extra-toes: a new mutant gene causing multiple ...malformations of the brain, central nervous system...
TRANSCRIPT

/ . Embryol. exp. Morph. Vol. 17, 3, pp. 543-81, June 1967 5 4 3With 3 plates
Printed in Great Britain
Extra-toes: a new mutant gene causing multipleabnormalities in the mouse
ByD. R.JOHNSON
M.R.C. Experimental Genetics Research Unit,Department of Animal Genetics, University College London
INTRODUCTION
The present paper deals with the genetics, morphology and developmentalanatomy of a new mutant gene in the mouse called extra-toes. Extra-toes(symbol Xt) derives its name from the fact that in the heterozygous conditionthere are extra digits on the preaxial side of both hind feet. Apart from this (andcorresponding changes in the forefeet) heterozygotes are externally normal. Thehomozygote dies at birth or in utero with a wide range of abnormalities. Theseinclude oedema, paddle-shaped feet with up to eight digits, hemimelia and grossmalformations of the brain, central nervous system and sense organs.
ORIGIN
Extra-toes arose in the control series of an irradiation experiment at the M.R.C.Radiobiological Research Unit at Harwell. Five Xtj + <$$ and two Xt\ + $$ weresent to the Animal Genetics Department at University College London inNovember 1962. Two of the $$ were mated with the ??, and the remaining threewith $? from the CB stock (the F1 of CBA/Gr and C57BL/Gr). An Xt\ + $ wasoutcrossed to two CB $$ in November 1963. With these exceptions the stock hasbeen brother/sister mated throughout.
GENETICS
Extra-toes is a semi-dominant gene with almost complete manifestation in theheterozygote and few normal overlaps. All four feet are usually affected. It isan embryonic or neonatal lethal when homozygous. This is reflected in thesegregation figures (Table 1). In backcross matings (1) the number of XtjXtanimals recovered was 45 whereas the expected number was 261 (i.e. XtjXt and+ /+ should be found in equal numbers). The numbers of Xt/+ and + / +animals are close to the expected 2:1 ratio. In outcross matings (2) the figures arein good agreement with the expected 1:1 ratio.
The deficiency of XtjXt animals in backcross matings together with the de-
Author's address: Department of Animal Genetics, University College, Gower Street,London, W.C.I, U.K.

544 D. R. JOHNSON
creased litter size suggest that homozygous abnormals are lost before or at birth.As Xt homozygotes can be identified from the 9th day of gestation onwardsviability may be studied directly. Table 2 gives details of 88 litters of embryosrecovered from backcross matings. The mean viability of XtjXt embryos is0-72, suggesting that a number ofXt/Xt zygotes die before they can be recognized.The number of homozygotes recovered at birth, however, is only 0-17 of theexpected total. The discrepancy between these figures reflects the nest tidyingactivity of the mother, who often consumes stillborn young.
Lyon, Morris, Searle & Butler (1967) have shown that Xt is situated inlinkage group XIV, near to the gene for crinkled (cr).
Table 1. The segregation of Extra-toes at birth
Matingtype /+ Xt/+ Xt/Xt
Unclass-ified Total
Meanlittersize
12
3
Xt/+xXt/ +Xt/+x+/ ++ /+xXt/++ / + x +/ +
261508
21
548522
—
45—
—
625
—
8601055
21
4-97-2
7 0
Table 2. Segregation of Extra-toes in litters of embryos
Age(days) Xt/ +
SolidXt/Xt moles Total
MeanNo. of litter Viabilitylitters size of Xt/Xt
910111213
14-18Total
1252
57586874
451
3496
191421158
31
108
35773
35
60
7977969657
214
619
111213159
28
88
7-16-47-46-46-37-6
7 0
0-970-720-920-610-520-630-72
MATERIAL AND METHODS
Xtj + $S and $$ were put together at 5 p.m. The females were examined forvaginal plugs at 9 a.m. the next morning. The date of the vaginal plug was takenas day 0 of gestation. Pregnant animals were killed with ether and their uteriremoved. If the pregnancy was in its 1 lth day or less the whole of the uterus wasfixed in Bouin's fluid and the embryos dissected out 24 h later. Older embryoswere dissected out in saline before fixation. Developmental age was checkedagainst the criteria of Griineberg (1943).
Sectioned material is listed in Table 3. It was not considered necessary tosection whole litters beyond the 9-day stage when homozygous abnormal ani-mals can be readily identified externally. Therefore embryos were sectioned inmatched pairs or trios of litter-mates. All material was dehydrated and double

Extra-toes: a new mutant 545embedded by the technique of Peterfi. Sections were cut at 5-10 jtc according toage. The heads of newborn and older mice were decalcified in 4 % nitric acid in90 % alcohol before dehydration and sectioned at 15 /i. All sectioned materialwas stained in Ehrlich's haematoxylin and eosin.
The skeletons of extra-toes mice and their normal litter-mates were studied asmethylene blue and alizarin red clearance preparations (Table 4). Methyleneblue preparations were made by Noback's modification of Van Wijhe's methodaccording to Griineberg's (1953) protocol. Alizarin red preparations were madeby a modification of the technique of Dawson (1926).
Table 3. Serial sections of embryos and young mice
Age(days)
8*8#99i
10111213141
161
Newborn*Newbornf
+ / +
002
19147
330282
14 days post partum* 4Total
* Heads only, f A
Table 4.
Type of preparation
Methylene blue
Alizarin red
91
Xt/ +
4416705
series of excised organs.
Materials used in
Age (days)
1518
Newborn4 days post
partumNewborn
4 days postpartum
the study
+ /+
3134
63
29
Xt/Xt
001
1596751252
—
53
Unclass-
oftheXt
Xt/ +
6173
135
29
ifiable
81821000000000
29
skeleton
Xt/Xt
618
—
3——
Total
8185
35231314122
102049
173
Total
153
187
228
58
THE Xtj + MOUSE
External anatomy
The extra-toes heterozygote is fully viable and breeds freely. It is of the samesize and weight as its normal litter-mates: at birth it is slightly larger and heavier

546 D. R. JOHNSON
Table 5. Birthweights of 231 animals from the Xt stock
Xt/ +
No. Mean wt. (g) No. Mean wt. (g)
1 Xt/+xXt/ +2 Xt/+x+/+\
+ / + xXt/+)
48 1-365 ±0053 98 1-434 ±0-028
40 1-432 ±0-032 45 1-501 ±0051
"1 cm
Text-fig. 1. Plantar views of the feet of + / + and Xt/ + mice. A, + / + right forefoot;B-D, Xt/+ right forefeet; E, + /+ right hind foot; F-L, Xt/+ right hind feet.
(Table 5). Classification is possible from birth onwards by the presence of extrapre-axial digits on the hind feet.
Plantar views of the feet of mice from the Xt stock are shown in Text-figure 1.The forefoot of the normal mouse (A) carries four clawed toes and a pollexrepresented by a small knob of tissue bearing a nail. In Xt/ + mice (B-D) thepollex is enlarged or duplicated. Digit II may carry a double claw. On the post-axial side of the foot a small lump often appears opposite the base of digit V,which may also carry a double claw.

Extra-toes: a new mutant 547In the hind foot (F-L) a sixth or even a seventh toe is added preaxially. The
form of the extra digit is variable. In low grades the hallux is simply thickenedand may or may not carry a double claw. Progressively more abnormal micehave a duplicated hallux whose constituents are united by soft tissue syndactylism,or two or three entirely separate toes. Neither postaxial nubbins nor duplicationsof the claw of digit V has been observed in adults: the hind feet of newbornXt\ + mice, however, sometimes carry a post-axial rudiment which regresses inthe first few days of life.
Belly spots
The original Xtl + mice obtained from Harwell all had white belly spots. Thischaracteristic has persisted in the present Xt stock (Table 6). The largest bellyspots seen are roughly circular, approximately 1 cm in diameter and situated inthe midline. All grades between this and-a few white hairs only have been seen.In some cases the spot is elongated to form a streak. One Xtl + <$ developed anasymmetrical head blaze late in life, but apart from this no head dots have beenseen.
Table 6. Belly spotting in the Xt stock
Spot
18459-2%
Table 7. The
Xt/ +
No spot
12740-8%
Total
311
size and occurrence
Size of interfrontalbone
Xt/ +
0
151
Spot
72-9%
+ / +
No spot
23797-1%
of the interfrontal bone in
+ + +
5 911 8
+ + +
09
Xt/ +
Total
2929
Total
244
mice
* m j „ The axial skeleton1. The skull
The only abnormality seen in the Xt/ + skull is the presence of an inter-frontal bone (Truslove, 1952) in a high proportion of cases (Table 7). In normallitter-mates an interfrontal was present less frequently; where both members ofa pair of litter-mates had an interfrontal bone it was larger in the Xtl + mouse inall cases save one, where the bones were of equal size.
2. Hydrocephaly
Six hydrocephalic mice have been recorded in the Xt stock to date. All wereXtl + a n d occurred in one family. Xt 52/55 produced 4 known hydrocephalicsin 43 offspring. Of the 39 non-hydrocephalics 3 pairs were brother/sister matedand produced 1/25, 1/58 and 0/60 abnormals respectively.

548 D. R.JOHNSON
3. The vertebral column and thorax
A vertebral count of 29 Xtj + mice and their normal litter-mates shows thatthe total number of presacral vertebrae in Xtj + and + / + mice is identical.There is no shift in the lumbo-sacral border as seen in some other polydactylousmutants. The ribs and sternebrae of Xtj + mice are normal.
The appendicular skeleton
1. The cartilaginous skeleton
In the normal embryo of 15 days the cartilaginous precursors of all the limbbones can be seen (Text-fig. 2 A). There is some ossification of the long bones andthe limb girdles.
Text-fig. 2. Forelimbs of 15-day-old embryos. A, + / + right; B, Xtj+ right; C,Xt/+ left. Camera-lucida drawings of methylene-blue clearance preparations.
In the forelimbs the Xt heterozygote shows a series of duplications of the firstdigit and the first metacarpal (Text-fig. 2B, C). In more extreme cases digit IImay also be partially duplicated. The small supernumerary digits which some-times appear at the base of digit V have a single rod-like skeletal element.
In the hind limb (Text-fig. 3) duplications of the phalanges and metatarsal ofdigit I can be seen. The cartilaginous precursor of the incomplete metatarsal orsplint bone, which is usually found on the preaxial side of metatarsal II is alsopresent.
2. The osseous skeleton
The abnormalities of the osseous skeleton of the forelimbs of Xtj + mice donot correspond exactly with the pattern seen in their cartilaginous precursors.It is clear from examination of Text-figs. 4 and 5 and comparison with Text-fig. 2that the situation has been complicated by tertiary distal fusion of the osseous

Extra-toes: a new mutant 549elements on the preaxial side of the foot. In Text-fig. 5B, for example, fusion hasoccurred between the claws of both parts of digit I and between this doubleelement and the claw of the preaxial part of digit II. As the anatomy of the fore-foot becomes more abnormal these fusions make interpretation more difficult.
In Text-fig. 4B and D there is fusion of the Os centrale with the Os multangu-lum minus. Postaxial accessory ossifications (A, Text-fig. 4D) were present in10 out of 29 preparations examined. In four cases they occurred bilaterally,whilst six mice had a unilateral occurrence on the right side. The converse, withthe element present on the left side only, did not occur in the sample.
1 mm
Text-fig. 3. Left hind limbs of 15-day-old embryos. A, +/+;B, Xt/+.
The lowest grade of abnormality seen in the hind feet of Xtj + mice is thepresence of a small splint of bone adjacent to the base of Os metatarsale II(Text-fig. 6). This may be present alone in a foot which externally looks quitenormal or may be associated with other abnormalities. It may occur as a separateentity or be fused with the Os tarsale primum or (less frequently) the Os meta-tarsale secundum. The Os tarsale primum is thickened in low grades of poly-dactyly and may bear a pre-hallux consisting of a single phalanx and claw;alternatively the metatarsal may be completely duplicated (Text-fig. 7C). Inmore extreme expressions it is the more medial of the two Ossa metacarpaliaprima which divides or tends to divide to form a seventh digit. The Os tarsaletertium may be fused with the Os tibiale; this condition tends to be restrictedto feet with a high grade of polydactyly (Text-fig. 7B, C). No other fusions wereobserved amongst the tarsalia.
Hind-foot polydactylism in Xtj+ is remarkably symmetrical (Table 8):62 % of the animals classified were symmetrically affected, and in cases ofasymmetry the right exceeded the left as often as vice versa.

550 D. R. JOHNSON
3. Hemimelia in heterozygotes
This has been seen in the Xt stock upon two occasions. In the first case,a $, there was total absence of the tibia and a reduced bowed fibula in the lefthind limb. In the foot digit I was absent. The right hind limb was normal withfive toes. The forefeet were typical of Xtj + mice, with preaxial polydactyly. The
C+tA-
Text-fig. 4. Left forefeet of + / + (A) and Xt/+ (B-D) mice. Extensor surface, 28days old. Camera-lucida drawings of alizarin clearance preparations. A, Accessorypostaxial digit. C+ M, Fused centrale and multangulum minus.

Extra-toes: a new mutant 551
Table 8. Symmetry of hind foot polydactyly at weaning in Xt/+ mice
L only
30-45 %
L>R
10017-4%
L = R
35762-1%
R > L
11319-6%
R only
30-45%
Total
576
0-25 cm
Text-fig. 5. Left forefeet of + /+ (A) and Xt/+ (B-D) mice. Lateral view.
vertebral column was abnormal with some wedge-shaped vertebrae andfusions. There was marked scoliosis in the lumbar region and multiple fusionsof the sacral vertebrae. The sternum was twisted, with irregular rib insertions.Genetically the mouse proved to be Xtj +. Mated with an Xtj + brother sheproduced 43 offspring (8 XtjXt, 24 Xtj + t 11 + / + ). None of the Xtj+ nor+ / + were hemimelic. She was killed at 450 days, and an alizarin red clearancepreparation was made.
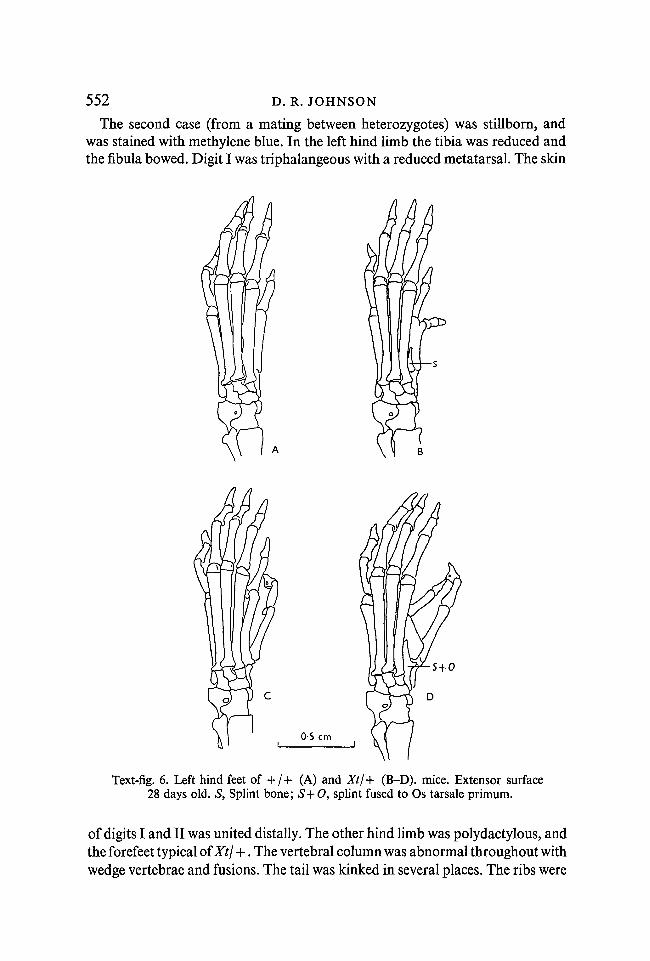
552 D. R. JOHNSON
The second case (from a mating between heterozygotes) was stillborn, andwas stained with methylene blue. In the left hind limb the tibia was reduced andthe fibula bowed. Digit I was triphalangeous with a reduced metatarsal. The skin
s+o
Text-fig. 6. Left hind feet of + / + (A) and Xt/+ (B-D). mice. Extensor surface28 days old. S, Splint bone; S+O, splint fused to Os tarsale primum.
of digits I and II was united distally. The other hind limb was polydactylous, andthe forefeet typical of Xtj +. The vertebral column was abnormal throughout withwedge vertebrae and fusions. The tail was kinked in several places. The ribs were

Extra-toes: a new mutant 553not inserted opposite each other on the sternum. The parents produced twenty-three other offspring, none of which was hemimelic.
0 + T
Text-fig. 7. The left (A, B) and right (C, D) hind feet of Xt/ + mice. O + T, Fused Ostibiale and Os tarsale tertium.
THE XtjXt MOUSE
External anatomy
Perhaps the most striking characteristic of the newborn XtjXt mouse, apartfrom its grossly abnormal limbs, is the retention of embryonic posture (Text-fig. 8). The normal mouse embryo has lost the C-shaped curve of its vertebral
35 J EE M 17

554 D. R. JOHNSON
column by the 18th day of gestation. The column then becomes S-shaped by aflexure appearing in the cervical region. This is even more pronounced at birth.In the extra-toes homozygote, however, the C-shape is retained and evenaccentuated. There is often a parietal brain hernia and ectopia of theviscera. The skin is less wrinkled than normal. The limbs carry eight or ninedistinct toes, always webbed. Supernumerary papillae on the forehead carryhairs. The abnormal arrangement of the mystacial vibrissae is shown in Text-figure 9.
One or two animals with neither pronounced ectopia nor protruding brainshave been born alive. They suffered respiratory distress and lived for only a fewminutes.
5 mm 2 mm
Fig. 8 Fig. 9
Text-fig. 8. Camera-lucida drawings of newborn mice. Left, + / + ; Right, Xt/Xt.Left fore- and hind-feet of Xt/+ litter-mate.
Text-fig. 9. Arrangement of mystacial vibrissae in newborn + / + (left) and Xt/Xt mice.
The viscera
Dissection of six newborn homozygotes (2 $$, 4 $$) revealed the followingvisceral abnormalities. The adrenal glands were displaced caudal so as to liemedial to the kidney. In one female the left adrenal gland was double.In all four females the right ovary was attached to the lateral side of theright kidney, whence the oviduct/uterus ran normally. In two animals theleft ovary lay beside the left adrenal gland. Males were not correspondinglyaffected.
In all cases the kidneys were irregular in outline with depressions seeminglycaused by the close proximity of other organs. For example, all kidneys hadindentations in the region lying adjacent to the bladder.
The following organs were removed from newborn homozygotes, sectioned

Extra-toes: a new mutant 555and compared to normal controls: adrenal gland, gonad, heart, kidney, liver,lung, spleen, stomach and thymus. No histological abnormalities were detected.
The brain
A single glance is enough to show the gross abnormality of the XtjXt brain(Text-figs. 10,11). In dorsal view the whole brain is twisted about its major axis.The olfactory lobes are absent. The cerebral hemispheres are small and partially
C£
Fig. 10 Fig. 11
Text-fig. 10. Dorsal and ventral views of the brains of newborn mice. A, Normaldorsal; B, Xt/Xt dorsal; C, normal ventral; D, Xt/Xt ventral. CE, cerebellum; CH,cerebral hemisphere; CQ, corpora quadrigemina; D, diencephalon; IN, infundibu-lum; MO, medulla oblongata; Nl, olfactory nerve; Nil, optic nerve; OL, olfactorylobe.
Text-fig. 11. Lateral views of the brains of newborn mice. A, Normal; B, Xt/Xt. Forabbreviations see Text-fig. 10.
hidden beneath the bulging mesencephalon. The pineal body is missing; thecorpora quadrigemina are approximately normal in size but abnormally dividedlaterally. The medulla oblongata is twisted and the cerebellum retains its un-differentiated folded form. Ventrally the optic chiasma is ill-defined (or absent in
35-2

556 D. R. JOHNSON
some cases). It is clear from Text-fig. 11 that the first flexure of the brain whichnormally runs forwards at an angle is almost vertical in abnormal animals,dividing the brain into two distinct parts.
Sections confirm the observations made externally. The cerebral hemispherescontain groups of pycnotic cells. They are separated by a median fluid-filledspace which extends back as far as the cerebellum, and into which invaginates asmall, superficial choroid plexus.
The nose
The anterior part of the XtjXt nose is laterally compressed between abnormallylarge maxillae and premaxillae (Plate 1, figs. A, B). The dorsal part of the nasalchamber may be almost obliterated, and its ectodermal lining represented onlyby a strand of tissue. The nasal septum is broad and short and the paraseptalcartilages deformed.
The posterior nasal region, in contrast, is broad and flat (Plate 1, figs. C, D).The structure of the nasal cartilages is more complex than usual. The olfactorynerves are enormous. In parasagittal sections they can be seen to pass through thecribriform plate; they then turn cephalad and terminate in a space filled withconnective tissue ahead of the cerebral hemispheres. The only contact betweenbrain and olfactory organ is by a very few fibres which turn caudad at the cribri-form plate and enter the brain individually.
The nasal epithelium itself is abnormal. In Xtj + animals it is folded in someareas, overlying a thickened lamina propria. In the homozygote the process isextended; the epithelium is thrown into a series of folds which often raise itconsiderably from the underlying cartilage.
The eye
At birth the eye of homozygotes may be up to half the normal diameter. Theeyelids are always closed. In more extreme cases the eyes may be represented bya pigmented hollow ball containing a lens rudiment (Plate 1, fig. H), by a pigment
EXPLANATION OF PLATES
Abbreviations. AER, Apical ectodermal ridge; CE, cerebellum; CR, crus commune; ED,endolymphatic duct; F, subarcuate fossa; /, upper incisor; L, lens; LC, lateral semicircularcanal; LSO, sense organ of lateral semicircular canal; M, maxilla; MAC, utricular macula;N, nasal cavity; OE, oedematous area; OL, olfactory lobe of brain; ON, olfactory nerve; P,pigment mass; R, retina; SC, superior semicircular canal; UT, utriculus.
PLATE 1
Figs. A, B. Transverse sections through the anterior part of the noses of normal (A) and Xt/Xt(B) newborn mice.Figs. C, D. Transverse sections through the posterior nasal regions of normal (C) and Xt/Xt(D) newborn mice.Figs. E, F, G, H. Left eyes of newborn mice. Transverse sections. E, + /+ ; F, Xt/+ ; G, H,Xt/Xt.

J. Embryo/, exp. Morph., Vol. 17, Part 3
D. R. JOHNSON

J. Embryol. exp. Morph., Vol. 17, Part 3 PLATE 2
D. R. JOHNSON

Extra-toes: a new mutant 557mass, or they may have disappeared without trace. Compensatory hyperplasiaof the Harderian glands helps to fill the orbit. When the eye is very muchreduced contact with the brain may be lost by atrophy of the optic nerve.
In three out of seven Xt\ + newborn heads sectioned the retinae of both eyeswere folded (Plate 1, fig. F). The folding was also recognizable in better developedXt/Xt eyes (Plate 1, fig. G). The retina here is very thick and surrounds the lensin all sections, there being no pupil.
The ear
Of the three semi-circular canals of the normal labyrinth the posterior is bestrepresented in Xt/Xt (Plate 2, fig. J). It is normal in position and although it maybe small in diameter has a lumen throughout. The superior canal has a lumen onlyposteriorly, where it leaves the crus commune, and anteriorly where it joins theampulla. Between those points it is seen only as a solid strand of tissue or as anon-cartilaginous region of the capsule. The lateral semicircular canal is absentin all cases, being represented only by its sense organ which is abnormallysituated adjacent to the utricular macula (Plate 2, fig. L).
* rr, , „ The axial skeleton1. The skull
The skulls of XtjXt mice are always abnormal to a high degree. The normalchondrocranium at 15 days is shown in Text-fig. 12 A. The method of closure ofthe foramen opticum differs from that described by Griineberg (1953). This seemsto be a minor variant of the genetic background, occurring in the majority ofchondrocrania examined.
In Xt\Xt embryos (Text-fig. 12B) there is a complete division into anterior andposterior parts by the failure of the hyphophysial cartilage and the trabecularregion of the central stem to unite. In the posterior part of the chondrocraniumthe otic capsules are poorly differentiated. The occipital cartilages are heavy andhave an indistinct boundary over the foramen magnum. Anteriorly the alaeorbitales are heavy and help to form a broad pan upon which rest the cerebralhemispheres of the abnormal brain.
2. The cervical vertebrae
In Xt\Xt mice there is a tendency for the neural arches of the cervical vertebraeto fuse with their neighbours (Text-fig. 13). In B this has happened in three placesand the spatulate nature of the unfused neural arch rudiments is apparent. In C
PLATE 2
Figs. I, J. Transverse sections through the ears of newborn normal (I) and Xt/Xt (J) mice at thelevel of the subarcuate fossa. Figs. K, L. Transverse sections through the ears of newbornnormal (K) and Xt/Xt (L) mice at the level of the utricular macula.

558 D. R. JOHNSON
almost all of the cervical vertebrae are involved and each side of the neck forms asolid ridge. The vertebral bodies in all cases are quite normal with no signs offusion.
2 mm
Text-fig. 12. Dorsal views of the chondrocranium of a normal (A) and an XtjXtembryo (B), 15 days old. A, Ala orbitalis; FM, foramen magnum; FO, foramenopticum; M, Meckel's cartilage; O, otic capsule; P, parachordal cartilage; PA,processus alaris; PN, paranasal cartilage; PT, parieto-tectal cartilage; 5", supra-occipital cartilage.
otSSS
V ' • ' " 'l''*
323
1 mm
Text-fig. 13. Dorsal views of the cervical vertebrae of a normal (A) and two Xt/Xtembryos (B, C) 15 days old.
3. The sternum
Text-fig. 14 shows the ventral aspect of the thorax of a normal mouse and twoXtjXt litter-mates. Their developmental age is a little over 15 days as the sternal

Extra-toes: a new mutant 559bands of the normal have united in the midline. In the abnormals, however, theyare still widely separated over most of their length.
4. The sacrum
In the XtjXt sacrum (Text-fig. 15) the situation is similar to that present in thecervical region: there is a tendency for vertebral arches to fuse with their neigh-bours rather than with their fellows on the opposite side of the midline.
Text-fig. 14. Thorax of a normal 15-day-old mouse (A) and two XtjXt litter-mates (B, C).
1 mm
Text-fig. 15. Dorsal views of the sacral vertebrae of a normal (A) and an Xt/Xt embryo(B), 15 days old.
The appendicular skeleton
The fore-and hind-limbs of XtjXt embryos show multiple abnormalities(Text-figs, 16, 17). The number of complete digits rises as high as eight or nine.The metacarpals and metatarsals may split distally or be represented only attheir distal extremities. The terminal phalanx of any finger or toe is often whollyor partially duplicated, or absent. The separation between digits is less completethan usual and the digits may be webbed. There is a variable amount of fusion

560 D. R. JOHNSON
amongst the carpals and tarsals, so much so that it is often difficult to identifyindividual elements with any certainty. It is usual for one or more carpal ortarsal to be united with a corresponding metacarpal or metatarsal by a narrowisthmus of cartilage.
Text-fig. 16. Forelimbs of 15-day-old XtjXt embryos. A, Right; B, C, left. F, Fora-men in fossa infraspinata; 0, ossified area.
Text-fig. 17. Hind limbs of 15-day-old Xt/Xt embryos. A, B, Right; C, left.
In the forelimbs the radius is occasionally represented by a proximal rudi-ment which may be no more than a spherical knob of bone adjacent to thehumerus, or extend half-way down the ulna. As a result of this hemimelia theforelimb is more or less bent at the wrist so that the middle digits point across themidline of the body. Usually, however, the radius is present and unreduced. Thehumerus is short and thick; the crista deltoidea is large and coarse or absent.

Extra-toes: a new mutant 561'The scapula often has a foramen in the fossa infraspinata or an indentation uponits cervical margin. The acromion may be reduced or bowed.
In the hind limbs bilateral defect of the tibia is the rule. This, like the radius, isthen represented by a proximal knob of bone. The fibula is somewhat bowed inextreme cases. The femur is short and stout. In the pelvic girdle the Os pubis maybe distorted forming a re-entrant in the outline of the foramen obturatum.
Hemimelia, when present, seems to be biased neither to left nor right (Table 9).Ossification is retarded throughout the limbs of XtjXt embryos.
Table 9.
ForelimbsHind limbs
Incidence
Hemimeliaabsent
190
of hemimelia in 23 Xt/Xt embryos
Left only Right only Bilateral
3 1 01 1 21
Total
2323
1 mm
Text-fig. 18. Dorsal views of normal (left) and Xt/Xt embryos 9£ days old.
EMBRYOLOGY
It has not been possible to classify Xt/Xt embryos in segregating litters lessthan 9 days old, either externally or from sections. At 9 days the homozygote canbe identified on the basis of two consistent abnormalities, a tendency for theneural tube to remain open and the overgrowth of the first pharyngeal arch.
9\ days
At this stage the neural tube of Xt/Xt embryos (identified by their abnormalpharyngeal region) may still be open from the otic vesicle forwards, sometimes

-562 D. R.JOHNSON
as far as the anterior neuropore. If the neural tube is closed (Text-fig. 18) theroof of the rhombencephalon is abnormal, looking like the spade of playingcards rather than being kite-shaped. Behind the otic vesicle the neural tube iswavy to the level of the forelimb bud. The forebrain is pointed rather than
Text-fig. 19. Lateral views of the embryos drawn in Text-fig. 18.
1 mm
Text-fig. 20. Dorsal views of the limb-buds of 9-day-old embryos. Left, normal;right, Xt/Xt.
rounded (Text-fig. 19) and the eyes may be elliptical rather than circular in out-line, the major axis of the ellipse passing through the nasal region. The oticvesicles are situated more dorsally than in normal litter-mates, and are thus veryclose to the neural tube. They may still be open to the exterior. The pharyngealarches are enlarged, especially the mandibular arch which is directed laterally.The anterior limb-buds present an abnormal outline when viewed dorsally(Text-fig. 20); they appear lobed, adjacent lobes being separated by a slightfissure.
In transverse sections (Text-fig. 21) the abnormal shape of the forepart of thehead can be seen to be due to the failure of the forebrain to expand. In the

Extra-toes: a new mutant 563normal 9-day embryo the telencephalon is oval in cross-section; in Xt/Xtanimals its walls are closely approximated. The optic vesicles are situated withtheir antero-posterior axis parallel to the overlying ectoderm and are thus notorientated parallel to the midline. The lumen of the optic stalk is constricted.The rhombencephalon, in this case, is widely opened.
OS mm
Text-fig. 21. Transverse sections through the region of the optic vesicles of 9-day-oldembryos. A, Normal; B, Xt/Xt. Projection drawings. O, Optic vesicle; T, telence-phalon.
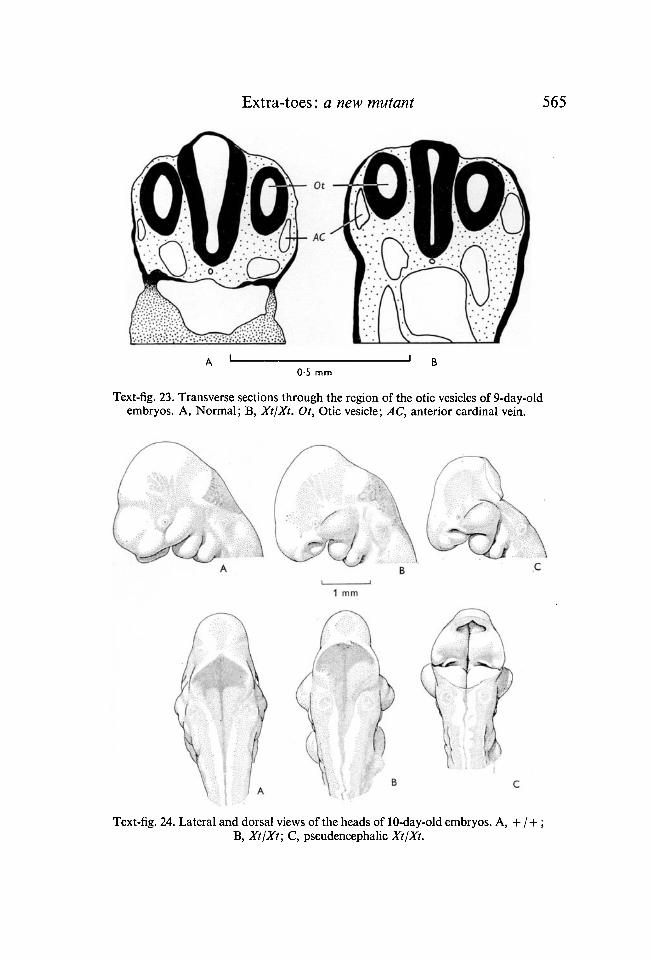
In frontal sections the rhombomeres are clearly visible (Text fig. 22). InXtjXt embryos they are much enlarged, with the exception of the fifth whichseems to be constrained between the otic vessels. This is confirmed by referenceto transverse sections at this level (Text-fig. 23) where the neural tube is alwaysclosed and lacks a roof plate. The anterior cardinal veins are somewhat en-larged and run lateral to the otic vesicles, rather than ventro-laterally.
10 days
At 10 days the external nasal processes of XtjXt embryos are reduced and theforehead lacks the re-entrant normally seen in side view (Text-fig. 24B, C). Thefirst pharyngeal arch has differentiated into a maxillary and a mandibular portion;the former is much enlarged. The pericardium is swollen and usually displaced.The forelimb buds no longer show the lobes present at 9 days and no correspond-

564 D. R. JOHNSON
ing abnormality of the hind limb has been seen. The hind brain may still be open(Text-fig. 24C) or may have been closed by an extensive roof (Text-fig. 24B).
In transverse sections of the normal 10-day embryo (Text-fig. 25) the telen-cephalic vesicles are more marked than at 9 days; the optic vesicle has becomean optic cup and lens induction is in progress. In the XtjXt embryo there is a
B0-5 mm
Text-fig. 22. Frontal sections through the rhombencephalon of 9-day-old embryos.A, Normal; B, Xt/Xt. Ot, Otic vesicle; R, rhombomeres; G, cranial ganglia.
bare suggestion of telencephalic vesicles. The wall of the prosencephalon betweenthese and the optic cup is folded; the optic vesicles are still flattened and theiraxes displaced. Lens induction occurs only posteriorly; the rest of the opticvesicle is no longer in contact with the overlying ectoderm. The nasal placode ismore extensive than normal, but invagination occurs only near its posterior end.In the forelimbs the apical ectodermal ridges are increased in size (Text-fig. 26;Plate 3, figs. M, N).
Extra-toes: a new mutant 565
0-5 mm
Text-fig. 23. Transverse sections through the region of the otic vesicles of 9-day-oldembryos. A, Normal; B, Xt/Xt. Ot, Otic vesicle; AC, anterior cardinal vein.
Text-fig. 24. Lateral and dorsal views of the heads of 10-day-old embryos. A, + /+ ;B, Xt/Xt; C, pseudencephalic XtjXt.

566 D. R. JOHNSON
11 days
By the 11th day all surviving Xt homozygotes have performed some kind ofhind-brain closure, although this is always abnormal (Text-fig. 27). There isstill no invagination between the putative cerebral hemispheres. The opticstalks have broad lumina and carry small optic cups, which usually have folds
Text-fig. 25. Transverse sections through the region of the optic vesicle of 10-day-oldembryos. A, + /+ ; B, Xt/Xt. Abbreviations as in Text-fig. 21.
II0-25 mm
Text-fig. 26. Projection drawings of the AER of a normal (A) and an Xt/Xt (B) embryo.11 days old. Dorso-ventral sections 7-5 ji thick. Every 5th section drawn, the middlesection of the five being the central section of the limb.

J. Embryol. exp. Morph., Vol. 17, Part 3
J
PLATE 3
D. R. JOHNSON facing p. 567

Extra-toes: a new mutant 567in their outer walls. In some embryos the eye cup has lost contact with the ecto-derm and is filled with mesoderm.
The maxillary process, having grown out laterally, starts to extend forwards;the ectoderm of its anterior face is thicker than normal.
Text-fig. 27. Transverse sections through the region of the optic vesicle of 11-day-oldmouse embryos. A, + /+ ; B, Xt/Xt. F, Fold in wall of optic vesicle.
12 days
In the homozygous abnormal there is a distinct bulge in the mesencephalicregion. The roof of the mesencephalon stands proud of the surrounding tissues,covered only by a layer of ectoderm. Behind the otic vesicles the neural tube isirregular with folded walls (Text-fig. 28).
In parasagittal sections of embryos of this age (Plate 3, figs. O, P) the develop-ing nasal region is abnormal. Normally the roof of the nasal chamber is arched.
PLATE 3
Figs. M, N. Transverse sections through the anterior limb-buds of 11-day-old normal (M)and Xt/Xt (N) embryos.Figs. O, P. Parasagittal sections through the nasal regions of 13-day-old normal (O) andXt/Xt (P) embryos. The roof of the nasal chamber is arrowed in each case.Figs. Q, R. Transverse sections through the trunks of 13-day-old normal (Q) and Xt/Xt(R) embryos at the level of the umbilical hernia.
Figs S, T. Transverse sections through the ear regions of 13-day-old normal (S) and Xt/Xt(T) embryos. The vertical axis is arrowed in each case.

568 D. R. JOHNSON
ooaaatuo
i i1 mm
Text-fig. 28. Transverse sections at 100/* intervals through the neural tube of a12-day-old Xt/Xt embryo. The first section is at the level of the second pharyngealarch, the last at the level of the anterior limb bud.
Text-fig. 29. Outline drawings of the limb buds of 12-day-old embryos. Cameralucida drawings. A, + / + ; B, Xt/+ ; C, Xt/Xt right forefeet. D, + / + ; E, Xt/+,F, Xt/Xt right hind feet.

Extra-toes: a new mutant 569In Xt homozygotes, however, it is much flatter, and the nasal chamber thuspartially occluded. The olfactory nerve is less well developed than in normalembryos of this age.
The eyes are reduced and rotated ventrally so that they look downwards intothe cheek. In some cases the maxillary process has grown forward over the eyeso that the latter is partially or completely obscured.
Text-fig. 30. Superimposed outline drawings of limb-buds; outlines as in Text-fig. 29.Solid line, normal; dotted line, abnormal. A, Right forefoot, Xt/ + on + /+ ; C, righthind foot, Xt/ + on + /+ ; B, Right forefoot, XtjXt on + /+ ; D, right hind footXt/Xt on + /+ .
At this stage the heterozygote can first be recognized by its footplates (Text-fig. 29). The normal 12 day footplate has an indented outline, and regionscorresponding to five digits may be recognized. In the Xt\ + limb-bud the areadestined to become digit I is enlarged. The XtjXt limb-bud is much wider thannormal. The indentations between digits are less well marked. There is obviouspreaxial enlargement and superimposition of the outlines of normal and XtjXtlimb-buds (Text-fig. 30) shows that there is postaxial enlargement also. In thehind limb the picture is similar. The Xt/Xt footplate has a pre- and a postaxialwidening, which seems to have occurred at the expense of proximo-distalelongation.
13 days
The 13-day XtjXt embryo has developed considerable subcutaneous oedema(Text-fig. 31). This takes the form of a swelling on either side of the neural tubewhich extends from the cervical region to the root of the tail and laterally as far asthe proximal part of the limbs. In sections (Plate 3, figs. Q, R) it can be seen thatthe oedema is due to expansion of the subdermal mesoderm which is less densethan normal. In the thoracic region a second oedematous area is seen betweenthe lungs and the ribs. There is an enlarged umbilical hernia which includespart of the liver. Mild oedema is sometimes evident in heterozygotes (8 of34 examined) and in one case a clear fluid-filled bleb was present in the shoulder
36 J E E M 17

570 D. R. JOHNSON
region. The homozygote is given a hunch-backed appearance by its distendedhind brain. On one or two occasions a bleb was seen surmounting this hump.The forehead and nose have an irregular array of epidermal papillae resemblingthe follicles of sensory hairs, which in fact they represent. Up to 10 have beenseen, but 2 or 3 is more usual. The supra-orbital papillae are often displaced andmay be three rather than two in number. By contrast the post-orbital papillaewere reduced or absent in 17 out of 28 heads examined. The maxillary region isenlarged and carries more follicles (which give rise to mystacial vibrissae) thanis usual. Normally five rows are visible; in XtfXt a partial sixth row is ofteninserted and the number of follicles per row increased.
2 mm
Text-fig. 31. Camera-lucida drawings of + /+ (left) and XtjXt 13-day-old embryos.
The number of nipples visible in XtjXt mice at this stage is reduced from fourto two pairs, the most anterior and posterior ones persisting. Plate 3, figs. S, T,shows transverse sections through the otic regions of normal and XtjXt embryosrespectively. In the normal there is an evagination in the wall of the pre-sumptive utricular region opposite the point of entry of the endolymphaticduct, which will later become the lateral semi-circular canal. In the abnormalthis evagination is absent. The seventh and the cochlear part of the eighthcranial ganglia are enlarged.
14-16 days
The 14-day XtjXt embryo is still oedematous. The abdominal hernia hasreached an abnormally large size. Extra digits can be seen both pre- and post-axially on the feet of the heterozygotes, and some have an oval blood clot in thefrontal region of the head.
At 15 days the oedema is less marked but the skin appears taut all over thebody.

Extra-toes: a new mutant 571At 16 days the mesencephalon may be exposed via an aperture in the cranium
which corresponds to the midcerebral bump found in younger animals. Whenthis occurs the amniotic fluid is always bloody and the embryo anaemic.
In sections of 14- to 16-day embryos small subcutaneous fluid-filled blebsare seen, in locations likely to be overlooked on superficial examination. Suchblebs have been found on the tip of the lower jaw, on the underside of the upperlip and in the external auditory meatus, at the base of the pinna. Heterozygotesmay also have blebs in the last position, and between the digits. Corneal blebsare seen, in homozygotes only, when the eyes are much reduced and there ispartial premature closure of the eyelids.
DISCUSSION
The syndrome described in this paper is one of the most complex known fromthe study of mammalian developmental genetics. In order to place the multipleeffects of the extra-toes gene in perspective it is necessary to understand theactions and interactions of abnormal parts during development. From the manyabnormalities described above three groups of related disturbances emerge:those due to (1) the maldevelopment of the nervous system, (2) the overgrowthof the pharyngeal arches and limb buds, and (3) oedema.
The brain and CNS
The late closure of the neural tube is one of the earliest abnormalities seen inXt. The subsequent elongation of the forebrain and folding of the neural tubepoint to an increase in volume of the neural tissue.
The forebrain, by failing to expand and continuing to elongate determines theshape of the forepart of the head. The optic vesicles which are attached to theforebrain must perforce be involved. The pointed forebrain leads to the apparentdecrease in the size of the nasal processes and probably to the abnormal in-vagination of the nasal placode. The failure of the roof of the telencephalon toinvaginate leads to the formation of small cerebral hemispheres abnormallypositioned with respect to each other. The anterior choroid plexus is formed verylate and is small and superficial. This may account for the pycnoses seen in thecerebral hemispheres at birth.
Farther back, at the level of the otic vesicles, the roof of the neural tube isless extensive than usual at the 9-day stage. Again this seems to be due to non-closure of the forebrain, and consequent failure to expand owing to lack ofhydrostatic pressure. The folding of the neural tube is not uniform. It appearsfirst and reaches its greatest extent in three regions, cervically and at the thoracicand lumbar flexures. This is the situation expected, if the folding is due toovergrowth of neural tissue.
The neural tube may fail to close because of an intrinsic defect in the neuraltissue or an extrinsic factor which fails to allow the edges of the neural folds to
36-2

572 D. R. JOHNSON
meet and fuse. Examples of both mechanisms are found in the literature. In theLoop-tail mouse Smith & Stein (1962) have suggested that the failure of theneural folds to unite is due to the failure of the primitive streak (and hence theneural plate) to elongate correctly. This is clearly an extrinsic dysraphism. InSplotch, on the other hand, Auerbach (1954) considers that there is a primarydefect of the neural plate, and Smith & Stein suggest that the difficulty in closingis due to an incomplete separation of the neural crest from the neural tube.Patten (1952, 1953) has described a series of pseudencephalic embryos whosebrain defects are similar to those of extreme Xt homozygotes. He ascribes thenon-closure of the neural tube to overgrowth of neural tissue. Dekaban &Bartelmez (1964), however, suggest that non-closure of the neural folds couldlead to an overgrowth of neural tissue. The work of Fowler (1953) supportsthis hypothesis. She operated on chick embryos, slitting the roof of the neuraltube and thus producing artificial spina bifida. Embryos thus treated showedneural overgrowth.
The eruption of the brain in the parietal region is considered to be a separateevent from the early failure of the neural tube to close. Exencephalic embryosare not seen between 11 and 16 days. The inference is that those embryos seen at16 days with exteriorized neural tissue have acquired it as a result of a secondarydisturbance acting upon the closed neural tube. Further, the protruding brainis not turned inside out as in pseudencephalic mice, but simply protrudesthrough the cranium. The Xt condition thus seems to resemble the mid-cerebrallesions seen by Carter (1959) in my rather than the pseudencephalic micedescribed by Bonnevie (1936) in the same stock.
, rrj The sense organs1. The nose 6
The first significant factor in the development of the XtjXt nose is the ab-normal invagination of the nasal placode seen at the 10-day stage. At 12 daysthe roof of the nasal chamber is flat rather than vaulted due to the excessive sizeof the maxillary processes which form it. The posterior part of the nose is laterflattened by the abnormal brain, whilst its anterior region is compressed verti-cally by the huge maxillae. A little later still the forepart is further compressedby the developing incisors. This may result in a loss of contact between thenasal chamber and the exterior.
Another group of abnormalities is due to the excessive growth of the epi-thelium lining the nasal chamber, first visible at the 15-day stage. The olfactorynerve consists of fibres which have their origin in the olfactory epithelium andgrow back towards the brain. At the 12-day stage this tract of fibres is poorlydeveloped in Xt\Xt\ at birth it is huge, notwithstanding the fact that contactwith the brain has not been made. It seems probable that the overgrowth of thenasal epithelium gives rise to a correspondingly enlarged olfactory tract.

Extra-toes: a new mutant 573
2. The eye
The developing eye is first affected by the elongation of the forebrain at the9-day stage. The proximal part of the optic stalk is carried forwards by the fore-brain; the distal part with its attached optic vesicle lags behind. The latter is oftendeformed by the force applied to it, and tends to become elliptical in cross-section. To a greater or lesser extent contact is lost between the surface of thevesicle and the ectoderm. The eye which develops from such a vesicle must be ofreduced size and, if separation of the eye cup from the ectoderm continues, willbe invaded by mesenchyme cells.
The part of the optic vesicle not involved in lens induction, which is largerthan normal, is incorporated into the optic stalk or appears as a fold in the outerlayer of the eye cup. By the 12-day stage the developing eye is threatened furtherby the maxillary process which grows over its lower part. Later still, at 15 daysthe retina may start to proliferate and fold. Finally a corneal bleb may be presentin some cases. At a stage depending upon the net result of these variables the eyeceases to develop further; hence in newborn XtjXt mice all stages are seen fromvirtual absence of the eye to a small reasonably developed eye with a crystallinelens and a folded retina.
3. The ear
The first sign of abnormality of the ear is the anomalous position of the oticvesicles at the 9-day stage, probably due to slow development. From thenonwards they lie adjacent to an abnormal hind brain.
Deol (1964, 1966) has demonstrated the correlation between abnormal hindbrain and abnormal ear in a number of mutants. It seems that in Xt also the poordifferentiation of the ear is a direct result of the abnormal hind brain.
4. Oedema and blebs
The origin of the oedema which arises in XtjXt embryos at the 13-day stage isnot clear. A similar oedema in talpid3 mutant in the chicken was ascribedby Ede & Kelly (1964a) to a mesodermal defect. In discussing the situation inPatch, in the mouse, Griineberg & Truslove (1960) suggest that hydrops and blebs'may be comparatively unspecific and rather far removed from the primary geneaction', pointing out that 'disturbances of liquid balance (oedema, blebs) areoften encountered in amphibian embryos which have been subjected to surgi-cal interference.' On the other hand M. S. Deol (unpublished) has also pointedout the association between blebs and neurological disturbances in themouse; Xt may well be yet another blebby/neurological mutant.
Whatever the origin of the oedema its results are clear cut. The force exertedby the fluid contents of the swellings lateral to the spine disrupts the ventralclosure of the embryo, leading to umbilical hernia. The presence of oedematoustissue in the dorsal part of the body leads to a reduction of space in the abdomen.

574 D. R. JOHNSON
The effects of the oedema are first on the sternum and secondly on theviscera.
The abnormal position of the ovaries may be explained in the following way.The mammalian gonad arises from the mesonephros, cephalad to the meta-nephros, and migrates caudad. The mechanism by which this migration isachieved is quite different in the two sexes. In the male it is active, through thecontraction of the gubernaculum relative to the surrounding tissue. In the femalethe ovaries migrate passively over a shorter distance under the influence of theirown weight. It is clear that in XtjXt males the process is normal, but that in thefemales passive migration of the ovaries is hampered by cramped conditions inthe abdomen.
No direct sequelae to the presence of blebs in the form of haematoma orthrombi of the type found in my and Ph have so far been discovered in Xt. How-ever, the blebs shown by Xt homozygotes are much smaller and less widespreadin distribution than those present in conjunction with the other two genes.
5. Vibrissae
The number of primary and secondary vibrissae in the mouse is an almostinvariant character. Fraser & Kindred (1960) worked on the sex-linked geneTabby which affects the coat and reduces the number of secondary vibrissae.After intensive selection they were able to produce lines of mice where the numberof vibrissae was greater or smaller than normal. When extra vibrissae werepresent they were always limited to a pre-existing site, e.g. the number of supra-orbital sinus hairs was sometimes increased from two to three.
Extra-toes shows abnormalities in the number of mystacial and supra-orbitalsinus hairs and in their pattern. There are also extra vibrissae present on thedorsal part of the snout and on the forehead. There are therefore two elementsat work. One of these is the interference with the number of vibrissae at anestablished site, as noted by Fraser & Kindred, and the other is the induction ofvibrissae at a site where none normally exist. The site in this instance includespractically the whole of the forehead, with no recognized preference for specificareas.
The skeletal system
The morphology of the skeletal system is largely subordinate to that of sur-rounding tissues. All the anomalies of the skeleton of XtjXt and Xtj + mice canbe related to pre-existing abnormalities in other structures.
The limbs
Polydactylism and hemimelia seem to be consequences of abnormal develop-ment of the limb-buds. The differences seen between the anterior limb-buds inXtjXt and normal embryos predates the first limb-bud abnormality seen in anyother mutant (Griineberg, 1961) by 2 days. It is probable that a more detailed

Extra-toes: a new mutant 575study of the early development of Xt limb-buds will reveal further changes instructure.
The work carried out so far sheas no light on the controversial roles of meso-derm and ectoderm in limb development. It can only be stated that the distribu-tion of mesoderm at 9 days is abnormal and that the AER is larger than normalat 10 days, when this structure approaches its maximum development.
The polydactyly which occurs in Xt cannot be classified with relation to thelimb axis in the usual way as both pre- and postaxial digits are regularly addedeven in the heterozygote. That the former are always larger in size and greater innumber than the latter is perhaps due to the postaxial-preaxial sequence ofmesodermal condensations in the footplate (Forsthoefel, 1963; Milaire, 1965).However, the type of polydactyly found in the heterozygote accords quite wellwith previous descriptions of polydactylous limbs.
Polydactyly seems to be divided into three morphological types. In the first(e.g. Duplicate in the chick, Warren, 1941; Strong's luxoid in the mouse, Strong,1961), part of a mirror image hand is formed, the digital sequence being V, IV,III, II, I-I, II, III, etc. New elements appear preaxially to all existing materialin each case, and the hallux remains more or less normal. This type of poly-dactyly may be explained by postulating a splitting of the AER into two parts.
In the second type (which includes luxate (Carter, 1951) and fidget (Truslove,1956)) and is characterized by the presence of incomplete metatarsals, the digitalsequence is less clear. Here digits are not added to the preaxial side of the footbut arise from the splitting of digit I or II, even in the presence of a pre-hallux.It is suggested that the organizing centre of the AER is enlarged but intact in thiscase. The polydactyly of extra-toes heterozygotes is basically of this type, if thepost-axial digit is ignored.
The third type of abnormality is that shown uniquely by eudiplopodia in thechick (Rosenblatt, Kreutziger & Taylor, 1959) where two-plane polydactyly iscorrelated with the appearance of two separate AER's.
The hemimelia in Xt homozygotes could be explained following Forsthoefel(1959). He suggested that the association of preaxial polydactyly and hemimeliamight be due to blastemal competition. The anterior portion of the footplate maybe able to 'take over' mesoderm from the anterior side of the forelimb anlagefor digit formation, thus leaving the radial region deficient: this is expressedlater as hemimelia. This argument, however, does not account for the presenceof hemimelia associated with ectrodactyly, as in Dh (Searle, 1964) and occasionalXt heterozygotes, where there is a preaxial tissue deficiency in both limb andfoot.
The involvement of the limb-girdles is of a type previously described in rela-tion to luxoid mutants. The presence of a foramen in the fossa infraspinata hasbeen described by Curry (1959) in droopy-ear and by Searle (1964) in postaxialhemimelia (px). Curry showed that this defect is present in the membranousskeleton and is therefore the result of a blastemal abnormality. Inpx the foramen

576 D. R. JOHNSON
probably dates from the same stage, and is certainly present in the cartilaginousscapula. In XtjXt the Os pubis may be reduced or bowed. This is also seen inDh, 1st and Ix (Carter, 1951; Strong & Hardy, 1956; Strong, 1961; Searle, 1964).These defects of the limb-girdles may be attributed to the redistribution ofskeletogenic material in the limb-bud.
The extensive bony f usions]of the digits on the preaxial side of the forelimb areclearly tertiary in nature. No fusion of cartilaginous elements has been seen inthis region up to birth. During postnatal growth the proportions of the forefootchange quite markedly, with the central digits elongating much more rapidly thanthe first, which is rudimentary in the adult. It seems that during this periodthe extra elements in the region of digit I become compressed and that fusiontakes place amongst them.
The two Xt\-v hemimelic animals which have occurred in the stock mostprobably represent a rare variation of the Xt genotype; as both these mice weredescended from the outcross mating Xt5/CB 1125 it seems likely that the hemi-melic form arose due to a change in genetic background.
2. The skull and axial skeleton
The abnormalities of the brain and spinal cord of Xt\Xt mice lead inexorablyto skeletal malformations. The abnormal skull is moulded upon the abnormalbrain. The cervical vertebrae fail to enclose the enlarged neural tissue and thesame situation, albeit to a lesser extent, is found in the sacral region. The fusionsbetween adjacent vertebrae in the cervical region are clearly secondary and arelimited to the processes destined to form the neural arch. There is no sign ofirregularities of the vertebral bodies (cf. talpid3, Ede & Kelly, 1964 a, b).
3. Belly spots
The data presented in Table 6 leave no doubt as to the association between Xtand belly spotting. Similar effects of a so-called 'major' gene upon a spottinggene have been previously described by Griineberg (1948).
Griineberg found that when mice heterozygous for microphthalmia (mi) werecrossed with his grey-lethal stock + \mi individuals showed head dots and bellyspotting absent from + / + litter-mates. Griineberg was able to show that mlin this situation is acting as a specific genetic modifier of spotting genes.
A similar situation has recently arisen in our laboratory (G. M.Truslove,personal communication). In syndactylism (sm, Griineberg, 1956) a head dot,confined to animals of the sm/sm genotype, has recently arisen. This occurredonly in certain families of the stock and has now died out.
It therefore seems most probable that on the genetic background of the Xtstock the gene acts as a genetic modifier of pre-existing spotting genes.

Extra-toes: a new mutant 577
Relationship o/Xt to other syndromes
Few syndromes known to developmental genetics provide such a range ofabnormalities as does extra-toes. The presence of a large number of concordantabnormalities is limited to a few genes, pollex (guinea-pig), blebs (mouse),patch (mouse) and the talpids (chicken).
The similarity to pollex is striking. Most of the features described by Scott(1937,1938) have parallels in the mouse. The youngest monsters described by himare aged 18£ days which corresponds to the 9^-day stage in the mouse. Thecharacteristic features of XtjXt are present in Scott's illustrations. The ace-of-spades shape of the roof of the rhombencephalon is clear, as is the displacementof the mandibular arch and the ovality of the eye. The description of the brainfits Xt/Xt well. In all subsequent stages there is good agreement of abnormalities.
Scott mentions the overgrowth of the pharyngeal arches only once, in con-junction with the presence of a fifth arch. It is clear that overgrowth of the mandi-bular arch must have been much less pronounced in Px than in Xt. Scott'sfailure to note the late closure of the neural tube may be attributed to the samecause. Cleft palate and harelip, reported in Px, have not been seen in Xt.
Table 10
myPhXt
+ ,
. Comparison
CNSdefect
of some features of the my, Ph and Xt
Blebs/oedema Polydactyly
Present; —, unrecorded or absent;
Kidneydefect
syndromes
Pigmenteffect
* — +
*, triphalangy of the pollex.
The reinvestigation of the X-rayed stock of Bagg & Little (blebs, my) byCarter (1956, 1959) and the description of patch by Griineberg & Truslove(1960) have shown that these genes have much in common with Xt. Carter'sdemonstration that previously described 'sporadic' defects are in fact basicallyattributable to one major gene enabled him to construct four 'pedigrees ofcauses' based upon CNS abnormalities, blebs/oedema, kidney defects andfaulty morphogenesis of the limbs. Patch and extra-toes share many of thesefeatures (Table 10). The pigment defects include spotting in patch and a shiftingof the pigment border in black-and-tan my\my mice. This may really be a hairdefect (as Carter assumes) or a secondary effect of a dorsal bleb on skin structure.The + sign opposite extra-toes refers to belly spotting seen in Xt heterozygotes.
It is not to be assumed, however, that the mechanisms underlying my, Ph andXt are identical or even similar. The defects described are common, as Carterpoints out, in embryos damaged at the 8^-9^ day stages by teratogens. If nothingmore, however, it seems that the lesions of my, Ph and Xt are produced by genesacting at a common stage of development.

578 D. R. JOHNSON
The talpid group of alleles in the chicken (Cole, 1942; Abbott, Taylor &Abplanalp, 1959, 1960; Ede & Kelly, 1964a, b) are polydactylous monsterssuperficially similar to the group described above. Ede & Kelly, however,describe mesodermal abnormalities such as irregular vertebral bodies and failureof mesenchymal condensations, and regard the syndrome as mesodermal inorigin.
CONCLUSIONS
Most of the abnormalities of the XtjXt phenotype can be ultimately tracedback to one of three anomalies: the disturbed neural tube, overgrowth of thepharyngeal arches and limb-buds and, later, oedema. As the first two abnormali-ties both become visible at the 9-day stage it is not possible to suggest a causalrelationship on temporal grounds. Three points of view are possible. First, theovergrowth of the pharyngeal arches may mechanically affect the closure of theneural tube. This is a possibility, as the area of non-closure is in the region of thepharyngeal arches and no other region of the neural tube is persistently open.On the other hand, the hind brain is one of the two regions most commonlyaffected by non-closure, and many cases of hind-brain dysraphism not associatedwith spina bifida are known. Secondly, there may be an abnormality of theneural plate which delays its closure and in some way affects the pharyngealarches and the limb-buds. An abnormality of the CNS could easily account forthe derangements seen in axial structures. Involvement of the appendicularskeleton is less easily explained. However, M. S. Deol (unpublished) has sug-gested that in some mutants the abnormalities of the nervous system may not beconfined to the neural tube but expand over the ganglia and nerves as well, andwhen this happens in the pectoral or pelvic region the limbs may be affected inconsequence. The present paper contains no evidence to confirm or deny thishypothesis. Thirdly, an unknown pre-existing factor may affect the developmentof limb-buds, pharyngeal arches and neural tube.
If we accept that the overgrowth of the neural tube is primary rather thansecondary, then both this and the enlargement of the limb-buds and pharyn-geal arches could be due to overgrowth, possibly as a result of a derangement ofthe reciprocal ectodermal/mesodermal interaction present during organogenesis.This could also explain the late proliferation of the nasal and retinal epitheliaand the appearance of extra vibrissae. Also in favour of this hypothesis is thelack of abnormalities where ectoderm and endoderm interact without theintervention of mesoderm; the pituitary body and the VER of the tail are ex-amples of this. Both are normal in XtjXt mice.
The position reached by this line of reasoning is similar to that of Scott (1938)in Px. He postulated the production in PxjPx embryos of an intracellular growthfactor 'for a short time after approximately 17i days copulation age'. Carter(1959) in reviewing his four 'pedigrees of causes' for my remarked that no causalconnexions between them had been traced, but goes on to postulate that con-

Extra-toes: a new mutant 579nexions may exist because of the significant statistical associations between partsof the syndrome. Griineberg & Truslove (1960) are inclined to ascribe thedefects of PhjPh to hydrops which is present from the 8-day stage onwards.
If we accept the concept of unitary primary gene action we also accept thatthe parts of extensive syndromes such as those produced by the genes my, Ph, Pxand Xt are related. In the absence of factual evidence to support this hypothesis,however, the investigator is liable to fall between two stools: either to admitdefeat (as did Carter) or to propose a blanket explanation (as have Scott andmyself) which runs the risk of being so general as to explain nothing. It is hoped,however, that further studies of Xt mice at present contemplated will allow a moreprecise explanation to be offered at a later date.
SUMMARY
1. Extra-toes (Xt) is a new semi-dominant gene in the mouse. XtjXt animalsdie in utero or at birth. They have multiple abnormalities including paddle-shaped feet with up to eight or nine digits, hemimelia, disturbed spinal cord andbrain, nose, eye and ear, and oedema. The heterozygote has pre- and post-axial polydactylism of all four feet.
2. The Extra-toes homozygote can first be recognized as a 9-day embryo byexcessively large pharyngeal arches and persistently open neural tube.
3. Many abnormalities of the brain and sense organs can be traced back tomalformation of the neural tube.
4. Malformations of the upper jaw region and limbs can be traced back toovergrowth from the 9-day stage onwards.
5. A third series of malformations due to oedema (first seen at 13 days) isdescribed.
6. The possible interrelations of these three groups of abnormalities to eachother, and the relationship of the Xt syndrome to other gene effects in the mouse,chicken and guinea-pig are discussed.
RESUME
Extra-toes: un nouveau gene mutant provoquant denombreuses anomalies chez la Souris
1. Extra-toes (Xt): nouveau gene semi-dominant chez la Souris. Lesanimaux Xt/Xt meurent dans l'uterus ou a la naissance. Us presentent denombreuses anomalies parmi lesquelles des pieds en forme de palette avec plusde 8 ou 9 orteils, de l'hemimelie, l'alteration de la moelle epiniere et du cerveau,du nez, de l'oeil, de l'oreille et de l'cedeme. Les quatre pieds des sujets hetero-zygotes presentent de la polydactylie pre et postaxiale.
2. On recommit le type homozygote de la mutation extra-toes chez l'embryonde 9 jours par la presence d'arcs branchiaux tres larges et d'un tube neuralouvert.

580 D.R.JOHNSON
3. De nombreuses anomalies du cerveau et des organes des sens peuventetre dues aux malformations du tube neural.
4. Les malformations de la region de la machoire superieure et des membrespourraient provenir d'excroissances apparaissant apres le 9eme jour.
5. Description d'une troisieme serie de malformations due a un cedeme(premiere observation a 13 jours).
6. La discussion porte sur les relations possibles entre les trois groupesd'anomalies et la relation entre le syndrome Xt et d'autres effets du gene chezla souris, le poulet et le cobaye.
I am indebted to Professor H. Griineberg, F.R.S., for his constant encouragement andcriticism, and to Drs Gillian M. Truslove and M. S. Deol for much useful discussion andadvice. I am much indebted to Mr A. J. Lee who prepared most of the Text-figures. The workwas supported by a grant from the Medical Research Council which is gratefully acknowledged.
REFERENCES
ABBOTT, U. K., TAYLOR, L. W. & ABPLANALP, H. (1959). A second talpid-like mutation in thefowl. Poult. Sci. 38, 1185.
ABBOTT, U. K., TAYLOR, L. W. & ABPLANALP, H. (1960). Studies with talpid2, an embryoniclethal of the fowl. / . Her ed. 51, 195-202.
AUERBACH, R. (1954). Analysis of the developmental effects of a lethal mutation in the housemouse. J. exp. Zool. 121, 305-30.
BONNEVIE, K. (1936). Pseudencephalie als spontane recessive (?) Mutation bei der Hausmaus.Skr. Norske Vidensk-Akad. Oslo, Math-nat kl. no 9, 39 pp.
CARTER, T. C. (1951). The genetics of luxate mice. I. Morphological abnormalities of heterozy-gotes and homozygotes. / . Genet. 50, 277-99.
CARTER, T. C. (1956). Genetics of the Little & Bagg X-rayed mouse stock. / . Genet. 54, 311-26.CARTER, T. C. (1959). Embryology of the Little & Bagg X-rayed mouse stock. / . Genet. 56,
401-35.COLE, R. K. (1942). The 'talpid lethal' in the domestic fowl. / . Hered. 33, 82-6.CURRY, G. A. (1959). Genetical and developmental studies on droopy-eared mice. / . Embryol.
exp. Morph. 7, 39-65.DAWSON, A. B. (1926). A note on the staining of the skeleton of cleared specimens with
Alizarin Red. S. Stain Tech. 1, 123-4.DEKABAN, A. S. & BARTELMEZ, G. W. (1964). Complete dysraphism in 14 somite human
embryo. Am. J. Anat. 115, 27-41.DEOL, M. S. (1964). The abnormalities of the inner ear in kreisler mice. / . Embryol. exp.
Morph. 12, 475-90.DEOL, M. S. (1966). Influence of the neural tube on the differentiation of the inner ear in the
mammalian embryo. Nature, Lond. 209, 219-20.EDE, D. A. & KELLY, W. A. (1964a). Developmental abnormalities in the head region of the
talpid3 mutant of the fowl. / . Embryol. exp. Morph. 12, 161-82.EDE, D. A. & KELLY, W. A. (19646). Developmental abnormalities in the trunk and limbs
of the talpid3 mutant of the fowl. / . Embryol. exp. Morph. 12, 339-56.FORSTHOEFEL, P. F. (1959). The embryological development of the skeletal effects of the
luxoid gene in the mouse, including its interactions with the luxate gene. J. Morph. 104,89-142.
FORSTHOEFEL, P. F. (1963). Observations on the sequence of blastemal condensations in thelimbs of the mouse embryo. Anat. Rec. 147, 129-38.
FOWLER, I. (1953). Responses of the chick neural tube in mechanically produced spina bifida./ . exp. Zool. 123, 115-51.

Extra-toes: a new mutant 581FRASER, A. S. & KINDRED, B. M. (1960). Selection for an invariant character, vibrissa number,
in the house mouse. Aust. J. Biol. Sci. 13, 48-58.GRUNEBERG, H. (1943). The development of some external features in mouse embryos.
/ . Hered. 34, 88-92.GRUNEBERG, H. (1948). Some observations on the microphthalmia gene in the mouse.
/ . Genet. 49, 1-13.GRUNEBERG, H. (1953). Genetical studies on the skeleton of the mouse. VII. Congenital
hydrocephalus. J. Genet. 51, 327-58.GRUNEBERG, H. (1956). Genetical studies on the skeleton of the mouse. XVIII. Three genes
for syndactylism (with an appendix by D. S. Falconer). / . Genet. 54, 113-45.GRUNEBERG, H. (1961). Genetical studies on the skeleton of the mouse. XXVII. The develop-
ment of Oligosyndactylism. Genet. Res. 2, 33-42.GRUNEBERG, H. & TRUSLOVE, G. M. (1960). Two closely linked genes in the mouse. Genet.
Res. 1, 69-90.LYON, M. F., MORRIS, T., SEARLE, A. G. & BUTLER, J. (1967). Occurrences and linkage
relations of the mutant 'extra-toes' in the mouse. Genet. Res. (in the Press).MILAIRE, J. (1965). fitude morphogenetique de trois malformations congenitales de l'autopode
chez la souris (Syndactylisme-Brachypodisme- Hemimelie dominante) par des methodescytochimiques. Mem. Acad. r. Belg. Cl. Sci. 16, 1-120.
PATTEN, B. M. (1952). Overgrowth of the neural tube in young human embryos. Anat. Rec.113, 381-93.
PATTEN, B. M. (1953). Embryological stages in establishing myeloschisis with spina bifida.Am. J. Anat. 93, 365-95.
ROSENBLATT, L. S., KREUTZIGER, B. O. & TAYLOR, L. W. (1959). Eudiplopodia. PoultrySci. 38, 1242.
SCOTT, J. P. (1937). The embryology of the guinea pig. III. The development of the poly-dactylous monster. / . exp. Zool. 11, 123-57.
SCOTT, J. P. (1938). The embryology of the guinea pig. II. The polydactylous monster./ . Morph. 62, 299-321.
SEARLE, A. G. (1964). The genetics and morphology of two 'luxoid' mutants in the housemouse. Genet. Res. 5, 171-97.
SMITH, L. J. & STEIN, K. F. (1962). Axial elongation and its retardation in homozygouslooptail mice. / . Embryol. exp. Morph. 10, 73-87.
STRONG, L. C. (1961). The Springville mouse, further observations on a new 'luxoid' mouse./ . Hered. 52, 122-4.
STRONG, L. C. & HARDY, L. B. (1956). A new 'luxoid' mutant in mice. / . Hered. 47, 277-84.TRUSLOVE, G. M. (1952). Genetical studies on the skeleton of the mouse. V. 'Interfrontal'
and 'Parted frontals'. / . Genet. 51, 115-22.TRUSLOVE, G. M. (1956). The anatomy and development of the fidget mouse. / . Genet. 54,
64-86.WARREN, D. C. (1941). A new type of polydactyly in the fowl. / . Hered. 32, 2-5.
{Manuscript received 23 January 1967)