extracellular polysaccharides of azotobacter …jb.asm.org/content/88/2/329.full.pdf ·...
TRANSCRIPT

JOURNAL OF BACTERIOLOGYVol .88, No. 2, p. 329-338 August, 1964Copyright ( 1964 American Society for Microbiology
Printed in U.S.A.
EXTRACELLULAR POLYSACCHARIDES OFAZOTOBACTER VINELANDII1
GARY H. COHEN' AND DONALD B. JOHNSTONE
Department of Agricultural Biochemistry, University of Vermont, Burlington, Vermont
Received for publication 11 March 1964
ABSTRACTCOHEN, GARY H. (University of Vermont, Bur-
lington), AND DONALD B. JOHNSTONE. Extracel-lular polysaccharides of Azotobacter vinelandii.J. Bacteriol. 88:329-338. 1964.-Extracellular poly-saccharides synthetized by Azotobacter vinelandiistrains 155, 102, and 3A were shown to be car-boxylic acid heteropolysaccharides of apparenthigh molecular weight. Cells were grown in anitrogen-free, mineral broth medium with 2%sucrose. Extracellular slime was recovered bycentrifugation and purified by repeated alcoholprecipitation and Sevag deproteinization. Cap-sular polysaccharide was recovered from washedcells by mild alkaline digestion. Methods of isola-tion and purification appeared to provide poly-saccharide showing no evidence of heterogeneitywhen examined by chemical and physical methods.Infrared analysis of purified slime from the threestrains suggested fundamental structural simi-larities. Colorimetric, paper chromatographic, andenzymatic analyses on both intact and acid-hydrolyzed slime polysaccharide indicated thatthe polymers contained in common galacturonicacid, [a] D-glucose, and rhamnose at a ratio ofapproximately 43:2:1, as well as a hexuronic acidlactone, probably mannurono-lactone. However,as shown by chemical and infrared analysis, minordifferences did exist; namely, slime from strain155 and 102 contained o-acetyl groups, whereasslime from strain 3A contained none. A sialic acid-like component (1.5% of dry weight of the poly-saccharide, calculated as N-acetyl neuraminicacid), was found only in the slime of strain 155.Capsular polysaccharide composition closelyresembled that for slime. It is of interest that themajor slime components were identical whetherthe energy source provided for the cells was su-crose, glucose, fructose, or ethanol.
1 From a dissertation submitted by the seniorauthor in partial fulfillment of the requirementsfor the Ph.D. degree. Contribution from the Uni-versity of Vermont, Agricultural ExperimentStation, Journal Article No. 136.
2 Present address: School of Veterinary Medi-cine, University of Pennsylvania, Philadelphia.
Azotobacter vinelandii, under most culturalconditions, synthesizes copious amounts ofextracellular polysaccharide. This polysaccharideoccurs morphologically in two distinct forms:(i) discrete capsules adhering to the cell walls,and (ii) as amorphous slime found loosely at-tached to the capsules and in the culture mediumfree from the cells.
Within recent years, reports have appearedimplicating A. vinelandii polysaccharide withvarious phenomena associated with this species.Wyss, Neuman, and Socolofsky (1961) reportedthat A. vinelandii cysts, which conceivably wereformed from extracellular polysaccharide syn-thesized by this species, consisted primarily ofrhamnose and glucose components. Eklund andWyss (1962) demonstrated the presence of acapsule-digesting enzyme formed when A. vine-landii cells were infected with bacteriophage.Preliminary tests by these authors on the chemi-cal composition of the capsules indicated thepresence of glucuronic acid and an unidentifiedcomponent. Recently, Cohen and Johnstone(1963a) presented evidence that the acid reactionproduced in culture medium by A. vinelandii wasassociated with extracellular slime synthesis.They reported galacturonic acid as the majorcomponent of the slime.
Earlier reports on the chemical composition ofA. vinelandii polysaccharide indicated that thecapsules consisted of glucose and rhamnose(Kaufman and Rapaske, 1958), whereas the cell-free slime, as stated by Kaufman (1960), con-sisted of glucose, a trace of uronic acid, and nomethyl pentose.
Knowledge pertaining to the chemical com-position of the extracellular polysaccharide ofA. vinelandii is basic to an understanding of thepolymer's functional role in this species. Further-more, such information may provide additionaldistinguishing characteristics for the differentia-tion of the often confused species, A. vinelandiiand A. agilis (Johnstone, 1962).
329
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

COHEN AND JOHNSTONE
Results presented here concern some studiesof the physical and chemical properties of extra-cellular polysaccharide produced by severalstrains of A. vinelandii. A preliminary report ofpart of this work was presented (Cohen andJohnstone, 1963b).
MATERIALS AND METHODS
Growth of the organisms. The organisms usedthroughout this study were A. vinelandii strains3A (ATCC 12837), 102 (ATCC 9046), and 155(ATCC 12518). Burk's nitrogen-free broth(Wilson and Knight, 1952) at pH 7.0 supple-mented with 2% sucrose was inoculated with theappropriate strain growing in the logarithmicphase. Cultures were incubated at 31 C in 7.5-liter New Brunswick fermentors with sterilemoist air supplied at the rate of 4 liters per min,and an impellor rotation of 130 rev/min. Incu-bation was discontinued and the polysaccharidewas harvested when the reaction of the culturemedium reached pH 5.5 (Cohen and Johnstone,1963a). This usually occurred in about 5 days.
Isolation and purification of extracellular poly-saccharide. The viscous culture medium wasdiluted onefold with distilled water, and the cellswere removed in a Servall continuous-flow cen-trifuge at 18,400 X g. The slime polysaccharide,designated hereafter as SP, was precipitated fromthe culture supernatant fluid by the addition of4 volumes of cold (-10 C) acidified (HCl) ab-solute ethanol. The precipitated SP was removed,washed with absolute ethanol, air-dried, andredissolved in water. Dissolution of SP wasfacilitated by mixing in water for 12 hr at 4 C.This was then centrifuged at 18,400 X g for 1 hrat 4 C to remove undissolved material. Theabove purification procedures were repeatedthree times. SP recovered from A. vinelandii155 and 102 was purified by these methods.To obtain a highly purified preparation forstudying chemical and physical properties ingreater detail, SP from strain 3A was subjectedto the following additional procedures. Proteinwas removed by Sevag (Sevag, Lackman, andSmolens, 1938) extraction. Sevag deproteiniza-tion was discontinued when protein was nolonger detected by the method of Lowry et al.(1951). The turbid, opalescent nature of aqueoussolutions of SP was completely removed by thistreatment, leaving a water-clear solution. At thispoint, 1% (w/v) NaCl was added to insure
complete precipitation, and SP was precipitatedwith cold acidified ethanol. The precipitate wasredissolved, centrifuged at 18,400 X g for 3 hrat 4 C to remove undissolved material, and re-precipitated. Precipitated SP from each strainwas washed successively in 80% ethanol, abso-lute ethanol, acetone, and ether, and was driedto constant weight in vacuo at room temperature.
Capsular polysaccharide was removed fromwashed cells by adjusting the reaction of themedium to pH 9.0 with NaOH and incubatingthe cells on a rotary shaker for 1 hr at roomtemperature. The alkaline digestion mixturewas centrifuged, the cell mass was discarded,and the polysaccharide was isolated and purifiedby the above methods. For studies of SP syn-thesized from ethanol, strain 3A was grown on 1%(v/v) ethanol as the sole carbon and energysource with an additional 1% added after 16hr of incubation as described by Kaufman (1960).
Methods for determining polysaccharide homo-geneity. The crude supernatant fluid containingSP was titrated with a 3% (w/v) solution ofethylhexadecyldimethyl ammonium bromide un-til precipitation of acidic polysaccharide nolonger occurred. The culture medium was clearedof precipitated SP by centrifugation and refrig-erated overnight, and the titration procedure wasrepeated. After all traces of precipitate wereremoved, 4 volumes of cold (-10 C) ethanolwere added to precipitate any remaining poly-saccharide. Detergent was removed from SPby washing with methanol.SP was analyzed electrophoretically with 5%
acrylamide gel as a supporting medium, accord-ing to the methods of Davis (1963). The poly-saccharide was located on acrylamide gel with0.06% (w/v) Toluidine Blue in 7% (v/v) aceticacid. Destaining was accomplished with 7%(v/v) acetic acid.
Ultracentrifugal studies were carried out in a2.5 to 5.0% (v/v) linear glycerol density gradientat 39,000 rev/min with a SW 39 swinging-bucketrotor of a Spinco L preparative ultracentrifuge.The orcinol test (Mejbaum, 1939) was used toestimate polysaccharide concentration in thepresence of glycerol.
Physical methods. Infrared analyses of SPincorporated into KBr pellets were done in aPerkin-Elmer model 21 double-beam spectro-photometer. For these analyses, SP was pre-cipitated from neutral solution and air-dried.
330 J. BACTERIOL.
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 88, 1964 EXTRACELLULAR POLYSACCHARIDES OF A. VINELANDII
All other spectrophotometric measurements wereperformed in a Beckman model DU spectro-photometer with cuvettes having a light path of1.0 cm. Molecular weight estimation of SP,dissolved in 0.5 M NaCl, was performed, by use ofSephadex gel filtration, according to Flodin(1962).Chemical methods. Phosphorus and nitrogen
determinations were carried out by the Fiskeand SubbaRow (1925) and micro-Kjeldahl tech-niques, respectively. Protein concentration wasestimated as described by Lowry et al. (1951)with purified A. vinelandii 3A protein as astandard. The protein standard was preparedaccording to Racusen and Johnstone (1961),except that the method of Lowry et al. (1951)was employed instead of the biuret reaction.Hexuronic acid was detected and identified bythe following methods: carbazole method, modi-fied carbazole test for galacturonic acid, the reac-tion of glucuronic acid with thioglycolic acid andsulfuric acid in the presence of mannose, and thereaction of galacturonic acid with L-cysteine andsulfuric acid according to the methods of Dische(1962). Quantitative determinations of hexoseand methylpentose were accomplished by twodifferent procedures. (i) Determinations werecarried out on the intact SP by use of the primaryL-cysteine-sulfuric acid reaction for hexose(Dische, Shettles, and Osnos, 1949) and the 10-min modification of the reaction of methylpen-tose with L-cysteine and sulfuric acid (Discheand Shettles, 1948). Adequate internal controlswere included to account for both nonspecificcolor reactions caused by sulfuric acid treatmentof the polysaccharide and the carbohydrate con-tent of the polymer. (ii) The sugar moieties,obtained from acid hydrolysis of SP, were sepa-rated by paper chromatography, extracted fromthe paper, as recommended by Smith and Mont-gomery (1959), and assayed by the phenol-sulfuricacid method of Dubois et al. (1951).
O-acetyl groups were estimated according toHestrin (1949). The thiobarbituric acid assaymethod of Warren (1959) was employed forthe determination of sialic acid. A purified sampleof IV-acetyl neuraminic acid isolated from bovinesubmaxillary gland was the gift of R. C. Wood-worth, University of Vermont. Total carbohy-drate was estimated by the phenol-sulfuric acidmethod of Dubois et al. (1951), and reducingsugar was determined by the colorimetric pro-
cedure of Nelson (1944) and Somogyi (1952).Glucose was used as a standard for both methods.
Chromatography. Paper chromatography wascarried out by descending and ascending tech-niques on Whatman no. 1 and 3 MM paper.The following solvent systems were used: (A)n-butanol-pyridine-water (9:5:8, v/v); (B)ethyl acetate-pyridine-water (12:5:4, v/v); (C)isopropanol-butanol-water (14:2:4, v/v); (D)ethyl acetate-pyridine-acetic acid-water (5:5:1:3,v/v). The sugars and sugar derivatives werelocated and identified with aniline hydrogenphthalate (Partridge, 1949), aniline diphenylamine (Smith, 1958), hydroxamic acid spray forlactones and esters (Abdel-Akher and Smith,1951), glucose oxidase reagent (WorthingtonBiochemical Corp., Freehold, N.J.; Salton, 1960),and 0.2% (w/v) ninhydrin in acetone for aminosugars.Acid hydrolysis. Acid hydrolyses were carried
out in Teflon-lined screw-capped tubes. Hy-drolysates to be used for paper chromatographywere prepared as follows. Polysaccharide washydrolyzed in 2 N H2SO4 at 100 C for 5 hr. (Atime course was run to obtain maximal reducingvalues.) The hydrolysate was neutralized withsolid Ba(OH)2, the precipitated BaSO4 wasremoved by centrifugation, and excess Ba++ wasremoved by passing the supernatant fluid througha column containing Dowex 50 (hydrogen form).
For large-scale separation of SP componentsSP was hydrolyzed in 2 N HCl at 100 C for 5hr, and the hydroylsate was filtered. The hy-drolysate was evaporated to dryness under re-duced pressure and redissolved; the procedurewas repeated four times to remove HCl.The acidic sugars were adsorbed on Dowex
1-2X (acetate form); the neutral sugars (neutralfraction) were eluted with distilled water, chro-matographed on Whatman 3 MM with solventB, and isolated according to Smith and Mont-gomery (1959). The acidic sugars (acidic frac-tion), after elution from the resin with 4 N aceticacid, were isolated by the same chromatographictechniques.
RESULTS
Homogeneity of polysaccharide. The addition ofalcohol to the culture supernatant fluid of strain3A, previously depleted of SP by detergent treat-ment, resulted in the formation of a white floc-culant precipitate. This second precipitate (43
331
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

COHEN AND JOHNSTONE
mg per liter of supernatant culture medium) wasalso demonstrated after alcohol precipitation ofSP, but in both cases occurred only after 12 to 24hr at 4 C. Analysis of the dialyzed second precipi-tate indicated a carbohydrate to protein ratio of2.2, no uronic acid, and the presence of glucose.The precipitate was associated with the greenfluorescent pigment of A. vinelandii and may besimilar to the fluorescent peptide described byBulen and Le Comte (1962). The yield of crudeSP recovered after treatment with alcohol ordetergent was 740 mg per liter of supernatantculture medium. These figures are relative to theculture batch, and represent an average of poly-saccharide from several isolations. Recovery ofSP immediately after alcohol precipitation ap-peared to separate the two precipitates, sincethe second formed only after a period of time inthe cold.The homogeneity of highly purified SP was
examined by acrylamide gel "disc" electrophore-sis. After electrophoresis, the gels were stainedwith Toluidine Blue. A slow-moving anionic bandexhibiting metachromasy, which is indicative ofacid polysaccharides, was observed along with afaintlystained area preceding the heavy band. Thefaster-moving area probably represented speciesof the acidic polymer of lower molecular weight,since it also exhibited metachromatic properties.
Sedimentation profiles of highly purified SPin a linear glycerol density gradient revealed asingle symmetrical peak.From these data, it would appear that the SP
under investigation was homogeneous in com-position.
Physical analysis. The infrared absorptionspectra of SP in the dissociated form from strains3A, 155, and 102 are illustrated in Fig. 1. Thefollowing functional group assignments weremaed: OH, 3.0 ; C-H, 3.45 ,; -CH2CO-
800w
< 60
z 40
20
INFRARED SPECTRA
O-R, 5.77 and 8.1 ,u; and O=C-0, 6.25 and7.1 ,u (Levine, Stevenson, and Kabler, 1953).These spectra were quite similar, especially inthe region 8.5 to 14.0 ,A, which indicated a funda-mental structural similarity. However, thespectra for SP 3A differed by the absence ofester linkage absorption at 8.1 j,, and only aslight shoulder at 5.77 ,u which corresponded tothe carbonyl band. Subsequent chemical tests(Hestrin, 1949) failed to detect o-acetyl groups inSP 3A, whereas positive results, as describedbelow, were obtained for SP 155 and 102.The ultraviolet spectrum of a 0.1% (w/v)
aqueous solution of SP 3A revealed no maximain the region 230 to 300 m,u. Molecular weightstudies with various types of Sephadex (G-25 toG-200) suggested that the largest molecularweight of a species of the polymer was greaterthan 200,000, since it is excluded from a columnof Sephadex G-200. This should be interpretedwith caution, since one must assume that SPfollows the principle of exclusion from Sephadexgel pores by size alone.The highly viscous polysaccharide precipitated
in the presence of multivalent cations, or whenthe pH reaction was adjusted to pH 3.5. Theseattributes, as pointed out by Whistler (1959),are indicative of carboxylic acid polysaccharides.Exhaustive dialysis of SP against water resultedin the formation of a firm gel.
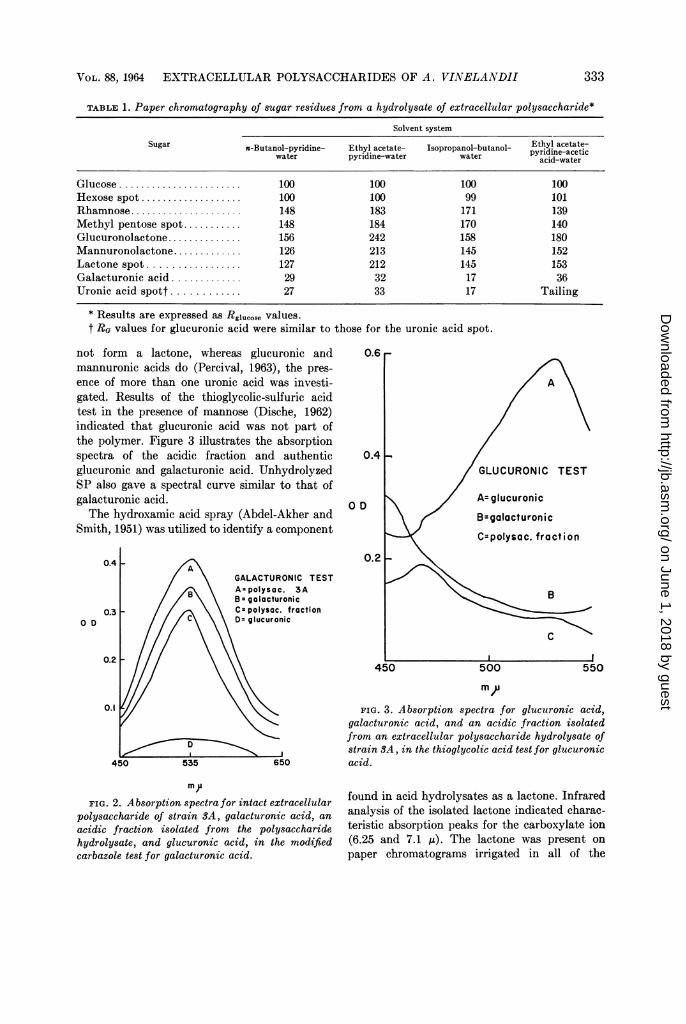
Chemical analysis. SP possessed no reducingproperties (Nelson, 1944; Somogyi, 1952) anda negligible amount of phosphorus (Fiske andSubbaRow, 1925). Paper chromatography ofacid hydrolysates revealed four major compo-nents which were identified by the following meth-ods as galacturonic acid, a lactone, rhamnose,and [a]D-glucose (Table 1).The results of the carbazole test (Dische, 1962)
on SP indicated a high concentration of uronicacid. Furthermore, the modified carbazole method(Dische, 1962) demonstrated that galacturonicacid was present. Figure 2 illustrates the absorp-tion spectra for unhydrolyzed SP, an acidicfraction isolated from an acid hydrolysate, andauthentic samples of glucuronic and galacturonicacid.
Further presumptive evidence for galacturonicacid was indicated by a positive L-cysteine-sul-furic acid test (Dische, 1962). However, since auronic acid lactone was present (as describedbelow), and presumably galacturonic acid does
3 4 5 6 7 a 9 0 1I1 12 13 14 IS
MICRONS
FIG. 1. Infrared absorption spectra for extra-cellular polysaccharides produced by Azotobactervinelandii strains 155, 102, and SA.
332 J. BACTERIOL.
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
VOL. 88, 1964 EXTRACELLULAR POLYSACCHARIDES OF A. VINELANDII
TABLE 1. Paper chromatography of sugar residues from a hydrolysate of extracellular polysaccharide*
Solvent system
Sugar n-Butanol-pyridine- Ethyl acetate- Isopropanol-butanol- Ethyl acetate-water pyridine-water water pyridine-aceticwater ~~~~~~~~~~~~~acid-water
Glucose ....................... 100 100 100 100Hexose spot ................... 100 100 99 101Rhamnose..................... 148 183 171 139Methyl pentose spot........... 148 184 170 140Glucuronolactone.............. 156 242 158 180Mannuronolactone............. 126 213 145 152Lactone spot ................. 127 212 145 153Galacturonic acid ............. 29 32 17 36Uronic acid spott ............ 27 33 17 Tailing
* Results are expressed as Rglucose values.t RG values for glucuronic acid were similar to those for the uronic acid spot.
not form a lactone, whereas glucuronic andmannuronic acids do (Percival, 1963), the pres-ence of more than one uronic acid was investi-gated. Results of the thioglycolic-sulfuric acidtest in the presence of mannose (Dische, 1962)indicated that glucuronic acid was not part ofthe polymer. Figure 3 illustrates the absorptionspectra of the acidic fraction and authenticglucuronic and galacturonic acid. UnhydrolyzedSP also gave a spectral curve similar to that ofgalacturonic acid.The hydroxamic acid spray (Abdel-Akher and
Smith, 1951) was utilized to identify a component
0.4 A
GALACTURONIC TESTA-polysac. 3A
B/\ \ B = golacturonic
0.3 _ C-polysoc. fractionO D C D glucuronic
0.2
0.1
450 535 650
MryFIG. 2. Absorption spectra for intact extracellular
polysaccharide of strain SA, galacturonic acid, anacidic fraction isolated from the polysaccharidehydrolysate, and glucuronic acid, in the modifiedcarbazole test for galacturonic acid.
0.6
0.4GLUCURONIC TEST
A= alucuronic0 D
U. .%Pill%,
B=galacturon i c
C=polysac. fraction
0.2
450 500 550
my
FIG. 3. Absorption spectra for glucuronic acid,galacturonic acid, and an acidic fraction isolatedfrom an extracellular polysaccharide hydrolysate ofstrain SA, in the thioglycolic acid test for glucuronicacid.
found in acid hydrolysates as a lactone. Infraredanalysis of the isolated lactone indicated charac-teristic absorption peaks for the carboxylate ion(6.25 and 7.1 ,u). The lactone was present onpaper chromatograms irrigated in all of the
333
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

COHEN AND JOHNSTONE
chromatographic techniques, as well as identicalA;\ color reactions with developing sprays, showed
this component to be identical with rhamnose.0.4 Hexose was indicated on the basis of chro-
matographic evidence (Table 1) and the primary
B \ METHYL PENTOSE TEST cysteine-sulfuric acid method of Dische et al.A= polysoc. 3A (1949). Figure 5 illustrates identical absorptionB= rhamnose spectra for a hexose isolated from the neutralC= pol ysac. f r a ct ion fraction of an acid hydrolysate and an authentic
0.3 sample of D-glucose. The isolated hexose frac-tion was identified by paper chromatographywith authentic D-glucose (Table 1). Since glucose
0 D and galactose have similar RF values in imostsolvents, cochromatography, multiple ascending(solvent A), and descending (solvent B) paperchromatographic techniques were utilized to
0.2 separate these two sugars. Furthermore, theC/c \ \ \ isolated fraction gaxve a p)ositive glucose oxidase
test (Salton, 1960), which strongly indicated[a]D-glucose.Thus far, the data presented have been for
\\ SP 3A, unless otherwise noted. However, by use370 400 450 of the above methods, results were obtained for
MPFIG. 4. Absorption spectra for intact extracellu lar 0.6
-
polysaccharide of strain 3.4, rhamnose, and a nmethylpentose isolated from the neutral fraction of thepolysaccharide hydrolysate, in the cysteine-suilfuiricacid test for methyl pentose. A
solvent systems. When the lactone was eluted 0from paper and rechromatographed in solvents / BA, B, C, or D, it dissociated into two compo-nents, one of which moved at the same iate asthe lactone and a second slower-migrating com- 0 Dponent. Eluates of this second spot gav-e apositive carbazole test (Dische, 1962) for hexu-ronic acid and a negative modified carbazole test 0.2 - HEXOSE TEST(Dische, 1962) for galacturonic acid. In addition, A=glucosethe lactone migrated slower than glucurono- B=polysoc. fractionlactone in all solvents used and appeared toresemble closely the RF values of mannurono-lactone (Table 1).Methyl l,entose was indicated on the basis of
chromatographic evidence (Table 1) and the _L-eysteine-sulfuric acid test of Dische and 350 400 450Shettles (1948). Figure 4 illustrates identical m ,absorption spectra for unhydrolyzed SP, a FIG. 5. Absorption spectra for glucose, and amethyl pentose isolated from the neutral fraction hexose isolated fromii the neutral fraction of a hy-of an acid hydrolysate, and an authentic sample drolysate of extracellullar polysaccharide from strainof L-rhamnose. Rates of movement in four solvent 3A, in the primlary cysteine-sulfuiric acid test forsystems, by ascending, descending, and co- hexose.
334 J. BACTERtIOL.
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
lVOL. 88, 1964 EXTRACELLULAR POLYSACCHARIDES OF A. VINELANDII
SP 102 and 155 identical to those obtained forSP 3A. Consequently, the polysaccharides ofA. vinelandii under investigation had the fourmajor components listed above in common.
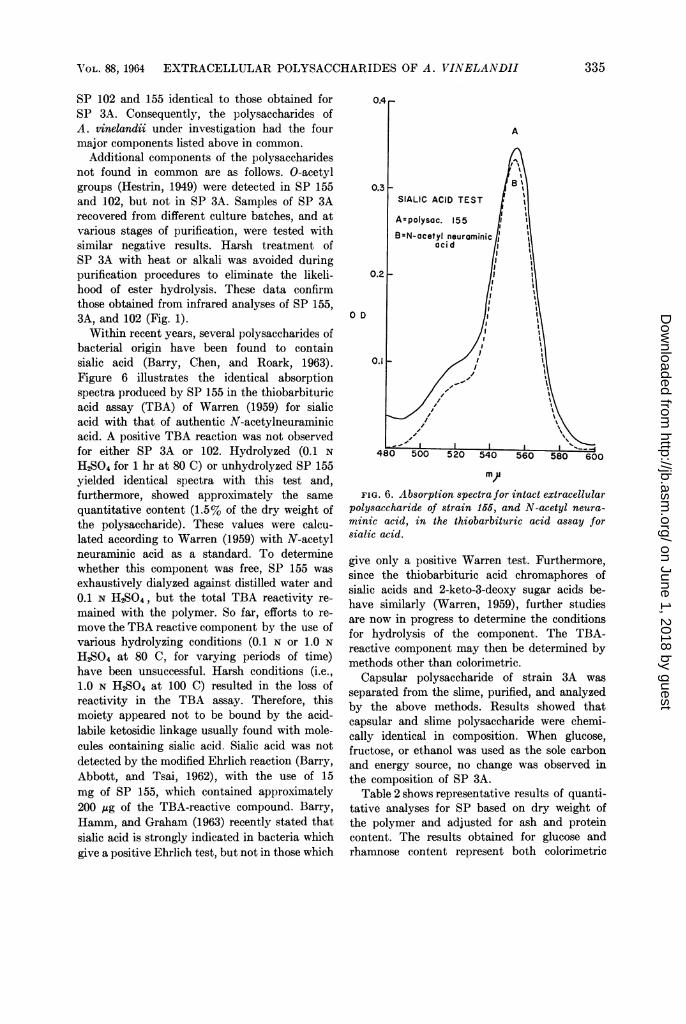
Additional components of the polysaccharidesnot found in common are as follows. O-acetylgroups (Hestrin, 1949) were detected in SP 155and 102, but not in SP 3A. Samples of SP 3Arecovered from different culture batches, and atvarious stages of purification, were tested withsimilar negative results. Harsh treatment ofSP 3A with heat or alkali was avoided duringpurification procedures to eliminate the likeli-hood of ester hydrolysis. These data confirmthose obtained from infrared analyses of SP 155,3A, and 102 (Fig. 1).Within recent years, several polysaccharides of
bacterial origin have been found to containsialic acid (Barry, Chen, and Roark, 1963).Figure 6 illustrates the identical absorptionspectra produced by SP 155 in the thiobarbituricacid assay (TBA) of Warren (1959) for sialicacid with that of authentic N-acetylneuraminicacid. A positive TBA reaction was not observedfor either SP 3A or 102. Hydrolyzed (0.1 NH2SO4 for 1 hr at 80 C) or unhydrolyzed SP 155yielded identical spectra with this test and,furthermore, showed approximately the samequantitative content (1.5% of the dry weight ofthe polysaccharide). These values were calcu-lated according to Warren (1959) with N-acetylneuraminic acid as a standard. To determinewhether this component was free, SP 155 wasexhaustively dialyzed against distilled water and0.1 N H2SO4, but the total TBA reactivity re-mained with the polymer. So far, efforts to re-move the TBA reactive component by the use ofvarious hydrolyzing conditions (0.1 N or 1.0 NH2SO4 at 80 C, for varying periods of time)have been unsuccessful. Harsh conditions (i.e.,1.0 N H2SO4 at 100 C) resulted in the loss ofreactivity in the TBA assay. Therefore, thismoiety appeared not to be bound by the acid-labile ketosidic linkage usually found with mole-cules containing sialic acid. Sialic acid was notdetected by the modified Ehrlich reaction (Barry,Abbott, and Tsai, 1962), with the use of 15mg of SP 155, which contained approximately200 ,ug of the TBA-reactive compound. Barry,Hamm, and Graham (1963) recently stated thatsialic acid is strongly indicated in bacteria whichgive a positive Ehrlich test, but not in those which
O D
0.3
0.2
0.I
A
SIALIC ACID TEST
A=polysac. 155
m)jFIG. 6. Absorption spectra for intact extracellular
polysaccharide of strain 155, and N-acetyl neura-minic acid, in the thiobarbituric acid assay forsialic acid.
give only a positive Warren test. Furthermore,since the thiobarbituric acid chromaphores ofsialic acids and 2-keto-3-deoxy sugar acids be-have similarly (Warren, 1959), further studiesare now in progress to determine the conditionsfor hydrolysis of the component. The TBA-reactive component may then be determined bymethods other than colorimetric.
Capsular polysaccharide of strain 3A wasseparated from the slime, purified, and analyzedby the above methods. Results showed thatcapsular and slime polysaccharide were chemi-cally identical in composition. When glucose,fructose, or ethanol was used as the sole carbonand energy source, no change was observed inthe composition of SP 3A.
Table 2 shows representative results of quanti-tative analyses for SP based on dry weight ofthe polymer and adjusted for ash and proteincontent. The results obtained for glucose andrhamnose content represent both colorimetric
335
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

COHEN AND JOHNSTONE
TABLE 2. Chemical analyses of extracellular polysaccharides of Azotobacter vinelandiia
Strain Nitrogenb Carbohydrateb Galacturonic Glucose b, c Rhamnoseb,c O-acetylb Ratiodacidb ai
3A 0.14 50.5 41.1 1.2 1.4 Not 41:1:1detected
102 0.76 45.5 37.9 1.5 1.7 2.9 38:2:2155 0.38 46.0 49.5 4.1 0.9 6.5 50:4:1
a All figures are calculated on a dry weight basis adjusted for ash and protein.Analysis performed on intact polysaccharides.
c Analysis performed on acid hydrolysatesd Per cent ratio of galacturonic acid-glucose-rhamnnose.
analyses on the intact polymer and quantitativepaper chromatography, since these methodsyielded virtually identical results.Attempts were made to account for the re-
mainder of the dry weight of SP. Various con-ditions for acid hydrolysis were employed in con-junction with sugar detection methods; however,only the four reported components were observed.Hexosamine was not found in the polysacca-rides investigated. Furthermore, Kjeldahl nitro-gen in SP was accounted for as l)rotein by themethod of Lowry et al. (1951).
DISCUSSION
These studies confirm, in part, the results ofKaufman and Rapaske (1958), and Kaufman(1960), who indicated that the capsular l)oly-saccharide consisted of glucose and rhamnoseunits. The evidence reported here, however,indicates that galacturonic acid is also l)resentand is a major component of the lpolysaccharide.In addition, a hexuronic acid lactone, p)robablymannurono-lactone, is also present in acid hy-drolysates. Whereas Kaufman (1960) reportedthat capsules and slime possess different chemi-cal coml)osition, the present study shows themto possess the same four major components.This is consistent with Wilkinson's (1958) state-ment that if cells produce both capsules andslime, they are of similar eomposition. Above-mentioned differences may have been due inpart to the resistance of the polysaccharide toacid hydrolysis. For example, after 5 hr of hy-drolysis with 2 N H2SO4, only traces of uronicacid wAere detected by paper chromatography.However, colorimetric methods of Dische forsugar analysis on intact p)olysaccharide providedevidence for the composition of A. vinelandii
polymers not easily detected by paper chroma-tography.
In addition to the four major componentscommon to the strains, o-acetyl was found in onlytwo of the three strains, one of which alsocontained a thiobarbituric acid assay (Warren,1959) -positive component. Therefore, it appearsthat the polysaccharides are similar in manyrespects, but are not identical.
Although evidence suggests that the lactoneis mannurono-lactone, further studies are neededfor confirmation. The lactone resembles theunknown component, Xl, described by Zaitseva,Belozerskii, and Afanas'eva (1959), who studiedthe gross lpolysaccharide content of -4. vinelandii.If, indeed, two uronic acid residues are lpresent,it would represent one of the few such reportsfor bacterial extracellular lpolysaccharides. Aero-bacter aerogenes type 64 extracellular polysac-charide was found to contain glueuronic andmannuronic acid (Barker et al., 1958).Two components, glucose and rhamnose, were
found in common with cyst polysaccharide (Wysset al., 1961). Further work is needed to deter-mine whether cyst polysaccharide does, in fact,contain all of the components shown here inextracellular polysaccharide of the vegetativecell.
ACKNOWLEDGMENTS
We wish to thank the Department of Regula-tory Chemistry, University of Vermont, for theinfrared analyses, and N. J. Calvanico for adviceon disc electrophoresis.
LITERATURE CITED
ABDEL-AKHER, M., AND F. SMITH. 1951. The de-tection of carbohydrate esters and lactones
336 J . BATCTER{IOL .
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

VOL. 88, 1964 EXTRACELLULAR POLYSACCHARIDES OF A. VINELANDII
after separation by paper chromatography.J. Am. Chem. Soc. 73:5859-5860.
BARKER, S. A., A. B. FOSTER, I. R. SIDDIQUI, ANDM. STACEY. 1958. Structure of an acidic poly-saccharide elaborated by Aerobacter aerogenes.Nature 181:999.
BARRY, G. T., V. ABBOTT, AND T. TSAI. 1962. Re-lationship of colominic neuraminic acid. J.Gen. Microbiol. 29:335-352.
BARRY, G. T., F. CHEN, AND E. ROARK. 1963.Isolation of N-acetyl neuraminic acid and4-oxo-norleucine from a polysaccharide ob-tained from Citrobacter freundii. J. Gen. Mi-crobiol. 33 :97-116.
BARRY, G. T., J. D. HAMM, AND M. G. GRAHAM.1963. Evaluation of colorimetric methods inthe estimation of sialic acid in bacteria.Nature 200:806-807.
BULEN, W. A., AND J. R. LECOMTE. 1962. Isolationand properties of a yellow-green fluorescentpeptide from azotobacter medium. Biochem.Biophys. Res. Commun. 9:523-528.
COHEN, G. H., AND D. B. JOHNSTONE. 1963a. Acidproduction by Azotobacter vinelandii. Nature198:211.
COHEN, G. H., AND D. B. JOHNSTONE. 1963b. Theinfluence of the extracellular polysaccharideslime of Azotobacter vinelandii on pH. Bac-teriol. Proc., p. 11.
DAVIS, B. J. 1963. Disc electrophoresis. II. Mate-rial and methods. Distillation Products In-dustries, Rochester, N.Y.
DISCHE, Z. 1962. Color reactions of hexuronicacids, p. 497-501. In R. L. Whistler and M.L. Wolfrom [ed.], Methods in carbohydratechemistry, vol. 1. Academic Press, Inc., NewYork.
DISCHE, Z., AND L. B. SHETTLES. 1948. A specificcolor reaction of methylpentoses and a spec-trophotometric micromethod for their deter-mination. J. Biol. Chem. 175:595-603.
DISCHE, Z., L. B. SHETTLES, AND M. OSNOS. 1949.New specific color reactions of hexoses andspectrophotometric micromethods for theirdetermination. Arch. Biochem. 22:169-184.
DUBOIS, M., K. A. GILLES, J. K. HAMILTON, P. A.REBERS, AND F. SMITH. 1951. A colorimetricmethod for the determination of sugars. Na-ture 168:167.
EKLUND, C., AND 0. WYSS. 1962. Enzyme associa-ted with bacteriophage infection. J. Bacteriol.84:1209-1215.
FISKE, C. H., AND Y. SUBBAROW. 1925. The colori-metric determination of phosphorus. J. Biol.Chem. 66:375-400.
FLODIN, P. 1962. Dextran gels and their applica-tion in gel filtration. Dissertation, A B Phar-macia, Uppsala, Sweden.
HESTRIN, S. 1949. The reaction of acetyl cholineand other carboxylic acid derivatives withhydroxylamine and its analytical application.J. Biol. Chem. 180:249-261.
JOHNSTONE, D. B. 1962. Azotobacter agilis or A.vinelandii? Soil Microbiological MethodsNewsletter 1:6-8.
KAUFMAN, B. 1960. The physiology of carbohy-drate formation by Azotobacter vinelandii.Ph.D. Thesis, Indiana University, Blooming-ton.
KAUFMAN, B., AND R. REPASKE. 1958. Polysac-charide synthesis by Azotobacter vinelandii.Bacteriol. Proc., p. 126.
LEVINE, S., H. J. R. STEVENSON, AND P. W. KAB-LER. 1953. Qualitative studies of pneumo-coccal polysaccharides by infrared spectro-photometry. Arch. Biochem. Biophys. 45:65-73.
LOWRY, 0. H., N. J. ROSEBROUGH, A. L. FARR,AND R. J. RANDALL. 1951. Protein measure-ment with the Folin phenol reagent. J. Biol.Chem. 193:265-275.
MEJBAUM, W. 1939. tYber die Bestimmung kleinerPentosemengen, insbesondere in Derivatender Adenylsaiure. Z. Physiol. Chem. 258:117-120.
NELSON, N. 1944. A photometric adaption of theSomogyi method for the determination ofglucose. J. Biol. Chem. 153:375-380.
PARTRIDGE, S. M. 1949. Aniline hydrogen phtha-late as a spraying reagent for chromatographyof sugars. Nature 164:443.
PERCIVAL, E. 1963. Aldonic, uronic, oxoaldonicand ascorbic acids, p. 67-112. In M. Florkinand E. H. Stotz [ed.], Comprehensive bio-chemistry, vol. 5. Elsevier Publishing Co.,New York.
RACUSEN, D., AND D. B. JOHNSTONE. 1961. Estima-tion of protein in cellular material. Nature191:492-493.
SALTON, M. R. J. 1960. Specific detection of glucoseon paper chromatograms. Nature 186:966-967.
SEVAG, M. G., D. B. LACKMAN, AND J. SMOLENS.1938. The isolation of the components ofstreptococcal nucleoproteins in serologicallyactive form. J. Biol. Chem. 124:425-436.
SMITH, F., AND R. MONTGOMERY. 1959. The chem-istry of plant gums and mucilages. ReinholdPublishing Corp., New York.
SMITH, I. 1958. Sugars, p. 164-177. In I. Smith[ed.], Chromatographic techniques-clinicaland biochemical applications. IntersciencePublishers, Inc., New York.
SOMOGYI, M. 1952. Notes on sugar determination.J. Biol. Chem. 195:19-23.
WARREN, L. 1959. The thiobarbituric acid assayof sialic acids. J. Biol. Chem. 234:1971-1975.
337
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

338 COHEN ANI
WHISTLER, R. L. 1959. Factors influencing gumcosts and applications, p. 1-13. In R. L. Whist-ler and J. N. BeMiller [ed.], Industrial gums.Academic Press, Inc., New York.
WILKINSON, J. F. 1958. The extracellular poly-saccharides of bacteria. Bacteriol. Rev. 22:46-73.
WILSON, P. W., AND S. G. KNIGHT. 1952. Experi-ments in bacterial physiology. Burgess Pub-lishing Co., Minneapolis.
WYSS, O., M. G. NEUMANN, AND M. D. SOCOLOF-
JOHNSTONE J. BACTERIOL.
SKY. 1961. Development and germination ofthe azotobacter cyst. J. Biophys. Biochem.Cytol. 10:555-565.
ZAITSEVA, G. N., A. N. BELOZERSKII, AND T. P.AFANAS'EVA. 1959. Chemistry of azotobacter.VII. A study of the polysaccharides of threespecies of Azotobacter in relation to the com-position of the medium and the source of ni-trogen feeding. Microbiology USSR (EnglishTransl.) 28:52-57.
3 i
on June 1, 2018 by guesthttp://jb.asm
.org/D
ownloaded from