fruit size in a tropical tree species: variation ... · pdf filefruit size in a tropical tree...
TRANSCRIPT

163Vegetatio 107/108: 163-174, 1993.T. H. Fleming and A. Estrada (ed!"). Fnlgivory and Seed Disper.~al: Ecological and Evolutionary.' A.~pects.@ 1993 Kluwer Academic Publishers. Printed in Belgnml.
Fruit size in a tropical tree species: variation, preference .by birds, and
heritability
Nathaniel T. WheelwrightDepartment of Biology, Bowdoin College, Brnnswick, ME 04011, USA
Keywords: Ocotea tenera, Lauraceae, Seed dispersal, Frugivory, Plant reproduction
Abstract
Ocotea tenera (Lauraceae), an understory bird-dispersed tree, produces single-seeded fruits that vary indiameter from 1.4 to 2.4 cm. Much of the variation within a population at Monteverde, Costa Ricaoccurred within individual trees. The relative size of fruits produced by diffcrcnt trees remained gcncr-ally constant over an II-year period despite slight differences between years in the average size of fruitsproduced by a given tree.
Fruit-eating birds could thus express their preferences for particular fruit size characteristics bychoosing among trees that have distinct distributions of fruit diameters, and between individually var-iable fruits within trees. In a field study of individually marked fruits, birds removed 46.2 % of fruits:the rest of the fruits were destroyed by invertebrate (25.3 %) and vertebrate (4.3 %) pulp-feeders oraborted by the plant after remaining ripe but uneaten for as long as 100 days (24.2 %). The four majoravian seed dispersers of O. tenera each have gape widths exceeding all but the largest fruits. Birdspreferred plants with greater-than-average-sized fruits; within trees, they favored larger fruits, apparentlybecause net pulp mass increases with fruit diameter. Fruits that ripened early in the season were morelikely to be removed and were removed more quickly.than late-ripening fruits.
Based on mother-offspring regressions of mean fruit size, tile phenotypic variation in fruit diameterin o. tenera is highly heritable, indicating the potential for an evolutionary response to selection by birds.Nonetheless, directional selection on fruit size or shape is likely to be inconsistent, constrained by gc-netic correlations, and weak compared to selection on traits like fecundity or phenology.
Introduction
When flocks of emerald toucanets (Aulacor-hynchus prasinus) forage through the lower mon-tane forests of Costa Rica, they encounter a di-versity of fruits. On a given day, the birds mightstop to feed bn the clustered orange fruits of Ureraelata, which measure only 3 mm in diameter, thenmove on to eat a few 22-mm diameter blackdrupes of Beilschmeidia pendula before swallow-ing 6-mm Sapium oligoneuron seeds covered by
red arils (Wheelwright et al. 1984). In a temperate-zolle forest the variety of fruits scell by birds isless than in the tropics, but nolletheless birds suchas American robins (Turdus migratorius) consumefruits as distinct as the3-mm furry scarlet drupesof Rhus copa/lina and the lO-mm watery fruits ofPyrus sp. (Wheelwright 1986a; White & Stiles
1991).Fruit variation within species, though less ex-
treme than variation between species, can also bestriking. Individual plants of the same soecies

164
preferences, specifically the role of plant featuressuch as fruit size, coloration, structure and f~cun-dity, as well as extr.insic, non-heritable featuressuch as habitat or the characteristics of neighbor-ing plants (Howe 1983; Gautier-Hion et 01. 1985;Wheelwright & Janson 1985; Denslow 1987; Jor-daDo 1987a; Debussche & Isenmann 1989; Sar-gent 1990). We know far less about where birdsdrop seeds and how seed dispersal patterns in-fluence seedling survival and establishment (Es-trada & Fleming 1986, Murray 1988), much lessplant fitness or the interaction between prefer-ences and patterns of dispersal. With respect lothe genetic basis of variation in fruit traits, weknow next lo nothing about non-commcrcial,
long-lived plant species.Nonetheless, there is abundant (albeit indirect)
evidence that plant reproductive traits have re-spondcd lo natural selection by birds and olllcranimals. In this paper I concentrate on one suchtrait, fruit size, in an attempt to illustrate pheno-typic variation in natural populations at differentscales and show how seed dispersers potentiallyinfluence the evolution of fruit traits. The juslifi-cation for focusing on fruit size is that, of thevariety of traits that have been investigated, fruitsize seems to affect handling ,md profitability. di-rectly and to bear a relatively predictable relation-ship lo the reeding behavior ,md morpllology ofbirds. Birds inn,llure ,md in c,tplivily v,try in how-they select or handle fruits of dilferenl sizes. Thereasons are various, but they include the physicaldifficulty of picking or swallowing large fruits be-cause of gape limitations (Pralt & Stiles 1985;Wheelwright 1985a; Jordano 1987b; L,unbert1989), differences in profitability between (,trgeand small rruits due lo dift"crcnccs in pulp:secdr,tlios (1Icrrcr,t Il)R I; Ilowc & Vande Kcrl:khovc1981; Howe 1983; Jordano 1987; Sno\y & Snow1988) or handling rates (While & Stiles 1991),and digestive problems associ,lted with large ver-sus small seeds (Levey 1987; Murray et £11. 1992).Most of the studies cited above examined birds'fruit size preferences between plant species, but ,lfew have shown that birds respond to fruit sizevariation between populations (Herrera 1981) orindividual plants (Howe 1983; Wheelwright
often produce fruits that differ from those of othermembers of the population (Gorchov 1985). Evenwithin a plant, fruits may vary. Fruit-eating birdsmust therefore make a series of choices at differ-ent scales: between plant species, between indi-vidual plants of a single species, and betweenindividual fruits within the canopy of a particulartree.
Most recent research on fruit-eating birds andfruiting plants has tried to make sense of thesechoices by describing variation in plant reproduc-tive traits and evaluating how different traits af-fect fruit selection and handling by birds (Willson1986; Jordano 1992). The problems of fruit vari-ation and avian preferences have implications forunderstanding the diversity of diets among fruit-eating birds, particularly dietary specializationversus generalization, tiS 'fell tiS the degree ofmutual dependence between birds t1l1d plants, andthe strength of their interaction. Birds' choicestranslate into different rates of fruit removal fromplants and influence seed dispersal patterns.Therefore, birds may affect plant fitness and di-rect the evolution of certain plant traits, but onlywhen the following conditions are met. Theremust be variation in a particular reproductive traitwithin plant populations, birds must pick or han-dle fruits non-randomly with respect to that trait,their preferences must influence the probability ofsuccessful seed dispersal and seedling establish-ment, and at least some of the variation in theplant reproductive trait must be heritable. Oncewe determine to what extent these conditionsapply in natural populations, we will htive a IJ1uchbetter understanding of the diversity of plant re-productive features and the degree to which theyhave been influenced evolutionarily by interac-tiol1s with sced dispersers.
Of these conditions, recent work has clearlydocumented variation in fruit traits at various lev-els -between species, between populations, be-tween individuals, and within individuals (Her-rera 1981; Howe & Vande Kerckhove 1981;Janson 1983; Wheelwright et a/. 1984; Gorchov1985; Johnson eta/. 1985; Pratt & Stiles 1985;Dowsett-Lemaire 1988; Foster 1990). We havealso learned a great deal about the bases of birds'

measure correlations between fruit size and othertraits of relevance to foraging birds, such as seedcharacteristics or crop size; 3) evaluate the con-stancy of the trait in different seasons andthroughout the life of individual plants; 4) deter-mine if fruits of distinct size differ in their prof-itability to potential seed dispersers; 5) show thatbirds can distinguish and actually choose amongfruits on the basis of size at the level of species,individual plants, or individual fruits within acanopy; 6) demonstrate that birds' preferencesresult in differential survival and recruitment ofthe seeds of fruits of distinct sizes; and 7) deter-mine how much of the variation is heritable ver-sus induced by environmental factors such as cli-mate or microhabitat. In other words, we need toask whether fruit size really matters to birds, andif so, whether their choices influence plant litness.
This paper addresses some of these questionsby describing the nature of fruit v,lriatlon inOcotea tel/era, a tree species which I have stud-ied in Costa Rica since 1979. Diameter is used asthe measure of fruit size most meaningful to avianseed dispersers (Pratt & Stiles 1985; Wheelwright1985a; Lambert 1989). I describe variation at twolevels: within a natural population and within in-dividual plants. 1 also present the results of astudy of fruit removal as a function of mean fruitsize within plants and individual fruit size. Fi-nally, I provide estimates of the heritability (h1) offruit diameter based on parent-offspring regres-sIons.
1985a; Piper 1986; see also Boriaccorsco 1979).In several temperate zone studies the relationshipbetween a bird's gape or body size and the num-ber or size of fruits that it ate was weak or non-existent (Johnson et al. 1985; Malmborg & Will-son 1988, but see J ordano 1987b), presumablybecause temperate zone fruits tend to be smallrelative to birds' gapes, and because the range offruit sizes is limited compared to tropical habitats(Wheelwright 1988a). In general, though, moststudies agree that fruit size alone is one of thesures.t predictors of whether or not a bird willconsume a fruit and disperse its seeds (Jordano
1992).Within habitats, the distribution of gape widths
of fruit-eating birds often mirrors the size distri-bution of the fruits on which they depend forfood, further evidence that frui~ size is an impor-tant reature of plants, one that could direct co-evolution between species and determine commu-nity-wide patterns (Wheelwright 1985a; Pratt &Stiles 1985; Dowsett-Lemaire 1988).. Moreover,the allometry of fruit forn1 -specifically, the ten-dency for larger fruits to be more elongated thansmaller fruits (Mazer & Wheelwright, 1993) -
indicates that the evolution of fruit shape. may beconstrained by the ability of potential seed dis-persers to swallow fruits.
The common interpretation of such patterns ofsize and shape in the fruits of bird-dispersedplants is that they reflect mutual adaptation withcontemporary seed dispersers. Such patternscould be influenced by selection at the level ofspecies as well as genotype. For example, paral-lel distributions of fruit sizes and gape widths orfruit-eating birds, as well as upper limits on fruitsize, could be the result of a process of speciesselection producing broad patterns. Contempo-rary fruit forms may have arisen in response tohistorical dispersers (J anzen & Martin 1982;Wheelwright 1988b). Furthermore, fruit size andshape conceivably could have evolved in responseto selection on genetically correlated characterssuch as flower or leaf traits (Primack 1987).
To understand the evolution of traits such asfruit size, it is impol"tant to 1) quantify phenotypicvariability among and within individual plants; 2)
Methods
S/l/l(I' .\'pecie.\'
Ocotea tel/era (Lauraceae) is a small, dioecious,bird-dispersed tree which reaches a m~lximumheight of 12 m and diameter-at-breast-height(DB H) of 20 cm. O. tenera occurs in clumpeddistributions along forest edges and in small lightgaps, although once established, saplings can sur-vive for many years in dense shade in the under-story (Fig. 1). Endemic to Costa Rica, the spe-cies' range extends from an elevation of 1700 m

166
c
.1,:
4
I I
100m'.~
:::".. ..
~~~~lJJ'~~:1
::;,~:,,~
:~::;:~~::~¥~:;;'~
,
';~lff§f~~1Itt T;
t) --,..~~... ..,- ,.
.~. o~~'1~~i~~~ :~:.:'-Plot H ;: :..
r. .' ':;'"' ::
!}:)::~-
~{;~.';'
/(
>
,
~°f:\
/, .::c' t;.f}~~~;;~;~ .
...
-,.
"'c..
'-~;:l~t~~:!..:;;:;i'...1,'-, ,:..,~~.~;11
-.c":.,~~:;;;;
'.,;A~~\
",.:'::":~j';
Monteverde, Costa Rica
~
Fig. I. Map or the study site. Stars indicate the k>cations or all reproductively mature Ocotea tel/era trees kllown in 1980. Darklines indicate dirt roads or major trails; dashed lilIes indicatc minor trails. Opcn arcus designatc pasturcs. wId stipplcd arcus.rorests. Plot T and Plot H wcre establishcd in 1981 and 1984. rcspectively; in each plot. 32 trccs or known matcrnity survivcdto reproductive maturity.
nators are small, generalist wasps, bees, and flies.The fruits, borne in shallow receptacles at the endof enlarged, scarlet pedice.ls, become full-sizeabout 6 months after pollination. As they ripen,they change in color from green to black over a2-3 day period. The principal dispersers of O.tenera are resplendent quetzals (PllaromachnlSmocilmo), three-wattled bellbirds (Procnia.r tri-carnl1cltlata), emerald toucanets, keel-billed tou-cans (Ramphastos slllfuratus) and black guans
\~"~
in the central highlands to as low as 50 m on theCaribbean slope; it is commonest between 1200and 1350 m (Burger & van der Werff 1990). Inearlier publications (Wheelwright et al. 1984,Wheelwright 1986b) the species was referred to asO. bernouliana (sic), now considered by Burger &van der Werff (1990) to be a distinct species.
O. tenera begins to flower at a median age of5 years, height of 1.5 m, and DBH of 2-3 cm(Wheelwright & Bruneau, 1992). Its major polli-
~
" :\\;,:~.~:'::::'i;i:.

167
(Chamaepetes unicofor) (Wheelwright et af. 1984).Birds swallow the fruits whole and regurgitate thelarge single seeds after about 50 min (Wheelwright1991 ).
Study site and procedures
In 1980, as part of a study of the phenology of theLauraceae (Wheelwright 1986b), I marked everyreproductively mature individual of O. tenera en-countered in 15 km2 of lower montane wet andrain forests around Monteverde, Costa Rica (10018 N, 840 48' W; see Lawton and Dryer (1980)& Wheelwright (1985b) for a more complete de-scription of the site) (Fig. 1). Thereafter, all treeshave been measured and their fruits counted in-dividually at least once a year dflring the peak offruit production in mid-dry season (mid-Februaryto early March). For most of the female trees,haphazard samples of ripe fruits, sometimes com-prising all that were produced by the tree, werecollected in 1981, and their dimensions (lengthand maximum diameter of fruits and seeds) weremeasured to 0.1 mm using dial calipers; mass wasdetermined to 0.1 g using portable balances. Atleast 10 fruits were measured per plant, except forthose with crop sizes less than 10 fruits. I mea-sured the fruits of several plants in at least 5different years spanning an II-year period.
To determine the factors influencing fruit re-moval by birds, I monitored the disapp~arance ofripe fruits from 29 trees in 1980-1. For trees withcrop sizes less than the median for the population(30 fruits), I observed all fruits; for trees withlarger crop sizes, I followed a subset of accessi-ble fruits (those borne at heights lower than 3 m).Fruits were identified by writing a number in blackpermanent marker on the pedicel. For 524 fruitsthe diameter at the widest point was measuredwith calipers, as was the length of the exposedrruit (from the margin of the receptacle to the tip).From late December, 1980 until late May, 1981,597 fruits were censused at 3-day intervals. Thedates of ripening and removal (or damage causedby animals or desiccation) were determined. Be-cause the stages of ripening were signaled by
gradual color changes, 3-day censuses were fre-quent enough to determine the exact day whel;1fruits became fully ripe.. Removal was obviousbecause of the empty receptacle left on the tree;I checked beneath plants for evidence that fruitshad b.een dropped rather than dispersed. Pulp-feeding animals left characteristic marks of theiractivity (pericarp disks excised or totally nakedseeds left by leaf-cutter ants (AcrO1llj'rme,y spp.),patchy holes in the pericarp excavated by weevils(Curculionidae), and incisor scars produced byarboreal rodents. Virtually all removal of fruitswas by birds, as indicated by direct observationsof feeding behavior by birds (Wheelwright 1991),negligible fruit removal rates during the night, ab-sence of seeds in mammal feces or beneath batroosts, and examination of isolated dispersedseeds showing that the pulp had been cleanly rc-moved.
In 1981 and 1984 I established experimentalpopulations consisting of 185 O. tenera seedlingsgrown from seeds of known maternity, represent-ing 11 maternal families; 64 plants survived until1991. Of those, most proved to be males and afew had not reached reproductive maturity by1991. For 25 female F. plants whose maternalphenotypes were known, fruits were collected andmeasured. The mean fruit diameters of siblingswere then averaged within sibships and regressedagainst maternal mean fruit diameters to estimateheritability (h2). Because pollen donors were un-known, the slope of the single parent-offspringregression equation was multiptied by 2 to calc'u-late h2 (Falconer 1989). Note that this methodpotentially overestimates h2 for several reasons.First, it overlooks possible maternal genetic ef-fects (e.g., cytoplasmic genes; long-leflll ellcctsc.lused by matern.ll provisioning of thc sccd .lrcless likely). Second, it assumes that F I trees grownfrom seeds taken from the same parent tree werehalf-sibs, when their relatedness may actually begreater if they shared the same pollen donor. An-other potential problem is that fruits produced byF I trees may have resulted from mating betweensiblings in the experimental plots. In spite of thesepotential confounding factors, a strong parent-offspring resemblance in fruit traits almost cer-

168
tainly would indicate an appreciable genetic basisto phenotypic variation. Regressions and otherstatistical analyses were performed on Statview(Abacus 1988) on a Macintosh computer. Unlessnoted otherwise, descriptive statistics are pre-sented as x ( :!: 1 SD).
14, N=46
Results
Variation in fruit diameter 1.4 1.6 1.8 2 2.2 2.4 2.6X Fruit Diameter (cm)
Fig. 3. Frequency distribution of tile mean fruit diameters for46 Dcolea leI/era trees.
poor predictor of seed mass/fruit mass (r2 = 0.0 I;P < 0.05). A relatively small increase in diameterwas associated with a relatively large gain in cd-ible pulp. For exatnple, a 6 ~~,;, dilfercnce in rruitdiameter (e.g., 1.8em to 1.9 cm in diameter) meantan 18% increase in pulp mass (2.61 to 3.08 g).
Trees differed substantially in the size of thefruits they produced. Some trees had character-istically small fruits (x = 1.64 cm in diameter)whereas others produced large fruits (x = 2.22 cm;x of tree means = 1.88:!: 0,15 cm; one-wayANOYA comparing trees: P<O.OOI) (Fig. 3).Fruit diameter within trees varied significantlyamong years; the magnitude of year-to-year dif-ferences varied between trees (two-way ANOY A:among years: P = 0.004; among trees: P < 0.00 I;year-by-tree interaction: P = 0.002). Nonetheless,absolute average fruit size difference within treesbetween years were slight (Fig. 4), and a tree's
Within the natural O. tenera population, therewas wide variability in fruit size. The smallestfruits (1.41 cm in diameter) were only 60% thesize of the largest (2.37 cm; x= 1.82:t 0.14 cm,coefficient of variation [CV] = 7. 7~~; N = 674fruits). Although none of the fr~its was too smallto be eaten profitably by the major seed dispers-ers of O. tenera, the bigger fruits were too broadto be swallowed by bellbirds and perhaps touca-nets (Wheelwright 1985a).
Fruit diameter was positively correlated withother traits, including fruit length, fruit mass, seeddiameter, seed length, and seed mass (linear re-gression; ?=0.21, 0.72, 0.77, 0.21, and 0.60,re-spectively; P < 0.001). From the perspective offruit profitability (Herrera 1981), fruit diameterwas positively correlated with pulp mass (fruitmass -seed mass) (r2 = 0.56; N = 415 fruits;
P<O.OOI) (Fig. 2), although it was a relatively
'00'-''"'"cu~Po
;f
i'-'Bu
~is-£~
1-i"i
11- J
!=::~I::="- H-5---4~ ~ 72-A
80 82 84 86 88 90
Year
Fig. 4. Mean fruit diameter in three Ocotea tenera trees whosefruits were measured in at least five years between 1981 and1991. Error bars represent:!: I SE.
1.4 1.5 1.6 1.7 1.8 1.9 2 2.1 2.2 2.3Fruit Diameter (cm)
Fig. 2. Relationship between fruit diameter and pulp mass(fresh fruit mass -seed mass) in OCOlea lenera.
2.2 , i-..
2.1 ;
2.0
J1.9
1.8
1.71
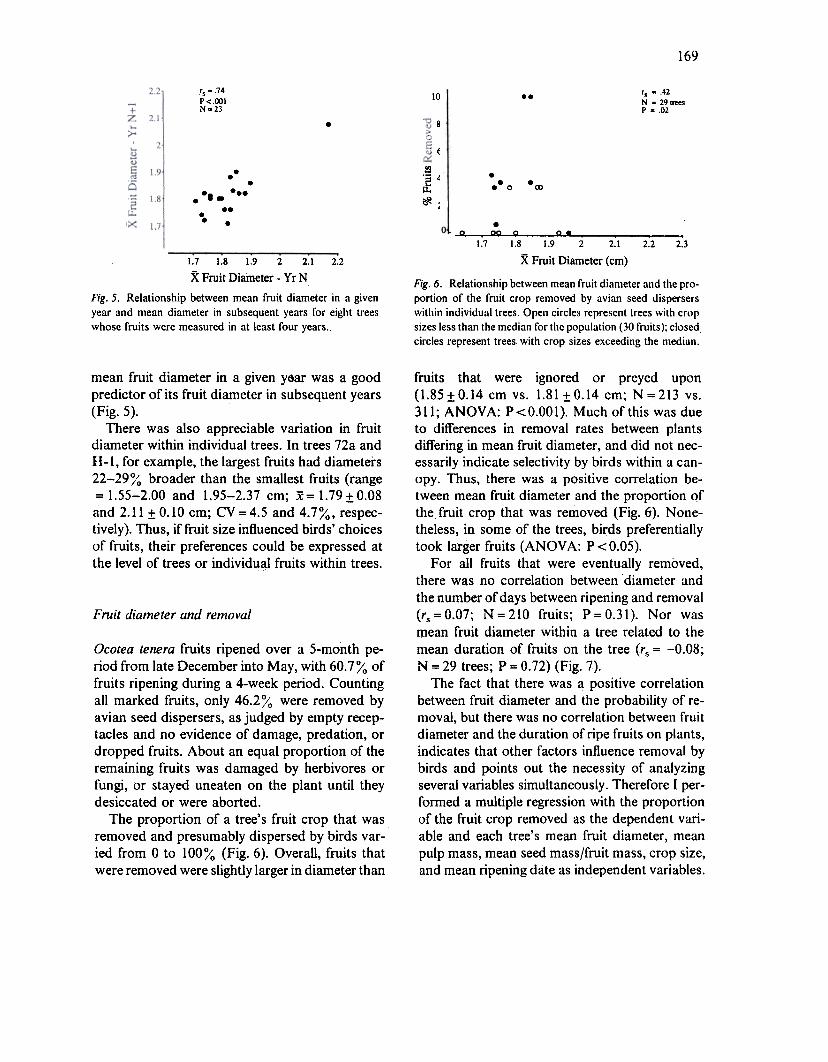
169
rs = .74
P<.OOINg23
's = .42
N z 29 treesP z .02
100,
..
80
.
60
~.-40,
£~ 20,
........ .
.
..0 8(1)
.000008...
1.7 1.8 1.9 2 2.1 2.2 2.3
X Fruit Diameter (cm)
Fig. 6. Relationship between mean fruit diameter and the pro-portion of the fruit crop removed by avian seed disperserswithin individual trees. Open circles represenl trees with cropsizes less than the median for tile population (30 fruits); closedcircles represent trees with crop sizes exceeding the median.
1.7 1.8 1.9 2 2.1 2.2
X Fruit Diameter -Yr N
Fig. 5. Relationship between mean fruit diameter in a givenyear and mean diameter in subsequent years for eight treeswhose fruits were measured in at least four years.
mean fruit diameter in a given year was a goodpredictor of its fruit diameter in subsequent years
(Fig. 5).There was also appreciable variation in fruit
diameter within individual trees. In trees 72a andH-I, for example, the largest fruits had diameters22-29% broader than the smallest fruits (range=1.55-2.00 and 1.95-2.37 cm; x=I.79::t0.08and 2.11 :t 0.10 cm; CV = 4.5 and 4.7%, respec-tively). Thus, if fruit size influenced birds' choicesof fruits, their preferences could be expressed atthe level of trees or individu~1 fruits within trees.
Froit diameter and removal
fruits that were ignored or preyed upon(1.85:1:0.14 cm vs. 1.81 :1:0.14 cm; N =213 vs.311; ANQYA: P<O.OOl). Much of this was dueto differences in removal rates between plantsdiffering in mean fruit diameter, and did not nec-essarily indicate selectivity by birds within a can-opy. Thus, there was a positive correlation be-tween mean fruit diameter and the proportion qfthe fruit crop that was removed (Fig. 6). None-theless, in some of the trees, birds preferentiallytook la:rger fruits (ANOY A: P < 0.05).
For all fruits that were eventually removed,there was no correlation between diameter andthe number of days between ripening and removal(r. = 0.07; N=210 fruits; P=0.31). Nor was
mean fruit diameter within a tree related to themean duration of fruits on the tree (r. = -0.08;N = 29 trees; P = 0.72) (Fig. 7).
The fact that there was a positive correlationbetween fruit diameter and the probability of re-moval, but there was no correlation between fruitdiameter and the duration of ripe fruits on plants,indicates that other factors influence removal bybirds and points out the necessity of analyzingseveral variables simultaneously. Therefore I per-formed a multiple regression with the proportionof the fruit crop removed as the dependent vari-able and each tree's mean fruit diameter, meanpulp mass, mean seed mass/fruit mass, crop size,and mean ripening date as independent variables.
Ocotea tenera fruits ripened over a 5-month pe-riod from late December into May, with 60.7% offruits ripening during a 4-week period. Countingall marked fruits, only 46.210 were removed byavian seed dispersers, as judged by empty recep-tacles and no evidence of damage, predation, ordropped fruits. About an equal proportion of theremaining fruits was damaged by herbivores orfungi, Dr stayed uneaten on the plant until theydesiccated or were aborted.
The proportion of a tree's fruit crop that wasremoved and presumably dispersed by birds var-ied from 0 to 100% (Fig. 6). Overall, fruits thatwere removed were slightly larger in diameter than
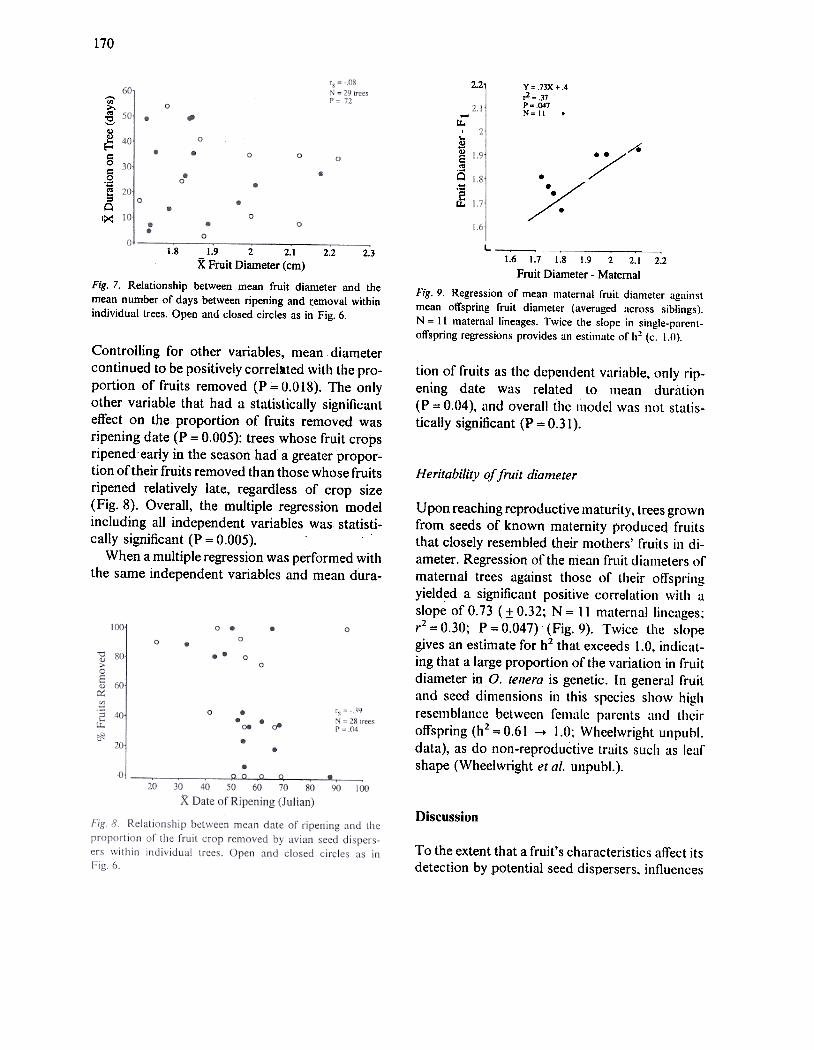
170
2.21 y= .73X".4,2 = .37P =047N= 11 .
""''"...,:S~~c0c
,9
~Q
I~
~,..£e(OJ
Q...
'2~
"'"
;;'".
yL1.8 1.9 2 2.1 2.2 2.3
.it Fruit Diameter (cm)
Fig. 7. Relationship between mean fruit dianleter and themean number of days between ripening and r:emoval withinindividual trees. Open and closed circles as in Fig. 6.
1.6 1.1 1.8 1.9 2 2.1 2.2
Fruit Diameter -Maternal
Fig. 9. Regression of mean maternal fruit diameter againslmean offspring fruit diameter (averaged across siblings).N = II maternal lineages. Twice the slope in single-parcnt-
offspring regressions provides an estimatc of h~ (c. 1.0).
tion of fruits as the dependent variable. only rip-ening date was related to mean duration(P = 0.04), and overall the model was not st~ltis-tic ally significant (P = 0.31).
Heritability of fruit diameter
Upon reaching reproductive maturity, trees grownfrom seeds of known maternity produced fruitsthat closely resembled their mothers' fruits in di-ameter. Regression of the mean fruit diameters ofmaternal trees against those of their offspringyielded a significal1t positive correlation with aslope of 0.73 (:!: 0.32; N = 11 maternal lineages;r2 = 0.30; P = 0.047) (Fig. 9). Twice the slope
gives an estimate for h2 that exceeds 1.0, indicat-ing that a large proportion of the variation in fruitdiameter in o. tenera is genetic. In general fruitand seed dimensions in this species show highresemblance between female ptlrcl1ts tl11d lilciroffspring (h2 = 0.61 -.1.0; Wheelwright unpuhl.
data), as do non-reproductive traits such as letlfshape (Wheelwright et at. unpubl.).
Controlling for other variables, mean diametercontinued to be positively correlated with the pro-portion of fruits removed (P = 0.018). The onlyother variable that had a statistically significanteffect on the proportion of fruits removed wasripening date (P = 0.005): trees whose fruit cropsripened early in the season had a greater propor-tion of their fruits removed than those whose fruitsripened relatively late, regardless of crop size(Fig. 8). Overall, the multiple regression modelincluding all independent variables was statisti-cally significant (P = 0.005).
When a multiple regression was performed withthe same independent variables and mean dura-
Discussion
To the extent that a fruit's characteristics affect itsdetection by potential seed dispersers, influences

71
its probability of being eaten or the time betweenripening and removal, or determines how it ishandled externally or processed internally, suchtraits may be the target of natural selection. Thcimplications of such a conclusion are that theday-to-day interactions between fruit-eating ani-mals and fruiting plants that biologists commonlydocument can have evolutionary significance.Moreover, much of the richness of reproductivefeatures among animal-dispersed plants may le-gitimately be considered 'adaptations,' shaped bygenerations of interactions with animals acting asseed vectors. This requires, of course, that fruittraits have a genetic basis. Research in the fieldof seed dispersal and plant-animal interactionshas explicitly or implicitly assumed that fruitcharacters influence the probability of dispersal,and that much of the phenotypic ~riation in thosecharacters is heritable, but the longevity of mosttrees and the brevity of most studies have pre-vented a full assessment of such assumptions.
This study demonstrates that in at least onetree dispersed by birds, a fundamental fruit char-acter, diameter, shows wide variation betweenand within individuals, in contrast to earlier stud-ies which emphasized the constancy of seed (andby implica.tion, fruit) size within species (Harperet at. 1970). Other species in the Lauraceae areequally variable in fruit morphology (Mazer &Wheelwright, 1993). Faced with a hierarchy ofchoices between trees that have distinct fruittraits, and within trees that produce fruits of vary-ing sizes, avian seed dispersers appear to be dis-criminating foragers. Birds favored fruits oflarger-than-average diameter, presumably be-cause such fruits offered more pulp per fruit (al-though not necessarily a more profitable pulp-:seed ratio) (Herrera 1981; Howe and VandeKerckhove 1981; Piper 1986; Levey 1987; Mur-ray et af. 1992). Birds' fruit-size preferences meanthat plants with less-favored fruits disperse few orno seeds, and that, within canopies, some fruitsare ignored and exposed for longer periods topulp- or seed-damaging herbivores. If birds' pref-erences are relatively stable, they should have acompounding effect on a plant's seed dispersalsuccess year after year, because mean fruit diam-
eter changes little throughout the course of a tree'slife, at least in O. tenera. ,
Mean fruit diameter also changes little betweengenerations within a lineage; in other words, it ishighly heritable in O. tenera. Heritability expressesthe percentage of phenotypic variation of a traitthat is attributable to genotype (the additive ge-netic variance). Although my estimate of h2 maybe biased upward because of maternal effects andthe assumption that F I trees were only half-:sibs(see Methods), the results clearly show a stronggenetic basis to fruit morphology. The significanceof demonstrating that fruit diameter in o. tenerais heritable is that relatively small selective pres-sures can quickly drive evolutionary change insuch traits. Selection on traits that are geneticallycorrelated with fruit diameter could have effectssimilar to direct selection on fruit diameter (Pri-mack 1987). Traits that are most closely relatedto fitness, as fruit diameter might be expected tobe, are ordinarily expected to have relatively lowh2 because most of the additive genetic variancehas already been fixed by natural selection (Fal-coner 1989), but sufficient variability may be in-troduced by mutation or maintained by fluctuat-ing selection to result in high heritabilities of traitssuch as fruit diameter.
If Ocotea tenera is representative of other spe-cies in the Lauraceae, which have fruits of simi-lar size and Corm, share the saine avian seed dis-persers, and are an important component orneotropical forests (Wheelwright et al. 1984),there may be the potential for evolutionary re-sponses on the part of those plants to birds' fruit-size preferences. Whether other plant taxa showequivalent variation in reproductive traits, inter-act with their seed dispersers in comparable ways,and exhibit high heritabilities in such traits is stillunknown, but it is not unreasonable to expectqualitatively similar patterns (cf. Mazer & Wheel-wright 1993).
The results of this study should not be con-strued to imply that directional selection on fruitsize or any other reproductive character is likelyto be strong or constant. Depending upon whetherone views the amount of variation in fruit sizefound in this study as wide or narrow, the high

72
heritability of fruit traits could be interpreted asreflecting stabilizing selection (Gorchov, pers.comm.). In any case, the maintenance of exten-sive phenotypic variation in fruit size within theO. tenera population appears to be inconsistentwith strong directional selection, and it may beexplained by a variety of factors. First, this studyhas not answered a crucial question: does speedyfruit removal in fact result in measurably im-proved plant fitness? If it does not, directionalselection on fruit diameter would be weak, despiteapparent fruit size preferences by birds. Second,there is clearly ample phenotypic variation in fruitform in O. tel1era, and regressions of maternaltraits against that of their daughters establishedthat the variation is heritable, but to what extentis variation within fruit crops caused by differ-ences in pollen donors, and what is the geneticbasis of those effects? A recent study using mo-lecular techniques to examine population geneticstructure and investigate paternity and maternityin seedlings and saplings in O. tel1era may be ableto fill in many important details about the rela-tionship between fruit removal by birds and plantfitness, and shed light on the paternal effects onfruit form (Gibson and Wheelwright, unpubl.).Third, even though this study shows that fruitdiameter hardly fluctuates between years, andother studies suggest that interactions with seeddispersers, or at least assemblages of disperscrs,remain quite constant in the face of wide annualvariation (Howe 1983), other parameters changeextensively, such as a plant's fecundity (Wheel-wright 1986b, unpubl.) and, perhaps more impor-tantly, the fecundity of its neighbors (Howe 1983;Sargent 1990). Fourth, for many reasons plantsmay be constrained in their evolutionary responseto selection by avian seed dispersers. Amongthese are fluctuating selection in space and time,opposing selection, asymmetries in generationlengths of animals and the plants whose seedsthey disperse, and genetically correlated charac-ters (Wheelwright & Orians 1982; Herrera] 985;Wheelwright 1988b; Mazer & Wheelwright 1993).Slight preferences on the part of birds for a traitsuch as fruit size, for example, may be overshad-owed by their response to differences in fecundity
between plants (Howe 1983; Murray 1987; Jor-dano 1987a). ,
General ratller th~ specific adaptation ofplants to their seed dispersers is the more likelyoutcome of evolutionary interactions, and mayexplain both diversity and convergence in fruittraits within habitats and between taxonomicallydistinct plant communities. The general premisesof models of coevolution between mutualists look111 ore credible in light or this study because, in atleast one neotropical tree species, reproductivetraits are highly variable, they matter to birds, andthey have a genetic basis.
Acknowledgements
I th~tnk Montcvcrdc residents, cspcci~llly D. I.cc,S. Perly, the Hogcs, Holl~lI1dcrs, Stuckcys ~lI1dTrostles, ror permitting me to study on their 1~lI1d;F. Joyce, Jr., S. Sargent, J. Wolre, and W. Zu-chowski for helping to establish and monitor theexperimental plots; D. Gorchov, T. Fleming andA. Estrada for helpful commen~s on an earlierdraft; V. Sork for illuminating discussions aboutmaternal genetic effects; and the Organiz~ltion rorTropical Studies, the Douroucouli Foundation,the National Science Foundation, tl1e Council forInternation~tI Exch~lI1ge or Scholars, ~lI1d Bow-doin College ror support or my rcsc~lrl.:lt.
References
Bonaccorsco, F.J. 1979. Foraging and reproductive ccoll>gyin a Panamanian bat community. Bull. Florida Stale Mus.24: 359-408.
Burgcr. W. & H. van dcr WcriT. 1990. Lauraccac. Fillra Co~-taricen~is. Ficldiana (Botany New Series) 23: 1-121.
Dcbusschc, M. & P. Iscmllann. 1989. I-lcshy Iruit charuclcr~and lhe chqiccs or bird and mammal seed dispcrscrs in amedilerraneat1 region.. Oikos 56: 327-338.
Denslow, J.S. 1987. Fruit removal rates rrom aggregated andisolated bushes or the red elderberry. Salllhllcl/X !'l/hell,f.Can. J. Bot. 65: 1229-1235.
Dowsctt-Lemaire, F. 1988. Fruit choice and sccd disscmina-tion by birds and mammals in the evergreen rorcsts or Up-land Malawi. Revue d'Ecologie (Terre Vie) 43: 251-286.
Eslrada, A. & T.H, Flcming (eds.). 1986. Frugivorcs andSeed Dispersal. W, Junk, Amslerdam.

.
173
Falconer, D.S. 1989. Introduction to quantitative genetics.Longman, Essex.
Fleming, T.H. 1986. Opportunism versus specialization: theevolution of feeding strategies in frugivorous bats. In: Es-trada. A. & T.H. Fleming (eds.), Frugivores and Seed Dis-persal. W. Junk, Amsterdam. Pp. 105-118.
Foster, M.S. 1990. Factors influencing bird foraging prefer-ences among conspecific fruit trees. Condor 92: 844-854.
Gautier-Hiim, A., Duplantier J .M., Quris R., Feer F.. SourdC., Decoux J.P., Dubost G., Emmons L., Erard C. &Hecketsweiler P. 1985. Fruit characters as a basis of fruitchoice and seed dispersal in a tropical forest vertebratecommunity. Oecologia 65: 324-337.
Gorchov, D.L. 1985. Fruit ripening asynchrony is related tovariable seed number in Ame/anchier and Vaccinilllll. Amer.J. Bot. 72: 1939-1943.
Harper. J.L., Lovell P.H. & Moore K.G. 1970. The shapesand sizes of seeds. Ann. Rev. Ecol. S,>:st. I: 327-356.
Hcrrera. C,M. 1981a. Fruit variation and competition fordispersers in natural populations of Smi/a.\: a.\'pera. Oikos36: 51-58.
Hcrrera. C.M. 1981b. Are tropic.1l frJits more rewarding todispersers than temperate ones? Amer. N.ltur. II!\: !\96-907.
Herrera, C.M. 1985. Determinants of plant-animal coevolu-tion: the case of mutualistic dispersal of seeds by verte-brates. Oikos 44: 132-141.
Herrera. C.M. 1988. The fruiting ecology of O.\'}'ri.\' quadripar-tita: individual variation and evolutionary potential. Ecol-ogy 69: 233-249.
Herrera C.M. 1989. Frugivory and seed dispersal by carniv-orous mammals and associated fruit characteristics in un-disturbed mediterranean habitats. Oikos 55: 250-262.
Hoppes, W.G. 1987. Pre- and post-foraging movements offrugivorous birds in an eastern deciduous forest woodland.USA. Oikos 49: 281-290.
Howe, H.F. 1983. Annual variation in a neotropical seed-dispersal system. In: Sutton. S.L.. Whitmore T.C. & Chad-wick A.C. (eds.) Tropical rainforest ecology and n1anage-ment. Blackwell Scientific Publications. London. Pr. 211-227.
Howe. H.F. & Richter W.M. 1982. Effccts of seed size ~'ndseedling size in Vir(J/a .\'urinamensi.\': a within and betweentree analysis. Oecologia 53: 347-351.
Ilowc. I-I.F. & V~lnde Kcrckhovc G.A. 19!\ I. l{emov~11 of wildnutmeg (Vir()/a .furinamen.\'i.\') crops by birds. Ecology 62:1093-1106.
Janson, C.H. 1983. Adaptation of fruit morphology todispersal agents in a neotropical forest. Science 219: 187-189.
Johnson, R.A., Willson M.F., Thompson J.N. & Bertin R.I.1985. Nutritional values of wild fruits and consumption bymigrant frugivorous birds. Ecology 66: 819-827.
Jordano. P. 1987a. Avian fruit removal: effects of fruit vari-ation, crop size, and insect damage. Ecology 68: 1711-1723.
J ordano. P. 1987b. Frugivory, external morphology and di-gestive system in mediterranean sylviid warblers S)1,'ia sp,P.Ibis 129: 175-189. .
Jordano, P. 1989. Pre-dispersal biology or Pi.~lacia 11?1I1i.~('u.~(Anacardiaceae): cumulative effects on seed removal bybirds. Oikos 55: 375-386.
Jordano, P. 1992. Fruits and rrugivory. In: Fenner. M. (cd.).Seeds: the ecology or regeneration in plant communities.CAB International, London. III prl?~.~.
Katusic Malmborg, P. & Willson M.F. 1988. Foraging ecol-ogy or avian rrugivores and some consequences ror seeddispersal in an Illinois woodlot. Condor 90: 173-186.
Lambert, F. 1989. Fig-eating birds in a Malaysian lowlandrain rorest. J. Trop. Ecol. 5: 401-412.
Lee. W.G., Wilson J.B. & Johnson P.N. 1988. Fruit color inrelation to the ecology and habit or Copro.w/a spp. (Rubi-aceae) in Ncw Zcaland. Oikos 53: 325-331.
Levey. D.J. 1987. Sced size and rruit-handling tcchniqucs I»avian rrugivores. American Naturalist 129: 471-485.
Lcvey. D.J. & Mocrmond T.C. 1984. Fruil choice in ncotro-pical birds: thc cncct I» uistancc hctwccn rruils on prcli:r-cncc pattcrns. Ecology 65: 84850.
u,iscllc. B.A. ..~ Blakc J.G. 19l){). Dicts I» unucrst\Jry li1lit-eating birds in Costa Rica: seasonalily and resource abun-dance. Studies Avian Bioi. 13: 91-103.
Mazer. S.J. & Wheelwright N.T. Fruit size and shape: allom-etry at different taxonomic levels in bird-dispersed plants.Evolutionary Ecology. In press.
Murray, K.G. 1987. Selcction ror oplimal rruil crop size inbird-dispersed plants. Amer. Nalur. 129: 19-31.
Murray, K.G. 1988. Avian seed dispersal orthrce neolropicalgap-dependent plants. Ecol. Monogr. 58: 271-298. .
Murray, K.G., Winnett-Murray K., Cromie E.A. & Minor M.1993. Feeding prererences or rrugivorous birds: the inl1u-ence or rruit color. seed packaging. and sugar conccntra-lion. Vcgclalio. III p"I!.~.~.
Obeso, J.R. 1989. Fruit removal and polential seed dispersalin a southern Spanish population or Bl?rbl?ri.~ I'ulgaris (Ber-beridaceae). Acta Oecol. Oecol. Plant 10: 321-328.
Piper. J.K. 1986. Seasonality I» rruit characters and sccdrcmoval by birds. Oikos 46: 3113-310.
Pipcr, J. K. 1986. Effccts \)r habiult and sizc or Iruit display onremoval or Smilt/(.illa .~ll?llala (Liliaccac) rruits. Can. J. Bot.
64: 1050-1054.Prlltt. T.K & Stiles E.W. I')X5. .nlC inl1ucn\:c 1»li1lil size alld
structure on composilion or rrugivore assemblages in NcwGuinea. Biotropica 17: 314-321.
Primack, R. 1987. Relationships among flowers. rruits, andseeds. Ann. Rev. Ecol. Syst. 18: 409-430.
Sargent, S. 1990. Neighborhood effects on rruit removal bybirds: a field experiment with Vihurllum demulum (Capriro-liaceae). Ecology 71: 1289-1298.
Snow, B.K. & Snow D~ W. 1988. Birds and berries. T. & A.D.Poyser, Calton.
van der Pijl, L. 1982. Principles or dispersal in higher plants.Springer-Verlag, Berlin.

.174
Wheelwright, N.T. & Bruneall, A., 1992. Population sex ra-tios and spatial distribution of Oralell lellera (Lauraceae)trees in a tropical forest. .lournal of Ecology 80: 425-432.
Wheelwright, N.T., W.A. Haber, K.G. Murray, and C. Guin-don. 1984. Tropical fruit-eating birds and their food plants:a survey of a Costa Rican lower montane forest. Biotropica16: 173-191.
Wheelwright, N.T. & Orians G.H. 1982. Seed dispcrsal byanimals: contrasts with pollen dispersal, problems of ter-minology, and constraints on coevolution. American Nat-uralist 119: 150-164.
White, D.W. & Stiles E.W. 1990. Co-occurrences of foods instomachs and feces of fruit-eating birds. Condor 92: 291-303.
White, D. W. & Stiles E. W. 1991. Fruit harvesting by Amer-ican Robins: innucncc of fruit size. Wils. Bllli. /11 prex.I'.
Willson, M.F. 1986. Avian frugivory and seed dispersal incastcrn NI)rth Amcric:l. Currcnt Ornithology 3: 223-279.
Worthington, A.H. 1989. Adaptations for avian rrugivI)ry:assimilation cfficicncy and gllt transit timc of MIIIIII('rl,t ri-lel/i/lll,t :\I1d I';P,./I /II('IIIt//ix. Occok!gia SO: 31\ 1-31\9.
Wheelwright, N. T. 1985a. Fruit size, gape width, and the dietsof fruit-eating birds. Ecology 66: 808-818.
Wheelwright, N. T. 1985b. Competition for dispersers, and thetiming of flowering and fruiting in a guild of tropical trees.Oikos 44: 465-477.
Wheelwright, N.T. 1986a. The diet of American Robins: ananalysis of U.S. Biological Survey records. Auk 103: 710-725.
Wheelwright, N.T. 1986b. A seven-year study of individua[variation in fruit production in tropical bird-dispersed treespecies in the family Lauraceae. In: Estrada, A. & FlemingT.H. (eds.) Frugivores and Seed Dispersal. W. Junk, Am-sterdam. Pp. 19-35.
Wheelwright, N.T. 1988a. Fruit-eating birds and fruitingplants in the tropics and temperate zone. Trends in Ecol-ogy and Evolution 3: 270-274.
Wheelwright, N.T. 1988b. Four constraints on coevolutionhctwccn plant!\ and their !\ccd di!\pcr!\cr!\: a tropical ca!\chistory. Proceedings of the XIX International Ornitholog-ical Congrc!\s, pp. 827-845.
Wheelwright, N.T. 1991. How long ~o fruit-cating bird!\ stayin thc plant!\ whcrc they fccd? Biolropica 23: 29-40.