fulltext.pdf
TRANSCRIPT

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
Journal of Chemical Ecology, Vol. 30, No. 2, February 2004 (©C 2004)
MALE AGGREGATION PHEROMONE OF DATE PALM FRUITSTALK BOREROryctes elegans
DIDIER ROCHAT,1,∗ KAZEM MOHAMMADPOOR,2
CHRISTIAN MALOSSE,1 ARMAN AVAND-FAGHIH, 3
MARTINE LETTERE,1 JOSIANE BEAUHAIRE,1 JEAN-PAUL MORIN,4
ADELINE PEZIER,1 MICHEL RENOU,1
and GHOLAM ABBAS ABDOLLAHI3
1INRA, Unite de Phytopharmacie et Mediateurs ChimiquesCentre de Versailles, Route de Saint-Cyr
78026 Versailles Cedex, France2Saravan Plant Pests & Diseases Research Laboratory
P.O. Box 364, 99515 Saravan, Iran3Harmful Insects to Plants Research Department
Plant Pests & Diseases Research Institute, P.O. Box 145419395 Tehran, Iran
4CIRAD-CP, TA 80/02, Avenue Agropolis34398 Montpellier Cedex 5, France
(Received May 26, 2003; accepted September 8, 2003)
Abstract—Laboratory and field investigations were carried out to characterizethe chemical communication system of the date palm fruit stalk borer,Orycteselegans, and to develop pheromone-based trapping in Eastern Iran. Adults ofboth sexes feeding on date palm pieces attracted conspecifics, whereas datepalm alone was minimally attractive. Males were twice as attractive as females.More beetles were captured at the palm crown than at ground level. Odorsfrom adults feeding on sugarcane were sampled and analyzed by gas chro-matography and mass spectrometry. Whereas females did not emit sex specificvolatiles, males emitted a blend of 4-methyloctanoic acid (1: major component)and ethyl 4-methyloctanoate (2), occasionally mixed with minor components:4-methyloctanyl acetate (3), methyl 4-methyloctanoate (4), 4-methyloctanol (5),and nonanyl acetate (6). Electroantennography and field trapping experimentsdemonstrated that compound1 is an essential component of the male aggregationpheromone ofO. elegans.It was barely attractive by itself but synergistic withfresh date palm odor. It attracted many more beetles than any of compounds2–6. The addition of one or several of compounds2–6 to 1 did not improvetrap captures. During the course of 2 years, we captured 4000 beetles, with a
∗To whom correspondence should be addressed. E-mail: [email protected]
387
0098-0331/04/0200-0387/0C© 2004 Plenum Publishing Corporation

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
388 ROCHAT ET AL.
weekly average of 6.3 beetles/trap, and were able to monitor the seasonal flightof O. elegans. Our results provide the basis for developing mass trapping forcontrol of this pest.
Key Words—Coleoptera, Scarabaeidae,Oryctes, aggregation pheromone, EAG,field trapping, date palm, synergy.
INTRODUCTION
Rhinoceros beetles constitute a group of medium- to large-sized scarabs of theDynastinae subfamily whose males are characterized by a large cephalic horn.Among rhinoceros beetles, the genusOryctesis specific to the Old World andincludes about 40 species (Lepesme, 1947; Endr¨odi, 1985). Many rhinoceros bee-tles are associated with palm trees and are severe pests of coconut, oil, and datepalms (Lepesme, 1947; Bedford, 1980). Adult beetles burrow galleries in the freshtissues of palms for feeding. The damage is particularly severe when the apicalbud is attacked in the young trees or the fruit-stalk in producing trees. Indirectdamage often occurs in areas occupied by palm weevils (Rhynchophorusspp.),which lay eggs in the galleries made by the dynastine beetles. Palm weevil infes-tations are often fatal to palms. Larvae have no direct impact on plant productionbecause most of them feed on dead tissue. Despite insecticides active against mostrhinoceros beetles, efficient and acceptable methods of controlling them are stilllacking in many cases, either because of economic reasons, or more generally,because adults spend most of their life hidden in galleries, and are excellent flyers,capable of rapidly colonizing new feeding and breeding sites.
The idea of manipulating adult behavior, especially luring beetles into traps,was investigated in the 1970s. Some attractants were reported but subsequentlyabandoned (Julia and Mariau, 1976; Vander Meer et al., 1979; Vander Meer andMcGovern, 1983). Recently, male aggregation pheromones were reported fromfour rhinoceros beetles attacking palm trees:Oryctes monoceros(Olivier) in Africa(Gries et al., 1994),Oryctes rhinocerosL. (Hallet et al., 1995; Morin et al., 1996)in the Asia-Pacific area (further referred to as “the Tropical Oryctes”),Scapanesaustralis Boisduval in the New Guinea region, andStrategus aloeusL. in theNew World (Rochat et al., 2000b, 2002). Synthetic pheromone makes it possibleto capture these species in bucket traps. Applications on a large scale have beenreported forO. rhinocerosandScapanes australis(Hallet et al., 1995; Ho, 1996;Chung, 1997; Purba et al., 2000) but not forO. monocerosandS. aloeus. Forboth O. rhinocerosandS. australis, attraction to the pheromone was enhancedby, or synergistic with, plant odors from decaying wood inO. rhinoceros(Halletet al., 1995; Alfiler, 1999; Sudharto et al., 2001), or from fresh palm tissues inS.australis(Rochat et al., 2002).
ThreeOryctesspecies develop specifically in date palms and are present in theNear- and Middle-East and in North Africa:O. elegansPrell [=sinaicusPetrovitz

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 389
(non Walker)], O. agamemnonBurmeister (=desertorumArrow, =persicusEndrodi), andO. richteri Petrovitz (Endr¨odi and Petrovitz, 1974). The first two arecommon throughout the Near- and Middle-East and seem to be sympatric in manyplaces. They are univoltine, flying from spring to autumn. Adults cause damageby mining the stalks of fruit bunches. In the Tropical Oryctes, larvae develop inthe wood of dead trees, and adults feed on living trees. The two feeding habitatsare separate in space. In contrast, larvae and adults of the date palmOryctesspp.live in the same living and standing trees. Larvae develop in the crown and at theperiphery of the stem, feeding at the interface of dead and living tissues. Thoughnot reported as major pests,O. elegansandO. agamemnoninflict chronic dam-age in date palm groves and cause economic losses in Iran and Iraq (Hurpinand Fresneau, 1969; Gharib, 1970; Hussain, 1974). Insecticide applications in thecrown against emerging adults are commonly made in Iran in spring and summer.Winter pruning and burning of the dead fronds and of the dried frond bases justbelow the crown is recommended and applied by some growers to eliminate lar-vae and pupae. Both measures require considerable labor and their impact is notknown.
As ethyl 4-methyloctanoate had been reported as the male aggregationpheromone of the Tropical Oryctes (Gries et al., 1994; Hallet et al., 1995; Morinet al., 1996) and also identified in a preliminary analysis of airborne effluvia fromO. elegansmales (Rochat and Malosse, unpublished), we tested this compoundas an attractant for trappingO. elegansin Southern and Eastern Iran in 1996–97.Several weeks of trapping in highly infested areas, without using plant material asa co-attractant led to the capture of just two adults ofO. elegans(Avand-Faghihand Rochat, unpublished). To understand this apparent contradiction and developa trapping strategy against this insect, we undertook the study of the chemicalcommunication ofO. elegans. The work was based on a field approach withexperimental trapping using live insects followed by screening of synthetic pu-tative pheromone compounds selected from the results of volatile collections, gaschromatography–mass spectrometry analyses, organic synthesis, and electroan-tennographic screening.
Here we report the identification and activity of the male aggregationpheromone ofO. elegansand provide practical data to develop an efficient trap-ping system. The result of a 6-month population monitoring study is presented, andthe differences between the chemical communication systems of the rhinocerosbeetles are discussed.
METHODS AND MATERIAL
Synthetic Chemicals.Chiral molecules were used as racemic mixes.4-Methyloctanoic acid (1) and ethyl 4-methyloctanoate (2) were purchased fromE.G.N.O.-Chimie (Tancarville, France) and served as starting material to prepare

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
390 ROCHAT ET AL.
4-methyloctanyl acetate (3), methyl 4-methyloctanoate (4), and 4-methyloctanol(5).4was prepared from a mixture of methanol,1, and sulfuric acid;5was obtainedby reduction of2 with LiAlH 4 in anhydrous diethyl ether;3 and nonanyl acetate(6) were prepared from5 and nonanol, respectively using acetic anhydride in pyri-dine. 1–6 were>98% pure by gas chromatography (GC) and3–5 were free ofdetectable traces of1 and2. Octanoic (=caprylic;7) and nonanoic (=pelargonic;8) acids were purchased from Aldrich (7: UK and8: Germany) and were 99.5 and96% pure, respectively.
Insects. O. eleganswere collected in date palm groves in the Saravan area,shipped by air to France, and maintained on sugarcane at 23–28◦C, 75% RH, andunder a 13L:11D photoperiod. Sugarcane (commercial source) was used as foodto replace date palm core not available in France, as for other palm rhinocerosbeetles (Morin et al., 1996; Rochat et al., 2000b, 2002).
Collection of the Natural Pheromone. O. eleganspheromone was collectedfrom effluvia by passing a stream of air (purified over activated charcoal; 100 ml/min)into 1-l glass flasks containing four to six adult insects. Odors were adsorbed onto350 mg SupelpakTM-2 (SUPELCO, USA) for 4–7 d. Effluvia were trapped frombatches of males, or females, or both sexes placed on two 20 cm halved sugarcanepieces, and from sugarcane controls. Five series of collections were carried outbetween 1996 and 2002, yielding extracts from males (N = 7), females (N = 6),and males plus females (N = 2) with sugarcane, and sugarcane control (N = 4)odors. The adsorbent was eluted with 1.5 ml 99.5% dichloromethane (1996 and1998) or hexane (2000–2002). The extracts were stored at−40◦C and used for GCand/or GC–MS.
GC–MS analyses were carried out using a Varian 3300 gas chromatographcoupled to a Nermag R10-10C quadrupole mass spectrometer. Electron impactspectra were obtained at 70 eV. Additional spectra were obtained in chemicalionization mode using NH3 as reactant gas. The chromatograph was typicallyequipped with a WCOT fused silica column (30 m× 0.32 mm id, 0.5-µm MDN-5S phase; Supelco, Bellefonte, PA, USA) operated from 50◦C (1 min) to 300◦C at10◦C/min. Additional GC analyses were carried out using a Varian 3400 CX GCwith an equivalent apolar column or a polar column with same dimensions (RTX-Wax phase; Restek, Bellefonte, PA, USA) and operated from 50◦C (1 min), thenheated at 15◦C/min to 80◦C/(9 min), then heated to 240◦C at 6◦C/min. Helium wasused as carrier gas at 13 psi. Samples were injected using a SPI injector at 250◦C.The amount of each male-specific compound was estimated by the percentage ofits FID (GC) or TIC (GC–MS) response in the total response recorded to all themale-specific compounds (Table 1).
Electroantennography.Electroantennograms (EAGs) were recorded usingthe same equipment as described forO. rhinoceros(Morin et al., 1996), fromdissected antennae. The antennal lamellae were held apart using small insect pins.The recording electrode was placed on the central lamella, while the reference

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 391
TAB
LE1.
CH
AR
AC
TE
RIS
TIC
SA
ND
AB
UN
DA
NC
EO
FT
HE
MA
LE-S
PE
CIF
ICC
OM
PO
UN
DS
IDE
NT
IFIE
DB
YG
AS
CH
RO
MA
TO
GR
AP
HY
AN
DM
AS
S
SPE
CT
RO
ME
TR
YF
RO
MO
ryct
es
ele
ga
nsE
FF
LUV
IAT
RA
PP
ED
ON
SUP
ELP
AKT
M-2
GC
rete
ntio
ntim
es(m
in)
Mol
ecul
arA
pola
rP
olar
EIm
ass
spec
trum
at70
eVc
%ra
tioan
dfr
eque
ncy
Com
poun
dw
eigh
tR
TX
-5M
SaR
TX
-Wax
bch
arac
teris
ticio
nsm
/z(%
)in
mal
eef
fluvi
ad
1:4-
Met
hylo
ctan
oic
acid
158
14.8
5–15
.35
e29
.22
41,
43,
55,
57(1
00),
60,
69,
73,
83,9
9,10
1,12
953±
11(8
)
2:E
thyl
4-m
ethy
loct
anoa
te18
615
.24
14.7
041
,43
,45
,55
,57
,60
,61
,70
,73
,83
,88
(100
),99
,10
1,12
3,12
9,14
1,15
7
37±
10(8
)
3:4-
Met
hylo
ctan
ylac
etat
e18
615
.61
16.3
341
,43
(100
),56
,61
,69
,70
,84
,98
,101
,126
8±
4(4
)
4:M
ethy
l4-m
ethy
loct
anoa
te17
213
.19
12.7
941
,43
(100
),55
,57
,59
,69
,74
,87
,99,
115,
123,
141,
143
1±
1(3
)
5:4-
Met
hylo
ctan
ol14
411
.85
19.0
741
,43
,45
,56
,69
(100
),70
,84
,98
,126
1±
1(3
)
6:N
onan
ylac
etat
e18
617
.05
17.9
841
,43
(100
),55
,56
,61
,70
,84
,98
,116
,126
0±
1(2
)
a25
m×
0.32
mm
id×
0.5µ
mdf
(Res
tek)
oper
ated
from
50◦ Cfo
r1
min
,50–
100◦ C
at15◦ C
/min
,1m
inat
100◦ C
,and
100–
280◦ C
at5◦
C/m
in.
b30
m×
0.32
mm
id×
0.5µ
mdf
(Res
tek)
oper
ated
from
50◦ Cfo
r1
min
,50–
80◦ C
at15◦ C
/min
,9m
inat
80◦ C
,and
80–2
40◦C
at6◦
C/m
in.
c Ner
mag
R10
-10C
quad
rupo
lem
ass
spec
trom
eter
.dM
ean±
S.E
.bas
edon
tota
lFID
orT
ICre
spon
sem
easu
red
for
the
mal
e-sp
ecifi
cpe
aks.
The
num
ber
ofoc
curr
ence
sis
give
nin
brac
kets
(n=
9).
e Elu
tes
asa
broa
das
ymm
etric
peak
over
lapp
ing
with
the
peak
sof
2an
d3
whe
nla
rge
amou
nts
are
inje
cted
.

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
392 ROCHAT ET AL.
electrode was pushed into the lumen of the scape. Each antenna was bathed by aflow of moist air (400 ml/min) and stimulated with compounds1–4 and7–8. Eachcompound was delivered at four doses, selected randomly from serial dilutionsin hexane (0.1–100µg; decadic step) The antenna was stimulated by hexane(solvent) at the beginning, the end, and between each dose series. Compoundswere presented as 1µl of hexane solutions deposited on filter papers inserted inPasteur pipettes just prior to the stimulation (0.5 sec; 200 ml/min). Raw EAGvalues for a given antenna were normalized to relative EAGs (percent) by dividingthem by the average response recorded to the complete stimulation series.
Field Trapping Experiments: Chemical Dispensers.Synthetic chemicalswere emitted from heat-sealed sachets made of polyethylene films (ALPLAST,France), the characteristics and dimensions of which were adapted to the chem-icals and the targeted release rates after laboratory calibration (e.g., 28× 40×0.2 mm filled with 0.7 ml to emit1 at 3 mg/d at 35◦C). Calibration was achievedby preparing dispensers of various sizes and different film thicknesses, filled withthe pure chemicals, and left in a room thermostated at 30, 35, and 40◦C for one wkat each temperature. The dispensers were weighed daily, and the slopes obtainedfrom the linear adjustment of the weight loss versus time furnished the estimationsof the evaporation rates. The dispensers were stored at−10◦C and weighed (mgprecision) prior to and at the end of each replicate or assay to determine the dailyrelease rates of the chemicals (reported in the figures and tables).
Location and General Features.All assays were carried out close to thePakistan border in Saravan or Zaboli, a place about 100 km west from Saravan.Both areas are located in Sistan and Balootchestan province of Iran and constitutethe eastern limit of the area of date palm production in Iran, with a mean elevationof ca. 1200 m above sea level and having cold winters. The areas consist of scatteredoases, with a traditional farming system of mixed vegetables, pasture, and fruitcrops, with few or no chemical inputs, either fertilizer or pesticide. Date palms aremostly Mazafati variety, in highly heterogeneous plots.
Traps consisted of 24-l plastic buckets whose lids were equipped with eightradial 8× 5 cm openings. All treatments (except Assay 8) included one piece ofdate palm core (ca. 1 kg) freshly cut from a shoot. The palm piece was wrappedin a punctured plastic bag to limit dehydration (synthetic lures) or put in a 1-lperforated plastic box with two liveO. elegans. In Assay 8, date palm core testedwithout beetle was prepared as described above or by chopping the palm piece andcrushing the tissue with a hammer.
In Assays 1–2, traps were put on the ground at the base of a date palm. FromAssay 3 onwards, traps were hung from the stem at 2.5–3 m above ground level,generally positioned just below the crown of the palms. Soapy water was pouredinto the bottom of the bucket to prevent beetles from escaping. No insecticide wasused. Assays 1–8 were carried out in randomized complete block designs. Trapswere spaced≥80 m apart. The two blocks were separated by≥200 m. One assay

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 393
consisted ofn consecutive periods designated as replicates. During each replicate,the date palm pieces remained unchanged and the traps were visited at regularintervals (every 3 or 7 d) to collect trapped beetles, check the health of theO.elegansused as bait, and replace dead bait insects and empty chemical dispensersas needed. Date palm tissue was renewed and the treatments were re-randomizedat each replicate (Table 2).
Chronology and Experimental Choices.The experiments are summarized inTable 2. Host plant was systematically included as bait (except specific Assay 8)because compound2 alone had captured almost no beetles. The attraction to con-specifics was investigated first. Then we searched for the best trap positioning(Assays 1–3). Synthetic chemicals were further evaluated as pheromone candi-dates in various situations (Assays 4–8). Finally, Assay 9 evaluated the feasibilityof monitoringO. eleganspopulation over one flight season.
Statistical Analysis.(Minitab, 1998). Assays 4–8: The total captures per trapper visit were analyzed by a three-way ANOVA (F1: bait; F2: age of date palmtissue–no. of visit within replicate; F3: replicate) on Ln(x + 1) transformed datausing a general linear model (GLM) procedure. Means were compared by multiplecomparison (Tukey test) or comparison to a reference (Dunnett test) withP = 0.05.The data from the four assays were pooled and subjected to another commonANOVA to evaluate the effect of the age of the date palm tissue. In assays 1–3,we could not apply ANOVA to the raw data because too many values were zero.We performed an ANOVA on cumulative captures per trap: the total per replicateand the mean daily total per assay for Assays 3 and 1–2, respectively. Results ofAssays 1–2 were analyzed together because both had the same structure and gavesimilar results. The sex ratio of the captures was analyzed similarly in all assays.Details of these analyses are not presented because the sex ratio appeared not tobe correlated to the experimental variables and to be variable across the trappingperiods (e.g., Figure 1).
RESULTS
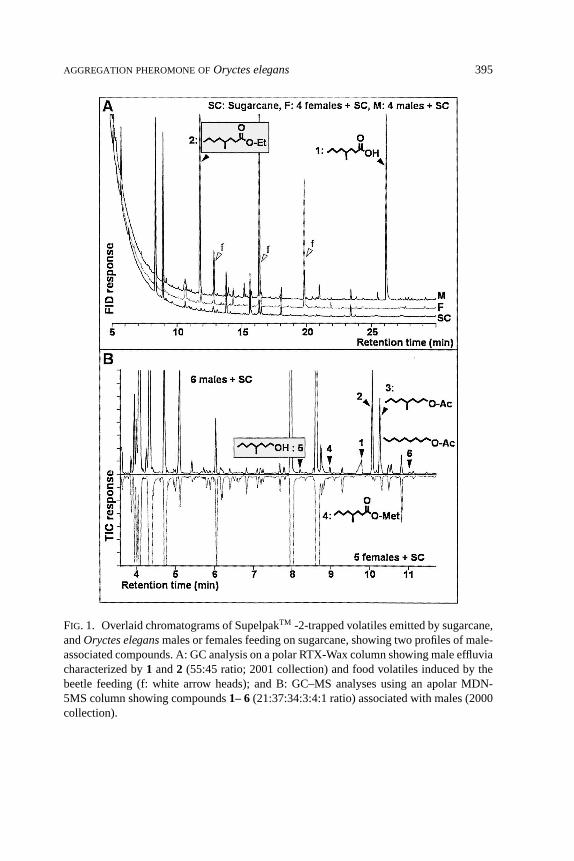
Effluvia Characteristics and Pheromone Structural Identification.Feedingby beetles of either sex induced the emission of various molecules (f; Figure 1A).Comparisons of the odors emitted by only sugarcane, and males, females, or bothsexes feeding on sugarcane showed that the presence of males was correlated withat least six volatile molecules occurring in variable proportions (Figure 1; Table 1).In contrast, no female-produced chemicals were detected. Based on their massspectra and retention times by GC, and comparisons with reference compounds,the male-specific molecules were identified as1: 4-methyloctanoic acid,2: ethyl4-methyloctanoate,3: 4-methyloctanyl acetate,4: methyl 4-methyloctanoate,5: 4-methyloctanol, and6: nonanyl acetate (Table 1). At least one of these compounds
P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
394 ROCHAT ET AL.
TAB
LE2.
GE
NE
RA
LC
HA
RA
CT
ER
IST
ICS
OFN
INE
FIE
LDE
XP
ER
IME
NT
SC
AR
RIE
DO
UT
TO
TR
AP
Ory
cte
se
leg
an
sUS
ING
NA
TU
RA
LO
R
SYN
TH
ET
ICA
TT
RA
CT
AN
TS
INE
AS
TE
RN
IRA
NF
RO
M19
98T
O20
02W
ITH
RE
FE
RE
NC
ET
OT
HEF
IGU
RE
SA
ND
TAB
LES
SHO
WIN
GT
HE
RE
SU
LTS
Cha
ract
eris
tics:
bloc
ks,
Pur
pose
(no.
trea
tmen
ts)
no.×du
ratio
nof
repl
icat
eE
xpt.
#B
aits
(add
edto
date
palm
core
exce
ptA
ssay
8)(f
requ
ency
ofvi
sits
)a
Dat
ean
dpl
ace
1A
ttrac
tion
byco
nspe
cific
san
dse
xef
fect
(4)
4bl
ocks
,4×14
dM
ay21
–Jul
20,1
998,
Zab
oli
Ba
its:2
,2,2
+2
,no
beet
les
(c)
(7d)
Zab
oli
2A
ttrac
tion
byco
nspe
cific
san
dse
xef
fect
(4)
4bl
ocks
,10
×9
dM
ay5–
Aug
3,19
99,S
arav
anB
aits
:2,2
,2+
2,n
obe
etle
s(c
)(3
d)3
Effe
ctof
trap
heig
htfr
omgr
ound
(9)
4bl
ocks
,2×9
dJu
l9–S
ep10
,200
0,S
arav
anB
aits
:2at
0,2,
5m
,2at
0,2,
5m
,no
beet
les
at0,
2,5
m(3
d)S
ynth
etic
com
po
un
ds
(ma
le-s
pe
cific
:1–
6o
ra
na
log
s:7
–8
)4
Test
ofco
mpo
unds1
–3em
itted
bysa
me
disp
ense
rm
odel
(6)
5bl
ocks
,4×9
dJu
n14
–Jul
20,2
001,
Sar
avan
Ba
its:1
,2,3
,1:2
(1:1
v/v)
,1:2
:3(1
:1:1
),2
(c)
(3d)
5Te
stof
com
poun
ds1-8
atsa
me
targ
etdo
seof
5m
g/da
y(9
)4
bloc
ks,2×9
dJu
n20
-Jul
8,20
02,S
arav
anB
aits
:1,2
,3,4
,5,6
,7,8
,2(c
)(3
d)6
Effe
ctof
addi
ngco
mpo
unds2
–6to
com
poun
d1:t
esto
ffive
5bl
ocks
,2×
9d
Jul2
6-A
ug13
,200
2,S
arav
an1–
6m
ixes
(B1–
B5)
atta
rget
dose
of5
mg/
day
vs.o
nly
1(9
)(3
d)B
aits
:B1,
B2,
B3,
B4,
B5,
1(3
and
6m
g/da
yta
rget
dose
s),2
(c)
7D
ose-
resp
onse
toco
mpo
und
1(5
)6
bloc
ks,2×
9d
Jul8
–26,
2002
,Sar
avan
Ba
its:f
our
dose
sof
1,2
(c)
(3d)
8S
yner
gybe
twee
nco
mpo
und1an
dda
tepa
lmco
re-D
P-
(5)
5bl
ocks
,2×9
dA
ug13
–31,
2002
,Sar
avan
Ba
its:1
,DP,
crus
hed
DP,1
+D
P,1+
crus
hed
DP,
2+
DP
(c)
(3d)
9S
easo
nalp
opul
atio
nm
onito
ring
(1)
10tr
aps,
25×7
dA
pr7–
Sep
1,20
02,S
arav
anB
ait:
com
poun
d1(t
arge
tdos
eof
3m
g/d)
(7d)
Sar
avan
No
te.(
c):c
ontr
ol.
aF
orex
ampl
e:A
ssay
3(f
our
com
plet
ebl
ocks
)la
sted
4×
9d=
36d.
The
num
ber
ofvi
sits
(bas
icda
tafo
rA
NO
VA)
was
4×(9
d/3
d)=
12.I
nA
ssay
8,th
e10
trap
s(s
ingl
etr
eatm
ent)
wer
evi
site
dw
eekl
yfo
r25
wk.
P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 395
FIG. 1. Overlaid chromatograms of SupelpakTM -2-trapped volatiles emitted by sugarcane,andOryctes elegansmales or females feeding on sugarcane, showing two profiles of male-associated compounds. A: GC analysis on a polar RTX-Wax column showing male effluviacharacterized by1 and2 (55:45 ratio; 2001 collection) and food volatiles induced by thebeetle feeding (f: white arrow heads); and B: GC–MS analyses using an apolar MDN-5MS column showing compounds1– 6(21:37:34:3:4:1 ratio) associated with males (2000collection).

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
396 ROCHAT ET AL.
FIG. 2. Relative electroantennogram (EAG) responses byOryctes elegansmales and fe-males (mean+S.E.;N = 4 of each sex) to six synthetic molecules (four compounds isolatedfrom the male effluvia:1–4; and two related molecules:7–8) at four doses. The horizontallines within the graphs indicate the mean responses to solvent.
was detected from eight of the nine batches with males.1 and2 were the mostabundant and regular components (respectively, 53± 11 and 37± 10% total male-specific volatiles; mean±S.E.).1was emitted at a daily rate estimated between 0.1and 5µg/male. In six batches,1 was more abundant than2 (80 to 50%) followedby 2. In two cases,2 was the most abundant male-specific molecule (88 and 37%)either mixed with only2 (12%) or3 (35%) and other minor components.3–6 weredetected in less than half the samples.3 was the third most abundant component(8 ± 4%) whereas4–6 appeared as sporadic trace components, accounting for<1% of the male-specific volatiles.
Electroantennography.(Figure 2). EAG responses recorded to the six testedmolecules differed between sexes. In females, most of the responses to the chem-icals were higher than to the solvent. The most stimulatory compound was the
P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 397
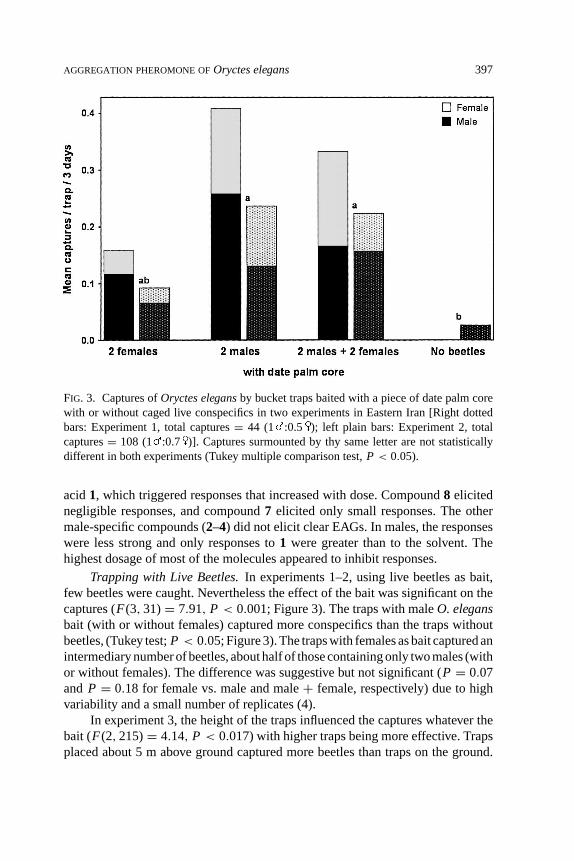
FIG. 3. Captures ofOryctes elegansby bucket traps baited with a piece of date palm corewith or without caged live conspecifics in two experiments in Eastern Iran [Right dottedbars: Experiment 1, total captures= 44 (1 :0.5 ); left plain bars: Experiment 2, totalcaptures= 108 (1 :0.7 )]. Captures surmounted by thy same letter are not statisticallydifferent in both experiments (Tukey multiple comparison test,P < 0.05).
acid1, which triggered responses that increased with dose. Compound8 elicitednegligible responses, and compound7 elicited only small responses. The othermale-specific compounds (2–4) did not elicit clear EAGs. In males, the responseswere less strong and only responses to1 were greater than to the solvent. Thehighest dosage of most of the molecules appeared to inhibit responses.
Trapping with Live Beetles.In experiments 1–2, using live beetles as bait,few beetles were caught. Nevertheless the effect of the bait was significant on thecaptures (F(3, 31)= 7.91, P < 0.001; Figure 3). The traps with maleO. elegansbait (with or without females) captured more conspecifics than the traps withoutbeetles, (Tukey test;P < 0.05; Figure 3). The traps with females as bait captured anintermediary number of beetles, about half of those containing only two males (withor without females). The difference was suggestive but not significant (P = 0.07and P = 0.18 for female vs. male and male+ female, respectively) due to highvariability and a small number of replicates (4).
In experiment 3, the height of the traps influenced the captures whatever thebait (F(2, 215)= 4.14, P < 0.017) with higher traps being more effective. Trapsplaced about 5 m above ground captured more beetles than traps on the ground.

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
398 ROCHAT ET AL.
FIG. 4. Effect of the position of the trap on captures ofOryctes elegansusing a pieceof date palm core with or without caged live conspecifics as bait (Eastern Iran Assay 3).Whatever the bait, captures associated with the same letter are not statistically different.Catches by males and females were statistically equivalent and greater than by no beetles(Tukey multiple comparison test,P < 0.05).
At about 2 m high, traps captured an intermediate number of insects. Attractionto odors of males and females was equivalent and greater than to traps with nobeetles whatever the trap position (Tukey test,P < 0.05; Figure 4).
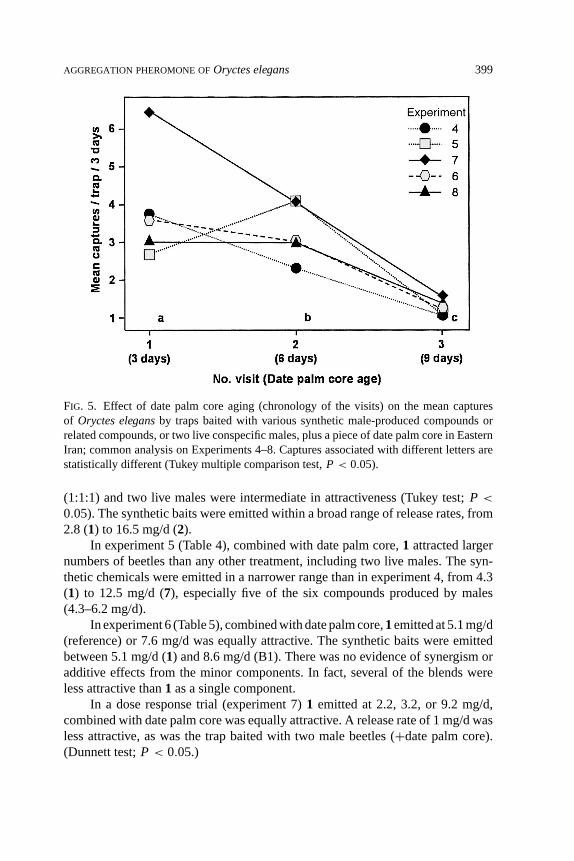
Evaluation of Synthetic Chemicals.Whatever the synthetic bait and the repli-cate, trap catches decreased as the date palm tissue aged (F(2, 1115)= 67.23, P <
0.001; Figure 5) in experiments 4–8. The mean captures were significantly lower ata given visit than at the previous one. They decreased by 20 and 66% respectivelyat the second (day 6) or third (day 9) visit as compared to the first (day 3).
The effect of the replicate (time) was significant in experiments 4, 5, and7, (F(3, 354)= 40.23, F(1, 208)= 43.32, andF(1, 179)= 5.23, respectively;P < 0.001), indicating that the level of catches changed with time whatever thebaits. In experiments 6 and 8, the level of capture was equivalent in the tworeplicates.
In experiment 4 (Table 3), the effect of the bait was significant (F(5, 354)=7.56, P < 0.001). Combined with date palm core, compound1was most attractive,significantly more than compounds2, 3, and the2:3 blend (1:1). The1:2:3 mix
P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 399
FIG. 5. Effect of date palm core aging (chronology of the visits) on the mean capturesof Oryctes elegansby traps baited with various synthetic male-produced compounds orrelated compounds, or two live conspecific males, plus a piece of date palm core in EasternIran; common analysis on Experiments 4–8. Captures associated with different letters arestatistically different (Tukey multiple comparison test,P < 0.05).
(1:1:1) and two live males were intermediate in attractiveness (Tukey test;P <
0.05). The synthetic baits were emitted within a broad range of release rates, from2.8 (1) to 16.5 mg/d (2).
In experiment 5 (Table 4), combined with date palm core,1 attracted largernumbers of beetles than any other treatment, including two live males. The syn-thetic chemicals were emitted in a narrower range than in experiment 4, from 4.3(1) to 12.5 mg/d (7), especially five of the six compounds produced by males(4.3–6.2 mg/d).
In experiment 6 (Table 5), combined with date palm core,1emitted at 5.1 mg/d(reference) or 7.6 mg/d was equally attractive. The synthetic baits were emittedbetween 5.1 mg/d (1) and 8.6 mg/d (B1). There was no evidence of synergism oradditive effects from the minor components. In fact, several of the blends wereless attractive than1 as a single component.
In a dose response trial (experiment 7)1 emitted at 2.2, 3.2, or 9.2 mg/d,combined with date palm core was equally attractive. A release rate of 1 mg/d wasless attractive, as was the trap baited with two male beetles (+date palm core).(Dunnett test;P < 0.05.)

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
400 ROCHAT ET AL.
TABLE 3. COMPARATIVE CAPTURES OFOryctes elegansBY THREESYNTHETIC
COMPOUNDS(1–3: THE MAJORCOMPONENTS OF THEMALE EFFLUVIA), ALONE OR
MIXED, AND EMITTED BY THE SAME TYPE OFDISPENSER, AND TWO LIVE CONSPECIFIC
MALES IN BUCKET TRAPSCONTAINING A PIECE OFDATE PALM CORE(EASTERNIRAN,EXPERIMENT 4)
Mean± S.E. release rateBait added to date palm corea of the chemical bait (mg/day)b Mean± S.E. captures/trap/3 dc
1: 4-Methyloctanoic acid 2.8± 0.4 4.0± 0.6b
2: Ethyl 4-methyloctanoate 16.5± 1.3 1.4± 0.2a
3: 4-Methyloctanyl acetate 13.6± 0.3 2.2± 0.5a
2:3 (1:1) 15.6± 1.0 1.5± 0.3a
1:2:3 (1:1:1) 8.5± 0.4 2.6± 0.5ab
Two males (control) — 2.5± 0.5ab
Notes. Total catches= 844 (1 : 1.4 ). S.E.: standard error of the mean.aValues in brackets indicate the relative proportion of the components within mixes.bActual rates during the experiment (n = 25). Values for each component of the mixes undetermined.cCaptures followed by the same letter are not significantly different (Tukey multiple comparison test,
P < 0.05).
In the 8th experiment (Table 6), captures by all the baits were significantlydifferent from zero. Crushed date palm core tended to attract more beetles, eitheralone (×2.6) or combined with1 (×1.4), than a standard date palm core piece. Asbefore,1 combined with date palm core was more attractive than two live maleson date palm core, alone, or date palm core alone (Dunnett test;P < 0.05.)
TABLE 4. COMPARATIVE CAPTURES OFOryctes elegansBY EIGHT SYNTHETIC
MOLECULES AT THESAME TARGET DOSE OF5 Mg/Day (SIX COMPOUNDSISOLATED
FROM THEMALE EFFLUVIA: 1–6AND TWO RELATED MOLECULES: 7–8),AND TWO LIVE
CONSPECIFICMALES IN BUCKET TRAPSCONTAINING A PIECE OFDATE PALM CORE
(EASTERNIRAN, EXPERIMENT 5)
Mean± S.E. release rateBait added to date palm core of the chemical bait (mg/day)a Mean± S.E. captures/trap/3 db
1: 4-Methyloctanoic acid 4.3± 0.2 9.6± 2.5a
2: Ethyl 4-methyloctanoate 5.5± 0.2 2.3± 1.0b
3: 4-Methyloctanyl acetate 8.2± 0.1 2.7± 0.3b
4: Methyl 4-methyloctanoate 4.8± 0.2 0.9± 0.9b
5: 4-Methyloctanol 6.2± 0.4 2.7± 0.8b
6: Nonanyl acetate 5.4± 0.2 0.5± 0.7b
7: Octanoic acid 12.7± 2.0 1.0± 0.2b
8: Nonanoic acid 9.5± 0.5 1.7± 0.6b
Two males (control) — 1.8± 0.6b
Notes: Total catches= 653 (1 : 0.5 ). S.E.: standard error of the mean.aActual rates during the experiment (6≤ n ≤ 8).bCaptures followed by the same letter are not significantly different (Tukey multiple comparison test,
P < 0.05).

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 401
TABLE 5. COMPARATIVE CAPTURES OFOryctes elegansBY FIVE SYNTHETIC MIXES
(B1-B5) OF THESIX COMPOUNDSISOLATED FROM THEMALE EFFLUVIA 1–6, TWO DOSES
OF 1, THE MAJORCOMPOUND OF THEMALE EFFLUVIA, AND TWO LIVE CONSPECIFIC
MALES IN BUCKET TRAPSCONTAINING A PIECE OFDATE PALM CORE IN EASTERNIRAN
(EXPERIMENT 6)
Mean± S.E. release rate Mean± S.E. captures/Bait added to date palm corea of the chemical bait (mg/day)b trap/3 dc
B1: 1:2:3 (1:1:1) 8.6± 1.2 1.8± 0.4∗B2: 1:2:3 (10:1:1) 5.2± 0.1 2.0± 0.3∗B3: 1:2:3:4:5:6 (100:10:10:1:10:1) 5.3± 0.3 3.3± 0.6B4: 1:2:3:4:5:6 (10:10:10:1:10:1) 7.3± 0.8 3.3± 0.6B5: 1:2:3:4:5:6 (10:10:10:1:1:1) 7.6± 0.8 1.5± 0.3∗1: 4-Methyloctanoic acid 5.1± 0.2 →3.9± 0.61: 4-Methyloctanoic acid 7.6± 0.5 4.5± 0.7Two males (control) — 0.7± 0.2∗
Notes. Total catches= 623 (1 : 1.3 ). S.E.: standard error of the mean.a2: Ethyl 4-methyloctanoate,3: 4-methyloctanyl acetate,4: methyl 4-methyloctanoate,5: 4-
methyloctanol, and6: nonanyl acetate). Values in brackets indicate the relative proportion of thecompounds within the mixes B1–B5.
bActual rates during the experiment (9≤ n ≤ 10). Values for each component of the mixes undeter-mined.
cCaptures asterisked are significantly lower than compound1 at 5.1 mg/d (Dunnett test,P < 0.05).
Population Monitoring over 25 wk(experiment 9: Figure 6). A total of 841beetles (55% female) was captured in 10 traps. Weekly captures per trap, whichwere nil in mid-April, increased regularly until the beginning of June (5.3± 1.4beetles). Then, they oscillated around that level until mid-September with a sharp
TABLE 6. SYNERGISTICCAPTURES OFOryctes elegansBY COMBINED DATE PALM CORE
(CRUSHED ORNOT) AND SYNTHETIC 1, THE MAJORCOMPONENT OF THEMALE
AGGREGATIONPHEROMONE ASCOMPARED TOTWO LIVE MALE CONSPECIFICS ONDATE
PALM CORE IN BUCKET TRAPS(EASTERNIRAN EXPERIMENT 8)
Mean± S.E. release rate Mean± S.E. captures/Bait of the chemical bait (mg/day)a trap/3 db
1: 4-Methyloctanoic acid 3.0± 0.1 0.3± 0.1b
Date palm core — 0.2± 0.1b
Crushed date palm core — 0.6± 0.3b
1+ Date palm core 3.5± 0.1 5.2± 0.7a
1+ Crushed date palm core 3.3± 0.1 7.4± 0.9a
Two males+ date palm core — 1.0± 0.3b
Notes. Total catches= 435 (1 : 1.5 ). S.E.: standard error of the mean.aActual rates during the experiment (n = 8).bCaptures followed by the same letter are not significantly different (Tukey multiple comparison test,
P < 0.05).

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
402 ROCHAT ET AL.
FIG. 6. Weekly captures ofOryctes elegansper trap—bottom, left scale—with correspond-ing sex-ratios—top, right scale—from 10 traps baited with a piece of date palm core and1(4-methyloctanoic acid, the major component of the male aggregation pheromone) emittedat a rate of 2.2± 0.1 mg/d (mean± S.E.) for 25 wk between April and September 2002 inEastern Iran (Experiment 9). Marked points indicate that the represented values are averagesdue to a single visit on July 7 with captures from 2 wk accumulated in traps.
peak of capture at the beginning of August (11.5± 1.9 beetles) and decreasedduring the two final weeks of the trial. Considering the mean captures per trapcalculated on a monthly basis, the population reached a maximum in summer(mid-July–mid-August: 5.9± 0.8 beetles). The sex ratio of the catches fluctuatedthroughout the trial, being approximately balanced in spring and then showing anexcess of females through summer, especially in August.
DISCUSSION
Our data demonstrate that maleO. elegansproduce an aggregation pheromonelike all the dynastid species studied to date. Its active component is 4-methyloctanoic acid, a molecule previously reported from the TropicalOryctes,O. monocerosandO. rhinoceros(Gries et al., 1994; Hallet et al., 1995; Morin etal., 1996). However, besides these common features, theO. eleganspheromonecommunication system differs from the pheromone communication system of theTropical Oryctesin two major aspects: (1)O. elegans’most active pheromonecompound is not active in the TropicalOryctes, and (2)O. eleganspheromone isweakly attractive by itself but highly synergistic with the odor emitted by freshlycut and macerated palm tissue. This is not the case for the aggregation pheromonesof the TropicalOryctes, which are highly attractive without plant chemicals.

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 403
Attraction to Conspecifics in Natural Conditions.Both sexes ofO. elegansattracted conspecifics when placed on date palm core, suggesting the emissionof (a) pheromone(s) and/or plant volatiles induced by beetle feeding. The latterhypothesis is in agreement with the catches by either sex feeding on palm tissuerecorded in the initial assays. GC analyses also showed that feeding of beetles ofeither sex on sugarcane resulted in the emission of several compounds.
Trap captures in traps baited with males were never significantly greater thanin traps baited with females and the initial field assays did not conclusively supportthe emission of a male aggregation pheromone. Evidence for a male-producedpheromone was provided by the analyses of the beetles’ effluvia and subsequentassays of synthetic chemicals. Conversely, the existence of a female aggregationpheromone seems unlikely because no female-specific compounds were detectedfrom the female effluvia. Thus, attraction of beetles to females feeding on datepalm may only reflect a response to palm volatiles induced by beetle feeding.In addition, our first assays were performed in harsh environmental conditions(>50◦C, very low humidity). These conditions were fatal to many bait insects asshown by the high mortality recorded on the weekly visits (experiments 1–2), andmales probably did not emit pheromone during the major part of the assays.
Male Pheromone Composition.The male pheromone secretion ofO. eleganswas more complex and variable than reported in the TropicalOryctes. In additionto the major compounds1 and2 that were reported from them,O. elegansmalesproduced the new compounds3–5, which have obvious structural relationshipwith 1 and2, and with ethyl 3-methylheptanoate, reported fromO. rhinoceros(Hallet et al., 1995). These molecules share a methyl-branched carbon skeleton,suggesting a common biogenetic pathway. The trace amounts of3–6detected fromcertain batches of maleO. eleganscould be artifacts of our sampling proceduredue to reactions between fermentation products from sugarcane with compounds1–2. However, SPME sampling showed that two males placed in silanized glassvials without food could emit 0.1–1µg of 1, 2, and5 simultaneously (Rochat andPezier, unpublished). The noticeable amounts of the acetate3 detected in at leasttwo batches of male effluvia also support production of3 and5 by the insect itself.
O. elegansmale-specific effluvia were variable both in the quantities emittedand in their qualitative composition. This is possibly related to our sampling con-ditions because we used wild beetles, of unknown age and physiological status,at different seasons, and grouped in different numbers with only males or mixedsexes. Pheromone collection from more standardized samples, coupled to an ap-propriate method for GC quantification will be required to determine the biologicalfactors which modulate pheromone emission.
Pheromone Activity. Insects responded to racemic1, indicating that chiralitywas not a critical point in the species or suggesting that it uses a racemic pheromone.Hallet et al. (1995) reported 4S-2 to be as attractive as (±)-2 in O. rhinocerosandconcluded that this species produces either the S enantiomer or a racemic mix. We

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
404 ROCHAT ET AL.
did not investigate the enantiomeric composition of the molecules produced byO.elegans.
A strong response to the acid1 was the constant feature of the comparativetrials performed, in accordance with the prevalence of this molecule in the malepheromone secretion. The specificity of the response was correlated to the methyl-branched carbon skeleton because responses to the acid analogs7 and 8 werenegligible. In the field, 1–10 mg/d release rates of1 were active. Higher doseswere not evaluated because release rates of 2–3 mg/d were as good as the higherdose of 10 mg/d.
All the other male-specific chemicals were less active than1, either as sin-gle compounds or blends (experiment 3), and even tested at higher doses. Theseobservations are in agreement with the strong EAG responses recorded to1, incontrast to those recorded to the other molecules. The presence of large amountsof 2 (ethyl ester) or3 (acetate) with1 (experiments 4–5) seemed to reduce thetrapping efficiency with the exception of the mix1:2:3:4:5:6 (10:10:10:1: 1:1) inexperiment 5. The good activity of this mix may be due to the large amount of5, a compound that was not tested by EAG. However, because no clear effect, ei-ther inhibitory or enhancing, was observed to the blends, and because most minormale-specific chemicals appeared as erratic components in the male effluvia, theresponses to the molecules other than1–2 in the field may be due to their structuralrelationship with1. Alternately, these compounds might be involved in reproduc-tive isolation withO. agamemnon, which is sympatric withO. elegansin lowlandareas of the Middle East; in that case it would not be expected that their presencewould improve the efficiency of the traps.
The strong activity of1 in O. elegansand the weak activity of2 is opposite towhat happens in the TropicalOryctes: 2 proved to be the only or essential activepheromone compound in these species while1 resulted in negligible captures ofO. rhinocerosat 30 mg/d (Hallet et al., 1995).1, inactive when evaluated by GC-EAD of natural male effluvia, was not evaluated in the field withO. monoceros(Gries et al., 1994). InO. rhinoceros, a third molecule, ethyl 3-methylheptanoatewas also reported from males and attracted about four times fewer beetles than2at 30 mg/d (Hallet et al., 1995), indicating the species was moderately responsiveto another molecule produced by males, with a close structural relationship to themajor pheromone compound.
The sex ratio of the beetles captured was not correlated to the pheromonebait but varied during the flight period of the beetle. An equal ratio of males andfemales was captured over the whole set of experiments with some variations,the origin of which is unknown. Both sexes seemed to emerge simultaneously inspring according to the 2002 monitoring. The female excess recorded in summermay reflect a shorter life span of the males or a greater mobility of females.
Plant Co-Attraction.Although1 as a single component and date palm odorwere attractive alone catches dramatically increased when both odors were

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 405
combined. The effect was strong with fresh palm material, becoming weaker as thetissue aged, and the effect tended to be enhanced when the palm tissue was crushed.Whereas this phenomenon is classical in palm weevils (Giblin-Davis et al., 1996;Rochat et al., 2000a), it has not been reported in other dynastid beetles, or, if so,to a much lesser extent. It may be correlated to the ecology ofO. elegans, whichcolonizes live trees for mating and egg laying in the same way as the palm weevils.
In the TropicalOryctes, mating sites are separate from feeding sites. Males arealways found singly in feeding galleries, an observation fully in agreement withthe absence of pheromone emission from galleries (Morin et al., unpublished).Their aggregation pheromone is attractive by itself (Hallet et al. 1995; Morin andAllou, unpublished). An enhanced or synergistic response, not as dramatic as inO. elegans, can be recorded by adding decaying woody material to the pheromoneof O. rhinoceros, an egg laying substrate where the beetles hide and the malesemit their pheromone (Hallet et al., 1995; Alfiler, 1999; Sudharto, 2001).
The dynastS. australispresents an intermediate case betweenO. elegansand the TropicalOryctes. Rochat et al. (2000b) showed that the male aggregationpheromone was quite attractive by itself but also synergistic with odors emitted byfreshly cut coconut or sugarcane tissues. MaleS. australisemit their aggregationpheromone from the feeding galleries burrowed in such fresh tissue, attractingfemales and mating with them, but also attracting other males which they get ridof by fighting (Prior et al., 2000). The egg-laying sites are separate from the feedingsites, as in the TropicalOryctesand contrary toO. elegans. Therefore, a synergybetween the male pheromone and plant odors appears particularly intense when itinvolves fresh tissues where the males are emitting pheromone. The mechanismof the plant–insect interaction, especially in the synergistic response and in thepheromone emission, remains unknown.
Applied Perspectives.More than 4000O. eleganswere caught over thetwo trapping seasons during which the synthetic pheromone was evaluated. Thecaptures averaged 6.3 beetles/trap/wk considering only the traps baited with1 anddate palm, which emerged as the operational bait for further trapping. The 25-wktrapping campaign showed that seasonal monitoring of the pest can be carried outefficiently by this means. The trap catch data agreed with data available in theliterature and obtained from farmers:O. elegansadults appear in spring and can befound until the temperature drops in late fall. The maximum abundance is recordedin summer, based on the individuals observed at lights.
Trapping could be used as a monitoring and decision tool to improve thetiming of insecticide application, which is generally performed when farmers pruneleaves around fruit bunches and place them in dried leaf nets in order to protect themfrom wind and predators. Insecticide is applied as a prophylatic measure, and thereis no simple way to evaluate the population level. Trapping could also be integratedwith insecticide applications to directly control the pest, with a double benefit: amore efficient and safer elimination of the beetles, which can be done using soapy

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
406 ROCHAT ET AL.
water without insecticide in traps as reported forS. australis(Rochat et al., 2002).This method is particularly attractive considering the low economic level of thefarmer, the human health risks from insecticides, and the fragile environment ofthe oases.
Besides the cost of the pheromone, one critical point of this trapping strategyremains, i.e., the necessity to use fresh date palm tissue to synergize the pheromone.Date palm tissue must be renewed weekly in order to maintain a satisfactory levelof captures. However, such material is precious for farmers because shoots are soldfor the multiplication of the palms. We need to find cheap substitutes for the datepalm shoots, either natural or synthetic, before trapping can be fully developed.
Finally, the development of a common trapping ofO. elegansand the redpalm weevil,Rhynchophorus ferrugineusOliv., now sympatric with the dynast inmany places, is also planned for a better and more economic general control ofdate palm beetle pests.
Acknowledgments—We thank Hooshang Boroomand from the Insect Taxonomy Research De-partment, Coleoptera Laboratory, in Tehran for help and visits to theOryctescollections from theHayk Mirzayans Museum, all the farmers and technicians from Saravan, especially Reza Erfanian andFaridoddin Yousefi without whom the field work could not have been possible, Fr´ederic Marion-Pollfor English revision, and two anonymous reviewers for helpful comments.
REFERENCES
ALFILER, A. R. R. 1999. Increased attraction ofOryctes rhinocerosaggregation pheromone, ethyl4-methyloctanoate, with coconut wood.Coconut Res. Dev.15:131–149.
BEDFORD, G. O. 1980. Biology, ecology, and control of palm Rhinoceros beetles.Annu. Rev. Entomol.25:309–339.
CHUNG, G. F. 1997. The bioefficacy of the aggregation pheromone in mass trapping of rhinocerosbeetles (Oryctes rhinocerosL.) in Malaysia.The Planter (Kuala Lumpur)73:119–127.
ENDRODI, S., 1985. The Dynastinae of the World. Dr. W. Junk Publishers, the Hague, The Netherlands,ISBN 90-6193-138-X, Series Entomologica 28, 800 pp.
ENDRODI, S. and PETROVITZ, R. 1974. Die Arten und Rassen der GattungOryctesIlliger in Iran.Entomol. Phytopathol. Appl. (Iran)36:4–19.
GHARIB, A. 1970.Oryctes elegansPrell. (Coleoptera—Dynastidae).Entomol. Phytopathol. Appl.(Iran)29:10–12.
GIBLIN -DAVIS, R. M., OEHLSCHLAGER, A. C., PEREZ, A., GRIES, G., GRIES, R., WEISSLING, T. J.,CHINCHILA , C. M., PENA, J. E., HALLET, R. H., PIERCE, H. D. JR., and GONZALEZ, L. M., 1996.Chemical and behavioral ecology of palm weevils (Curculionidae: Rhynchophorinae).Fla. En-tomol.79:153–167.
GRIES, G., GRIES, R., PEREZ, A. L., OEHLSCHLAGER, A. C., GONZALEZ, L. M., PIERCE, H. D., JR.,ZEBEYOU, M., and KOUAME, B. 1994. Aggregation pheromone of the African rhinoceros beetles,Oryctes monoceros(Olivier) (Coleoptera: Scarabaeidae).Z. Naturforsch.49c:363–366.
HALLET, R. H., PEREZ, A. L., GRIES, G., GRIES, R., PIERCE, H. D., JR., YUE, J., OEHLSCHLAGER,A. C., GONZALEZ, L. M., and BORDEN, J. H. 1995. Aggregation pheromone of the coconutrhinoceros beetles,Oryctes rhinoceros(L.) (Coleoptera: Scarabaeidae).J. Chem. Ecol.21:1549–1570.

P1: IAZ
Journal of Chemical Ecology [joec] pp1129-joec-478533 February 20, 2004 22:23 Style file version June 28th, 2002
AGGREGATION PHEROMONE OFOryctes elegans 407
HO, C. T. 1996. The integrated management ofOryctes rhinoceros(L.) populations in the zero burningenvironment,in Proceedings of the PORIM International Oil Palm Congress, Kuala Lumpur,Malaysia. pp. 336–368.
HURPIN, B. and FRESNEAU, M. 1969. Contribution `a l’etude deOryctes elegans[Col. Dynastinae].Ann. Soc. Entomol. Fr. (N.S.)5:595–612.
HUSSAIN, A. A., 1974. Date Palms and Dates with Their Pests. University of Baghdad, Baghdad, Iraq,pp. 93–95.
JULIA , J.-F. and MARIAU, D. 1976. Research onOryctes monocerosOl. in Ivory Coast. III. Olfactorytrapping with ethyl chrysanthemate.Oleagineux31:263–272.
LEPESME, P. 1947. Les insectes des palmiers. Paul Lechevalier, Paris, pp. 447–473.MINITAB , 1998. Statistical software, version 12.2 in French, Minitab Inc., France.MORIN, J.-P., ROCHAT, D., MALOSSE, C., LETTERE, M., DESMIER DE CHENON, R., WIBWO, H.,
and DESCOINS, C. 1996. Ethyl-4-methyloctanoate, major component ofOryctes rhinoceros(L.)(Coleoptera, Dynastidae) pheromone.C.R. Acad. Sci. Paris, Life Sci.319:595–602
PRIOR, R., MORIN, J.-P., ROCHAT, D., BEAUDOIN-OLLIVIER, L., STATHERS, T., KAKUL , K., EMBUPA,S., and NANGUAI, R. 2000. New aspects of the biology of the Melanesian rhinoceros beetleScapanes australis(Col., Dynastidae) and evidence for field attraction to males.J. Appl. Entomol.124:41–50.
PURBA, R., PRAWIROSUKARTO, S., DESMIER DE CHENON, R., MORIN, J.-P., and ROCHAT, D. 2000.Effect of Oryctes rhinocerospheromone (ethyl 4-methyloctanoate) diffusion rate on the size ofpest catches,in Proceedings of the XXIth International Congress of Entomology. Foz do Iguaçu,Brasil, August 20–26, 2000, Vol. I, p. 181.
ROCHAT, D., MORIN, J.-P., KAKUL , T., BEAUDOIN-OLLIVIER, L., PRIOR, R., RENOU, M., MALOSSE, I.,STATHERS, T., EMBUPA, S., and LAUP, S. 2002. Activity of male pheromone of the Melanesianrhinoceros beetleScapanes australis. J. Chem. Ecol.28:479–500.
ROCHAT, D., NAGNAN-LE MEILLOUR, P., ESTEBAN-DURAN, J. R., MALOSSE, C., PERTHUIS, B., MORIN,J.-P., and DESCOINS, C. 2000a. Identification of pheromone synergists in American palm weevil,Rhynchophorus palmarum, and attraction of relatedDynamis borassi(Coleoptera, Curculionidae).J. Chem. Ecol.26:155–187.
ROCHAT, D., RAMIREZ-LUCAS, P., MALOSSE, C., ALDANA , R., KAKUL , T., and MORIN, J.-P. 2000b. Roleof solid-phase microextraction in identification of highly volatile pheromones of two rhinocerosbeetlesScapanes australisandStrategus aloeus(Coleoptera, Scarabaeidae, Dynastinae).J. Chrom.A 885:433–444.
SUDHARTO, P. S., PURBA, R. Y., ROCHAT, D., and MORIN, J.-P. 2001. Synergy between empty oil palmfruit bunch and synthetic aggregation pheromone (ethyl 4-methyloctanoate) for mass trapping ofOryctesrhinoceros beetles in the oil palm plantation in Indonesia,in Proceedings of the PORIMInternational Oil Palm Congress, Kuala Lumpur, Malaysia, pp. 661–664.
VANDER MEER, R. K. and MCGOVERN, T. P. 1983. Structure-activity correlations for derivatives ofsiglure: Attractants forOryctes rhinocerosL. (Coleoptera: Scarabaeidae).J. Econ. Entomol.76:723–727.
VANDER MEER, R. K., GHATAK , U. R., ALAM , S. K., and CHAKRABORTI, P. C. 1979. (Plus or minus)-Des-N-morphinan: A unique bridged hydrocarbon attractant for the rhinoceros beetle,Oryctesrhinoceros, and development of an olfactometer.Environ. Entomol.8:16–10.