flexibility and fine structure of smooth-muscle α-actinin
TRANSCRIPT

Eur. J. Biochem. 248, 193-199 (1997) 0 FEBS 1997
Flexibility and fine structure of smooth-muscle a-actinin Jorg WINKLER', Heinrich LUNSDORF' and Brigitte M. JOCKUSCH'
' Cell Biology, Zoological Institute, Technical University of Braunschweig, Braunschweig, Germany ' Department of Microbiology, GBF-National Research Center for Biotechnology, Braunschweig, Germany
(Received 2 May 1997) - EJB 97 0636/3
The microfilament protein a-actinin exists as a dimer. The N-terminal regions of both polypeptides, arranged in antiparallel orientation, comprise the actin-binding regions, while the C-terminal, larger parts consist of four spectrin-like repeats that interact to form a rod-like structure. To elucidate the fine structure of smooth-muscle a-actinin, we used energy-filtered transmission electron microscopy in conjunction with negative staining. Survey pictures of the protein purified from chicken gizzard revealed discrete, elongated particles whose length and width varied with the ionic strength of the buffer. It was determined to to 29.3 nmX4.8 nm in 0.05 M KC1 and 32.6 nmX4.4 nm in 0.15 M KC1. Both ends of the molecule displayed hook-like structures consisting of globular domains, which were highly variable in their orienta- tion with respect to the long axis of the molecule. Their location at the ends of the molecule, and the finding that these hooks were missing from particles obtained by thermolysin digestion indicated that they probably correspond to the N-terminal actin-binding regions. The rod-like center of the molecule revealed discrete globular masses which probably comprise the spectrin-like repeats. Their arrangement was compatible with the interpretation that three spectrin repeats of each polypeptide chain can form pairs with the respective sequences of the other chain. The rod-like 53-kDa fragment obtained after thermolysin digestion largely retained this structural organization but appeared wider (22.5 nmX 5.9 nm). Our results help to clarify previous discrepancies on the quatenary organization of a-actinin and suggest that effective actin-binding and cross-linking of a-actinin is based on the high flexibility of the terminal hooks.
Keywords: cytoskeleton; a-actinin; structure ; electron microscopy ; electron-spectroscopic imaging.
a-Actinin is an F-actin cross-linking protein highly enriched in the Z-disk of striated muscle (Masaki et al., 1967; Lazarides and Granger, 1978; Yamaguchi et al., 1985), and in the cyto- plasmic and membrane-associated dense plaques of smooth muscle (Geiger et al., 1981 ; Small, 1985). In non-muscle cells, a-actinin is located in focal adhesions and in stress fibers as well as in the cortical web (Lazarides and Burridge, 1975; Sanger et al., 1983). In addition to F-actin, the muscle and non-muscle isoforms of a-actinin bind to various proteins of focal adhesion sites including vinculin (McGregor et al., 1994; Kroemker et al., 1994), P,-integrin (Otey et al., 1990) and zyxin (Crawford et al., 1992). Based on sequencing data, the a-actinin polypeptide can be divided into three main regions: an evolutionary well-con- served N-terminal F-actin binding domain, four internal 122- amino-acid repeats (spectrin-like repeats), and a short C-terminal calcium-binding motif containing two EF-hands, which is dys- functional in the muscle isoforms (Baron et al., 1987; Noegel et al., 1987; Blanchard et al., 1989). In the dimeric molecule, the polypeptides are oriented in an antiparallel fashion (Wallraff et al., 1986).
Several reports dealt with the structural organization of muscle a-actinin. Electron microscopy provided images of rod-
Correspondence to J. Winkler, Cell Biology, Zoological Institute, Technical University of Braunschweig, Spielmannstrasse 7, D-38092 Braunschweig, Germany
Fax: +49 531 391 8203. E-mail: [email protected] Abbreviations. ESI, electron spectroscopic imaging; EFTEM, energy
filtered electron microscopy ; ABD, actin-binding domain.
shaped molecules, measuring 30-40 nm by 2-4.5 nm (Podlub- naya et al., 1975; Suzuki et al., 1976; Imamura et al., 1988). Regarding the quatenary organization, there are conflicting re- ports with respect to the arrangement of the spectrin-like repeats. Projection images obtained from electron micrographs of two- dimensional crystals of smooth muscle a-actinin revealed six density peaks for each polypeptide (Taylor and Taylor, 1993). These probably comprise the two F-actin binding domains and the four spectrin-like repeats. Within the dimer, the eight spectrin-like repeats were proposed to be arranged in three central pairs, leaving the first repeat unpaired at either end. In contrast, Flood et al. (1995) provided evidence that in aqueous solution the eight spectrin-like repeats of a dimer interact with each other, giving rise to four pairs, rather than three.
To obtain further information on the ultrastructural organiza- tion of smooth-muscle a-actinin and to possibly resolve the discrepancy described above, we analyzed the intact protein and its 53-kDa thermolysin-derived fragment by energy-filtered transmission electron microscopy (EFTEM). Images obtained by this technique revealed that a-actinin is composed of discrete globular domains. The terminal ones on each side are arranged in highly flexible, hook-like structures, while the central ones contribute to the rod-like central region. Our results favour a model proposing the pairing of only three spectrin-like repeats in the central rod.
MATERIALS AND METHODS Purification of intact a-actinin and its thermolysin-de-
rived fragments. Chicken gizzard a-actinin was purified ac-

194 Winkler et al. ( E m J. Biochem. 248)
a b c 1004
53-
27-
Fig. 1. Coomassie blue stained profiles of purified chicken gizzard a- actinin (lane a), and its 27-kDa (lane b) and 53-kDa fragments (lane c) generated by thermolysin cleavage, after SDSPAGE on 12 % gels.
cording to Feramisco and Burridge (1980). Briefly, the purified protein was dialyzed against buffer B, consisting of 0.02 M Tris/ HCI, pH 7.5, 0.05 M KCl, 0.1 mM EGTA, 0.2 mM phenyl- methylsulfonyl fluoride and 0.2 mM dithioerythritol. To obtain the 53-kDa fragment, the protein (1 mg/ml) was digested for 2 h with 0.03 mg/ml thermolysin at 37°C in buffer B containing 1 mM CaC1,. The reaction was terminated by the addition of 5 mM EGTA (final concentration). The cleavage products were loaded onto a HR 10/10 Mono Q column and eluted with a linear gradient of 0 to 0.5 M NaCl. Protein concentration was determined by the method of Bradford (1976). SDSPAGE was performed using 12 % polyacrylamide slab gels, as described by Laemmli (1970). Fig. 1 shows the polypeptide profile of purified a-actinin and its proteolytic fragments.
Electron microscopy. The purified protein was diluted in buffer B to a final concentration of 50 pg/ml. To obtain physio- logical salt conditions, the KC1 concentration was increased to 0.15 M. Unfixed protein samples were negatively stained with uranyl acetate (4 % mass/vol.) according to the diffusion tech- nique described by Valentine et al. (1968). Since it was shown
that prolonged staining with uranyl acetate may alter the width of a-actinin (Suzuki et al., 1976), the staining procedure was limited to approximately 5 s. Ultrathin carbon foils were mounted on perforated carbon-collodion films (Lunsdorf and Spiess, 1986). The samples were analyzed with a CEM 902 transmission microscope equipped with an integrated electron energy-loss spectrometer (Zeiss) in the elastic bright-field at an energy loss of A E = 0 eV and in the inelastic dark-field at AE = 115 eV, specific for the uranium edge O,, (Bauer, 1988). For rotary shadowing, protein samples at 0.2 mg/ml in 40% (mass/ vol.) glycerol were sprayed onto freshly cleaved mica. The samples were dried in vacua at room temperature for 1 h. Shad- owing was performed in a (BAL-TEC MED 020) with platinum- carbon at an elevation angle of 7". Exposures were carried out at X30000 and X50000 primary magnifications using an accel- eration voltage of 80 kV, and an objective aperture of 30 pm.
Measurements of molecular length and width. The length and width of a-actinin molecules were measured on images pro- jected to a final magnification of 450000X. To estimate the mean width of a given population, each molecule was measured three times at the widest point of its internal region. To obtain reliable data, measurements were restricted to molecules whose structure appeared well preserved under intermediate or deep stain conditions with clearly discernible contours (Mayer et al., 1988). Care was taken to include all molecules of a given area for statistical evaluations.
RESULTS
ESI reveals structural details in a-actinin molecules. When purified chicken smooth muscle a-actinin was negatively stained and examined in the electron microscope using the ESI (Fig. 2a-c) or the conventional bright-field mode (Fig. 2d), elongated particles were seen as the most prominent structures.
Fig.2. Field views of negatively stained a-actinin purified from chicken gizzard smooth muscle. (a, b) Dark-field images at A E = 115 eV showing protein samples i n 0.05 M KC1 (a, low-salt conditions) and 0.15 M KCI (b, high-salt conditions). (c, d) Corresponding images taken either at AE = 115 eV (c), or in the elastic bright-field mode at AE = 0 eV (d). Identical molecules of both views are encircled. (e) Replica of platinum- carbon rotary shadowed a-actinin (arrowheads) imaged in reversed contrast. ds = deep stain, is = intermediate stain. Bars (a, b) = 40 nm. Bars (c-e) = 20 nm.

195
- 5, . - 8 10
2
5
0 20 n 8 . 0
25 30 c 35 11 40
Winkler et al. (EUK J. Biochem. 248)
a
b
c
Molecule length (nm)
Fig. 3. Diagram showing the length distribution of chicken smooth muscle a-actinin. Black bars, a-actinin in 0.15 M KCI. White bars, a- actinin in 0.05 M KCI.
d a b
I
Fig. 4. Negatively stained a-actinin imaged in the uranium dark-field under different salt conditions. The gallery emphasizes different mo- lecular lengths of individual molecules obtained under (a) low-salt (0.05 M KCl) or (b) high-salt conditions (0.15 M KCl). The terminal centers of mass, which appeared either roughly globular or oval-shaped are marked by arrows. The internal density peaks are marked by ar- rowheads. Bar = 10 nm.
Based on previous evidence, it was assumed that each of these particles represented a homodimer of a-actinin polypeptides (Su- zuki et al., 1976; Imamura et al., 1988). A closer look at single particles (encircled in Fig. 2a,b) revealed that each one com- prises several masses. These structural features became more apparent in specimens obtained from low salt preparations (Fig. 2a), as compared with a-actinin particles prepared under physiological conditions (Fig. 2 b). A comparison between corre- sponding images, taken either in the EST mode, or the elastic bright-field, is shown in Fig. 2c and 2d, respectively. The con- trast ratio of stain salt versus protein is significantly enhanced in the uranium image. Especially those molecules that were imaged
Fig. 5. Gallery of negatively stained a-actinin revealing a variety of molecular profiles of its rod-like central region. (a) The central por- tion of these molecules presents themselves as thin rods without internal substructures. (b) The rod-like center displays a regular pattern of globu- lar masses (arrowheads), partially merged with each other. The mole- cules shown in (c) are composed of six discrete globular masses arranged like pearls on a string. (d) Thicker molecules with globular or dumbbell- shaped masses arranged side by side in a slightly shifted or diagonal arrangement (arrowheads). In shallow stain, some molecules reveal an internal structure which appears roughly zigzag-shaped (d7,X). This may be an artifact due to insufficient sustaining and subsequent flattening of single globular masses during air drying. Bar = 10 nm.
under intermediate or shallow stain conditions (Fig. 2c, marked is) revealed significantly more fine-structural information than the corresponding bright-field image (Fig. 2d, marked is*). Such differences between both imaging modes were less obvious with molecules projected under deep stain conditions (Fig. 2c, d ; molecules marked ds/ds*). For comparison, we also examined parallel preparations obtained after rotary shadowing (Fig. 2 e). Printed at the same magnification, the particles also appear rod- like but larger and without distinct structural details, which is probably the result of the metal coating. These results prompted us to concentrate only on the ESI technique for further analysis.
The overall dimensions depend on the ionic strength of the buffer. Length measurements of individual molecules seen in the uranium dark-field revealed an average molecular length of 32.6%3.7 nm (n = 75) under physiological ionic strength (0.15 M KCl), but only 29.3 23.2 nm (n = 85) in low salt (0.05 M KCl). Fig. 3 shows the statistical distribution of the par-

196 Winkler et al. ( E m J. Biochem. 248)
a b 1
C
2
- Fig. 6. Gallery of molecules showing the variability of the terminal regions. (a) Molecules show terminal kinks either in the same (a, cis-like) or in opposite (b, trans-like) orientation (arrows). Furthermore, these terminal regions are composed of single masses (c, arrows) and may appear hook-like (cl, c2), or in the form of open (c,) or closed (c,) triangles. Bar = 10 nm.
ticle length under both conditions. In Fig. 4, molecules corre- sponding to extreme values were selected to emphasize the length variability of a-actinin under both salt conditions. Struc- tural details were generally more discernible in the shorter mole- cules imaged under low salt conditions (Fig. 4a). Both ends of the particles displayed two globular centers of mass (arrows), which were tentatively interpreted as the actin-binding sites lo- cated at the N-terminus (Taylor and Taylor, 1993). In some par- ticles, the central rod-like region of the molecules displayed globular masses grouped in pairs (arrowheads). The longer par- ticles obtained in high salt (Fig. 4 b) displayed more extended terminal hooks and appeared thinner. Again, under favorable conditions, globular or oval-shaped protein masses were visible (arrowheads).
Different orientations of a-actinin result in different mole- cule profiles. In both types of buffers, the diameter of the rod- shaped central region appeared variable. Width measurements of particles, as determined in the Materials and Methods section, yielded a mean diameter of 4.8 ? 0.8 nm (n = 58) for low salt preparations and 4.420.7 nm (n = 67) for physiological ionic strength. Thus, the deviation from the mean was quite large. Furthermore, the internal organization appeared quite variable even within one buffer system. This is evident from the gallery of individual molecules seen in Fig. 5. Part of the molecules depicted appear without any internal structures (Fig. 5 a), others display several regulary spaced globules (Fig. 5 b, arrowheads). In addition, a minor fraction seemingly consisted of discrete globular masses arranged like a string-of-beads (Fig. 5 c). In Fig. 5 d, particles are seen displaying a larger diameter, with an average value of 5.8 ?0.3 nm (n = 29), compared with 3.6 -C 0.2 nm (n = 28) obtained from molecules in Fig. 5 a-c. Such a heterogeneous appearance, observed under identical experi- mental conditions may be caused by different orientations of the particles relative to the electron beam. To elucidate the contribu- tion of different staining conditions to the structural appearance we collected a gallery of particles in similar orientation, but ima- ged under different staining conditions (Fig. 5dl-d. From di to
the conditions gradually change from deep to shallow stain. Under deep stain conditions, which correspond to a brighter
Fig. ,. 53-kDa thermolysin fragment of gizzard (a) Survey view, showing rod-shaped particles (mows), (b, c. Gallery of single molecules, showing a pattern of discrete lobes (b), which form gaps, allowing stain penetration between the protein masses (c, arrowheads). Bar = 10 nm.
Winkler et al. ( E M J . Biochem. 248) 197
a
b
C
d ABd
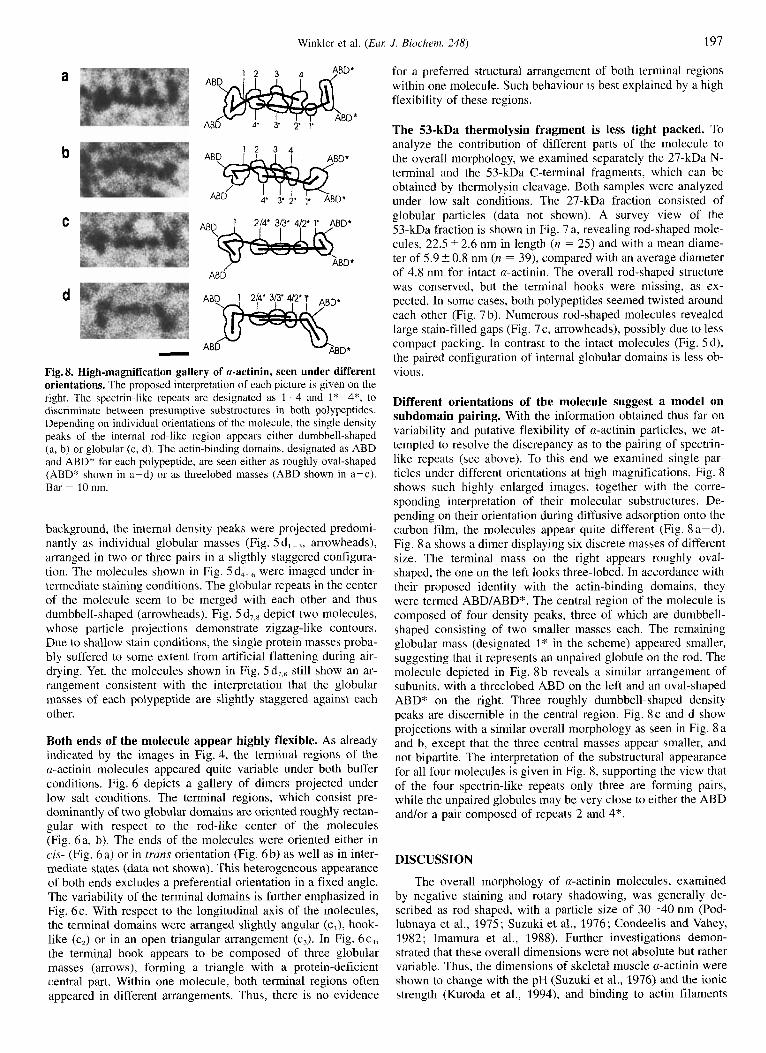
- Fig. 8. High-magnification gallery of a-actinin, seen under different orientations. The proposed interpretation of each picture is given on the right. The spectrin-like repeats are designated as 1-4 and I*-4*, to discriminate between presumptive substructures in both polypeptides. Depending on individual orientations of the molecule, the single density peaks of the internal rod-like region appears either dumbbell-shaped (a, b) or globular (c, d). The actin-binding domains, designated as ABD and ABD* for each polypeptide, are seen either as roughly oval-shaped (ABD* shown in a-d) or as threelobed masses (ABD shown in a-c). Bar = 10 nm.
background, the internal density peaks were projected predomi- nantly as individual globular masses (Fig. 5 d, - -3r arrowheads), arranged in two or three pairs in a sligthly staggered configura- tion. The molecules shown in Fig. 5 d,. were imaged under in- termediate staining conditions. The globular repeats in the center of the molecule seem to be merged with each other and thus dumbbell-shaped (arrowheads). Fig. 5 d,,8 depict two molecules, whose particle projections demonstrate zigzag-like contours. Due to shallow stain conditions, the single protein masses proba- bly suffered to some extent from artificial flattening during air- drying. Yet, the molecules shown in Fig. 5d,,, still show an ar- rangement consistent with the interpretation that the globular masses of each polypeptide are slightly staggered against each other.
Both ends of the molecule appear highly flexible. As already indicated by the images in Fig. 4, the terminal regions of the a-actinin molecules appeared quite variable under both buffer conditions. Fig. 6 depicts a gallery of dimers projected under low salt conditions. The terminal regions, which consist pre- dominantly of two globular domains are oriented roughly rectan- gular with respect to the rod-like center of the molecules (Fig. 6a, b). The ends of the molecules were oriented either in cis- (Fig. 6a) or in trans orientation (Fig. 6b) as well as in inter- mediate states (data not shown). This heterogeneous appearance of both ends excludes a preferential orientation in a fixed angle. The variability of the terminal domains is further emphasized in Fig. 6c. With respect to the longitudinal axis of the molecules, the terminal domains were arranged slightly angular (c,), hook- like (c,) or in an open triangular arrangement (cJ. In Fig. 6cq, the terminal hook appears to be composed of three globular masses (arrows), forming a triangle with a protein-deficient central part. Within one molecule, both terminal regions often appeared in different arrangements. Thus, there is no evidence
for a preferred structural arrangement of both terminal regions within one molecule. Such behaviour is best explained by a high flexibility of these regions.
The 53-kDa thermolysin fragment is less tight packed. To analyze the contribution of different parts of the molecule to the overall morphology, we examined separately the 27-kDa N- terminal and the 53-kDa C-terminal fragments, which can be obtained by thermolysin cleavage. Both samples were analyzed under low-salt conditions. The 27-kDa fraction consisted of globular particles (data not shown). A survey view of the 53-kDa fraction is shown in Fig. 7a, revealing rod-shaped mole- cules, 22.5 ? 2.6 nm in length (n = 25) and with a mean diame- ter of 5.9 +- 0.8 nm (n = 39), compared with an average diameter of 4.8 nm for intact a-actinin. The overall rod-shaped structure was conserved, but the terminal hooks were missing, as ex- pected. In some cases, both polypeptides seemed twisted around each other (Fig. 7 b). Numerous rod-shaped molecules revealed large stain-filled gaps (Fig. 7c, arrowheads), possibly due to less compact packing. In contrast to the intact molecules (Fig. 5d), the paired configuration of internal globular domains is less ob- vious.
Different orientations of the molecule suggest a model on subdomain pairing. With the information obtained thus far on variability and putative flexibility of a-actinin particles, we at- tempted to resolve the discrepancy as to the pairing of spectrin- like repeats (see above). To this end we examined single par- ticles under different orientations at high magnifications. Fig. 8 shows such highly enlarged images, together with the corre- sponding interpretation of their molecular substructures. De- pending on their orientation during diffusive adsorption onto the carbon film, the molecules appear quite different (Fig. 8 a-d). Fig. 8 a shows a dimer displaying six discrete masses of different size. The terminal mass on the right appears roughly oval- shaped, the one on the left looks three-lobed. In accordance with their proposed identity with the actin-binding domains, they were termed ABD/ABD*. The central region of the molecule is composed of four density peaks, three of which are dumbbell- shaped consisting of two smaller masses each. The remaining globular mass (designated 1" in the scheme) appeared smaller, suggesting that it represents an unpaired globule on the rod. The molecule depicted in Fig. 8 b reveals a similar arrangement of subunits, with a threelobed ABD on the left and an oval-shaped ABD* on the right. Three roughly dumbbell-shaped density peaks are discernible in the central region. Fig. 8 c and d show projections with a similar overall morphology as seen in Fig. 8 a and b, except that the three central masses appear smaller, and not bipartite. The interpretation of the substructural appearance for all four molecules is given in Fig. 8, supporting the view that of the four spectrin-like repeats only three are forming pairs, while the unpaired globules may be very close to either the ABD andlor a pair composed of repeats 2 and 4*.
DISCUSSION
The overall morphology of a-actinin molecules, examined by negative staining and rotary shadowing, was generally de- scribed as rod-shaped, with a particle size of 30-40 nm (Pod- lubnaya et al., 1975; Suzuki et al., 1976; Condeelis and Vahey, 1982 ; Imamura et al., 1988). Further investigations demon- strated that these overall dimensions were not absolute but rather variable. Thus, the dimensions of skeletal muscle a-actinin were shown to change with the pH (Suzuki et al., 1976) and the ionic strength (Kuroda et al., 1994), and binding to actin filaments

198 Winkler et al. ( E m J. Biochem. 248)
b
C
Fig. 9. Structural models of smooth muscle a-actinin. (a) Model with three paired interactions between the spectrin-like repeats (1 -4 and 1"- 4*, respectively) as proposed by (Taylor and Taylor, 1993). (b) Alternative model with four paired repeats, according to Flood et al. 1995. (c) Model as deduced from our data, showing three paired repeats positioned in a slightly staggered arrangement. The N-terminal regions with the two presumptive actin-binding domains (ABD and ABD") are depicted as flexible hooks (c, -c3). The C-terminal calcium-binding motifs are not included in this scheme.
a
b
c
Fig. 10. Three-dimensional model of smooth-muscle a-actinin. PAC- tinin seen in a face-on view (a) and after a 60" (b) and 90" (c) rotation. Due to a rotation of the dimer around its longitudinal axis the two rows of spectrin-like repeats as shown in the face-on view (a) may approxi- mate (b) and finally fuse after a 90" rotation, to give an eclipsed pro- jection (c). The variability of the images of the terminal regions cannot he totally accounted for by different projections but must also reside in a high flexibility of these domains.
leads to shortening of a-actinin from Acanthamoeba and Dic- tyostelium (Meyer and Aebi, 1990). The data presented in this study show that the particle length of smooth muscle a-actinin can also vary with the ionic strength of the buffer (Figs 3 and 4). As a molecular explanation for this behaviour, Kuroda et al. (1994) proposed that the two polypeptide chains in the dimeric a-actinin molecule are twisted around each other, conferring a certain degree of elasticity to the central rod. Indeed, some of our pictures (Fig. 7 b) might support the view of a structure com- posed of two polypeptides twisted around each other. However, that the a-actinin molecules varied considerably in their width (Fig. 5 ) argues against this interpretation. The majority of the particle profiles shown in this study favour the assumption that smooth-muscle a-actinin consists of two series of discrete globu- lar masses comprising the central rod domain, with highly flexi- ble masses attached at both ends. As evident from molecule pro- jections shown in Figs 5 and 8, the appearance of the internal domains depends critically on the orientation of the particles and the individual staining conditions. The terminal regions, whose subdomains may adopt various orientations, can unfold to dif- ferent degrees and thus contribute primarily to the observed length differences described. It is conceivable that the shorter of a-actinin molecules seen in a-actinin-actin filament networks
(Meyer and Aebi, 1990) are caused by a conformational change in these terminal domains upon F-actin binding.
The organization of a-actinin in discrete subunits has also been proposed by Taylor and Taylor (1993). They showed that images obtained from negatively stained two-dimensional crys- tals, formed on positively charged lipid layers, were composed of 12 density peaks within one unit cell of a dimer. The topo- graphical arrangement of these 12 masses, as proposed by these authors, is depicted in Fig. 9 a. In this model, it is suggested that only three repeats of each chain pair with an equivalent mass in the opposite chain. In contrast, Flood et al., (1995) provided evidence that the eight spectrin-like repeats of an a-actinin dimer are aligned in four pairs, rather than three. This type of arrange- ment is shown Fig. 9 b. These authors used recombinant frag- ments of the a-actinin rod rather than the complete molecules for their analysis that was based on chemical cross-linking and analytical equilibrium sedimentation. Our images, as presented here, combine high-resolution electron microscopy with a speci- men preparation performed in aqueous solution. Thus, we avoided artifacts possibly induced by a two-dimensional crystal lattice, as analyzed by Taylor and Taylor (1993). From examin- ing the a-actinin particles at high magnifications in the ESI mode, we obtained insights into the substructural composition of a-actinin, in analogy to our studies on vinculin and talin per- formed with the same technique (Winkler et al., 1996, 1997). We provide evidence that the rod-like center of a dimer contains up to three pairs of globular masses arranged predominantly in a slightly staggered orientation. Thus, our data rather support the mode of pairing of the spectrin-like repeats as proposed in Fig. 9 a (Taylor and Taylor, 1993), but proposes a slight stagger of the globular masses as an additional parameter, as seen in Fig. 9c. In contrast, the pairing of all four repeats, as reported by Flood (1 995) might be explained by the finding that in their study, recombinant C-terminal fragments were used. Such head- less molecules might be prone to partial structural rearrange- ment, as is also indicated by our data showing a less compact packing of the 53-kDa proteolytic fragment.
The staggered arrangement of the repeats, together with the flexibility of the terminal domains (Fig. 9c, c , - ~ ) might allow for another interaction, given by the close proximity between repeats 1, 2, and 4*, as defined in Fig. 9c. Thus, one could envi- sion that repeat 4* interacts directly with repeat(s) 1 andlor 2. This view is supported by chemical cross-linking studies which revealed that the fourth repeat can be cross-linked to a polypep- tide consisting of the first and second (Imamura et al., 1988). An exceptional role for repeat 1 in the structural organization of a-actinin is supported by sequencing data, which show that this repeat is most highly conserved during evolution and less similar when compared with other repeats (Blanchard et al., 1989).

Winkler et al. (Eur: J. Biochem. 248) 199
Hence, the variability of the a-actinin images as seen in our study has probably two reasons. First, the molecules themselves are variable, due to the flexibility of the terminal masses and the possibility of multiple interactions between the globular spectrin-like repeats, as described above. Second, due to dif- ferent orientations on the grid, they may give rise to different projections, as is demonstrated in Fig. 10.
The flexibility and variability of a-actinin, giving the impres- sion of a breathing molecule may be important for the ability of this protein to form different types of networks with actin fila- ments (Jockusch and Isenberg, 1981, 1982; Meyer and Aebi, 1990; Korneeva and Jockusch, 1996). Three discrete suprastruc- tures were described : tight bundles, where a-actinin molecules are probably packed between actin filaments in parallel orienta- tion; ladder-like structures, where a-actinin is oriented perpen- dicular to the actin filaments; and a third structure with ob- liquely oriented a-actinin molecules, which may be a transient intermediate between the first and the second type (Meyer and Aebi, 1990). It is plausible that this switch from a perpendicular to a parallel orientation with respect to the actin filaments re- sides in the flexible N-terminal hooks. This flexibility, as de- scribed here, may indeed represent an essential structural pre- requisite for the three-dimensional variability of actin-a-actinin networks found in cells.
We express our appreciation to Eva Saxinger for exellent technical assistance. This work was financially supported by the Deutsche For- schungsgemeinschaft.
REFERENCES Baron, M. D., Davison, M. D., Jones, P. & Critchley, D. R. (1987) The
sequence of chick a-actinin reveals homologies to spectrin and cal- modulin, J. Biol. Chem. 262, 17 623- 17 629.
Bauer, J. (1988) Electron spectroscopic imaging : an advanced technique for imaging and analysis in transmission electron microscopy, Meth- ods Microbiol. 20, 113-146.
Blanchard, A,, Ohanian, V. & Critchley, D. (1989) The structure and function of a-actinin, J. Muscle Res. Cell Motil. 10, 280-289.
Bradford, M. M. (1976) A rapid and sensitive method for the quantifica- tion of microgram quantities of protein utilizing the principle of pro- tein-dye binding, Anal. Biochem. 72, 248-254.
Condeelis, J. & Vahey, M. (1982) A calcium and pH regulated protein from Dictyosteliurn discoideum that cross-links actin filaments, J. Cell. Bid. 94, 466-471.
Crawford, A. W., Michelsen, J. W. & Beckerle, M. C. (1992) An interac- tion between zyxin and a-actinin, J. Cell Biol. 116, 1381-1393.
Feramisco, J. R. & Burridge, K. (2980) A rapid purification of a-actinin, filamin and a 130000-dalton protein from smooth muscle, J. Biol. Chem. 255, 1194-1199.
Flood, G., Kahana, E., Gilmore, A. P., Rowe, A. J., Gratzer, W. B. & Critchley, D. R. (1995) Association of structural repeats in the a- actinin rod domain. Alignment of inter-subunit interactions, J. Mol. Bid. 252, 227-234.
Geiger, B., Dutton, A. H., Tokuyasu, K. T. & Singer, S. J. (1981) Immu- noelectron microscope studies of the membrane-microfilament in- teractions: distributions of a-actinin, tropomyosin, and vinculin in interstinal epithelial brush border and chicken gizzard smooth muscle cells, J. Cell Biol. 91, 614-628.
Imamura, M., Endo, T., Kuroda, M., Tanaka, T. & Masaki, T. (1988). Substructure and higher structure of chicken smooth muscle a-actinin molecule, J. Biol. Chem. 263, 7800-7805.
Jockusch, B. M. & Isenberg, G. (1981) Interaction of a-actinin and vin- culin with actin: opposite effects on filament network formation, Proc. Natl Acad. Sci. USA 78, 3005-3009.
Jockusch, B. M. & Isenberg, G. (1982) Vinculin and a-actinin: interac- tion with actin and effect on microfilament network formation, Cold Spring Harb. Symp. Quant. Biol. 46, 613-623.
Korneeva, N. L. & Jockusch, B. M. (1996) Light microscopy analysis of ligand-induced actin filament suprastructures, Eul: J. Cell Biol. 71, 351-355.
Kroemker, M., Rudiger, A. H., Jockusch, B. M. & Rudiger, M. (1994) Intramolecular interactions in vinculin control a-actinin binding to the vinculin head, FEBS Lett. 355, 259-262.
Kuroda, M., Kohira, Y. & Sasaki, M. (1994) Conformational changes of skeletal muscle a-actinin induced by salt, Biochim. Biophys. Actu.
Laemmli, U. K. (1970) Cleavage of structural proteins during the assem- bly of the head of bacteriophages T4, Nature 227, 680-685.
Lazarides, E. & Burridge, K. (1975) a-actinin: immunofluorescent local- ization of a muscle structural protein in nonmuscle cells, Cell 6, 289-298.
Lazarides, E. & Granger, B. L. (1978) Fluorescent localization of mem- brane sites in glycerinated chicken skeletal muscle fibers and the relationship of these sites to the protein composition of the Z disc, Proc. Natl Acad. Sci. USA 75, 3683-3687.
Lunsdorf, H. & Spiess, E. (1986) A rapid method for preparing perfo- rated supporting foils for the thin carbon films used in high resolution transmission electron microscopy, J. Microsc. (Ox$) 144, 21 1-213.
Masaki, T., Endo, M. & Ebashi, S. (1967) Location of 6s component of a-actinin in Z-band, J. Biochem. (Tokyo) 62, 630-632.
Mayer, F., Rohde, M., Salzmann, M., Jussofie, A. & Gottschalk, G. (1988) The methanoreductosome: a high-molecular-weight enzyme complex in the methanogenic bacterium strain GO1 that contains components of the methylreductase system, J. Bacteriol. 170, 1438- 1444.
McGregor, A,, Blanchard, A. D., Rowe, A. J. & Critchley, D. R. (1994) Identification of the vinculin-binding site in the cytoskeletal protein a-actinin, J. Biochem. (Tokyo) 301, 225-233.
Meyer, R. K. & Aebi, U. (1990) Bundeling of actin filaments by (1-
actinin depends on its molecular length, J. Cell Bid. 110, 2013- 2024.
Noegel, A., Witke, W. & Schleicher, M. (1987) Calcium sensitive non- muscle a-actinin contains EF-hand structures and highly conserved regions, FEBS Lett. 221, 391 -396.
Otey, C. A,, Pavalko, F. M. & Burridge, K. (1990) An interaction be- tween a-actinin and the beta 1 integrin subunit in vitro, J. Cell Biol.
Podlubnaya, Z. A,, Tskhovrebova, L. A,, Zaalishvili, M. M. & Stefa- nenko, G. A. (1975) Electron microscopic study of a-actinin, J. Mol. Biol. 92, 357-359.
Sanger, J. W., Sanger, J. M. & Jockusch, B. M. (1983) Differences in stress fibers between fibroblasts and epithelial cells, J. Cell Biol. 96,
Small, J. V. (1985) Geometry of actin-membrane attachments in the smooth muscle cell: the localization of vinculin and a-actinin, EMBO
Suzuki, A,, Goll, D. E., Singh, I., Allen, R. E., Robson, R. M. & Stromer, M. H. (1976). Some properties of purified skeletal muscle a-actinin, J. Biol. Chem. 251, 6860-6870.
Taylor, K. A. & Taylor, D. W. (1993) Projection image of smooth muscle a-actinin from two-dimensional crystals formed on positively charged lipid layers, J. Mol. Bid. 230, 196-205.
Valentine, R. C., Shapiro, B. M. & Stadtman, E. R. (1968) Regulation of glutamine synthetase. XII. Electron microscopy of the enzyme from Escherichia coli, Biochemistry 7 , 2143 -2152.
Wallraff, E., Schleicher, M., Modersitzki, M., Rieger, D., Isenberg, G. & Gerisch, G. (1986) Selection of Dictyosteliurn mutants defective in cytoskeletal proteins: use of an antibody that binds to the ends of a- actinin rods, EMBO J. 5, 61 -67.
Winkler, J., Lunsdorf, H. & Jockusch, B. M. (1996) The ultrastructure of chicken gizzard vinculin as visualized by high-resolution electron microscopy, J. Struct. Biol. 116, 270-277.
Winkler, J., Lunsdorf, H. & Jockusch, B. M. (1997) Energy-filtered electron microscopy revealed that talin is a highly flexible protein composed of a series of globular domains, Eur: J. Biochem. 243, 430-436.
Yamaguchi, M., Izumimoto, M., Robson, R. M. & Stromer, M. H. (1985) Fine structure of wide and narrow vertebrate 2-lines. A proposed model and computer simulation of Z-lines architecture, J. Mol. Biol. 184, 621-644.
1205, 97-104.
111, 721 -729.
961 -969.
J. 4, 45-49.