fourth nawqa taxonomy workshop on harmonization of …diatom.acnatsci.org/nawqa/pdfs/fourth nawqa...
TRANSCRIPT

Fourth NAWQA Taxonomy Workshop
on Harmonization of Algal Taxonomy October 2000
Report No. 00-21F
The Patrick Center for Environmental Research
held 16-18 October 2000
at
The Academy of Natural Sciences1900 Benjamin Franklin Parkway
Philadelphia, PA 19103-1195
Prepared byEduardo A. Morales
February 7, 2001

THE ACADEMY OF NATURAL SCIENCES 1 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
INTRODUCTION
The Fourth NAWQA Diatom Taxonomy Workshop was held at the Academy of NaturalSciences of Philadelphia (ANSP) on 16-18 October 2000. Specialists participating in theworkshop were: Dr. R. Jan Stevenson and Kalina Manoylova from Michigan State University;Dr. Rex L. Lowe from Bowling Green State University; Dr. Sophia I. Passy from the U.S.G.SEcological Survey and the New York State Department of Environmental Conservation; WilliamR. Cody, environmental consultant based in Ohio; Dr. Loren L. Bahls, environmental consultantbased in Helena, Montana; Todd A. Clason, environmental consultant based in Seattle,Washington; and from the Patrick Center for Environmental Research’s Phycology Section at theAcademy of Natural Sciences of Philadelphia, Dr. Marina Potapova, Dr. Eduardo A. Morales,Dr. Donald F. Charles, Diane M. Winter, Karin C. Ponader and Frank W. Acker.
The three previous NAWQA Taxonomy Workshops had the overall objectives ofharmonizing taxa names used in the ANSP and University of Louisville/University of Michiganlaboratories, identifying reference images for each taxon, and agreeing on up-to-datenomenclature to use when analyzing NAWQA 1997-start samples (see ANSP, 1999; 2000a,b). The fourth NAWQA Diatom Taxonomy Workshop focused on issues concerning the taxonomyof problematic Navicula and Gomphonema species. Taxa that received most attention during theworkshop occur commonly in NAWQA material and are often difficult to identify during routinelight microscopy. Future workshops will focus in a similar way on other genera.
Dr. Potapova selected taxa in need of further taxonomic investigation based on her reviewof existing NAWQA data. During her preparation of data sets for ecological analyses, she hadmany sample counts in which some taxa seemed to have been either misidentified or lumpedunder a single name. On the other hand, she reviewed many slides containing type material andfound that several taxa listed in NAWQA counts were in fact misidentified. Based on theseobservations, she compiled a list of “problematic” taxa within Navicula and Gomphonema. Inmost of the cases, “problematic” taxa corresponded to groups of species that may be easilyconfused with each other because they have similar diagnostic characteristics. Such problematictaxa were arranged into units referred to as “complexes”:
Complex 1- Navicula tripunctata- N. tripunctata var. schizonemoides- N. recens- N. sp. 5 ANS WRC- N. margalithii- N. erifuga- N. cari- Bill Cody’s N. sp. 5

THE ACADEMY OF NATURAL SCIENCES 2 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
Complex 2- Navicula cryptotenella- N. menisculus- N. radiosa var. tenella- N. cincta var. rostrata
Complex 3- Navicula notha- N. leptostriata- N. heimansioides
Complex 4- Navicula luzonensis- N. biconica- N. subminuscula
Complex 5- Gomphonema pumilum spp.
Complex 6- Gomphonema minutum- G. kobayasii
An electronic version of the above list was sent to non-ANSP workshop participants, and theywere asked to prepare plates with photographs of specimens that they believed were difficult todetermine and fell within one of the complexes listed above. These plates were distributedelectronically among participants so they could raise hypotheses as to the identity of depictedtaxa.
Each participant was also asked to prepare a short presentation on any one of thecomplexes. Presentations were to incorporate the historical background of the taxa covered inthat complex, different views in the literature as to their taxonomic position, affinities withclosely related taxa, features used for identification, and relevant ecological information thatmight be helpful in the characterization of the taxon in question. Each presentation was to last15 minutes and be followed by brief discussions.
During the workshop, laboratory sessions were held following each one or twopresentations and concentrated on examination of type slides from the Diatom Herbarium atANSP and/or permanent slides from NAWQA study units. During laboratory sessions, intensivediscussion and literature search led, in many cases, to the application of correct names toproblematic taxa or to the conclusion that more research was needed in order to be confidentabout the identity of certain species and their varieties. During observation of type slides andlaboratory discussions, Dr. Charles W. Reimer’s participation was often crucial in reaching a

THE ACADEMY OF NATURAL SCIENCES 3 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
decision regarding the application of a name. He also provided references, material, andtranslations from German for many of the taxa covered during the workshop.
At the end of the final day of the workshop, all participants met to discuss the format andcontents of the workshop report, names to be applied to specimens depicted on plates preparedprior to the event, changes to be made to NAWQA counts in light of the evidence generatedduring the discussion sessions, and the possible topics of future diatom taxonomic workshops.
This report includes the outcomes of discussions arranged by complex and plates tosupport decisions made during such discussions. These plates should be used by NAWQAtaxonomists as a reference for future identification of depicted taxa. Plates include imagespresented before the workshop and during presentations, images scanned from bibliographicalreferences, and images taken during and after the workshop. Morphological terminology used inthis report follows Barber and Haworth (1981) and Cox (1996).

THE ACADEMY OF NATURAL SCIENCES 4 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
WORKSHOP OUTCOMES
Complex 1- Navicula tripunctata- N. tripunctata var. schizonemoides- N. recens- N. sp. 5 ANS WRC- N. margalithii- N. erifuga- N. cari
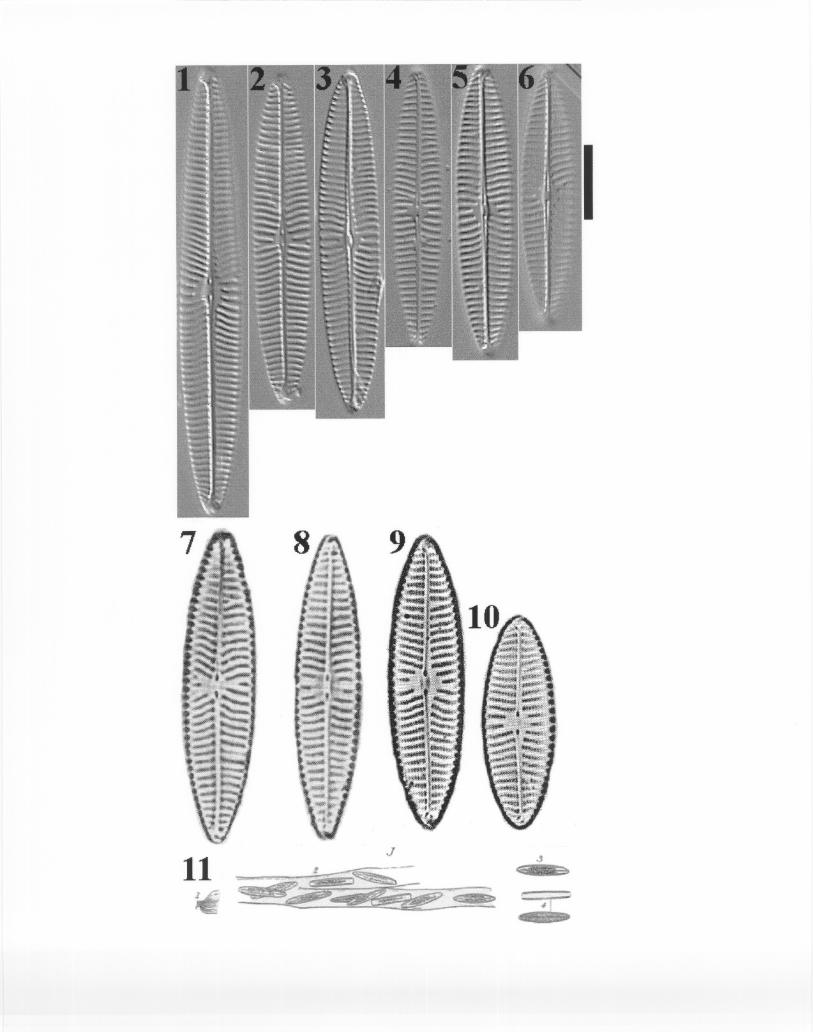
The main reasons for considering these taxa were that, in many instances, each namelisted had been applied to more than one morphological group, and/or a single morphologicalgroup had been identified using more than one of these names. For example, at least twodifferent morphological groups were identified as N. tripunctata var. schizonemoides –hereinreferred to as Groups I and II. Group I (Plate 1, Figs. 1-19) resembles material presented byKrammer and Lange-Bertalot (1986) as N. recens. The latter is a second name used by someNAWQA taxonomists to refer to this Group I. Additionally, a third name –Navicula sp. 5 ANSWRC- was applied to this same group. Group II (Plate 2, Figs. 1-9) also received three differentnames: N. tripunctata var. schizonemoides, N. heufleri var. leptocephala, and N. erifuga.
The taxonomic status of N. tripunctata var. schizonemoides is largely unclear in theliterature. Specimens depicted in Plate 1 have been traditionally considered by taxonomistsbased in the U.S. to belong to this taxon (e.g., Patrick and Reimer, 1966). The combination N.tripunctata var. schizonemoides was erected by Patrick (1959) based on specimens identified byVan Heurck (1880-1887, Text, p. 83; Atlas, plate VII, Figs. 9 and 10) as N. gracilis var.schizonemoides. Van Heurck considered Schizonema neglectum (Thwaites, 1848) to be thebasyonym of N. gracilis var. schizonemoides. However, observations made by Lange-Bertalot(pers. comm.) on type material of Schizonema neglectum led him to conclude that the latter taxonis simply a synonym of N. tripunctata (nominate variety). Hence, the synonymy established byVan Heurck, and thus Patrick’s combination, might be incorrect. Since only drawings ofSchizonema neglectum are available in the literature at this time, the true identity of this taxonremains unresolved and merits further research.
On the other hand, European workers have often included specimens resembling thosepresented in Plate 1 (Figs. 1-19) in N. recens (Krammer and Lange-Bertalot, 1986; Lange-Bertalot, 1980). Frustule dimensions and characteristics of NAWQA specimens depicted in Plate1 fit the description of N. recens. However, N. recens is also a species that needs further revisionas pointed out by its own author (Lange-Bertalot in Krammer and Lange-Bertalot [1986]). Manydiagnostic features are shared with other taxa and the only character that seems unique; i.e., the

THE ACADEMY OF NATURAL SCIENCES 5 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
convergence of the striae at the valve ends, might be a state of either a purely geneticallydetermined or an environmentally induced character.
The literature contains no reference to the relationship between N. tripunctata var.schizonemoides and N. recens. Production of mucilage tubes by N. tripunctata var.schizonemoides has been reported (Patrick, 1959; Patrick and Reimer, 1966). Evidently, this wasa key feature in separating this variety from N. tripunctata var. tripunctata (also mentioned byCox [1979]). However, Lange-Bertalot (pers. comm.) states that the nominate variety of N.tripunctata is also able to produce mucilaginous tubes under certain environmentalcircumstances. Hence, the usefulness of mucilaginous-tube production as a character forclassification of this group remains unclear. It is not known at this stage whether mucilaginoustubes are simply a facultative feature produced when organisms are subjected to changingenvironmental conditions. Culture studies may prove useful in solving this matter (Patrick, 2000pers. comm.).
NAWQA specimens from Groups I and II differ from N. gracilis var. schizonemoides (A-V.H. 234) (Plate 3, Figs. 1-6). Specimens in the mentioned slide are larger in size, have slightlylower density of striae and areolae, and there are also slight differences in valve shape and striaepattern at the central area. Thwaites’ drawings of S. neglectum (Plate 3, Fig. 11) bear even lessresemblance to organisms in NAWQA samples. These drawings depict valves with parallelstriae and lacking the somewhat acute-angled subfacia (due to shorter striae) characteristic ofNAWQA specimens.
Krammer and Lange-Bertalot’s illustrations of N. recens (Plate 3, Figs. 7-10) are verysimilar to NAWQA specimens allocated in Group I (Plate 1). For the time being, and since therelationship between N. recens and N. tripunctata var. schizonemoides needs clarification, bothnames will be maintained in NAWQA counts. As for the name Navicula sp. 5 ANS WRC, thistaxon should be transferred to N. recens, as agreed by workshop participants.
As stated before, Group II, referred to as N. tripunctata var. schizonemoides, alsoreceived the names N. erifuga and N. heufleri var. leptocephala in NAWQA counts. It was clearto Workshop participants that for this Group II none of the taxonomic names discussed for thefirst group could be applied. A literature search for N. erifuga and N. heufleri var. leptocephalarevealed that these two taxa are synonyms. In 1986, Krammer and Lange-Bertalot concluded thatN. heufleri var. leptocephala was in fact a separate entity since transitional forms between thisand the nominate variety of N. heufleri could not be found. These authors then raised N. heuflerivar. leptocephala to the species level under the name N. erifuga.
Workshop participants agreed to use the name N. erifuga for Group II (Plate 2, Figs. 1-9). In NAWQA counts, the name N. heufleri var. leptocephala was also applied to a group that, inreality, belongs to N. libonesis. Workshop participants agreed that the latter should be applied toorganisms such as those depicted on Plate 2, Figs. 10-13.

THE ACADEMY OF NATURAL SCIENCES 6 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
Although N. tripunctata (nominate variety) was not discussed in depth during theworkshop, none of the participants considered it to be a problematic taxon. It is easilydistinguished from the var. schizonemoides and from N. recens by the straight nature of the striaeand by the symmetry of the subfacies (central area).
Dr. Potapova noted that the taxon illustrated in Plate 4, Figures 1-5 has beenmisidentified as N. tripuncata. This taxon has an asymmetric central area, proximal raphe endsbent unilaterally toward the primary side of the valve, and only superficially resembles N.tripunctata in valve outline and striae pattern. This taxon should be kept separate from N.tripunctata, but a name for it has not been found in the literature. For now, the name Naviculasp. 5 SCTX MP should be used.
K. Manoylova found a taxon (Plate 4, Figs. 6-8) that also resembles taxa in Complex 1and is probably closely related to the N. recens-tripunctata var. schizonemoides group. Thistaxon is distinguishable from all the above taxa by a number of characteristics. Valve outline isclearly lanceolate and the width of the valve is narrower. Valve ends are cuneate and althoughthe striae have similar patterns of orientation and distribution similar to N. recens and N.tripunctata var. schizonemoides, they are finer due to smaller-sized lineolae. This taxon couldnot be found in the literature and may also correspond to a new species. For the time being, itshould be kept separate from the rest of the species in this complex under the name: Navicula sp.1 ACAD NAWQA (NADED ID 203004).
N. margalithii was not discussed during the workshop. Its morphology closely resemblesthat of N. tripunctata (nominate variety), but N. margalithii differs, among other features, in itspreference for brackish to marine habitats.
Navicula cari, although not discussed in depth, is sufficiently distinct and does not posedifficulties. Both N. margalithii and N. cari can be identified using Krammer and Lange-Bertalot(1991, 1986).
Complex 2Navicula cryptotenella
N. menisculus
N. radiosa var. tenella
N. cincta var. rostrata
Many inconsistencies were found in NAWQA counts regarding species in this complex. Such inconsistencies may have had their origin in the apparent existence of transitional formsamong these species. This is illustrated by the case of N. cryptotenella and N. menisculusculus,in which smaller varieties of the latter species (i.e., grunowii, obtusa, and upsaliensis; see below)can resemble N. cryptotenella in both the valve outline and diagnostic valve ornamentation.

THE ACADEMY OF NATURAL SCIENCES 7 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
NAWQA records show that names mentioned above have been used interchangeably and that, inmany instances, more than one morphologically distinct group has received the same name.
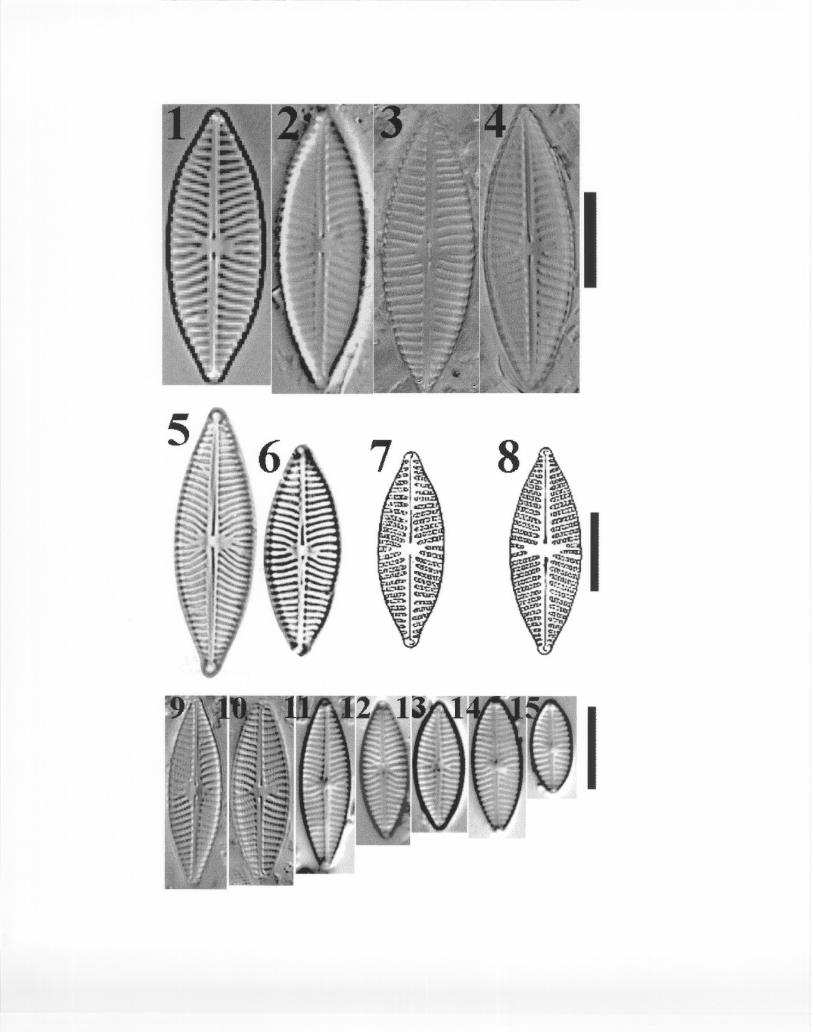
Krammer and Lange-Bertalot (1986) studied populations of N. radiosa var. tenella andconcluded that there are no transitional forms between this and the nominate variety. Hence, theyraised the variety tenella to the species level naming it N. cryptotenella. NAWQA taxonomistswill follow Krammer and Lange-Bertalot’s study and use the name N. cryptotenella. This taxon(Plate 5) is characterized by narrower valves (when compared to N. menisculus and its varieties),a very narrow central nodule, and radiate striae becoming parallel toward the valve ends. Characteristically, a longer stria flanked by two smaller ones is present on at least one side of thecentral area. Although in smaller specimens this feature may be distorted, it may be observed inthe majority of the specimens encountered during routine counts. However, and as pointed outby W. Cody in his presentation of this complex, similar taxa share this diagnostic feature, butfrequently differ from each other in a number of other characteristics. These include differencesin valve outline, thickness of striae (hence, stria number per 10 :m), raphe structure, etc. N.cryptotenella and associated taxa can be found in plates presented by Krammer and Lange-Bertalot (1991, 1986) and they should be used as sources of identification. Plate 5 illustratesspecimens found in NAWQA material and a SEM photograph from the Thames River Estuary,CT.
N. menisculus (nominate variety) is a much more robust taxon than N. cryptotenella(Plate 6, Figs. 1-4). Striae in the former taxon are very conspicuous and lineolae are often visibleduring routine analyses. The shape of the valves is broadly lanceolate with frequently perfectlycuneate ends. Axial area is narrow, but more visible than that of N. cryptotenella. Striae areradiate and sometimes somewhat curved at the central portion of the valve due to the fact that theraphe lies on a depression (ridge) that runs along the axial area of the valve. The striae areslightly radiate to parallel at the valve ends. Although this taxon may occasionally have a shortstria flanked by smaller ones at one side of the central area (e.g., Plate 6. Fig. 1), the reverse isusually true; i.e., a smaller stria is surrounded by two longer ones.
N. menisculus var. upsaliensis (Plate 6, Figs. 5 and 6) differs from the nominate variety inthat the valves possess slightly rostrate ends. Striae are strongly radiate and a higher number ofshorter striae are present in the central axial area. A further difference between this and thenominate variety is that the striae at the poles tend to vary from parallel to slightly convergent inthe var. upsaliensis. Drawings presented by Patrick and Reimer (1966) for this taxon (Plate 6,Figs. 7 and 8) were based on A-V. H. 190 (a specimen from this latter slide is presented in Plate6, Fig. 2). However, Van Heurck’s specimens differ substantially from valves in the typematerial of N. menisculus var. upsaliensis (Krammer and Lange-Bertalot, 1986) (Plate 6, Figs. 5and 6). The latter possess extremely radiate and sigmoid striae in the central area and the striaeat the apices might vary from parallel to slightly convergent. In contrast, specimens in A-V. H.190 are, in general, more robust and have radiate and slightly curved striae in the central area. Striae at the apices are slightly radiate to parallel, though in a few instances they might be slightlyconvergent. This is to say that specimens in A-V. H. 190 bear more resemblance to the nominatevariety of N. menisculus than to the variety upsaliensis (compare Fig. 2 with Figs. 1, 3, and 4 and

THE ACADEMY OF NATURAL SCIENCES 8 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
then with Figs. 5 and 6 in Plate 6). Hence, identification of drawings in Patrick and Reimer(1966) as N. menisculus var. upsaliensis might be incorrect.
N. menisculus var. grunowii is a much smaller taxon than the nominate and theupsaliensis varieties (Plate 6, Figs. 9-15). The ends of the valve vary from slightly cuneate tobroadly rounded. Striae are much finer, radiate in the middle portion to parallel toward the endof the valves. The central axial area presents shorter striae which vary in number from one tofour. The axial area is much narrower than in the nominate and the upsaliensis varieties.
N. menisculus var. obtusa incorporates organisms with elliptical valves. Apices of thevalves are rounded or slightly cuneate. Striae are parallel and end at the same level at the axialarea throughout the length of the valve, although in rare cases a smaller stria can be observed atthe central axial portion of the valve. Lineolae are much longer than in the grunowii variety,hence, striae are much wider in N. menisculus var. obtusa. A plate for this taxon can be found inSimonsen (1987; Plate 329, Figs. 25 and 26).
In NAWQA counts, N. menisculus var. menisculus has been sometimes misidentified asN. menisculus var. upsaliensis. Many other records of N. menisculus var. upsaliensis should bechanged to N. menisculus var. grunowii as revealed by observations of NAWQA material madeby Dr. Potapova. Future identification of the upsaliensis variety should not be based on diagramspresented in Patrick and Reimer (1966). Apparently, the occurrence of this variety is morerestricted in U.S. rivers than that of the variety grunowii. Similarly, the abundance of N.menisculus var. obtusa in these ecosystems is yet to be determined. Currently, NAWQA countscontain a few records of this taxon, but further research is needed to verify its identity. Wemention it here to avoid its misidentification in the future.
Type material for N. cincta var. rostrata was analyzed during the workshop (Plate 7, Figs.1-8). This taxon seems to be sufficiently different from other taxa to support its maintenance as aseparate taxon. In NAWQA counts, this name has been misapplied and in all cases records of N.cincta var. rostrata should be changed to N. veneta. The latter taxon can be identified based onKrammer and Lange-Bertalot (1986, Plate 32, Figs. 1-4, p. 505).
Complex 3Navicula notha
N. leptostriata
N. heimansioides
The NAWQA database contains records of these species. However, the fact that all threetaxa are very closely related at the morphological level made it difficult to separate them intodistinct groups. Additionally, past differences among NAWQA taxonomists about the identity of

THE ACADEMY OF NATURAL SCIENCES 9 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
N. notha and N. leptostriata made it clear that a review of the taxonomy of these taxa wasneeded.
Examination of plates and discussion presented by Krammer and Lange-Bertalot (1986)for N. heimansioides and by Wallace (1960) for N. notha led to the conclusion that these twotaxa actually represent morphological variations of a single species. The only basis used byKrammer and Lange-Bertalot for separation of these two taxa is the larger size of the frustules ofN. heimansioides. Examination of the type material of N. notha (Plate 7, Figs. 9-17) revealedthat the range of length given by Wallace was much shorter than many of the valves present inhis permanent slide (A. G. C. 4613b). Larger specimens in such material are morphologicallysimilar to smaller representatives of the same population, and thus, cannot be separated fromspecimens diagrammed by Wallace (compare Figs. 9-17 with Figs. 22-25 in Plate 7).
Previous changes made in NAWQA counts from N. notha to N. leptostriata seemtherefore not to be correct. An example of the occurrence of N. notha in NAWQA material isgiven in Plate 7, Figs. 18-21. N. leptostriata and N. notha can be distinguished from each otherbased on a number of characteristics. N. leptostriata is much less silicified, the endings of theraphe at the central nodule are closer to each other, striae are much finer (due to smallerlineolae), and the central area is much wider (Krammer and Lange-Bertalot, 1991, Plate 70, Figs.9-14, p. 388). The extent of the occurrence of N. leptostriata in NAWQA material is yet to bedetermined. Available data suggest that this taxon is not common in U.S. rivers.
Van Dam and Kooyman (1982) described N. heimansii, but it was later determined byKrammer and Lange-Bertalot (1986) that this taxon should be a synonym of N. leptostriata.
Complex 4Navicula biconica
N. luzonensis
N. subminuscula
Dr. Potapova found that the names N. luzonensis and N. biconica were applied to a singlemorphological group. Also, the names N. luzonensis and N. subminuscula were usedinterchangeably to identify the morphological group illustrated in Plate 8, Figs. 1-11.
The confusion between N. luzonensis and N. biconica seems to have had its origin in thedrawing presented by Patrick and Reimer (1966) for the latter taxon (Plate 8, Fig. 12). Thisdrawing depicts a much wider valve and much coarser striae than the type specimen (Plate 8, Fig.13), characteristics that resemble more those of N. luzonensis (compare Figs. 1-11 with 13-18 inPlate 8).

THE ACADEMY OF NATURAL SCIENCES 10 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
In the specific case of the Central Nebraska study unit (Figs. in Plate 8, except Figs. 12and 13), both N. luzonensis and N. biconica are present. In many samples from this study unit,however, counts of N. biconica should reflect lower percentages than those recorded byNAWQA taxonomists.
An additional observation of specimens of N. biconica in the Central Nebraska study unitis that they seem to represent transitional forms between the type specimen of N. biconica (Plate8, Fig. 13) and representatives of N. molestiformis, as presented in Krammer and Lange-Bertalot(1986, Plate 45, Figures 1-9, p. 531). Thus, the relationship between these latter taxa should bereviewed.
As demonstrated by Krammer and Lange-Bertalot (1986), N. luzonesis (Hustedt 1942) isa synonym of N. subminuscula (Manguin, 1941). Therefore, future NAWQA counts should referto specimens such as those presented in Plate 8, Figs. 1-11 as N. subminuscula.
Workshop participants agreed that most records of N. biconica should be changed to N.subminuscula, and that all records of N. luzonensis should be changed to N. subminuscula. Asfor records of N. biconica in the Central Nebraska study unit, they should be kept as they are fornow until the relationship of this population and N. molestiformis is clarified.
Complex 5Gomphonema pumilum spp.
The taxonomic position of this taxon was not discussed in depth and should beconsidered as a topic for a future workshop. Although this taxon is sufficiently distinct fromclosely related taxa, NAWQA taxonomists have either confused it with other closely relatedgomphonemoid taxa or have applied the name G. pumilum to different morphological groups. Inlight of these problems, correcting the taxonomy of this taxon will be a difficult endeavor. Fornow, most records for G. pumilum will be left as they are. In the future, however, NAWQAtaxonomists must differentiate this species from other taxa using Kociolek and Kingston (1999),Kociolek and Stoermer (1990, 1991), and Reichardt (1997).
Complex 6Gomphonema minutum
G. kobayasii
Many NAWQA counts contain records of these two taxa. In many cases these two nameshave been used to refer to a single morphological group. However, these two species are easily

THE ACADEMY OF NATURAL SCIENCES 11 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
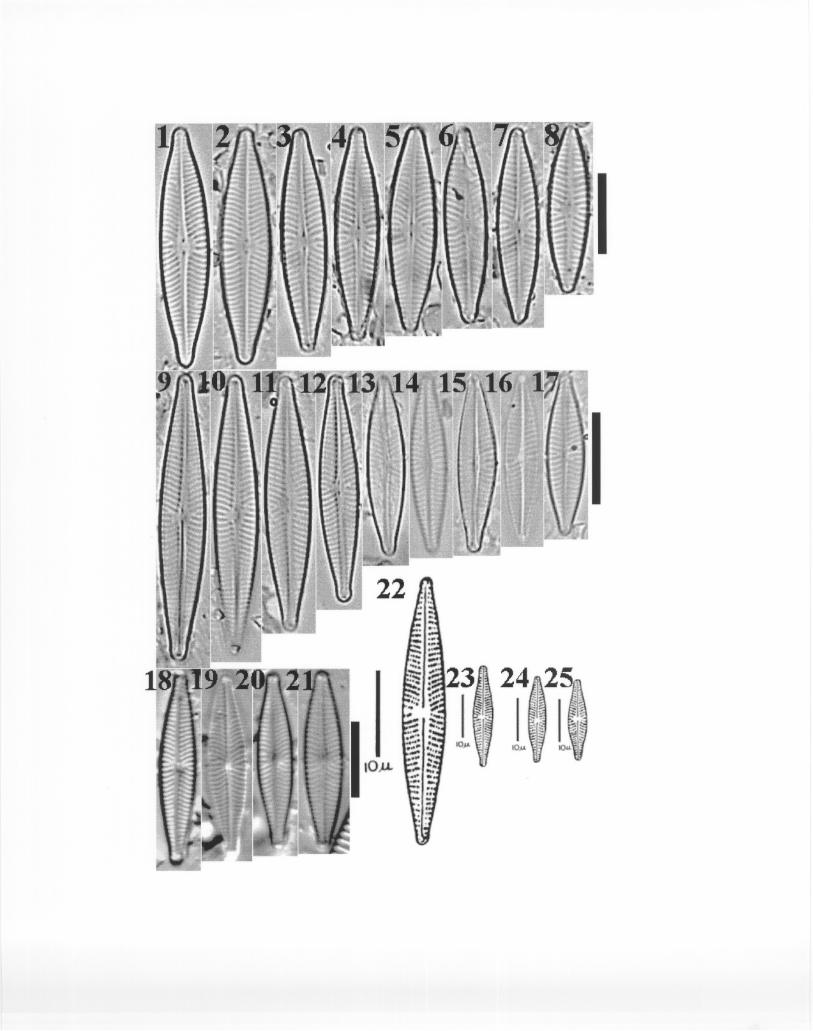
distinguished from each other in both their valve and girdle views. In G. minutum (Plate 9, Figs.1-19) valves are broadly clavate, striae are much coarser due to the existence of double rows ofporoids (SEM), and the arching of the striae at the central nodule is very conspicuous. In girdleview, this taxon is distinctly wedge-shaped and a single row of poroids on the valvocopulae canbe seen easily (Krammer and Lange-Bertalot, 1991, Tafel 81, Fig. 4, pg. 411).
NAWQA counts of this taxon also include an entity that is closer to G. tenellum, aspresented in Patrick and Reimer (1975). The latter presents an incipient capitate head pole that isbroader than that of G. minutum. For now, these two morphs will be kept together, but furtherstudies should be conducted to elucidate their true identity.
In G. kobayasii valve contours are not broadly clavate, but lanceolate (Plate 10, Figs. 1-11). Striae are parallel to slightly radiate in arrangement; striae on the opposite side of the stigmaare distinctly disordered and irregular. The striae are much more sparse than in G. minutum andthey are also less conspicuous (composed of lineolae as revealed by SEM). The axial area isbroader than in G. minutum. Girdle views of G. kobayasii have the shape of rectangular boxesand also show distinct, but much coarser, rows of poroids on the valvocopulae. Plates for G.minutum and G. kobayasii are presented here (Plates 9 and 10), but also refer to Kociolek andKingston (1999) for further details on these taxa.
Although not in this complex, the taxonomic position of Gomphoneis herculeana and G.minuta was discussed (Plate 11, Figs. 1-7). Valve dimensions and striae number for both ofthese taxa are similar to each other and the only basis for their separation at this point are themore linear valve outline and the absence of an inflated portion in the valves of G. minuta (Kociolek and Stoermer, 1988) (compare Figs. 1 and 3 with 2 and 4 in Plate 11). Moreinformation on the morphology of these two entities, as well as their ecology, should beinvestigated to establish whether there is a clear separation between them.
Additionally, gomphonemoid diatoms without stigmata from the Arcadian-Pontchartrainstudy unit were presented by K. Manoylova (Plate 11, Figs. 8-12). Her specimens closelyresemble Gomphonema patrickii (Kociolek et al., 1995), and as such should be inserted in theNAWQA database.

THE ACADEMY OF NATURAL SCIENCES 12 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
ADDITIONAL TAXA CONSIDERED DURING THE WORKSHOP
In addition to the species above, the taxonomic status of several other taxa wasconsidered. These taxa were included in discussions due to their resemblance to some of the taxaoriginally included in the six complexes presented above.
In the case of N. symmetrica, workshop participants agreed not to follow the groupingproposed by Krammer and Lange-Bertalot (1991) in which N. symmetrica is placed as a varietyin N. schroeteri. N. symmetrica is distinct from N. schroeteri and associated taxa and shouldstand on its own. As for the rest of the varieties in N. schroeteri presented by Krammer andLange-Bertalot (1991), i.e., the varieties schroeteri and escambia, NAWQA taxonomists willfollow the mentioned authors’ taxonomy. The retention of the name N. symmetrica will notcause inconsistencies in existing NAWQA data because all taxonomists distinguished this taxonfrom N. schroeteri in the past.
N. lanceolata was also discussed during the workshop. This taxon had been discussedduring the third workshop, but Dr. Bahls suggested a re-consideration of its taxonomic position. Participants examined type material from the Agardh Collection in the ANSP Diatom Herbariumand concluded that Krammer and Lange-Bertalot’s decision to synonymize Frustulia lanceolataAgardh with N. lanceolata (Agardh) Ehrenberg is in fact correct (Krammer and Lange-Bertalot,1986). Also, the same authors were correct in synonymizing N. lanceolata (Agardh) Kützingwith N. trivialis Lange-Bertalot. For NAWQA purposes, the name N. viridula var. avenacea willbe changed to N. lanceolata (Agardh) Ehrenberg and analysts will continue using the name N.trivialis.
The use of the name N. vandamii (Krammer and Lange-Bertalot, 1991) should be avoidedand N. canalis is preferred in its place. The reason for this is that N. canalis (Patrick, 1944) haspriority in time over N. vandamii (Schoeman and Archibald, 1986).
Examination of the type slide of N. canalis from the ANSP Diatom Herbarium (Plate 12,Figs. 1 to 4)) led to the conclusion that both of these names have been applied to a single species. Additionally, K. Manoylova presented specimens which she originally had identified as Naviculasp. 1 0B UL NAWQA 96 KM, but now should be changed to N. canalis (Plate 12, Figs. 5 and 6). All records of N. vandamii in the NAWQA database should be changed to N. canalis.
N. veneta, N. exilis, and N. reichardtiana were also considered during the workshop. InNAWQA counts N. reichardtiana was commonly grouped together with N. veneta and/or N.cryptocephala var. veneta, whereas N. exilis was often included in N. veneta.
N. veneta, N. exilis, and N. reichardtiana are distinct entities and should be kept separatein NAWQA counts. Thus, appropriate changes should be made and future identification of allthese taxa should be based on plates presented by Krammer and Lange-Bertalot (1991, 1986). Krammer and Lange-Bertalot (1986) raised N. cryptocephala var. veneta to the species level

THE ACADEMY OF NATURAL SCIENCES 13 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
under the name N. veneta. This decision was based on the absence of transitional forms betweenN. cryptocephala var. veneta and the nominate variety of N. cryptocephala. Such a decision willbe adopted by NAWQA taxonomists.
Workshop participants adopted the decision made by Germain (1981) indicating that N.salinarum var. intermedia is not a variety of closely related taxa such as N. salinarum or N.cryptocephala. For this reason, it should be considered as a different species and the name N.capitatoradiata should be used to refer to it.
Finally, the case of N. secreta var. apiculata and N. gregaria was also considered duringthe Workshop. These two taxa were determined to be sufficiently different as to maintain theircurrent status. N. secreta var. apiculata is a much more robust taxon and its pattern of striation isdifferent from that of N. gregaria (compare Figs. 7 and 8 with Figs. 9 and 10 in Plate 12). Inboth taxa the striae are composed of lineolae, but in N. secreta var. apiculata these structures arelarger, and therefore, the striae are more conspicuous and their density is lower (Plate 12, Figs. 7and 8). There are also, differences in the structure of the raphe. In N. gregaria, the raphe lies ona ridge and the ends of this structure at the central nodule bend unilaterally (Plate 12, Figs. 9 and10). Although the raphe also lies on a ridge in N. secreta var. apiculata, the proximal raphe endsare directly opposite to each other and do not bend to the sides.
Both taxa should continue to be kept separate during analyses of NAWQA material andtheir identification should be based on photographs presented herein. Navicula gregaria hasbeen observed to be far more common in NAWQA samples than N. secreta var. apiculata.

THE ACADEMY OF NATURAL SCIENCES 14 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
REFERENCES
Academy of Natural Sciences of Philadelphia (ANSP). 1999. First NAWQA Diatom TaxonomyHarmonization Workshop. Clason, T.A. and D.F. Charles. Patrick Center forEnvironmental Research. The Academy of Natural Sciences of Philadelphia. Report No.99-14.
_________. 2000a. Second NAWQA Diatom Taxonomy Harmonization Workshop. Clason,T.A. and D.F. Charles. Patrick Center for Environmental Research. The Academy ofNatural Sciences of Philadelphia. Report No. 00-7.
_________. 2000b. Third NAWQA Diatom Taxonomy Harmonization Workshop. E.A.Morales and M. Potapova. Patrick Center for Environmental Research. The Academy ofNatural Sciences of Philadelphia. Report No. 00-8.
Barber, H. G. and Haworth, E. Y. 1981. A Guide to the Morphology of the Diatom Frustule. Freshwater Biological Association. Sci. Publ. No. 44. Titus Wilson & Son Ltd. England. 112 pp.
Cox, E.J. 1979. Taxonomic studies of the diatom genus Navicula Bory: The typification of thegenus. Bacillaria 2:137-154.
_________. 1996. Identification of Freshwater Diatoms from Live Material. Chapman & Hall. England. 158 pp.
Germain, H. 1981. Flore des diatomées. Diatomophycées eaux douces et sumâtres du MassifArmoricain et des contrées voisines d’ Europe occidentale. Société Nouvelle desÉditions Boubée. Paris, France. 444 pp.
Hustedt, F. 1942. Süsswasser-Diatomeen del indomalayischen Archipiels und der Hawaii-Inseln. Internat. Rev. Hydrobiol. 42:1-252.
Kociolek, J.P. and J.C. Kingston. 1999. Taxonomy, ultrastructure, and distribution of somegomphonemoid diatoms (Bacillariophyceae: Gomphonemataceae) from rivers in theUnited States. Can. J. Bot. 77:686-705.
Kociolek, J. P. & Stoermer, E. F. 1988. Taxonomy, ultrastructure and distribution ofGomphoneis herculeana, G. eriense and closely related species (Naviculales:Gomphonemataceae). Proc. Acad. Nat. Sci. Phil. 140: 24-97.
Kociolek, J.P. and E.F. Stoermer. 1990. A new, highly variable Gomphonema(Bacillariophyceae) species from Laurentian Great Lakes. Pages 139-144 in M. Ricard

THE ACADEMY OF NATURAL SCIENCES 15 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
and M. Coste, eds. Ouvrage dédié à la Mémorie du Professeur Henry Germain (1903-1989). pp. 139-144. Koeltz Scientific Books, Germany.
_________. 1991. Taxonomy and ultrastructure of some Gomphonema and Gomphoneis taxafrom the upper Laurentian Great Lakes. Can. J. Bot. 169:1557-1576.
Kociolek, J.P., E.F. Stoermer and M.A. Edlund. 1995. Two new freshwater diatom species. Pages 9-19 in J.P. Kociolek and M.J. Sullivan, eds. A Century of Diatom Research inNorth America: A tribute to the Distinguished Careers of Charles W. Reimer and RuthPatrick. Koeltz Scientific Books, Germany.
Krammer, K. and Lange-Bertalot, H. 1986. Bacillariophyceae. 1. Teil: Naviculaceae.In: H. Ettl,J. Gerloff, H. Heynig and D. Mollenhauer, eds. Süsswasserflora von Mitteleuropa. Vol.2(1):1-876. Gustav Fisher Verlag, Germany.
_________. 1991. Bacillariophyceae. 4. Teil: Achnanthaceae. Kritische Ergänzungen zuNavicula (Lineolatae) und Gomphonema. In: H. Ettl, J. Gerloff, G. Gärtner, H. Heynigand D. Mollenhauer, eds. Süsswasserflora von Mitteleuropa. Vol. 2(4):1-437. GustavFisher Verlag, Germany.
Lange-Bertalot, H. 1980. Zur taxonomischen Revision einiger ökologische wichtiger“Naviculae lineolatae” Cleve. Die Formenkreise um Navicula lanceolata, N. viridula, N.cari. Cryptogamie Algalogie 1:29-50.
Manguin, E. 1941. Contribution à la flore des diatomées d’eau douce de Madagascar. ReviewAlgologie 12:153-157.
Patrick, R. 1959. New changes and nomenclatural changes in the genus Navicula. Proc. Acad.Nat. Sci. Philadelphia 111:91-108.
Patrick, R. and C.W. Reimer. 1966. The Diatoms of the United States. Exclusive of Alaska andHawaii. Vol. 1. Monographs of the Academy of Natural Sciences of Philadelphia 13. 688 pp.
_________. 1975. The Diatoms of the United States. Exclusive of Alaska and Hawaii. Monographs of the Academy of Natural Sciences of Philadelphia 2(13):1-213.
Reichardt, E. 1997. Taxonomische revision des artenkomplexes um Gomphonema pumilum(Bacillariophyceae). Nova Hedwigia 65:99-129.

THE ACADEMY OF NATURAL SCIENCES 16 PATRICK CENTER FOR ENVIRONMENTAL RESEARCH
Schoeman, F. R. & Archibald, R. E. M. 1986. Navicula vandamii nom. nov. a new name forNavicula acephala Schoeman, and a consideration of its taxonomy. Nova Hedwigia 44:479-487.
Simonsen, R. 1987. Atlas and Catalogue of the Diatom Types of Friedrich Hustedt. Vols. 1-3. J. Crammer, Germany.
Thwaites, G.H.K. 1848. Further observations on the diatomaceae with descriptions of newgenera and species. Ann. Mag. Nat. Hist. 1(2):161-172.
Van Dam, H. and H. Kooyman. 1982. A new diatom from Dutch moorland pools: Naviculaheimansii. Acta Bot. Neerlandica 31:1-4.
Wallace, J.H. 1960. New and Variable Diatoms. Notulae Natura 331:1-8.