features ofsmallintestinal pathology(epithelial kinetics
TRANSCRIPT

Gut, 1982, 23, 498-506
Features of small intestinal pathology (epithelial cellkinetics, intraepithelial lymphocytes, disaccharidases) ina primary Giardia muris infectionJ GILLON, D AL THAMERY,* and ANNE FERGUSONt
From the Gastro-Intestinal Unit, University of Edinburgh and Western General Hospital, Edinburgh
SUMMARY In an attempt to correlate host and parasite-related events occurring during the courseof a primary Giardia infection in the mouse we have measured epithelial cell kinetics, enzymes,and intraepithelial lymphocytes at different stages of the infection. New methods were developedfor the accurate measurement of parasite numbers and distribution within the gut. In jejunum amodest decrease in villus length and intraepithelial lymphocytes at week 1 preceded apronounced disaccharidase deficiency at week 2, the time of maximum trophozoite numbers,whereas crypt lengthening and increased cell production became maximal at week 3. Astrophozoite numbers fell the intraepithelial lymphocyte count and disaccharidase values rose.With the exception of the intraepithelial lymphocyte count, which followed the same pattern as injejunum but two weeks later, the changes seen in the ileum were the opposite of those injejunum, suggesting rapid ileal adaptation. The results indicate that the disaccharidase deficiencyassociated with giardiasis is likely to represent a direct effect of the parasite on the brush borderrather than enterocyte immaturity, whereas the intraepithelial lymphocyte response reflects hostimmunity to the parasite. Profound adaptive changes occur throughout the small intestine inresponse to a relatively localised insult.
Infection with Giardia lamblia may be entirelyasymptomatic, may produce a mild, self-limitingillness, or chronic diarrhoea with or without mal-absorption. The reasons for this variation in severityare unknown at present, but among the possiblecauses to be considered are variations in parasitevirulence, associated bacterial overgrowth in thesmall intestine, nutritional status, and other hostfactors such as the type of the immune response andits effect on the intestinal mucosa. Various ab-normalities of small bowel pathology have beenidentified consistently in giardiasis, and theseinclude disaccharidase deficiencies 12 increasedintraepithelial lymphocyte counts3 4 and, in patientswith malabsorption, crypt hyperplasia with shortvilli and increased lamina propria cellularity.5 6 Theseverity of the mucosal changes in a biopsy ofproximal jejunum appears to correlate with the
* On leave from the Department of Paediatrics, Basrah Medical College, Iraq.t Address for correspondence and reprint requests: Dr J Gillon, Gastro-intestinal Unit, Western General Hospital, Edinburgh EH4 2XU.Received for publication 19 October 1981
overall degree of malabsorption,2 7 8despite the factthat the distribution of the parasite along the smallintestine in man is unknown, and the proximaljejunum may not necessarily be representative ofthe mucosa of the whole small bowel.
Giardiasis affects many other species, includinglaboratory rodents. Murine giardiasis is a usefulexperimental system for studies of host parasiterelationships, for it is possible to document themagnitude and time-course of infection by cyst andtrophozoite counts.9 We have previously reportedl'that in chronic murine giardiasis the intraepitheliallymphocyte count is increased, epithelial cellkinetics are altered, and tissue sucrase activity isreduced. These features are similar to those in thejejunum in 'mild' human disease.
In the experiments described in this paper wehave examined the progression of changes inepithelial cell kinetics, enzymes, and intraepitheliallymphocytes during a primary Giardia muris infec-tion. The objectives have been to establish which, ifany, of the pathological features appear to becorrelated with the magnitude of the parasite
498

Features of small intestinal pathology in a primary Giardia muris infection
infection at that site, and whether any of thesefeatures are likely to be due to mechanisms otherthan the direct effect of the parasite. In particular,we were interested in the possible role of theimmune response as a mechanism of tissue damage.
Methods
ANIMALSCBA mice (male and female) between 8 and 10weeks of age at the time of infection were used.They had free access to tap water and pelletedrodent diet (Strats mouse pellets providing 4.2calories/g, protein content 21.3%). Faecal speci-mens and intestinal contents of stock CBA micewere checked regularly to confirm the absence ofprotozoal and helminth infections in the colony.
GIARDIA MURIS INFECTIONGiardia muris cysts were provided by Dr I Roberts-Thompson and flown from Australia to the UnitedKingdom by airmail. Stock mice were infected byoral inoculation of 1000 cysts, and the infectionmaintained by weekly inoculation of two or threeadult CBA mice. For all experiments, CBA miceaged 8 to 10 weeks were infected by intragastricadministration of 1000 Giardia muris cysts in 0.2 mltap water. Cysts were isolated and counted by themethod of Roberts-Thompson et a19 describedbelow.
CYST COUNTSA two-hour faecal collection was obtained fromeach mouse by isolating it within a plastic tub fortwo hours. Faeces were then broken up in tap waterand the faecal suspension layered on molar sucroseof specific gravity 1.11, and centrifuged at 400 g for15 minutes. Cysts, concentrated at the water-sucroseinterface, were removed, washed in normal salineand resuspended in a known volume beforecounting in a haemocytometer. With this techniquethe limit of detection is 1000 cysts per specimen.
TROPHOZOITES: HISTOLOGYIn animals which were used for measurements ofepithelial cell kinetics and disaccharidases, thepresence of trophozoites in the intestine was con-firmed histologically or by direct smear of jejunalcontents.
TROPHOZOITE COUNTS: VIBRATION METHODAccurate trophozoite counts were performed inother batches of animals, by a vibration method.The animal is killed, the small intestine removedintact, and the mesentery peeled off. The gut is theneverted on a spiral rod of glass or steel, exposing the
luminal surface, and the ends tied with silk sutures.The top part of the spiral rod is then connected to aChemap AG Vibromix Model Type El, which has avibration frequency of 50 Hz. The spiral is thenplaced in a dish containing 100 ml of fluid (98 mlnormal saline, 2 ml acetyl cysteine Airbron) at 4°C.The intestine is then vibrated in medium for 10minutes at the full power of the Vibromix, a 10 mlsample of medium removed, centrifuged at 400 g for10 minutes, the supernatant discarded, the pelletresuspended in 0*5 ml of medium and thetrophozoites counted in a haemocytometer. Thetotal trophozoite count/100 ml - that is, per intestine- is calculated. With this technique the limit ofdetection is 10 000 trophozoites/animal.
TROPHOZOITE COUNT: SIEVE METHODIn order to define the distribution of trophozoitesalong the intestine, another technique was used in asmall number of animals. The whole small intestinewas removed and its length measured. Beginning atthe gastroduodenal junction, segments of wholeintestine 1 cm in length were taken, discarding 2 cmbetween each. Thus counts were performed onone-third of the entire intestine. Each 1 cm segmentwas mashed through a stainless steel grid with holesof 50 ,um diameter, using a glass rod, and washedthrough with 1 ml normal saline from a syringe andneedle. Aliquots of the resulting suspension werethen examined and a trophozoite count performedin a Neubauer haemocytometer.
MEASUREMENTS OF EPITHELIAL CELL KINETICSAND INTESTINAL ARCHITECTUREA stathmokinetic technique, using colchicine block-age and microdissection, was used. A group of mice(10-15 in most experiments) was taken andcolchicine (BDH) injected, 5 mg/kg body weight,intraperitoneally. Mice were killed by ether over-dosage or cervical dislocation at intervals from 30minutes to 24 hours later and a piece of jejunum, 10cm from the pylorus, and a piece of ileum, 5 cmfrom the ileocaecal valve, were dissected out andfixed in a 75% absolute alcohol, 25% glacial aceticacid mixture, for 24 hours. Tissues were thentransferred to 75% alcohol until processed formeasurements of villus length, crypt length, andmetaphases/crypt.For measurements of crypts and villi, tissues were
stained in bulk by an adaptation of the Feulgantechnique of Wimber and Lamerton. 12 13 The tissueswere stained as follows: 50% alcohol 10 minutes; tapwater 10 minutes; hydrolysis in molar HCI at 60°Csix minutes; tap water 10 minutes; Schiff reagent 20minutes; then stored in tap water until examined.With a dissecting microscope the tissue was
499

Gillon, Al Thamery, and Ferguson
orientated and one or two villi with their surround-ing crypts were cut from the edge by carefuldissection with a scalpel and fine forceps, placed ona slide in 45% acetic acid, and lengths of villi andcrypts measured by a calibrated micrometer eye-piece. For each specimen 10 measurements of villuslength and of crypt length were made and the meanvalue obtained.
In order to count metaphases, a fragment of thetissue was placed on a slide in 45% acetic acid andgently squashed with a coverslip. The number ofmetaphases/crypt was counted. Metaphase blockagewas found in every specimen examined. For eachspecimen the number of metaphases in 10 crypts wascounted and the mean value obtained. The crypt cellproduction rate was calculated from the slope of theregression line, drawn using the method of leastsquares, of metaphase count against time aftercolchicine injection.
HISTOLOGY AND INTRAEPITHELIAL LYMPHOCYTECOUNTSAt the same time as specimens were taken forepithelial cell kinetic measurements, adjacent seg-ments of jejunum and ileum were fixed in formolsaline and paraffin embedded, 5 gm sections werecut and stained with haematoxylin and eosin.Intraepithelial lymphocyte counts were performedby a differential count in the epithelium covering thevilli, in well-orientated H and E stained sections.Results were expressed as lymphocytes/100 epi-thelial cells.15 In addition, an assessment of theheaviness of Giardia infection was made, scoringeach section as containing many trophozoites(confluent sheets of trophozoites in the mucus overthe villi), few trophozoites (single organisms, atleast one or two per villus, and readily seen), or notrophozoites (no parasites seen in six to eight highpower fields.
DISACCHARIDASE ASSAYTissues for disaccharidase assay were taken fromjejunum around 10 cm from the pylorus, and ileum,5 cm from the ileocaecal valve. The full thicknessspecimen of intestine was weighed, homogenised inice-cold maleate buffer, and the disaccharidaseslactase, sucrase, and maltase assayed by the tech-nique of Dahlqvist.16
STATISTICAL ANALYSISIn all studies the overall significance of the resultswas first assessed by parametric and non-parametricone way analyses of variance (F-test and Kruskall-Wallis respectively). Where appropriate individualgroups were then compared using Student's t test.
Results
TIME-COURSE OF INFECTION AND DISTRIBUTIONOF TROPHOZOITESIn order to establish the typical time-course ofinfection in the strain of mice used for the experi-ments, the pattern of cyst excretion after infectionwith 1000 cysts was examined in eight CBA mice atone, two, three, four, and six weeks post-infection.In addition cyst counts were carried out in 50 CBAmice which had been infected between three and 11months previously, for the maintenance of a stock ofinfected animals.
Thirty-seven infected mice were killed at intervalsup to six weeks post-infection (six to nine animals/group) and in each animal the total small intestinaltrophozoite load was measured by the vibrationtechnique. A further 18 stock CBA mice, three to 11months post-infection, were killed and trophozoitescounted by this vibration technique.
Results are shown in Fig. 1. Results of cyst countsshow the pattern of cyst excretion to be very similarto previous reports, with peak cyst excretion at twoweeks, and cysts undetectable in the majority ofanimals after four weeks. The number of tropho-zoites also reached a peak at two weeks post-infection, with values in some animals as high as15-18x106. Trophozoites were also found in sub-stantial numbers at six weeks post-infection, whencysts were no longer detected in faeces, and werealso present in 50% of the animals studied at three
Trophozoiteor cystcount 7.0(l 1910)
6.0
5.0
4.0
3.0
f9A18 positivel
~ -_ [.o5 positive
1 2 3 4 5 6 >12Weeks post-infection
Fig. 1 Cyst excretion (two hour collection) and intestinaltrophozoite count (vibration method) in CBA mice atintervals after infection with 1000 cysts ofGiardia muris.Mean ± standard error, six to nine animals per group. Thenumbers at >12 weeks indicate proportion ofanimals inwhich either trophozoites or cysts were detected, >12 weeksafter a primary infection. (Values for positive cyst countsranged 1000-8000; values for positive trophozoite countsranged 10 000-170 000.) * Trophozoites (per animal). 0Cysts (two hourfaeces collection).
500

Features of small intestinal pathology in a primary Giardia muris infection
or more months post-infection. The distribution ofparasites along the small intestine was examined atintervals from six to 35 days after infection, and theresults in three representative animals, infected six,14, and 35 days previously, are summarised in Fig.2. The findings confirm the previously reportedobservations, that the greatest number of tropho-zoites are to be found in the proximal small bowel,although early in infection substantial numbers(several thousand trophozoites/cm segment) were
present in the ileum. As the infection progressed thelocalisation became more obviously confined to theupper jejunum.
In Table 1 the results of studies in histologicalsections of jejunum and ileum are summarised, overthe 10 weeks after primary infection of CBA micewith 1000 Giardia muris cysts. It is clear that, just aswas shown by the quantitative assay, the highestnumbers of trophozoites are present in the jejunumin the first four weeks post-infection, with few or no
parasites visible at eight and 10 weeks. In the ileum,confluent sheets of trophozoites were seen in onlyfour of the 13 mice examined at one week post-infection, and few trophozoites were present in themajority of animals at one to four weeks post-infection.
60Q000
40Q000-
20QOOO
U)
z
M 600.0000
0400
W 20QOOO0
MASHING TECHNIQUE6 DAYS
.. 91 1. v
35 DAYS
4 1 4 7 10 13 16 19 22 25 28 31 34 iDUODENUM 1 cm SAMPLES EVERY ILEUM
3rd cm
Fig. 2 Distribution of Giardia trophozoites along thelength ofthe small intestine in three CBA mice, six, 14, and35 days after infection with 1000 cysts ofGiardia muris (formethod see text).
EFFECTS OF PRIMARY GIARDIA MURIS INFECTIONON THE LENGTHS OF VILLI AND CRYPTSFor this experiment, batches of 10-15 mice/groupwere infected with 1000 Giardia muris cysts orally; agroup of animals was killed at one, two, three, four,six, eight, and 10 weeks post-infection. One batch ofcontrol, uninfected mice was also studied. Eachmouse was given intraperitoneal colchicine andkilled at a measured time interval thereafter, andspecimens of jejunum and ileum taken for microdis-section, and for histology as described below.
Results of the measurements of crypts and villi in
jejunum and ileum are summarised in Figs. 3 and 4.Results for jejunum show significant shortening ofvilli only at one week post-infection (p<0.01), meanvillus length being reduced from 690 ,m to 630 ,m.Villi were significantly longer (p<0.05) thancontrols at four, six, eight, and 10 weeks post-infection. Significant crypt lengthening was alsofound between one and four weeks post-infection.
Results for the ileum showed a different pattern.Information is available for only five groups ofanimals (because of deterioration of other speci-
Table 1 Trophozoites (Tz) in histological sections ofjejunum and ileum in CBA mice infected with 1000 Giardia muriscysts
Jejunum Ileum
Experimental Total Number of Total Number ofgroup number of specimens with: number of specimens with:(weeks post- specimens specimensinfection) examined Many Tz Few Tz None examined Many Tz Few Tz None
1 14 12 2 - 13 4 7 22 15 15 - - 15 - 13 23 15 14 1 - 14 - 8 64 15 14 - 1 16 - 6 106 16 8 8 - 15 - - 128 13 1 5 7 12 - - 1210 10 1 2 7 12 - - 12
L- . .
501

Gillon, Al Thamery, and Ferguson
2 4 6 8 10weeks post-infection
Fig. 3 Diagrammatic representation ofarchitecture ofjejunal mucosa in groups ofmice either uninfected, or atintervals ofone to 10 weeks after infection with Giardiamuris. Ten to 15 mice per group. (Mean ± SD. In this andthefollowingfigures single asterisk = p<005, doubleasterisk = p<0.01).
mens in storage) and show significant shortening ofboth villi and crypts at eight and 10 weeks post-infection.
EFFECT OF PRIMARY GIARDIA MURIS INFECTION ONCRYPT CELL PRODUCTION RATE (CCPR) INJEJUNUM AND ILEUMResults of the measurement of epithelial cellkinetics are summarised in Table 2. Metaphaseaccumulation was linear in all groups over theperiod studied, and the jejunal CCPR for the batchcontrol animals was 6-2 mitoses/crypt/hour. Valueswere higher in jejunum of mice at one, two, three,four, and eight weeks post-infection, but the onlyresult which approached statistical significance wasthat at three weeks with p value between 0.1 and0*05.The crypt cell production rate was higher in the
Villus 600-lengthI JAM) 4007
200-
Cryptlength 200-(FLm)
* *
Uninfected 2 4 6 8 10
weeks post-infection
Fig. 4 Diagrammatic representation ofarchitecture ofilealmucosa in groups ofmice either uninfected, or at intervalsofone to 10 weeks after infection with Giardia muris. Ten to15 mice per group.
ileum than in the jejunum of control animals, being8.6 mitoses/crypt/hour. In the ileum significantlylower values were present in the batches of micekilled at three, eight, and 10 weeks post-infection.
EFFECT OF PRIMARY GIARDIA MURIS INFECTION ONDISACCHARIDASE CONTENT OF JEJUNUM ANDILEUMAssays of tissue disaccharidases were carried out ina different batch of mice from those used in theepithelial cell kinetic study. There were nine un-infected animals, and groups of four mice which hadbeen infected with 1000 Giardia muris cysts one,two, three, four, six, eight, and 10 weeks previously.In contrast with the minimal effect of the infectionon intestinal architecture, striking and significantdisaccharidase changes were obtained (Figs. 5 and6).At two weeks post-infection, lactase, sucrase, and
maltase levels in the jejunum were significantlylower than in control animals. At six, eight, and 10weeks the disaccharidase content was higher than in
Table 2 Crypt cell production rate (CCPR) injejunum and ileum ofCBA mice infected with 1000 Giardia muris cysts
CCPR- jejunum CCPR - ileumExperimentalgroup Accumulated Accumulated(weeks post- metaphasesl p (versus metaphasesl p (versusinfection) cryptlhour uninfected) cryptlhour uninfected)
6-2 8-61 7-1 NS -
2 7-0 NS -
3 8-2 O05<p<O-1 5.5 OO5<p<O.14 7-5 NS -
6 559 NS 6-6 NS8 7-0 NS 5-0 p<00210 5-8 NS 5.3 p<0.05
502

Features of small intestinal pathology in a primary Giardia muris infection
°°Or
MALTASE
mean + SD* P (0.051 P(0.01
so5
20p
cm
a, 10CL.5_ 5
U
: 2
*0
UUa
a 0.5
0.2
A A
Uninfected 2 4 6 8weeks post-infection
10
MALTASE
Uninfected 2 4 6 8weeks post-infection
Fig. 5 Effects ofprimary Giardia muris infection ondisaccharidase content ofjejunal mucosa, in CBA miceeither uninfected, or at intervals from one to 10 weeks afterGiardia muris infection.
the control mice. In the ileum the only abnormalityin the early phase post-infection was a significantincrease in lactase at one week and sucrase at twoweeks post-infection. Significantly low values forlactase and sucrase, however, were obtained inspecimens taken at six, eight, and 10 weeks post-infection (Fig. 6).
EFFECTS OF PRIMARY GIARDIA MURIS INFECTION
ON INTRAEPITHELIAL LYMPHOCYTE COUNTS IN
JEJUNUM AND ILEUM
Counts of intraepithelial lymphocytes were per-formed in six uninfected mice, and in groups of sixinfected one, two, three, four, six, eight, and 10weeks previously. The results are summarised inFig. 7. In the jejunum there was a significantreduction in the intraepithelial lymphocyte count atone week, but by three weeks the counts were
significantly raised, and remained so thereafter. Inthe ileum a significant fall occurred at three weeks,and raised counts were obtained from six weeksonwards.
Fig. 6 Effects ofprimary Giardia muris infection ondisaccharidase content of ileal mucosa, in CBA mice eitheruninfected, or at intervals from one to 10 weeks afterGiardia muns infection.
SUMMARY OF THE ABNORMAL FEATURESIn Table 3 the features of abnormality of intestinalarchitecture, disaccharidases, and lymphocytecounts in these experiments are summarised. Injejunum, peak trophozoite numbers were present attwo weeks with many parasites in the jejunum up tofour weeks post-infection. A modest reduction invillus length and intraepithelial lymphocytesoccurred at week 1, and striking disaccharidasedeficiencies at week 2. This was followed bylengthening of the crypts of Lieberkuhn and a raisedcrypt cell production rate, peaking at week 3. Laterafter infection, as trophozoite numbers fell, theintraepithelial count rose, and there was an increasein both the length of the villi and the disaccharidaseactvity of the jejunum.
In contrast, in the ileum few or no trophozoiteswere present during the course of this infection. Theonly changes in the mucosa of the ileum in the earlyphase were an increase in the content of sucrase andlactase at weeks 1 and 2 respectively, and a decreasein crypt cell production rate and intraepithelial
°°0r
soF
20-
& 10IA
c-5
.0
U
c' 2
._
o 1
U
aU.
10C) 0.51
0.2 _
10a a E E 503
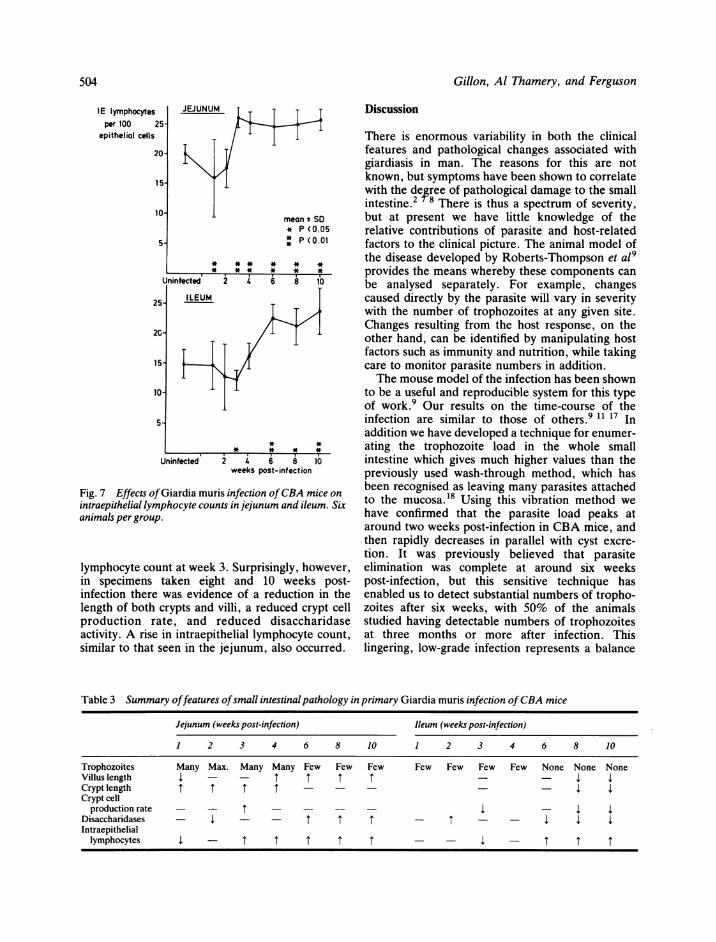
Gillon, Al Thamery, and Ferguson
IE lymphocytesper 100 25
epitheliol cells
20
151
10
25
25-
2G
15-
10-
5-
JEJUNUM
Unintected
ILEUI
_
Uninfected
Fig. 7 Effects ofGiardia mintraepithelial lymphocyte cc
animals per group.
lymphocyte count at weein specimens taken eiinfection there was evidlength of both crypts anproduction rate, andactivity. A rise in intraelsimilar to that seen in th
Discussion
1 1 There is enormous variability in both the clinicalfeatures and pathological changes associated withgiardiasis in man. The reasons for this are notknown, but symptoms have been shown to correlatewith the degree of pathological damage to the smallintestine.2 78 There is thus a spectrum of severity,
mean t SD but at present we have little knowledge of the* P (0.05 relative contributions of parasite and host-related
P<0. factors to the clinical picture. The animal model ofthe disease developed by Roberts-Thompson et al9provides the means whereby these components can
2 4 6 8 10 be analysed separately. For example, changes
M caused directly by the parasite will vary in severitywith the number of trophozoites at any given site.
Changes resulting from the host response, on theother hand, can be identified by manipulating hostfactors such as immunity and nutrition, while takingcare to monitor parasite numbers in addition.The mouse model of the infection has been shown
to be a useful and reproducible system for this typeof work.9 Our results on the time-course of theinfection are similar to those of others.9 11 17 Inaddition we have developed a technique for enumer-
* * * * ating the trophozoite load in the whole small2z 6 8 10 intestine which gives much higher values than theweeks post-infection previously used wash-through method, which has
been recognised as leaving many parasites attachedluris infection of CBA mice on 1iunts injejunum and ileum. Six to the mucosa.18 Using this vibration method we
have confirmed that the parasite load peaks ataround two weeks post-infection in CBA mice, andthen rapidly decreases in parallel with cyst excre-
tion. It was previously believed that parasite,k 3. Surprisingly, however, elimination was complete at around six weeksight and 10 weeks post- post-infection, but this sensitive technique haslence of a reduction in the enabled us to detect substantial numbers of tropho-id villi, a reduced crypt cell zoites after six weeks, with 50% of the animalsreduced disaccharidase studied having detectable numbers of trophozoites
pithelial lymphocyte count, at three months or more after infection. Thisie jejunum, also occurred. lingering, low-grade infection represents a balance
Table 3 Summary offeatures ofsmall intestinal pathology in primary Giardia muris infection ofCBA mice
Jejunum (weeks post-infection) Ileum (weeks post-infection)
1 2 3 4 6 8 10 1 2 3 4 6 8 10
Trophozoites Many Max. Many Many Few Few Few Few Few Few Few None None NoneVillus length 4 - - 1 1 1 1 - - 4 4Cryptlength 1 1 1 - - _ _ - 4 4Crypt cell
production rate - - 1 - - - - 4 - 4 4Disaccharidases - 4 - - I I I - I - - 4 4 4Intraepitheliallymphocytes 4 - I I I I I - - 4 - I I I
504
i-
1-
i-
1-

Features of small intestinal pathology in a primary Giardia muris infection
between parasite virulence and host immunity, andwe have shown in a separate study that cortico-steroid treatment of these mice can cause recrudes-cence of the infection.1' There is, however, nodoubt that the rapid diminution in parasite numbersat around six weeks is immunologically mediated, asit does not occur in T cell depleted animals. 17 It hasnot yet been established whether the reduction inparasite numbers is effected by T cells themselves, Tdependent antibody or T cell stimulatedmacrophages.The aim of the present study was to examine small
intestinal epithelial cell kinetics and villus and cryptarchitecture, brush border enzyme activity, andintraepithelial lymphocyte counts throughout thecourse of infection, in order to establish which of thechanges could be attributed directly to the parasiteload and which were likely to be secondary to thehost response. The most striking effect clearlyattributable to the parasite itself was a reduction inbrush border disaccharidase activity, particularlythat of lactase, in the jejunum at two weekspost-infection. This result was in conflict with ouroriginal hypothesis that the changes in disaccha-ridases known to occur in giardiasis reflected thepresence of an immature enterocyte population. 19 Infact, the changes in intestinal architecture andepithelial cell kinetics were small in comparison withthe drop in disaccharidase activity, and by week 3,when changes in the crypts were maximal, disaccha-ridase activity had returned to normal. It has beenrecognised that the presence of Giardia adjacent toa sheet of columnar epithelial cells is accompaniedby damage to the microvilli covering the luminal cellsurface.20 This phenomenon has also been found inbacterial overgrowth of the small bowel.21 Asgiardiasis and bacterial overgrowth frequently occurtogether,22 23 the precise mechanism whereby thebrush border enzyme activity is reduced needsfurther investigation.The changes in intestinal architecture early in the
course of the infection consisted mainly of a verylimited increase in crypt mitosis with villi of normallength. This is similar to the pattern of intestinalarchitecture which we found in chronic murinegiardiasis,10 and also is similar to the chroniclow-grade graft-versus-host disease of mice.24Mucosal changes in graft-versus-host disease and inallograt rejection are caused by cell-mediated hyper-sensitivity. 14 24 We have proposed that T cellssecrete 'enteropathic' lymphokines which actdirectly on the crypt cells and stimulate mitosis. In amore vigorous cell-mediated reaction they may alsolead to villus damage. This hypothesis has beensupported by the work of Roberts-Thompson andMitchell,17 who showed that the changes in the
villus-crypt ratio of giardia infected athymic (nude)mice were augmented when the mice were reconsti-tuted with lymphoid cells from non-infected donormice.There were significant changes in the intra-
epithelial lymphocyte counts in both jejunum andileum. In the jejunum there was a significantreduction in the intraepithelial lymphocyte count atweek 1, but by week 3, when parasite numbers werefalling, the intraepithelial lymphocyte count wasraised and remained so throughout the study.Similar changes occurred in the ileum but thechanges appeared two to three weeks later. Intra-epithelial lymphocytes are mainly T cells25 and theseresults provide further evidence for involvement ofT cell mediated immune reaction during the expul-sion phase in giardiasis. Intraepithelial lymphocytecounts have been found to be raised in the majorityof adults and children with giardiasis who have beeninvestigated for malabsorption.25 Our findings of asustained rise in intraepithelial lymphocyte countafter trophozoites have largely been eliminatedsuggests that the occasional isolated finding of a highintraepithelial lymphocyte count in the jejunalbiopsy of a patient investigated for diarrhoea mayindicate recent, self-cured protozoal infection. Thefall in intraepithelial lymphocyte count early in thecourse of the infection is unexplained and wouldmerit further study.The changes in small bowel architecture late after
infection (weeks 8 and 10) consisting, in thejejunum, essentially of an expansion in villus sizewith increased disaccharidase activity, and in theileum of a diminution in villi and crypts with reduceddisaccharidases, were entirely unexpected findings.The ileal changes tend to run in the oppositedirection from those in the jejunum, and would beconsistent with the remarkable adaptive propertiesof the ileum. As with the intraepithelial lymphocytecounts, these features illustrate how a relativelyminor insult, albeit more or less localised to one partof the small bowel, may lead to prolonged altera-tions in epithelial cell turnover and in mucosallymphoid cell populations throughout the intestine.
We are grateful to the staff of the Animal Unit,Western General Hospital, and to Miss G Paul andMiss F Allan, Mr A Sutherland, and Mr F Donnellyfor technical assistance. This work was supported inpart by a grant from the National Fund for Researchinto Crippling Diseases.
505

506 Gillon, Al Thamery, and Ferguson
References
1 Jennings W, Rowland R, Hecker R et al. The signifi-cance of lowered disaccharidase levels. Aust NZ J Med1976; 6: 556-60.
2 Duncombe VM, Bolin TD, Davis AE et al. Histo-pathology in giardiasis: a correlation with diarrhoea.Aust NZ J Med 1978; 8: 392-6.
3 Ferguson A, McClure JP, Townley RRW. Intra-epithelial lymphocyte counts in small intestinal biopsiesfrom children with diarrhoea. Acta Paediatr Scand1976; 65: 541-6.
4 Wright SG, Tomkins AM. Quantification of thelymphocyte infiltrate in jejunal epithelium in giardiasis.Clin Exp Immunol 1977; 29: 408-12.
5 Yardley JH, Takano J, Hendrix TR. Epithelial andother mucosal lesions of the jejunum in giardiasis.Jejunal biopsy studies. Bull Johns Hopkins Hosp 1964;115: 389-406.
6 Barbieri D, De Brito T, Hoshino S et al. Giardiasis inchildhood. Arch Dis Child 1970; 45: 466-72.
7 Wright SG, Tomkins AM, Ridley DS. Giardiasis:clinical and therapeutic aspects. Gut 1977; 18: 343-50.
8 Wright SG, Tomkins AM. Quantitative histology ingiardiasis. J Clin Pathol 1978; 31: 712-16.
9 Roberts-Thompson IC, Stevens DP, Mahmoud AAF,Warren KS. Giardiasis in the mouse: an animal model.Gastroenterology 1976; 71: 57-61.
10 MacDonald IT, Ferguson A. Small intestinal epithelialcell kinetics and protozoal infection in mice. Gastro-enterology 1978; 74: 496-500.
11 Nair KPV, Gillon J, Ferguson A. Corticosteroidtreatment increases parasite numbers in murinegiardiasis. Gut. 1981; 22: 475-80.
12 Clarke RM. Mucosal architecture and epithelial cellproduction in the small intestine of the albino rat. JAnat 1970; 107: 519-29.
13 Wimber DR, Lamerton LF. Cell population studies onthe intestine of continually irradiated rats. Radiat Res1963; 18: 137-46.
14 MacDonald TT, Ferguson A. Hypersensitivity reac-tions in the small intestine. 3. The effects of allograftrejection and graft-versus-host disease on epithelial cellkinetics. Cell Tissue Kinet 1977; 10: 301-12.
15 Ferguson A, Murray D. Quantification of intra-epithelial lymphocytes in human jejunum. Gut 1971;12: 988-94.
16 Dahlquist A. Assay of intestinal disaccharidases.Analyt Biochem 1968; 22: 99-107.
17 Roberts-Thompson IC, Mitchell GF. Giardiasis inmice. I. Prolonged infections in certain mouse strainsand hypothymic (nude) mice. Gastroenterology 1978;75: 42-6.
18 Owen RL, Nemanic P, Stevens DP. Ultrastructuralobservations on giardiasis in a murine model. I.Intestinal distribution, attachment and relationship tothe immune system of Giardia muris. Gastroenterology1979; 76: 757-69.
19 Rey J, Schmitz J, Rey F, Jos J. Cellular differentiationand enzymatic defects. Lancet 1971; 2: 218.
20 Takano J, Yardley JH. Jejunal lesions in patients withgiardiasis and malabsorption. An electron microscopicstudy. Bull Johns Hopkins Hosp 1965; 116: 413-29.
21 Wehman HJ, Lifschitz F, Terchberg S. Effects ofenteric microbial overgrowth on small intestinalultrastructure in the rat. Am J Gastroenterol 1978; 70:249-58.
22 Tandon BN, Tandon RK, Satpathy BK, Shriniwa S.Mechanism of malabsorption in giardiasis: a study ofbacterial flora and bile salt deconjugation in upperjejunum. Gut 1977; 18: 176-81.
23 Tomkins AM, Wright SG, Drasar BS, James WPT.Bacterial colonisation of jejunal mucosa in giardiasis.Trans R Soc Trop Med Hyg 1978; 72: 33-6.
24 Mowat AMcI, Ferguson A. Hypersensitivity reactionsin the small intestine. 6. Pathogenesis of the graft-versus-host reaction in the small intestinal mucosa ofthe mouse. Transplantation 1981; 32: 238-43.
25 Ferguson A. Intraepithelial lymphocytes of the smallintestine. Gut 1977; 18: 921-37.