families of nuclear receptors in vertebrate models ... · families of nuclear receptors in...
TRANSCRIPT

Families of Nuclear Receptors inVertebrate Models: Characteristic andComparative Toxicological PerspectiveYanbin Zhao1, Kun Zhang1, John P. Giesy2,3,4 & Jianying Hu1
1MOE Laboratory for Earth Surface Processes, College of Urban and Environmental Sciences, Peking University, Beijing 100871,China, 2Department of Veterinary Biomedical Sciences and Toxicology Centre, University of Saskatchewan, Saskatoon,Saskatchewan, Canada, 3Department of Zoology, and Center for Integrative Toxicology, Michigan State University, East Lansing,MI, USA, 4Department of Biology & Chemistry and State Key Laboratory in Marine Pollution, City University of Hong Kong, Kowloon,Hong Kong, SAR, China.
Various synthetic chemicals are ligands for nuclear receptors (NRs) and can cause adverse effects invertebrates mediated by NRs. While several model vertebrates, such as mouse, chicken, western clawed frogand zebrafish, are widely used in toxicity testing, few NRs have been well described for most of these classes.In this report, NRs in genomes of 12 vertebrates are characterized via bioinformatics approaches. Althoughnumbers of NRs varied among species, with 40–42 genes in birds to 66–74 genes in teleost fishes, all NRs hadclear homologs in human and could be categorized into seven subfamilies defined as NR0B-NR6A.Phylogenetic analysis revealed conservative evolutionary relationships for most NRs, which were consistentwith traditional morphology-based systematics, except for some exceptions in Dolphin (Tursiopstruncatus). Evolution of PXR and CAR exhibited unexpected multiple patterns and the existence of CARpossibly being traced back to ancient lobe-finned fishes and tetrapods (Sarcopterygii). Compared to themore conservative DBD of NRs, sequences of LBD were less conserved: Sequences of THRs, RARs and RXRswere $90% similar to those of the human, ERs, AR, GR, ERRs and PPARs were more variable withsimilarities of 60%–100% and PXR, CAR, DAX1 and SHP were least conserved among species.
Nuclear receptors (NRs) are one of the largest groups of transcription factors in vertebrates, and serveimportant functions in regulation of a range of physiological functions including growth and differenti-ation of cells, metabolic processes, reproduction, development and overall homeostasis. Transcriptional
activities of NRs are regulated by binding of endogenous small lipophilic compounds1,2. There is growingevidence that diverse chemicals that occur in the environment, including synthetic molecules such as pharma-ceuticals, endocrine disrupting chemicals and some industrial compounds, can mimic endogenous small com-pounds that can bind to ligand binding domains (LBDs), activate NR-mediated signals that then lead to toxicresponses3,4. Typically, interactions of some pesticides and industrial chemicals with estrogen (ER) and androgen(AR) receptors have been linked to a number of adverse effects including birth defects, developmental neuro-toxicity, both male- and female-factor reproductive health, such as decreased quality of sperm, and increasedincidences of cancers5–7.
A series of in vitro bioassays, based on signaling of endocrine receptors including well-studied steroid hormonereceptors such as ER, AR, glucocorticoid receptors (GRs), and progesterone receptor (PR) and the less well-studied retinoic acid receptor (RAR), retinoid X receptor (RXR), and thyroid hormone receptor (THR), have beenestablished or are under assessment by OECD and/or US EPA8–10. Due to their relatively clear physiologicalfunctions and responses to environmentally-relevant organic micropollutants, these NR-based assays have beenused in assessment of toxicological effects of chemicals in the environment. For example, ERs, AR and THRs,involved in development and maintenance of the endocrine system, have been demonstrated to be targets ofalkylphenols, phthalates (PAEs), dichlorodiphenyltrichloroethane and some metabolites of polychlorinatedbiphenyls (PCBs) and polybrominated diphenyl ethers (PBDE)11–13. Besides endocrine receptors, PXR andCAR, NRs that participate in metabolism of both endobiotics and xenobiotics to detoxify or bioactivate chemicals,can be activated by a variety of pharmaceuticals such as rifampicin, pesticides such as chlorpyrifos and methoxy-chlor, and other synthetic chemicals used in industry, such as PBDEs and BPA14–17 In addition to these well-known NRs, there are more NRs, that, during the past decade, have been identified in genomes of several
OPEN
SUBJECT AREAS:
EVOLUTIONARYECOLOGY
ENVIRONMENTAL SCIENCES
Received23 October 2014
Accepted21 January 2015
Published25 February 2015
Correspondence andrequests for materials
should be addressed toY.Z. (zhaoyb@pku.
edu.cn) or J.H. ([email protected])
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 1

vertebrates. These include 48 NR genes in human (Homo sapiens), 47genes in rat (Rattus norvegicus), 49 genes in mouse (Mus musculus)and 68 genes in the teleost puffer fish Fugu rubripes18,19. Specifically,structures of 48 NRs in the human have been identified and categor-ized, based on sequence homology, into seven different subfamiliesNR0B-NR6A20. Except for two NRs in the subfamily NR0B whichlack a DNA binding domain (DBD), all 46 NRs contain the followingsix functional domains: (A–B) variable N-terminal regulatorydomain; (C) conserved DNA-binding domain; (D) variable hingeregion; (E) conserved ligand binding domain (LBD) and (F) variableC-terminal domain20. In addition, sets of NRs described in humansoffered a better understanding of characteristics of NRs, and pro-vided insight for uncovering novel molecular and signal targets andmechanisms of action of synthetic toxicants. For instance, it has beenfound that some widely used pharmaceutical drugs that are found inthe environment, including thiazolidine diones, trichloroacetic acidand toxaphene are ligands for human RORa, PPARa and ERRa,respectively21–23. Compared with the extensive understanding ofNRs in human, fewer NRs have been identified in other vertebratesused as models to screen chemicals for toxic potencies, such as rep-tiles, amphibians and teleost fishes. While in recent years, due toextensive information about their developmental biology andmolecular genetics and now the availability of completed sequencingof their genomes, these vertebrate species have been much used astoxicological models such as western clawed frog (X. tropicalis), zeb-rafish (Danio rerio), and freshwater Japanese medaka (Oryziaslatipes)24–26, information on NRs in these vertebrates were still lim-ited to ERs, AR, GR, PXR, RARs and PPARs, though studies on somenovel NRs, such as VDR, FXR and NURR are in progress27–29.Additionally, since sets of NRs in human, mouse and rat that havebeen identified in previous studies were based on their genomesassembled a decade ago18, there is also a need to reevaluate the char-acteristics of NRs in these genomes due to the constantly updatedsequence data and annotations. In addition to the sequences of gen-omes, predicted transcriptomes and proteomes, now available for allof these species in Genebank and Ensembl, provide useful databasesthat can be further used to uncover and characterize additional NRs.Therefore, comprehensive descriptions of NRs and their families forthese vertebrates used as models to screen for toxic potencies ofchemicals, will be helpful for their further development and inter-pretation of results of studies of synthetic chemicals of envir-onmental significance.
In this study, complete sets of NRs were described for genomes of12 vertebrates used as models in studies of toxic potency andmechanisms of action of chemicals. Several bioinformaticsapproaches were applied to four mammals (human, Homo sapiens;mouse, Mus musculus; rat, Rattus norvegicus and dolphin, Tursiopstruncatus), two birds (chicken, Gallus gallus and mallard (wild duck),Anas platyrhynchos), a reptile (Chinese softshell turtle, Pelodiscussinensis), an amphibian (Western clawed frog, Xenopus tropicalis)and four teleost fishes (zebrafish, Danio rerio; medaka, Oryziaslatipes; tilapia, Oreochromis niloticus and stickleback, Gasterosteusaculeatus). The locations of NRs on chromosomes, phylogeneticanalysis and DBD and LBD sequence conservations among specieswere also analyzed to better understand the characteristics of theseNRs in these vertebrates.
Results and DiscussionIdentification of NRs in 12 vertebrates. Substantial and continuousinformation gathered from developmental biology and moleculargenetics, together with the complete sequencing of genomes hasplaced a series of vertebrate species in attractive positions for usein toxicological research. Twelve species were chosen for descriptionand complete sets of NR genes within their genomes were identifiedby use of a systemic bioinformatics approach. In total, 42–74 NRgenes were uncovered within these vertebrates and a large number of
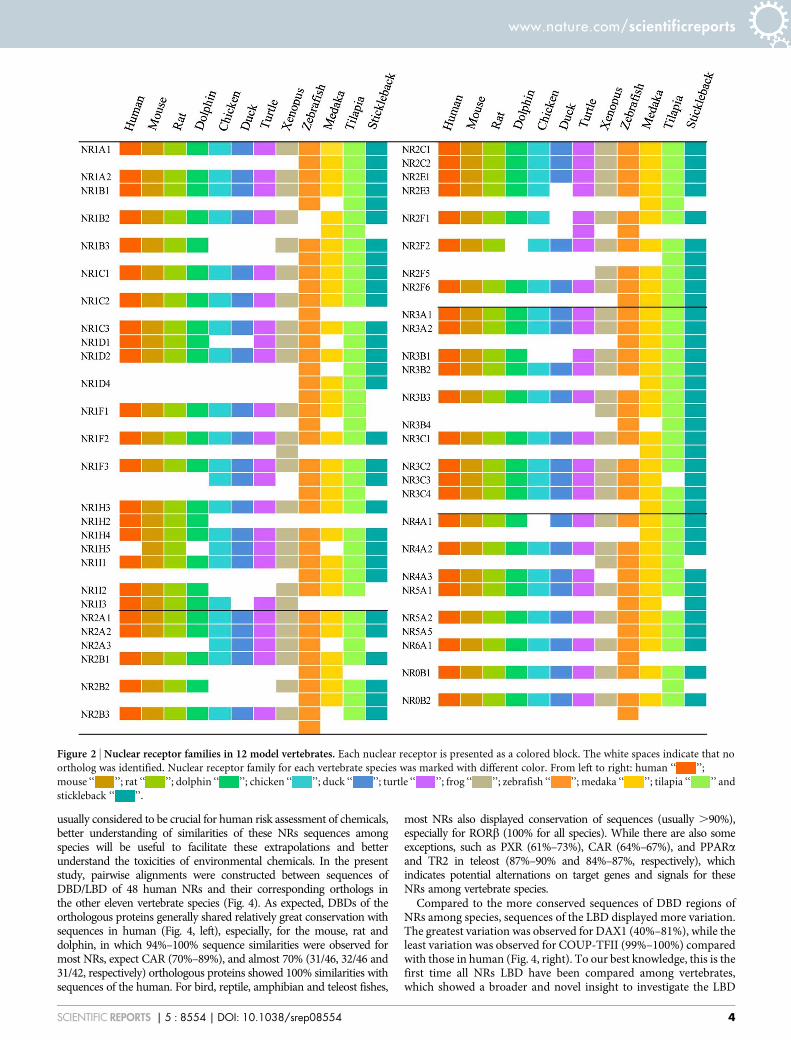
variations were observed among classes (Fig. 1A, Table S2).Comparisons of sequences showed that all of these NRs displayedsignificant similarity to NRs of the human and could be categorizedinto the seven subfamilies NR0B-NR6A, with no novel subfamilies.For mammals, there were 48, 49, 49 and 47 NRs identified in human,mouse, rat and dolphin genomes, respectively (Fig. 1A). Comparedto the human, one more gene (NR1H5) was observed for mouse andrat and one (NR2F2) was absent from dolphin (Fig. 2). Sets of NRs inhuman and mouse were consistent with previous reports18, while twomore NRs (NR1D2 and NR2E3) were newly identified for the rat.The absences of these two NRs in rat in previous study18 were due tothe existence of sequence gaps in the rat genome which wasassembled in 2003.
The numbers of NRs in birds were less than those in human,though there were some unique genes observed. There were sevenNRs (NR1B3, NR1D1, NR1H2, NR1I2, NR2B2, NR3B1 and NR4A1)present in the human that were absent from the chicken. Similarly,there were nine NRs (NR1B3, NR1D1, NR1H2, NR1I2, NR1I3,NR2B2, NR2E3, NR2F1 and NR3B1) present in the human that wereabsent from the mallard, though there were three new NRs (NR1F3,NR1H5 and NR2A3) were identified that were unique to chicken andmallard (Fig. 2). Similar absences were observed in the genomes ofturkey (Meleagris gallopavo), flycatcher (Ficedula albicollis) andzebra finch (Taeniopygia guttata), where 9, 5 and 6 NRs, respectively,that are present in the human genome were absent from these birds(Fig. 3C). These results demonstrated that a cluster of NRs wereindeed absent from genomes of the class aves, especially in galloan-serae, that were deleted during the course of evolution.
Some NRs present in the human were absent from turtle andwestern clawed frog while some others were unique in these species.In the one species of turtle, 48 NRs were identified with four genesabsent (NR1B3, NR1H2, NR1I2 and NR2B2) and four new genesgained (NR1F3, NR1H5, NR2A3 and NR2F1) compared with thosein human. Similarly, 52 NRs were identified in western clawed frogwith 2 genes absent (NR1H2 and NR4A3) and six additional genes(NR1F2, NR1H5, NR2A3, NR2F5, NR3B3 and NR4A2) appearedwhich were not present in the human (Fig. 2).
For the four teleost fishes studied, there were many additional NRsuncovered in this study. Specifically, 73 and 74 NRs were identified inzebrafish and tilapia, respectively (Fig. 1A), which were consistent withthose reported for Fugu rubripes (68 NRs identified)19. The additionalNRs were mainly due to the paralogue genes exist in their sets of NRs(Fig. 1C). In zebrafish, two or more paralogues were identified tocorrespond with one of 20 NRs in human and with one of 18, 22and 17 NRs in medaka, tilapia and stickleback, respectively. Existencesof paralogue genes in teleost fishes were not random but focused onsome specific NR units. For instance, NR1F3 (RORc) was the mostabundant NR, with a total of seven paralogue gene copies in these fourteleost fishes. The NRs NR1A1, NR1B3, NR1C1, NR1I1, NR2B2,NR2F6, NR3A2, and NR3B3 were also rich in paralogues, with oneparalogue gene copy in each of the four teleosts (Fig. 3D).
Characteristics of NRs families. Genomic locations of NRs in sevenvertebrate genomes (human, mouse, rat, chicken, zebrafish, medakaand stickleback) were retrieved via the Ensemble annotations. Ingeneral, distributions of NRs on chromosomes were morewidespread in teleost fishes than those of mammals and birds(Fig. 1B). This is possibly due to the existence of more paraloguegenes in teleosts. For example, NRs in zebrafish, medaka andstickleback were distributed throughout their genomes except for1–2 chromosomes. The most abundant clusters of NRs wereobserved on chromosomes 8 and 16 in zebrafish, each with 6 NRs;on chromosomes 7 and 16 in medaka, each with 7 NRs; and onchromosome 12 in stickleback, with 8 NRs. The narrowestdistribution of NRs was observed for species of chicken, in which44 NRs were distributed in 61% (19/31) chromosomes.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 2

Phylogenetic analyses, based on their full amino acid sequencesand DBD plus LBD compositions of NRs, were performed for 48types of NRs among these 12 vertebrates. The Neighbor-Joining (NJ)and Maximum-Likelihood (ML) phylogenetic analyses showed sim-ilar patterns, while the Neighbor-Joining algorithm gave better reso-lution at the base of the phylogram. Conservative evolutionaryrelationships were observed for most NRs, i.e. the evolutionary rela-tionships were generally consistent with the traditional morphology-based systematics (Fig. S1). As exemplified for NR3A1 (ERa), closerrelationships were observed within each class and the traditionalteleost-amphibian-reptile-bird and mammal evolutionary relation-ships were followed (Fig. 3A). This was verified by the similarity ofsequences of the LBD of ERa among species (Fig. 4). In details, about82–93% sequence similarities among teleost, 99% between birds and98–99% among mammals was observed and the sequence similaritiesamong classes were relatively small (Fig. 5). Some exceptions wereobserved in Dolphin such as NR2A1 and NR2A2 (Fig. S1). Thoughdolphin, diverged from artiodactyls approximately 50 million yearsago30, was thought to show the closest relationship with humanamong the 12 vertebrates, there were 32% NRs that showed closerrelationships between rodents and human compared with those indolphin. Similarities between sequences of the DBD and LBD alsoconfirmed this likely historical divergence. In rodents, 13% ofsequences of amino acids of DBD and 26% of those of the LBDexhibited relationships more similar to those of the human thandolphin (Fig. 3B). These variations in NRs in dolphin were possiblydue to the results of positive Darwinian selection, the major drivingforce for adaptive evolution and diversification among species, toadapt their radical habitat transition from land to a marine envir-onment. Though increasing toxicological research has been pre-formed using dolphins and extrapolations from dolphin to humanwere thought to be more significant, results of the present study
demonstrated more variations, indicating more genetic characteris-tics should be taken into account when assessing toxicities of chemi-cals based on results of studies with dolphins. In addition, since PXRand CAR displayed the largest variations and were absent in severalvertebrates used in this study (Fig. 2 and 4), more comparisonsamong species were conducted. Existence of NR1I (VDR, PXR andCAR) genes were demonstrated in 35 vertebrate species (20 mam-mals, 5 birds, 2 reptile, 1 amphibian and 7 teleost fishes) with forwhich complete sequences of genomes were available and unexpec-ted patterns were showed for their evolutions. VDR genes appearedin all vertebrate genomes, a result which was consistent with those inprevious reports that VDR could be detected in mammals, birds,amphibians, reptiles, teleost fishes, and even the sea lamprey31.PXR appeared in most teleost fishes (expect for stickleback), amphi-bians and mammals (also known as SXR), but were totally absentfrom reptiles and birds. Though CAR also appeared in all mammals,it exhibited quite different patterns in other classes. CAR was mostlyabsent in birds (expect for chicken), but retained in reptiles andamphibians, and appeared in lobe-finned fishes and tetrapods(Sarcopterygii) (Fig. 3E). Since Sarcopterygii appeared nearly 400million years ago during the Devonian, and are widely accepted asancestors of all tetrapoda, including amphibians, reptiles, birds andmammals32, the appearance of CAR in Sarcopterygii possibly indi-cated that the existence of CAR was much earlier than previouslythought. In general, these results revealed a novel evolutionary rela-tionship for PXR/CAR. These two NRs likely coexisted in ancientSarcopterygii, first due to the duplication events, descended intoamphibians and then to mammals, but one of them was absent fromreptiles and both were absent from most birds (Fig. S2).
Alignment of sequences of DBD and LBD. Since cross-speciesextrapolations from surrogate vertebrate species to humans are
Figure 1 | Identification of NRs in genomes of 12 toxicological vertebrate models. (A) Total number of NRs in each vertebrate genome (B) the
genomic distributions of NRs in seven vertebrate species (C) the number of NRs for each type (NR0B-NR6A) and the paralogous gene numbers (P.G.) in
total.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 3
usually considered to be crucial for human risk assessment of chemicals,better understanding of similarities of these NRs sequences amongspecies will be useful to facilitate these extrapolations and betterunderstand the toxicities of environmental chemicals. In the presentstudy, pairwise alignments were constructed between sequences ofDBD/LBD of 48 human NRs and their corresponding orthologs inthe other eleven vertebrate species (Fig. 4). As expected, DBDs of theorthologous proteins generally shared relatively great conservation withsequences in human (Fig. 4, left), especially, for the mouse, rat anddolphin, in which 94%–100% sequence similarities were observed formost NRs, expect CAR (70%–89%), and almost 70% (31/46, 32/46 and31/42, respectively) orthologous proteins showed 100% similarities withsequences of the human. For bird, reptile, amphibian and teleost fishes,
most NRs also displayed conservation of sequences (usually .90%),especially for RORb (100% for all species). While there are also someexceptions, such as PXR (61%–73%), CAR (64%–67%), and PPARaand TR2 in teleost (87%–90% and 84%–87%, respectively), whichindicates potential alternations on target genes and signals for theseNRs among vertebrate species.
Compared to the more conserved sequences of DBD regions ofNRs among species, sequences of the LBD displayed more variation.The greatest variation was observed for DAX1 (40%–81%), while theleast variation was observed for COUP-TFII (99%–100%) comparedwith those in human (Fig. 4, right). To our best knowledge, this is thefirst time all NRs LBD have been compared among vertebrates,which showed a broader and novel insight to investigate the LBD
Figure 2 | Nuclear receptor families in 12 model vertebrates. Each nuclear receptor is presented as a colored block. The white spaces indicate that no
ortholog was identified. Nuclear receptor family for each vertebrate species was marked with different color. From left to right: human ‘‘ ’’;
mouse ‘‘ ’’; rat ‘‘ ’’; dolphin ‘‘ ’’; chicken ‘‘ ’’; duck ‘‘ ’’; turtle ‘‘ ’’; frog ‘‘ ’’; zebrafish ‘‘ ’’; medaka ‘‘ ’’; tilapia ‘‘ ’’ and
stickleback ‘‘ ’’.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 4

differences between species and between multiple NRs units. In thepresent study, three groups were identified in general based on sim-ilarities in sequences of NRs. The first group contained 13 NRsincluding THRa, THRb, RARa, RARb, RARc, RORa, RXRa,RXRb, RXRc, COUP-TFII, ERRc, NURR1 and LRH1 (except someorthologs for RARa, RORa, RXRb, RXRc and NURR1) with $90%similarity of sequences of the LBDs for all eleven vertebrates com-pared with those of the human (Fig. 4, right). As observed for RXRa,97–100% similarities in sequences, for the best alignment orthologs,were observed from multiple sequence alignment (Fig. 5). Variationsin conservation of sequences, window averaged across 10 amino acidresidues, found that there were fewer than 5 variations in amino acidresidues among these 12 vertebrate species, and most of them wereobserved in a-helix 3 to a-helix 6 of the LBD structures (Fig. 5).RXRa commonly functions as a heterodimers with other NRs andmainly mediates signaling of hormones derived from vitamin A(retinol) such as 9-cis retinoic acid, and are involved in multiplephysiological functions of vertebrates such as embryonic patterningand organogenesis, proliferation of cells and differentiation of tis-sues33. It has been reported that among vertebrates, such as mouse
and human, LBDs of RXRa interacted with similar types of ligandswith similar binding affinities34,35. Sequence similarities of these 13NRs among vertebrates suggested potential straightforward interspe-cies extrapolations when assessing toxicity of chemicals via theseNRs. Approximately 77% of NRs such as the well-known ERs, AR,PR, PPARs and VDR can be sorted into the second group, exhibiting60–100% similarities of sequences (for the best aligned orthologs)compared with those of human. Similarities in sequences of theseNRs among four fishes were substantially the same and usually$90% in mouse, rat and dolphin, showing apparent differences insequences of amino acids between teleosts and mammals.Specifically, LBDs of NRs in the second group, such as ERa andPPARc, always shared the same variations in amino acids withinfour fishes, which were quite different from those of mammals(Fig. 5 for ERa). ERa is a well-studied NR, activated by endogenousand exogenous estrogens, and plays a variety of central physiologicalroles, such as maintenance of reproductive, cardiovascular and cent-ral nervous systems in vertebrates36. Potencies of binding of ligandsto LBDs of ERa were different for fishes when compared to mam-mals. It has been reported that widespread chemicals like 4-t-octyl-
Figure 3 | Characteristics of the 12 NRs families. (A) Phylogenetic tree for 12 NR3A1 (ERa) genes (B) The evolutionary relationships of NRs among
dolphin, rodents and human species. Left: the proportions of dolphin NRs with closer relationships with human compared to rodents are presented as
percent/number and blue colour. The proportions of rodents NRs with closer relationships with human are presented as percent/number and orange
colour. Green colour represents the NRs numbers with equivalent sequence similarities with human for dolphin and rodents. Right: phylogenetic tree for
NR2C1 and NR2A1 represents the different positions of NRs for dolphin. (C) Comparative searches for the ten lacked NRs in five bird species (D)
Paralogous gene copy numbers for each type of NRs (E) Comparative searches for NR1I genes (VDR, PXR and CAR) in 35 vertebrates, including 20
mammals, 5 birds, 2 reptiles, 1 amphibian and 7 teleost (details are described in Table S4). Phylogenetic tree was developed utilizing 35 full amino acid
sequences of VDR.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 5

Figure 4 | Pairwise alignments between DBD/LBD amino acid sequences of 48 human NRs and the corresponding orthologs in other eleven vertebratespecies. Left for the DBD sequence comparisons and right for the LBD. The sequence similarities are presented as the percentage (%) and relevant
color. NRs, with incomplete amino acid sequences of DBD/LBD, were not included in this comparison.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 6

phenol and bisphenol A (BPA) bound with greater avidity to rainbowtrout ER than that of human or rat. Also, types of ligands werevarious: of 34 chemicals tested, 29 can bind to ER of rainbow trout,while only 20 of them can bind to ER of human/rat37. PPARc is also awell-studied transcription factor, which could be activated by fattyacids and is involved in lipid and glucose metabolism38. Reports onbinding strengths of LBDs for PPARc were rare, but interspeciesextrapolations on LBD binding activities can be likely to estimate,due to the similar sequence characteristics between PPARc and ERa.
In the third group, with less than 85% similarities in sequences ofeleven vertebrate species compared with those in human, four NRsincluding PXR, CAR, DAX1 and SHP (Fig. 4) were classified as beingdifferent from human. DAX1 and SHP, which belong to the subfam-ily NR0B, displayed the greatest variations among NRs and amongvertebrates (Fig. 4 and 5), a result which is consistent with thosereported previously that NRs in the NR0B group were a unique classof NRs with among-species variability in sequences and lacking DBDdomains18. PXR and CAR were also assigned to this group, and
Figure 5 | Variations in LBD sequence conservation across the sequence of RXRa, ERa and SHP. Left: LBD sequences for eleven vertebrates compared
to the related human nuclear receptors. All sequences were window averaged across 10 residues. Right: multiple sequence alignments among the 12
vertebrates. The sequence similarities are presented as the percentage (%) and relevant color. The LBD sequence of ERa in Dolphin was not included in
this comparison due to the incomplete amino acid sequences.
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 7

exhibited apparent differences among vertebrates and even amongfishes. PXR and CAR can be activated by xenobiotics and have rela-tively broad abilities to bind ligands39. The unusually great diversityin sequences of the LBD among species could be related to diversityin binding activities among species. This is exemplified by the factthat phenobarbital, a pharmaceutical that is generally detectable ineffluents of municipal waste water plants (WWTP), was a moderateactivator of the zebrafish PXR and exhibited greater binding affinitywith human PXR, while it did not bind to PXR of mouse39. Thesedifferences among species might be due to the differences in diet andphysiology among vertebrates, and such largely differences ofsequences of PXR and CAR among vertebrates complicated the insilico extrapolations.
Here, for the first time, genes that code for NRs and their relativecharacteristics are provided for 12 vertebrate species used as modelanimals in screening of toxic potencies of chemicals. These resultswill help understanding of the NRs in vertebrates and will be usefulfor clarifying mechanisms of toxic effects of environmental chemi-cals on these model species and also the extrapolations from theeffects on these surrogates to human.
MethodsIdentification of NRs in 12 vertebrate genomics. Identification of sequences forNRs was performed as described previously40,41 with slight modifications. In brief, theputative NRs for each vertebrate were identified through a combination of BLASTnand BLASTp searches of the genome and protein databases, which were obtainedfrom NCBI and Ensembl. The nucleotide and protein sequences of 165 described NRsin three vertebrates (48 in human, 49 in mouse and 68 in Fugu rubripes) weredownloaded from GenBank and used as templates for interrogating the vertebratedatabases. Nucleotide homology searches were performed using the full nucleotidesequences of each of the 165 NRs against these 12 genomic sequences database atNCBI by use of nucleotide BLAST with a blastn algorithm and an e value cut off of 1e-04. Protein sequences were then used to construct multiple sequence alignments byClustalX2 (http://www.clustal.org/clustal2/) and then the DNA-binding domain(DBD) and the ligand-binding domain (LBD) amino acid sequences weredemonstrated. BLASTp searches were performed using the conserved DBD plus LBDdomains against the non-redundant vertebrate protein sequence database at NCBI byuse of protein BLAST with a blastp algorithm and an e value cut off of 1e-25. The ecut-off values were set to be just loose enough to find all the Fugu NRs when usinghuman NRs as queries. Genes identified by BLASTn and BLASTp searches were thencombined and individual putative genes were sorted according their unique DNA andamino acid sequences. All these putative genes were verified by online softwareNRpred and iNR-PhysChem to remove the false-positive hits, and the NR0B1 andNR0B2, which are known to lack the DBD region, were added to the final sets of NRs.Details for the sequence searches were shown in Table S1. Finally, complete sequencesfor each NR in each vertebrate species were loaded into Ensembl database. Thenomenclatures of NRs were based on Ensembl’s GeneTree and Orthologyannotations.
Genomic distributions. Genomic location for each nuclear receptor in sevenvertebrate genomes (human, mouse, rat, chicken, zebrafish, medaka and stickleback)were retrieved via the Ensembl annotations, and then mapped onto completevertebrate karyograms.
Analyses of sequences of DBD and LBD. Sequences of peptides in the DBD and LBDdomains for each NR were identified by use of Pfam software (http://pfam.sanger.ac.uk/, Pfam 27.0) and modified manually, based on characteristics of DBD and LBDregions reported previously. The sequence of DBD, which is classified as a type-II zincfinger motif, corresponds to a 75–80 amino acid residue segment, starting at thelocation of two amino acid residues before the first conserved cysteine andencompassing both C4 zinc fingers and the LBD, a flexible unit made of a-helicescontaining of 170 to 210 amino acid residues, begin at the 12th residue of a-helix 3and extended through a-helix 1042,43.
The pairwise alignments between sequences of the DBD and LBD of humanprotein and corresponding orthologs in the other 11 vertebrates were constructed byuse of the NCBI BLASTp software with default parameters. Similarities in sequenceswere calculated based on the numbers of identical residues over the total numbers ofaligned residues in human.
Phylogenetic analysis. Phylogenetic trees were constructed by use of amino acidsequences of 48 types of NRs downloaded from Ensembl based on the set ofhomologous NRs in the human. Only full- length molecules were included for theanalysis. Some genes without complete amino acid sequences in the Ensembl databasewere retrieved from NCBI/EMBL/DDBJ databases (Table S3). They were alsoincluded. The Ensembl ID of each NR used in the analyses is available in SI Table S2.Conserved sequences of DBD and LBD for each NR were also isolated and used as asupportive analysis. Sequences of DBD and LBD were combined and then aligned,
except for NR0B1 and NR0B2. Multiple alignments of sequences of amino acids weregenerated by use of ClustalX2 software with default parameters, and the results usedfor construction of phylogenetic trees by implementation of the Neighbour-Joiningand Maximum-Likelihood algorithms with a Poisson model in MEGA6 software(http://www.megasoftware.net/mega.php). Confidence for branching patterns wasassessed by bootstrap analysis (1000 replicates). For NR1I1 (VDR) analysis, the fullamino acid sequences of NR1I1 in 35 vertebrates, including 20 mammals, 5 birds, 2reptiles, 1 amphibian and 7 teleost fishes (Table S4), were downloaded from theEmsenbl database. These full amino acid sequences were then aligned and applied forgene phylogenetic analysis by use of the same method described above.
1. Mangelsdorf, D. J. et al. The nuclear receptor superfamily: the second decade. Cell83, 835–839 (1995).
2. Pardee, K., Necakov, A. S. & Krause, H. Nuclear receptors: small molecule sensorsthat coordinate growth, metabolism and reproduction. Subcell. Biochem. 52,123–153 (2011).
3. Janosek, J., Hilscherova, K., Blaha, L. & Holoubek, I. Environmental xenobioticsand nuclear receptors - interactions, effects and in vitro assessment. Toxicol. inVitro 20, 18–37 (2006).
4. Grun, F. & Blumberg, B. Environmental obesogens: organotins and endocrinedisruption via nuclear receptor signaling. Endocrinology 147, S50–S55 (2006).
5. Damstra, T., Barlow, S., Bergman, A., Kavlock, R. & Van Der Kraak, G. eds.International programme on chemical safety global assessment: the state-of-the-science of endocrine disruptors. Geneva: World Health Organization (2002).Available at: http://www.who.int/ipcs/publications/new_issues/endocrine_disruptors/en/ (Accessed: 23th December 2014).
6. Huang, R. et al. Chemical genomics profiling of environmental chemicalmodulation of human nuclear receptors. Environ. Health. Perspect. 119,1142–1148 (2011).
7. Toppari, J. et al. Male reproductive health and environmental xenoestrogens.Environ. Health. Perspect. 104, 741–803 (1996).
8. Kortenkamp, A. et al. State of the art assessment of endocrine disrupters, finalreport. 2011. Available at: http://ec.europa.eu/environment/chemicals/endocrine/documents/studies_en.htm (Accessed: 23th December 2014).
9. Kavlock, R. et al. Update on EPA’s ToxCast Program: Providing high throughputdecision support tools for chemical risk management. Chem. Res. Toxicol. 25,1287–1302 (2012).
10. Martin, M. T. et al. Impact of environmental chemicals on key transcriptionregulators and correlation to toxicity end points within EPA’s ToxCast program.Chem. Res. Toxicol. 23, 578–590 (2010).
11. Kuiper, G. G. J. M. et al. Interaction of estrogenic chemicals and phytoestrogenswith estrogen receptor beta. Endocrinology 139, 4252–4263 (1998).
12. Sonnenschein, C. & Soto, A. M. An updated review of environmental estrogen andandrogen mimics and antagonists. J. Steroid Biochem. Mol. Biol. 65, 143–150(1998).
13. Zoeller, R. T. Environmental chemicals as thyroid hormone analogues: newstudies indicate that thyroid hormone receptors are targets of industrialchemicals? Mol. Cell. Endocrinol. 242, 10–15 (2005).
14. Jacobs, M. N., Nolan, G. T. & Hood, S. R. Lignans, bacteriocides andorganochlorine compounds activate the human pregnane X receptor (PXR).Toxicol. Appl. Pharmacol. 209, 123–133 (2005).
15. Chang, T. K. & Waxman, D. J. Synthetic drugs and natural products as modulatorsof constitutive androstane receptor (CAR) and pregnane X receptor (PXR). DrugMetab. Rev. 38, 51–73 (2006).
16. Zhao, Y. B., Luo, K., Fan, Z. L., Huang, C. & Hu, J. Y. Modulation of benzo [a]pyrene-induced toxic effects in Japanese medaka (Oryzias latipes) by 2, 29, 4, 49-tetrabromodiphenyl ether. Environ. Sci. Technol. 47, 13068–13076 (2013).
17. DeKeyser, J. G., Laurenzana, E. M., Peterson, E. C., Chen, T. & Omiecinski, C. J.Selective phthalate activation of naturally occurring human constitutiveandrostane receptor splice variants and the pregnane X receptor. Toxicol. Sci. 120,381–391 (2011).
18. Zhang, Z. et al. Genomic analysis of the nuclear receptor family: new insights intostructure, regulation, and evolution from the rat genome. Genome Res. 14,580–590 (2004).
19. Maglich, J. M. et al. The first completed genome sequence from a teleost fish (Fugurubripes) adds significant diversity to the nuclear receptor superfamily. NucleicAcids Res. 31, 4051–4058 (2003).
20. Germain, P., Staels, B., Dacquet, C., Spedding, M. & Laudet, V. Overview ofnomenclature of nuclear receptors. Pharmacol. Rev. 58, 685–704 (2006).
21. Missbach, M. et al. Thiazolidine diones, specific ligands of the nuclear receptorretinoid Z receptor/retinoid acid receptor related orphan receptor alpha withpotent antiarthritic activity. J. Biol. Chem. 271, 13515–13522 (1996).
22. Maloney, E. K. & Waxman, D. J. Trans -activation of PPARa and PPARc bystructurally diverse environmental chemicals. Toxicol. Appl. Pharmacol. 161,209–218 (1999).
23. Yang, C. & Chen, S. Two organochlorine pesticides, toxaphene and chlordane, areantagonists for estrogen-related receptor alpha-1 orphan receptor. Cancer Res. 59,4519–4524 (1999).
24. Showell, C. & Conlon, F. L. The western clawed frog (Xenopus tropicalis): anemerging vertebrate model for developmental genetics and environmental
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 8

toxicology. Cold Spring Harb. Protoc. 2009, pdb.emo131 (2009); DOI:10.1101/pdb.emo131.
25. Hill, A. J., Teraoka, H., Heideman, W. & Peterson, R. E. Zebrafish as a modelvertebrate for investigating chemical toxicity. Toxicol. Sci. 86, 6–19 (2005).
26. Ankley, G. T. & Johnson, R. D. Small fish models for identifying and assessing theeffects of endocrine-disrupting chemicals. Inst. Lab Anim. Res. 45, 469–483(2004).
27. Ciesielski, F., Rochel, N., Mitschler, A., Kouzmenko, A. & Moras, D. Structuralinvestigation of the ligand binding domain of the zebrafish VDR in complexeswith 1alpha, 25(OH)2D3 and Gemini: purification, crystallization andpreliminary X-ray diffraction analysis. J. Steroid Biochem. Mol. Biol. 89–90, 55–59(2004).
28. Howarth, D. L. et al. Two farnesoid X receptor a isoforms in Japanese medaka(Orzias latipes) are differentially activated in vitro. Aquat. Toxicol. 98, 245–255(2010).
29. Kapsimali, M., Bourrat, F. & Vernier, P. Distribution of the orphan nuclearreceptor Nurr1 in medaka (Oryzias latipes): cues to the definition of homologouscell groups in the vertebrate brain. J. Comp. Neurol. 431, 276–292 (2001).
30. Meredith, R. W. et al. Impacts of the cretaceous terrestrial revolution and KPgextinction on mammal diversification. Science 334, 521–524 (2011).
31. Whitfield, G. K. et al. Cloning of a functional vitamin D receptor from the lamprey(Petromyzon marinus), an ancient vertebrate lacking a calcified skeleton andteeth. Endocrinology 144, 2704–2716 (2003).
32. Georges, D. & Blieck, A. Rise of the earliest tetrapods: an early Devonian originfrom marine environment. PLoS ONE 6, e221362011 (2011); DOI:10.1371/journal.pone.0022136.
33. Szanto, A. et al. Retinoid X receptors: X-ploring their (patho) physiologicalfunctions. Cell Death Differ. 11, S126–S143 (2004).
34. Heyman, R. A. et al. 9-cis retinoic acid is a high affinity ligand for the retinoid Xreceptor. Cell 68, 397–406 (1992).
35. Mangelsdorf, D. J. et al. Characterization of three RXR genes that mediate theaction of 9-cis retinoic acid. Genes Dev. 6, 329–344 (1992).
36. Heldring, N. et al. Estrogen receptors: How do they signal and what are theirtargets. Physiol. Rev. 87, 905–931 (2007).
37. Matthews, J., Celius, T., Halgren, R. & Zacharewski, T. Differential estrogenreceptor binding of estrogenic substances: a species comparison. J. SteroidBiochem. Mol. Biol. 74, 223–234 (2000).
38. Lee, C. H., Olson, P. & Evans, R. M. Minireview: lipid metabolism, metabolicdiseases, and peroxisome proliferator-activated receptors. Endocrinology 144,2201–2207 (2003).
39. Moore, L. B. et al. Pregnane X receptor (PXR), constitutive androstane receptor(CAR), and benzoate X receptor (BXR) define three pharmacologically distinctclasses of nuclear receptors. Mol. Endocrinol. 16, 977–986 (2002).
40. Thomson, S. A., Baldwin, W. S., Wang, Y. H., Kwon, G. & Leblanc, G. A.Annotation, phylogenetics, and expression of the nuclear receptors in Daphniapulex. BMC Genomics 10, 500 (2009); DOI:10.1186/1471-2164-10-500.
41. Vogeler, S., Galloway, T. S., Lyons, B. P. & Bean, T. P. The nuclear receptor genefamily in the Pacific oyster, Crassostrea gigas, contains a novel subfamily group.BMC Genomics 15, 369 (2014); DOI:10.1186/1471-2164-15-369.
42. Wurtz, J. M. et al. A canonical structure for the ligand-binding domain of nuclearreceptors. Nat. Struct. Biol. 3, 87–94 (1996).
43. Greschik, H. et al. Characterization of the DNA-binding and dimerizationproperties of the nuclear orphan receptor germ cell nuclear factor. Mol. Cell Biol.19, 690–703 (1999).
AcknowledgmentsThis study supported by the National Natural Science Foundation of China [41330637 and41171385] and the 111 Project (B14001). Prof. Giesy was supported by the Canada ResearchChair program, a Visiting Distinguished Professorship in the Department of Biology andChemistry and State Key Laboratory in Marine Pollution, City University of Hong Kong.
Author contributionsY.B.Z. and J.Y.H. designed the experiments, Y.B.Z. and K.Z. performed the experiment andanalyzed the data, Y.B.Z., K.Z., J.P.G. and J.Y.H. wrote the manuscript. All authorscontributed to scientific discussions of the manuscript.
Additional informationSupplementary information accompanies this paper at http://www.nature.com/scientificreports
Competing financial interests: The authors declare no competing financial interests.
How to cite this article: Zhao, Y., Zhang, K., Giesy, J.P. & Hu, J. Families of NuclearReceptors in Vertebrate Models: Characteristic and Comparative Toxicological Perspective.Sci. Rep. 5, 8554; DOI:10.1038/srep08554 (2015).
This work is licensed under a Creative Commons Attribution 4.0 InternationalLicense. The images or other third party material in this article are included in thearticle’s Creative Commons license, unless indicated otherwise in the credit line; ifthe material is not included under the Creative Commons license, users will needto obtain permission from the license holder in order to reproduce the material. Toview a copy of this license, visit http://creativecommons.org/licenses/by/4.0/
www.nature.com/scientificreports
SCIENTIFIC REPORTS | 5 : 8554 | DOI: 10.1038/srep08554 9

Supplementary information for: 1
Families of Nuclear Receptors in Vertebrate Models: Characteristic and Comparative 2
Toxicological Perspective 3
Yanbin Zhao1, Kun Zhang
1, John P. Giesy
2,3,4, and Jianying Hu
1 4
1MOE Laboratory for Earth Surface Processes, College of Urban and Environmental Sciences, 5
Peking University, Beijing 100871, China 6
2Department of Veterinary Biomedical Sciences and Toxicology Centre, University of 7
Saskatchewan, Saskatoon, Saskatchewan, Canada 8
3Department of Zoology, and Center for Integrative Toxicology, Michigan State University, East 9
Lansing, MI, USA 10
4Department of Biology & Chemistry and State Key Laboratory in Marine Pollution, City 11
University of Hong Kong, Kowloon, Hong Kong, SAR, China 12
13
Address for Correspondence 14
Dr. Yanbin Zhao; Prof. Dr. Jianying Hu 15
College of Urban and Environmental Sciences 16
Peking University, Yi Fu Second Building 17
Beijing 100871 China 18
TEL & FAX: 86-10-62765520 19
Email: [email protected]; [email protected] 20

Figure S1. Phylogenetic analysis for 48 types of nuclear receptor genes in twelve vertebrate 21
species. Numbers at branches indicate the bootstrap probabilities (≥90%) with 1,000 replicates. 22
Neighbour-Joining trees of ClustalX-aligned full amino acid/DBD plus LBD sequences were 23
constructed and displayed for the majority of NRs. For some trees, which displayed better 24
topological structures in Maximum-Likelihood analysis, the ML trees were constructed instead. 25
26
99
100
100 95
93
Human_0B1
Dolphin_0B1
Mouse_0B1
Rat_0B1
Chicken_0B1
Duck_0B1
Xenopus_0B1
Turtle_0B1
Tilapia_0B1a
Zebrafish_0B1
Stickleback_0B1
Medaka_0B1
Tilapia_0B1b
99
Human_0B2
Dolphin_0B2
Mouse_0B2
Rat_0B2
Chicken_0B2
Duck_0B2
Turtle_0B2
Zebrafish_0B2a
Tilapia_0B2
Medaka_0B2
Stickleback_0B2
100
100
100
100
90 Human_1A1
Mouse_1A1
Rat_1A1
Chicken_1A1
Xenopus_1A1
Zebrafish_1A1b
Zebrafish_1A1a
Medaka_1A1a
Tilapia_1A1a
Stickleback_1A1b
Tilapia_1A1b
Medaka_1A1b
Stickleback_1A1a
100
98 98
99 99
Human 1A2
Dolphin 1A2
Mouse 1A2
Rat 1A2
Duck 1A2
Chicken 1A2
Turtle 1A2
Xenopus 1A2
Zebrafish 1A2
Medaka 1A2
Tilapia 1A2
Stickleback 1A2
96
100
99 99
Human_1B1
Dolphin_1B1
Rat_1B1
Mouse_1B1
Chicken_1B1
Xenopus_1B1
Zebrafish_1B1a
Medaka_1B1
Tilapia_1B1a
Zebrafish_1B1b
Tilapia_1B1b
Stickleback_1B1a100
98
90
Human_1B2
Rat_1B2
Mouse_1B2
Chicken_1B2
Duck_1B2
Turtle_1B2
Xenopus_1B2
Medaka_1B2b
Tilapia_1B2b
Medaka_1B2a
Tilapia_1B2a
Stickleback_1B2
96
99
100
Dolphin_1B3
Human_1B3
Rat_1B3
Mouse_1B3
Xenopus_1B3
Medaka_1B3b
Tilapia_1B3b
Stickleback_1B3a
Zebrafish_1B3a
Zebrafish_1B3b
Medaka_1B3a
Tilapia_1B3a
Stickleback_1B3b
100
97
Human_1C1
Dolphin_1C1
Mouse_1C1
Rat_1C1
Chicken_1C1
Duck_1C1
Turtle_1C1
Xenopus_1C1
Medaka_1C1a
Zebrafish_1C1b
Tilapia_1C1a
Stickleback_1C1b
Zebrafish_1C1a
Stickleback_1C1a
Medaka_1C1b
Tilapia_1C1b
100
100 100
100 100 100
94 100
Human_1C2
Dolphin_1C2
Mouse_1C2
Rat_1C2
Turtle_1C2
Chicken_1C2
Duck_1C2
Zebrafish_1C2a
Zebrafish_1C2b
Stickleback_1C2
Medaka_1C2
Tilapia_1C2
Xenopus_1C2
99
100 90
97 99
99

27 Human_1C3
Dolphin_1C3
Mouse_1C3
Rat_1C3
Chicken_1C3
Duck_1C3
Turtle_1C3
Xenopus_1C3
Zebrafish_1C3
Medaka_1C3
Tilapia_1C3
Stickleback_1C3
100
100 100
100 98
99
Human 1D2
Dolphin 1D2
Mouse 1D2
Rat 1D2
Chicken 1D2
Duck 1D2
Turtle 1D2
Xenopus 1D2
Zebrafish 1D2b
Tilapia 1D2b
Stickleback 1D2a
Zebrafish 1D2a
Medaka 1D2
Tilapia 1D2a
Stickleback 1D2b
100 100
100 100 100
100
100
100 100
100 98
99
Human_1D1
Mouse_1D1
Rat_1D1
Dolphin_1D1
Xenopus_1D1
Zebrafish_1D1
Tilapia_1D1
Stickleback_1D1
100
100
98
100
Human 1F2
Mouse 1F2
Rat 1F2
Dolphin 1F2
Duck 1F2
Chicken 1F2
Turtle 1F2
Xenopus 1F2b
Zebrafish 1F2
Medaka 1F2
Tilapia 1F2
Stickleback 1F2
98
97
100
100
Human 1H3
Dolphin 1H3
Mouse 1H3
Rat 1H3
Chicken 1H3
Duck 1H3
Turtle 1H3
Xenopus 1H3
Stickleback 1H3
Zebrafish 1H3
Medaka 1H3
Tilapia 1H3
100
100
100
100
100
99
95 99
Human 1H4
Dolphin 1H4
Mouse 1H4
Rat 1H4
Chicken 1H4
Duck 1H4
Turtle 1H4
Xenopus 1H4
Zebrafish 1H4
Tilapia 1H4
Medaka 1H4
Stickleback 1H4
100 100
98
98
96
100
100
Chicken_1F1
Duck_1F1
Turtle_1F1
Human_1F1
Dolphin_1F1
Mouse_1F1
Rat_1F1
Zebrafish_1F1a
Medaka_1F1
Tilapia_1F1a
Xenopus_1F1
Tilapia_1F1b
Zebrafish_1F1b
93
93 99
97
Human_1F3
Dolphin_1F3
Mouse_1F3
Rat_1F3
Chicken_1F3a
Duck_1F3a
Turtle_1F3b
Zebrafish_1F3b
Stickleback_1F3b
Medaka_1F3b
Tilapia_1F3c
Zebrafish_1F3c
Stickleback_1F3a
Medaka_1F3c
Tilapia_1F3a
Zebrafish_1F3a
Medaka_1F3a
Tilapia_1F3b
99
99
100
100
100
100
99
99 98
100 100
Human_1H2
Dolphin_1H2
Mouse_1H2
Rat_1H2100
Human 1I1
Dolphin 1I1
Mouse 1I1
Rat 1I1
Chicken 1I1
Duck 1I1
Turtle 1I1
Xenopus 1I1
Zebrafish 1I1b
Tilapia 1I1a
Medaka 1I1b
Stickleback 1I1b
Zebrafish 1I1a
Medaka 1I1a
Tilapia 1I1b
Stickleback 1I1a
100
100
100
100 100
91
97
Human 1I2
Dolphin 1I2
Mouse 1I2
Rat 1I2
Xenopus 1I2
Zebrafish 1I2
Medaka 1I2
Tilapia 1I2
100
100
100 100
99
100
100
Human_1I3
Dolphin_1I3
Mouse_1I3
Rat_1I3
Turtle_1I3
Xenopus_1I3

28 Human 2A1
Mouse 2A1
Rat 2A1
Dolphin 2A1
Chicken 2A1
Duck 2A1
Turtle 2A1
Xenopus 2A1
Zebrafish 2A1
Tilapia 2A1
Stickleback 2A1
100 100
100 99
99 100
Human_2A2
Mouse_2A2
Rat_2A2
Dolphin_2A2
Chicken_2A2
Duck_2A2
Turtle_2A2
Xenopus_2A2
Medaka_2A2
Tilapia_2A2
Stickleback_2A2
100 98
100
100 100
99
90
100
Human 2B1
Mouse 2B1
Rat 2B1
Dolphin 2B1
Chicken 2B1
Duck 2B1
Turtle 2B1
Xenopus 2B1
Zebrafish 2B1b
Zebrafish 2B1a
Medaka 2B1a
Tilapia 2B1
Stickleback 2B1
100
100
97
95
Human 2B2
Dolphin 2B2
Mouse 2B2
Rat 2B2
Xenopus 2B2
Zebrafish 2B2a
Tilapia 2B2b
Medaka 2B2a
Stickleback 2B2b
Zebrafish 2B2b
Medaka 2B2b
Tilapia 2B2a
Stickleback 2B2a
100 99
100
100
100
Human 2B3
Dolphin 2B3
Mouse 2B3
Rat 2B3
Chicken 2B3
Duck 2B3
Turtle 2B3
Xenopus 2B3
Zebrafish 2B3a
Zebrafish 2B3b
Medaka 2B1b
Tilapia 2B3
Stickleback 2B3
100
100
100
100 96
96
99
Human 2C1
Dolphin 2C1
Mouse 2C1
Rat 2C1
Chicken 2C1
Duck 2C1
Turtle 2C1
Xenopus 2C1
Zebrafish 2C1
Stickleback 2C1
Medaka 2C1
Tilapia 2C1
100
100 100
97
100
100
98
Human 2C2
Dolphin 2C2
Mouse 2C2
Rat 2C2
Chicken 2C2
Duck 2C2
Turtle 2C2
Xenopus 2C2
Zebrafish 2C2
Stickleback 2C2
Medaka 2C2
Tilapia 2C2
100
100
100 100
100
98 99 94
Human 2E3
Mouse 2E3
Rat 2E3
Dolphin 2E3
Chicken 2E3
Turtle 2E3
Xenopus 2E3
Zebrafish 2E3
Medaka 2E3a
Tilapia 2E3a
Stickleback 2E3
Medaka 2E3b
Tilapia 2E3b
100
100
100 90
91
96 Human_2E1
Dolphin_2E1
Mouse_2E1
Rat_2E1
Chicken_2E1
Duck_2E1
Turtle_2E1
Xenopus_2E1
Zebrafish_2E1
Stickleback_2E1
Medaka_2E1
Tilapia_2E193 99
100
Human_2F6
Dolphin_2F6
Mouse_2F6
Rat_2F6
Turtle_2F6
Xenopus_2F6
Zebrafish_2F6a
Zebrafish_2F6b
Stickleback_2F6b
Medaka_2F6b
Tilapia_2F6b
Tilapia_2F6a
Medaka_2F6a
Stickleback_2F6a
100
100
90 90
94
94
98
Human_2F1
Mouse_2F1
Rat_2F1
Dolphin_2F1
Xenopus_2F1
Chicken_2F1
Zebrafish_2F1a
Tilapia_2F1
Stickleback_2F1
100
99 90
100
Human_2F2
Mouse_2F2
Rat_2F2
Chicken_2F2
Turtle_2F2
Xenopus_2F2
Duck_2F2
Tilapia_2F2b
Zebrafish_2F2
Medaka_2F2
Stickleback_2F2b
Tilapia_2F2a
Stickleback_2F2a
100
100
100
99

29 Human 3A1
Dolphin 3A1
Mouse 3A1
Rat 3A1
Chicken 3A1
Duck 3A1
Turtle 3A1
Xenopus 3A1
Zebrafish 3A1
Medaka 3A1
Tilapia 3A1
Stickleback 3A1
100 100
100 100
100 99
99
92
Human 3A2
Dolphin 3A2
Mouse 3A2
Rat 3A2
Chicken 3A2
Duck 3A2
Turtle 3A2
Xenopus 3A2
Zebrafish 3A2b
Medaka 3A2b
Tilapia 3A2b
Zebrafish 3A2a
Stickleback 3A2a
Medaka 3A2a
Tilapia 3A2a
100 100
100
100
100 100 90
99
97
Human 3B1
Dolphin 3B1
Mouse 3B1
Rat 3B1
Turtle 3B1
Xenopus 3B1
Zebrafish 3B1
Medaka 3B1
Tilapia 3B1
Stickleback 3B1
100
100
99
99
92
100
100
100
100
99
100
99
Human_3B2
Mouse_3B2
Rat_3B2
Dolphin_3B2
Chicken_3B2
Duck_3B2
Turtle_3B2
Zebrafish_3B2
Tilapia_3B2a
Medaka_3B2a
Stickleback_3B2a
Stickleback_3B2b
Medaka_3B2b
Tilapia_3B2b
100
Human_3B3
Dolphin_3B3
Mouse_3B3
Rat_3B3
Turtle_3B3
Chicken_3B3
Duck_3B3
Xenopus_3B3a
Zebrafish_3B3a
Tilapia_3B3b
Medaka_3B3a
Stickleback_3B3b
Medaka_3B3b
Tilapia_3B3a
Zebrafish_3B3b
Xenopus_3B3b
Stickleback_3B3a
100
100
90
100
100
100
100 100
99
Human_3C1
Dolphin_3C1
Rat_3C1
Mouse_3C1
Chicken_3C1
Duck_3C1
Turtle_3C1
Xenopus_3C1
Tilapia_3C1b
Medaka_3C1b
Stickleback_3C1b
Zebrafish_3C1
Tilapia_3C1a
Medaka_3C1a
Stickleback_3C1a
100
100
92
98 99
95
Human_3C2
Dolphin_3C2
Mouse_3C2
Rat_3C2
Chicken_3C2
Duck_3C2
Turtle_3C2
Xenopus_3C2
Zebrafish_3C2
Medaka_3C2
Tilapia_3C2
Stickleback_3C2
100
100 100
100
100 100
99
99
98
Human_3C3
Dolphin_3C3
Mouse_3C3
Rat_3C3
Chicken_3C3
Turtle_3C3
Xenopus_3C3
Stickleback_3C3
Zebrafish_3C3
100
100
100
100
99
Human_3C4
Mouse_3C4
Rat_3C4
Dolphin_3C4
Turtle_3C4
Chicken_3C4
Duck_3C4
Xenopus_3C4
Zebrafish_3C4
Stickleback_3C4b
Medaka_3C4b
Tilapia_3C4b
Stickleback_3C4a
Medaka_3C4a
Tilapia_3C4a
100
100
100 100
100 100
100 98
100
95

30
Human 5A2
Dolphin 5A2
Mouse 5A2
Rat 5A2
Chicken 5A2
Duck 5A2
Turtle 5A2
Xenopus 5A2
Zebrafish 5A2
Stickleback 5A2
Medaka 5A2
Tilapia 5A2
100 99
99
99
97 98
100
Human_5A1
Dolphin_5A1
Mouse_5A1
Rat_5A1
Chicken_5A1
Xenopus_5A1
Zebrafish_5A1b
Zebrafish_5A1a
Stickleback_5A1a
Medaka_5A1a
Tilapia_5A1
Medaka_5A1b
Stickleback_5A1b
100 100
100
100
100
98
99
Human_6A1
Mouse_6A1
Rat_6A1
Dolphin_6A1
Chicken_6A1
Turtle_6A1
Xenopus_6A1
Zebrafish_6A1a
Tilapia_6A1
Stickleback_6A1
Zebrafish_6A1b
100 90
96 100
100 97
90
Human 4A1
Dolphin 4A1
Mouse 4A1
Rat 4A1
Turtle 4A1
Xenopus 4A1
Duck 4A1
Zebrafish 4A1
Stickleback 4A1a
Medaka 4A1b
Tilapia 4A1a
Medaka 4A1a
Tilapia 4A1b
Stickleback 4A1b
100
100
100
100
100
94
91
Human 4A3
Dolphin 4A3
Mouse 4A3
Rat 4A3
Chicken 4A3
Duck 4A3
Turtle 4A3
Zebrafish 4A3
Medaka 4A3
Tilapia 4A3
Stickleback 4A3
100 100
99
95
100
100
100
Human_4A2
Dolphin_4A2
Rat_4A2
Mouse_4A2
Turtle_4A2
Chicken_4A2
Xenopus_4A2a
Zebrafish_4A2a
Stickleback_4A2
Medaka_4A2a
Tilapia_4A2a
Zebrafish_4A2b
Medaka_4A2b
Tilapia_4A2b
Xenopus_4A2b
91
96
99 95
100
100
99 99
100

Figure S2. Schematic diagram depicts the evolution of PXR and CAR in vertebrates. 31
32

Table S1. Details for nuclear receptor sequence searches in 12 model vertebrates. 33
34
BLASTn
Hits
BLASTp
Hits Sum
After
sortation
Verified by
software
NR0B
Subfamily
Final sets
of NRs.
Human 33849 24967 58816 57 46 2 48
Mouse 23014 12540 35554 62 47 2 49
Rat 8312 8896 17208 70 47 2 49
Dolphin 2834 2752 5586 74 45 2 47
Chicken 3712 3761 7473 50 42 2 44
Duck 2381 4034 6415 48 40 2 42
Turtle 2922 3421 6343 48 46 2 48
Xenopus 2289 3850 6139 53 50 2 52
Zebrafish 9788 9230 19018 72 70 3 73
Medaka 3601 4090 7691 78 65 2 67
Tilapia 7586 6630 14216 83 71 3 74
Stickleback 571 268 839 64 64 2 66
35

Table S2. Sequence ID. for each nuclear receptor gene in Ensembl database. 36
37
Human Mouse Rat Dolphin Chicken Duck Turtle Xenopus Zebrafish Medaka Tilapia Stickleback
NR1A1 ENSG0000012
6351
ENSMUSG00
000058756
ENSRNOG00
000009066
ENSTTRG000
00016893
ENSGALG00
000000270
ENSAPLG000
00016001
ENSPSIG0000
0012754
ENSXETG000
00024399
ENSDARG00
000000151
ENSORLG000
00016941
ENSONIG000
00018247
ENSGACG000
00003766
ENSDARG00
000052654
ENSORLG000
00012005
ENSONIG000
00006456
ENSGACG000
00006540
NR1A2 ENSG0000015
1090
ENSMUSG00
000021779
ENSRNOG00
000006649
ENSTTRG000
00001859
ENSGALG00
000011294
ENSAPLG000
00006081
ENSPSIG0000
0008182
ENSXETG000
00003871
ENSDARG00
000021163
ENSORLG000
00008122
ENSONIG000
00010312
ENSGACG000
00007996
NR1B1 ENSG0000013
1759
ENSMUSG00
000037992
ENSRNOG00
000009972
ENSTTRG000
00016901
ENSGALG00
000005629
ENSAPLG000
00006377
ENSPSIG0000
0002372
ENSXETG000
00024390
ENSDARG00
000056783
ENSORLG000
00004373
ENSONIG000
00019915
ENSGACG000
00012955
ENSDARG00
000034893
ENSONIG000
00006314
ENSGACG000
00005297
NR1B2 ENSG0000007
7092
ENSMUSG00
000017491
ENSRNOG00
000024061
ENSTTRG000
00010874
ENSGALG00
000011298
ENSAPLG000
00006432
ENSPSIG0000
0007930
ENSXETG000
00007272
ENSORLG000
00008502
ENSONIG000
00010320
ENSGACG000
00007999
ENSORLG000
00016394
ENSONIG000
00006493
NR1B3 ENSG0000017
2819
ENSMUSG00
000001288
ENSRNOG00
000012499
ENSTTRG000
00002778
ENSXETG000
00012670
ENSDARG00
000034117
ENSORLG000
00015382
ENSONIG000
00012223
ENSGACG000
00009372
ENSDARG00
000054003
ENSORLG000
00007861
ENSONIG000
00019165
ENSGACG000
00000612
NR1C1 ENSG0000018
6951
ENSMUSG00
000022383
ENSRNOG00
000021463
ENSTTRG000
00004136
ENSGALG00
000022985
ENSAPLG000
00010641
ENSPSIG0000
0018221
ENSXETG000
00023454
ENSDARG00
000031777
ENSORLG000
00002413
ENSONIG000
00016715
ENSGACG000
00018958
ENSDARG00
000054323
ENSORLG000
00011091
ENSONIG000
00008831
ENSGACG000
00003703
NR1C2 ENSG0000011
2033
ENSMUSG00
000002250
ENSRNOG00
000000503
ENSTTRG000
00009416
ENSGALG00
000002588
ENSAPLG000
00004751
ENSPSIG0000
0005889
ENSXETG000
00015121
ENSDARG00
000044525
ENSORLG000
00006636
ENSONIG000
00011871
ENSGACG000
00008288
ENSDARG00
000009473

NR1C3 ENSG0000013
2170
ENSMUSG00
000000440
ENSRNOG00
000008839
ENSTTRG000
00016565
ENSGALG00
000004974
ENSAPLG000
00009031
ENSPSIG0000
0011100
ENSXETG000
00017422
ENSDARG00
000031848
ENSORLG000
00004432
ENSONIG000
00014331
ENSGACG000
00001665
NR1D1 ENSG0000012
6368
ENSMUSG00
000020889
ENSRNOG00
000009329
ENSTTRG000
00016894
ENSPSIG0000
0014806
ENSXETG000
00024397
ENSDARG00
000033160
ENSONIG000
00009283
ENSGACG000
00009356
NR1D2 ENSG0000017
4738
ENSMUSG00
000021775
ENSRNOG00
000046912
ENSTTRG000
00010829
ENSGALG00
000011291
ENSAPLG000
00005753
ENSPSIG0000
0008488
ENSXETG000
00003869
ENSDARG00
000003820
ENSORLG000
00016431
ENSONIG000
00008699
ENSGACG000
00012958
ENSDARG00
000009594
ENSONIG000
00010308
ENSGACG000
00007986
NR1D4 ENSDARG00
000031161
ENSORLG000
00007837
ENSONIG000
00012213
ENSGACG000
00000614
ENSDARG00
000059370
ENSORLG000
00015399
ENSONIG000
00019164
NR1F1 ENSG0000006
9667
ENSMUSG00
000032238
ENSRNOG00
000027145
ENSTTRG000
00007718
ENSGALG00
000003759
ENSAPLG000
00005866
ENSPSIG0000
0011314
ENSXETG000
00021123
ENSDARG00
000031768
ENSORLG000
00007645
ENSONIG000
00015289
ENSDARG00
000001910
ENSONIG000
00015603
NR1F2 ENSG0000019
8963
ENSMUSG00
000036192
ENSRNOG00
000013413
ENSTTRG000
00008387
ENSGALG00
000015150
ENSAPLG000
00007187
ENSPSIG0000
0005579
ENSXETG000
00031251
ENSDARG00
000033498
ENSORLG000
00012441
ENSONIG000
00010762
ENSGACG000
00011556
ENSXETG000
00008148
NR1F3 ENSG0000014
3365
ENSMUSG00
000028150
ENSRNOG00
000046831
ENSTTRG000
00003151
ENSGALG00
000025988
ENSAPLG000
00013051
ENSPSIG0000
0008995
ENSXETG000
00002131
ENSDARG00
000087195
ENSORLG000
00009486
ENSONIG000
00004686
ENSGACG000
00012280
ENSGALG00
000001035
ENSAPLG000
00011493
ENSPSIG0000
0016262
ENSDARG00
000057231
ENSORLG000
00003765
ENSONIG000
00010247
ENSGACG000
00015341
ENSDARG00
000017780
ENSORLG000
00014886
ENSONIG000
00006222
NR1H3 ENSG0000002
5434
ENSMUSG00
000002108
ENSRNOG00
000013172
ENSTTRG000
00014149
ENSGALG00
000008202
ENSAPLG000
00010925
ENSPSIG0000
0010360
ENSXETG000
00000307
ENSDARG00
000043170
ENSORLG000
00001286
ENSONIG000
00005828
ENSGACG000
00017167

NR1H2 ENSG0000013
1408
ENSMUSG00
000060601
ENSRNOG00
000019812
ENSTTRG000
00002416
NR1H5 ENSMUSG00
000048938
ENSRNOG00
000023073
ENSGALG00
000002170
ENSAPLG000
00008338
ENSPSIG0000
0003828
ENSXETG000
00021443
ENSDARG00
000031046
ENSONIG000
00009252
ENSGACG000
00004938
NR1H4 ENSG0000001
2504
ENSMUSG00
000047638
ENSRNOG00
000007197
ENSTTRG000
00016373
ENSGALG00
000011594
ENSAPLG000
00013289
ENSPSIG0000
0005774
ENSXETG000
00030372
ENSDARG00
000057741
ENSORLG000
00011270
ENSONIG000
00014678
ENSGACG000
00011745
NR1I1 ENSG0000011
1424
ENSMUSG00
000022479
ENSRNOG00
000008574
ENSTTRG000
00012578
ENSGALG00
000026166
ENSAPLG000
00005087
ENSPSIG0000
0018108
ENSXETG000
00010658
ENSDARG00
000043059
ENSORLG000
00001063
ENSONIG000
00009200
ENSGACG000
00004763
ENSDARG00
000070721
ENSORLG000
00016402
ENSONIG000
00019378
ENSGACG000
00007975
NR1I2 ENSG0000014
4852
ENSMUSG00
000022809
ENSRNOG00
000002906
ENSTTRG000
00016650
ENSXETG000
00018029
ENSDARG00
000029766
ENSORLG000
00017953
ENSONIG000
00014385
NR1I3 ENSG0000014
3257
ENSMUSG00
000005677
ENSRNOG00
000003260
ENSTTRG000
00009227
ENSGALG00
000028624
ENSPSIG0000
0004437
ENSXETG000
00031759
NR2A1 ENSG0000010
1076
ENSMUSG00
000017950
ENSRNOG00
000008895
ENSTTRG000
00013004
ENSGALG00
000004285
ENSAPLG000
00008950
ENSPSIG0000
0012689
ENSXETG000
00001775
ENSDARG00
000021494
ENSORLG000
00016380
ENSONIG000
00016515
ENSGACG000
00011485
NR2A3 ENSGALG00
000015670
ENSAPLG000
00011331
ENSPSIG0000
0017650
ENSXETG000
00016389
ENSDARG00
000012764
ENSONIG000
00005911
NR2A2 ENSG0000016
4749
ENSMUSG00
000017688
ENSRNOG00
000008971
ENSTTRG000
00003691
ENSGALG00
000005708
ENSAPLG000
00011794
ENSPSIG0000
0003756
ENSXETG000
00017845
ENSDARG00
000071565
ENSORLG000
00006996
ENSONIG000
00014490
ENSGACG000
00002422
NR2B1 ENSG0000018
6350
ENSMUSG00
000015846
ENSRNOG00
000009446
ENSTTRG000
00009492
ENSGALG00
000002626
ENSAPLG000
00013150
ENSPSIG0000
0011977
ENSXETG000
00012733
ENSDARG00
000057737
ENSORLG000
00012155
ENSONIG000
00013076
ENSGACG000
00018189
ENSDARG00
000035127
ENSORLG000
00016690
NR2B2 ENSG0000020
4231
ENSMUSG00
000039656
ENSRNOG00
000000464
ENSTTRG000
00004291
ENSXETG000
00020416
ENSDARG00
000078954
ENSORLG000
00006476
ENSONIG000
00020007
ENSGACG000
00000096
ENSDARG00
000002006
ENSORLG000
00007020
ENSONIG000
00002873
ENSGACG000
00007982

NR2B3 ENSG0000014
3171
ENSMUSG00
000015843
ENSRNOG00
000004537
ENSTTRG000
00003653
ENSGALG00
000003406
ENSAPLG000
00004831
ENSPSIG0000
0004871
ENSXETG000
00004750
ENSDARG00
000005593
ENSONIG000
00002143
ENSGACG000
00011685
ENSDARG00
000004697
NR2C1 ENSG0000012
0798
ENSMUSG00
000005897
ENSRNOG00
000006983
ENSTTRG000
00016305
ENSGALG00
000011327
ENSAPLG000
00006253
ENSPSIG0000
0017190
ENSXETG000
00023840
ENSDARG00
000045527
ENSORLG000
00004114
ENSONIG000
00008566
ENSGACG000
00010174
NR2C2 ENSG0000017
7463
ENSMUSG00
000005893
ENSRNOG00
000010536
ENSTTRG000
00009876
ENSGALG00
000008519
ENSAPLG000
00007538
ENSPSIG0000
0008928
ENSXETG000
00004817
ENSDARG00
000042477
ENSORLG000
00010877
ENSONIG000
00017240
ENSGACG000
00002941
NR2E1 ENSG0000011
2333
ENSMUSG00
000019803
ENSRNOG00
000050550
ENSTTRG000
00008863
ENSGALG00
000015305
ENSAPLG000
00010675
ENSPSIG0000
0006035
ENSXETG000
00014853
ENSDARG00
000017107
ENSORLG000
00013426
ENSONIG000
00013281
ENSGACG000
00008934
NR2E3 ENSG0000003
1544
ENSMUSG00
000032292
ENSRNOG00
000050690
ENSTTRG000
00009410
ENSGALG00
000002093
ENSPSIG0000
0017480
ENSXETG000
00005219
ENSDARG00
000045904
ENSORLG000
00000011
ENSONIG000
00007109
ENSGACG000
00004739
ENSORLG000
00007175
ENSONIG000
00015396
NR2F1 ENSG0000017
5745
ENSMUSG00
000069171
ENSRNOG00
000014795
ENSTTRG000
00001519
ENSGALG00
000027907
ENSPSIG0000
0009818
ENSXETG000
00011594
ENSDARG00
000052695
ENSORLG000
00010191
ENSONIG000
00011840
ENSGACG000
00010385
ENSPSIG0000
0010198
ENSDARG00
000017168
NR2F2 ENSG0000018
5551
ENSMUSG00
000030551
ENSRNOG00
000010308
ENSGALG00
000007000
ENSAPLG000
00010629
ENSPSIG0000
0017164
ENSXETG000
00022346
ENSDARG00
000040926
ENSORLG000
00008429
ENSONIG000
00015133
ENSGACG000
00013235
ENSONIG000
00003070
ENSGACG000
00014846
NR2F5 ENSXETG000
00011046
ENSDARG00
000033172
ENSORLG000
00016315
ENSONIG000
00008594
ENSGACG000
00013191
NR2F6 ENSG0000016
0113
ENSMUSG00
000002393
ENSRNOG00
000016892
ENSTTRG000
00003132
ENSGALG00
000027294
ENSAPLG000
00003193
ENSPSIG0000
0013773
ENSXETG000
00013531
ENSDARG00
000003607
ENSORLG000
00008749
ENSONIG000
00010512
ENSGACG000
00007766
ENSDARG00
000003165
ENSORLG000
00008911
ENSONIG000
00010104
ENSGACG000
00015583

NR3A1 ENSG0000009
1831
ENSMUSG00
000019768
ENSRNOG00
000019358
ENSTTRG000
00002996
ENSGALG00
000012973
ENSAPLG000
00004585
ENSPSIG0000
0004166
ENSXETG000
00012364
ENSDARG00
000004111
ENSORLG000
00014514
ENSONIG000
00013354
ENSGACG000
00008711
NR3A2 ENSG0000014
0009
ENSMUSG00
000021055
ENSRNOG00
000005343
ENSTTRG000
00000517
ENSGALG00
000011801
ENSAPLG000
00011895
ENSPSIG0000
0018210
ENSXETG000
00007257
ENSDARG00
000016454
ENSORLG000
00017721
ENSONIG000
00005633
ENSGACG000
00007514
ENSDARG00
000034181
ENSORLG000
00018012
ENSONIG000
00001710
ENSGACG000
00000213
NR3B1 ENSG0000017
3153
ENSMUSG00
000024955
ENSRNOG00
000021139
ENSTTRG000
00010296
ENSPSIG0000
0016751
ENSXETG000
00007211
ENSDARG00
000069266
ENSORLG000
00010624
ENSONIG000
00001778
ENSGACG000
00020287
NR3B2 ENSG0000011
9715
ENSMUSG00
000021255
ENSRNOG00
000010259
ENSTTRG000
00001302
ENSGALG00
000010365
ENSAPLG000
00012470
ENSPSIG0000
0017916
ENSXETG000
00013217
ENSDARG00
000040151
ENSORLG000
00016581
ENSONIG000
00015282
ENSGACG000
00010561
ENSORLG000
00009126
ENSONIG000
00020192
ENSGACG000
00007542
NR3B3 ENSG0000019
6482
ENSMUSG00
000026610
ENSRNOG00
000002593
ENSTTRG000
00006004
ENSGALG00
000009645
ENSAPLG000
00005309
ENSPSIG0000
0005595
ENSXETG000
00020932
ENSDARG00
000004861
ENSORLG000
00011528
ENSONIG000
00000573
ENSGACG000
00013426
ENSXETG000
00016948
ENSDARG00
000011696
ENSORLG000
00016819
ENSONIG000
00017162
ENSGACG000
00016275
NR3B4 ENSDARG00
000015064
ENSONIG000
00001134
ENSGACG000
00004898
NR3C1 ENSG0000011
3580
ENSMUSG00
000024431
ENSRNOG00
000014096
ENSTTRG000
00003260
ENSGALG00
000007394
ENSAPLG000
00007318
ENSPSIG0000
0015245
ENSXETG000
00001879
ENSDARG00
000025032
ENSORLG000
00006022
ENSONIG000
00017907
ENSGACG000
00018209
ENSORLG000
00001565
ENSONIG000
00008483
ENSGACG000
00020725
NR3C2 ENSG0000015
1623
ENSMUSG00
000031618
ENSRNOG00
000034007
ENSTTRG000
00014440
ENSGALG00
000010035
ENSAPLG000
00015146
ENSPSIG0000
0006383
ENSXETG000
00026061
ENSDARG00
000037025
ENSORLG000
00007530
ENSONIG000
00010029
ENSGACG000
00017193
NR3C3 ENSG0000008
2175
ENSMUSG00
000031870
ENSRNOG00
000006831
ENSTTRG000
00000030
ENSGALG00
000017195
ENSAPLG000
00003887
ENSPSIG0000
0013654
ENSXETG000
00005482
ENSDARG00
000035966
ENSORLG000
00002651
ENSGACG000
00012162
NR3C4 ENSG0000016
9083
ENSMUSG00
000046532
ENSRNOG00
000005639
ENSTTRG000
00004230
ENSGALG00
000004596
ENSAPLG000
00006566
ENSPSIG0000
0010176
ENSXETG000
00005089
ENSDARG00
000067976
ENSORLG000
00008220
ENSONIG000
00012854
ENSGACG000
00018525
ENSORLG000 ENSONIG000 ENSGACG000

00009520 00017538 00020332
NR4A1 ENSG0000012
3358
ENSMUSG00
000023034
ENSRNOG00
000007607
ENSTTRG000
00002817
ENSAPLG000
00014123
ENSPSIG0000
0018018
ENSXETG000
00000579
ENSDARG00
000000796
ENSORLG000
00015557
ENSONIG000
00016717
ENSGACG000
00010788
ENSORLG000
00015279
ENSONIG000
00019260
ENSGACG000
00000045
NR4A2 ENSG0000015
3234
ENSMUSG00
000026826
ENSRNOG00
000005600
ENSTTRG000
00005740
ENSGALG00
000012538
ENSAPLG000
00012071
ENSPSIG0000
0008054
ENSXETG000
00031753
ENSDARG00
000017007
ENSORLG000
00016692
ENSONIG000
00008976
ENSGACG000
00005831
ENSXETG000
00024016
ENSDARG00
000044532
ENSORLG000
00000050
ENSONIG000
00012131
NR4A3 ENSG0000011
9508
ENSMUSG00
000028341
ENSRNOG00
000005964
ENSTTRG000
00007458
ENSGALG00
000013568
ENSAPLG000
00011263
ENSPSIG0000
0012281
ENSDARG00
000055854
ENSORLG000
00008732
ENSONIG000
00006026
ENSGACG000
00009027
NR5A1 ENSG0000013
6931
ENSMUSG00
000026751
ENSRNOG00
000012682
ENSTTRG000
00017390
ENSGALG00
000001080
ENSAPLG000
00004548
ENSPSIG0000
0006131
ENSXETG000
00011456
ENSDARG00
000017704
ENSORLG000
00016486
ENSONIG000
00020218
ENSGACG000
00003539
ENSDARG00
000023362
ENSORLG000
00013196
ENSGACG000
00018317
NR5A2 ENSG0000011
6833
ENSMUSG00
000026398
ENSRNOG00
000000653
ENSTTRG000
00003256
ENSGALG00
000002182
ENSAPLG000
00009302
ENSPSIG0000
0003632
ENSXETG000
00000314
ENSDARG00
000042556
ENSORLG000
00006933
ENSONIG000
00012517
ENSGACG000
00008896
NR5A5 ENSDARG00
000039116
ENSORLG000
00006019
ENSONIG000
00001686
ENSGACG000
00009952
NR6A1 ENSG0000014
8200
ENSMUSG00
000063972
ENSRNOG00
000013232
ENSTTRG000
00017391
ENSGALG00
000001073
ENSAPLG000
00004788
ENSPSIG0000
0006445
ENSXETG000
00008578
ENSDARG00
000018030
ENSORLG000
00016492
ENSONIG000
00020217
ENSGACG000
00003560
ENSDARG00
000014480
NR0B1 ENSG0000016
9297
ENSMUSG00
000025056
ENSRNOG00
000003765
ENSTTRG000
00013272
ENSGALG00
000016287
ENSAPLG000
00003894
ENSPSIG0000
0009740
ENSXETG000
00015374
ENSDARG00
000056541
ENSORLG000
00011824
ENSONIG000
00012111
ENSGACG000
00002817
ENSONIG000
00006662
NR0B2 ENSG0000013
1910
ENSMUSG00
000037583
ENSRNOG00
000007229
ENSTTRG000
00016680
ENSGALG00
000000887
ENSAPLG000
00010744
ENSPSIG0000
0017134
ENSXETG000
00011771
ENSDARG00
000044685
ENSORLG000
00004442
ENSONIG000
00006772
ENSGACG000
00007198

Table S3. Genes with incomplete/without DBD/LBD regions in the Ensembl database. Genes 38
marked in red means the full sequences were retrieved in NCBI/ EMBL/DDBJ databases. 39
40
Gene and related Ensembl ID.
Human —
Mouse —
Rat NR2E3 (ENSRNOG00000050690); NR3A1 (ENSRNOG00000019358); NR3C3 (ENSRNOG00000006831);
NR5A2 (ENSRNOG00000000653)
Dolphin
NR1A1 (ENSTTRG00000016893); NR1B2 (ENSTTRG00000010874); NR1C1 (ENSTTRG00000004136);
NR1F3 (ENSTTRG00000003151); NR1I2 (ENSTTRG00000016650); NR2A1 (ENSTTRG00000013004);
NR2B1 (ENSTTRG00000009492); NR2B3 (ENSTTRG00000003653); NR2F6 (ENSTTRG00000003132);
NR3A1 (ENSTTRG00000002996); NR3B2 (ENSTTRG00000001302); NR4A1 (ENSTTRG00000002817);
NR4A3 (ENSTTRG00000007458)
Chicken NR1B1 (ENSGALG00000005629); NR2F6 (ENSGALG00000027294);
Duck
NR1A1 (ENSAPLG00000016001); NR1B1(ENSAPLG00000006377); NR1F2 (ENSAPLG00000007187);
NR1F3b (ENSAPLG00000011493); NR1H4 (ENSAPLG00000013289); NR1I1 (ENSAPLG00000005087);
NR2F6 (ENSAPLG00000003193); NR3A1 (ENSAPLG00000004585); NR3C3 (ENSAPLG00000003887);
NR4A1 (ENSAPLG00000014123); NR4A2 (ENSAPLG00000012071); NR5A1 (ENSAPLG00000004548);
NR6A1 (ENSAPLG00000004788); NR0B1 (ENSAPLG00000003894)
Turtle
NR1A1 (ENSPSIG00000012754); NR1B1 (ENSPSIG00000002372); NR1D1 (ENSPSIG00000014806);
NR2A2 (ENSPSIG00000003756); NR2E3 (ENSPSIG00000017480); NR2F1b (ENSPSIG00000010198);
NR3B1 (ENSPSIG00000016751); NR4A2 (ENSPSIG00000008054); NR5A1 (ENSPSIG00000006131);
NR6A1 (ENSPSIG00000006445)
Xenopus NR1C2 (ENSXETG00000015121); NR1C3 (ENSXETG00000017422); NR1F3 (ENSXETG00000002131);
NR2A1 (ENSXETG00000001775); NR4A2b (ENSXETG00000024016)
Zebrafish NR1C3 (ENSDARG00000031848); NR1I1 (ENSDARG00000043059)
Medaka NR1B1 (ENSORLG00000004373); NR2A1 (ENSORLG00000016380); NR2F1 (ENSORLG00000010191);
NR3C3 (ENSORLG00000002651); NR6A1 (ENSORLG00000016492)
Tilapia —
Stickleback —
Table S4. The vertebrate species used for NR1I1 (VDR) gene phylogenetic analysis. 41
42
Common name Scientific name Common name Scientific name
Human Homo sapiens Flycatcher Ficedula albicollis
Gibbon Nomascus leucogenys Zebra Finch Taeniopygia guttata
Gorilla Gorilla gorilla gorilla Duck Anas platyrhynchos
Macaque Macaca mulatta Chicken Gallus gallus
Marmoset Callithrix jacchus Turkey Meleagris gallopavo
Bushbaby Otolemur garnettii
Cat Felis catus Anole lizard Anolis carolinensis
Dog Canis lupus familiaris Chinese softshell turtle Pelodiscus sinensis
Ferret Mustela putorius furo
Hedgehog Erinaceus europaeus Xenopus Xenopus tropicalis
Rabbit Oryctolagus cuniculus
Dolphin Tursiops truncatus Coelacanth Latimeria chalumnae
Pig Sus scrofa Tilapia Oreochromis niloticus
Opossum Monodelphis domestica Zebrafish Danio rerio
Cow Bos taurus Tetraodon Tetraodon nigroviridis
Sheep Ovis aries Medaka Oryzias latipes
Mouse Mus musculus Platyfish Xiphophorus maculatus
Rat Rattus norvegicus Stickleback Gasterosteus aculeatus
Guinea Pig Cavia porcellus
Squirrel Ictidomys tridecemlineatus
43