experimental study of suspension-feeding activity in the ...directory.umm.ac.id/data...
TRANSCRIPT

Journal of Experimental Marine Biology and Ecology252 (2000) 57–74
www.elsevier.nl / locate / jembe
Experimental study of suspension-feeding activity in the¨serpulid polychaete Ditrupa arietina (O.F. Muller)
*ˆ ´Esther Jordana, Jean-Claude Duchene, Francois Charles , Antoine Gremare,Jean-Michel Amouroux
´Observatoire Oceanologique de Banyuls, CNRS UMR 7621, BP 44, F-66651, Banyuls-sur-Mer Cedex,France
Received 23 March 1999; received in revised form 23 May 2000; accepted 31 May 2000
Abstract
As time spent feeding can be an index of the feeding intensity, we attempted to describe thefiltering activity in the suspension-feeding polychaete Ditrupa arietina. This description was basedon the detection of the opening of the gill-crown by an automated image analysis system. Thecommon pattern of activity could be described as the succession of filtering events and restingperiods of which the number and the mean duration differed greatly from one worm to another.Filtering activity in D. arietina was neither a continuous process nor even a process having aparticular rhythm. Within a same batch of worms, total filtration durations could represent between12.5 and 87.5% of the total experimental time. Despite a strong inter-individual variability, ourresults showed the existence of pronounced seasonal variations in the activity of the gill-crown. InMay, worms spent less than 25% of time feeding compared to more than 50% during the rest ofyear. These temporal changes appeared to result from the physiological state of the worms(reproductive period and ageing) at the time of the experiment. 2000 Elsevier Science B.V. Allrights reserved.
Keywords: Suspension-feeding; Polychaete; Image analysis; Ditrupa arietina
1. Introduction
The serpulid polychaete Ditrupa arietina has recently increased all along the north´western coasts of the Mediterranean Sea (Gremare et al., 1998a,b). The abundances
2reported, up to 3000 individuals (ind) /m , and the wide geographical distribution
*Corresponding author.E-mail address: [email protected] (F. Charles).
0022-0981/00/$ – see front matter 2000 Elsevier Science B.V. All rights reserved.PI I : S0022-0981( 00 )00231-8

58 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
suggest that through processes such as nutrition, this species affects the functioning ofthe coastal benthic ecosystem in this region.
As for all the other serpulids (Jorgensen, 1966), D. arietina is a suspension feeder.Therefore, through the benthic trophic network, one of its major impacts concerns theremoval of particles from the water mass overlaying the sediment. Quantitativeinformation about the filtration and clearance rates in suspension-feeding polychaetesremains rather scarce. One commonly held belief is that this group of suspension-feedersprocesses small water volumes and is generally less efficient than other suspension-feeding (Dales, 1957). Consequently suspension-feeding polychaetes have more or lessbeen considered as organisms relying on environmental factors to get the energy theyneed (Merz, 1984). More recently, the ecological role of suspension-feeding inpolychaetes has been demonstrated in some systems (Merz, 1984; Davies et al., 1989;Riisgard et al., 1996). Moreover, suspension-feeding polychaetes are described as highlyefficient suspension-clearers, relying on very dilute food resources to live and grow(Riisgard and Ivarsson, 1990). Laboratory experiments, especially those conducted toassess physiological performances (e.g. filtration, clearance, and oxygen consumptionrates) of suspension feeders, were often run on isolated specimens (Shumway et al.,1988; Riisgard and Ivarsson, 1990). Consequently, values measured generally corre-spond to the greatest performance rates of the organisms. Extrapolation of such values todetermine, for instance, the global feeding pressure of a natural population of wormsrequires knowledge of the average time actually spent feeding by each individual.
In suspension feeders, the intensity of feeding basically depends on filtering capacityand filtering activity (i.e. time spent pumping; Foster-Smith, 1976). Such a parametersometimes may be affected by factors which that induce co-ordinated responses andeven rhythm of activity (Leonard, 1989; Sanford et al., 1994; Vedel et al., 1994; Achituvand Yamaguchi, 1997; Thorin et al., 1998). The assessment of the feeding activity atdifferent time scales and under various environmental conditions is thus essential forextrapolating feeding rates (clearance and ingestion rates) to the field.
The aim of the present study is to describe the filtering activity of the serpulid Ditrupaarietina. This description is based on the utilisation of an automated video system thattracks the opening of the gill fan (e.g. indicating that worms were processing water). Welooked (i) for temporal changes in the filtration behaviour of worms from differentpopulations, and (ii) at the effect of variations in environmental factors on the filteringactivity.
2. Material and methods
2.1. The automated video system
The automated video system was developed by one of us (J.-C.D.) to quantify variousactivities of benthic epifauna. This tool permits automated image analysis of videoimages taken at the sediment surface. Briefly, it consists (see Fig. 1) of a video sensorcomposed of a charged coupled device (CCD; type ICX059 AI from VLSI Vision Ltd.)connected to an ADSP 2181 signal microprocessor, a memory board and an interface

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 59
Fig. 1. The automated video system. (A) Basic set-up for the experiments. (B) Main components of the videosensor (arrows represent image-analysis processing by the automated video system).
board. The microprocessor is driven by real-time routines uploaded in the permanentmemory. These routines control picture acquisition frequency and run images analysis.Images of the sediment surface are collected by the CCD and transferred to the videoworking memory of the microprocessor. In the present study, image analysis wasperformed on a pixel-by-pixel basis and consisted of computing the differences in greylevels between an image recorded at a given time and a reference image correspondingto no filtering activity (i.e. worms withdrawn inside their tubes). These differences werethen stored as numerical objects in the memory board. At the end of a recording session,data were uploaded through the RS232 port of the interface board to a microcomputerwhere they were analysed using a set of post-treatment programs. These programsallowed the recovery of all differences due to the activity of each worm, including whenthe worms were filtering.
This system was calibrated for the study of filtering activity of D. arietina. Methodvalidation was achieved by comparing the filtering activities measured using theautomated video system with those observed by conventional video recordings. This

60 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
procedure showed that with a frame sampling frequency of 60 s, the automated videosystem was a convenient and suitable tool for studying the filtering activity in D.arietina.
2.2. Biological material and experimental set-up
Worms used during this work were collected by dredging (25 , depth , 30 m) eitherinside the bay of Banyuls-sur-Mer (428299235N, 3889500E) or 20 km north of this bay in
`front of the city of Argeles-sur-Mer (428349607N, 3849847E). Back at the laboratory,worms were placed on a thin layer (2 mm) of well-sorted fine sand in tanks (length(L) 3 width (W) 3 height (H): 2 3 1 3 0.2 m) supplied with running ambient sea water.All experiments conducted during this work, except those dedicated to the study of theeffect of hydrodynamics which involved the use of a recirculating flow tank, took placein an aquarium (L 3 W 3 H: 60 3 40 3 20 cm) filled with 30 l of filtered seawater (1.5mm). Batches of either 12 or 15 worms were placed upright in a thick layer (10 cm) ofsand so that the tube openings were 1 cm above the sediment surface. Observationsmade in the field and from undisturbed sediment cores sampled at the study sites showedthat D. arietina occurred either lying on or standing up in the sediment. The automatedvideo system localised (x, y coordinates in the picture) and characterised (time, durationand size of the image modification) any movement occurring inside the area of sedimentcovered by the video sensor. This process provides a rapid and convenient description ofthe filtering activity of D. arietina.
2.3. Description of the filtering activity
Standard filtration pattern (i.e. in absence of any stimulus) was characterized inseveral experiments run at different periods of the year and on several groups of wormsfrom different populations. We looked at filtration duration for each worm every hour todetermine if the filtering activity of Ditrupa arietina was either regular or irregularduring a 24-h period. Worms in batches of 12 or 15 specimens were handled as describedin Section 2.2. and their filtering activity analysed by the automated video sensor during24 h.
2.4. Monitoring of filtration activity
2.4.1. Comparison between populationsTwo experimental time series were run on individuals originating from two different
populations to assess whether there were temporal or spatial variations in filteringactivity. Each time series consisted of measurements once a month of filtering activity ofone batch of 15 freshly collected worms. The two time series were run from October1998 to September 1999 and, respectively involved worms collected in the bay of
`Banyuls-sur-Mer and off Argeles-sur-Mer. Worms were acclimated to laboratoryenvironment 24 h prior the beginning of the experiments. Each experiment lasted 5 hand was run at field temperature (near-bottom water temperature was measured once a
week by using a CTD Seabird SBE 19). The experiments were run on specimens that

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 61
were representative of the frequency size–distribution observed in the corresponding`population. The population of Argeles-sur-Mer was dominated by large individuals with
a mean tube length of 30 mm. The presence of epibiotics on the tubes and the absence ofsignificant increases in the mean length of the tubes during the entire time seriessuggested that the worms were in their second year of life (Medernach and Gremare,1998; Merdenach et al., 2000). The population in the bay of Banyuls-sur-Mer wasexclusively comprised of young individuals (,1 year old) with a mean tubes’ length of22 mm. These tubes always remained clear of any epibiotics and their mean lengthsteadily increased to reach a mean value of 27 mm by the end of the study. For theBanyuls-sur-Mer population, the presence of oocytes in the coelomic cavity of 150randomly collected worms was assessed fortnightly to determine whether the activity ofthe gill-crown went through changes during the breeding season.
2.4.2. Effect of worm ageAnother time series was run during 1997 on a sub-population (428299379N, 3889632E)
of the bay of Banyuls-sur-Mer. This sub-population presented was comprised exclusive-ly of 2-year-old individuals. The purpose of the experiment was to determine whetherand how the activity of the gill-crown was affected by worm age. Once a month, fromMarch to December, the filtration activity of 12 worms freshly collected at thecorresponding station was measured by using the automated video system. The age ofthe population was assessed indirectly by measuring changes in worm density and themean individual dry weight.
2.5. Effects of environmental factors
Preliminary studies on the filtering activity of D. arietina (i.e. observations madeduring the calibration and the validation of the automated video system) showed theexistence of strong inter-individual variability. A conventional experimental plan basedon replication was thus unsuitable to detect significant treatment effects. Therefore wesubmitted batches of worms to the successive changes in the values of the test parameterand then used pairwise comparison tests to infer statistically the effect of each treatmentfactor. The occurrence of rhythms, as well as synchronicity between individuals, in thefiltering activity were checked using spectral analysis.
2.5.1. Effect of the presence of food
2.5.1.1. Experiment 1 Worms were collected on March 1997 within the bay ofBanyuls-sur-Mer. In the laboratory, worms were placed in an aquarium supplied withunfiltered running seawater. One experiment was carried out on: 0, 1, 3, 8, 10, 14, 16,23, 35, 43, 52, and 58 days after worms collection. Each of these experiments consistedof determining the filtering activity of a batch of 12 worms that were successivelyconfronted with the absence (i.e. filtered 1.2-mm seawater) and then to the presence offood in the surrounding water. Confrontations lasted 5 h. The food, the flagellate
4 21microphyte Dunaliella tertiolecta, was provided at a concentration of 1310 cells ml .

62 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Changes in the filtering activity during the two periods were compared using Wilcoxonsigned-rank tests.
2.5.1.2. Experiment 2 Experiments were run during June 1997 on worms from the bay`of Banyuls-sur-Mer and during October 1997 on worms collected off of Argeles-sur-Mer
to assess the effect of food concentration on filtering activity. Batches of 12 worms weresuccessively exposed to increasing concentrations of D. tertiolecta (i.e. 0, 1000, 5000,
21and 10 000 cells ml ). During June, worms were exposed to each concentration for 8versus 4 h in October. Friedman two-way analysis of variance was performed to detectsignificant changes in the filtering activity with increasing food concentrations. Signifi-cant differences between matched samples were tested using Wilcoxon signed-rank tests.
3. Results
3.1. Description of the filtering activity
The filtering activity of Ditrupa arietina is presented in Fig. 2 for 15 worms collected`in February 1998 at the site off Argeles-sur-Mer. This example illustrates what was
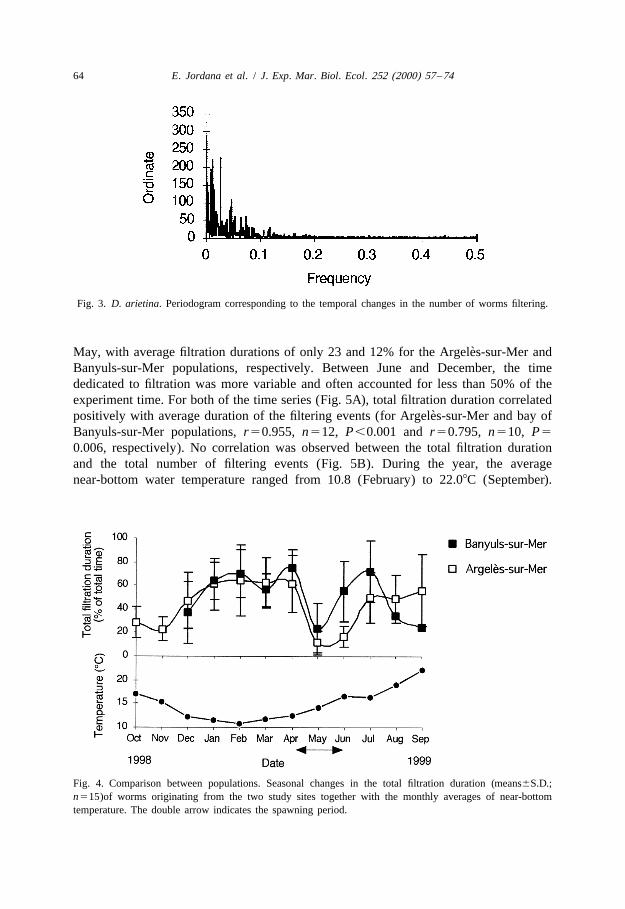
observed in experiments conducted to describe the standard filtering activity (i.e. inabsence of any stimulus) of D. arietina. The basic pattern of filtration in D. arietina (Fig.2A) consisted of a succession of active periods (i.e. gill-crown openings) and rest (i.e.gill-crown retracted inside the tube). Such patterns are described by the following threevariables: (i) the total filtration duration (i.e. total time during which the gill-crown waswide-open), (ii) the number of filtering events (i.e. number of times the gill-crown wentout), and (iii) the average duration of each filtering event (Fig. 2B). During thetime-course of the survey, gill-crowns stayed widely opened for 3 to 21 h (i.e. 12.5 and87.5% of the total experiment time), and worms filtered between 23 and 135 times ofwhich the average duration ranged from 2 to 48 min. The number of simultaneouslyfiltering worms (Fig. 2C) changed very frequently but was usually comprised betweenbetween six and nine and never exceeded thirteen. Spectral analysis (see correspondingperiodogram; Fig. 3) did not reveal any clear periodicity in temporal changes of thenumber of filtering worms. The cumulative filtration time (Fig. 2D) increased steadilyduring experiment. These results suggest that, on a daily basis, there was no particularrhythm in the filtering activity of D. arietina.
3.2. Monitoring of filtration activity
3.2.1. Comparison between populationsThe time series of total duration of filtration for worms originating from the two study
sites (Fig. 4) sites showed a significant date but not location effect (two-way ANOVA,P,0.001 and P50.281, respectively). There was also a significant interaction betweenthe two factors (P,0.001) which means that the date effect differed at the two sites.From January to April the filtration durations were rather high (around 60% of theexperiment time) and constant for both populations. The lowest values were observed in

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 63
`Fig. 2. D. arietina. Standard filtering activity in 15 worms collected in February 1998 off Argeles-sur-Mer. (A)Filtering pattern for a single worm (dark bars account for filtering). (B) Box plots showing total filtrationdurations, the number of filtering events and the average duration of the filtering events run by the batch ofworms (boxes encompass 25 and 75% quartiles; the central line represents the median and bars encompass95% limits of the obtainable values). (C) Temporal changes in the number of worms filtering. (D) Cumulatedfiltration time per worm.
64 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Fig. 3. D. arietina. Periodogram corresponding to the temporal changes in the number of worms filtering.
`May, with average filtration durations of only 23 and 12% for the Argeles-sur-Mer andBanyuls-sur-Mer populations, respectively. Between June and December, the timededicated to filtration was more variable and often accounted for less than 50% of theexperiment time. For both of the time series (Fig. 5A), total filtration duration correlated
`positively with average duration of the filtering events (for Argeles-sur-Mer and bay ofBanyuls-sur-Mer populations, r50.955, n512, P,0.001 and r50.795, n510, P5
0.006, respectively). No correlation was observed between the total filtration durationand the total number of filtering events (Fig. 5B). During the year, the averagenear-bottom water temperature ranged from 10.8 (February) to 22.08C (September).
Fig. 4. Comparison between populations. Seasonal changes in the total filtration duration (means6S.D.;n515)of worms originating from the two study sites together with the monthly averages of near-bottomtemperature. The double arrow indicates the spawning period.

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 65
Fig. 5. Comparison between populations. Relationships between total filtration durations and (A) averagedurations of each filtering event, and (B) numbers of filtering events, for the populations of Banyuls-sur-Mer
`(j) and Argeles-sur-Mer (h).
There was no correlation between near-bottom temperature and the activity of thegill-crown of D. arietina (P.0.05 for both population).
Temporal changes in the proportion of worms carrying oocytes (Fig. 6) shows that, inJanuary 1998, the number of gravid females accounted for 20% of the total population ofthe bay of Banyuls-sur-Mer. This proportion increased to 50% at the beginning ofMarch, suggesting that the whole population was sexually mature at this time (sex ratioin D. arietina is 1:1). Release of oocytes started during April and ended in July; most ofthe oocytes depletion occurred in April and May. Spawning period coincided with theperiod of decrease total filtration duration (Fig. 4).

66 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Fig. 6. Monitoring of the filtration activity. Temporal changes in the proportion of individuals carrying oocytesin the coelomic cavity. Values correspond to samples of 150 worms collected every 2 weeks from January toSeptember 1999 within the bay of Banyuls-sur-Mer population. The region in grey indicates the period ofspawning.
3.2.2. Effect of worm ageThe time series to assess changes in filtering activity in a declining population (Fig.
227A) showed a worm density close to 1500 ind m in May, that regularly decreased22during the survey to less than 200 ind m in December. Mean individual dry weights
confirmed that worms were old adults as the individual biomass remained constant´(Medernach and Gremare, 1998). Simultaneously (Fig. 7B), the total time spent by the
worms in suspension feeding decreased dramatically (one-way ANOVA, P,0.001). Bythe end of the survey worms were filtering on average less than 7% of the totalexperiment time. The number as well as the average duration of the filtering events alsochanged over time (one-way ANOVA, P,0.001, for each variable). They were drasticdecreases in total filtration durations resulting from the joint reduction of the number andthe average duration of the events of filtration. In December, worms had an average oftwo filtering events of which the mean duration lasted only 2 min.
3.3. Effects of environmental factors
3.3.1. Effect of the presence of food
3.3.1.1. Experiment 1 To determine effect of food on filtration activity, we first had toevaluate a potential date effect. Temporal changes in filtering activity (Fig. 8) showedthat the total filtration duration in the absence of food was significantly affected by the

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 67
Fig. 7. Monitoring of the filtration activity. Decreasing filtering activity in a declining population. (A)Temporal changes in density (j) and individual dry weight (h). (B) Effect of worm age on filtering activity.Box plots showing total filtration durations, the number of filtering events, and the average duration of thefiltering events run by the batch of worms (boxes encompass 25 and 75% quartiles; the central line representsthe median and bars encompass 95% limits of the obtainable values. Stars and dots indicate observations,respectively, outside the 95% limits and outside the 99% limits).

68 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Fig. 8. Effect of the presence of food. Temporal changes in the effect of the presence of food on the totalduration filtration for 12 batches of randomly selected worms. Values correspond to mean6S.D. (n512).
experiment date (n5141, df511, P,0.001). For both of the treatments, the lowestvalues were measured 35 days after the worm collection (i.e. on 20th May).
The characteristics of the 141 filtration record pairs (i.e. absence vs. presence of food;three records were unexploitable) (Fig. 9A) showed that the presence of food enhancedthe total time dedicated by the worms to filtration (Wilcoxon signed ranks test, n5141,P,0.001). In addition, the total duration of filtration following food addition correlated
2positively with values measured in filtered seawater (n5141, r 50.679, P,0.001). Theenhancement of the total duration of the filtration could reflect two kinds of changes inthe filtering behaviour of the worms: either an increase in the frequency of the filteringevents (Fig. 9B), or an increase in the average duration of filtration events (Fig. 9C).Pairwise comparison tests indicated that the presence of food led to the lengthening ofthe duration of individual events of filtration (Wilcoxon signed-ranks test, n5141,P50.001) rather than to an increase of the number of filtration events (Wilcoxonsigned-ranks test, n5141, P50.339).
Individual spectral analysis of filtering behaviour of the worms did showed neither aparticular rhythm in the filtering activity nor synchronicity in the activity betweenworms.
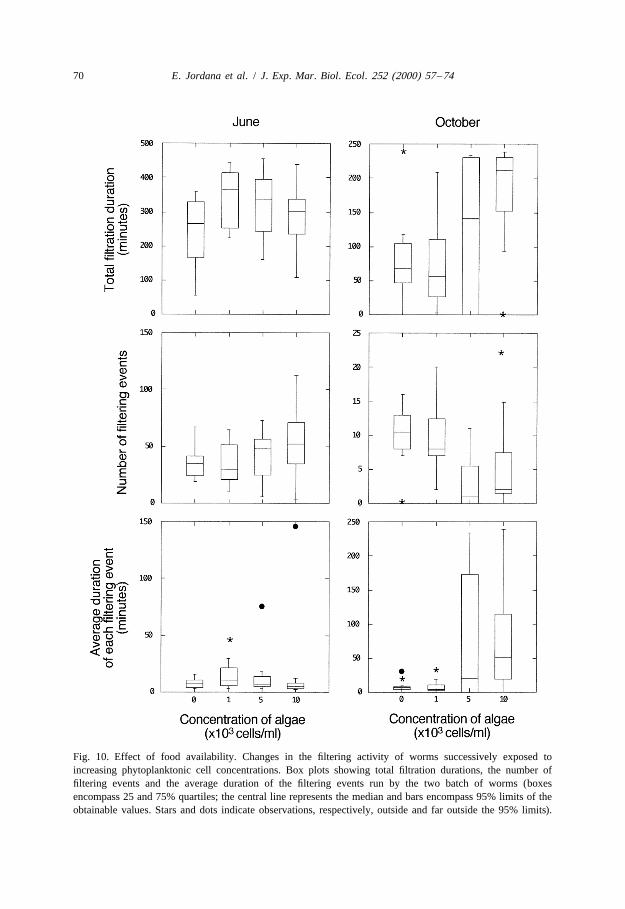
3.3.1.2. Experiment 2 In June, the average time spent filtering by the worms under eachof the food concentrations did not vary significantly (Fig. 10; Friedman two-way
21ANOVA, P50.296). Times ranged from 241 (no food) to 341 min (1000 cell ml ).The mean number of filtration events, as well as the average duration of these eventsremained unchanged for all cell concentrations (Friedman two-way ANOVAs, P50.139and, P50.331, respectively).
In October, the mean standard filtering activity (i.e. in the absence of food) was lowerthan in June (Fig. 10). The mean duration of the total filtration time ranged from 80 (no

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 69
Fig. 9. Effect of the presence of food. Individual effect of the presence of food on (A) the total duration offiltration, (B) the total number of filtering events, (C) the average duration of the filtering events. Straight linescorrespond to the first bisecting line (i.e. no changes in the filtering parameters).
70 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Fig. 10. Effect of food availability. Changes in the filtering activity of worms successively exposed toincreasing phytoplanktonic cell concentrations. Box plots showing total filtration durations, the number offiltering events and the average duration of the filtering events run by the two batch of worms (boxesencompass 25 and 75% quartiles; the central line represents the median and bars encompass 95% limits of theobtainable values. Stars and dots indicate observations, respectively, outside and far outside the 95% limits).

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 71
21food) to 173 min (10 000 cells ml ). Four worms did not start or stopped filtering forseveral hours, explaining the large dispersion of the data. In spite of such variability,there was a significant effect of food particle concentration on filtration duration(Friedman two-way ANOVA, P50.021). The average duration of the filtering eventswas also significantly affected by the concentration of food (Friedman two-wayANOVA, P50.024). Wilcoxon signed rank tests indicated that filtering events lastedlonger at the highest concentration than at the two lowest concentrations (Wilcoxonsigned-rank tests, P50.008 and P50.009) but that the number of filtering eventsremained unchanged (Wilcoxon signed-rank tests, P50.084 and P50.099).
4. Discussion
Results of the present study showed that there was a strong inter-individual variabilityin the filtering activity of Ditrupa arietina. Worms filtered neither continuously noraccording to a particular rhythm. Moreover, there was no synchrony in the filteringactivity between individuals. This trend was illustrated by the large dispersion of thedata around the average values.
4.1. Effects of life history
There was a seasonal pattern of variation in the filtering activity of D. arietina. Threemain periods were characterised in this study. In the first period, lasting from January toApril, total filtration durations were high and relatively constant. In the second period, inMay and June, there was a dramatic drop in the filtering activity. Lastly, in the thirdperiod, from June to December, the total time dedicated by the worms to filtration washighly variable. Such seasonal changes in the feeding activity already have been reportedin bivalves, and were related to changes either in the food availability (Newell andBayne, 1980) or in the water temperatures (Kamermans, 1994). In the present case, theactivity of the gill-crowns of D. arietina was independent of the water temperature, butnot of food availability (see paragraph below). During the first two periods, worms ofboth populations behaved similarly. Thus, filtering activity may be controlled by onestrong factor or even by a combination of several factors. Through the year (excepted inFebruary), the phytoplanktonic content of the waters of the bay of Banyuls-sur-Mer israther low and constant (Jacques, 1970; Jordana, personal communication). This suggeststhat food availability alone can not entirely explain the seasonal variations in thefiltration activity of D. arietina.
So, other factors must affect the filtering activity such as endogenous factors likereproductive activity. The filtering activity would first be high to insure gonad growthand then would drop drastically in May after the spawning period. At that time of theyear, worms appeared exhausted. They were thin, and lot of them had regenerating gillfilaments (Charles, personal observation). The drastic drop of the filtering activity in
`May was previously observed during 1997 on worms of the population of Argeles-sur-Mer (Jordana, unpublished data). The experiments run here to determine the effect of thepresence of food on these worms was to ascertain the cause of the occurrence of a slack

72 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
period of the filtering activity in May. During the third period (from June to December),the total duration of filtration varied with time but it also seemed to change according topopulation. Such differences can be related to local environmental conditions which maybe specific for both populations, but also to population aging. Ditrupa arietina has a2-year lifespan (Medernach et al., 2000). At the beginning of the study the population of
`Argeles-sur-Mer was mainly composed of 2-year-old worms (1,age#2 years old). Thispopulation aged during the course of the study. In June, their filtering activity was stillfairly low in comparison with that of younger worms (,1 year) of the bay ofBanyuls-sur-Mer. This difference in the filtering activity (Fig. 6) may be explained bythe effect of the senescence of one part of the population. In this case changes in filteringactivity can be related to the physiological state of the worms. As proportionally moreworms aged, the number of filtering events decreased and the duration of these eventswas reduced.
4.2. Effects of environmental factors
The experiments dealing with presence of food showed that the filtering activity of D.arietina was indeed affected by variations in external factors. The results of theseexperiments also showed that recent physiological state had a significant effect on theresponse of worms facing changes in their environment. Thus, according to the level ofthe standard filtering activity (i.e. in absence of stimulus), total filtration durationsincreased or remained stable.
The range of food concentrations tested here was similar to those experienced by theworms in the field (Jacques, 1970, Jordana et al. submitted). Our experiments showedthat worms kept their gill-crowns wide-open even when food concentration reached
4 211310 cells ml . However, no relationship was noticed between filtration activity andfood availability. When presence of food enhanced filtering activity, in most of the casesthis increase was associated with a lengthening of the average duration of the filteringevents. In suspension feeding polychaetes, filtration rate is assumed to be constant whengill-crown is wide-open (Riisgard and Ivarsson, 1990), suggesting that for the range offood concentrations tested in the present study, a decrease in the clearance rate withincreasing food concentration would be due to an overloading of the feeding system andnot to a reduction of the activity of the gill-crown. Davies et al. (1989) showed, foranother serpulid Ficopomatus enigmaticus, that ingestion rates increased with particle
21concentrations ranged from 1 to 10 mg ml . These results suggest that, during thepresent study in which particle concentrations were lower, clearance rates probablyremained unchanged and so ingestion rate of D. arietina increased steadily with foodconcentrations.
5. Conclusion
The filtering activity of D. arietina changed with time of the year but also accordingto the considered worm population. Our results suggested that the activity of thegill-crown was mainly controlled by endogenous factors such as worm aging and status

E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74 73
of sexual maturity. Environmental factors could also affect the filtering behaviour but, inthat case, the effect was largely dependent of the initial level of activity of the testedworms.
Acknowledgements
E. Jordana was supported by EEC doctoral fellowships (contract no. MAS3-CT96-5006 and MAS3-CT98-5063-PL980769).We greatly appreciated the helpful comments ofthe reviewers. This work was carried out within the framework of the Laboratoire
´Europeen des Sciences de la Mer. [RW]
References
Achituv, Y., Yamaguchi, T., 1997. Water pumping in the pendiculate barnacle Conchodermata auritum. J. Mar.Biol. Assoc. UK 77, 1073–1082.
Dales, P., 1957. Some quantitative aspects of feeding in sabellids and serpulids fan worms. J. Mar. Biol. Ass.UK. 36, 309–316.
Davies, B.R., Stuart, V., de Villiers, M., 1989. The filtration activity of a serpulid polychaete population(Ficopomatus enigmaticus Fauvel) and its effects on water quality in a coastal marina. Estuar. Coast. ShelfSci. 29, 613–620.
Foster-Smith, R.L., 1976. Some mechanisms for the control of pumping activity in bivalves. Mar. Behav.Physiol. 4, 41–60.´ ´Gremare, A., Amouroux, J.M., Vetion, G., 1998a. Long term comparison of macrobenthos within the softbottoms of the bay of Banyuls-sur-Mer (North-western Mediterranean Sea). Neth. J. Sea Res. 40, 281–302.´Gremare, A., Sarda, R., Medernach, L., Jordana, E., Pinedo, S., Amouroux, J.M., Martin, D., Nozais, C.,
¨Charles, F., 1998b. On the dramatic increase of Ditrupa arietina O.F. Muller (Annelida: Polychaeta) alongboth the French and the Spanish Catalan coasts. Estuar. Coast. Shelf Sci. 47, 447–457.
´Jacques, G., 1970. Aspects quantitatifs du phytoplankton de la region de Banyuls-sur-Mer (Golfe du Lion).` ´These de Doctorat des Sciences naturelles, Faculte des Sciences de Paris, pp. 210.
Jorgensen, C.B. (Ed.), 1966. Biology of Suspension-feeding. Pergamon Press, Oxford, p. 357.Kamermans, P., 1994. Similarity in food source and timing of feeding in deposit- and suspension-feeding
bivalves. Mar. Ecol. Prog. Ser. 104, 63–75.Leonard, A.B., 1989. Functional response in Antedon mediterranea (Lamarck) (Echinodermata: Crinoidea):
the interaction of prey concentration and current velocity on a passive suspension feeder. J. Exp. Mar. Biol.Ecol. 84, 81–103.
Newell, R.I.E., Bayne, B.L., 1980. Seasonal changes in the physiology, reproductive condition andcarbohydrates content of the cockle Cardium (5Cerastoderma) edule (Bivalvia: Cardiidae). Mar. Biol. 56,11–19.
´ ´ ´Medernach, L., Gremare, A., 1998. Comparaison de methodes d’evaluation de la production secondaire d’une´ `population de l’annelide polychete Ditrupa arietina (O.F. Muller). Oceanol. Acta 22, 337–352.
´Merdenach, L., Jordana, E., Gremare, A., Nozais, C., Charles, F., Amouroux, J.M., 2000. Population dynamics,¨secondary production and calcification of Ditrupa arietina O.F. Muller (Annelida: Polychaeta) within the
bay of Banyuls-sur-Mer (Western Mediterranean). Mar. Ecol. Prog. Ser. (in press).Merz, R.A., 1984. Self-generated versus environmentally produced feeding currents: a comparison for the
sabellid polychaete Eudistylia vancouveri. Biol. Bull. 167, 200–209.Riisgard, H.U., Ivarsson, N.M., 1990. The crown-filament pump of the suspension-feeding polychaete Sabella
penicillus: filtration, effects of temperature, and energy cost. Mar. Ecol. Prog. Ser. 62, 249–257.

74 E. Jordana et al. / J. Exp. Mar. Biol. Ecol. 252 (2000) 57 –74
Riisgard, H.R., Poulsen, L., Larsen, P.S., 1996. Phytoplankton reduction in near-bottom water caused byfilter-feeding Nereis diversicolor: Implications for worm growth and population grazing impact. Mar. Ecol.Prog. Ser. 141, 47–54.
Sanford, E., Bermudez, D., Bertness, M.D., Gaines, S.D., 1994. Flow, food supply and acorn barnaclepopulation dynamics. Mar. Ecol. Prog. Ser. 104, 49–62.
Shumway, S.E., Bogdanowicz, C., Dean, D., 1988. Oxygen consumption and feeding rates of the sabellidpolychaete, Myxicola infundibulum (Renier). Comp. Biochem. Physiol. 90, 425–428.
Thorin, S., Bourdages, H., Vincent, B., 1998. Study of siphon activity in Mya arenaria (L.) in the intertidalzone by means of an underwater video camera. J. Exp. Mar. Biol. Ecol. 224, 205–244.
Vedel, A., Andersen, B.B., Riisgard, H.U., 1994. Field investigations of pumping activity of the facultativelyfilter-feeding polychaete Nereis diversicolor using an improved infrared phototransducer system. Mar. Ecol.Prog.Ser. 103, 91–101.