faculty of bioscience engineering interuniversity
TRANSCRIPT

1
Katholieke
Universiteit
Leuven
FACULTY OF BIOSCIENCE ENGINEERING
INTERUNIVERSITY PROGRAMME
MASTER OF FOOD TECHNOLOGY (IUPFOOD)
TOWARDS A BETTER UNDERSTANDING OF THE PECTIN
STRUCTURE-FUNCTION RELATIONSHIP IN CARROT AND TOMATO
PUREE
Promoter: Prof.Dr.ir.M. Hendrickx
Department of Food and Microbial
Technology.
Laboratory of Food Technology.
May 2011
Master dissertation submitted in
partial fulfilment of the requirements
for the Degree of Master of Science
in Food Technology.
By: Davis Chaula

i
ACKNOWLEGMENTS
I am glad to express my heartfelt appreciation to everyone whose advice, support and
encouragement has contributed to accomplishment of this work.
Special thanks to my supervisor Stefanie Christiaens for her tireless support in day to day activities in
the Laboratory. It is through her patience that I managed to do all of my experiments within the
allowed time frame. I admire her systematic way of doing things and hard working. She was never
angry when things went wrong, instead she encouraged me.
I sincerely appreciate the help from Sandy Van Buggenhout during report writing. Indeed this
document would have been incomplete without her support. I am proud of you. Your assistance has
nourished my scientific report writing skills and critical interpretation of scientific data and literature.
I thank all technical staffs of the Laboratory of Food Technology for their help during the laboratory
work. They were always available and ready to guide me on how to use different equipments and
rectify any fault.
Last but not least I acknowledge the invisible hand of my promoter Prof. Marc Hendrickx. It is
through his high level of organization of all resources in the Laboratory of Food Technology that for
the first time in my life I have the real feel of science. He is the core of my experience in
team-working, organization and planning.

ii
TABLE OF CONTENTS
ACKNOWLEGMENTS ................................................................................................................................. i
TABLE OF CONTENTS ................................................................................................................................ii
LIST OF SYMBOLS AND ABBREVIATIONS .................................................................................................. v
ABSTRACT ................................................................................................................................................ vi
LITERATURE REVIEW ............................................................................................................................... 1
1. PECTIN ................................................................................................................................................. 1
1.1 Chemical Structure ........................................................................................................................ 1
1.1.1 Homogalacturonan ................................................................................................................. 1
1.1.2 Rhamnogalacturonan-I ........................................................................................................... 2
1.1.3 Rhamnogalacturonan-II .......................................................................................................... 3
1.1.4 Macromolecular organization of pectin ................................................................................. 3
1.2 PECTIN STRUCTURAL MODIFICATIONS ............................................................................................. 4
1.2.1 Enzymatic pectin conversion ...................................................................................................... 5
1.2.1.1 Pectinmethylesterase .......................................................................................................... 5
1.2.1.2 Polygalacturonase ............................................................................................................... 8
1.2.2 Non enzymatic pectin conversion .............................................................................................. 9
1.3 PECTIN FUNCTIONAL PROPERTIES .................................................................................................. 11
1.3.1 Pectin and fruit ripening ........................................................................................................... 11
1.3.2 Pectin as a gelling argent ......................................................................................................... 12
1.3.3 Pectin as emulsifier .................................................................................................................. 12
1.3.4 Pectin in juice extraction and cloud stability ........................................................................... 13
1.4 PECTIN IN THERMALLY PROCESSED FRUITS AND VEGETABLES ....................................................... 13
1.5 THE USE OF HIGH PRESSURE IN THE CONTEXT OF FRUIT AND VEGETABLE PROCESSING .............. 15
1.6 ELUCIDATING THE CHEMICAL STRUCTURE OF HG BY USING ANTI-HG ANTIBODIES .................... 16

iii
1.6.1 Monoclonal antibodies ............................................................................................................. 16
1.6.2 Monoclonal antibodies against specific pectin structures ....................................................... 17
1.6.3 The use of anti-HG antibodies in food-related context............................................................ 18
1.7 Objective of this study ..................................................................................................................... 20
MATERIALS AND METHODS .................................................................................................................. 21
2.1. EXPERIMENTAL SET UP ................................................................................................................... 21
2.2. PLANT MATERIALS .......................................................................................................................... 21
2.3. PRE-TREATMENTS .......................................................................................................................... 22
2.3.1 Low temperature blanching .................................................................................................... 22
2.3.2 High temperature blanching ................................................................................................... 22
2.3.3 High pressure pre-treatment ................................................................................................... 22
2.4. PREPARATION OF PUREES .............................................................................................................. 23
2.4.1 Blending .................................................................................................................................... 23
2.4.2 High pressure homogenization (at 100 bar) ............................................................................. 23
2.5. BOSTWICK RHEOMETRY ................................................................................................................ 24
2.6. WET SIEVING .................................................................................................................................. 24
2.7. EXTRACTION OF ALCOHOL INSOLUBLE RESIDUE ........................................................................... 25
2.8. FRACTIONATION OF AIR ................................................................................................................ 26
2.9.1 DETERMINATION OF DEGREE OF ESTERIFICATION OF AIR AND PECTIN FRACTIONS ................. 26
2.9.1. 1.Determination of methyl ester groups ................................................................................. 26
2.9.1.2 Determination galacturonic acid content ............................................................................. 27
2. 9.2 MOLAR MASS DISTRIBUTION. ..................................................................................................... 28
2.9.3 IMMUNO-DOT-ASSAYS ................................................................................................................ 28
2.9.4 MICROSCOPY ................................................................................................................................ 29
2.9.4.1 Preparation of sample for microscopy .................................................................................. 29
RESULTS AND DISCUSSION .................................................................................................................... 30
PECTIN STRUCTURE-FUNCTION RELATIONSHIP IN CARROT PUREE .................................................. 30
3.1. The influence of pretreatments, blending and high pressure homogenization on the
bostwick consistency of carrot purees. ........................................................................................ 30

iv
3.2. The influence of pretreatments, blending and high pressure homogenization on the particle
size distribution of carrot purees .................................................................................................. 31
3.3. Degree of esterification of alcohol insoluble residue ............................................................ 33
3.4 Fractionation of AIR. ................................................................................................................ 35
3.5 Degree of esterification of water and chelator soluble pectin fractions. ............................. 39
3.6 Immuno-dot assays ................................................................................................................ 40
3.7. Immunolabeling of non-pretreated and pretreated carrot tissue particles. ......................... 42
3.8 Molar mass distribution .......................................................................................................... 44
3.9 Conclusion ............................................................................................................................... 45
PECTIN STRUCTURE-FUNCTION RELATIONSHIP IN TOMATO PUREE ................................................ 47
4.0. Selection of pretreatments for tomato samples. .................................................................. 47
4.1 The influence of pretreatments and blending versus high-pressure homogenization on the
bostwick consistency of tomato purees ........................................................................................ 47
4.2.The influence of pretreatment and blending versus high-pressure homogenization on the
particle size distribution of tomato purees ................................................................................... 49
4.3. Degree of esterification of alcohol insoluble residue ............................................................ 51
4.4. Fractionation of AIR ................................................................................................................ 52
4.5. Degree of esterification of water and chelator soluble pectin .............................................. 54
4.6. Immno-dot assays .................................................................................................................. 55
4.7. Immunolabeling of non-pretreated and pretreated carrot tissue particles .......................... 56
4.8. Molar mass distribution ......................................................................................................... 59
4.9. Conclusion .............................................................................................................................. 59
GENERAL CONCLUSIONS ....................................................................................................................... 61
REFERENCES .......................................................................................................................................... 64

v
LIST OF SYMBOLS AND ABBREVIATIONS
AIR Alcohol insoluble residue
CDTA Cyclohexane-trans-1,2-diamine tetra-acetic acid
CSP Chelator soluble pectin
DE Degree of esterification
FITC Fluorescencein isothiocyanate
GalA Galacturonic acid
HG Homogalacturonan
HP High-pressure
HPH High-pressure homogenization
HTB High temperature blanching
LTB Low temperature blanching
MPBS Milk-PBS
NSP Sodium carbonate soluble pectin
PBS Phosphate buffer saline
PG Polygalacturonase
PME Pectinmetylesterase
RG-I Rhamnogalacturonan-I
RG-II Rhamnogalacturonan-II
RWF Rest water fraction
WSP Water soluble pectin

vi
ABSTRACT The pectin structure-function relationship in carrot and tomato purees was investigated in the
current work. Carrots were selected as a plant based food in which PME-induced pectin changes
plays an important role in structural/textural changes, whereas tomato were selected as a source
rich in PME as well as PG. To achieve the objective, pretreatment conditions were selected to
stimulate or to inactivate endogenous PME and/or PG, two important enzymes involved in pectin
conversions during processing of plant based food. For puree preparation, two types of mechanical
disruption, blending and high pressure homogenization (HPH), were used. The flow properties of
purees from non-pretreated and pretreated carrot and tomato samples were measured by using the
bostwick consistometer. Via wet sieving, the particle size distribution of the samples was
determined. Different pectin properties, such as the degree of esterification (DE) of the alcohol
insoluble residue (AIR), the solubility of pectin in AIR via fractionation, the DE of the different pectin
fractions and the molecular mass distribution of some of these fractions were analyzed. These
chemimal analyses were supplemented with analyses (immuno-dot assays as well as microscopy)
using different anti-pectin antibodies.
Puree prepared by blending non-pretreated carrots showed a high consistency, but also large
separation between the liquid and particle phase (syneresis). A low temperature blanching (LTB - 40
min at 60°C) pretreatment was applied prior to blending that aimed to stimulate PME activity. This
resulted in a puree containing a lower average degree of esterification (DE = 51.9%) and a higher
amount of chelator soluble pectin (CSP = 38.5%) and slightly larger tissue particles compared to the
non-pretreated puree. The flow properties of this puree were comparable to those of the non-
pretreated puree. When a high temperature blanching (HTB - 5 min at 95°C) pretreatment, that
should inactivate PME, was applied prior to the puree preparation step, the consistency of the
resulting puree slightly increased, but the separation between the liquid and particle fraction could
be limited. This change in flow properties, compared to the non-pretreated sample, was
accompanied by following pectin changes: a higher amount of WSP (and a lower amount of CSP and
NSP), which could be the result of thermosolubilization and depolymerization of the WSP fraction. In
all carrot samples, HPH resulted in purees with smaller tissue particles and with a lower consistency
compared to the respective blended samples. In case carrot were HTB prior to HPH, syneresis was

vii
again limited, the WSP fraction was again relative high in amount and thermosolubilization and
depolymerization of the WSP fraction was noticed.
For the tomato samples, the pretreatments conditions (HTB - 8 min at 95°C and high pressure (HP) -
10 min at 550MPa, 25°C) were selected respectively to inactivate PME and PG and to selectively
inactivate tomato PG. Blending or HPH of both non-pretreated and pretreated tomato samples
resulted in puree with almost no syneresis. The consistency of the different purees however clearly
differed. The non-pretreated, blended tomato puree had a very low consistency. This puree
contained small tissue particles and pectin with a low DE (22.4 %), of which a high portion was water
soluble (28%) and chelator soluble (24%). Depolymerization of the WSP and pectin solubilization
were demonstrated by HPSEC and immunolabeling experiments. A homogeneous distribution of low
esterified pectin epitopes as well as of pectin-calcium cross-links were visualized in this sample. Both
the HTB and HP pretreated samples resulted in blended purees with a higher consistency than the
non-pretreated sample. Both purees also contained larger particles than the non-pretreated puree.
However, the pectin properties in these samples were different: pectin in the HTB sample was
medium-high esterified (DE = 67.5 %) and rich in NSP (20%) while pectin in the HP sample was low
esterified (DE = 15.9 %) and rich in CSP (30%). HPH of non-pretreated tomatoes resulted in a puree
with a higher consistency than the blended sample. This change in consistency was not accompanied
by remarkable changes in particle size or in pectin properties measured in this study.

1
CHAPTER ONE
LITERATURE REVIEW
1. PECTIN Pectin is a naturally occurring biopolymer among the important components of plant cell walls. It is a
family of complex polysaccharides composing the primary cell walls of all plants. The study of
location, distribution and structural changes of these polysaccharides during cell and tissue
development has been of particular interest to both plant and food scientists. Regarding the
structure of these biopolymers, variations occur among plant species, tissue type and plant
developmental stage. The biological roles of these molecules within the cell wall are diverse. The
structural complexity of pectin polysaccharides and their reasonably high proportion (one-third) in
dry matter of the primary cell walls coupled with presence of cell wall-based pectin modifying
enzymes, suggest their involvement in plant growth, morphology and development.
1.1 Chemical Structure Pectic polysaccharides, abundantly found in the plant primary cell walls and middle lamellae, are
diverse in their structure. Three pectic polysaccharide domains referred to as galacturonans, are
found in plants and have been isolated from primary cell walls (O’Neil and others, 1990). These are
homogalacturonan (HG), rhamnogalacturonan-I (RG-I) and rhamnogalacturonan-II (RG-II).
1.1.1 Homogalacturonan
Homogalacturonan (HG) is a widespread linear homopolymer. Its basic structure consists of a chain
of α-D-galacturonic acid residues (GalA) linked by α(1→4) glycosidic bonds and is thought to contain
some 100-200 GalA residues (Zhan and others, 1998). HG domains are referred to as the smooth
region of pectin (Schols and Voragen, 1996). The carboxyl groups of galacturonic acid residues in HG
(Figure 1) are to varying degrees esterified with methyl alcohol or O-acetylated at C-3 or C-2 (Ishii,
1995, 1997a). The pattern and degree of methylesterification vary from one plant to another and
within a given species with developmental stage and location in the cell wall. This means that, plants
are capable of synthesizing and eventually modify pectin molecules for their survival. Highly
methylesterified pectin molecules are synthesized in the Golgi apparatus and secreted into the plant
cell wall where they are subjected to modifications (Ridley and others, 2001). In order to achieve
required functionalities within individual cell walls during plant development, the degree of HG

2
methylesterification changes as a result of endogeneous enzyme pectinmethylesterase (PME) activity
(Willats and others, 2001). De-esterification of HG appears to be a complex regulated process that
does not occur uniformly throughout tissues or cell walls (Liberman and others, 1999). This can be
expected because different plant tissues have different functions. On the other hand, un-esterified
carboxyl groups (negatively charged groups) in adjacent HG molecules may interact in presence of
divalent cation like Ca+2 forming a stable gel important for maintaining the cell walls integrity. HG
cross-linked via Ca+2 has impact on cell wall mechanical properties and cell wall matrix porosity
(Somerville and others, 2004). The interaction of HG with Ca+2 is favored in the presence of long un-
esterified galacturonic acid chains and depends on the need at a particular moment. The amount of
methyl groups, acetyl esters and their distribution in the polymer largely determine its chemical
properties and hence its suitability for particular industrial applications.
Figure 1. The primary structure of homogalacturonan . The structure of the pectic polysaccharide HG
as a linear polymer of (1,4) linked GalA residues. Representative sites of methylesterification at the
C-6 and O-acetylation at C-2 or C-3 of the carbohydrate ring are shown (Ridley and Others, 2000).
1.1.2 Rhamnogalacturonan-I
Rhamnogalacturonan-I (RG-I) is a another family of pectic polysaccharides composed of a backbone
of a repeating disaccharide *→4)- α-D-GalA-(1→2)- α-L-Rhamnose-(1→+ to which a variety of glycan
chains (principally arabinan and galactan) are attached. The sugar composition of RG-I is highly

3
heterogeneous. Because the rhamnosyl residues (20%-80%) are substituted at C-4 with neutral and
acidic side chains, RG-I is large and highly variable family of polysaccharides (Albersheim and others,
1996). RG-I is known as the hairy region of pectin (Schols and Voragen,1996).
1.1.3 Rhamnogalacturonan-II
Rhamnogalacturonan-II (RG-II) is a pectic domain which belongs to a group of polysaccharides
referred to as substituted galacturonans. They contain a backbone of seven up to ten linear 1,4-
linked α-D-GalA residues with complex side chains attached to GalA residues (O’Neill and others,
1990). RG-II is present in all higher plant cell walls and has the same structure irrespective of the
source.
1.1.4 Macromolecular organization of pectin
Further developments in structural analysis of pectic polysaccharides have revealed that in the
primary cell wall, the three pectic domains (HG, RG-I and RG-II) are covalently linked together giving
rise to a macromolecular pectic network (molecular organization). Different models have been
proposed for distribution of RG-I and HG in pectin (Matsuhashi and others 1983; Caffall and Mohnen,
2009). Pectic molecules are also involved in linkages with other cell wall components. Suggested
pectin cross-links within a cell wall and the intercellular adhesion include ionic bridges, borate-diol
esters, uronyl esters, hydrophobic interactions and ferulic acid linkages. HG is the predominant
anionic polymer in the middle lamella and primary cell walls of many plants. Calcium cross-linking of
HG chains result in formation of junction zones linking parallel or antiparallel chains (Powell and
others, 1982). Hydrophobic interactions between methoxyl groups and hydrogen bonds between
undissociated carboxyl and secondary alcohol groups may also be involved in holding pectic
polysaccharides within the plant cell wall (Oakenfull and Scott, 1984). Therefore, no dought that the
chemical structure, distribution and interaction of pectic polysaccharides at cellular level have
influence on the plant physiology and characteristics of plant-based foods.

4
1.2 PECTIN STRUCTURAL MODIFICATIONS
Being one of the cellular components, pectic polysaccharides undergo chemical/or biochemical
conversions important in plant physiology. This has triggered researchers to find more information to
understand the relationship between pectin structure and its function. These researches have been
of particular interest to plant scientists and food technologists as well. Pectin conversion reactions
can be enzymatic or nonenzymatic. Figure 2 shows the schematic presentation of possible conversion
reactions of homogalacturonan.
Figure 2. Schematic presentation of possible homogalacturonan conversion reactions in plant-based
foods and possible routes for tailoring quality parameters: PME = pectinmethylesterase,
Ca+2 = calcium crosslinking, PG = polygalacturonase, PL = pectate lyase, T = temperature,
OMe = methoxy-esters, R1/R2 = initial/terminal fragment of the pectin polymer (Sila and others,
2009).

5
1.2.1 Enzymatic pectin conversion Pectinases are hydrolytic polysaccharide-degrading enzymes abundantly found in nature. This family
of enzymes can also be synthesized by bacteria and fungi. Conversion of pectin by pectinases is
important in plant physiology and has technological applications in food and pharmaceutical
industries. In fruits and vegetables processing, action of pectinases on pectin can be exploited to
increase the extraction yield of juices and concentrates and to tailor juice cloud stability and clarity
(Demir and others 2007), controlling the rheological properties of plant based food products and
engineering the texture of fruits and vegetables (Van Buggenhout and others, 2009). HG enzymatic
conversion is widely exploited and has acquired commercial significance in food industry. The major
two HG degrading enzymes used in fruit and vegetable processing are the endogenous plant
pectinmethylesterase (PME) and polygalacturonase (PG). The catalytic activity of these two enzymes
can be influenced by intrinsic and extrinsic parameters. Pretreatments and/or processing conditions
can promote or slow down the activity of these enzymes. Preparations can stimulate enzyme activity
in intact tissue, although during some physiological processes like fruit ripening enzyme are active.
Otherwise they are activated following tissue damage and temperature changes during food
processing.
1.2.1.1 Pectinmethylesterase
PME catalyses the de-esterification of HG, creating free negatively charged carboxyl groups in the
pectin structure (Figure 2) while releasing methanol and H3O+. This reaction is of particular
importance because changes in degree of pectin methylesterification are associated with changes in
functional properties interesting for technological applications in fruits and vegetables processing. In
the course of removing the methoxyl groups, PME has two types of action patterns. The so called
block-wise action (single chain mechanism) is displayed when the enzyme removes methyl groups
from contiguous galacturonic acid residues resulting to relatively long stretches or blocks of de-
esterified GalA residues. Plant PME is thought to have block-wise action (Willats and others, 1999a).
This action pattern is presumed to be a prerequisite for effective calcium cross-linking of HG chains
(Guillotin and others, 2005). By using different methods available to map pectin, studies have
indicated that de-esterified blocks of HG occur in distinct regions of cell walls. This was observed in

6
developing carrot tissue (Knox and others, 1990) and in raw and processed carrots (Christiaens and
others, 2010). Microbial PME shows non-block-wise action leading to removal of single or limited
number of methoxyl ester groups at a time (Denès and others, 2000). PME activity yields a substrate
for PG action. The more the PME-induced de-esterification, the more the non-methoxy-esterified
galacturonic acid is liberated by endo-polygalacturonase action (Guillotin and others, 2005).
PME has been isolated from different plants. The characteristics of this enzyme vary from one plant
to another and within plant species. In general, optimal temperatures for PME catalysis vary between
45 and 55°C and at higher temperatures, the enzyme is denatured. Most plant PMEs have pH optima
between pH 4 and 8. For example carrot PME with molar mass 34.5kDa has been isolated and
characterized with its isoelectric pH >9, optimum temperature 40-50°C and optimum pH 7.3-7.4 (Ly
Nguyen and others, 2003), whereas tomato PME with molar mass 33.6kDa has isoelectric pH > 9.3,
optimum temperature 55°C and optimum pH 8.0 (Fachin and others, 2002). The activity of purified
tomato PME as a function of temperature at atmospheric pressure at pH 4.4 in presence and absence
of PG is shown in Figure 3. HG demethoxylation by PME action can thus be favored by increasing
temperature to optimum value. PME activity is also influenced by the presence of cations. Increasing
cation concentration results in increased PME activity (Nari and others, 1991).
PMEs from different plant materials differ in thermal stability. The thermal stability of PME also
depends on the matrix (buffer, juice, tissue). In tissue-type systems, PME is less vulnerable to thermal
inactivation as opposed to the purified form (Sila and others, 2007). In general, most PMEs are
inactivated at temperatures below 70°C. Tomato PME in its purified form in 40mM sodium acetate
buffer (pH 4.4) is for example completely inactivated at 70°C in 5 minutes (Figure 4). However,
different PME isoenzymes have been isolated from different plant sources. Their presence in plants is
thought to be important for modification of matrix properties at a local level in specific regions of cell
wall (Willats and others, 2001). Multiple isoenzymes have been purified from tomato fruits and
vegetative tissues (Gaffe and others, 1994). These isoenzymes differ in catalytic properties.

7
Figure 3. Normalized tomato PME activity (V0’) at atmospheric pressure as a function of temperature
at pH 4.4: in presence of tomato PG (∆) and in absence of tomato PG (×) (Verlent and others, 2006)
Most PMEs can survive high pressure treatment. At a fixed temperature, the pressure stability of
PMEs vary from moderately pressure sensitive types (>600Mpa) like carrot PME (Balogh and others,
2004) to the extremely barotolerant ones (>1GPa) like tomato PME (Crelier and others, 2001).
Activity of purified PME from different tomato varieties is reduced to 50% after 15 minutes
treatment at 850MPa (Rodrigo and others, 2005). On the other hand, pressure-temperature
combinations within a given range can have stimulatory effect on PME activity and hence pectin
conversion. The acceleration of PME catalytic activity has been noted for carrot (Sila and others,
2007) and tomato (Verlent and others, 2004). For carrot PME, the optimal conditions for substrate
conversion were 50°C in combination with 200 to 400MPa, in the case of tomato, enhanced PME
activity was reported in juice at 300 to 500MPa in combination with various temperatures (Hsu and
others, 2008).

8
1.2.1.2 Polygalacturonase
PG cleaves the α-1,4-glycosidic bonds in HG by hydrolysis. The enzyme has been found in some fungi,
bacteria, yeasts and several higher plants including tomato. De-esterified HGs are preferentially
hydrolysed by PG as compared to methylesterified HGs. This means that PME action yields a
substrate for the action of PG. It has been shown that in tomato fruit, PG exist in two isoforms, PG1
and PG2. Several studies have confirmed this. When crude extracts of PG from different tomato
varieties were assayed at pre-set temperatures for 5 minutes, Rodrigo and others (2005) observed
two inactivation phases. One phase starting at around 55°C, which correspond to PG2, the
thermolabile isoform, while the second phase starts at around 80°C corresponding to PG1, the
thermalstable isoform. In their purified form (in 40mM Sodium acetate buffer pH 4.4), PG1 and PG2
have large differences in thermal stability (Fachin, 2003; Rodrigo and others, 2005; Peeters and
others, 2004). In their study, Fachin and others (2002) found that PG in tomato juice (pH 4.4) could
be completely inactivated at temperatures higher than 90°C. Figure 4 shows the thermal stability of
purified tomato PG1 and PG2 compared to that of PME. From this figure, PG2 is thermally less stable
than PME. PGs are also sensitive to ionic environment and their pH optima range between 4 and 6,
depending on the PG isoform.
Figure 4. Thermal stability of purified tomato PG1, PG2 and PME (Rodrigo and others, 2005).
9
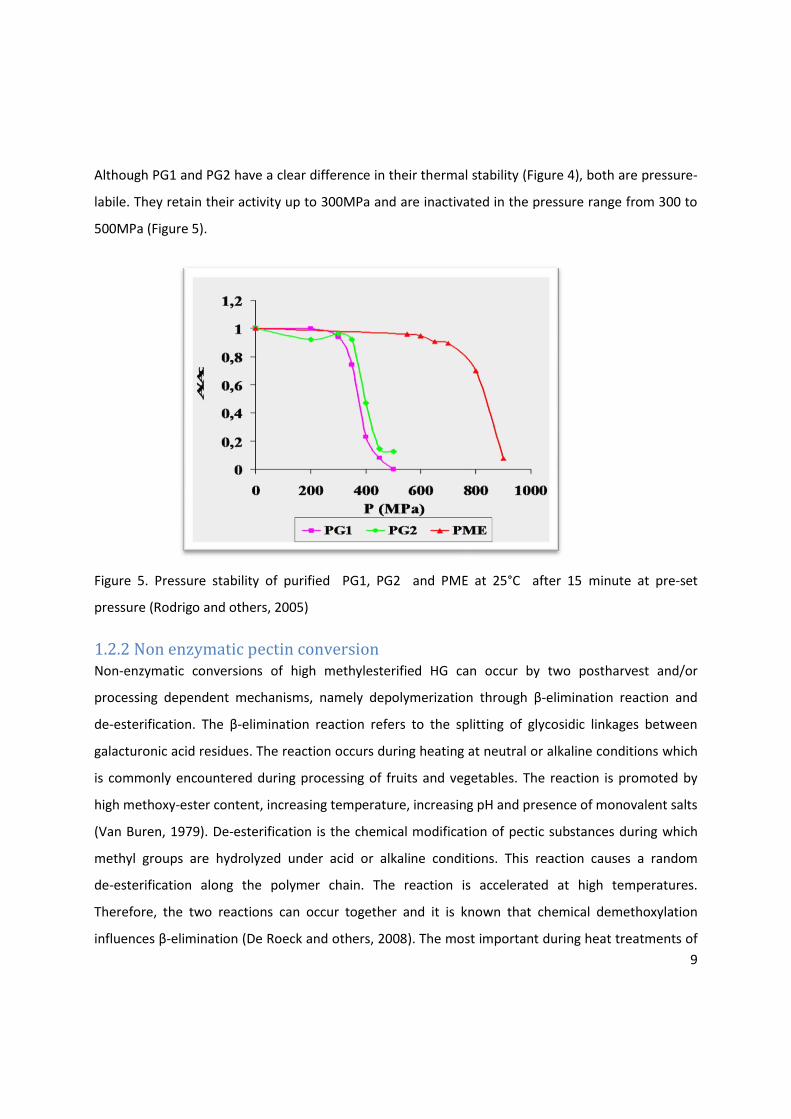
Although PG1 and PG2 have a clear difference in their thermal stability (Figure 4), both are pressure-
labile. They retain their activity up to 300MPa and are inactivated in the pressure range from 300 to
500MPa (Figure 5).
Figure 5. Pressure stability of purified PG1, PG2 and PME at 25°C after 15 minute at pre-set
pressure (Rodrigo and others, 2005)
1.2.2 Non enzymatic pectin conversion Non-enzymatic conversions of high methylesterified HG can occur by two postharvest and/or
processing dependent mechanisms, namely depolymerization through β-elimination reaction and
de-esterification. The β-elimination reaction refers to the splitting of glycosidic linkages between
galacturonic acid residues. The reaction occurs during heating at neutral or alkaline conditions which
is commonly encountered during processing of fruits and vegetables. The reaction is promoted by
high methoxy-ester content, increasing temperature, increasing pH and presence of monovalent salts
(Van Buren, 1979). De-esterification is the chemical modification of pectic substances during which
methyl groups are hydrolyzed under acid or alkaline conditions. This reaction causes a random
de-esterification along the polymer chain. The reaction is accelerated at high temperatures.
Therefore, the two reactions can occur together and it is known that chemical demethoxylation
influences β-elimination (De Roeck and others, 2008). The most important during heat treatments of

10
fruits and vegetables is the β-eliminative depolymerization reaction (Sila and others, 2006). Because
most plant based foods have a pH above 4.5 and are processed at temperatures above 80°C, their
pectic substances can undergo β-elimination reaction (Sila and others, 2005). The reaction leads to
pectin solubilization, decreased cell-cell adhesion and eventually tissue softening.
Efforts are geared towards limiting pectin degradation during processing of fruits and vegetables in
order to improve or preserve texture. De Roeck and others (2008) studied the influence of
temperature and combined high pressure/high temperature on pectin degradation in model systems.
They found that the rate constants of both pectin demethoxylation and β-elimination reactions
increased with increasing temperature. The activation energy for β-elimination reaction in several
studies has been found to be higher than that of de-esterification reaction, implying that any
temperature rise result in stronger acceleration of β-elimination than of de-esterification (De Roeck
and others, 2008; Fraeye and others, 2007). With respect to high pressure in model systems, the rate
of β-elimination decreases when high pressure is combined with elevated temperature. However
the exact cause of low rate of β-elimination during high pressure treatment is difficult to establish
because many parameters influence the reaction rate (De Roeck and others, 2008). On the other
hand, it has been shown that tailoring the methoxy-ester content of pectin can control the β-
elimination reaction (Vu and others, 2006; De Roeck and others, 2008). The de-esterification reaction
in food systems can be achieved by in situ activation of endogenous or exogenous PME (see
paragraph 1.2.1.1) using pretreatment conditions. The activation of PME by pretreatments during
thermal processing of carrot resulted in pronounced de-esterification and significant texture
retention ( Sila and others, 2006).
Another mechanism leading to pectin degradation is acid hydrolysis (see Figure 2) which occurs
during heat treatment in acid conditions (pH<3.0). The reaction is influenced by degree of
methylesterification and is less important during regular food processing.

11
1.3 PECTIN FUNCTIONAL PROPERTIES Besides being an important cellular component which plays different roles in plant development,
pectin is the main contributing factor to quality attributes of many plant based foods, particularly
texture (Van Buren, 1979) and rheological properties (Fraeye and others, 2009). The fact that pectic
substances are more soluble and more chemically reactive than other cell wall polymers and that
they constitute one-third of dry matter of the primary cell wall, explains their dominance in
influencing textural quality characteristics of many plant based foods. This implies that most textural
and/or rheological changes encountered during food handling and processing are accompanied by
chemical/biochemical reactions in which the structure of pectic substances is modified.
Understanding pectin conversions in plant based foods and factors regulating the process is
important to food technologists, whose main role is to control reactions during processing of foods
to meet a given set of quality characteristics. This can be achieved when enzymatic activity is
carefully controlled by intelligent postharvest handling and/or processing.
1.3.1 Pectin and fruit ripening During fruit development, softening is one of the most familiar and pronounced changes associated
with ripening. This is a process of practical value because it confers specific characteristics on fruit
and in some cases it is used as an index of maturation for making harvest decisions. Sometimes, the
shelf life of fruits can be strongly limited by the loss of firmness. Fruit softening is related to cell
separation, due to hydrolytic structural changes in the cementing intercellular wall material (middle
lamella) and the degradation of cell wall material (Fischer and others, 1991). Cell wall degradation is
catalyzed by endogenous and/or exogenous hydrolytic enzymes, including pectate lyases (PLs),
polygalacturonases (PGs) and pectinmethylesterases (PMEs) (Brummel and Harpster, 2001).
Softening in tomatoes is often well correlated with PG activity. Zainon and others (2004) observed
varying levels of PG activity in unripe and ripe tomato tissues. The enzyme activity increased with
ripening and a 500% increase in activity in tomato was recorded.

12
1.3.2 Pectin as a gelling argent In food technology, pectin is well known for its involvement in gel formation. This functional property
has been exploited in the area of food technology as well as in the area of pharmaceutical
applications. The gelling and stabilizing properties of pectin make it an important ingredient of jams,
jellies and confectionary products. The quality attributes of these products like viscosity can differ
depending on the source of pectin, among other factors. It has been found that the suitability of
pectin for any application is governed by its structural features such as neutral sugar content, molar
mass, ferulic acid substitution, proportion of smooth and hairy regions, amount of methoxy acetyl
esters and distribution of ester groups in the polymer (Daas and others, 1999; Braccin and others,
1999). These structural features can be altered by conditions of the system (such as ionic strength,
temperature, pH, dissolved salts and so on) and the type of post-harvest handling processes,
resulting in changes in pectin functionality. Furthermore, the extraction procedure used and
endogenous/exogenous enzyme present, highly influence the pectin gelling ability (Strom and others,
2007). Pectin with degree of methylesterification (DE) above 50% form gels at pH≤3.5 in presence of
more than 55% sugar and mainly hydrophobic interactions and hydrogen bonds are involved
(Oakenfull and Scott, 1984). Pectin with DE<50% can form gels in presence of Ca2+ and absence of
sugar. The time required for gel formation and the molecular interactions involved also depend on
the degree of methylesterification (DE) and the distribution of methyl esters of pectin. Decreasing DE
for high methylesterified pectin (DE>50%) and adding Ca2+ result in faster gel formation (Löfgren and
others, 2005). Highly acetylated pectin with relatively small molar mass has poor gelling ability
(Renard and Thibault, 1993).
1.3.3 Pectin as emulsifier Pectin is among the substances known to reduce the interfacial tension between oil and water
phases, hence can be used as emulsifier. In their study, Leroux and others (2003) found that the
emulsifying properties of pectin were significantly influenced by its molecular weight, protein and
acetyl content. Highly acetylated pectin with relatively small molar mass shows good emulsion
properties (Drusch, 2007). The pectin emulsifying properties depend on its interaction with other
components present in a particular system. For example the pectin fine structure allows interaction
with whey protein resulting in improved emulsifying properties of this milk protein. In their study,

13
Neirynck and others (2004) found that the heat-induced covalent binding of low methylesterified
(DE<50%) pectin to whey protein resulted in substantial improvement of the emulsifying properties
at pH 5.5. Among different pectin sources, sugar beet has pectin with good emulsifying properties
(Leroux and Others, 2003).
1.3.4 Pectin in juice extraction and cloud stability Pectinases are used as processing aid in juice extraction to facilitate juice recovery. Their action in
combination with other endogenous and/or exogenous enzymes during juice extraction are also
associated with improved juice quality by allowing extraction of more soluble solids and flavor
compounds. For example, when incubated with apple pulp, pectinases degrade soluble pectin
leading to increased juice extraction (Kashyap and others, 2001). If one aims at producing clear apple
juice, the use of pectin degrading enzymes can help removing the suspended matter in pulp. PME
activity followed by PG activity (Figure 2) prior to pressing and centrifugation can serve this purpose,
however complete breakdown of pectin requires correct proportion of different enzymes (Versari
and others, 1999). For cloudy products, these enzymatic reactions can result cloud loss, negatively
affecting product acceptability.
1.4 PECTIN IN THERMALLY PROCESSED FRUITS AND VEGETABLES The textural and rheological properties of many plant based foods can partly be explained by the
variation in their pectin content and composition and the subsequent processing steps applied. In
both traditional and novel food processing technologies, the focus is to use available knowledge in
basic food components and optimize processes to meet a given set of product quality attributes. The
knowledge in basic pectin-structure function has been exploited in food processing to achieve some
desired process-induced textural/rheological changes.
Thermal processes (blanching, pasteurization, cooking and sterilization) applied to plant-based foods
are associated with textural changes, related to structural characteristics of the food.
Thermally-induced texture loss partly results from solubilization and depolymerization of pectic
polymers involved in cell-cell adhesion (Waldron and others, 2003). During thermal processing of
plant based foods, depending on the pH and DE, pectin degradation can either be through acid
hydrolysis or via β-elimination reaction. The latter reaction (see also paragraph 1.2.2) is favored at

14
elevated temperatures (>80°C) and pH≥4.5 (Greve and others, 1994). In their study, Sila and others
(2006a) found that β-elimination reaction is the main contributing factor to texture degradation
during thermal processing of carrots. The dependence of this reaction on the DE of pectin in a given
plant food material offers an opportunity for its control, since targeted DE manipulation can be
achieved through activation of endogenous/exogenous PME (see paragraph 1.2.1.1). This can be
achieved by preheating the food material at temperatures between 50 and 70°C for ≥30 minutes
(Roy and others, 2001). The activated PME de-esterifies pectic polysaccharides which then cross-link
in presence of divalent cations (Figure 2). As a consequence, the β-elimination reaction is also
reduced. The combined effect being reduced texture degradation.
As processing conditions and pretreatments applied to plant based foods are associated with pectin
structural changes, they influence pectin solubility. Sila and others (2006a) found that low
temperature blanching and high pressure (paragraph 1.6) applied to carrot samples result in
significant reduction in DE and a concomitant increase in sodium carbonate soluble pectin (NSP) and
decrease in water soluble pectin (WSP). When pretreated samples are subsequently thermally
processed, this results in conversion of NSP and chelator soluble pectin (CSP) into WSP (Sila and
others, 2006b).
During conventional thermal processing of tomato, coldbreak (65°C) or hotbreak (77-93°C) processes
are applied. During coldbreak processes, endogenous PME activity is promoted resulting in
de-esterified pectin which can be depolymerized by tomato PG. The combined action of PME and PG
results in dramatic decrease in consistency of tomato product (Lopez and others, 1997). On the other
hand, during hotbreak processes both enzymes are inactivated which results in products with high
viscosity (Goodman and others, 2002). This example shows that careful control of the activity of
endogenous enzymes is an important issue towards quality preservation of tomato products.

15
1.5 THE USE OF HIGH PRESSURE IN THE CONTEXT OF FRUIT AND
VEGETABLE PROCESSING In fruits and vegetables processing , the major challenge is to produce a product with desired quality
given the quality attributes of raw materials. Texture of plant based foods, is a focus of modern food
processing technologies. Efforts are geared towards better understanding of how the texture of the
final product is related to plant cell wall structural components. The present knowledge on important
cell wall component (pectin) and its enzymatic and non-enzymatic degradation during processing has
been an important tool in improving texture of processed plant based foods. In this context of pectin
engineering, also the possibilities of high pressure processing have been explored.
In the context of using high pressure as a pre-heating treatment, increasing temperature and
pressure in specific ranges has been shown to have a stimulatory effect on PME activity. In their
study on intact carrot tissues, Sila and others (2007) noticed that PME activity increased with
increasing temperature (in the range 30-60°C) combined with pressure (in range 0.1- 400 MPa). The
optimum conditions for PME activity was found to be 60°C in combination with pressures between
100 and 400MPa. These high pressure pretreatment conditions were shown to lead to pronounced
texture retention during subsequent thermal processes when compared to not pretreated samples
(Sila and others, 2006).
The same principle can be used to improve the texture of fruits with low PME levels. In case of for
example strawberries, high pressure processing was successfully applied in case the fruits were
infused with exogenous PME and Ca+2 prior to the processing step. In their study, Fraeye and others
(2009) found differences in firmness among the PME and Ca+2 infused strawberries after thermal
(70°C-0.1MPa), high pressure (25°C-550MPa) and combined thermal-high pressure (70°C-550MPa)
processes. They observed that the strong decreases in DE (from 73.3 to 24.7% and from 66.2 to
13.0%) after respectively high pressure and thermal-high pressure processing of infused strawberries
was associated with higher tissue firmness when compared to thermally treated ones. This
observation could be explained partly by PME activation during high pressure processing that

16
induced pectin structural changes (DE%) which in turn enhanced texture (firmness) due to pectin
crosslinking in presence of Ca+2.
In the context of the rheological quality of tomato based products, high pressure processing is an
interesting tool for enzyme inactivation. Given the stability of tomato PME and PG at different
combinations of temperature and pressure, it is possible to selectively inactivate PG, while retaining
PME activity. For tomato puree, Verlent and others (2006) found that the combination of 40°C and
400MPa is optimal for this selective enzyme inactivation. Upon using these conditions, tomato
products with good rheological properties can be obtained.
1.6 ELUCIDATING THE CHEMICAL STRUCTURE OF HG BY USING ANTI-HG
ANTIBODIES Since the functionalities of HG are related to its structure, detailed understanding of the HG chemical
structure has been of particular interest. Extraction methods are available for structural analysis of
pectin. However, by using these methods, it is impossible to extract pectin without possible
destruction of its structure. Therefore, ex situ analysis of the structure of pectin can give an
incomplete or misleading picture of pectin. Immunolocalization techniques use monoclonal
antibodies developed against cell wall polymers to locate polymers in situ. This has emerged as an
important tool in providing more information about the pectin structure.
1.6.1 Monoclonal antibodies An antibody is a large Y-shaped protein (figure 6) which is part of the immune response of
vertebrates to identify and neutralize foreign objects (antigen). The variable small region at the tip of
the protein allows different antigens to bind. Monoclonal antibodies are antibodies produced by only
one clone and therefore, they have only one type of antigen-binding site (Goding, 1996). Because of
that, such antibodies are very specific for the type of antigen they recognize. Different methods are
available for making antibodies. There are cell-based methods which generate hybridoma
monoclonal antibodies (VandenBosch and others, 1989; Knox and others, 1990). Recent
developments describe the gene-bases approaches for generation of monoclonal antibodies (Willats
and others, 1999).

17
Figure 6. Schematic representation of an antibody ( Mian and others, 1991).
1.6.2 Monoclonal antibodies against specific pectin structures A number of monoclonal antibodies have been developed specifically to different HG structures
(table 1). Anti-HG antibodies can recognize pectic molecules with different degree of
methylesterification and can be used to analyze HG structure in context of cell wall architecture and
plant cell development (Willats and others, 2001b). The epitopes of antibodies JIM5 and JIM7 are
partially methylesterified HGs, whereby JIM5 binding increases with decreasing degree of
methylesterification (Willats and others, 2000). Knox and others (1990) found that the epitopes for
these two antibodies are not uniformly distributed in cell walls of carrot root apex. JIM5 epitope was
located to the inner surface of the primary cell wall adjacent to the plasma membrane, in the middle
lamella and abundantly to the outer surface of the intercellular space whereas the JIM7 epitope was
located evenly throughout the cell wall. Different other studies have concluded that, the JIM5
epitopes probably contains both methyl-esterified and un-esterified galacturonic acid residues
(Willats and others, 2000; Clausen and others, 2003). Another monoclonal antibody, namely PAM1,
with specificity for de-esterified stretches of HG resulting from the blockwise action of plant PME has
been isolated (Willats and others, 1999). Furthermore, in attempt to fully explore HG structure and
its variations within cell walls, other monoclonal antibobies, designated LM18, LM19 and LM20, have
been isolated, characterized and tested for their recognition of HG pectic domain (Verhertbruggen

18
and others, 2009). With regard to their binding, LM18 and LM19 bind preferentially to de-
esterified/saponified HG similar to JIM5 while LM20 binds to epitope similar to JIM7 (Verhertbruggen
and others, 2009).
Table 1. Specifications of anti-HG antibodies.
1.6.3 The use of anti-HG antibodies in food-related context Antibodies raised against specific polysaccharides have potential to be used in food-related
applications. They can be used for specific localization of polysaccharides in plant based foods. Also
in other products, such as dairy products, they are used to investigate the structure of pectin
molecules. For example JIM7, was used to localize pectin in yoghurt in which the polysaccharides are
used as stabilizer (Arltoft and others, 2006).
In the context of processing of plant based foods, few studies have been performed in which
antibodies were applied to study pectin structure. For example, in case of PME-infusion studies,
JIM5 and JIM7 were successfully used to visualize changes in cell wall pectin after vacuum infusion
processing and during storage of egg plant (Bonjongsinsiri, Shields and Wicker, 2004). In another
study, Christiaens and others (2010) used anti HG antibodies to understand pectin structural changes
induced by PME activation during thermal pre-processing of carrots and broccoli samples. They
found that decreasing DE by PME thermal stimulation could be visualized using the anti HG
antibodies PAM1 and LM18, both recognizing pectin molecules with low DE. Their binding was found
to be high in long time (48h at 60°C) blanched carrot and that de-esterification by PME took place in
Monoclonal
antibody
Type Antigen References
JIM5 Rat IgG2a HG VandenBosch and others, (1989); Knox and
others, (1990)
JIM7 Rat IgA HG Knox and others, (1990)
LM7 Rat IgM HG Willats and others, (2001b)
LM18 Rat IgG HG Verhertbruggen and others, (2009)
LM19 Rat IgM HG Verhertbruggen and others, (2009)
LM20 Rat IgM HG Verhertbruggen and others, (2009)
PAM1 scFv De-esterified
HG blocks
Willats and others, (1999); Manfield and others,
(2005)
2F4 Mouse IgG1 Ca+2
-cross-linked HG Liners and others, (1989)

19
discrete regions of the inner face of the cell wall adjacent to plasma membrane and in tricellular
junctions and middle lamella for broccoli.

20
1.7 Objective of this study It is the objective of this study to gain insight into the structure –function relationship of pectin in
tomato and carrot purees. Anti–HG antibodies in conjunction with chemical pectin characterization
methods, will be used to provide more information on pectin structural changes during food
processing, in particular, during low temperature and high temperature blanching pretreatments
and during blending as compared to high-pressure homogenization.
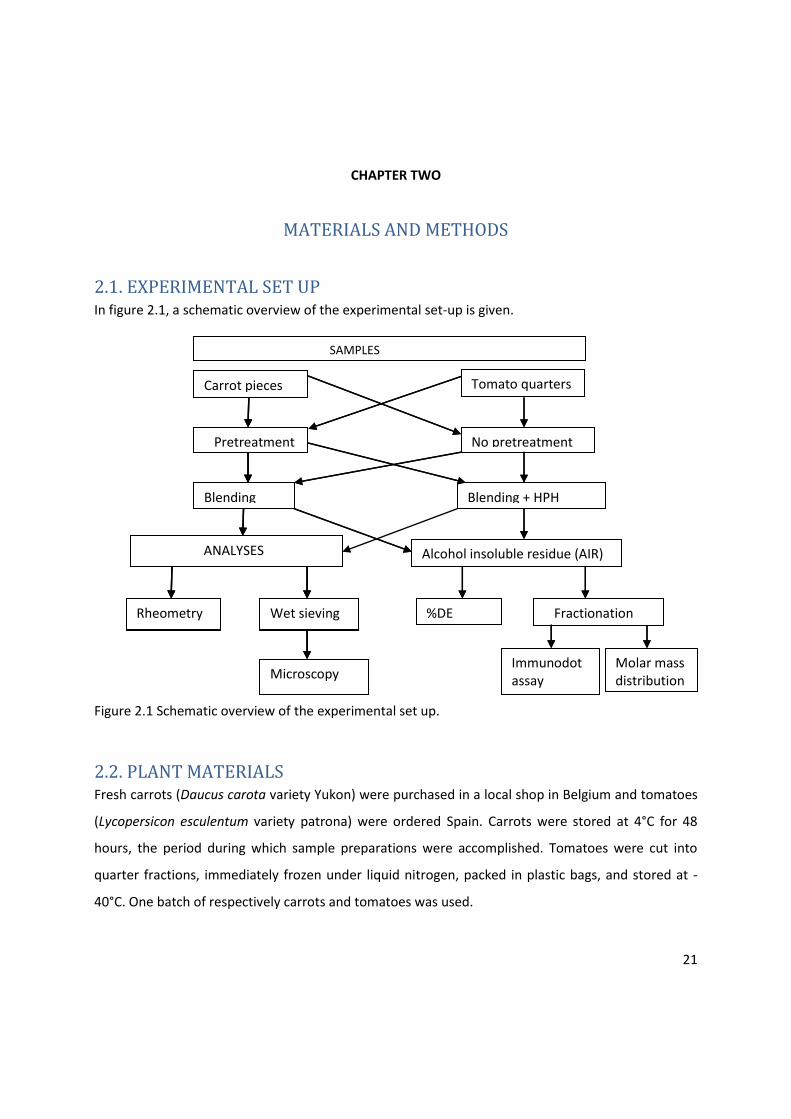
21
CHAPTER TWO
MATERIALS AND METHODS
2.1. EXPERIMENTAL SET UP In figure 2.1, a schematic overview of the experimental set-up is given.
Figure 2.1 Schematic overview of the experimental set up.
2.2. PLANT MATERIALS Fresh carrots (Daucus carota variety Yukon) were purchased in a local shop in Belgium and tomatoes
(Lycopersicon esculentum variety patrona) were ordered Spain. Carrots were stored at 4°C for 48
hours, the period during which sample preparations were accomplished. Tomatoes were cut into
quarter fractions, immediately frozen under liquid nitrogen, packed in plastic bags, and stored at -
40°C. One batch of respectively carrots and tomatoes was used.
SAMPLES
Carrot pieces Tomato quarters
Pretreatment No pretreatment
Blending Blending + HPH
ANALYSES Alcohol insoluble residue (AIR)
%DE Fractionation
Molar mass distribution
Immunodot assay
Rheometry Wet sieving
Microscopy
SAMPLES
Carrot pieces Tomato quarters
Pretreatment
Blending
ANALYSES
Rheometry Wet sieving
Microscopy
No pretreatment
Blending + HPH
Alcohol insoluble residue (AIR)
%DE Fractionation
Molar mass distribution
Immunodot assay
SAMPLES
Carrot pieces Tomato quarters
Pretreatment
Blending
ANALYSES
Rheometry Wet sieving
Microscopy

22
2.3. PRE-TREATMENTS Carrot samples were subjected to both low temperature blanching (LTB) and high temperature
blanching (HTB), whereas for tomatoes samples, HTB and high pressure (HP) pre-treatments were
done.
2.3.1 Low temperature blanching Carrots were peeled and cut into small non-calibrated pieces, spread out into one uniform layer in
polyethylene bags, vacuum sealed and then heated at 60°C for 40 minutes in a temperature
controlled water bath (Julabo UC, Merck Belgolabo, Belgium). The samples were then cooled in an
ice bath.
2.3.2 High temperature blanching Small non-calibrated pieces of peeled carrots were placed into polyethylene bags, vacuum sealed and
heated at 95°C for 5 minutes in a temperature controlled water bath (Julabo UC, Merck Belgolabo,
Belgium) and subsequently cooled in an ice water bath. For tomatoes, quarter fractions of fresh
tomatoes were put into the polyethylene bags followed by vacuum sealing prior to heating at 95°C
for 8 min in a temperature controlled water bath. Then the samples were cooled in ice water, frozen
using liquid nitrogen and stored at -40°C for further experiments.
2.3.3 High pressure pre-treatment High pressure (HP) pre-treatment of tomatoes was carried out at laboratory scale in a single vessel
(volume: 590mL, internal diameter: 50mm, height: 300mm) HP apparatus (Engineered Pressure
Systems International, EPSI, Temse, Belgium). The equipment allows pressures up to 600MPa in
combination with temperatures between -30°C and 100°C. Tomatoes in quarter fractions (~180g)
packed in double polyethylene bags were treated at 25°C and 550MPa for 10 minutes. Pressure
build up and pressure release were almost instantaneous and treated samples were immediately
cooled in an ice water bath.

23
2.4. PREPARATION OF PUREES
2.4.1 Blending To 200g of raw, low temperature and high temperature blanched carrots, water was added in a 1:1
weight ratio and blended in a laboratory blender (Waring blender, Torrington. Connecticut, USA) for
1min, 20s at low speed and 40s at high speed. For tomatoes, blending was done using the BUCHI
mixer (Buchi Labortechnik AG, Flawil Switzerland) and no water was added. After thawing (15min at
25°C) pre-treated and non-pretreated tomatoes, were placed in a jar and mixed 3 times for 5s. Seeds
and large skin pieces were removed by passing the blended sample through a sieve (1 mm pore
size).
2.4.2 High pressure homogenization (at 100 bar) High pressure homogenization (HPH) of blended carrots and tomatoes was done at 100bar by using
high pressure homogenizer (GEA Nitro Soavi S.P.A, Parma, Italy) (Figure 2.2 ).
Figure 2.2 High pressure homogenizer (GEA Nitro Soavi S.P.A, Parma, Italy).

24
2.5. BOSTWICK RHEOMETRY With the reservoir gate closed, 100g of puree was placed in a reservoir of a clean and dry Bostwick
rheometer (Figure 2.3). Immediately after filling, the trigger was pushed down to open the gate
allowing the puree to flow along the trough and simultaneously a stop watch started. After 30s,
readings on the rheometer ruler were recorded for both the pulp and the liquid fraction. For each
sample the procedure was repeated twice.
Figure 2.3 Bostwick rheometer
2.6. WET SIEVING Sieves of different pore sizes (1mm, 500µm, 250µm, 125µm, 80µm and 40µm) staked on a
mechanical shaker were used to fractionate purees in classes with different tissue particle sizes.
Before installation, weights of clean and dry sieves were measured and recorded. 200g of sample
was sieved using constant flow rate of deionized water, 1.5mm amplitude and a running time of 2
minutes. The rest water fraction was collected and its volume recorded. Sieves were dried and re-
weighted to determine the dry matter retained on each. The relative mass fraction of puree material
washed out of the sieve column denoted as rest water fraction (MWO*) was calculated as the
difference between the amount of sample sieved (200g) and the sum of the material retained in all
the sieves. This fraction contains tissue particles of size less than 40µm. Also a small amount of dry
matter from each sieve was preserved in 10 ml of 10% ethanol and stored at 4°C for immunolabeling
experiments.

25
Figure 2.4 Installed sieves on shaker
2.7. EXTRACTION OF ALCOHOL INSOLUBLE RESIDUE Cell wall material was extracted in form of alcohol insoluble residue (AIR) as described by McFeeters
and Armstrong (1984). 15g of puree was homogenized 3 times 6 seconds in 96ml of 95% ethanol
using a mixer (Buchi mixer B-400, Flawil, Switzerland ). The suspension was filtered by using filter
paper (MN 615 Ø 90mm, Macherey-NAGER GmbH & Co, Düren, German). The residue was re-
homogenized in 48 ml of 95% ethanol and filtered again. The residue was then homogenized in 48 ml
of acetone at 4°C for 10 minutes and filtered. The final residue was dried overnight at 40°C to get
AIR. The AIR obtained was ground using a motor and pestle and stored in a desiccators until analysis.

26
2.8. FRACTIONATION OF AIR AIR was fractionated into water soluble (WSP), chelator soluble (CSP) and sodium carbonate soluble
pectin (NSP). WSP was fractionated by using the extraction procedure described by Braga and others
(1998). In this method, 0.25g of AIR was suspended by stirring in 45ml of hot deionized water (100°C)
for 5min. The resulting solution was cooled under tap water, filtered using filter paper (MN 615 Ø
90mm, Macherey-NAGER GmbH & Co. Düren, German), the filtrate (WSP) was collected and
adjusted with deionized water to 50ml. The residue was re-suspended in 45ml of 0.05M cyclohexane-
trans-1,2-diamine tetra-acetic acid (CDTA) in 0.1M potassium acetate (pH 6.5) in a shaking water
bath at 28°C for 6h, preceded by 15 min stirring at room temperature (Chin and others, 1999). After
filtration, the filtrate (CSP) was adjusted with CDTA in 0.1M potassium acetate (pH 6.5) to 50ml. The
residue was re-incubated in 45ml 0.05M sodium carbonate (Na2CO3) containing 0.02M NaBH4 for
16h at 4°C with constant stirring, followed by 6h at 28°C in shaking water bath (Chin and others,
1999). After filtration, the filtrate (NSP) was adjusted to 50ml with Na2CO3 . All the pectin fractions
were frozen using liquid nitrogen and stored at -40°C. Later, all the fractions were analyzed for
galacturonic acid content prior to lyophilization followed by determination of molar mass
distribution. Lyophilization was done using a freeze-drier (Christ alpha 2-4, Osterode, German), after
freezing 25mls of each fraction using liquid nitrogen. The powder was kept in a desiccators until
subsequent analysis.
2.9.1 DETERMINATION OF DEGREE OF ESTERIFICATION OF AIR AND
PECTIN FRACTIONS The DE of the pectin was determined as the ratio of moles of methyl ester groups to the moles of
anhydrous galacturonic acid in AIR, WSP and CSP.
2.9.1. 1.Determination of methyl ester groups The pectin methyl ester groups content was determined after hydrolyzing the ester bonds of the
pectin with 2M sodium hydroxide (NaOH) as described by Ng and Waldron (1997). In this procedure,
to 20mg of AIR, 8ml of demineralized water was added. The mixture was sonicated for 10 min to
suspend the AIR. 3.2ml of 2M NaOH was added to hydrolyze the samples for 1h at 20°C . The samples
were neutralized by adding 3.2ml of 2M hydrochloric acid and equlibriated at 25°C for 15 min.

27
Subsequently, the samples were adjusted to 50ml with 0.0975M phosphate buffer (pH 7.5). The
procedure was done in duplicate for each sample.
The released methanol was quantified calorimetrically as described by Klavons and Benett (1986). In
this procedure, to 1ml of hydrolyzed samples, 1 unit alcohol oxidase activity was used to oxidize
methanol to formalaldehyde at 25°C for 15 min. Formalaldehyde reacted with 2,4-pentanedione in
ammonium acetate and acetic acid at 58°C for 15 min, resulting into a yellow-colored compound
(3,5-diacetyl-1,4-dihydro-2,6-dimethylpyridine). The absorbance of the yellow colored compound
was measured by a spectrophotometer (Utraspec 1100 pro UV/Visible Spectrophotometer, GE
Healthcare, Uppsala, Sweden) at 412nm and 25°C. The procedure was done in triplicates for each
hydrolyzed sample. The concentration of methanol (µg/ml phosphate buffer) for each sample was
determined by using a pre-prepared methanol standard curve.
2.9.1.2 Determination galacturonic acid content To determine the moles of galacturonic acid (GalA), 10mg of AIR was hydrolyzed in duplicate with
8ml of concentrated sulphuric acid (98%) in presence of water with a constant stirring in an ice
bath for complete hydrolysis (1h), according to the method of Ahmed and Labavitch (1977). After
hydrolysis, the samples were diluted to 25ml or 50ml with demineralized water depending on
expected amount of GalA content. GalA acid content of pectin was determined calorimetrically as
described by Blumenkrantz and Asboe-Hansen (1973). In this procedure, 3.6ml of cold sulphuric acid-
sodium tetraborate solution (0.0125M sodium tetraborate in 98% sulphuric acid) was added to 0.6ml
of hydrolyzed samples in an ice bath and heated at 100°C for 5 min. Then to each sample, 60µl of
m-hydroxydiphenyl-solution (0.15% 3-phenylphenol in 0.5% NaOH) was added and the absorbance
was measured at 520nm and 25°C using spectrophotometer (Utraspec 500 pro UV/Visible
Spectrophotometer, GE Healthcare, Uppsala, Sweden). This procedure was done in triplicate for each
hydrolyzed sample. Finally, the concentration of GalA (µg/ml water) was determined using a
pre- prepared GalA standard curve.
For the liquid pectin fractions (WSP and CSP), the same procedures were followed for determination
of methyl ester groups and galacturonic acid content. In this case, 2 ml of each of the liquid pectin
fractions (WSP and CSP) was hydrolysed during methanol and galacturonic acid analyses. The

28
hydrolysed samples were diluted to 25 ml with distilled water for galacturonic acid analysis and with
phosphate buffer for methanol analysis.
2. 9.2 MOLAR MASS DISTRIBUTION. The molecular mass distribution of WSP, CSP and NSP was studied using high-performance size
exclusion chromatography (HPSEC). This was done by using the Akta purifier system equipped with a
mixed bed column of Bio-gel TSK (length: 300mm, diameter: 7.5mm, pole size: 100-1000Å, particle
size: 13µm, maximum pressure: 300psi) (Biorad Labs, Richmond, CA) in combination with a TSK guard
column. Elution was done at 35°C with 0.05 NaNO3 at flow rate of 0.7mL/min for 25 min. The eluent
was monitored by a Shodex R10-101 refractive index detector (Showa Denko, K.K., Tokyo, Japan.
Pullulan standards (MW range 188-788000), which have a structure and hydrodynamic
characteristics similar to polygalacturonic acid, were used.
2.9.3 IMMUNO-DOT-ASSAYS Serial dilutions of the pectin fractions WSP and CSP (1/10, 1/100 and 1/1000) in demineralised water
were made. Thereafter, 1µl aliquots for each dilution were dotted onto a piece of nitrocellulose
membrane (Hybond ECL, GE Healthcare, Uppsala, Sweden). The nitrocellulose membrane was then
air-dried for 30min prior to 1h blocking with 5% MPBS. The membrane was washed 6 times with
running demineralised water and 3 times for 5min with PBS and incubated with primary antibody for
1h and 30 min at room temperature. Hybridoma supernatant of JIM5 and JIM7 were diluted 1/10 in
1%MPBS, whereas hybridoma supernatant of LM18, LM19 and LM20 were diluted 1/20 in 1%MPBS.
JIM5 and PAM1 were used at concentration of 25µg/ml and 10µg/ml respectively. After washing, the
membranes were incubated with secondary antibodies. An anti-rat IgG horseradish peroxidase
conjugate (Nordic Immunology, Tilburg, The Netherlands) diluted 1/1000 in 1%MPBS was used as
secondary antibody for JIM5, JIM7, LM18, LM19 and LM20. For PAM1, anti-polyhistidine peroxidase
antibody (Sigma-Aldrich, St. Louis, United States), diluted 1/2000 in 1%MPBS was used as a
secondary antibody. The membrane was again washed with water and PBS. Finally, a color reagent
(25ml PBS, 5ml chloronapthol, 16.7µl 27%(v/v) hydrogen peroxide) was added and left in dark for 40
min for color development. Positive blots were stained blue-purple.

29
2.9.4 MICROSCOPY For both tomato and carrot samples, microscopic examination was done for selected fractions
recovered after wet sieving. Anti-homogalacturonan antibodies JIM5, JIM7, LM18, LM19, LM20, 2F4
and PAM1 (PlantProbes, Leeds, United Kingdom) were used for immunolabeling of pectin epitopes.
Table 1 shows the specifications and primary references for the generation of these monoclonal
antibodies.
2.9.4.1 Preparation of sample for microscopy Ethanol in preserved samples (see paragraph 2.6) was removed by centrifugation (5 minutes, 3000g
and 22 °C). The samples were washed twice by centrifugation (5 minutes, 3000g and 22 °C) using 1
ml of phosphate buffer saline (PBS: 140mM Nacl, 2.7mM Kcl, 8.0 mM Na2HPO4, 1.5 mM KH2PO4, pH
7.35) prior to incubation with primary antibodies. The primary antibodies were diluted in phosphate
buffer saline containing 3% milk (MPBS) and incubated with the sample for 1h and 30 min at room
temperature with continuous gentle shaking. JIM5 and JIM7 were used as 5-fold dilutions of
hybridoma supernatant, whereas LM18, LM19 and LM20 were used as 10-fold dilutions. PAM1 was
used at a concentration of 20 µg/ml. Thereafter, samples were washed 3 times 5 min in 1 ml PBS by
centrifugation (400g, 22°C). For the visualization of JIM5, JIM7, LM18, LM19 and LM20, secondary
antibody anti-rat IgG coupled with fluorescein isothiocynate (anti-rat FITC) (Nordic Immunology,
Tilburg, The Netherlands) diluted 1/20 in MPBS was used. PAM1 was visualized using a three stage
labeling procedure whereby the samples after incubation with primary antibody were subsequently
incubated with an anti-polyhistidine (anti-polyHIS) antibody (Sigma-Aldrich, St. Louis, United States)
followed by an anti-mouse antibody coupled to FITC (anti-mouse FITC) (Nordic Immunology, Tilburg,
The Netherlands). Anti-polyHIS was diluted 1/1000 and anti-mouse FITC was diluted 1/5 in 5% MPBS.
Incubation of samples with anti-rat FITC and anti-mouse FITC was done in the dark. After washing
with PBS, 1 drop of an anti-fade agent (Citifluor, Agar Scientific) was added to samples prior to
examination with the Olympus B-41 microscope (Olympus, Optical Co. Ltd) equipped with
epifluorescence illumination (X-CiteR Fluorescence Illumination, series 120Q, EXFO Europe).
Micrographs were taken using image analysis software (cell*, Soft Imaging System).

30
CHAPTER THREE
RESULTS AND DISCUSSION
PECTIN STRUCTURE-FUNCTION RELATIONSHIP IN CARROT PUREE
3.1. The influence of pretreatments, blending and high pressure homogenization on the
bostwick consistency of carrot purees.
The flow behavior of purees obtained from non-pretreated and pretreated carrot samples was
studied by using the Bostwick consistometer. Purees were prepared by either blending or blending
followed by high pressure homogenization (HPH) of the samples. The results (Figure 3.1) show a clear
difference in flow properties between purees prepared by blending and HPH. The separation
between the liquid fraction and the pulp (syneresis) is more pronounced in purees prepared by
blending when compared to purees prepared by HPH for all samples. Generally, purees prepared by
HPH are less consistent compared to those prepared by blending for all samples.
High temperature blanching of carrots prior to blending or HPH results in purees with almost no
syneresis, whereas low temperature blanched carrot samples results in purees with the highest
degree of separation between liquid and pulp after blending and HPH.

31
Figure 3.1. Bostwick consistency measurements for purees prepared by blending and HPH of
non-pretreated (Non.) and pretreated carrot samples. LTB = low temperature blanching, HTB = high
temperature blanching, HPH = high pressure homogenization (at 100 bar). Error bars indicate the
standard deviation of two independent measurements.
3.2. The influence of pretreatments, blending and high pressure homogenization on the
particle size distribution of carrot purees
To assess the particle size distribution of the different carrot purees analysis by wet sieving was
performed. The results obtained are presented in bar charts ( Figures 3.2 and 3.3).
In general, the tissue particles in carrot purees have a unimodal distribution both in blended and
HPH samples. The highest relative mass of the tissue particles is between 250 and 500 µm for
blended and HPH samples, but tissue particles between 500 and 1000 µm are more abundant in the
blended purees whereas the particles between 125 and 250µm are more abundant in the HPH puree
compared to the blended samples.
For the blended samples, both LTB and HTB result in an increased amount of tissue particles retained
in the sieve column (Figure 3.2). This means a decreased amount of material washed out from the
sieve column (MWO*) as compared to the non-pretreated sample. For the LTB sample, this seems to
occur alongside a notable increase in the amount of tissue particles retained on 500 µm sieve
0 5 10 15 20
Bostwick consistency (cm)
Pulp
Liquid fraction

32
whereas the HTB sample shows a small increase of tissue material on the 125 and 250 µm sieve. This
could means that LTB carrot tissues were more resistant to physical disintegration upon blending as
compared to non-pretreated and HTB tissue. In literature, LTB has been shown to result into
improved cell adhesion and hence tissue firming (Sila and others, 2004; Vu and others, 2004).
Figure 3.2 Relative mass (%) of tissue material of purees prepared by blending of the non-pretreated
and pretreated carrot samples retained on different sieves. LTB = low temperature blanching,
HTB = high temperature blanching. MWO*= material washed out of the sieve column calculated as:
MWO* = Weight of sieved sample – weight of tissue material retained in all sieves.
HPH also results in a remarkable increase in materials retained on 125 µm sieve (Figure 3.3) and a
decrease in material on the 500 µm sieve as compared to blended samples. After HPH, the relative
amount of material washed out of the sieve column (RWF*) increased sharply (from 34.4 to 40.8%
for non-pretreated, from 25 to 34.5% for LTB and from 21.3 to 36.6% for HTB). The tissue particles
contained in this fraction are smaller than 40 µm. These differences between blended and HPH
carrot purees can be expected as HPH has a more intense shear input compared to simple blending.
0
5
10
15
20
25
30
35
40
45
50
Re
lati
ve m
ass
(%)
Non. + blending
LTB + blending
HTB + blending
MWO* 40 80 125 250 500 1000
Sieve size (µm)

33
Figure 3.3. Relative mass (%) of tissue material of purees prepared by HPH of the non-pretreated and
pretreated carrot samples, retained on different sieves. LTB = low temperature blanching,
HTB = high temperature blanching, HPH = high pressure homogenization (at 100 bar).
MWO* = weight of sieved sample – weight of tissue material retained in all sieves.
3.3. Degree of esterification of alcohol insoluble residue
The degree of esterification (DE) of alcohol insoluble residue (AIR) from different carrot samples was
determined by analyzing its GalA content and the amount of methoxyl groups. In these analyses,
standard curves were prepared. Presented in Figures 3.4a and 3.4b are the standard curves for
methanol and GalA analyses respectively.
The standard curve for determining the amount of methoxyl groups (Figure 3.4a) is:
y = 0.046x + 0.0398 (R2 = 0.9993)
Where x = concentration of methoxyl groups (µg methanol/ml phosphate buffer)
Y = absorbance at 412nm and 25°C
0
5
10
15
20
25
30
35
40
45
Re
lati
ve m
ass
(%)
Non. + HPH
LTB + HPH
HTB + HPH
RWF* 40 80 125 250 500 1000
Sieve size (µm)
MWO* 40 80 125 250 500 1000
Sieve size (µm)

34
Figure 3.4 (a). Standard curve for the determination of the amount of methoxyl groups.
The standard curve for determining GalA content (Figure 3.4b) is :
y = 0.0137x + 0.0183 (R2 = 0.995)
Where x = concentration of galacturonic acid (µg/ml H2O)
Y = absorbance at 520nm and 25°C
Figure 3.4 (b). Standard curve for the determination of GalA content.
0
0.2
0.4
0.6
0.8
1
1.2
0 5 10 15 20 25
Ab
sorb
ance
Concentration (µg methanol/ml phosphate buffer)
0
0.2
0.4
0.6
0.8
1
1.2
1.4
1.6
0 20 40 60 80 100 120
Ab
sorb
ance
Concentration (µg galacturonic acic/ml water)

35
The degree of esterification (DE) of AIR obtained from non-pretreated and pretreated carrot samples
is shown in Table 3.1. For the non-pretreated carrot, the DE was estimated at 59.5% which is in close
agreement with the values reported for fresh carrot (DE = 60-69%) (Ng and Waldron, 1997; Sila and
others, 2006a). It is also observed that there is no remarkable difference in DE between non-
pretreated and HTB samples. However, from Table 3.1, it is observed that LTB showed a pronounced
reduction in DE compared to non-pretreated sample. This might be due to stimulation of carrot
PME. Sila and others (2004) also reported a decrease in DE of AIR obtained from carrot after LTB. Van
Buren (1979) also suggested that PME is activated at temperatures between 50 and 80°C. HPH
seemed not to affect the DE of non-pretreated and pretreated samples.
Table 3.1. Degree of esterification of AIR from non-pretreated and pretreated, blended and HPH
carrot puree samples. LTB = low temperature blanching, HTB = high temperature blanching,
HPH = high pressure homogenization (at 100 bar).
Sample DE (%) of AIR
Blended HPH (100 bar)
Raw 59.5 ± 2.3 a 62.4 ± 1.8
LTB 51.9 ± 2.3 52.5 ± 2.0
HTB 60.8 ± 3.5 60.8 ± 3.5
astandard deviation
3.4 Fractionation of AIR.
AIR of different carrot puree samples was fractionated in water soluble pectin (WSP), chelator
soluble pectin (CSP) and sodium carbonate soluble pectin (NSP).
The effect of pretreatment and blending/HPH on the relative amount of the WSP is presented in
Figure 3.5a. In general, HPH results in increased relative amounts of WSP for both non-pretreated
(raw) and pretreated carrot samples. This effect is more pronounced for the non-pretreated sample
where the proportion of WSP increased from 24.6 to 44.2% . For LTB, the relative amount of WSP
increased from 20.8 to 27.8%, whereas in the HTB sample, the increase is from 38.2 to 44.1%. Upon
comparing the three samples, LTB sample has the lowest relative amount of WSP while the HTB

36
sample has the highest. In literature it has been found that a decrease in DE after LTB lowers the
solubilization of pectin in water ( Sila and others, 2006).
Figure 3.5a: Relative amount of WSP fraction for blended and HPH carrot purees.
Non. = Non-pretreated, LTB = low temperature blanching, HTB = high temperature blanching,
HPH = high pressure homogenization (at 100 bar), WSP = water soluble pectin.
HPH results in a decrease of chelator soluble pectin (CSP) proportions in purees prepared from both
non-pretreated and LTB carrots (Figure 3.5b). The figure also shows that the proportion of CSP for
HTB remains almost unchanged after HPH. In LTB, blended samples and non-pretreated, blended
samples, the proportion of CSP is higher than in the other samples.
0
5
10
15
20
25
30
35
40
45
50
Non. LTB HTB
WSP
(%
)
blended
HPH

37
Figure 3.5b: Relative amount of CSP fraction for blended and HPH carrot purees.
Non.= Non-pretreated, LTB = low temperature blanching, HTB = high temperature blanching,
HPH = high pressure homogenization (at 100bar), CSP = chelator soluble pectin.
From Figure 3.5c, it is observed that HPH is associated with a decrease in relative amount of NSP for
all samples. In non-pretreated sample, the relative amount of NSP dropped from 16 to 9.3% after
HPH. The decrease in proportion of NSP for LTB sample is from 18 to 12.7% and for HTB the change is
from 9.6 to 6% after HPH.
With regard to pretreatments, it is remarkable that the HTB samples have the lowest relative amount
of NSP.
0
5
10
15
20
25
30
35
40
45
Non. LTB HTB
CSP
(%
)
blended
HPH

38
Figure 3.5c: Relative amount of NSP fraction for blended and HPH carrot purees.
Non. = Non-pretreated, LTB = low temperature blanching, HTB = high temperature blanching,
HPH = high pressure homogenization (at 100 bar), NSP = sodium carbonate soluble pectin.
These changes in proportions of the three pectin fractions in different samples are also presented in
Figure 3.5d. In this figure, additional information on the amount of pectin which was not extracted by
the three solvents is given. Thus, the proportion of pectin lost during fractionation can be estimated.
As shown in Figure 3.5d, the fractionation yield in all samples amounted to above 70% including the
galacturonic acid recovered from the residue. LTB samples have the highest proportion of pectin in
the residue.
0
2
4
6
8
10
12
14
16
18
20
Non. LTB HTB
NSP
(%
)
blended
HPH

39
Figure 3.5d. Relative amounts of GalA in different pectin fraction and fractionation yield for different
carrot samples. Non. = non-pretreated, LTB = low temperature blanching, HTB = high temperature
blanching, HPH = high pressure homogenization (at 100 bar), WSP = water soluble pectin, CSP =
chelator soluble pectin, NSP = sodium carbonate soluble pectin.
3.5 Degree of esterification of water and chelator soluble pectin fractions.
Differences in the DE of the pectin fractions were observed (Table 3.2). Water soluble pectin was
highly esterified with a DE in the range 68.8 – 77.9%. The CSP was lowly esterified with a DE in the
range 38.7 – 48.1%. The DE of the NSP fraction could not be determined because sodium carbonate,
used during fractionation, hydrolyses all the methyl esters.
0102030405060708090
Re
lati
ve a
mo
un
t (%
)
Residue
NSP
CSP
WSP

40
Table 3.2. Degree of esterification of WSP and CSP fractions from non-pretreated (Non.) and
pretreated carrot samples. LTB = low temperature blanching, HTB = high temperature blanching,
HPH = high pressure homogenization (at 100 bar), WSP = water soluble pectin, CSP = chelator soluble
pectin.
Sample DE of WSP (%) DE of CSP (%)
Non.+ blending 74.4 ± 2.5 a 48.1 ± 0.9 LTB + blending 70.4 ± 1.6 38.7 ± 1.6 HTB + blending 70.2 ± 3.9 39.0 ± 1.5 Non.+ HPH 68.8 ± 1.4 46.2 ± 4.0 LTB + HPH 70.4 ± 2.8 44.4 ± 1.2 HTB + HPH 77.9 ± 2.8 39.0 ± 3.4
aStandard deviation
3.6 Immuno-dot assays
Pectin structure was further assessed by immuno-dot-assays using anti-HG antibodies with particular
binding specificities. Figure 3.6 shows the binding of anti-HG antibodies JIM5, JIM7, LM18, LM19,
LM20 and PAM1 to water soluble pectin (WSP) and chelator soluble pectin (CSP) of different carrot
samples. From the figure, it can be seen that all the antibodies bind to the WSP fraction of the non-
pretreated and LTB carrot purees. This means that, although the WSP fraction has a relatively high DE
(68.8-77.9%), it contains pectins with a low, middle and high DE. To the WSP fractions of HTB carrot
purees, JIM7 binds intensely, whereas the antibodies LM19 and LM20 bind to a lesser extent and the
antibodies JIM5 and PAM1 only minimally bind. This could mean that the distribution of DE in HTB
carrot samples is much smaller compared to non-pretreated or LTB samples. It is also noticed from
the immune-dot results that, binding of the WSP of all samples to JIM5, LM18, LM19 and PAM1 is
limited to the central dots while JIM7 and LM20 bind to the central dot as well as to an outer ring
material. This means that, the WSP is rich in pectic components with abundant side chains and in
more linear pectin components that are highly esterified.
In the immune-dot assay with CSP, it can be seen that the CSP of all samples binds to all antibodies
used during the assay. However, the relative high binding with antibodies LM18, LM19 and PAM1
and the relative low binding with LM20, suggest that the CSP fraction is relatively low esterified. For
the CSP fraction, binding is not only limited to the central dot, indicating that the CSP fraction of

41
carrot puree contains pectic components with abundant side chains as well as linear pectic
components.
Non
. x b
lend
ed
LTB
x b
lend
ed
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
HT
B x
ble
nded
Non
. x H
PH
LTB
x H
PH
HT
B x
HP
H
Carrot CSP
Non
. x b
lend
ed
LTB
x b
lend
ed
HT
B x
ble
nded
Non
. x H
PH
LTB
x H
PH
HT
B x
HP
H
Carrot WSP
JIM5
JIM7
LM18
LM19
LM20
PAM1
Figure 3.6. Immuno-dot-assays of JIM5, JIM7, LM18, LM19, LM20, and PAM1 binding to WSP and
CSP of non-pretreated (Non.) and pretreated carrot samples. Samples were applied in dilution series
at levels indicated. LTB = low temperature blanching, HTB = high temperature blanching, HPH = high
pressure homogenization (at 100 bar),

42
3.7. Immunolabeling of non-pretreated and pretreated carrot tissue particles.
Anti-HG antibodies with different binding specificities were used to localize pectin in carrot tissue
particles. The particles were obtained from wet sieving of non-pretreated and pretreated carrot
purees. Figure 3.7a shows the immunofluorescence labeling of carrot tissue particles retained on
80 and 125µm sieves with antibodies JIM5, JIM7 and LM20.
From the figure, it is observed that in all samples, cells appear in clusters. Both JIM5 and JIM7 labeled
pectin and their epitopes seem to be uniformly distributed on cell walls. In general, there are no
remarkable differences among samples, although non-pretreated is less intensely labeled with JIM5
and HTB is less intensely labeled with JIM7 . On the other hand, LM19 displays a dotted binding
pattern in the HTB samples while showing higher labeling intensity for LTB sample. This can indicate
that the LTB sample contains more lowly esterified pectin epitopes, accessible to LM19 than the non-
pretreated and the HTB samples.
Figure 3.7a. Representative pictures resulting from immunofluorescence labeling of non-pretreated,
low temperature blanched (LTB), high temperature blanched (HTB) carrot tissue particles with anti-
HG antibodies JIM5, JIM7 and LM19. Tissue Particles retained on 80 and 125µm sieves were used.
Scale bars=100µm.

43
Figure 3.7b shows the immunfluorescence labeling of carrot tissue particles with antibodies LM20,
PAM1 and 2F4. The antibody LM20 shows higher labeling intensity in non-pretreated and HTB
samples than in the LTB sample indicating the presence of high esterified LM20 epitopes. PAM1
shows relatively high binding intensity in all samples. Interesting with PAM1 is that its epitopes
appear to be concentrated in specific locations of cell walls (the cell junctions). This is most clearly
visible in LTB sample and might indicate that process-induced de-esterification is very pronounced in
the cell junctions. 2F4, an antibody directed towards Ca+2-cross-linked pectins, display faint labeling
in all samples indicating that the level of Ca-pectin cross-link in the carrot samples is low.
Figure 3.7b. Immunofluorescence labeling of non-pretreated, low temperature blanched (LTB), high
temperature blanched (HTB) carrot tissue particles with anti-HG antibodies LM20, PAM1 and 2F4.
LTB = low temperature blanching, HTB = high temperature blanching. Scale bars =100µm.
2F4
Raw
LM20
PAM1
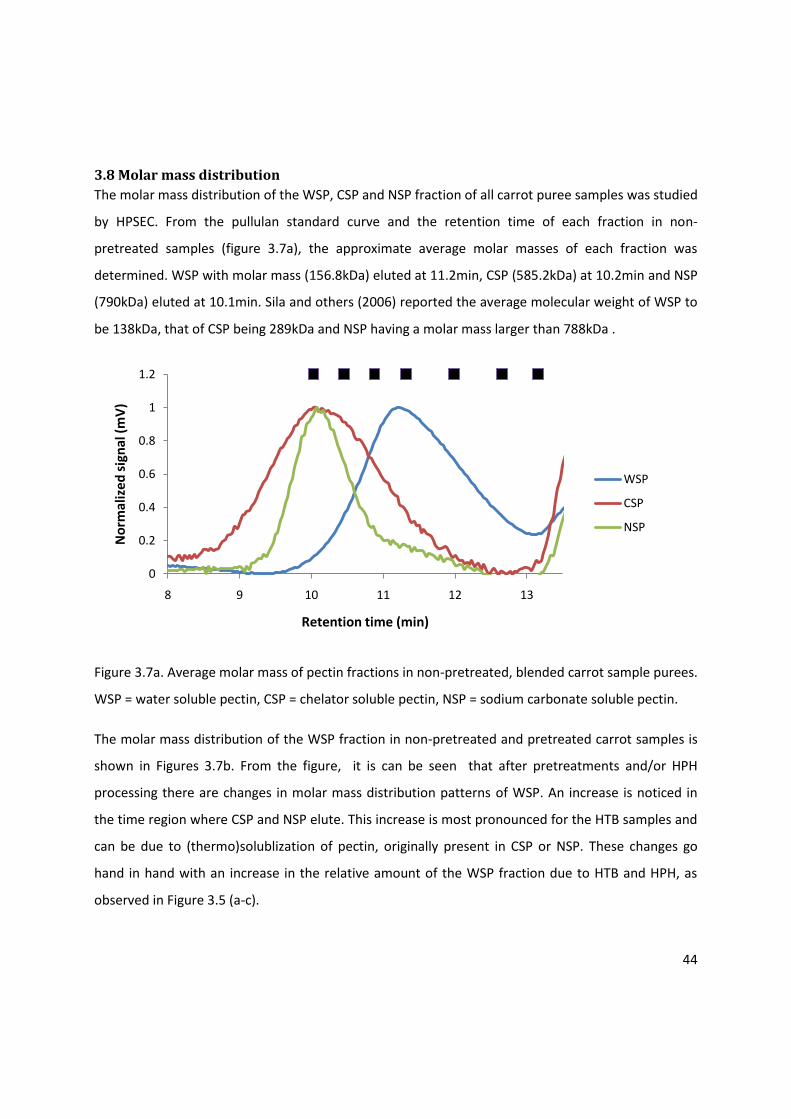
44
3.8 Molar mass distribution
The molar mass distribution of the WSP, CSP and NSP fraction of all carrot puree samples was studied
by HPSEC. From the pullulan standard curve and the retention time of each fraction in non-
pretreated samples (figure 3.7a), the approximate average molar masses of each fraction was
determined. WSP with molar mass (156.8kDa) eluted at 11.2min, CSP (585.2kDa) at 10.2min and NSP
(790kDa) eluted at 10.1min. Sila and others (2006) reported the average molecular weight of WSP to
be 138kDa, that of CSP being 289kDa and NSP having a molar mass larger than 788kDa .
Figure 3.7a. Average molar mass of pectin fractions in non-pretreated, blended carrot sample purees.
WSP = water soluble pectin, CSP = chelator soluble pectin, NSP = sodium carbonate soluble pectin.
The molar mass distribution of the WSP fraction in non-pretreated and pretreated carrot samples is
shown in Figures 3.7b. From the figure, it is can be seen that after pretreatments and/or HPH
processing there are changes in molar mass distribution patterns of WSP. An increase is noticed in
the time region where CSP and NSP elute. This increase is most pronounced for the HTB samples and
can be due to (thermo)solublization of pectin, originally present in CSP or NSP. These changes go
hand in hand with an increase in the relative amount of the WSP fraction due to HTB and HPH, as
observed in Figure 3.5 (a-c).
0
0.2
0.4
0.6
0.8
1
1.2
8 9 10 11 12 13
No
rmal
ize
d s
ign
al (
mV
)
Retention time (min)
WSP
CSP
NSP
Raw
LM20
PAM1
1

45
In addition, it can be noticed that the main WSP peak shifts to the right upon pretreating (HTB or
HPH) of the carrot puree. This could mean that, pectins in the WSP fraction have a lower molar mass
compared to the non-pretreated, blended puree. This depolymerization effect is most pronounced
for the HTB samples. Heat-induced solubilization and depolymerization of carrot pectin has been
reported also by Sila and others, (2005; 2006).
Figure 3.7b. Molar mass distribution of water soluble pectin in non-pretreated and pretreated carrot
samples. LTB = low temperature blanching, HTB = high temperature blanching, HPH = high pressure
homogenization (at 100bar).
There were no noticeable changes in molar mass distribution for the NSP and CSP fractions of all
carrot puree samples.
3.9 Conclusion
Based on the results of the present work, HPH was shown to decrease the consistency and the
syneresis of carrot purees. On the other hand, it was also observed that the HPH carrot purees
contained smaller tissue particles and high amount of WSP compared to the respective blended
carrot puree samples.
0
0.2
0.4
0.6
0.8
1
1.2
8 9 10 11 12 13
No
rmal
ize
d s
ign
al (
mV
)
Retention time (minutes)
Non. + blending
LTB + blending
HTB + blending
Non.+ HPH
LTB + HPH
HTB + HPH

46
A low temperature blanching step resulted in blended and HPH carrot purees with higher consistency
and more syneresis as compared to the non-pretreated blended and HPH samples. In the low
temperature blanched samples, it was also noticed that tissue particles were larger and pectin was
less water-soluble when compared to non-pretreated samples purees. The high CSP of low
temperature blanched samples indicates a high probability for ionically cross-linked pectin, which in
turn might be related to a better cell-cell adhesion in the carrot tissue and thus the high resistance of
this tissue to physical disintegration upon blending/homogenization.
A high temperature blanching treatment prior to blending/homogenization step resulted in carrot
purees samples in which no syneresis occurred. In these samples, a high proportion of pectin was
water-soluble, which is probably the consequence of pectin solubilization during pretreatment. The
effect of high temperature blanching on the consistency and the tissue particle size distribution were
not clear-cut.
In summary, it can be stated that pectin engineering seems to be a helpful tool to control the
consistency and syneresis of carrot purees. Hereby, (pre)treatments that solubilize pectin, such as
HPH and in particular high temperature blanching, can be used to limit syneresis phenomena. The
consistency of carrot puree seems to be correlated with the particle size distribution of the puree.
Hereby, low temperature blanching can be used as a tool to increase the cell-cell adhesion, the
resistance to tissue particle disintegration and thus the tissue particle size. HPH on the other hand,
has been shown to systematically reduce the particle size of carrot puree tissue particles compared
to simple blending.

47
PECTIN STRUCTURE-FUNCTION RELATIONSHIP IN TOMATO PUREE
4.0. Selection of pretreatments for tomato samples.
To understand the pectin structure-function relationship in tomato puree, non-pretreated, high
temperature blanched (HTB) and high pressure (HP) pretreated tomato samples were used to make
puree. The purees were prepared by blending or blending followed by high pressure homogenization
(HPH). Based on available literature, the conditions for the HTB pretreatment were selected to
inactivate tomato PME and PG, so that pectin molecules remain unchanged during subsequent
thawing, blending/HPH of the HTB pretreated samples. On the other hand, HP pretreatment aimed
at selective inactivation of tomato PG .
4.1 The influence of pretreatments and blending versus high-pressure homogenization on
the bostwick consistency of tomato purees
The consistency of tomato purees of non-pretreated and pretreated samples was measured by using
the bostwick consistometer. The results show differences in flow properties of purees prepared by
blending and high pressure homogenization (HPH) of non-pretreated and pretreated tomato
samples. Almost no separation between the liquid fraction and the pulp (syneresis) in tomato purees
of all samples was observed (Figure 3.8). HTB and HP pretreated samples have higher consistency
than the non-pretreated sample after blending.

48
Figure 3.8. Bostwick consistency of purees prepared by blending and HPH of non-pretreated (Non.)
and pretreated tomato samples. HTB = high temperature blanching, HP = high pressure pretreatment
(10 min at 550 MPa, 25°C), HPH = high pressure homogenization (at 100 bar).
HPH results in a remarkable increase in consistency for non-pretreated sample compared to
blending. This observation was also reported in previous study ( Colle and others, 2010 ). They found
that increasing homogenization pressure resulted in a the tomato pulp with a stronger fiber network
and increased viscosity.
The low consistency of non-pretreated, blended puree, might be explained by the fact that tomato
PME and PG can be active in this sample. As the sample did not undergo any enzyme-inactivation
step prior to thawing and blending, both PME and PG can alter tomato pectin molecules. During the
thawing and blending process, the tomato tissue is moreover disintegrated, enhancing the contact
between substrate and the enzymes. This could mean that pectin molecules can be depolymerized
by the action of PG, that can be even stimulated by low DE of pectin as a result of PME activity.
On the other hand, HTB conditions should inactivate both PME and PG (Rodrigo and others, 2006).
Apparently, this inactivation goes hand in hand with a high consistency puree.
0 5 10 15 20 25
Bostwick consistency (cm)
pulp
Liquid fraction

49
HP pretreatment of tomato resulted into puree with the same consistency as the HTB pretreated
sample. This result is consistent with the result of Krebbers and others (2003) who also noticed that
by increasing the pressure level ( in range 300 to 700Mpa) at 20°C during processing of tomato
puree, the viscosity of the puree increased. The HP pretreatment of tomato was done for 10min at
550MPa, 25°C, conditions to selectively inactivate PG while retaining PME activity (Rodrigo and
others, 2006). On the other hand, disruptions of cell wall and membrane during high pressure
processing (Van Bugghenhout and others, 2005) is accompanied by release of cell wall bound PME
and can thus enhance the contact between enzyme and its substrate. The high consistency of puree
from HP pretreated, blended tomato sample might be related to the fact that during HP
pretreatment, PG was inactivated while PME remained active. This might implicate that pectin
molecules can be demethoxylated and could be less prone to enzymatic depolymerization.
4.2.The influence of pretreatment and blending versus high-pressure homogenization on
the particle size distribution of tomato purees
In general, the tomato tissue particles in both blended and HPH samples have a unimodal
distribution. The highest relative amount of tissue particles for blended samples is retained on the
125 and 250µm sieves (Figure 3.9a). From the figure, it can also be seen that after blending of HTB
and HP pretreated tomato samples, the resulting purees are characterized by a decrease in relative
amount of materials washed out of the sieve column (MWO*) compared to the non-pretreated
sample. This was accompanied by a pronounced increase in tissue particles retained on 125 and
250µm sieves. This might be due to PG activity in non-pretreated tomato sample which resulted in
pectin depolymerization and hence cells were easily disrupted upon blending compared to HTB and
HP samples. Small changes in tissue particles on other sieves were also observed.

50
HPH resulted in a dramatic increase in relative amount of puree materials washed out of the sieve
column (MWO*) for the HTB and HP treated samples (Figure 3.9b). The tissue particles retained in
this fraction are less than 40µm in size and accounted for at least 50% of the weight of puree sieved
for all samples. This is expected due to particle size reduction during HPH. For the non-pretreated
sample, the relative amount MWO* is high, but this was also the case after blending.
0
10
20
30
40
50
60
Re
lati
ve m
ass
(%)
Non.+ blending
HTB + blending
HP + blending
MWO* 40 80 125 250 500 1000
Sieve size (µm)
Figure 3.9a. Relative mass of tissue particles in tomato purees prepared by blending of
non-pretreated (Non.) and pretreated tomato samples. HP = high pressure pretreatment (10 min at
550MPa, 25°C ), HTB = high temperature blanching. MWO* is calculated as:
MWO* = weight of sieved sample – weight of tissue particles retained in all sieves.

51
Figure 3.9b. Relative mass of tissue particles in tomato purees prepared by blending followed by high
pressure homogenization (at 100bar) (HPH) of non-pretreated (Non.) and pretreated tomato
samples. HP = high pressure pretreatment (10 min at 550MPa, 25°C ), HTB = high temperature
blanching. MWO* = weight of sieved sample – weight of tissue particles retained in all sieves.
4.3. Degree of esterification of alcohol insoluble residue
The degree of esterification (DE) of AIR obtained from non-pretreated and pretreated tomato
samples is shown in Table 3.3. It can be seen that there is no shift in DE as a result of HPH. The DE of
non-pretreated sample was estimated at 22.4%, which is lower than that of HTB sample. This low
value of DE suggests that PME was active during thawing and subsequent blending/HPH of the
non-pretreated tomato sample.
Furthermore, during thawing and blending/HPH PME is brought in contact with its substrate as a
result of tissue damage. The HTB tomato sample have a medium-high average DE (67.5%). As the
enzymes, and thus PME, in these samples should be inactivated prior to freezing, this value should
also represent the average DE of pectin in the intact tomato fruits. On the other hand, HP
pretreatment of tomato samples resulted into a significant decrease in DE. This might also be due the
activity of tomato PME as well as possibility for its stimulation during HP.
0
10
20
30
40
50
60
70
Re
lati
ve m
ass
(%
)
Non. + HPH
HTB + HPH
HP + HPH
MWO* 40 80 125 250 500 1000
Sieve size (µm)

52
Table 3.3. Degree of esterification (DE) of AIR from non-pretreated and pretreated, blended and HPH
tomato samples. HTB = high temperature blanching, HP = high pressure pretreatment (10min at
550MPa, 25°C), HPH = high pressure homogenization (at 100bar).
Sample DE of AIR (%)
Blended HPH
Non-pretreated 22.4 ± 1.8a 22.9 ± 3.1
HTB 67.5 ± 3.0 63.7 ± 2.4
HP 15.9 ± 1.1 23.8 ± 1.1
astandard deviation
4.4. Fractionation of AIR
AIR of non-pretreated and pretreated tomato puree was fractionated in water soluble pectin (WSP),
chelator soluble pectin (CSP) and sodium carbonate soluble pectin (NSP).
The effects of pretreatment and blending/HPH on the relative amount of pectin fractions and residue
are presented in Table 3.4. HPH results in a small increase in relative amount of WSP in HTB and HP
treated tomato samples. A small decrease in relative amount of WSP due to HPH is observed for
non-pretreated tomato sample. Blended, HP treated tomato samples have a remarkable lower
relative amount of WSP compared to blended, non-pretreated samples.

53
Table 3.4: Relative amount of WSP, CSP, NSP and residue in non-pretreated (Non.) and pretreated,
blended and HPH tomato purees. HTB =high temperature blanching, HP = high pressure
pretreatment (10min at 550MPa, 25°C), HPH = high pressure homogenization (at 100bar), WSP =
water soluble pectin, CSP = chelator soluble pectin, NSP = sodium carbonate soluble pectin.
Sample WSP (%) CSP (%) NSP (%) Residue (%)
Non.+ blending 27.7 ± 0.6a 24.2 ± 0.8 6.1 ± 0.8 4.7 ± 0.3
HTB + blending 22.8 ± 1.2 10.4 ± 0.7 19.8 ± 1.0 7.7 ± 0.2
HP + blending 12.5 ± 0.6 30.2 ± 3.4 12.0 ± 0.9 10.6 ± 0.6
Non.+ HPH 26.2 ± 0.7 30.4 ± 0.9 5.5 ± 0.5 6.3 ± 0.6
HTB + HPH 26.9 ± 0.8 21.4 ± 0.6 14.3 ± 1.1 7.8 ± 0.3
HP + HPH 15.9 ± 0.4 31.4 ± 2.0 13.1± 1.1 9.9 ± 0.9
astandard deviation
From the table, it can also be seen that HPH results in a remarkable increase of chelator soluble
pectin (CSP) proportions in non-pretreated and HTB tomato samples. After HPH, the relative amount
of CSP in HTB samples increased from 10.4 to 21.4% while in non-pretreated tomato the increase
was from 24.2 to 30.4%. For the HP sample, the increase was small (from 30.4 to 31.4%) as HP
treated, blended samples already contain a high proportion of CSP. HTB sample had the lowest
relative amount of CSP.
HPH also resulted in decreased relative amount of NSP in both non-pretreated and HTB tomato
samples. However, HPH resulted in an increase in relative amount of NSP from 12 to 13.1% in the HP
treated tomato sample. Among all tomato samples, non-pretreated, HPH had the smallest relative
amount of NSP whereas the HTB, blended sample had the highest relative amount of NSP.
The total relative amount of galacturonic acid recovered in the three pectin fractions (fractionation
yield) is presented in Figure 4. This figure gives an overview of the different pectin fractions in the
different samples. The HTB samples (in which the enzymes PME and PG are presumably completely
inactivated) in general contain a high proportion of NSP and a relatively low proportion of CSP.
Compared to HTB samples, both the non-pretreated and the HP treated sample contain a high

54
portion of CSP. For the non-pretreated samples, this is accompanied by a decrease in NSP and by a
small increase in WSP. For the HP treated samples, this is accompanied by a decrease in NSP as well
as a decrease in WSP. Also in this figure, the relative amount of un-recovered galacturonic acid
(residue) is presented. The fractionation yield in all tomato samples was between 55 and 62%. The
highest relative amount of residue was 10.6%, found in the HP sample.
Figure 4. Fractionation yield and un-extracted pectin (residue) of different tomato samples.
Non.= Non-pretreated, HTB = high temperature blanching, HP = high pressure pretreatment (10 min
at 550 MPa, 25°C), HPH = high pressure homogenization (at 100 bar), WSP = Water soluble pectin,
NSP = sodium carbonate soluble pectin, CSP = Chelator soluble pectin.
4.5. Degree of esterification of water and chelator soluble pectin
Table 3.4 shows the DE of WSP and CSP in non-pretreated and pretreated tomato samples. The
pectin molecules in WSP and CSP of the HTB samples are highly esterified while those in WSP and
CSP of non- pretreated and HP treated samples are lowly esterified. The DE of WSP of the HTB,
blended is almost the same as that of its CSP while the DE of the HTB, HPH sample is however
lowered to 49.6%. It is however remarkable that the DE of WSP of the non-pretreated sample is
lower than the DE of WSP of the HP treated sample whereas for CSP, the DE is lower for the HP
treated sample.
01020304050607080
Pe
rcen
t (%
)
Residue
NSP
CSP
WSP

55
Table 3.5. Degree of esterification (DE) of WSP and CSP from non-pretreated and pretreated tomato
samples. HTB = high temperature blanching, HP = high pressure pretreatment (10min at 550MPa,
25°C), HPH = high pressure homogenization (at 100bar), WSP = water soluble pectin, CSP = chelator
soluble pectin.
Sample DE of WSP (%) DE of CSP (%)
Non-pretreated + blending 17.3 ± 0.4 a 18.1 ± 1.2 HTB + blending 78.6 ± 4.0 80.6 ±5.6 HP + blending 24.6 ± 1.2 12.9 ± 1.4 Non-pretreated + HPH 15.8 ± 1.2 17.1 ± 0.6 HTB + HPH 79.0 ± 2.9 49.6 ± 1.4 HP + HPH 22.9 ± 0.9 12.6 ± 0.9
astandard deviation
4.6. Immno-dot assays
Figure 4.1 shows the binding of anti-HG antibodies JIM5, JIM7, LM18, LM19, LM20 and PAM1 to
water soluble pectin (WSP) and chelator soluble pectin (CSP) of different tomato samples. From the
figure, it is observed that CSP binds to all antibodies with JIM7 displaying the highest binding
strength. This shows that there is very broad distribution in DE among the pectin molecules in CSP
fraction including the pectins with an average DE as low as 12-13%. LM18, LM19 and PAM1 show
more intense binding to CSP of non-pretreated and HP treated tomato samples compared to the HTB
samples. This correlates with the higher DE of the HTB samples.
Also the WSP of all tomato samples display binding to all antibodies. This observation suggest that
WSP has a broad distribution in DE. The binding of JIM7 to WSP of all tomato samples is more intense
compared to other antibodies. PAM1 shows high binding intensity for non-pretreated, blended
samples, HP, blended samples and HP, HPH samples correlating to the low DE of these samples. The
binding of LM18, LM19, LM20 to is relatively low compared to their binding to CSP. Maybe their
epitopes in this sample were not accessible or possibly a factor necessary for their binding is not
present.
In general it was observed that the high viscosity of samples, especially the highly concentrated
samples result in practical limitation as the pectin material in such samples did not stick very well to
the nitrocellulose membrane.

56
Tomato WSP
Raw
x b
lend
ed
HT
B x
ble
nded
HP
x bl
ende
d
Raw
x H
PH
HT
B x
HP
H
HP
x H
PH
Raw
x b
lend
ed
HT
B x
ble
nded
HP
x bl
ende
d
Raw
x H
PH
Tomato CSP
HT
B x
HP
H
HP
x H
PH
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
5 µg
0.5 µg
50 ng
5 ng
JIM5
JIM7
LM18
LM19
LM20
PAM1
Figure 5.1. Immuno-dot-assays of JIM5, JIM7, LM18, LM19, LM20, and PAM1 binding to WSP and
CSP of non-pretreated (raw) and pretreated tomato samples. Samples were applied in dilution series
at levels indicated. HPH = high pressure homogenization (at 100bar), HP = high pressure
pretreatment (10min at 550MPa, 25°C), HTB = high temperature blanching.
4.7. Immunolabeling of non-pretreated and pretreated carrot tissue particles
In Figure 5.2, different pectin structures are localized in tomato tissue particles retained on the 125
µm sieve. Both JIM5 and JIM7 intensely label the HTB and HP treated tomato particles. Hereby, the
epitopes of JIM7 seem to be homogeneously distributes on the cell walls in both HTB and HP
pretreated tomato samples. Sometimes, a ‘spotty’ labeling pattern is however found, indicating a
local heterogeneity in particular pectin structures. Both JIM5 and JIM7 show a weak binding to
non-pretreated tomato samples which might be related to pectin solubilization in these samples.
JIM5 labeling in non-pretreated sample is concentrated on the cell junctions.

57
All tomato puree samples show (very)poor binding to the anti-HG antibodies LM19 (Figure 5.2) and
LM20 (Figure 5.3), except the HP treated sample which is labeled by LM19. The LM19 epitopes seem
to be uniformly distributed on the cell wall. The immunofluorescence experiments with LM18 did not
give positive results. Apparently, the pectin epitopes of the antibodies LM18,LM19 and LM20 are not
(abundantly) present or not (easy) accessible in the different tomato samples.
Figure 5.2. Representative pictures resulting from immunofluorescence labeling of non-pretreated,
high temperature blanched (HTB), high pressure pretreated(HP) tomato tissue with anti-HG
antibodies JIM5, JIM7 and LM19. Tissue particles retained on 80 and 125 µm sieves were used. Scale
bars =100µm.
From Figure 5.3, it can also be seen PAM1 displays intense binding to all tomato samples. Its
epitopes, lowly esterified pectins, seem to be distributed very heterogeneously in the HTB sample. In
both the non-pretreated and HP treated samples however, a homogeneous labeling pattern can be

58
noticed. These pictures suggest that PME in the intact tomato fruits can locally act on pectin, while
PME in thawed, blended samples demethoxylates the pectin all over the tomato cell walls.
The antibody 2F4 shows weak binding intensity to the HTB sample and shows more intense similar
binding pattern to the non-pretreated and HP treated samples. Its epitopes seem to be concentrated
in certain parts of cell wall, in particular the cell junctions. Pectin-calcium cross-link seem to be
present in the intact tomato fruits used to make purees. However, their presence can be increased
by PME activity duding thawing and subsequent blending in non-pretreated and HP treated samples.
Figure 5.3. Representative pictures resulting from immunofluorescence labeling of non-pretreated,
high temperature blanched (HTB), high pressure pretreated(HP) tomato tissue with anti-HG
antibodies JIM5, JIM7 and LM19. 2F4. 80 µm and 125 µm fractions were used. Scale bars =100µm.

59
4.8. Molar mass distribution
The molar mass distribution of the WSP of all tomato puree samples was studied by HPSEC. The
pullulan standards were used to estimate the average molar masses of the water soluble pectin
fraction. Figure 6 shows the molar mass distribution of WSP fraction in non-pretreated and
pretreated tomato samples. The WSP fraction of HTB samples have a molar mass corresponding to
retention time of 10.4 min. From the figure it can be seen that the peak for WSP of non-pretreated
and HP treated samples shift to higher retention times. For the non-pretreated sample, this shift in
molar mass could be related to the depolymerizing action of PG.
Figure 6. In situ changes in molecular weight distribution of water soluble pectin in non-pretreated
(Non.) and pretreated tomato samples. HTB = high temperature blanching, HP = high pressure
pretreatment (10min at 550MPa, 25°C), HPH = high pressure homogenization (at 100bar),
4.9. Conclusion
All tomato samples purees show almost no syneresis. The consistency of the tomato samples is
however dependent on the pretreatments and how the puree is prepared (blending or HPH).
0
0.2
0.4
0.6
0.8
1
1.2
8 9 10 11 12 13
No
rmal
ize
d s
ign
al (
mV
)
Retention time (minutes)
Non. + blending
HTB + blending
HP + blending
Non. + HPH
HTB + HPH
HP + HPH

60
The non-pretreated, blended tomato sample puree has the lowest consistency compared to
pretreated tomato samples. The low consistency of non-pretreated blended puree, might be
explained by the fact that in this sample the two enzymes (PME and PG) may be active. The low DE of
AIR, WSP and CSP and the low molar mass of WSP of the non-pretreated sample, indicate that both
PME and PG induced pectin changes during thawing and subsequent blending of the samples.
Immunolabeling experiments using the antibodies, PAM1 and 2F4, moreover clearly showed a
homogeneous, abundant presence of low esterified pectin and of pectin-calcium cross-links in the
non-pretreated samples, whereas these pectin structures only locally appeared in the cell walls of
the intact tomato fruit (it is the HTB tomato sample). The less intense labeling pattern of the non-
pretreated samples with JIM5 and JIM7, when compared to other tomato samples, could be probably
linked to pectin solubilization.
HTB conditions were selected to inactivate both PME and PG in tomato. Apparently this resulted in a
high DE of AIR, WSP and CSP, a molar mass of WSP around 404kDa, a relatively high amount of NSP
and a high consistency. Hence, the pretreatment can be used to improve the consistency of tomato
purees prepared by blending.
HP pretreatment of tomato resulted in blended puree with the same consistency as the HTB, blended
puree. HP pretreatment condition was selected to selectively inactivate tomato PG. The
pretreatment (followed by blending) resulted in relatively high amount of CSP, a low DE of AIR, WSP
and CSP and an average molar mass of 112kDa. The homogeneous presence of low esterified pectins
and pectin-calcium cross-links in this tomato puree sample could be visualized during
immunolabeling experiments using the antibodies PAM1 and 2F4. It is therefore possible to improve
the consistency of blended tomato puree by either HTB or HP pretreatments. However, although it
was not part of this study, the appearance (color) of the purees from the two pretreatments were
visually different.
Preparation of puree by HPH results in a remarkable increase in consistency for non-pretreated
tomato samples. Meanwhile, there is no change in particle size distribution, no change in DE, no
change in relative amounts of WSP, CSP and NSP and no change in molar mass. This suggest that the

61
high consistency of non-pretreated tomato sample might be due to factors other than pectin
properties measured in this study and particle size.
However, when applied to HTB and HP pretreated tomato samples, HPH results in pronounced
increase in relative amounts of small tissue particles washed out of the sieve column (MWO*), and a
lower consistency purees compared to HTB and HP pretreated, blended samples. This effect is more
pronounced in HTB tomato samples where it is also accompanied by an increase in CSP and WSP.
GENERAL CONCLUSIONS Puree prepared by blending non-pretreated carrot pieces showed a high consistency but also a high
separation between the liquid and particle fraction. This puree was rich in particles smaller than 40
µm and between 125 and 1000 µm and contained pectin with an average DE of 59.5%. The ratios of
water soluble pectin (WSP), chelator soluble pectin (CSP) and sodium carbonate soluble pectin (NSP)
were respectively 25%, 35% and 16% of the AIR and the molecular mass of the WSP, CSP and NSP
fraction was estimated respectively around 156.8kDa, 585.2kDa and 790kDa%.
Carrot tissues subjected to LTB resulted in purees with a high consistency and more syneresis and
larger particles compared to the non-pretreated carrot puree sample. This puree contained pectin
with a low average DE (51.9%) and a lower amount of WSP and a higher amount of CSP and NSP than
the non-pretreated samples.
HTB pretreatment, which should inactivate PME, followed by blending resulted in a puree containing
pectin with high average DE (60.8%), a high relative amount of WSP, accompanied by relatively low
amounts of CSP and NSP and larger particles than the puree prepared from the non-pretreated
carrots. Thermosolubilization and depolymerization of the WSP fraction of this puree was clearly
demonstrated by HPSEC. These puree properties could be linked to a low degree of syneresis.
Puree preparation by HPH was found to systematically reduce the size of the carrot puree tissue
particles and to, increase the pectin solubility in water. This resulted in low consistency purees in
which syneresis phenomena are limited compared to the respective blended samples. HPH however
did not cause remarkable changes in the DE of pectin or in molecular mass distribution of the WSP
fraction.

62
The anti-pectin antibodies JIM5, JIM7, LM19, LM20, PAM1 and 2F4 mapped different pectin
structures in the carrot tissue particles and demonstrated that pectin in all the puree samples have a
broad distribution in their DE.
The puree prepared by blending non-pretreated tomato samples was found to have a low DE of AIR
(22,4%), WSP (17.3 %) and CSP (18.1 %) and a low estimation of the molecular mass of WSP
(47.3kDa). The pectin properties of the non-pretreated tomato puree differed noticeably from the
pectin properties of the HTB puree samples, which presumably is a consequence of PME and PG
activity. The resulting puree had a very low consistency and small tissue particles when compared to
the pretreated purees.
Blending the HTB treated sample resulted in a tomato puree containing pectin with an average
medium-high DE of AIR (67.5 %) and a high DE of WSP (78.6 %) and CSP (80.6 %). The molecular mass
of WSP was estimated at 404kDa, which is considerably higher than the molecular mass of the WSP
of the non-pretreated puree. The sample also contained a remarkable high portion of NSP. The puree
with the above-mentioned pectin characteristics showed a high consistency.
Blending the HP treated tomato sample resulted in the same consistency as the HTB treated, blended
sample. However, the pectin properties of this sample were different: pectin had a low average DE of
15.9 %, a relative low portion of WSP and a relative high portion of CSP.
HPH resulted in a remarkable increase in consistency in case of non-pretreated tomatoes were used
to prepare the puree. However, for this sample, HPH did not result in remarkable changes in tissue
particle sizes and pectin properties measured in this study. Factors other than those studied in the
current study might influence the puree consistency of non-pretreated, HPH tomato samples.
Remarkably, almost no syneresis was observed for all the tomato purees which clearly shows that
tomato and carrot purees are two completely different matrices.
For the tomato samples, the anti-pectin antibodies were shown very useful to visualize the
homogenous distribution of medium and high esterified pectin structures and the local presence of
low esterified pectin and pectin-calcium cross-links in HTB samples. In non-pretreated and HP treated
samples, a completely different labeling pattern was observed, clearly showing the result of
enzyme-induced pectin changes.

63
Based on the results of the present work, it can be stated that (pre)treatments aiming at particular
(enzyme-induced) pectin changes influence the flow behavior of carrot and tomato purees. This case
study thus shows that pectin engineering in the context of food processing applications is an
important tool towards improving the textural/rheological properties of plant based foods. However,
it should be noted that the successful application of pectin engineering depends on the intrinsic
properties of the raw material, pretreatments involved and the processing technology applied.
The use of anti-pectin antibodies to map the distribution of pectin methylesterification provides
additional information important for targeted manipulation of processing dependent pectin
properties.

64
REFERENCES Ahmed, A.E.R, Labavit, J.M. (1997). A simplified method for accurate determination of cell wall
uronide content. Journal of Food Chemistry, 1: 361-365.
Arltoft, D., Ipsen, R., Christensen, N., & Madsen, F. (2006). Localizing pectin in dairy products using
direct immunostaining. In P. A. Williams, & G. O. Phillips (Eds.), Gums and Stabilizers for the Food
Industry 13 (pp. 41–50). The Royal Society of Chemistry.
Balogh, T., Van Loey, A.M., Smout, C., Hendrickx, M. (2004). Thermal and high-pressure inactivation
kinetics of carrot pectin methylesterase: from model system to real foods. Innovative Food Science
and Emerging Technologies, 5, 429-436.
Blumenkrantz, N., Asboe-Hansen, G. (1973). New method for quantitative determination of uronic
acids. Analytical Biochemistry, 54, 484-489.
Bonjongsinsiri, P., Shields, J., and Wicker, L. (2004). Vacuum infusion of plant of fungal
pectinmethylesterase and calcium effects the texture and structure of egg plant. Journal of
Agricultural and Food Chemistry, 52, 8214-8223.
Braccini I., Grasso R., Perez S. (1999). Conformational and configurational features of acidic
polysaccharides and their interactions with calcium ions: a molecular modeling investigation.
Carbohydrate Research, 317(1–4), 119–30.
Braga, M., Pessoni, R., Dietritch, S. Cell wall polysaccharide composition of leaf of tropical rubiaceae
differing in phytoalexin response. Revista Brasileira de Fisiologia Vegetal (1998), 10 (2), 71-78.
Brummel, D.A., and Harpster, M.H.(2001). Cell wall metabolism in fruit softening and quality and its
manipulation in transgenic plants. Plant Molecular Biology,47(1-2), 311-340.
Caffall, K.H., Mohnen, D. (2009). The structure, function, and biosynthesis of plant cell wall pectic
polysaccharides. Carbohydrate Research, 344, 1879 – 1900.

65
Chin, L.H., Ali, Z.M., Lazan, H. (1999). Cell wall modifications, degrading enzymes and softening of
carambola fruit during ripening. Journal of Experimental Botany, 50, 767-775.
Christiaens, S., Van Buggenhout, S., Ngouémazong, E. D., Vandevenne, E., Fraeye, I., Duvetter, T., Van
Loey, A., and Hendrickx, M. (2010). Anti-homogalacturonan antibodies: a way to explore the effect of
processing on pectin in fruits and vegetables? Food Research International, 44 (1), 225-234.
Clausen, M.H., Willats, W.G.T., Knox, J.P. (2003). Synthetic methyl hexagalacturonate hapten
inhibitors of anti-homogalacturonan monoclonal antibodies LM7, JIM5 and JIM7. Carbohydraye
Research, 338, 1797-1800.
Colle, I., Van Buggenhout, S., Van Loey, A.M., Hendrickx, M. (2010). High pressure homogenization
followed by thermal processing of tomato pulp: Influence on microstructure and lycopene in vitro
bioaccessility. Food Research International, 43, 2193-2200.
Crelier, S., Robert, M.C., Claude, J., Juillerat, M.A. (2001). Tomato (Lycospersicon esculentum) pectin
methylesterase and polygalacturonase behaviors regarding heat and high-pressure induced
inactivation. Journal of Agricultural and Food Chemistry, 49(11), 5566-5575.
Daas, P.J.H., Meyer-Hansen, K., Schols, H.A., De Ruiter, G.A., Voragen, A.G.J. (1999). Investigation of
the non-esterified galacturonic acid distribution in pectin with endo-polygalacturonase.
Carbohydrate Research, 318, 135 – 145.
De Roeck, A., Sila, D. N., Duvetter, T., Van Loey, A., Hendrickx, M. (2008). Effect of high pressure/
high-temperature processing on cell wall pectic substances in relation to firmness of carrot tissue.
Food Chemistry 107 (3), 1225-1235.
Dénes, J., Baron, A., Drilleau, J. 2000. Purification, properties and heat inactivation of
Pectinmethylesterase from apple (Golden delicious). Journal of the Science of Food and Agriculture,
80,1503-9.
Drush, S. (2007). Sugar beet pectin: a novel emulsifying wall component for microencapsulation of
lipophillic food ingredients by spray-drying. Food Hydrocolloids, 21(7), 1223-1238.

66
Fachin, D. (2003). Temperature and pressure inactivation of tomato pectinases: a kinetic study.
Doctoral dissertation, Department of Food Technology, Faculty of Agriculture and Applied Biology
Sciences, Katholieke Universiteit Leuven, Leuven, Belgium.
Fachin, D., Van Loey, A., Nguyen, B.L., Verlent, I., Indrawati, I., and Hendrickx, M. (2002). Comparative
study of the inactivation kinetics of pectin methylesterase in tomato juice and purified form.
Biotechnology Progress, 18(4), 739-744.
Fischer L. Robert, Bennet B. Alan 1991. Role of cell wall hydrolases in fruit ripening. Annual Review
Plant physiology.
Fraeye, I., De Roeck, A., Duvetter, T., Verlent, I., Sila, D.N., Van Loey, A., Hendrickx, M. (2007).
Comparison of enzymatic de-esterification of strawberry and apple pectin at elevated pressure of
fungal pectinmethylesterase. Innovative Food Science and Emerging Technologies, 8 (1), 93-101.
Fraeye, I., Ines, C., Vandevenne, E., Duvetter, T., Van Buggenhout, S., Moldenaers, P., Van Loey, A.,
Hendrickx, M. (2009). Influence of pectin structure on texture of pectin–calcium gels. Innovative
Food Science and Emerging Technologies, 11, 401 – 409.
Gaffe, J., Tieman, D.M., Handa, A.K., 1994. Pectin methylesterase isoforms in tomato (Lycopersicon
esculentum) tissues—effects of expression of a pectin methylesterase antisense gene. Plant
Physiology. 105 (1), 199-203, Plant biology 42,675-703.
Goding, J. W. (1996). Monoclonal antibodies: Principle and practice (3rd ed). London, UK: Academic
Press.
Goodman, C.L., Fawcett, S., Barringer, S.A. (2002). Flavor, viscosity and color analyses of hot and cold
break tomato juices. Journal of Food Science, 404-408.
Greve, L., Shackel, K. A., Ahmadi, H., McArdle, R. N., Gohlke, R., and Labavitch, J.M. (1994). Impact of
heating on carrot firmness: Contribution of cellular turgo. Journal of Agricultural and Food Chemistry,
42, 2896-2899.
Guillotin S.E, Bakx E, Boulenguer P, Mazoyer J, Schols HA, Voragen AGJ. (2005). Populations having
different GalA blocks characteristics are present in commercial pectins which are chemically similar
but have different functionalities. Carbohydr Research, 60(3),391–8.

67
Hsu K-C, Tan F-J, Chi H-Y.(2008). Evaluation of microbial inactivation and physicochemical properties
of pressurized tomato juice during refrigerated storage. Lebensm Wiss Technology 41(3):367-375.
Ishii, T., 1995. Pectic polysaccharides from bamboo shoot cell walls. Mokuzai Gakkaishi, 41, 669-676.
Ishii, T., 1999a O-acetylated oligosaccharides from pectins of potato tuber cell walls. Plant
Physiology, 113, 1265-1272.
Kashyap, D.R., Vohra, P.K., Chopra, S., Tewari, R. (2001). Applications of pectinases in the commercial
sector: A review. Bioresources Technology, 77(3), 215-227.
Klavons, J.A., Benett, R.D. (1986). Determination of methanol using alcohol oxidase and its
application to methyl ester content of pectin. Journal of Agriculture and Food Chemistry, 50: 597-
599.
Knox, J. P., Linstead, P. J., Cooper, C., and Roberts, K. (1990). Pectin esterification is spartially
regulated both within cell walls and between developing tissues of root apices. Planta, 181, 512-521.
Krebbers, B., Matser, A.M., Hoogerwerf,S.W.,Moezelaar, R., Tomassen, M.M., Van den Berg, R.W.
(2003). Combined high-pressure and thermal treatments for processing of tomato puree: evaluation
of microbial inactivation and quality parameters. Innovative Food Science and Emmerging
Technologies 4, 377-385.
Leroux, J., Langendorff, V., Schick, G., Vaishnav, V., Mazoyer, J. (2003). Emulsion stabilizing properties
of pectin. Food Hydrocolloids, 17, 455-462.
Libermans, M., Mutaftschiev, S., Jauneau, A., Vian, B., Catesson, A., Goldberg, R. 1999. Mung bean
hypocotyl homogalacturonan : localization, organization and origin. Annals of Botany -London
84,225-33.
Löfgren, C., Guillotin, S., Evenbratt, H., Schols, H.A., Hermanson A.M. (2005). Effect of calcium, pH,
and blockiness on kinetic rheological behavior and microstructure of pectin gels.
Biomacromolecules, 6 (2), 646-652.

68
Lopez, P.,Sanchez, A. C., Vercet, A., Burgos, J. (1997). Thermal resistance of tomato
polygalacturonase and pectin methylesterase at physiological pH. Zeitschrift Fur Lebensmittel-.
Untersuchung Und-Forschung. A Food Research and Technology 204(2), 146-150.
Lopez-Sanchez P., Njsse J., Blonk H.C.G., Lucy B., Schumm S., Langhan M., (2010): Effect of
mechanical and thermal treatments on the microstructure and rheological properties of carrots,
tomato and broccoli dispersions. Journal of the Science of Food and Agriculture, 91, 207-217.
LY-Nguyen, B., Van Loey, A.M., Smout, C., Verlent, I., Ozcan, S.E., Fachin, D., Truong, S.V., Duvetter,
T., Hendrickx, M.E.(2003). Mild-heat and high-pressure inactivation of carrot pectin methyl esterase:
A kinetic study. Journal of Food Science, 68(4), 1377-1383.
Matsuhasi, N. Nishikawa, T. Negishi and C. Hatanaka (1993). Enzymic-HPLC determination of the
amount and distribution of the galacturonan region in pectate molecules. Journal of Liquid
Chromatography, 16, 3203–3215.
McFeeters, R.F., Amstrong, S.A., (1984). Measurement of pectin methylation in plant cell walls.
Analytical Biochemistry, 139: 212- 217.
Mian I, Bradwell A, Olson A (1991). "Structure, function and properties of antibody binding sites".
Journal of Molecular Biology, 217 (1): 133–51.
Nari,J., Noat, G., Ricard, J. (1991). Pectin methylesterase, metal-ions and plant cell-wall extention-
Hydrolysis of pectin by plant cell-wall pectin methylesterase. Biochemical Journal, 279,343-350.
Neirynck, N., Van der Meeren, P., Gorbe, S.B., Dierckx, S., Dewettinck, K. (2004). Improved emulsion
stabilizing properties of whey protein isolate by conjugation with pectins. Food Hydrocolloids
18,949-957.
Ng, A., Waldron, K.W. (1997). Effect of cooking and pre-cooking on cell wall chemistry in relation to
firmness of carrot tissues. Journal of Science of Food and Agriculture, 73: 503-512.
Oakenfull, D.G., Scott, A. 1984. Hydrophobic interaction in the gelation of high methoxyl pectins.
Journal of Food Science 49, 1093-1098.

69
O'Neill, M., Albersheim, P. and Darvill ,A. The pectic polysaccharides of primary cell walls. In: D.M.
Dey,( Editor), Methods in Plant Biochemistry, Vol. 2, Academic Press, London (1990), pp. 415–441.
Peeters, L., Fachin, D., Smout, C., Van Loey, A., and Hendrickx, M. (2004). Influence of β-subunit on
thermal and high pressure process stability of tomato polygalacturonase. Biotechnology and
Bioengineering, 86, 543-549.
Powell, D.A., Moris, E.R., Gidley, M.J. and Rees, D.A. (1982). Conformations and interactions of
pectins. II. Influence of residue sequence on chain associations in calcium pectate gels. Journal of
Molecular Biology 155, 517-531.
Renard CM, Thibault J. F.(1993). Structure and properties of apple and sugar beet pectins extracted
by chelating argents. Carbohydrates Research, 244: 99-114.
Ridley, B.L., O’Neill, M.A., Mohnen, D. (2000). Pectins: Structure, biosynthesis and
Oligogalacturonide-related signaling. Phytochemistry,57 (6), 929-967.
Rodrigo, D., Cortés, C., Clynen, E., Schoofs, L., Van Loey, A., Hendrickx, M. (2005). Thermal and high-
pressure stability of purified polygalacturonase and pectinmethylesterase from four different tomato
processing varieties. Food Research International, 39,(4 ) 440-448.
Roy, S.S., Taylor, T.A., Kramer, H.L. (2001). Textural and ultrastructural changes in carrot tissue as
affected by blanching and freezing. Journal of Food Science, 66:176-180.
Schols, H.A., and Voragen, A.G.J. 1996. Complex pectins: Structure elucidation using enzymes. In: J.
Visser and A.J. Voragen(editors) Pectins and Pectinases, Elsevier Science, Amsterdam. pp 3-19.
Sila D.N., Elloit, F., Smout C., A. Van Loey, and M. Hendrickx. (2006b). non-enzymatic
depolymerisation of carrot pectin; Toward a better understanding of carrot texture during thermal
processing. Journal of Food Science, 71(1), E1-E9.
Sila D.N., S. Van Buggenhout, T. Duvetter, I. Fraeye, A. De Roeck, A. Van Loey, and Hendrickx, M.
(2009). Pectins in Processed Fruits and Vegetables: Part II – Structure-Function Relationships,
Comprehensive Reviews in Food Science and Food Safety Vol. 8: 86 – 104

70
Sila, D. N., Smout, C., Elliot, F., Van Loey, A., and Hendrickx, M. (2006a). Non-enzymatic
depolymerization of carrot pectin: Toward a better understanding of carrot texture during thermal
processing. Journal of Food Science.
Sila, D.N., Yue, X., Van Buggenhout, S., Smout, C., Van Loey, A., and Hendrickx, M.(2007). The
relation between (bio-)chemical, morphological, and mechanical properties of thermally processed
carrots as influenced by high-pressure pretreatment condition. European Food Research and
Technology, 226, 127-135.
Sila, D.N., Doungla, E., Smout, C., Van Loey, A., Hendrickx, M. (2006). Pectin fractions inter-
conversions: insight into understanding texture evolution of thermally processed carrots. Journal of
Agricultural and Food Chemistry, 54(22), 8471-8479.
Sila, D.N., Smout C, Satara, Y., Truong, V., Van Loey, A., Hendrickx, M. (2007). Combined thermal and
high pressure effects on carrot pectinmethylesterase stability and catalytic activity. Journal of Food
Engineering 78:755-64.
Sila, D.N., Smout C, Truong, V., Van Loey, A., Hendrickx, M. (2005).The influence of pretreatment
conditions on the texture and cell wall components of carrots during thermal processing. Journal of
Food Science 69(5):205-211
Sila, D.N., Smout, C., Vu, T.S., and Hendrickx, M. (2004). Effect of high-pressure pretreatment and
calcium soaking on the texture degradation kinetics of carrots during thermal processing. Journal of
Food Science, 69(5), 205-211.
Somerville, C., Bauer, S., Brininstool, G., Facette, M., Hamann, T., Milne, J.,Osborne, E., Paredez, A.,
Persson, S., Raab, T., Vorwerk, S., Youngs, H.,(2004). Toward a systems approach to understanding
plant-cell walls. Science 306
Strom, A., Ribelles, P., Lundin, L., Norton, I., Morris, E.R., Williams, M.A. (2007). Influence of pectin
fine structure on the mechanical properties of calcium-pectin and acid-pectin gels. Bio-
macromolecules 8(9): 2668-74.

71
Van Buggenhout, S., Messagie, I., Maes, V., Duvetter, T., Van Loey, A., and Hendrickx, M. (2006).
Minimizing texture loss of frozen strawberries: Effect of infusion with pectinmethylesterase and
calcium combined with different freezing conditions and effect of subsequent storage/thawing
conditions. European Food Research and Technology, 223 (3), 395-404.
Van Buggenhout, S., Sila, D.N., Duvetter, T., Van Loey, A., and Hendrickx, M. (2009). Pectins in
processed fruits and vegetables: Part III-Texture engineering. Comprehensive Reviews in Food
Science and Food Safety, 8 (2), 105-117.
Van Buren, J.P. (1979). The chemistry of texture in fruits and vegetables. Journal of texture studies,
10, 1-23.
VandenBosch, K.A., Bradley, D. J., Knox, J.P., Perotto, S., Butcher, G.W. and Brewin, N.J.(1989).
Common components of the infection thread matrix and the intercellular space identified by
immunocytochemical analysis of pea nodules and uninfected root. The European Molecular Biology
Organization Journal, 8, 335-342.
VandenBosch, K.A., Bradley, D.J., Knox, J.P., Perotto, S., Butcher, G.W., & Brewin, N.J.(1989). common
components of the infection thread matrix and the intercellular space identified by
immunocytochemical analysis of pea nodules and uninfected roots. The European Molecular Biology
Organization Journal, 8, 335-342.
Verhertbruggen, Y., Marcus, S. E., Haeger, A., Ordaz-Ortiz, J.J., Knox, J. P. (2009). An extended set of
monoclonal antibodies to pectin homogalacturanan. Carbohydrate Research, 344, 1858-1862.
Verlent, I., Hendrickx, M., Verbeyst, A., Van Loey, A. (2006). Effect of temperature and pressure on
combined action of purified tomato pectinmethylesterase and polygalacturonase in presence of
pectin. Enzyme and Microbial Technology, 40 (5), 1141-1146.
Verlent, I., Van Loey, A., Smout, C., Duvetter, T., Nguyen, B.L., Hendrickx, M. (2004) Changes in
purified tomato pectinmethylesterase activity during thermal and high pressure treatment. Joural of
the Science of Food and Agriculture, 84, 1839-1847.

72
Versari, A., Biesenbruch, S., Barabanti, D., Farnell, P., Galassi, S. (1999). HPAEC-PAD analysis of
oligogalacturonic acid in strawberry juice. Food Chemistry, 66, 257-261.
Vu, T.S., Smout, C., Sila, D.N., Ly Nguyen, B., Van Loey, A., and Hendrickx, M. (2004). Effect of
pretreating on thermal degradation kinetics of carrot texture. Innovative Food Science and
Emmerging Technologies, 5, 37-44.
Vu, T.S., Smout, C., Sila, D.N., Van Loey, A., and Hendrickx, M. (2006). The effect of brine ingredients
on carrot texture during thermal processing in relation to pectin depolymerization due to β-
elimination reaction. Journal of Food Science, 71(9), E370-E375.
Waldron, K. W., Parker, M. L., and Smith, A. C. (2003). Plant cell walls and food quality.
Comprehensive Reviews in Food Science and Food Safety, 2, 128-146.
Willats, W. G.T., Gilmartin, P.M., Mikkelsen, J. D., and Knox, J. P. (1999). Cell wall antibodies without
immunization: generation and use of de-esterified homogalacturonan block-specific antibodies from
naive phage display library. The Plant Journal, 18, 57-65.
Willats, W.G.T., Orfila, C., Limberg, G., Buchholt, H. C., Van Alebeek, G. J., Voragen, A. G., et al.
(2001). Modulation of the degree and pattern of methylesterification of pectic homogalacturonan in
plant cell walls. Implications for pectinmethylesterase action, matrix properties and cell wall
adhesion. The Journal of Biological Chemistry, 276 (22), 19404-19413.
Willats, W.G.T., Orfila, C., Limberg, G., Buchholti, H.C., van Alebeek, G-J.W.M., Voragen, A.G.J.,
Marcus, S.E., Christensen, T.M.I.E., Mikkelsen, J.D., Murray, B.S., Knox, J.P. (2001b). Modulation of
degree and pattern of methyl esterification of pectic homogalacturonan in plant cell
walls – Implications for pectin methyl esterase action, matrix properties, and cell adhesion. Journal of
Biological Chemistry 276, 19404-19413.
Willats, W.G.T., Steele-King, C. G., McCartney, L., Orfila, C., Marcus, S. E., Knox, J. P.,(1999). Making
and Using antibodies to study plant cell walls. Plant physiology Biochemistry, 38 (1/2) 27-36.

73
Willats,W. G., Rasmussen, S. E., Kristensen, T., Mikkelsen, J. D., & Knox, J. P. (2000). Sugar-coated
microarrays: A novel slide surface for the high-throughput analysis of glycans. Proteomics,2(12),
1666–1671.
Zhan, D., Janssen, P., Mort, A.J. 1998. Scarcity or complete lack of single rhamnose residues
interspersed within the homogalacturonan regions of citrus pectin. Carbohydrates. Research. 308,
373-380.