1 244
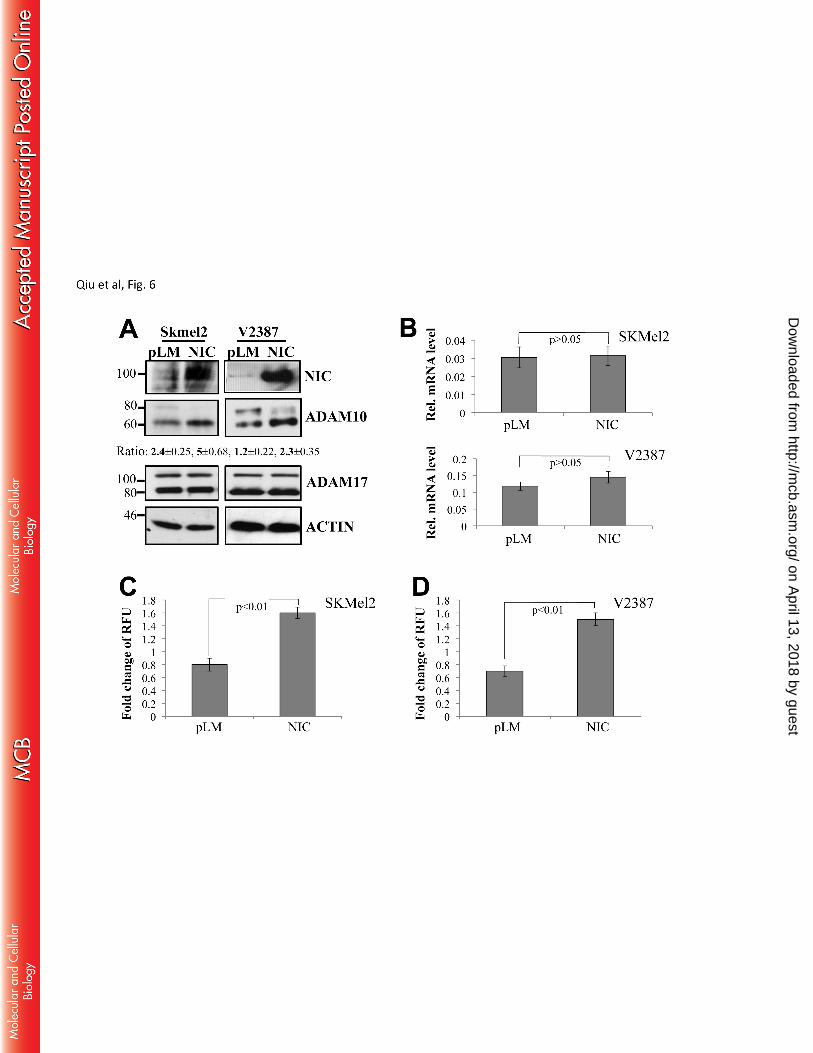
through Furin induction. As shown in Fig 6A, the active band of ADAM10 (60kDa, the bottom band) 245
increases in the NIC expressing cells, indicating an increase in processing. Similarly, in cells stimulated by 246
recombinant JAGGED1, ADAM10 processing is increased (Fig. 2). The same was not true for ADAM17, 247
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
9
however. We further measured the activity of ADAM10 in vitro using a commercially available activity 248
assay (ANASPEC), and found that overexpression of NIC significantly increased ADAM10 activity in both 249
SKMel2 and V2387 cells (Fig 6C, D). Of note, Notch1 did not affect ADAM10 gene expression (Fig 6B). 250
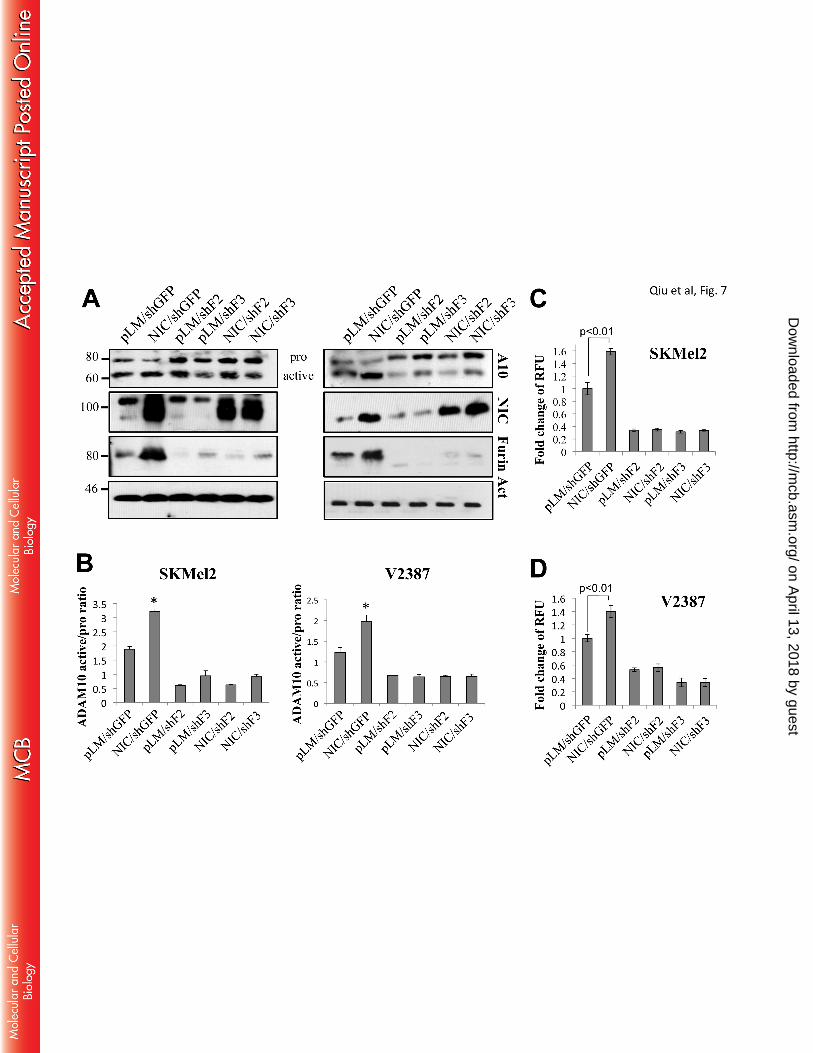
We next evaluated whether Notch1 played a role in the conversion of ADAM10 through Furin. As Fig 7 251
shows, the active band of ADAM10 increases in the NIC expressing cells, but decreases in the Furin knock 252
down cells. Again, inhibition of Furin abolished the ability of Notch1-NIC to process ADAM10, as 253
previously seen for MT1-MMP. The in vitro activity assay mirrored the western blot data, showing 254
induction of activation in the presence of NIC and lack of increase by Notch1 when Furin is inhibited (fig. 255
7C, D). 256
Inhibition of furin diminishes Notch1 processing – given that Furin is promotes the first cleavage of 257
Notch1 and such cleavage is required for proper Notch positioning at the plasma memnbrane and access to 258
the ligand (5, 6), we first wanted to determine whether interruption of Notch1 signaling at the DNA binding 259
levels would affect furin and consequently Notch1-NIC release. To do so, cells were transduced with the 260
dominant negative MAML. Expression of MAML-DN inhibited the expression of furin and HEY1, a known 261
direct Notch transcription target, and resulted in diminished processing of full length to cleaved Notch1 as 262
shown by an antibody that recognized both full length and processed Notch1 (Fig. 8A upper panel and graph) 263
and by one that specifically recognizes Notch1 cleaved by γ-secretase at valin 1744 (Fig 8A, NIC specific 264
panel). Inhibition of furin by a specific shRNA reduced Notch processing in a similar manner (Fig. 8B). 265
These data indicate Notch1 can promote its own activation through a signaling loop involving furin and the 266
proteases ADAM10 and MT1-MMP. 267
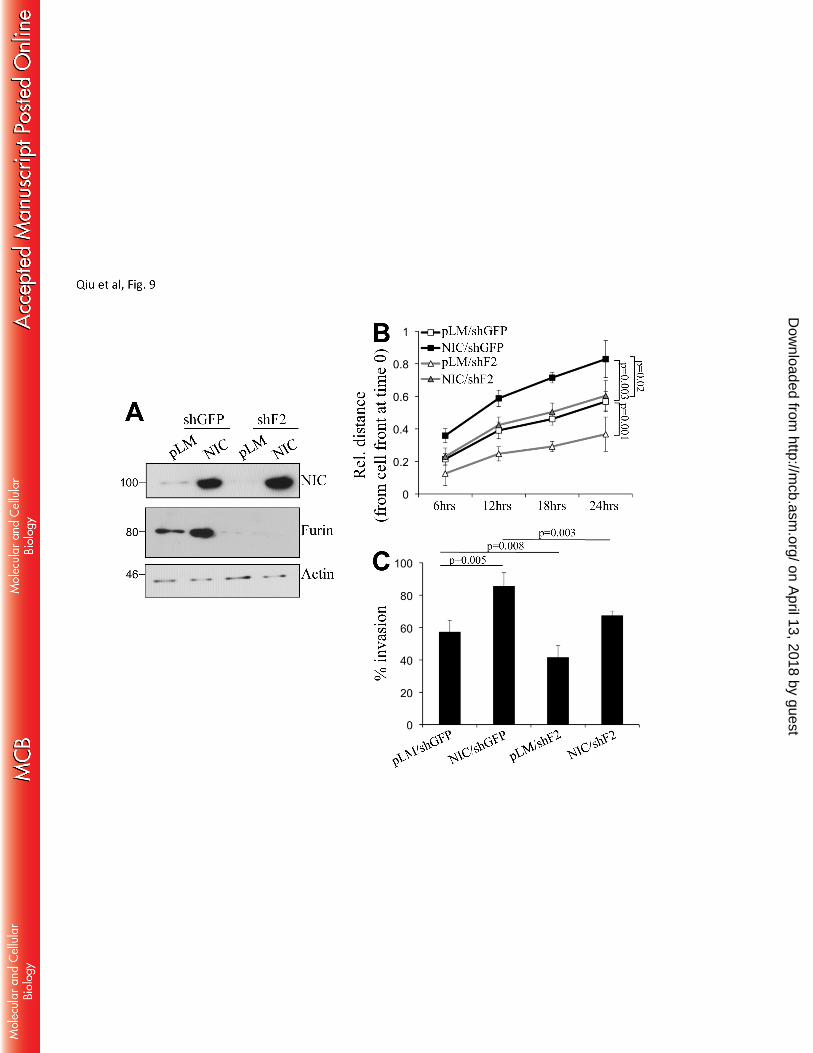
Inhibition of furin affects melanoma cell migration and invasion – We have previously shown that 268
MT1-MMP is required for melanoma cell migration and invasion (64). Given that Notch1 dependent furin 269
expression promotes MT1-MMP activation we next examined whether furin inhibition could affect 270
migration and invasion of cells downstream of Notch1. Furin was downregulated in SKMel2 cells 271
expressing constitutive Notch1-NIC (fig. 9A). Then a scratch assay and a matrigel invasion assay were 272
performed. Cells expressing Notch1-NIC migrated faster than controls, however, a 30% reduction in 273
migration (on average) was observed when furin was inhibited in both control (pLM) and NIC expressing 274
cells (fig. 9B and suppl. Fig 1). Similarly, cells expressing active Notch1 were more invasive than control 275
cells, but inhibition of furin reduced these invasion promoting effects (fig, 9C). 276
277
278
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
10
DISCUSSION 279
Notch processing is an unexplored and potentially therapeutically targetable mechanism of increased 280
Notch signaling in cancer. We have established that elevated Notch plays a critical role in the pathogenesis 281
of melanoma, and describe here a novel circuit by which Notch drives an autoregulatory positive feedback 282
loop initiating with the elevated expression of Furin. The full length precursor of Notch1 is cleaved by Furin 283
in the trans-Golgi network, prior to arriving to the plasma membrane as a heterodimer where it can be 284
activated by interaction with Delta-like and Jagged ligands expressed on the surface of adjacent cells (6, 9, 285
65, 66). The cleavage by Furin is required for the proper surface presentation of Notch1 and proper ligand 286
binding (65). In fact, studies show that Furin-resistant Notch1 receptors exhibit decreased surface expression 287
and ligand-mediated receptor activation (5). Although Furin can be regulated by a variety of factors 288
including Sox9 (67), TGFβ (68) and BMPs (69), here we find that Furin is a novel direct transcriptional 289
target of its own substrate Notch1. Furthermore, we show that the activity of two major proteases involved 290
in Notch1 cleavage following ligand binding, are induced by active Notch1 through the modulation of Furin. 291
Both MT1-MMP and ADAM10 are synthesized as inactive zymogens whose activation requires cleavage of 292
an inhibitory prodomain sequence (40, 70). Furin is an essential activator of pro-MT1-MMP that controls 293
the level of active MT1-MMP on the cell surface (34). On the other hand, previous studies demonstrated that 294
overexpression of Furin and protein convertase PC7 increased the levels of active ADAM10, and mutation 295
of the convertase dependent cleavage sites blocked the processing of ADAM10 to its active form (40). 296
Indeed, here we demonstrate that Notch1 promotes the processing of MT1-MMP and ADAM10 to their 297
active forms through Furin, as down regulation of Furin expression is sufficient to abolish the ability of 298
Notch1 to promote the processing of the proteases and to increase their activity. Based on these data we 299
suggest that Notch1, by regulating the expression of Furin, establishes a positive feedback loop, which not 300
only can promote Notch1 processing but also its activation by modulating the activity of MT1-MMP and 301
ADAM10. 302
MT1-MMP belongs to a family of membrane-tethered matrix metalloproteinase and it is considered one 303
of the most important MMPs in promoting cancer cell migration and invasion (33). Recently, we 304
demonstrated that MT1-MMP operates as a protease involved in Notch1 activation, and that MT1-MMP 305
dependent Notch1 activation mediates part of the pro-tumorigernic properties of MT1-MMP (32). 306
Melanomas express high levels of active Notch1 and its expression correlated with that of MT1-MMP (32, 307
54, 71, 72). Hence, considering the results presented here, it is likely that in tumors where the two proteins 308
are present, they functionally influence each other by promoting their reciprocal activation. The decrease in 309
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
11
cell migration and invasion downstream of active Notch1 upon furin inhibition would substantiate this 310
notion. 311
On the other hand, although ADAM10 and ADAM17 are both capable of processing Notch1 at the S2 312
site (10, 73), more and more evidence point to ADAM10 as the main enzyme responsible of such activity. 313
Firstly, the defects found in ADAM10 deficient mice phenocopy those of Notch1 deficient mice (74-76); 314
and secondly, Van Tetering et al. (10) demonstrated that only in ADAM10 knock out fibroblasts Notch1 315
cleavage is severely diminished, but not so in ADAM17 knock out cells. In melanoma, expression of 316
ADAM10 has been shown to be significantly increased in melanoma metastases compared to primary 317
tumors (37, 38). Knock down of ADAM10 inhibits melanoma cell growth and migration, while 318
overexpression of ADAM10 increases migration of melanoma cells (38). Our study shows that 319
overexpression of Notch1 increases the activity of ADAM10 through the modulation of Furin. In contrast, 320
Notch1 did not promote the processing of ADAM17. These data suggest that ADAM17, at least in 321
melanoma cells, is not under the influence of Notch1 signaling. 322
Together, the data presented here highlight a novel positive feedback signaling whereby Notch1, by 323
regulating Furin expression, promotes its own cleavage in the Golgi, a process that is required for the proper 324
localization of Notch1 on the plasma membrane; and stimulates the activation of ADAM10 and MT1-MMP, 325
which not only are involved in the cleavage and activation of Notch1, but that contribute to tumorigenesis by 326
modulating the processing of a variety of substrates including extracellular matrix and surface receptors. 327
These findings add further complexity to Notch signaling deregulation in cancer. Notch not only promotes 328
tumorigenesis by directly regulating genes involved in tumor development and progression (77), but can 329
also indirectly do so by influencing the activity of protumorigenic enzymes. We therefore propose that the 330
positive feedback loop between Notch1 and its proteases promotes the amplification of Notch1 signaling and 331
may represent a novel therapeutic target to treat melanoma. 332
333
334
335
336
337
338
339
340
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
12
REFERENCES 341
1. Siegel R, Naishadham D and Jemal A: Cancer Statistics, 2012. Ca-a Cancer Journal for 342 Clinicians 62: 10-29, 2012. 343 2. Weinstock MA: Reducing Death from Melanoma and Standards of Evidence. Journal Of 344 Investigative Dermatology 132: 1311-1312, 2012. 345 3. Chang AE, Karnell LH and Menck HR: The National Cancer Data Base report on cutaneous and 346 noncutaneous melanoma - A summary of 84,836 cases from the past decade. Cancer 83: 1664-1678, 347 1998. 348 4. Bray SJ: Notch signalling: a simple pathway becomes complex. Nature Reviews Molecular Cell 349 Biology 7: 678-689, 2006. 350 5. Gordon WR V-UD, L'Heureux S, Ashworth T, Malecki MJ, Sanchez-Irizarry C, McArthur DG, 351 Histen G, Mitchell JL, Aster JC, Blacklow SC.: Effects of S1 cleavage on the structure, surface 352 export, and signaling activity of human Notch1 and Notch2. PLoS One 4: 1/-12, 2009. 353 6. Logeat F, Bessia C, Brou C, LeBail O, Jarriault S, Seidah NG and Israel A: The Notch1 receptor 354 is cleaved constitutively by a furin-like convertase. Proceedings Of the National Academy Of 355 Sciences Of the United States Of America 95: 8108-8112, 1998. 356 7. Bozkulak EC and Weinmaster G: Selective Use of ADAM10 and ADAM17 in Activation of 357 Notch1 Signaling. Molecular And Cellular Biology 29: 5679-5695, 2009. 358 8. Sulis ML, Saftig P and Ferrando AA: Redundancy and specificity of the metalloprotease system 359 mediating oncogenic NOTCH1 activation in T-ALL. Leukemia 25: 1564-1569, 2011. 360 9. Tiyanont K, Wales TE, Aste-Amezaga M, Aster JC, Engen JR and Blacklow SC: Evidence for 361 Increased Exposure of the Notch1 Metalloprotease Cleavage Site upon Conversion to an Activated 362 Conformation. Structure 19: 546-554, 2011. 363 10. van Tetering G, van Diest P, Verlaan I, van der Wall E, Kopan R and Vooijs M: Metalloprotease 364 ADAM10 Is Required for Notch1 Site 2 Cleavage. Journal Of Biological Chemistry 284: 365 31018-31027, 2009. 366 11. Fortini ME: gamma-secretase-mediated proteolysis in cell-surface-receptor signalling. Nature 367 Reviews Molecular Cell Biology 3: 673-684, 2002. 368 12. Artavanistsakonas S, Matsuno K and Fortini ME: Notch Signaling. Science 268: 225-232, 1995. 369 13. Egan SE, St-Pierre B and Leow CC: Notch receptors, partners and regulators: From conserved 370 domains to powerful functions. Protein Modules In Signal Transduction 228: 273-324, 1998. 371 14. Hsieh JJD, Henkel T, Salmon P, Robey E, Peterson MG and Hayward SD: Truncated 372 mammalian Notch1 activates CBF1/RBPJk-repressed genes by a mechanism resembling that of 373 Epstein-Barr virus EBNA2. Molecular And Cellular Biology 16: 952-959, 1996. 374 15. Jarriault S, Brou C, Logeat F, Schroeter EH, Kopan R and Israel A: Signaling Downstream Of 375 Activated Mammalian Notch. Nature 377: 355-358, 1995. 376 16. Kopan R and Goate A: A common enzyme connects Notch signaling and Alzheimer's disease. 377 Genes & Development 14: 2799-2806, 2000. 378 17. Oswald F, Liptay S, Adler G and Schmid RM: NF-kappa B2 is a putative target gene of 379 activated notch-1 via RBP-J kappa. Molecular And Cellular Biology 18: 2077-2088, 1998. 380 18. Weng AP, Millholland JM, Yashiro-Ohtani Y, Arcangeli ML, Lau A, Wai C, Bianco C, 381 Rodriguez CG, Sai H, Tobias J, Li Y, Wolfe MS, Shachaf C, Felsher D, Blacklow SC, Pear WS and 382 Aster JC: c-Myc is an important direct target of Notch1 in T-cell acute lymphoblastic 383 leukemia/lymphoma. Genes & Development 20: 2096-2109, 2006. 384
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
13
19. Wu L, Sun T, Kobayashi K, Gao P and Griffin JD: Identification of a family of mastermind-like 385 transcriptional coactivators for mammalian notch receptors. Molecular And Cellular Biology 22: 386 7688-7700, 2002. 387 20. Zhou SF, Fujimuro M, Hsieh JJD, Chen L, Miyamoto A, Weinmaster G and Hayward SD: SKIP, 388 a CBF1-associated protein, interacts with the ankyrin repeat domain of NotchIC to facilitate NotchIC 389 function. Molecular And Cellular Biology 20: 2400-2410, 2000. 390 21. Zhou K, Huang L, Zhou Z, Hu C, Liu W, Zhou J and Sun H: Wnt and Notch signaling pathways 391 selectively regulating hematopoiesis. Ann Hematol 89: 749-57, 2010. 392 22. Brabletz S, Schmalhofer O and Brabletz T: Gastrointestinal stem cells in development and 393 cancer. J Pathol 217: 307-17, 2009. 394 23. Imayoshi I, Sakamoto M, Yamaguchi M, Mori K and Kageyama R: Essential roles of Notch 395 signaling in maintenance of neural stem cells in developing and adult brains. J Neurosci 30: 3489-98, 396 2010. 397 24. Haass NK and Herlyn M: Normal human melanocyte homeostasis as a paradigm for 398 understanding melanoma. Journal Of Investigative Dermatology Symposium Proceedings 10: 399 153-163, 2005. 400 25. Kumano K, Masuda S, Sata M, Saito T, Lee SY, Sakata-Yanagimoto M, Tomita T, Iwatsubo T, 401 Natsugari H, Kurokawa M, Ogawa S and Chiba S: Both Notch1 and Notch2 contribute to the 402 regulation of melanocyte homeostasis. Pigment Cell & Melanoma Research 21: 70-78, 2008. 403 26. Schouwey K and Beermann F: The Notch pathway: hair graying and pigment cell homeostasis. 404 Histology And Histopathology 23: 609-619, 2008. 405 27. Osawa M and Fisher DE: Notch and Melanocytes: Diverse Outcomes from a Single Signal. 406 Journal Of Investigative Dermatology 128: 2571-2574, 2008. 407 28. Balint K, Xiao M, Pinnix CC, Soma A, Veres I, Juhasz I, Brown EJ, Capobianco AJ, Herlyn M 408 and Liu ZJ: Activation of Notch1 signaling is required for beta-catenin-mediated human primary 409 melanoma progression. Journal Of Clinical Investigation 115: 3166-3176, 2005. 410 29. Liu ZJ, Xiao M, Balint K, Smalley KSM, Brafford P, Qiu RH, Pinnix CC, Li XL and Herlyn M: 411 Notch1 signaling promotes primary melanoma progression by activating mitogen-activated protein 412 kinase/phosphatidylinositol 3-kinase-Akt pathways and up-regulating N-cadherin expression. Cancer 413 Research 66: 4182-4190, 2006. 414 30. Pinnix CC, Lee JT, Liu ZJ, McDaid R, Balint K, Beverly LJ, Brafford PA, Xiao M, Himes B, 415 Zabierowski SE, Yashiro-Ohtani Y, Nathanson KL, Bengston A, Pollock PM, Weeraratna AT, 416 Nickoloff BJ, Pear WS, Capobianco AJ and Herlyn M: Active Notch1 Confers a Transformed 417 Phenotype to Primary Human Melanocytes. Cancer Research 69: 5312-5320, 2009. 418 31. Zhang K, Wong P, Zhang L, Jacobs B, Borden EC, Aster JC and Bedogni B: A 419 Notch1-neuregulin1 autocrine signaling loop contributes to melanoma growth. Oncogene 31: 420 4609-4618, 2012. 421 32. Ma J, Tang XY, Wong PK, Jacobs B, Borden EC and Bedogni B: Noncanonical Activation of 422 Notch1 Protein by Membrane Type 1 Matrix Metalloproteinase (MT1-MMP) Controls Melanoma 423 Cell Proliferation. Journal Of Biological Chemistry 289: 8442-8449, 2014. 424 33. Shaverdashvili K, Wong P, Ma J, Zhang KM, Osman I and Bedogni B: MT1-MMP modulates 425 melanoma cell dissemination and metastasis through activation of MMP2 and RAC1. Pigment Cell 426 & Melanoma Research 27: 2014. 427 34. Yana I and Weiss SJ: Regulation of membrane type-1 matrix metalloproteinase activation by 428 proprotein convertases. Molecular Biology Of the Cell 11: 2387-2401, 2000. 429 35. Turner SL, Blair-Zajdel ME and Bunning RAD: ADAMs and ADAMTSs in cancer. British 430 Journal Of Biomedical Science 66: 117-128, 2009. 431
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
14
36. Anderegg U, Eichenberg T, Parthaune T, Haiduk C, Saalbach A, Milkova L, Ludwig A, Grosche 432 J and Simon JC: Functions of ADAM10 in human melanoma cells. Experimental Dermatology 17: 433 275-275, 2008. 434 37. Gangemi R, Amaro A, Gino A, Barisione G, Fabbi M, Pfeffer U, Brizzolara A, Queirolo P, Salvi 435 S, Boccardo S, Gualco M, Spagnolo F, Jager MJ, Mosci C, Rossello A and Ferrini S: ADAM10 436 correlates with uveal melanoma metastasis and promotes in vitro invasion. Pigment Cell & 437 Melanoma Research 27: 1138-1148, 2014. 438 38. Lee SB, Schramme A, Doberstein K, Dummer R, Abdel-Bakky MS, Keller S, Altevogt P, Oh 439 ST, Reichrath J, Oxmann D, Pfeilschifter J, Mihic-Probst D and Gutwein P: ADAM10 Is 440 Upregulated in Melanoma Metastasis Compared with Primary Melanoma. Journal Of Investigative 441 Dermatology 130: 763-773, 2010. 442 39. Cireap N and Narita D: Molecular Profiling of ADAM12 and ADAM17 Genes in Human 443 Malignant Melanoma. Pathology & Oncology Research 19: 755-762, 2013. 444 40. Anders A, Gilbert S, Garten W, Postina R and Fahrenholz F: Regulation of the alpha-secretase 445 ADAM10 by its prodomain and proprotein convertases. Faseb Journal 15: 1837-+, 2001. 446 41. Srour N, Lebel A, McMahon S, Fournier I, Fugere M, Day R and Dubois CM: 447 TACE/ADAM-17 maturation and activation of sheddase activity require proprotein convertase 448 activity. Febs Letters 554: 275-283, 2003. 449 42. Schlondorff J, Becherer JD and Blobel CP: Intracellular maturation and localization of the 450 tumour necrosis factor alpha convertase (TACE). Biochemical Journal 347: 131-138, 2000. 451 43. Laprise MH, Grondin F, Cayer P, McDonald PP and Dubois CM: Furin gene (fur) regulation in 452 differentiating human megakaryoblastic Dami cells: involvement of the proximal GATA recognition 453 motif in the P1 promoter and impact on the maturation of furin substrates. Blood 100: 3578-3587, 454 2002. 455 44. Ayoubi TAY, Creemers JWM, Roebroek AJM and Vandeven WJM: Expression Of the Dibasic 456 Proprotein Processing Enzyme Furin Is Directed by Multiple Promoters. Journal Of Biological 457 Chemistry 269: 9298-9303, 1994. 458 45. Creemers JWM, Siezen RJ, Roebroek AJM, Ayoubi TAY, Huylebroeck D and Vandeven WJM: 459 Modulation Of Furin-Mediated Proprotein Processing Activity by Site-Directed Mutagenesis. 460 Journal Of Biological Chemistry 268: 21826-21834, 1993. 461 46. Gendron F, MacMahon S, Dubois C, Asselin C and Rivard N: Transcriptional regulation of the 462 proprotein convertase, furin, by the caudal-related homeodomain protein CDX2 in intestinal 463 epithelial cells. Molecular Biology Of the Cell 15: 86a-86a, 2004. 464 47. Gendron FP, Mongrain S, Laprise P, McMahon S, Dubois CM, Blais M, Asselin C and Rivard N: 465 The CDX2 transcription factor regulates furin expression during intestinal epithelial cell 466 differentiation. American Journal Of Physiology-Gastrointestinal And Liver Physiology 290: 467 G310-G318, 2006. 468 48. Khatib AM, Siegfried G, Chretien M, Metrakos P and Seidah NG: Proprotein convertases in 469 tumor progression and malignancy - Novel targets in cancer therapy. American Journal Of Pathology 470 160: 1921-1935, 2002. 471 49. Khatib AM, Siegfried G, Prat A, Luis J, Chretien M, Metrakos P and Seidah NG: Inhibition of 472 proprotein convertases is associated with loss of growth and tumorigenicity of HT-29 human colon 473 carcinoma cells - Importance of insulin-like growth factor-1 (IGF-1) receptor processing in 474 IGF-1-mediated functions. Journal Of Biological Chemistry 276: 30686-30693, 2001. 475 50. Basak A, Chen A, Scamuffa N, Mohottalage D, Basak S and Khatib AM: Blockade of Furin 476 Activity and Furin-Induced Tumor Cells Malignant Phenotypes By The Chemically Synthesized 477 Human Furin Prodomain. Current Medicinal Chemistry 17: 2214-2221, 2010. 478
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
15
51. Blanchette F, Day R, Dong W, Laprise MH and Dubois CM: TGF beta 1 regulates gene 479 expression of its own converting enzyme furin. Journal Of Clinical Investigation 99: 1974-1983, 480 1997. 481 52. Dickson MC, Slager HG, Duffie E, Mummery CL and Akhurst RJ: Rna And Protein 482 Localizations Of Tgf-Beta-2 In the Early Mouse Embryo Suggest an Involvement In Cardiac 483 Development. Development 117: 625-639, 1993. 484 53. Lee JE, Pintar J and Efstratiadis A: Pattern Of the Insulin-Like Growth Factor-Ii 485 Gene-Expression during Early Mouse Embryogenesis. Development 110: 151-&, 1990. 486 54. Bedogni B, Warneke JA, Nickoloff BJ, Giaccia AJ and Powell MB: Notch1 is an effector of Akt 487 and hypoxia in melanoma development. Journal Of Clinical Investigation 118: 3660-3670, 2008. 488 55. Razorenova OV, Agapova LS, Budanov AV, Ivanov AV, Strunina SM and Chumakov PM: 489 Retroviral reporter systems for assessing the activity of stress-inducible signal transduction pathways 490 controlled by the p53, HIF-1, and HSF-1 transcription factors. Molecular Biology 39: 253-259, 2005. 491 56. Palomero T, Lim WK, Odom DT, Sulis ML, Real PJ, Margolin A, Barnes KC, O'Neil J, 492 Neuberg D, Weng AP, Aster JC, Sigaux F, Soulier J, Look AT, Young RA, Califano A and Ferrando 493 AA: NOTCH1 directly regulates c-MYC and activates a feed-forward-loop transcriptional network 494 promoting leukemic cell growth. Proceedings Of the National Academy Of Sciences Of the United 495 States Of America 103: 18261-18266, 2006. 496 57. Nishimura M, Isaka F, Ishibashi M, Tomita K, Tsuda H, Nakanishi S and Kageyama R: 497 Structure, chromosomal locus, and promoter of mouse Hes2 gene, a homologue of Drosophila hairy 498 and Enhancer of split. Genomics 49: 69-75, 1998. 499 58. Proweller A, Pear WS and Parmacek MS: Notch signaling represses myocardin-induced smooth 500 muscle cell differentiation. J Biol Chem 280: 8994-9004, 2005. 501 59. Buas MF, Kabak S and Kadesch T: Inhibition of myogenesis by Notch: evidence for multiple 502 pathways. J Cell Physiol 218: 84-93, 2009. 503 60. Shaverdashvili K and Bedogni B: Matrix Metalloproteinase MT1-MMP Regulates Melanoma 504 Metastasis by Activation RAC1. Journal Der Deutschen Dermatologischen Gesellschaft 11: 44-44, 505 2013. 506 61. Knauper V, Will H, Lopez-Otin C, Smith B, Atkinson SJ, Stanton H, Hembry RM and Murphy 507 G: Cellular mechanisms for human procollagenase-3 (MMP-13) activation. Evidence that 508 MT1-MMP (MMP-14) and gelatinase a (MMP-2) are able to generate active enzyme. J Biol Chem 509 271: 17124-31, 1996. 510 62. Sato H, Takino T, Okada Y, Cao J, Shinagawa A, Yamamoto E and Seiki M: A matrix 511 metalloproteinase expressed on the surface of invasive tumour cells. Nature 370: 61-5, 1994. 512 63. Strongin AY, Collier I, Bannikov G, Marmer BL, Grant GA and Goldberg GI: Mechanism of 513 cell surface activation of 72-kDa type IV collagenase. Isolation of the activated form of the 514 membrane metalloprotease. J Biol Chem 270: 5331-8, 1995. 515 64. Shaverdashvili k. WP, Ma J., Zhang K., Osman I., Bedogni B.: MT1-MMP modulates 516 melanoma cell dissemination and metastasis through activation of MMP2 and RAC1. Pigment 517 Cell and melanoma Res 27: 287-96, 2014. 518 65. Nichols JT, Miyamoto A, Olsen SL, D'Souza B, Yao C and Weinmaster G: DSL ligand 519 endocytosis physically dissociates Notch 1 heterodimers before activating proteolysis can occur. 520 Journal Of Cell Biology 176: 445-458, 2007. 521 66. van Tetering G and Vooijs M: Proteolytic Cleavage of Notch: "HIT and RUN". Current 522 Molecular Medicine 11: 255-269, 2011. 523 67. Guimont P GF, Dubois CM.: Sox9-dependent transcriptional regulation of the proprotein 524 convertase furin. Am J Physiol Cell Physiol 293: C172-83, 2007. 525
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
16
68. Blanchette F RP, Grondin F, Attisano L, Dubois CM: Involvement of Smads in 526 TGFbeta1-induced furin (fur) transcription. J Cell Physiol 188: 264-73, 2001. 527 69. Chang HM CJ, Klausen C, Leung PC.: Recombinant BMP4 and BMP7 increase activin A 528 production by up-regulating inhibin βA subunit and furin expression in human granulosa-lutein cells. 529 J Clin Endocrinol Metab 100: E375-86, 2015. 530 70. Remacle AG, Chekanov AV, Golubkov VS, Rozanov DV, Fugere M, Day R and Strongin AY: 531 Proprotein convertases and glycosylation regulate MT1-MMP activity. Matrix Biology 25: S49-S49, 532 2006. 533 71. Bedogni B and Powell MB: Unique transforming properties of Notch1 in human melanocytes. 534 Pigment Cell & Melanoma Research 22: 702-703, 2009. 535 72. Bedogni B and Powell MB: Hypoxia, melanocytes and melanoma - survival and tumor 536 development in the permissive microenvironment of the skin. Pigment Cell & Melanoma Research 537 22: 166-174, 2009. 538 73. Brou C, Logeat F, Gupta N, Bessia C, LeBail O, Doedens JR, Cumano A, Roux P, Black RA 539 and Israel A: A novel proteolytic cleavage involved in Notch signaling: The role of the 540 disintegrin-metalloprotease TACE. Molecular Cell 5: 207-216, 2000. 541 74. Hartmann D, De Strooper B, Serneels L, Craessaerts K, Herreman A, Annaert W, Brabant V, 542 Luebke T, Illert AL, von Figura K and Saftig P: Deficiency for the disintegrin metalloprotease 543 ADAM10 causes disturbed alpha-secretase function and a notch deficiency-related phenotype in 544 mice. Neurobiology Of Aging 23: S183-S183, 2002. 545 75. Krebs LT, Xue YZ, Norton CR, Shutter JR, Maguire M, Sundberg JP, Gallahan D, Closson V, 546 Kitajewski J, Callahan R, Smith GH, Stark KL and Gridley T: Notch signaling is essential for 547 vascular morphogenesis in mice. Genes & Development 14: 1343-1352, 2000. 548 76. Limbourg FP, Takeshita K, Radtke F, Bronson RT, Chin MT and Liao JK: Essential role of 549 endothelial Notch1 in angiogenesis. Circulation 111: 1826-1832, 2005. 550 77. Ntziachristos P LJ, Sage J, Aifantis I.: From fly wings to targeted cancer therapies: a centennial 551 for notch signaling. Cancer Cell 25: 318-34, 2014. 552 553
554
555
556
557
FOOTNOTES: 558
This work was supported by a grant from the Harry Lloyd Charitable Trust and by a grant from the 559
Concern Foundation. 560
561
562
563
564
565
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
17
FIGURE LEGENDS 566
Figure 1. Notch1 promotes MT1-MMP activity. A. The active MT1-MMP fraction (55kDa, bottom 567
band) is increased in Notch1-NIC expressing cells. B. Notch1-NIC increases MT1-MMP in vitro 568
activity. Neg=negative control (buffer); pos= positive control (0.5 μg/ml recombinant active 569
MT1-MMP). C. Zymography image of MMP2 processing in pLM or NIC expressing cells. 570
Numbers represent fold induction of active band (bottom band) over controls (pLM) ±SD. Results 571
are the average between three repeats. *p<0.05, Student’s T test. D. Notch1 increases both mRNA 572
and protein levels of Furin, but doesn’t affect the expression of MT1-MMP. qRT-PCR and Western 573
blot of the cells in A showing expression changes of Furin and Furin like convertases. Differences in 574
expression are significant (p<0.001, Student’s T test). E. Inhibition of Notch1 reduces both Furin 575
mRNA and protein. *p<0.001, Student’s T test. β-actin is used as loading control. 576
Figure 2. Stimulation of endogenous Notch1. JAGGED1 dependent activation of endogenous 577
Notch1 increases furin and promotes MT1-MMP and ADAM10 processing in two melanoma cell 578
lines. 579
Figure 3. Furin is a direct target of Notch1. A. Schematic representation of the Furin promoter. 580
There are three putative CSL binding sequences TTCCCAC, located at -3556 (B1), -1236 (B2), and 581
+960 (B3). B. Western blot analysis in V2387 and WM266-4 cells showing Notch1 protein levels in 582
control and shNotch1 expressing cells. C. ChIP analysis showing association of Notch1 to the 583
binding site -1236 (B2) both in V2387 and WM266-4 cells (**, *** p<0.01, Student’s T test). D. 584
The HES1 promoter was used as a positive control for CSL/Notch-binding activity (**, *** p<0.01, 585
Student’s T test). All data are representative of three independent experiments. 586
Figure 4. Notch1 affects Furin expression through binding to the Furin promoter. A. Schematic 587
representation of the Luciferase reporter constructs containing a WT sequence TTCCCAC or the 588
mutated sequence TCACAGC. The construct SacI contains the sequence from PstI at the TSS 589
(transcription start site) to SacI. B. Luciferase reporter assay in V2387 cells in control (PLM) or 590
Notch1-NIC expressing cells. Constructs: -Basic indicates pGL2 basic – luciferase construct without 591
promoter region and represents the negative control for the luciferase assay; HES1 contains the 592
promoter region of the Notch1 target HES1 (positive control; *p<0.01, Student’s T test). A 593
significant induction of luciferase activity was observed in NIC expressing cells when using the WT 594
construct SacI (**p<0.01, Student’s T test). C. Induction of luciferase in NIC expressing cells is seen 595
in WT SacI expressing cells (*p<0.01, Student’s T test), and it is significantly reduced when the SacI 596
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
18
construct is mutated (**p<0.01, Student’s T test). Upper panel: Notch1-NIC protein levels in pLM 597
and NIC expressing cells. Experiments were done in three cell lines: WM115, Skmel2, V2387. Data 598
are the average of three independent experiments. 599
Figure 5. Notch1 induces MT1-MMP activation through Furin. A. Western blot analysis showing 600
the active fraction of MT1-MMP increases in NIC expressing cells, but decreases in Furin knockdown 601
cells. Experiments were done in three cell lines: SkMel2, V2387, and WM115, as shown. Values 602
reported below the blots are the average quantification of furin bands ±SD among three separate 603
experiments. B. Ratio between the active and inactive (pro-) MT1-MMP bands normalized to the 604
corresponding β-actin. Data are the average between three independent experiments (pSKMel2<0.01; 605
pV2387<0.01; pWM115<0.01, Student’s T test). 606
Figure 6. Notch1 promotes ADAM10 activity. A. In Skmel2 and V2387 cells, the active ADAM10 607
fraction (60kDa, bottom band) is increased in Notch1-NIC expressing cells. ADAM17 does not 608
change. Ratio between inactive (pro) and active ADAM10 normalized to their corresponding β-actin 609
is indicated. Values are the average ± standard deviation of three independent experiments. B. 610
qRT-PCR of ADAM10 in pLM or NIC expressing cells showing mRNA levels of ADAM10 611
normalized to GAPDH. C. ADAM10 in vitro activity assay in pLM or NIC expressing cells. Fold 612
change of RFU (relative fluorescence units) in SKMel2 and V2387 cells were detected. Statistical 613
significance is indicated (Student’s T test). 614
Figure 7. Notch1 increases the active fraction of ADAM10 through Furin. A. Western blot 615
analysis showing expression of pro and active ADAM10 and Notch1-NIC in NIC expressing cells 616
(SKMel2 – left panel; V2387 – right panel) in the presence of shGFP or shFurin (shF2 and shF3). B. 617
Ratio between active and inactive ADAM10 of the lanes (normalized to β-actin) in the blots in A. 618
*pSKMel2<0.01; pV2387<0.01, Student’s T test. Data are the average of four independent 619
experiments. C-D. ADAM10 in vitro activity assay in SKmel2 and V2387 expressing empty vector 620
(pLM) or active Notch1 (NIC) in the presence of shGFP or shFurin (shF2 and shF3). Statistical 621
significance is indicated (Student’s T test). Results are the average of four independent experiments. 622
Figure 8. Inhibition of Furin decreases Notch1 processing. A. Notch1 full length (FL) and 623
intracellular (NIC) (Ab: mN1A), Notch-NIC (Ab: Cleaved Val1744), MT1-MMP, ADAM10 and 624
Furin expression in cells stimulated with recombinant JAGGED1 and expressing either empty vector 625
(pLM) or dominant negative MAML (MAML-DN). B. Notch1 full length (FL) and intracellular 626
(NIC) (Ab: mN1A), Notch-NIC (Ab: Cleaved Val1744), MT1-MMP, ADAM10 and Furin expression 627
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from

Furin is a direct transcription target of Notch1
19
in cells stimulated with recJAGGED1 and expressing a specific furin shRNA (shF2) or control 628
(shGFP). The graphs are the means±SD of the ratio between NIC and FL Notch1 among two 629
independent experiments. P values were calculated by the Student’s T test. 630
Figure 9. Furin inhibition reduces melanoma cell migration and invasion. A. Expression levels 631
of Notch1-NIC and furin in cells expressing active Notch1 and furin shRNA. B. Migration (scratch 632
assay) over 24 hours of the cells in A. C. Invasion through Matrigel over a 24 hours period of the 633
cells in A. Reduction of migration and invasion in shFurin expressing cells is statistically significant 634
(Student’s T test). 635
on April 13, 2018 by guest
http://mcb.asm
.org/D
ownloaded from