fisiología y respuestas de comportamiento de escarabajos ... · más favorables, siendo el aumento...
TRANSCRIPT

Susana Pallarés Párraga
Dpto. de Ecología e Hidrología
Facultad de Biología
Directores: Josefa Velasco y Andrés Millán
TESINA DE LICENCIATURA
Fisiología y respuestas de comportamiento de
escarabajos acuáticos salinos frente al estrés térmico y
salino

i
Índice
Resumen ........................................................................................................................................ ii
1. Introducción .............................................................................................................................. 1
1.1 Justificación e interés del estudio ........................................................................................ 1
1.2 Objetivos .............................................................................................................................. 3
1.3 Hipótesis y predicciones de partida ..................................................................................... 4
2. Material y métodos ................................................................................................................... 5
2.1. Especies estudiadas ............................................................................................................ 5
2.2. Diseño experimental ........................................................................................................... 8
3. Resultados ............................................................................................................................... 12
3.1. Correlación entre la mortalidad y las respuestas de comportamiento ............................ 12
3.2. Efecto de la temperatura y la conductividad en las variables respuesta ......................... 12
3.3. Tolerancia al estrés: mortalidad ....................................................................................... 14
3.4. Respuestas de comportamiento de huida para evitar la exposición al estrés ................. 17
Emersión .................................................................................................................................. 17
Vuelo........................................................................................................................................ 20
4. Discusión ................................................................................................................................. 23
5. Conclusiones............................................................................................................................ 29
6. Referencias .............................................................................................................................. 31
Agradecimientos ......................................................................................................................... 38

ii
Resumen
El presente trabajo compara la tolerancia fisiológica y las respuestas de comportamiento frente al
estrés térmico y salino de tres pares de especies congenéricas de coleópteros acuáticos salinos
(Enochrus falcarius y E. bicolor, Nebrioporus baeticus y N. ceresyi, Ochthebius glaber y O. notabilis) con el
objetivo de explicar sus diferentes patrones de ocupación de hábitat y rango geográfico y su sensibilidad
a cambios en las condiciones de sus hábitats. Para ello se realizaron experimentos de laboratorio
sometiendo a individuos de cada especie a diferentes tratamientos combinados de conductividad y
temperatura del agua. Durante 30 minutos de exposición se midió la tolerancia fisiológica (mortalidad)
así como respuestas de comportamiento (emersión y vuelo) para evitar las condiciones estresantes. Los
resultados mostraron que el shock térmico agudo tiene un efecto más importante e inmediato sobre la
fisiología y el comportamiento de huida de los individuos que el estrés salino o la interacción de ambos
factores. La emersión y el vuelo fueron dependientes de la tolerancia al estrés de las especies.
Temperaturas entre 35-40ºC fueron críticas para las respuestas de comportamiento analizadas, siendo
estos valores los umbrales de máxima respuesta de vuelo o emersión en la mayoría de las especies
estudiadas. Temperaturas de 45ºC produjeron un daño fisiológico importante sobre los individuos,
resultando letales para las especies de Enochrus y Nebrioporus. Sin embargo, las especies de Ochthebius
resultaron más resistentes a dicha temperatura. En dos de los tres pares de especies estudiados
(Nebrioporus y Ochthebius) las especies que ocupan cuerpos de agua lóticos (N. baeticus y O.glaber)
fueron menos tolerantes al estrés térmico y en niveles críticos de estrés sus respuestas de emersión y
vuelo fueron de mayor magnitud que sus especies hermanas de medios leníticos (N. ceresyi y O.
notalibis), mostrando una mayor sensibilidad frente al aumento de temperatura. Dichos resultados
concuerdan con lo esperado, ya que los medios lóticos, en general y en un contexto evolutivo, sufren
menores fluctuaciones ambientales que los medios de aguas estancadas, más inestables. Sin embargo,
las dos especies de Enochrus presentaron similar tolerancia al estrés y respuesta de emersión, siendo E.
bicolor, la especie lenítica, la que mostró una respuesta de vuelo mayor a temperaturas críticas
subletales. En este caso, diferencias en la capacidad de dispersión, claramente menor en E. falcarius,
parecen determinar el distinto patrón biogeográfico de estas especies. Nuestro estudio, por tanto,
demuestra que, en general, las especies lóticas, especialmente las que tienen una capacidad de vuelo
reducida, son más sensibles que las leníticas frente al previsible aumento de temperaturas como
consecuencia del cambio climático.
Palabras clave: tolerancia fisiológica, respuestas de comportamiento, estrés térmico, estrés salino,
umbrales críticos de estrés, coleópteros acuáticos salinos, estabilidad del hábitat, cambio climático.

1
1. Introducción
1.1 Justificación e interés del estudio
El estudio de la fisiología de las especies constituye una herramienta muy útil
para algunas disciplinas generales como la ecología, biogeografía y biología evolutiva
(Spicer y Gaston 1999). Para entender los procesos pasados y presentes que actúan
sobre las especies es fundamental conocer cómo responden los organismos a la
variabilidad de su entorno (Chown 2001). Los rasgos fisiológicos definen el nicho de las
especies, delimitan sus rangos geográficos (e.g. Hoffmann y Blows 1994, Gaston 2003,
2009) y, además, juegan un papel esencial en aspectos básicos de su ecología, como
por ejemplo la ocupación de hábitat (Tomanek y Somero 1999, Stillman 2003). Así
mismo, en las últimas décadas ha surgido una urgente necesidad de predecir los
efectos potenciales de la alteración de los hábitats derivados del cambio climático y de
las diversas presiones humanas sobre las especies y poblaciones, dando un nuevo
ímpetus a la fisiología ecológica (e.g. Deutsch et al. 2008, Tewksbury et al. 2008,
Gaston et al. 2009, Helmuth 2009). En este contexto han surgido numerosos estudios
que examinan los efectos de factores estresantes sobre la fisiología de los organismos
con fines conservacionistas (e.g. Davis y Olla 2001, Homan et al. 2003, Swanson et al.
2000, Sardella et al. 2008b, Sánchez-Fernández et al. 2010).
Junto a los rasgos fisiológicos, la capacidad de las especies para adaptarse a los
cambios ambientales depende de ajustes del comportamiento que pueden modificar
sustancialmente las condiciones ambientales experimentadas por los individuos (Huey
1991, Marais y Chown 2008, Angilletta 2009). Por ejemplo, las especies pueden
adaptarse a cambios en las condiciones climáticas desplazándose hacia otras áreas
(Thuiller et al. 2005, Massot et al. 2008) o moviéndose dentro de sus actuales hábitats
en busca de condiciones microclimáticas favorables para mantener un ambiente
térmico constante (Feder 2010). En el contexto de la actual crisis de la biodiversidad y
en particular del cambio global, resulta especialmente necesario disponer de un mejor
conocimiento de este tipo de respuestas (Bozinovic et al. 2011), ya que determinan en
gran medida la supervivencia y persistencia de las poblaciones locales. Sin embargo,

2
hasta ahora pocos estudios han integrado medidas de tolerancia fisiológica y de
estrategias de comportamiento de organismos ectotermos en relación con el estrés
ambiental (e.g. Hazell et al. 2010).
Entre las múltiples dimensiones del nicho de las especies, la temperatura es
reconocida como uno de los factores más importantes ya que determina la actividad
metabólica y las historias de vida de las especies (Willott y Hassall 1998). Los efectos
del estrés térmico en insectos y las respuestas fisiológicas, morfológicas y de
comportamiento que los individuos emplean para contrarrestar estos efectos han sido
aspectos centrales en fisiología y ecología (e.g Bale 2002, Hoffmann et al. 2003, Sinclair
et al. 2003, Chown y Nicolson 2004, Korsloot et al. 2004). Sin embargo, la mayoría de
los estudios se han centrado en determinar los límites térmicos letales y subletales de
las especies o su tolerancia a temperaturas extremas (e.g Kay y Whitford 1978,
Pandolfo et al. 2010, Hughes et al. 2010, Chidawanyika y Terblanche 2011), dedicando
menos atención a otras respuestas subletales de comportamiento críticas para hacer
frente a los cambios ambientales. Por otro lado, los organismos no experimentan
variación en un solo factor abiótico independientemente, sino que en la naturaleza es
frecuente que múltiples factores cambien simultáneamente (Terblanche et al. 2011).
De este modo, es especialmente interesante el desarrollo de aproximaciones
multifactoriales que combinen la temperatura con otros factores ambientales de
estrés para evaluar el efecto de su interacción sobre las respuestas de los organismos.
Los medios salinos continentales mediterráneos son un interesante modelo para el
estudio de respuestas fisiológicas y de comportamiento frente a múltiples factores de
estrés. En estos ambientes, la combinación de altos niveles de salinidad y temperatura
es frecuente, actuando de forma aditiva, sinérgica o incluso antagónica sobre los
individuos que los ocupan. Estas condiciones tan extremas constituyen una barrera
infranqueable para la mayoría de especies acuáticas; solo las especies más
especializadas son capaces de colonizar estos medios (Millán et al. 2011). Estudios
recientes de tolerancia térmica en escarabajos acuáticos salinos han mostrado que la
salinidad influye en su fisiología térmica (e.g. Sánchez Fernández et al. 2010), al igual
que ocurre en otros taxones marinos (e.g. Kir y Kumlu 2008, Rajagopal et al. 2005,
Sardella et al. 2008a, b). Por lo tanto, es probable que las respuestas fisiológicas y de

3
comportamiento de las especies para compensar el estrés térmico y salino en estos
medios sean especialmente complejas y singulares, aunque hasta ahora han sido poco
estudiadas.
Entre los macroinvertebrados, los coleópteros, junto a los dípteros y hemípteros, son
los órdenes más comunes y ricos en especies en las aguas salinas continentales. En
especies taxonómicamente relacionadas de escarabajos salinos ibéricos se ha
encontrado una correlación entre el tamaño del rango geográfico y el tipo de hábitat
ocupado. Así, las especies que ocupan los medios leníticos (aguas estancas), sistemas
más inestables desde un punto de vista ecológico y geológico, presentan rangos
geográficos más amplios que especies congenéricas que viven en medios lóticos (aguas
corrientes) (e.g Ribera y Vogler 2000, Hof et al. 2006, Abellán et al. 2009, Damm et al.
2010, Arribas et al. en prensa). La tolerancia fisiológica, como un componente de la
capacidad de colonización de las especies, ha sido identificada como uno de los rasgos
fundamentales que determina este patrón. Debido a que los cuerpos de agua someros
leníticos están sujetos a mayores fluctuaciones térmicas diarias y estacionales que los
ríos o arroyos (Álvarez-Cobelas et al. 2005, Florencio et al. 2009), la selección para
resistir estas fluctuaciones locales ha podido resultar en una amplia tolerancia térmica
de las especies leníticas, permitiéndoles establecerse en un rango mayor de
localidades en comparación con las especies lóticas. Así, la fauna de los medios
acuáticos salinos constituye un grupo ideal para comparar respuestas frente al estrés
entre especies relacionadas que ocupan hábitats con diferente estabilidad ambiental.
1.2 Objetivos
El objetivo general del presente estudio es comparar las respuestas de
comportamiento frente al estrés térmico y salino en tres pares de especies
congenéricas de escarabajos acuáticos salinos, con diferente tamaño de sus rangos
geográficos y contrastada ocupación de hábitat, en relación con su tolerancia
fisiológica. Con los términos “respuestas de comportamiento” se hace referencia al
conjunto de estrategias, condicionadas por la tolerancia fisiológica, que los organismos
emplean frente a condiciones desfavorables o cambiantes en sus medios para evitar la
exposición al estrés ambiental, con el fin de mantener un estatus térmico y osmótico

4
óptimo. En concreto este estudio se centra en dos respuestas: vuelo y emersión del
agua (comportamiento de huida). La mayoría de especies de coleópteros en fase
adulta muestran capacidad para volar, lo cual les permite escapar de sus hábitats
cuando las condiciones se vuelven fisiológicamente subóptimas, en busca de hábitats
más favorables, siendo el aumento de la temperatura un factor clave del inicio de la
dispersión (Zalom et al. 1980, Velasco y Millán 1998). Por otro lado, los individuos
adultos acuáticos pueden emerger del agua cuando las condiciones del medio varían,
reduciendo el estrés al que están expuestos. Además la emersión en coleópteros
voladores es un paso previo necesario para el vuelo, ya que para desplegar las alas los
élitros deben estar secos.
El presente estudio pretende dar respuesta a las siguientes cuestiones: 1) ¿Afecta la
combinación de estrés térmico y salino a la tolerancia y el comportamiento de huida
escarabajos acuáticos salinos? 2) ¿Dependen las respuestas de comportamiento de
huida de la tolerancia fisiológica de las especies? 3) ¿Son las especies lóticas más
sensibles al estrés que las especies leníticas? 4) ¿Qué especies dentro de cada género
estudiado podría ser más susceptible a cambios en las condiciones de sus hábitats en
base a su fisiología y comportamiento frente al estrés ambiental?
1.3 Hipótesis y predicciones de partida
En relación a la cuestión 1, predecimos que la combinación de elevadas
temperaturas y salinidades disminuirá la capacidad de los individuos estudiados de
desarrollar respuestas de escape por el daño fisiológico acumulado, reduciendo la
supervivencia. En cuanto a las cuestiones 2 y 3, la predicción general es que las
respuestas de comportamiento de escape o huida están condicionadas por la
tolerancia fisiológica de las especies, y en base a ello se espera que dentro de cada par
de especies estudiadas, la especie menos tolerante al estrés muestre respuestas de
comportamiento críticas para su supervivencia (emersión y vuelo) en niveles de estrés
más bajos que las especies más tolerantes. Debido a que en general los arroyos salinos
son ecológicamente más estables que los humedales salinos, es esperable un patrón
de respuesta diferente entre especies lóticas y leníticas. Así, se prevé que las especies
lóticas sean más sensibles al estrés ambiental que las leníticas, ya que están adaptadas

5
a un menor rango de variación de las condiciones ambientales, y por ello sus
respuestas de escape se producirán en niveles de estrés térmico y salino más bajos que
las leníticas y/o serán de mayor magnitud. Por último, en relación a la cuestión 4, de
acuerdo con el planteamiento anterior, las especies lóticas podrían ser más
vulnerables frente a cambios ambientales que las leníticas.
2. Material y métodos
2.1. Especies estudiadas
Entre los coleópteros acuáticos salinos, las familias más representativas son
Hydraenidae, Hydrophilidae y Dytiscidae, pertenecientes a dos sub-órdenes
(Polyphaga las dos primeras y Adephaga la última). El presente estudio se centra en
tres pares de especies congenéricas de coleópteros típicos de medios acuáticos
mesosalinos e hipersalinos continentales, con contrastada ocupación de hábitat y
tamaño del rango geográfico. Estas especies pertenecen a tres géneros: Nebrioporus
(N. ceresyi (Aubé 1836) y N. baeticus (Schaum 1864), familia Dytiscidae), Enochrus (E.
bicolor (Fabricius 1792) y E. falcarius Hebauer 1991, familia Hydrophilidae) y
Ochthebius (O. notabilis Rosenhauer 1856 y O. glaber Montes & Soler 1988, familia
Hydraenidae).
Nebrioporus es un género de escarabajos acuáticos relativamente rico en especies, la
mayoría de ellas distribuidas por la región Paleártica, aunque el género también está
presente en zonas de África sub-sahariana (Toledo 2009). Algunas especies ocupan
arroyos y lagunas salinos (grupo N. ceresyi). N. ceresyi es una especie de distribución
circum-mediterránea que habita aguas estancas como humedales y pozas,
particularmente localizados en cotas bajas cerca de la costa (Fery et al. 1996).
Excepcionalmente ocupa zonas salinas de interior de origen marino. N. baeticus es
endémica del sureste de España y ocupa arroyos hipersalinos, en general alejados de la
costa (Fig. 1).
Enochrus es un amplio género de escarabajos acuáticos, con 189 especies distribuidas
en todas las regiones biogeográficas. Las especies mediterráneas de ambientes salinos
pertenecen al complejo Enochus bicolor (Schödl 1998). E. bicolor, la especie que da

6
nombre al complejo, ocupa cuerpos de agua salinos leníticos (humedales y pozas) en
Europa, Norte de África, este de Asia y Mongolia (Hansen 2004). Su especie
congenérica, E. falcarius, presenta una distribución más restringida, ocupando arroyos
salinos en el sur de la Península Ibérica, Túnez, Sicilia (Hansen 2004) y Marruecos (Fig.
1). Un estudio reciente ha revelado que esta especie tal y como se entiende
actualmente, realmente incluye un complejo de diferentes linajes, cada uno con
distribuciones disyuntas y restringidas a lo largo del área mediterránea (Arribas et al.
en prensa). En nuestro caso trabajamos con el linaje ibérico (“E. falcarius IP” según
Arribas et al., en adelante E. falcarius para simplificar).
Por último, Ochthebius es uno de los géneros más diversos de la familia Hydraenidae
(Jäch et al. 2005), con 66 especies citadas en la Península Ibérica. O. notabilis se
encuentra en la Península Ibérica, Islas Canarias y Norte de África (Fig. 1), habitando
lagunas salinas y cubetas salineras, mientras que O. glaber es endémico del sur de la
Península Ibérica, ocupando principalmente los arroyos hipersalinos (Abellán et al.
2009).

7
Figura 1. Distribución geográfica aproximada de los tres pares de especies estudiadas.

8
2.2. Diseño experimental
Para evaluar las respuestas de comportamiento (emersión y vuelo) y la tolerancia
al estrés térmico y salino en los tres pares de especies seleccionados se desarrolló un
diseño experimental en laboratorio con tres factores fijos (temperatura, conductividad
y especie).
En primer lugar se recolectaron aproximadamente 400 individuos de cada especie en
diferentes localidades (una localidad por especie) en el sureste ibérico (Tabla 1). Los
individuos se mantuvieron durante una semana en laboratorio, dentro de acuarios que
contenían agua filtrada procedente de los sitios de recolección con aireación artificial,
alimentándoles periódicamente (larvas de quironómidos para las especies
depredadoras - Nebrioporus-, Ruppia marítima para las especies herbívoras -Enochrus-
y biofilm para las especies de Ochthebius). Tras la semana de mantenimiento, los
individuos fueron aclimatados durante 24 h en una cámara climática (SANYO MLR-351)
en condiciones constantes de temperatura (20ºC), fotoperiodo (12h luz: 12h
oscuridad) e intensidad de luz de 15 μmol.m-2.s-1, sin proporcionarles alimento durante
este periodo.
Para llevar a cabo los experimentos, grupos de 10 individuos de cada especie
seleccionados aleatoriamente fueron sometidos a 12 tratamientos combinados de
conductividad y temperatura en el caso de las especies de Enochrus y Nebrioporus y 20
tratamientos para las especies de Ochthebius (Fig. 2). Los niveles de conductividad
testados se establecieron de acuerdo con el rango natural de salinidad en el que se
encuentran las especies en los hábitats que ocupan (Velasco et al. 2006): 20, 50, 80
mS.cm-1 para las especies de Enochrus y Nebrioporus y 20, 50, 80, 180, 240 mS.cm-1
para las especies de Ochthebius. Las disoluciones correspondientes a cada tratamiento
se prepararon disolviendo sal marina (Ocean Fish, Prodac®) en agua destilada. Los
niveles de temperatura testados son valores representativos de un gradiente que
abarca desde temperaturas habituales en los hábitats de las especies (20 y 35ºC) hasta
temperaturas extremas (40 y 45ºC) próximas a los límites térmicos subletales y letales
registrados para estas especies en estudios previos (Sánchez-Fernández et al. 2010,

9
Arribas et al. en prensa). Cada tratamiento combinado de temperatura y salinidad fue
replicado 3 veces.
Los acuarios experimentales se introdujeron en un baño húmedo con temperatura
controlada (Fig. 3) para mantener unas condiciones térmicas constantes durante el
experimento (±1ºC). Cada acuario contenía 100 ml de la correspondiente disolución y
una piedra artificial de cemento parcialmente emergida del agua (Fig. 3) cuya función
era facilitar a los individuos la salida del medio (emersión) y el vuelo para evitar las
condiciones de estrés. Cada conjunto de 10 individuos se extrajo de los acuarios de
aclimatación e inmediatamente se expuso al tratamiento asignado durante 30 min, es
decir, se trata de experimentos de choque térmico y salino de exposición corta. En este
periodo se registraron las respuestas de comportamiento de huida (emersión y vuelo)
y la mortalidad. Los individuos que volaron durante el experimento se contaron y
retiraron para que no interfirieran en el tratamiento; el vuelo se expresó como número
total de vuelos registrados en el tratamiento. Cada 2 min se registró el número de
individuos que se encontraba sobre la piedra, expresando la emersión como el número
medio de individuos que emergieron del agua durante el tiempo experimental. La
mortalidad, como medida de la tolerancia al estrés, se expresó como el número de
individuos muertos contados tras los 30 minutos de exposición.

10
Tabla 1. Rango de conductividad natural de los hábitats de las especies estudiadas y localidades de
recolección (posición geográfica y conductividad media).
Especies
Rango de
conductividad
(mS.cm-1
)
Localidad de muestreo Latitud Longitud
Conductividad
media
(mS.cm-1
)
N. ceresyi 2 - 190 Laguna Cotorrillo, Salinas
de San Pedro, Murcia
37.82516 -0.76196 60
N. baeticus 2 - 185 Río Chícamo, Murcia 38.21753 -1.05113 19
E. bicolor 4 - 90 Laguna del Mojón Blanco,
Albacete
38.47530 -1.25582 65
E. falcarius 7 - 160 Rambla Salada de Fortuna,
Murcia
38.16993 -1.12565 70
O. notabilis 50 - 220 Estrecho de la Salineta,
Alicante
38.43459 -0.78006 140
O. glaber 20 - 250 Rambla salada de Librilla,
Murcia
37.90656 -1.37102 180
Figura 2. Tratamientos combinados de temperatura y conductividad a los que fueron sometidos los
individuos.
20ºC 35ºC
40ºC 45ºC
20ºC 35ºC
40ºC 45ºC

11
Figura 3. Acuarios de mantenimiento (arriba izq.), baño húmedo con temperatura controlada
(abajo izq.) y acuarios experimentales (dcha.). Las flechas señalan individuos emergidos sobre la
piedra durante el experimento.
2.3. Análisis de datos
Dado que a priori las respuestas analizadas en los experimentos no son
independientes unas de otras (una de las predicciones de partida es que las respuestas
de comportamiento están en gran medida condicionadas por la tolerancia fisiológica),
se testó la relación entre las tres variables dependientes estimadas (emersión, vuelo y
mortalidad) mediante el cálculo de los coeficientes de correlación de Pearson para
cada par de especies. Tras comprobar la relación entre las variables, se realizó para
cada par de especies un MANOVA Multifactorial con la temperatura, conductividad y
especie como factores fijos. Se empleó el test multivariado de la lambda de Wilks para
estimar el efecto global de los factores y las interacciones entre factores en las
variables respuesta. Para determinar los efectos de cada factor e interacciones sobre
cada variable dependiente por separado se realizaron test univariados (ANOVA).

12
Se testó la homocedasticidad y normalidad de los datos y la homogeneidad de las
covarianzas. Sin embargo, debido a que algunos tratamientos no cumplieron estos
requisitos se analizó también el cumplimiento de los mismos con los residuos no
tipificados (Underwood 1997, Rutherford 2001). Para detectar diferencias significativas
en las variables respuesta entre los distintos tratamientos se realizaron análisis post-
hoc, aplicando el test de Bonferroni, más conservativo, para compensar el
incumplimiento de algunos de los requisitos del análisis. Todos los análisis estadísticos
se realizaron utilizando SPSS para Windows, Rel. 15.0.1. 2006. Chicago: SPSS Inc.
3. Resultados
3.1. Correlación entre la mortalidad y las respuestas de comportamiento
La emersión y la mortalidad estuvieron significativamente correlacionadas en
los tres pares de especies estudiadas. Esta relación fue negativa en las especies de
Enochrus (r= -0.386, p = 0.001) y Nebrioporus (r = -0.305, p = 0.009) y positiva en las
especies de Ochthebius (r = 0.253, p = 0.005). El vuelo y la mortalidad también
mostraron una correlación negativa en las especies de Enochrus (r = - 0.243, p = 0.040)
y Nebrioporus (r = -0.298, p = 0.011), aunque no en las especies de Ochthebius.
Finalmente, ambas respuestas de comportamiento (emersión y vuelo) tuvieron una
relación positiva en los tres pares de especies (r = 0.289, p = 0.014 en Enochrus; r =
0.440, p < 0.001 en Nebrioporus y r = 0.328, p < 0.001 en Ochthebius).
3.2. Efecto de la temperatura y la conductividad en las variables respuesta
Los test multivariantes de la lambda de Wilks mostraron que la temperatura
tuvo un efecto global significativo sobre las variables dependientes en los tres pares de
especies estudiadas (Tabla 2). Los test univariantes revelaron resultados similares (ver
Tabla 3 para mortalidad, Tabla 4 para emersión y Tabla 5 para vuelo).
En cambio, el efecto global de la conductividad y de la interacción conductividad x
temperatura solo fue significativo para las especies de Ochthebius (Tabla 2) en todas

13
las variables respuesta (Tablas 3, 4 y 5), aunque los test univariados (ANOVA)
mostraron que el efecto de estos factores fue diferente en O. glaber y O. notabilis.
Tabla 2. Resultados del MANOVA. Test de la lambda de Wilks del efecto global de la temperatura
y la conductividad sobre las variables dependientes en los tres pares de especies estudiadas.
Fuente Lambda de Wilks F Σ2 parcial p
Enochrus
Temperatura 0.003 119.425 0.853 ***
Conductividad 0.950 0.397 0.025 n.s
Especie 0.448 18.879 0.552 ***
Temperatura x Conductividad 0.768 0.708 0.084 n.s
Temperatura x Especie 0.532 3.686 0.190 ***
Conductividad x Especie 0.900 0.827 0.051 n.s
Temperatura x Conductividad x Especie
0.854 0.415 0.051 n.s
Nebrioporus
Temperatura 0.002 153.982 0.878 ***
Conductividad 0.916 0.690 0.043 n.s
Especie 0.447 16.788 0.523 ***
Temperatura x Conductividad 0.806 0.574 0.069 n.s
Temperatura x Especie 0.264 9.084 0.359 ***
Conductividad x Especie 0.948 0.419 0.027 n.s
Temperatura x Conductividad x Especie
0.657 1.162 0.131 n.s
Ochthebius
Temperatura 0.078 67.834 0.720 ***
Conductividad 0.499 8.203 0.293 ***
Especie 0.541 33.452 0.459 ***
Temperatura x Conductividad 0.472 2.995 0.313 ***
Temperatura x Especie 0.636 6.694 0.203 ***
Conductividad x Especie 0.720 3.528 0.152 ***
Temperatura x Conductividad x Especie 0.491 2.812 0.299 ***
* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, n.s no significativo

14
3.3. Tolerancia al estrés: mortalidad
Las dos especies de Enochrus mostraron una tolerancia similar al estrés térmico
agudo (Tabla 3). La supervivencia de E. bicolor y E. falcarius fue alta en todos los
niveles de temperatura testados excepto en el nivel más extremo, 45ºC, que resultó
letal para prácticamente todos los individuos expuestos (Fig. 4a).
Las especies de Nebrioporus fueron menos resistentes al estrés térmico; el incremento
de temperatura redujo la supervivencia de ambas especies y a 45ºC murieron la
mayoría de individuos (Fig. 4b). La mortalidad de N. baeticus fue significativamente
mayor que la de N. ceresyi (Tabla 3), siendo esta diferencia especialmente notable en
los tratamientos de 40ºC (post-hoc Temperatura x Especie 40ºC: p < 0.001; Fig. 4b).
Las especies de Ochthebius presentaron diferente tolerancia a ambos factores
estresantes, siendo O. glaber en general menos tolerante que O. notabilis (Tabla 3, Fig.
4c,d). La mortalidad de O. glaber aumentó progresivamente con la temperatura,
siendo moderadamente alta a 40ºC y alcanzando el valor máximo a 45ºC. Dentro de
los tratamientos de temperaturas más elevadas (40 y 45ºC), la mortalidad de esta
especie fue significativamente mayor en el nivel de conductividad más extremo (240
mS.cm-1) (Fig. 4c). En cambio, la mortalidad de O. notabilis fue muy baja o nula en las
temperaturas de 20, 35 y 40ºC; sólo el nivel de estrés térmico más extremo (45ºC)
redujo significativamente la supervivencia de esta especie (Fig. 4d). Para esta especie
no se observaron diferencias significativas en la mortalidad entre los diferentes niveles
de conductividad (p = 0.238).

15
Tabla 3. Resultados del ANOVA. Efecto de la temperatura y la conductividad en la
mortalidad en los tres pares de especies estudiadas.
Variable dependiente: Mortalidad
Fuente SS g.l. F p
Enochrus
Modelo corregido 918.000 23 79.826 ***
Intersección 338.000 1 676.000 ***
Temperatura 912.778 3 608.519 ***
Conductividad 0.583 2 0.583 n.s
Especie 0.056 1 0.111 n.s
Temperatura x Conductividad 2.639 6 0.880 n.s
Temperatura x Especie 0.500 3 0.333 n.s
Conductividad x Especie 0.528 2 0.528 n.s
Temperatura x Conductividad x
Especie 0.917 6 0.306 n.s
Error
24.000 48
Nebrioporus
Modelo corregido 1229.944 23 110.007 ***
Intersección 786.722 1 1618.400 ***
Temperatura 1168.056 3 800.952 ***
Conductividad 0.361 2 0.371 n.s
Especie 12.500 1 25.714 ***
Temperatura x Conductividad 0.194 6 0.067 n.s
Temperatura x Especie 44.500 3 30.514 ***
Conductividad x Especie 0.250 2 0.257 n.s
Temperatura x Conductividad x
Especie
4.083 6 1.400 n.s
Error
23.333 48
Ochthebius
Modelo corregido 612.125 39 17.439 ***
Intersección 316.875 1 352.083 ***
Temperatura 386.425 3 143.120 ***
Conductividad 34.583 4 9.606 ***
Especie 54.675 1 60.750 ***
Temperatura x Conductividad 37.617 12 3.483 ***
Temperatura x Especie 38.625 3 14.306 ***
Conductividad x Especie 23.950 4 6.653 ***
Temperatura x Conductividad x
Especie
36.250 12 3.356 ***
Error 72.000 80
SS, suma de cuadrados; g.l, grados de libertad.
* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, n.s no significativo

16
Figura 4. Mortalidad media ± ES de cada especie. Las diferencias significativas determinadas por los
análisis post-hoc con la corrección de Bonferroni se indican como sigue: con letras mayúsculas
junto a la leyenda para los niveles de conductividad, con letras minúsculas sobre las barras para los
niveles de temperatura, con números sobre las barras para los niveles de conductividad dentro de
un mismo nivel de temperatura y con asteriscos para diferencias entre especies congenéricas
dentro de un mismo tratamiento.

17
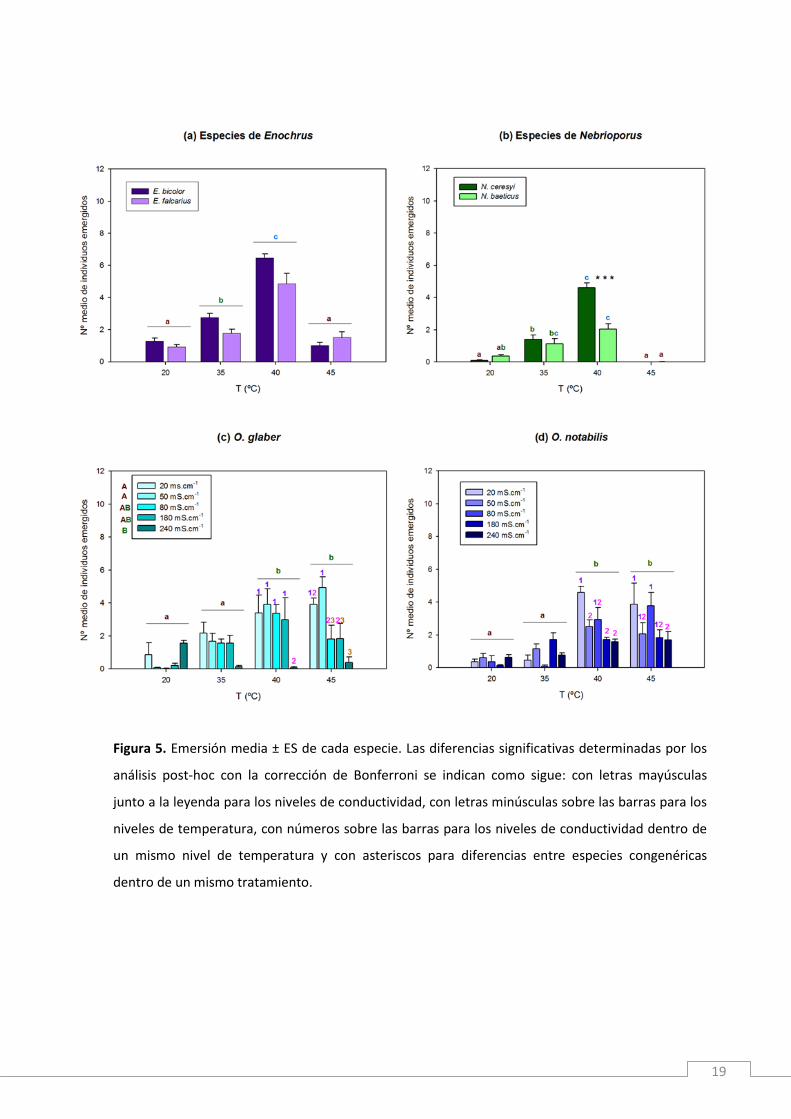
3.4. Respuestas de comportamiento de huida para evitar la exposición al estrés
Emersión
Las especies de Enochrus mostraron un patrón de respuesta de emersión
similar frente al estrés térmico, aunque se registraron más emersiones de E. bicolor
que de E. falcarius (Tabla 4). En ambas especies, la emersión fue más frecuente cuanto
mayor era la temperatura, siendo 40ºC el umbral crítico de máxima emersión y
decreciendo bruscamente a 45ºC (Fig. 5a) debido a la alta mortalidad registrada en
esta temperatura (Fig. 4a).
En las especies de Nebrioporus el estrés térmico produjo un resultado similar en la
respuesta de emersión, siendo 40ºC el umbral crítico de máxima actividad para las dos
especies (Fig. 5b). En esta temperatura, N. baeticus emergió menos que N. ceresyi
(post-hoc Temperatura x Epecie 40ºC: p < 0.001; Fig. 5b) debido a su mayor mortalidad
(Fig. 4b).
Entre O. glaber y O. notabilis no se encontraron diferencias significativas en el número
medio de emersiones (Tabla 4), mostrando ambas especies un patrón de respuesta
similar frente al estrés térmico. En las temperaturas más bajas se registraron pocas
emersiones y los umbrales críticos de máxima respuesta fueron 40 y 45ºC (Fig. 5c,d).
La emersión se vio significativamente afectada por la conductividad en O. glaber (p <
0.001), decreciendo conforme aumentaba el nivel de estrés salino. En las dos especies
la combinación de altas temperaturas (40 y 45ºC) y conductividad extrema (240
mS.cm-1) redujo significativamente la respuesta de emersión (Tabla 4), especialmente
en O. glaber, coincidiendo con la alta mortalidad de esta especie registrada en dichos
tratamientos.

18
Tabla 4. Resultados del ANOVA. Efecto de la temperatura y conductividad en la
emersión en los tres pares de especies estudiadas.
Variable dependiente: Emersión
Fuente SS g.l. F p
Enochrus
Modelo corregido 267.887 23 11.037 ***
Intersección 476.719 1 451.760 ***
Temperatura 242.262 3 76.526 ***
Conductividad 1.628 2 0.771 n.s
Especie 6.845 1 6.487 *
Temperatura x Conductividad 2.374 6 0.375 n.s
Temperatura x Especie 10.975 3 3.467 *
Conductividad x Especie 2.160 2 1.024 n.s
Temperatura x Conductividad x
Especie
1.643 6 0.260 n.s
Error
50.652 48
Nebrioporus
Modelo corregido 158.684 23 13.292 ***
Intersección 105.609 1 203.456 ***
Temperatura 124.697 3 80.076 ***
Conductividad 0.738 2 0.711 n.s
Especie 7.390 1 14.237 ***
Temperatura x Conductividad 1.481 6 0.475 n.s
Temperatura x Especie 22.964 3 14.747 ***
Conductividad x Especie 0.368 2 0.354 n.s
Temperatura x Conductividad x
Especie
1.047 6 0.336 n.s
Error
24.916 48
Ochthebius
Modelo corregido 228.429 39 6.194 ***
Intersección 364.241 1 385.183 ***
Temperatura 109.011 3 38.426 ***
Conductividad 35.375 4 9.352 ***
Especie 0.925 1 0.978 n.s
Temperatura x Conductividad 41.111 12 3.623 ***
Temperatura x Especie 1.833 3 0.646 n.s
Conductividad x Especie 9.234 4 2.441 n.s
Temperatura x Conductividad x
Especie
30.941 12 2.727 **
Error 75.650 80
SS, suma de cuadrados; g.l, grados de libertad.
* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, n.s no significativo
19
Figura 5. Emersión media ± ES de cada especie. Las diferencias significativas determinadas por los
análisis post-hoc con la corrección de Bonferroni se indican como sigue: con letras mayúsculas
junto a la leyenda para los niveles de conductividad, con letras minúsculas sobre las barras para los
niveles de temperatura, con números sobre las barras para los niveles de conductividad dentro de
un mismo nivel de temperatura y con asteriscos para diferencias entre especies congenéricas
dentro de un mismo tratamiento.

20
Vuelo
El patrón de actividad de vuelo frente al estrés térmico fue significativamente
diferente entre las especies de Enochrus (Tabla 5). E. bicolor voló más que E. falcarius
en todos los niveles de temperatura, siendo esta diferencia significativamente mayor
en 35 y 40ºC (post-hoc Temperatura x Especie 35ºC y 40ºC: p < 0.001; Fig. 6a), donde
se registró el máximo número de vuelos de ambas especies.
En ambas especies de Nebrioporus se registró un número significativamente alto de
vuelos en 35 y 40ºC, aunque en estas temperaturas N. baeticus voló más que N. ceresyi
(post-hoc Temperatura x Especie 35ºC: p < 0.001; 40ºC: p = 0.001; Tabla 5, Fig. 6b).
Las especies de Ochthebius iniciaron la respuesta de vuelo en niveles de estrés térmico
más altos que los otros pares de especies, siendo el umbral crítico de máximo vuelo
40ºC y mostrando una respuesta similar en 35 y 45ºC (Fig. 6c,d). La magnitud de la
respuesta de vuelo fue ligeramente superior en O. notabilis, aunque el nivel de
significación de esta comparación es próximo a 0,05 (Tabla 5). El efecto de la
conductividad sobre el vuelo solo fue significativo para O. notabilis (p < 0.001) aunque
la respuesta no mostró un patrón claro a lo largo del gradiente de estrés salino. Para
las dos especies la interacción de temperatura x conductividad tuvo un efecto
significativo sobre el vuelo, aunque de forma diferente (Tabla 5). Así, en las
temperaturas más extremas (40-45ºC) O. glaber voló más que O. notabilis en el nivel
mas bajo de conductividad (20 mS.cm-1) mientras que O. notabilis voló más que O.
glaber en las conductividades más altas (180 y 240 mS.cm-1) (Fig. 6c,d), lo cual
concuerda con la menor tolerancia que presentó O. glaber en estas condiciones más
extremas.

21
Tabla 5. Resultados del ANOVA. Efecto de la temperatura y conductividad en el vuelo en
los tres pares de especies estudiadas.
Variable dependiente: Vuelo
Fuente SS g.l. F p
Enochrus
Modelo corregido 99.778 23 2.814 **
Intersección 150.222 1 97.441 ***
Temperatura 20.333 3 4.396 **
Conductividad 0.861 2 0.279 n.s
Especie 43.556 1 28.252 ***
Temperatura x Conductividad 5.917 6 0.640 n.s
Temperatura x Especie 22.111 3 4.781 **
Conductividad x Especie 1.694 2 0.550 n.s
Temperatura x Conductividad x
Especie
5.306 6 0.574 n.s
Error
74.000 48
Nebrioporus
Modelo corregido 115.875 23 4.837 ***
Intersección 91.125 1 87.480 ***
Temperatura 63.042 3 20.173 ***
Conductividad 1.583 2 0.760 n.s
Especie 21.125 1 20.280 ***
Temperatura x Conductividad 7.083 6 1.133 n.s
Temperatura x Especie 10.819 3 3.462 *
Conductividad x Especie 1.083 2 0.520 n.s
Temperatura x Conductividad x
Especie
11.139 6 1.782 n.s
Error
50.000 48
Ochthebius
Modelo corregido 303.467 39 4.264 ***
Intersección 512.533 1 280.840 ***
Temperatura 56.267 3 10.277 ***
Conductividad 51.633 4 7.073 ***
Especie 7.500 1 4.110 *
Temperatura x Conductividad 70.233 12 3.207 ***
Temperatura x Especie 9.167 3 1.674 n.s
Conductividad x Especie 36.167 4 4.954 ***
Temperatura x Conductividad x
Especie
72.500 12 3.311 ***
Error 146.000 80
SS, suma de cuadrados; g.l, grados de libertad.
* p ≤ 0.05, ** p ≤ 0.01, *** p ≤ 0.001, n.s no significativo

22
Figura 6. Nº de vuelos medio ± ES de cada especie. Las diferencias significativas determinadas por los
análisis post-hoc con la corrección de Bonferroni se indican como sigue: con letras mayúsculas junto a la
leyenda para los niveles de conductividad, con letras minúsculas sobre las barras para los niveles de
temperatura, con números sobre las barras para los niveles de conductividad dentro de un mismo nivel
de temperatura y con asteriscos para diferencias entre especies congenéricas dentro de un mismo
tratamiento.

23
4. Discusión
¿Afecta la combinación de estrés térmico y salino a la tolerancia y el comportamiento
de huida de escarabajos acuáticos salinos?
La salinidad y la temperatura son dos de los factores abióticos que más influyen
en el crecimiento y la supervivencia de organismos acuáticos (Kir y Kumlu 2008). En los
ecosistemas marinos, varios estudios han demostrado que la salinidad influye en la
fisiología térmica de un amplio rango de animales, por ejemplo, corales (Coles y Jokiel
1978), crustáceos (Kir y Kumlu 2008), bivalvos (Rajagopal et al. 2005) y peces (Sardella
et al. 2008a, b). Recientemente se ha demostrado el efecto de la salinidad sobre los
límites térmicos en algunas de las especies estudiadas en el presente trabajo (Sánchez
Fernández et al. 2010, Arribas et al. en prensa). Estos estudios han encontrado un
efecto sinérgico de la exposición crónica a altas salinidades y temperaturas sobre los
límites térmicos letales en N. baeticus, N. ceresyi y E. falcarius, siendo dichos límites
mayores en individuos aclimatados a salinidades y temperaturas relativamente altas.
Sin embargo, en contra de lo esperado, en nuestros experimentos de exposición aguda
al estrés, ni la salinidad ni la interacción entre salinidad y temperatura tuvieron, en
general, un efecto significativo sobre la supervivencia ni las respuestas de
comportamiento de las especies estudiadas, excepto en las especies de Ochthebius,
especialmente O.glaber.
Nuestros resultados indican que la influencia de la salinidad sobre la tolerancia térmica
de las especies depende en gran medida del tiempo de exposición, siendo menos
importante en exposiciones cortas en las que los organismos son sometidos a cambios
bruscos en las condiciones térmicas y salinas que en exposiciones largas y graduales a
condiciones de estrés, donde la temperatura va progresivamente incrementándose
hasta alcanzar niveles extremos (experimentos de “ramping”). En exposiciones agudas,
el shock térmico parece causar un efecto más severo e inmediato sobre la fisiología y
la capacidad de desarrollar respuestas de huida de los organismos en comparación con
el estrés salino.

24
A diferencia del resto de especies, en O. glaber la combinación de altas
conductividades (≥ 180 mS.cm-1) con temperaturas extremas (40-45ºC) produjo un
daño fisiológico que impidió cualquier movimiento y por tanto desarrollar respuestas
de huida para evitar las condiciones de estrés, reduciendo su supervivencia. En este
caso, las interacciones entre ambos factores parecen ser más importantes cerca de los
límites de tolerancia, como han señalado otros autores (Coles y Jokiel 1978). En
individuos expuestos a temperaturas extremas los mecanismos osmoreguladores
resultan dañados ya que aumenta la tasa de pérdida de agua y la acumulación de
solutos en los organismos (Rezende et al. 2011). Es probable que en medios
hipersalinos el aumento de la temperatura acentúe los efectos del estrés sobre los
individuos, no obstante es necesario un conocimiento más profundo de las
consecuencias que la exposición a temperaturas próximas a los límites letales tiene
sobre la capacidad osmoreguladora de las especies.
¿Dependen las respuestas de comportamiento de huida de la tolerancia fisiológica de
las especies?
Los efectos de la variación de los factores abióticos del medio sobre un
organismo no solo dependen de su tolerancia fisiológica a dichos factores, sino
también de su capacidad de regular los niveles de estrés que experimenta a través de
ajustes del comportamiento (p.ej mediante la búsqueda de microhábitats). De este
modo, ambos rasgos deben ser interpretados conjuntamente determinando qué
niveles de estrés ambiental son críticos para la supervivencia de una especie bien por
causar un daño fisiológico irreversible o bien por inducir respuestas de
comportamiento para evitar la exposición a dicho estrés.
De acuerdo con nuestra predicción inicial, las respuestas de comportamiento
analizadas estuvieron condicionadas por la tolerancia al estrés de las especies. Esta
relación fue clara en las especies de Enochrus y Nebrioporus, cuya mortalidad y
respuestas de comportamiento mostraron una correlación negativa. En general, la
emersión y el vuelo aumentaron conforme lo hacía el nivel de estrés térmico en
temperaturas entre 20 y 40ºC, en las que no se produjo un daño fisiológico importante
y por tanto la mortalidad fue baja, siendo 35ºC el umbral de máximo vuelo y 40ºC el

25
umbral de máxima emersión. En cambio, en los tratamientos de 45ºC las respuestas de
comportamiento se vieron bloqueadas ya que el estrés térmico causó un daño severo
e irreversible sobre la fisiología de los individuos y en última instancia la muerte de
prácticamente todos los individuos expuestos.
En las especies de Ochthebius se observó un patrón diferente, siendo la relación entre
tolerancia y las respuestas de escape (emersión y vuelo) positiva. El nivel de estrés en
el rango de temperatura y salinidad testado no causó un daño fisiológico sobre O.
glaber y O. notabilis tan importante como en el caso anterior. La temperatura más
extrema (45ºC) redujo la supervivencia de ambas especies, sin embargo no fue letal
para todos los individuos. Por tanto, los individuos que sobrevivieron en el nivel de
estrés térmico más extremo fueron capaces de escapar de las condiciones adversas
mediante emersión y vuelo. Los umbrales térmicos críticos fueron 40ºC para el vuelo y
40-45ºC para la emersión. En este sentido, la mayor tolerancia de las dos especies de
Ochthebius en comparación con los otros pares de especies está en concordancia con
los hábitats más extremos que estas especies ocupan (Velasco et al. 2006, Abellán et
al. 2009; Millán et al. 2011).
¿Son las especies lóticas más sensibles al estrés que las especies leníticas?
De acuerdo con nuestras predicciones iniciales, en dos de los tres pares de
especies estudiadas (géneros Nebrioporus y Ochchtebius), las especies lóticas fueron
más sensibles al estrés térmico que las especies leníticas y por lo tanto en niveles
críticos de estrés sus respuestas de emersión y vuelo para escapar de las condiciones
desfavorables fueron de mayor magnitud.
Entre los Nebrioporus, la especie lótica N. baeticus fue menos tolerante al estrés
térmico que su especie hermana N. ceresyi. Este resultado concuerda con los
obtenidos por Sánchez-Fernández et al. (2010) en experimentos de aumento
progresivo de temperatura (“ramping”) para determinar los límites térmicos de este
par de especies, en los que N. ceresyi presentó un mayor rango térmico que N.
baeticus, aunque en este caso debido a su amplia tolerancia a temperaturas bajas. En
cuanto a las respuestas de comportamiento, en ambas especies se registraron los
máximos valores de vuelo y emersión en los mismos niveles de temperatura (35ºC

26
para el vuelo y 40ºC para la emersión), aunque N. baeticus, la especie menos
tolerante, mostró una mayor actividad de vuelo que N. ceresyi en estos umbrales
críticos de estrés térmico.
Entre las dos especies de Ochthebius se encontraron diferencias paralelas en la
tolerancia al estrés. O. glaber, la especie que ocupa cuerpos de agua lóticos, fue menos
resistente al estrés que su especie congenérica O. notabilis, que ocupa medios leníticos
con mayor variabilidad térmica y salina (Abellán et al. 2007, 2009). En los niveles de
estrés térmico más extremos (40-45ºC) y menor conductividad (20-50 mS.cm-1), O.
glaber emergió y voló más que O. notabilis, mientras que en las conductividades más
extremas (180-240 mS.cm-1) O. notabilis mostró una mayor actividad de emersión y
vuelo, de acuerdo con su mayor tolerancia al estrés.
Contrariamente a los resultados observados en las especies de Nebrioporus y
Ochthebius, E. bicolor y E. falcarius presentaron similares tolerancias al estrés térmico
y similar respuesta de emersión, aunque sí mostraron diferencias significativas en la
magnitud de la respuesta de vuelo. La especie lótica E. bicolor desarrolló una mayor
actividad de vuelo que E. falcarius en temperaturas subletales (35-40ºC), lo cual puede
estar relacionado con su mayor capacidad de dispersión. Análisis de la morfometría de
las alas de este par de especies han revelado que E. bicolor tiene mayor capacidad de
vuelo que su especie congenérica E. falcarius, por lo que se ha señalado la capacidad
de dispersión en vez de la tolerancia térmica como el principal factor determinante de
la contrastada distribución geográfica que muestran estas especies (Arribas et al. en
prensa). Por lo tanto, al someter a individuos de estas dos especies a condiciones de
estrés, dada su similar tolerancia fisiológica, la especie con mayor capacidad de
dispersión ha mostrado una mayor tendencia a escapar del medio para buscar
ambientes con condiciones más favorables.
La diferente estabilidad espacial y temporal de los medios lóticos y leníticos ha podido
dar lugar a la evolución de diferentes estrategias entre las especies. Así, las especies
que ocupan los medios leníticos, más inestables, se han visto forzadas a desarrollar
una mayor capacidad de colonización (tolerancia fisiológica y/o capacidad de
dispersión) que sus respectivas especies lóticas (Ribera 2008). Nuestros resultados

27
sugieren una relación positiva entre tolerancia al estrés, ocupación de hábitat y rango
geográfico en el par de especies estudiadas de Nebrioporus y Ochthebius, siendo las
especies lóticas con una distribución más restringida más sensibles a los extremos de
temperatura que sus correspondientes especies congenéricas leníticas de distribución
más amplia. Esta correlación entre tolerancia térmica y tamaño del rango de
distribución se ha encontrado en otras especies de escarabajos acuáticos (e.g. Calosi et
al. 2008, Calosi et al. 2010) y otros invertebrados (Ribera y Vogler 2000, Hof et al.
2006, Damm et al. 2010). Por otro lado, en el caso de E. bicolor y E. falcarius, como se
ha dicho anteriormente, la capacidad de dispersión y no la tolerancia térmica parece
determinar sus diferentes patrones biogeográficos (Arribas et al. en prensa).
¿Qué especies dentro de cada género estudiado podría ser más susceptible a cambios
en las condiciones de su hábitat en base a su fisiología y comportamiento frente al
estrés ambiental?
La integración de medidas de tolerancia fisiológica y respuestas de
comportamiento de especies relacionadas de escarabajos acuáticos no solo permite
describir su ecofisiología, sino que además proporciona una valiosa información sobre
las diferentes estrategias que las especies pueden adoptar en respuesta a cambios
ambientales tanto presentes como futuros. Por un lado, la mortalidad ofrece una
medida general de la tolerancia o resistencia de las especies a los factores estresantes
analizados; por otro, las respuestas de comportamiento (emersión y vuelo) dan una
idea de la habilidad de las especies para escapar de condiciones desfavorables o
variables.
Las predicciones de cambio climático prevén un incremento de las temperaturas y una
reducción de las precipitaciones en el área mediterránea (IPCC 2007), resultando en
unas condiciones más extremas (mayor temperatura y salinidad) para los organismos
que viven en los medios salinos continentales. A pesar de la amplia tolerancia de las
especies salinas a los cambios ambientales (Millán et al. 2011), en base a la fisiología y
las respuestas estudiadas en el presente trabajo, las especies lóticas N. baeticus y O.
glaber pueden ser más susceptibles que sus correspondientes especies leníticas N.
ceresyi y O. notabilis frente a cambios bruscos de temperatura. Especialmente en el

28
caso de O. glaber, la combinación de temperaturas y salinidades elevadas puede
comprometer su capacidad para adaptarse a cambios en las condiciones del medio
mediante dispersión y búsqueda de hábitats más favorables. Además, la filogeografía
de esta especie (Abellán et al. 2007, 2009), altamente amenazada en la Península
Ibérica (Sánchez-Fernández et al. 2008), ha revelado altos niveles de divergencia
genética entre poblaciones aisladas, con un bajo flujo génico y distancias de dispersión
muy cortas. Esto unido a la alta fragmentación de sus hábitats (los arroyos
hipersalinos) y la mayor sensibilidad que muestra la especie al estrés aumenta la
vulnerabilidad de O. glaber frente al cambio climático.
Los resultados obtenidos con las dos especies de Enochrus estudiadas sugieren que
ambas podrían mostrar una similar sensibilidad frente a un incremento de
temperatura de sus hábitats. Sin embargo, para E. falcarius, la especie del género más
amenazada en la Península Ibérica (Arribas et al. datos no publicados), su menor
capacidad de dispersión (Arribas et al. en prensa) puede suponer una desventaja que
podría dificultar la adaptación de la especie al cambio climático.
De acuerdo con los resultados obtenidos en este trabajo (Tabla 6), la fauna
acuática especializada de los medios salinos lóticos representa un componente
vulnerable frente al cambio global dentro de la biodiversidad de los ambientes áridos,
por lo que es recomendable centrar esfuerzos de conservación en estos hábitats tan
singulares y por otro lado amenazados e infravalorados. Sin embargo, futuros estudios
de los rasgos analizados en este trabajo y otros como la capacidad de dispersión, tasa
reproductiva o tolerancia a la composición iónica del agua serían muy útiles para
predecir con mayor rigor la vulnerabilidad de estas especies frente a cambios
ambientales presentes y futuros y para comprender mejor los divergentes patrones
biogeográficos que muestran.

29
5. Conclusiones
1. En general, no se ha observado efecto del estrés salino ni de la interacción entre
estrés térmico y salino en exposición corta sobre la tolerancia fisiológica y
comportamiento de huida en las especies estudiadas.
2. La tolerancia al estrés es un rasgo clave que condiciona la capacidad de los
individuos de desarrollar respuestas de huida cuando las condiciones del medio son
desfavorables.
3. En general las especies han mostrado una alta tolerancia al estrés térmico, aunque
una temperatura de 45ºC o superior produce un daño fisiológico importante sobre los
individuos, especialmente de las especies estudiadas de Nebrioporus y Enochrus.
Temperaturas entre 35 y 40º son claves para la emersión y el vuelo.
4. En los pares de especies estudiadas de los géneros Nebrioporus y Ochthebius, las
especies lóticas (N. baeticus y O. glaber) son más sensibles al estrés térmico que las
leníticas (N. ceresyi y O. notabilis) y en niveles críticos de estrés su respuestas de huida
son más patentes, de acuerdo con la mayor estabilidad ecológica y geológica de los
sistemas fluviales.
5. Las dos especies de Enochrus estudiadas presentan similares tolerancias al estrés
térmico, pero la respuesta de vuelo es mayor en la especie lenítica E. bicolor. En este
caso la distinta capacidad de dispersión parece ser el rasgo que determina el diferente
patrón biogeográfico y de ocupación de hábitat que muestran.
6. En los tres pares de especies estudiadas, aquellas que ocupan los medios lóticos
podrían ser más vulnerables frente a cambios en las condiciones de sus hábitat, debido
a una mayor sensibilidad al estrés térmico en el caso de N. baeticus y O. glaber y a la
menor capacidad de vuelo en el caso de E. falcarius.

30
Tabla 6. Tabla-resumen de los principales resultados: factores de estrés que han tenido efecto significativo en las respuestas estudiadas y
comparación entre la especie lótica-lenítica dentro de cada par de especies de la tolerancia al estrés, los umbrales críticos de máxima respuesta y la
magnitud de las respuestas de emersión y vuelo y la vulnerabilidad frente a cambios ambientales.
Especie
Factores
de estrés
Tolerancia
al estrés
Emersión Vuelo
Vulnerabilidad Umbrales críticos
Magnitud de
respuesta Umbrales críticos
Magnitud
de respuesta
E. bicolor T = 40ºC = 35ºC > <
E. falcarius T = 40ºC = - < >
N. ceresyi T > 40ºC > 35ºC < <
N. baeticus T < 40ºC < 35-40ºC > >
O. notabilis
T
T x C >
40ºC x 20 mS/cm
45ºC x 20 mS/cm
45º x 80 mS/cm
= 40ºC x 50 mS/cm
40ºC x 180-240 mS/cm
> <
O. glaber
T
C
T x C
< 40ºC x 20-180 mS/cm
45ºC x 20-50 mS/cm =
40ºC x 50 mS/cm
40ºC x 180 mS/cm
< >
T, temperatura; C, conductividad

31
6. Referencias
Abellán P, Gómez-Zurita J, Millán A, Sánchez-Fernández D, Velasco J, Galián J, Ribera
I. 2007. Conservation genetics in hypersaline inland waters: mitochondrial diversity
and phylogeography of an endangered Iberian beetle (Coleoptera: Hydraenidae).
Conserv Genet. 8 (1): 79-88.
Abellán P, Millán A, Ribera I. 2009. Parallel habitat-driven differences in the
phylogeographical structure of two independent lineages of Mediterranean saline
water beetles. Mol Ecol. 18: 3885-3902.
Álvarez-Cobelas M, Rojo C, Angeler DG. 2005. Mediterranean limnology: current
status, gaps and the future. J Limnol. 64: 13-29.
Angilletta MJ, Jr. 2009. Thermal adaptation. A theoretical and empirical synthesis.
Oxford. Oxford University Press.
Arribas P, Velasco J, Abellán P, Sánchez-Fernández D, Andújar C, Calosi P, Millán A,
Ribera I, Bilton DT. 2011. Dispersal ability rather than ecological tolerance drives
differences in range size between lentic and lotic water beetles (Coleoptera:
Hydrophilidae). J Biogeogr. En prensa.
Bale JS. 2002. Insects and low temperatures: from molecular biology to distributions
and abundance. Philos Trans R Soc Lond B. 357:849–861.
Bozinovic F, Calosi P, Spicer JI. 2011. Physiological correlates of geographic range in
animals. Annu Rev Ecol Evol Syst. 42: 155-179.
Calosi P, Bilton DT, Spicer JI, Atfield, A. 2008. Thermal tolerance and geographic range
size in the Agabus brunneus group of European diving beetles (Coleoptera: Dytiscidae).
J Biogeogr. 35: 295-305.

32
Calosi P, Bilton DT, Spicer JI, Votier SC, Atfield A. 2010. What determines a species’
geographical range? Thermal biology and latitudinal range size relationships in
European diving beetles (Coleoptera: Dytiscidae). J Anim Ecol. 79: 194–204.
Chidawanyika F, Terblanche JS. 2011. Rapid thermal responses and thermal tolerance
in adult codling moth Cydia pomonella (Lepidoptera: Tortricidae). J Insect Physiol.
57(1):108-117.
Chown SL, Nicolson S. 2004. Insect Physiological Ecology. Mechanism and patterns.
Oxford. Oxford University Press.
Chown SL. 2001. Physiological variation in insects: Hierarchical levels and implications.
J Insect Physiol. 47(7): 649-660.
Coles S, Jokiel PL. 1978. Synergistic effects of temperature, salinity and light on the
hermatypic coral Monbpora verrucosa. Mar Biol. 49: 187-195.
Damm S, Dijkstra KB, Hadrys H. 2010. Red drifters and dark residents: The phylogeny
and ecology of a plio-pleistocene dragonfly radiation reflects Africa's changing
environment (Odonata, Libellulidae, Trithemis). Mol Phylogenet Evol. 54(3):870-882.
Davis M, Olla B. 2001. Stress and delayed mortality induced in pacific halibut by
exposure to hooking, net towing, elevated seawater temperature and air: Implications
for management of bycatch. N Am J Fish Manage. 21(4): 725-732.
Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR.
2008. Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl
Acad Sci USA. 105(18): 6668-6672.
Feder ME. 2010. Physiology and global climate change. Annu Rev Physiol. 72:123-125.
Fery H, Fresneda J, Millán A. 1996. Bemerkungen zur Nebrioporus ceresyi-Gruppe
sowie Beschreibung von Nebrioporus schoedli n. sp. (Coleoptera: Dytiscidae). Entomol
Zeit. 106: 306–328.

33
Florencio M, Serrano L, Gómez-Rodríguez C, Millán A, Díaz-Paniagua C. 2009. Inter- and
intra-annual variations of macroinvertebrate assemblages are related to the
hydroperiod in mediterranean temporary ponds. Hydrobiologia 634(1):167-183.
Gaston KJ. 2003. The structure and dynamics of geographic ranges. Oxford. Oxford
University Press.
Gaston KJ, Chown SL, Calosi P, Bernardo J, Bilton DT, Clarke A, Clusella-Trullas S,
Ghalambor CK, Konarzewski M, Peck LS, Porter WP, Pörtner HO, Rezende EL, Schulte
PM, Spicer JI, Stillman JH, Terblanche JS, van Kleunen M. 2009. Macrophysiology: A
conceptual reunification. Am Nat. 174(5): 595-612.
Gaston KJ. 2009. Geographic range limits of species. Proc R Soc B-Biol Sci. 276(1661):
1391-1393.
Hansen M. 2004. Hydrophilidae. En: Löbl I, Smetana A, editores. Catalogue of
Palaearctic Coleoptera. Vol.2.Hydrophiloidea-Histeroidea-Staphylinoidea. Copenhagen.
Apollo Books. p. 44-67.
Hazell SP, Groutides C, Neve BP, Blackburn TM, Bale JS. 2010. A comparison of low
temperature tolerance traits between closely related aphids from the tropics,
temperate zone, and arctic. J Insect Physiol. 56(2): 115-122.
Helmuth B. 2009. From cells to coastlines: How can we use physiology to forecast the
impacts of climate change? J Exp Biol. 212(6): 753-760.
Hof C, Brandle M, Brandl R. 2006. Lentic Odonates have larger and more northern
ranges than lotic species. J Biogeogr. 33(1):63-70.
Hoffmann AA, Sorensen JG, Loeschcke V. 2003. Adaptation of Drosophila to
temperature extremes: Bringing together quantitative and molecular approaches. J
Therm Biol. 28(3): 175-216.
Hoffmann AA, Blows MW. 1994. Species borders: ecological and evolutionary
perspectives. Trends Ecol Evol. 9: 223–227.

34
Homan R, Regosin J, Rodrigues D, Reed J, Windmiller B, Romero L. 2003. Impacts of
varying habitat quality on the physiological stress of spotted salamanders (Ambystoma
maculatum). Anim Conserv. 6: 11-18.
Huey R. 1991. Physiological consequences of habitat selection. Am Nat. 137: S91-S115.
Hughes GE, Owen E, Sterk G, Bale JS. 2010. Thermal activity thresholds of the parasitic
wasp Lysiphlebus testaceipes and its aphid prey: Implications for the efficacy of
biological control. Physiol Entomol. 35(4): 373-378.
IPCC (Intergovernmental Panel on Climate Change). 2007. Climate Change 2007:
Synthesis Report. Contribution of Working Groups I, II and III to the Fourth Assessment
Report of the Intergovernmental Panel on Climate Change. Pachauri RK, Reisinger A,
editores. Geneva. Core Writing Team. p. 104.
Jäch MA, Beutel RG, Delgado JA, Díaz, JA. 2005. 11.1. Hydraenidae Mulsant, 1844. En:
Kristensen NP, editor. Handbook of Zoology, Vol. IV (Part 38), Coleoptera, Beetles, Vol.
1: Morphology and Systematics (Archostemata, Adephaga, Myxophaga, Polyphaga
partim). Berlin. Walter de Gruyter. p. 223-251
Kay C, Whitford W. 1978. Critical thermal limits of desert honey ants - possible
ecological implications. Physiol Zool. 51(2): 206-213.
Kir M, Kumlu M. 2008. Effect of temperature and salinity on low thermal tolerance of
Penaeus semisulcatus (Decapoda: Penaeidae). Aquacult Res. 39(10): 1101-1106.
Korsloot A, van Gestel C, van Straalen N. 2004. Environmental stress and cellular
response in arthropods. Boca Raton. CRC Press.
Marais E, Chown SL. 2008. Beneficial acclimation and the bogert effect. Ecol Lett.
11(10): 1027-1036.
Massot M, Clobert J, Ferriere R. 2008. Climate warming, dispersal inhibition and
extinction risk. Glob Change Biol. 14: 461-469.

35
Millán A, Velasco J, Gutiérrez-Cánovas C, Arribas P, Picazo F, Sánchez-Fernández D,
Abellán P. 2011. Mediterranean saline streams in southeast Spain: What do we know?
J Arid Environ. 75(12):1352-1359.
Pandolfo TJ, Cope WG, Arellano C. 2010. Thermal tolerance of juvenile freshwater
mussels (Unionidae) under the added stress of copper. Environ Toxicol Chem. 20
(3):691-699.
Rajagopal S, Van der Velde G, Van der Gaag M, Jenner H. 2005. Factors influencing the
upper temperature tolerances of three mussel species in a brackish water canal: Size,
season and laboratory protocols. Biofouling. 21(2): 87-97.
Rezende EL, Tejedo M, Santos M. 2011. Estimating the adaptive potential of critical
thermal limits: methodological problems and evolutionary implications. Funct Ecol. 25:
111-121.
Ribera I. 2008. Habitat constraints and the generation of diversity in freshwater
macroinvertebrates. En: Lancaster J, Briers RA, editores. Aquatic Insects: Challenges to
Populations. Wallingford. CAB International Publishing. p. 289-311.
Ribera I, Vogler A. 2000. Habitat type as a determinant of species range sizes: The
example of lotic-lentic differences in aquatic coleoptera. Biol J Linn Soc. 71(1): 33-52.
Rutherford A. 2001. Introducing ANOVA and ANCOVA a GLM approach. London. SAGE
Publications.
Sánchez-Fernández D, Calosi P, Atfield A, Arribas P, Velasco J, Spicer JI, Millán A, Bilton
DT. 2010. Reduced salinities compromise the thermal tolerance of hypersaline
specialist diving beetles. Physiol Entomol. 35(3):265-273.
Sánchez-Fernández D, Bilton DT, Abellán P, Ribera I, Velasco J, Millán A. 2008. Are the
endemic water beetles of the Iberian Peninsula and the Balearic Islands effectively
protected? Biol Conserv. 141: 1612–1627.

36
Sardella BA, Kultz D, Cech JJ, Jr., Brauner CJ. 2008a. Salinity-dependent changes in
Na+/K+-ATPase content of mitochondria-rich cells contribute to differences in thermal
tolerance of Mozambique tilapia. J Comp Physiol B-Biochem Syst Environ Physiol.
178(3): 249-256.
Sardella BA, Sanmarti E, Kultz D. 2008b. The acute temperature tolerance of green
sturgeon (Acipenser medirostris) and the effect of environmental salinity. J Exp Zool A-
Ecol Genet Physiol. 309A(8): 477-483.
Schödl S. 1998. Taxonomic revision of Enochrus (Coleoptera:Hydrophilidae) I. The E.
bicolor species complex. Entomol Probl. 29: 111-127.
Sinclair B, Vernon P, Klok C, Chown SL. 2003. Insects at low temperatures: An
ecological perspective. Trends in Ecol Evol. 18(5): 257-262.
Spicer JI, Gaston KJ. 1999. Physiological Diversity and its Ecological Implications.
Oxford. Blackwell Science.
Stillman J. 2003. Acclimation capacity underlies susceptibility to climate change.
Science. 301(5629): 65.
Swanson C, Reid T, Young P, Cech J. 2000. Comparative environmental tolerances of
threatened delta smelt (Hypomesus transpacificus) and introduced wakasagi
(H.nipponensis) in an altered California estuary. Oecologia. 123(3): 384-390.
Terblanche JS, Hoffmann AA, Mitchell KA, Rako L, Le Roux PC, Chown SL. 2011.
Ecologically relevant measures of tolerance to potentially lethal temperatures. J Exp
Biol. 214(22): 3713-3725.
Tewksbury JJ, Huey RB, Deutsch CA. 2008. Ecology - putting the heat on tropical
animals. Science. 320(5881): 1296-1297.
Thuiller W, Lavorel S, Araujo MB, Sykes MT, Prentice IC. 2005. Climate change threats
to plant diversity in Europe. Proc Natl Acad Sci USA. 102: 8245-8250.

37
Toledo M. 2009. Revision in part of the genus Nebrioporus Regimbart, 1906, with
emphasis on the N.laeviventris-group (Coleoptera: Dytiscidae). Zootaxa. 2040: 1-111.
Tomanek L, Somero GN. 1999. Evolutionary and acclimation-induced variation in the
heat shock responses of congeneric marine snails (genus Tegula) from different
thermal habitats: Implications for limits of thermotolerance and biogeography. J Exp
Biol. 202: 2925-2936.
Underwood AJ. 1997. Experiments in Ecology: Their logical design and interpretation
using Analysis of Variance. Cambridge. Cambridge University Press.
Velasco J, Millán A, Hernández J, Gutiérrez C, Sánchez D, Abellán P, Ruiz M. 2006.
Response of biotic communities to salinity changes in a Mediterranean hypersaline
stream. Saline Syst. 12 (2): 1-15.
Velasco J, Millán A. 1998. Insect dispersal in a drying desert stream: Effects of
temperature and water loss. Southwest Nat. 43(1):80-87.
Willott S, Hassall M. 1998. Life-history responses of British grasshoppers (Orthoptera:
Acrididae) to temperature change. Funct Ecol. 12(2): 232-241.
Zalom F, Grigarick A, Way M. 1980. Diel flight periodicities of some Dytiscidae
(Coleoptera) associated with California rice paddies. Ecol Entomol. 5(2): 183-187.

38
Agradecimientos
Gracias a todos los compañeros del grupo de investigación de Ecología Acuática del
departamento de Ecología e Hidrología que desde que surgió este trabajo han aportado de una
u otra forma su granito de arena para que por fin tanto esfuerzo pueda materializarse: a los
directores, Andrés y Pepa, que han contribuido en el trabajo de principio a fin, a Paula, por sus
útiles consejos y aportaciones, a Vanesa y Jose, por su imprescindible ayuda en la parte
experimental.
Al equipo del área de Biología Animal del departamento de Zoología y Antropología Física con
cuya colaboración hay un nuevo e interesante frente abierto para dar continuidad a este
trabajo.
También a los que desde fuera de la universidad siempre prestan una útil ayuda con su apoyo
e interés: familiares y amigos que siempre tienen palabras de ánimo cuando más se necesitan.