evolutionary algorithms based on non-darwinian theories of evolution: the peircean perspective
DESCRIPTION
Historically, Evolutionary Algorithms (EA) have been important for the Evolutionary Computation (EC) community for two primary reasons: 1) As a simulation of evolutionary processes, the way they happen in the natural world, and 2) using them to solve computationally hard problems, including optimization. With the passage of time EA have become increasingly focused on function optimization. Given this narrowing of vision in the EC community, it is worth revisiting a paper written in 1997 by Hans-Paul Schwefel on the future challenges for EC. In that paper the author argues that the more an algorithm models natural evolution at work in the universe, the better it will perform (even in terms of function optimization). The present dissertation tests Schwefel's hypothesis by designing an EA based on the evolutionary theory of Charles Sanders Peirce, founder of American school of thought known as pragmatism. Peirce's theory not only accounts for biological evolution on earth (as other theories of evolution do) but also offers an account of global, cosmological and universal evolution. In going beyond just biological evolution, Peirce's theory of evolution meets the criteria suggested by Schewefel in his 1997 paper. We follow this course because of three primary reasons. Firstly, Peirce has not been seriously tested in EC, although there have been EA based on other theories and sub-theories. Secondly, Peirce's universal theory, by not being restricted to biological evolution alone, qualities for Schwefel's hypothesis, perhaps more than most other theories that have already been modeled algorithmically. But most importantly because, in experimental terms, it lets us make an original claim that Peirce's insights are useful in improving the existing EA in computer science, as Peircean EA can potentially solve some of the major problems in this area such as the loss of diversity, stagnation, or premature convergence. In this thesis, we have provided a novel algorithm based on Peirce's theory of evolution, initiated it for the class of optimization problems, and tested it extensively against a benchmark set of mathematical problems of varying dimensions and complexity. Comparative results with classical and advanced EA form another significant part of the thesis, and help in strengthening the viability of Schwefel-Peirce hypothesis for EC. Besides the experimental and empirical findings, the theoretical aspects of Peircean EA's performance are also brought under discussion in the light of Holland's schema theorem and disruption analysis. It is then claimed that Peircean arrangements for an EA help minimize the disruptions potentially caused by the crossover operator and consequently help in the survival and continued evolution of good schemas.TRANSCRIPT

Evolutionary Algorithms based on non-Darwinian
Theories of Evolution:
The Peircean Perspective
PhD Thesis
Junaid Akhtar
2004-03-0019
Advisor: Dr. Mian Muhammad Awais
Co-Advisor: Dr. Basit Bilal Koshul
Department of Computer Science
School of Science and Engineering
Lahore University of Management Sciences

Dedicated to everyone and everything that brought me together

Lahore University of Management Sciences
School of Science and Engineering
CERTIFICATE
I hereby recommend that the thesis prepared under my supervision by Junaid Akhtar titled Evo-
lutionary Algorithms based on non-Darwinian Theories of Evolution: The Peircean
Perspective be accepted in partial fulfillment of the requirements for the degree of doctor of phi-
losophy in computer science.
Dr. Mian M. Awais (Advisor)
Recommendation of Examiners’ Committee:
Name Signature
Dr. Mian Muhammad Awais ——————————————
Dr. Basit B. Koshul ——————————————
Dr. Asim Karim ——————————————
Dr. Shafay Shumail ——————————————

Acknowledgements
It would not be unfair to thank Allah foremost at this point. I believe I am grateful to Him
mostly for giving me ideas that guide me. I want to thank my parents and family, that is the
least I can do in return for giving me their full support and peace of mind; my wife for motivating
me towards the completion of this milestone; my advisors, Dr. Mian Muhammad Awais and
Dr. Basit Koshul, for their most valuable advises over the years. And my friends for making
the time so memorable, especially Faheem, Umar Suleman, Zeeshan, Jahan, and Saqib. Higher
Education Commission, Pakistan, and Lahore University of Management Sciences, Pakistan, are
also acknowledged for the funding.

Contents
Abstract xiii
1 Introduction 1
1.1 Investigation 1: Research Directions within EC . . . . . . . . . . . . . . . . . . . . . 2
1.2 Investigation 2: Challenges Facing EC . . . . . . . . . . . . . . . . . . . . . . . . . . 6
1.3 Investigation 3: New Developments within Evolutionary Sciences . . . . . . . . . . . 8
1.4 Contributions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
1.5 Thesis Roadmap . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 12
2 Darwinian Evolutionary Algorithms: From Natural Reality to Problem Solving 13
2.1 Darwin’s Stature and Contribution in Science . . . . . . . . . . . . . . . . . . . . . . 14
2.1.1 Darwin, the Forward-looking Genius . . . . . . . . . . . . . . . . . . . . . . . 15
2.1.2 Darwin, the Backward-looking Intellect . . . . . . . . . . . . . . . . . . . . . 16
2.2 The Road from Darwin to Post Modern Synthesis . . . . . . . . . . . . . . . . . . . 20
2.3 From Evolutionary Theory to Evolutionary Computation . . . . . . . . . . . . . . . 23
2.3.1 Classical Evolutionary Algorithms . . . . . . . . . . . . . . . . . . . . . . . . 24
2.3.2 Classic Algorithms in EC are Foundationally Darwinian . . . . . . . . . . . . 25
2.4 Instantiation of an EA with an Optimization Problem . . . . . . . . . . . . . . . . . 27
2.5 Non-Classical Variant EAs . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
2.5.1 Non-Darwinian-type Ideas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 31
2.5.2 Peirce-type Ideas . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 32
v

3 Peircean Theory of Evolution 37
3.1 Understanding Peirce’s Framework through Evidence . . . . . . . . . . . . . . . . . . 40
3.1.1 Evidence for Peirce’s Thirdness in the biological world . . . . . . . . . . . . . 40
3.1.2 Evidence for Peirce’s Firstness in the biological world . . . . . . . . . . . . . 44
3.2 Understanding Peirce’s Evolutionary Framework through Definitions . . . . . . . . . 45
3.2.1 Firstness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
3.2.2 Secondness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 46
3.2.3 Thirdness . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 47
3.3 Understanding Peirce’s Evolutionary Framework through a Possible Synthesis . . . . 48
3.3.1 Definition :: Peircean Evolutionary Theory . . . . . . . . . . . . . . . . . . . 50
3.4 Nature of Chance and Laws in Light of Modern Physics and Cosmology . . . . . . . 50
3.4.1 Heisenberg on the Nature of Reality . . . . . . . . . . . . . . . . . . . . . . . 51
3.4.2 Weyl on the Historical understanding of Reality . . . . . . . . . . . . . . . . 51
3.4.3 Holton Concludes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53
4 Peircean Framework for Evolutionary Algorithms 57
4.1 Principles derived from Peirce’s theory for Evolutionary Algorithms . . . . . . . . . 57
4.1.1 Firstness as Spontaneity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
4.1.2 Secondness as Necessity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 58
4.1.3 Thirdness as Generalizing tendency . . . . . . . . . . . . . . . . . . . . . . . 58
4.1.4 Dynamic System . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 59
4.1.5 Reality at Multiple Levels . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 60
4.2 Peircean Evolutionary Algorithm . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 63
4.3 Comparison of Peircean EA with Related EA . . . . . . . . . . . . . . . . . . . . . . 67
4.3.1 IDEA 1 :: Diversity Retaining Measures . . . . . . . . . . . . . . . . . . . . . 68
4.3.2 IDEA 2 :: Distributed Population Dynamics . . . . . . . . . . . . . . . . . . 68
4.3.3 IDEA 3 = IDEA 1 + IDEA 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . 69
vi

5 Experimental Evidence 71
5.1 Experimental Setup . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
5.2 Results . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76
5.3 Results on an Extended Benchmark . . . . . . . . . . . . . . . . . . . . . . . . . . . 81
6 Analyses 85
6.1 Stagnation Analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 85
6.2 Cluster Analysis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 86
6.3 Schema Theorem . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88
6.3.1 Clusters as Schema - Effects on Disruption Analysis . . . . . . . . . . . . . . 90
6.4 Summary . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 91
7 Future of EC & our Conclusions 93
7.1 Hans-Paul Schwefel’s Future Directions for EC . . . . . . . . . . . . . . . . . . . . . 93
7.1.1 Equivalence between Peircean Framework and Schwefel’s Future Directions . 95
7.2 Kenneth De Jong’s Agenda for the 21stCentury and Peirce . . . . . . . . . . . . . . . 98
7.3 Conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 100
A Cooperation vs. Competition in Evolution 103
A.1 Cooperation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 103
A.1.1 Hypothesis 1: Only selfishness is real, cooperation is not . . . . . . . . . . . . 105
A.1.2 Hypothesis 2: Cooperation is as real as Selfishness . . . . . . . . . . . . . . . 107
vii

viii

List of Figures
2.1 Flow chart for Classical Evolutionary Algorithms . . . . . . . . . . . . . . . . . . . . 25
2.2 Image registration: A 2-D problem . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
2.3 Representation of EA Individuals for Image Registeration Problem . . . . . . . . . . 28
2.4 Crossover Operator’s Procedure - Parents 1 and 2 produce Children 1 and 2 . . . . . 30
4.1 Hypothetical 2D search space depicting a cluster formation within the population . . 61
4.2 Algorithm 1: Pseudo code for Peircean Evolutionary Algorithm . . . . . . . . . . . . 64
4.3 Algorithm 2: Pseudo-code for intra-cluster evolution . . . . . . . . . . . . . . . . . . 66
4.4 Algorithm 3: Pseudo-code for inter-cluster evolution . . . . . . . . . . . . . . . . . . 67
5.1 Multiple views of function f2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
5.2 Multiple views of function f3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 73
5.3 Multiple views of function f4 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 74
5.4 Convergence comparison: P-EA vs. D-EA . . . . . . . . . . . . . . . . . . . . . . . . 79
5.5 Extended Benchmark Functions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 82
6.1 Cluster Analysis: Effect of clustering on stagnation using Schwefel’s function . . . . 88
ix

x

List of Tables
4.1 Relationship of Peircian principles at different levels of algorithmic reality . . . . . . 63
5.1 Benchmark mathematical functions . . . . . . . . . . . . . . . . . . . . . . . . . . . . 72
5.2 Parameters for P-EA and D-EA . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 75
5.3 Compiled results for 50 runs of D-EA on f1, f2, f3 and f4, with stop-count = 500 . . 77
5.4 Compiled results for 50 runs of P-EA on f1, f2, f3 and f4, with stop-count = 500 . . 78
5.5 P-EA on f4 with varying stop-counts . . . . . . . . . . . . . . . . . . . . . . . . . . . 79
5.6 P-EA compared with other EA on f3 [30-D] . . . . . . . . . . . . . . . . . . . . . . . 80
5.7 P-EA compared with other EA on f4 [30-D] . . . . . . . . . . . . . . . . . . . . . . 81
5.8 Comparison between Peircean-EA and Darwinian-EA . . . . . . . . . . . . . . . . . 83
6.1 Number of generations before the population stagnates,20 experiments . . . . . . . . 86
7.1 Relation between Peircean framework and Schwefel’s future challenges . . . . . . . . 97
xi

xii

Abstract
Historically, Evolutionary Algorithms (EA) have been important for the Evolutionary Computa-
tion (EC) community for two primary reasons: 1) As a simulation of evolutionary processes, the
way they happen in the natural world, and 2) using them to solve computationally hard problems,
including optimization. With the passage of time EA have become increasingly focused on function
optimization. Given this narrowing of vision in the EC community, it is worth revisiting a paper
written in 1997 by Hans-Paul Schwefel on the future challenges for EC. In that paper the author
argues that the more an algorithm models natural evolution at work in the universe, the better
it will perform (even in terms of function optimization). The present dissertation tests Schwefel’s
hypothesis by designing an EA based on the evolutionary theory of Charles Sanders Peirce, founder
of American school of thought known as pragmatism. Peirce’s theory not only accounts for bio-
logical evolution on earth (as other theories of evolution do) but also offers an account of global,
cosmological and universal evolution. In going beyond just biological evolution, Peirce’s theory of
evolution meets the criteria suggested by Schewefel in his 1997 paper. We follow this course be-
cause of three primary reasons. Firstly, Peirce has not been seriously tested in EC, although there
have been EA based on other theories and sub-theories. Secondly, Peirce’s universal theory, by
not being restricted to biological evolution alone, qualifies for Schwefel’s hypothesis, perhaps more
than most other theories that have already been modeled algorithmically. But most importantly
because, in experimental terms, it lets us make an original claim that Peirce’s insights are useful in
improving the existing EA in computer science, as Peircean EA can potentially solve some of the
major problems in this area such as the loss of diversity, stagnation, or premature convergence. In
this thesis, we have provided a novel algorithm based on Peirce’s theory of evolution, initiated it
xiii

for the class of optimization problems, and tested it extensively against a benchmark set of math-
ematical problems of varying dimensions and complexity. Comparative results with classical and
advanced EA form another significant part of the thesis, and help in strengthening the viability of
Schwefel-Peirce hypothesis for EC. Besides the experimental and empirical findings, the theoretical
aspects of Peircean EA’s performance are also brought under discussion in the light of Holland’s
schema theorem and disruption analysis. It is then claimed that Peircean arrangements for an EA
help minimize the disruptions potentially caused by the crossover operator and consequently help
in the survival and continued evolution of good schemas.
xiv

Chapter 1
Introduction
We had been in correspondance with Hans-Paul Schwefel, one of the founders of Evolution Strate-
gies, regarding the design of Evolutionary Algorithms (EA) based on Charles Sanders Peirce’s
theory of evolution. All of a sudden he asks about the relevance of a “19th” century mathemati-
cian and evolutionary philosopher for the 21stcentury evolutionary sciences, especially Evolutionary
Computation (EC).1 The answer to Schwefel’s pertinent question seemed deceptively simple at first,
however, this entire thesis, in many ways, became an address to that sole question.
The simple answer is that Peirce is perhaps more necessary for evolutionary sciences today than
he was in the 19thcentury. But to reach to that conclusion or to answer the question satisfactorily,
there are three separate paths that would need to be investigated and developed towards an eventual
convergence. One path investigates the historical developments within EC, to make a legitimate
room for exploring Peircean ideas for an EA. Second path investigates the challenges faced by the
EC community as noted in the literature, and shows how Peircean framework has the potential
to act as a legitimate solution to those challenges. Having worked out the relevancy of Peirce
for 21stcentury EC specifically, the third path investigates the developments in the evolutionary
sciences generally, especially the cutting edge fields of biology. It is earnestly hoped that towards the
end of these three investigations it can be concluded, that the relevant research communities within
1Actual correspondance text: “I assume that not many people (and reviewers) are familiar with Pierce’s ideasabout evolution that may seem to many people some kind of outfashioned. He is better known as mathematician,mainly having done his work in the 19thcentury (we are in the 21stnow).”
1

the broader evolutionary sciences and especially EC are ripe for evolving their theoretical principles
to that of the Peircean evolutionary framework, rather than sticking to even older 19thcentury ones.
1.1 Investigation 1: Research Directions within EC
John Holland’s motivation behind his pioneering Genetic Algorithms (GA) model was to simulate
biological adaptive systems (Holland, 1975). In other words, Holland sought to model biological
evolution as proposed in Darwin’s theory. After Holland though, his students became increasingly
focused on designing GA for solving optimization problems (Goldberg, 1989). While the practical
need for optimization in GA is indeed important, it was only a marginal concern in Holland’s
original GA. Because his GA sought to model evolution in the natural world, Holland had to keep
in view the fact that for complex adaptive systems “improvement is usually much more important
than optimization.” This is an important point to keep in mind becasue - as (De Jong, 1993) notes:
“There is a subtle but important difference between ‘GAs as function optimizers’ and ‘GAs are
function optimizers’.” De Jong goes on to point out that there are important insights to be had
when this difference is understood and its implications are taken into account in developing GA
(and we may add EA in general).
This sentiment has been echoed in slightly different terms by two other pioneers in EA; (Schwefel,
1997) notes that “organic evolution certainly does not only aim at finding static optima just once
and with ultimate precision. Organic evolution happens within an ever-changing environment,
where evolvability is more important than precision”; Lawrence J. Fogel, pioneer in Evolutionary
Programming notes in (Back et al., 1997a, Section H1.2) that even though, “the solution of complex
engineering problems is important, but the use of evolutionary algorithms need not be restricted
to mere function optimization. The methods can also be used to gain an understanding of how
competitive or cooperative agents may interact given a variety of different available resources and
purposes.”
David B. Fogel aptly notes in the introduction of his Handbook of Evolutionary Computation
that “efforts in evolutionary computation commonly derive from one of four different motivations:
improving optimization, robust adaptation, machine intelligence, and facilitating a greater under-
2

standing of biology” (Back et al., 1997a, Section A1.1). On the one hand, this clearly indicates that
the spirit of EC is multi-faceted and cannot be reduced in its entirety to function optimization, or
any of the other three motivations for that matter. But at the same time it can lend itself to the
view that there is an either-or situation for the EC community–either one can be in EC to improve
function optimization or to understand the processes of natural evolution better. The two tasks
appear to be independent of each other and do not seem to be meaningfully related.
Perhaps it is because of this sectional view of EC that over the years the practical focus in EC is
increasingly “reduced” to factors such as efficiency, engineering applications, and standardizations.2
A careless reading of the foregoing could be taken as a suggestion that the importance of function
optimization or its efficiency in EA is being trivialized. We are, however, arguing something
very different! What is being suggested is that function optimization and its efficiency can be
enhanced by recognizing that there is a direct relation between understanding the natural processes
of evolution in greater detail and improvement in the working of evolutionary algorithms. In the
words of David B. Fogel:
Our challenge is, at least in some important respects, to not allow our own biases
to constrain the potential for evolutionary computation to discover new solutions to
new problems in fascinating and unpredictable ways. However, as always, the ultimate
advancement of the field will come from the careful abstraction and interpretation of
the natural processes that inspire it. (Back et al., 1997a, Section A1.1.6)
While the empirical reality of evolution occurring in nature is equally accessible to the entire
scientific community, yet slightly different understanding and perspective in theorizing the natural
processes have had different consequences. This consequential effect can be illustrated using data
from EC as well. Memetic algorithms are based on the notion of cultural evolution (Caraffini et al.,
2013; Moscato, 1989), inspired by Richard Dawkins’ proposal of meme as a unit of information that
reproduces itself as people interact and exchange ideas (Dawkins, 1976). Horizontal gene transfer
that takes place in bacterial organisms has inspired the design of specialized crossover operators
2Task Force on Future Directions in Evolutionary Computation (FDEC) had been part of the EvolutionaryComputation Technical Committee (ECTC), IEEE Computational Intelligence Society (CIS). The TF held an annualWorkshop as part of the IEEE Congress on Evolutionary Computation
3

that have spawned a new class of evolutionary algorithms called pseudo-bacterial genetic algorithms
(Balazs and Koczy, 2012; Nawa and Furuhashi, 1998). Lamarckian conception of evolution and
Baldwin effect on learning have been used by hybrid EA, and are reportedly more efficient than
their Darwinian counterparts (Fuhrmann et al., 2010; Mitchell and Taylor, 1999; Yuan et al.,
2010). Island Model EA are based on Eldridge and Gould’s punctuated equilibria theory (Segura
et al., 2011; Skolicki, 2005; Srinivasa et al., 2007), where subpopulations independently evolve while
infrequent migrations take place using some communication topology. There are Diffusion Model
EA as well which are based on population genetics idea of local (not global) selection and crossover.
Genetic makeup spreads from individual to population levels based on the diffusion model (Back
et al., 1997a; Jaimes and Coello, 2009). They are designed for parallel processing with one individual
per processor where the selection of parents is dependent on the underlying parallel architecture,
usually SIMD (Single Instruction, Multiple Data streams). The Genetic Algorithms itself is John
Holland’s “adaptation” of Darwin’s theory of evolution (Holland, 1975). This brief detour serves to
underscore the point that a different interpretation of natural evolution results in a different model
of EA.
According to Jacques Monod, Darwin’s theoretical explanation for evolution is an exquisite
mix of “chance and necessity” (Monod, 1971). In non-philosophical terms it is a combination of
a variety of chances and a variety of laws. In order for Darwinian evolution to work it takes as a
given, not only these two agents, but ironically the first batch of replicating life as well. Being a
naturalist, Darwin did not make an attempt to try and relate the two apparently warring agents
(chance & necessity), or how they “evolved” themselves before playing a role in the evolution of
the universe and its living forms. However, there is one man that did that after Darwin.
Charles Sanders Peirce, the 20th century evolutionary pragmaticist, has made major contribu-
tions to numerous fields such as logic and philosophy of science, formal and mathematical logic,
topology, linguistics, epistemology and semiotics. When Peirce looked at the empirical effects of
evolution, he came to the conclusion that there are three (not two) types of phenomenon, three basic
categories that are operative in the universe. First, chance; Second, necessity; Third, habit-taking.3
3Peirce’s use of habits is different from its Lamarckian usage: “For Peirce, habits are not provisional adaptiveresponses to fluctuating environmental conditions; they are steps on the universal road from indeterminacy to law,
4

From this, Peirce built a philosophically intricate system, and for its heart he installed semiosis, his
theory of signs, which attempts at describing the inter-relationship of his three categories. Stressing
unconditionally on the irreducibility of his semiotic triad, he said:
....by “semiosis” I mean, an action, or influence, which is, or involves, a cooperation of
three subjects, such as a sign, its object, and its interpretant, this tri-relative influence
not being in any way resolvable into actions between pairs. (Peirce, 1958, 5.484)
As far as the relation between a sign and its object is concerned, a sign, for Peirce, does not
properly function or signify outside the specific context of this triadic relation. This conception is
at odds with theories of signification that rely on a dyadic (or two part) relationship between signs
and the objects they signify. For Peirce, semiosis of processes of signification operate only when a
sign is considered in its triadic form, i.e. when a sign is a representation of its object, such that it
produces or modifies its interpretant.4 This effectively turns the interpretant into another sign of
the same object, and thus helps enable further interpretations. This dynamic semiotic process is
the cause for evolution and growth of meaning. It implies that organic matter learns to engage with
the world, not simply by forming ideas in response to stimuli, but by forming habits of responding
to non-organic material. For Peirce, habits are the ultimate interpretants of the world’s signs, and
he viewed habit-formation as the physiological manifestation of the sign-taking capacity (Ochs,
1993, pg. 68).
Mapping Peirce’s semiotic language on to his phenomenology it can be argued that evolution-
ary growth requires a cooperation of three evolutionary agencies; and for Peirce absolute chance,
mechanical necessity, and tendency to take habits are a sign that his three categories, Firstness,
Secondness and Thirdness are severally operative in the cosmos. Peirce shows that in logic they are
represented as beginning, end, and process. In psychology they are feelings, reaction-sensations,
and thought or reason. In the case of biology Peirce extends the almost linear two step evolutionary
process of random variation followed by natural selection into a non-linear triadic process. He says,
a road traveled by objects as well as by organisms...Habit-taking is a plastic faculty. The peculiar characteristic ofhabit is: “not acting with exactitude” ” (Menand, 2001, pg. 365)
4For Peirce, the interpretant can mean more than one thing: (a) The context in which a Sign relates to an Object(b) The relationship that an Object is able to establish with its context (Interpretant), because of Objects relationshipwith Sign (c) The potential consequences of the relationship of Sign with Object
5

First is the principle of individual variation or sporting; Second, the principle of heredity transmis-
sion; and Third, the process whereby the accidental characteristics become fixed (including, but
not limited to the elimination of unfavorable characters by natural selection.) (Peirce, 1958, 6.32)
Does the foregoing discussion have any implications for the EC community? In other words,
when the underlying theory of evolution evolves from a serial two-ness to a dynamic three-ness,
how does that affect the EC models consequently? Our work is an answer to this question. Taking
the lead from the pioneers in EC, our goal is to develop a framework for EA based on a theory
that offers a more intricate and explanatory account of the natural evolutionary process. Towards
this end, we are proposing a new framework for EA based on Peirce’s semiotic theory of evolution.
We have tested it on a benchmark of mathematical problems and compared the results with other
EAs.
1.2 Investigation 2: Challenges Facing EC
It is slightly ironic that our search for answering Schewefel’s concern regarding Peirce’s relevance
to EC leads us straight to Schwefel’s own research agenda. He fully understands the necessity
of developing a model which is closer to natural reality. In his essay “Challenges to and Future
Development of EA” he says:
Current evolutionary algorithms are certainly better models of organic evolution. Nev-
ertheless, they are still far from being isomorphic mappings of what happens in nature.
In order to perform better, an appropriate model of evolution would have to comprise
the full temporal and spatial development on the earth (a real global model) if not
within the whole universe. We must be more modest in order to understand at least a
little of what really happens – as always within natural sciences. (Schwefel, 1997)
We will refer to this passage as Schwefel’s hypothesis. Our call for revisiting Schwefel’s
research agenda is not because we are interested in the fulfillment of the initial promises of EC,
in and of themselves, or that we are not inclined towards looking at EA as function optimizers.
On the contrary, we bring to attention those unfulfilled promises only since they can potentially
6

help improve the function optimization capabilities of EA as well. The reason we chose Schwefel
to make a case is because of the clarity with which he has laid bare the relation between the two
things that are apparently viewed as being largely unrelated - performance of an EA and natural
understanding of reality. Some of the other future challenges according to Schwefel are:
1. Evolvability more important that precision
2. Dynamic interaction of agents
3. Multiple selection criteria
4. Cooperation as important as competition
5. Incorporation of social learning and epigenetic factors
If Schwefel had left out some open challenges to EA, Kenneth De Jong, the founding editor
of the premier Evolutionary Computation journal, while presenting a history of the field of GA
concludes with an extended “agenda for the 21stcentury” (De Jong, 2005). Some of the highlights
of this agenda are:
1. Developing a more general EC/EA framework
2. Decentralized and Speciation models
3. Self adapting and coevolutionary systems
4. Incorporating more biology into EA, especially Lamarckian ideas
5. Using EA to further our evolutionary understanding
In general our work is about revisiting Schewefel’s and De Jong’s research agenda for the
21stcentury. In particular it is about testing Schwefel’s hypothesis. It is in the pursuit of this end
that we introduce the evolutionary theory of Charles Sanders Peirce–the evolutionary philosopher,
mathematician, semiotician, and scientist par excellance (Peirce, 1958). As we saw above, Schwe-
fel’s hypothesis asks for a model of evolution that goes beyond biological evolution and encompasses
7

global, cosmological and universal evolution. Typically evolution has been confined to biological
processes, which help only explain the last few billion years of development within the universe.
Being a through-going evolutionary philosopher (and not a mere naturalist) Peirce sought to un-
derstand not only biological life, but also the emergence of all inanimate and animate matter as
well as the laws of nature shaping their behavior, in evolutionary terms. This dissertation tries
to demonstrate how Peircean principles have the potential to fit the bill for the above-mentioned
21stcentury research agenda for EA in general, and solve the problem of stagnation in particular,
caused by the loss of diversity in the population at early stages of classical EA paradigms (Deb,
2001; Fogel, 1994). Our claim is that the Peircean framework for EC achieves this naturally, with-
out having to resort to arbitrary and artificial arrangements that are commonly employed just
for demonstrating an improvement in the results (Lozano et al., 2008; Mahfoud, 1995; Sareni and
Krahenbuhl, 1998). In other words, Peirce can potentially provide the much needed theoretical
consistency to the future developments within EC.
1.3 Investigation 3: New Developments within Evolutionary Sci-
ences
Lawrence J. Fogel reminds us of the greatest challenge and promise the EC community had to
fulfill:
Perhaps the greatest challenge facing evolutionary computation is its use as a means
for gaining a greater understanding of natural evolution. This has been the promise of
the efforts of artificial life, but like many other such promises throughout the course of
computer science, they have been left mainly unfulfilled. (Back et al., 1997a, Section
H1.2)
Moreover, identifying reductionism as a problem for scientific understanding of complex pro-
cesses, he said:
An important step forward could be realized if attempts to perform such credit assign-
ment and related schema analysis in complex systems were abandoned in favor of more
8

holistic understandings of how selection acts on complex sets of behaviors in concert,
rather than in isolation. Just as no general understanding of the physics of flight can
come from assigning credit to feathers or flapping wings, no general understanding of
complex adaptive systems can come from piecemeal analysis of their ‘genes’. Refocus-
ing attention on ‘organisms’ rather than ‘genes’ represents a compelling and promising,
although old, direction for further investigation.
There is a surprising similarity between what Fogel had to say to his own community and what
(Goldenfeld and Woese, 2007), veterans in biology had to say to their own:
In the last several decades we have seen the molecular reductionist reformulation of
biology grind to a halt, its vision of the future spent, leaving us with only a gigantic
whirring biotechnology machine. Biology today is little more than an engineering dis-
cipline. Thus, biology is at the point where it must choose between two paths: either
continue on its current track, in which case it will become mired in the present, in
application, or break free of reductionist hegemony, reintegrate itself, and press for-
ward once more as a fundamental science. The latter course means an emphasis on
holistic, “nonlinear,” emergent biology–with understanding evolution and the nature of
biological form as the primary, defining goals of a new biology.
Fogel and Woese share with their respective community the potential consequences of working
on a reductionist paradigm in evolutionary sciences. Physics, the mother of natural sciences has
an advantage of hindsight that it can lend to the relatively younger evolutionary biology and even
younger EC. Having witnessed the transition from deterministic Newtonian physics to probabilistic
Quantum mechanics, the physicist-philosopher (Bohm, 1969) makes an interesting observation:
It does seem odd ... that just when physics is ... moving away from mechanism, biology
and psychology are moving closer to it. If the trend continues ... scientists will be
regarding living and intelligent beings as mechanical, while they suppose that inanimate
matter is too complex and subtle to fit into the limited categories of mechanism.
9

Fortunately, biology has already begun to see a movement away from the classical mechanistic
conceptions. Works of cutting edge groups like epigenetics, systems biology, symbiogenesis, emer-
gent biology and biosemiotics are at the frontiers and taking the same direction as seen in the
movement within physics. Unfortunately there is no equivalent EC group working in tandem with
the above-mentioned counterparts in biology. We would like to introduce this research work as an
effort to bring EC at par with the frontiers of evolutionary biology.
This new inertia demands explanation of old phenomena using a new perspective. We introduce
the framework of Charles Sanders Peirce as a non-reductionist explanation for evolution. The
classical Darwinian conception and its modern synthesis are based on two agents at the fundamental
level: blind chance and mechanistic laws. While Peirce acknowledges the role of both of these agents
in the evolutionary process, he identifies one additional agent, i.e. generalizing tendency. The
introduction of this third agent appropriates subtle but significant changes in our understanding
of the character of both chance and laws. In Peirce’s triadic scheme chance does not always follow
a blind uniform distribution but can follow distributions similar to those followed by laws of large
numbers. Laws on the other hand, are not encapsulated by the metaphor of machines, perfect and
precise in their output, but show tendencies of habits and hence variation and exceptions in their
delivery. This allows Peirce to describe the emergence of both chance and law as the outcomes
of an evolutionary process in the universe rather than as eternal givens outside the universe -
that work mysteriously. According to Peirce the main flaw of Darwinian and other philosophical
systems has been to either reduce this third regulatory element to one of the other two, or to miss
it altogether (Peirce, 1958, 6.303). It is surprising to see how many contemporary debates within
evolutionary sciences have a chance to be settled if this third Peircean agent is incorporated within
the evolutionary paradigm. The scope of this research work does not allow to venture into the
resolution of those debates. However we shall use the Peircean framework to show how it fares
in the wake of some of the new developments within the cutting edge evolutionary biology, in
general. In particular, we show how Peircean framework fits into the ongoing debate within the
evolutionary sciences regarding one of the most critical issue, that of an evolutionary explanation
for the empirical evidence of ’cooperation’.
10

1.4 Contributions
A brief overview of the main contributions of this work is listed below:
1. Bringing philosophy and (computer) science into a relationship
2. Realizing through experiments and theory the significance of seeing ‘EAs as function opti-
mizers’ rather than ‘EAs are function optimizers’
3. Highlighting the role of evolvability over precision for evolution, and the factors that con-
tribute towards it in an EA
4. Exploring the relationship between the efficiency of function optimization in an EA and the
interpretation of natural processes and theories of evolution
5. Significance of Charles Sanders Peirce and his work for the 21stcentury science
6. Presenting Peircean theory of evolution as a synthesis, and making it converse with contem-
porary developments withing evolutionary sciences
7. Bringing EC at par with its counterparts in other evolutionary sciences
8. Readying Peirce for EC by extracting a Peircean EA out of his philosophical volumes
9. Mapping Peircean EA’s characteristics with the future challenges listed by Schwefel and De-
Jong
10. Possible presentation of Peirce as a unifying framework within EC, that could help many
variant EAs
11. Testing Peircean EA on extensive benchmark mathematical functions
12. Comparing the results with Classical and advance EAs
13. Presenting a Peircean solution to a major problem in EC - stagnation
14. Demonstrating how Peirce can help resolve Darwinian controversies, especially ‘Cooperation’
11

15. Putting to test Peirce’s evolutionary hypotheses, and helping Peircean scholarship too, bring-
ing philosophy and science into a badly needed two-way relationship
16. Identification of further possibiliies to be explored by the EC community
1.5 Thesis Roadmap
The disseration roadmap is as follows: Chapter 2 shows how EAs solve optimization problems, but
more importantly, how they travel the road from Nature to theorization to algorithmic modeling.
The chapter also discusses the intellectual environment at the time of Darwin, and the number of
things that went in to making his theory. Finally we map various components of Darwin’s theory
to the respective elements of the classical EA (C-EA) model.
In chapter 3 the discussion is repeated, but for Peirce; mainly, Peircean conception of the
universe and Peircean evolutionary framework are discussed using evidence from Darwin, biology,
theoretical definitions, and quantum physics.
The discussion is finally brought home in chapter 4 where the ground work laid in the previous
chapter is used to build a general framework for EC, as well as a specific Peircean EA (P-EA).
Chapter 5 captures the experimental details of our work. An important segment is the compar-
ison of C-EA and P-EA over an extensive set of benchmark mathematical optimization functions.
The discussion then moves beyond the empirical grounds in chapter 6. The chapter investigates
the theoretical understanding of the effect of Peircean principles in an EA’s convergence. Some
interesting results are drawn using the classical schema theory of John Holland and the associated
disruption analysis.
Note: The three separate investigations (in response to Schwefel’s opening question) presented
in this introduction chapter are not given a serial treatment within this dissertation. They are rather
introduced intermittently as sub-discussions within different chapters, as well as in the appendices,
and also winded up in the conclusion - chapter 7.
12

Chapter 2
Darwinian Evolutionary Algorithms:
From Natural Reality to Problem
Solving
The objective of this chapter is to introduce the elements that go into the making of an evolutionary
algorithm. Notwithstanding the recent plethora of EA variants that hardly seem to even try and
engage with any evolutionary theory, classically speaking, EAs have had one design route: evolution
factually occurs in the natural reality, there are theories to explain evolutionary phenomena, and
that EAs are computational models of those theories that attempt to explain the processes or sub-
processes of natural evolution. This is a well-established reading and construction of an EA; and it
shortlists one of the defining objectives of the scientific domain known as Evolutionary Computation
- to understand the natural reality.
In revisiting the roots of EC, we hope to achieve an understanding of not just the history and
philosophy of EC, but also by instantiating the above-mentioned general definition through John
Holland’s classical model of GA, we intend to further understand the elements that go into the
making of the Darwinian paradigm - the grand theory upon which Holland based his adaptive com-
putational model. This chapter also aims to introduce Darwinian paradigm through the lens of its
historical development, reconstructing the theory’s truly evolutionary character and consequently
13

deconstructing any false finalistic images of it. We shall begin from the beginning then.
2.1 Darwin’s Stature and Contribution in Science
Charles Darwin is rightfully placed in Westminster Abbey near Newton’s last abode. Though
centuries apart in time, both men fundamentally set forth new foundations for science, one in biology
and the other in physics, respectively. In the wake of the success physics enjoyed after Newton,
what makes Darwin even greater is his resistance against not just creationism (the established
belief of his time that all the variety of species have always been a separate creation;) but also
against reductive and deterministic mechanism (the belief that perfect laws control the outcome
of every event perfectly, and that these universal laws could be perfectly captured by reducing
them to elements at much basic/fundamental level) - the established framework in the physics of
his era. In such a terrifying intellectual environment, for someone to almost single-handedly carve
out space for a theory that could not be sold as either naturally theistic or causally mechanistic,
was a feat which only a man like Darwin could have performed. At a time when most scientists
would have wanted to win legitimacy for their work by following the route of the physical sciences
(mathematical rigour + deterministic laws), Darwin fearlessly showed an independent scientific
legitimacy for biology and his ‘Origins of Species’ proved that the language of biology did not need
to be embedded with mathematical equations to present a valid scientific theory (Mayr, 2004).
Darwin managed to show that the living matter that naturalists (biologists) studied, did not or
could not lifelessly obey any set of universal laws, while at the same time showing that it was still
possible to study the regularities within the diversity of life without resorting to transcendental
theistic arguments ultimately. For this two-edged victory, Darwin will rightly stand out for his
contributions to science, perhaps always.
The above paragraph is not a mere lip-service! Then why is it that we want to explore non-
Darwinian evolutionary theories as a foundation for EA?1 To answer this question we will have
to show two tendencies - as if running in parallel - in Darwin’s thoughts and work. One is his
1Note that non-X does not necessarily mean anti-X, even if the force of culture pushes in favour of this interpre-tation!
14

forward looking creative genius tendency, and most people are already aware of it. The other
one is a backward tendency; where according to our reading, and in the hindsight of the scientific
development within biology, we show where Darwin slipped because of the remnants of the biases of
his intellectual era in his framework. The only legitimacy in choosing an alternative theory would
be, if that theory adopted the forward tendencies of Darwin’s theory, while making corrections in
the backward tendencies. Our claim is that Charles Peirce has that potential (substantiated in the
next chapter).
2.1.1 Darwin, the Forward-looking Genius
Darwin’s Origin of Species demolished the premise that God had immutably created all
the variety of species during the 6 days of His Creativity - and that it has been this way
ever since - as the 17thcentury churchmen had so carefully formulated. (Mayr, 2004)
There is no doubt that Darwin’s main address every now and then in his ‘Origin of Species’ is
towards the issue of independent creation of all varieties and species. He ends many of his illustrative
ideas by stating that independent creation hypothesis could not explain this or that particular
phenomenon. He resolves the issue through his theory of evolution by Natural Selection
preserving the fitter modified descendents w.r.t adaptation to their respective living
conditions; and this slow and continuous accumulation of fitter characteristics giving
rise to ever-divergent variations, that over time come to be classified as different
species.
There is little doubt left today that, in contrast to the belief in independent and fixed species of
creationism, evolution is the empirically proven scientific stance. We say ‘little’ because Intelligent
Design schools and Christian Creationists still exist today, staring coldly away from the ever-
accumulating evidence in support of evolution. We think their fate has been sealed by Darwin’s
theory.
The other, lesser explicit forward-looking tendency in Darwin’s work is his divergence from
the methods of reductive-mechanical physical sciences. This in fact earned his book the infamous
critique of “the law of the higgledy-piggledy” from John Herschel, the English matheatician and
15

astronomer Darwin really looked up to (Darwin, 1995, page 220). This thought has been developed
further by Ernst Mayr, considered by some as “the Darwin of the 20thcentury”, in his catalogue of
essays titled, “What Makes Biology Unique?” According to (Mayr, 2004), Darwin manages to do
this by being a true naturalist; furnishing a complete theory without feeling an impluse to detail a
single mathematical equation, and by being nuanced about almost every scenario, giving examples
of exceptions wherever possible, and by avoiding universal and absolutist language most of the
times. Here are a few examples to illustrate the point:
I am convinced that Natural Selection has been the most important, but not the exclu-
sive, means of modification.
This nuanced approach of Darwin has been ignored by the neo-Darwinists of today. Another
aspect that got lost in the transition to neo-Darwinism is the fact that even though Darwin did not
agree with Lamarck’s account in its entirety, yet he did not shy away from using the Lamarckian
language:
Changed habits produce an inherited effect, as in the period of the flowering of plants
when transported from one climate to another. With animals the increased use or disuse
of parts has had a more marked influence;
Before we get to the where and why of the departures of the proponents of the Modern Synthesis
from Darwin’s original theory, we would like to complete the section by discussing now the seemingly
backward looking intellectual aspects in Darwin.
2.1.2 Darwin, the Backward-looking Intellect
Notwithstanding all the true geniuses of the man, Darwin also belonged to the previous scientific
era, where certainty and determinism were the scientific ideals to be achieved. For instance, Darwin
often spoke of chance in a mechanistic sense:
I have hitherto sometimes spoken as if the variations so common and multiform in
organic beings under domestication, and in a lesser degree in those in the state of
16

nature - had been due to chance. This, of course, is a wholly incorrect expression, but
it serves to acknowledge plainly our ignorance of the cause of each particular variation.
(Darwin, 1859, page 131)
T. H. Huxley, who became the mouth-piece for Darwin, while clearing the allegations of chance
off Darwin’s work had this to say:
It is not a little wonderful that such an accusation as this should be brought against
a writer who has, over and over again, warned his readers that when he uses the word
“spontaneous,” he merely means that he is ignorant of the cause of that which is so
termed; and whose whole theory crumbles to pieces if the uniformity and regularity of
natural causation for illimitable past ages is denied. But probably the best answer to
those who talk of Darwinism meaning the reign of “chance,” is to ask them what they
themselves understand by “chance”? Do they believe that anything in this universe
happens without reason or without a cause? Do they really conceive that any event has
no cause, and could not have been predicted by any one who had a sufficient insight into
the order of Nature? If they do, it is they who are the inheritors of antique superstition
and ignorance, and whose minds have never been illumined by a ray of scientific thought.
The one act of faith in the convert to science, is the confession of the universality of
order and of the absolute validity in all times and under all circumstances, of the law
of causation. This confession is an act of faith, because, by the nature of the case, the
truth of such propositions is not susceptible of proof. But such faith is not blind, but
reasonable; because it is invariably confirmed by experience, and constitutes the sole
trustworthy foundation for all action.
If one of these people, in whom the chance-worship of our remoter ancestors thus
strangely survives, should be within reach of the sea when a heavy gale is blowing,
let him betake himself to the shore and watch the scene. Let him note the infinite vari-
ety of form and size of the tossing waves out at sea; or of the curves of their foam-crested
breakers, as they dash against the rocks; let him listen to the roar and scream of the
shingle as it is cast up and torn down the beach; or look at the flakes of foam as they
17

drive hither and thither before the wind; or note the play of colours, which answers a
gleam of sunshine as it falls upon the myriad bubbles. Surely here, if anywhere, he will
say that chance is supreme, and bend the knee as one who has entered the very pene-
tralia of his divinity. But the man of science knows that here, as everywhere, perfect
order is manifested; that there is not a curve of the waves, not a note in the howling
chorus, not a rainbow-glint on a bubble, which is other than a necessary consequence of
the ascertained laws of nature; and that with a sufficient knowledge of the conditions,
competent physico-mathematical skill could account for, and indeed predict, every one
of these “chance” events. (Huxley, 2010)
Affirming and reasoning about the regularities in nature is one thing, but this pre-modern
tendency to ascertain perfection in knowledge and also to place this certainty in its foundations,
while leaving no room for real chance or fallibilism is what we term as a backward tendency in
Darwin and most scientific men of his intellectual era. This apologetic defence brings Darwin back
into the folds of the very physical cartesianism which he intended to naturally break from.
Previously we mentioned the nuanced characteristics of Darwin’s observations positively, and
he was remarkable especially in collecting the examples in nature that acted as exceptions to his
otherwise general observations. Darwin seemed to be comfortable with the fact that the biological
entities do not seem to need to conform to any universal code or law like the way dead matter
does. Strangely this nuanced element is left behind when it comes to independence between laws
of variation and selection. Gould has summarized this Darwinian “necessity” aptly:
Textbooks of evolution still often refer to variation as “random.” We all recognize this
designation as a misnomer, but continue to use the phrase by force of habit. Darwini-
ans have never argued for “random” mutation in the restricted and technical sense of
“equally likely in all directions,” as in tossing a die. But our sloppy use of “random”
does capture, at least in a vernacular sense, the essence of the important claim that we
do wish to convey namely, that variation must be unrelated to the direction of evolution-
ary change; or, more strongly, that nothing about the process of creating raw material
biases the pathway of subsequent change in adaptive directions. This fundamental
18

postulate gives Darwinism its “two step” character, the “chance” and “necessity” of
Monod’s famous formulation – the separation of a source of raw material (mutation,
recombination, etc.) from a force of change (natural selection). (Gould, 2002, page 144)
Darwin also displays quite an uncharacteristic single-mindedness when it comes to presenting
natural selection as the only creative agency in evolutionary nature, and competition among both
inter and intra-species being the only real mode of relationship across the living world. Sometimes
this clearly lends itself on to the racial and imperial English worldview of his time.
At some future period, not very distant as measured by centuries, the civilised races
of man will almost certainly exterminate and replace throughout the world the savage
races. (Darwin, 1871)
Nietzsche, the great German philosopher has picked in Darwin another English tendecy, and
put it in his trademark style saying that European scholars have not been able to completely
out-grow their social conditions, and that these remain a part of almost each scholar’s intellectual
idiosyncrasy:
The wish to preserve oneself is the symptom of a condition of distress, of a limita-
tion of the really fundamental instinct of life which aims at the expansion of power
and, wishing for that, frequently risks and even sacrifices self-preservation. It should
be considered symptomatic when some philosophers–for example, Spinoza who was
consumptive–considered the instinct of self-preservation decisive and had to see it that
way; for they were individuals in conditions of distress.
That our modern natural sciences have become so thoroughly entangled in this Spinozis-
tic dogma (most recently and worst of all, Darwinism with its incomprehensibly onesided
doctrine of the “struggle for existence”) is probably due to the origins of most natural
scientists: In this respect they belong to the “common people”; their ancestors were
poor and undistinguished people who knew the difficulties of survival only too well at
firsthand. The whole of English Darwinism breathes something like the musty air of
19

English overpopulation, like the smell of the distress and overcrowding of small people.
But a natural scientist should come out of his human nook; and in nature it is not con-
ditions of distress that are dominant but overflow and squandering, even to the point
of absurdity. The struggle for existence is only an exception, a temporary restriction of
the will to life. The great and small struggle always revolves around superiority, around
growth and expansion, around power – in accordance with the will to power which is
the will of life. (Nietzsche, 1974)
Nietzsche was no naturalist, but he had a sharp eye for dogmas of all forms and shapes. In light
of Lynn Margulis’ work on symbiosis in nature (Margulis and Sagan, 2002), Nietzsche’s criticism
of Darwin does seem to be on the mark; as shall be discussed in appendix A. The objective behind
listing these backward tendencies is to hope for a construction of a truly modern evolutionary
theory. If we are to claim that Peirce’s theory has that potential, this dissertation would have to
show how Peirce manages to avoid these Darwinian pitfalls, while retaining the positives.
2.2 The Road from Darwin to Post Modern Synthesis
The Darwinian explanation of the evolutionary process, as popularly understood, is based on two
main factors: 1) variation through blind chance providing the raw material and 2) mechanistic
laws, primarily natural selection, sorting through the raw material to select the fittest; shaping the
evolutionary course. Darwin assumes that these two characteristics, blind chance and mechanistic
laws, have existed in the universe from eternity and will continue to exist for all eternity without any
change or modification (without any evolution). Even if this philosophical oversight is corrected and
a non-evolutionary basis is added to the otherwise evolutionary framework of Darwin, a growing
body of empirical evidence suggests that there are several shortcomings in the Darwinian theory of
evolution.
Historically speaking, the simple fact that a neo-Darwinian modern synthesis had to be proposed
suggests that there are at least some oversights in classical Darwinism. But the journey from
Darwinism to this modern synthesis was not a simple matter of rediscovery and incorporation of
Mendel’s genetic principles into the classical Darwinian theory. It is worth noting that it was
20

the very geneticists who rediscovered Mendel who challenged one of the fundamental principles
of classical Darwinism. The analysis of these post-Mendel geneticists suggested that the laws of
Mendelian genetics and the newly discovered phenomenon of discrete mutations did not fit well with
the Darwinian notion of blending of inheritance. A direct consequence was that due to discrete units
of traits being inherited, the gradual evolutionary process did not seem possible. These saltationists
eclipsed Darwinism (Bowler, 1983, page 14) until the introduction of the idea of population genetics.
It was shown that the gene pool of a population rather than an individual’s genome follows smooth
bell-curved distributions, and hence Mendelian genetics was reconciled with Darwinism. Effectively,
the architects of the modern synthesis performed three tasks: 1) they preserved some of the key
ideas of Darwin, such as gradualism, infinitesimal random variation, and adaptation through natural
selection; 2) they introduced Mendelian mechanisms of genetic inheritance, along with ideas of
population genetics; and 3) they discredited all alternative ideas at the time including Darwin’s
flirtations with use and disuse of organs, Darwin’s hereditary mechanisms, Lamarckian ideas of
inheritance, and the ideas of mutationists/saltationists (Mayr, 1993). The fact is that the modern
synthesis became the established framework in evolutionary biology:
The major tenets of the evolutionary synthesis, then, were that populations contain
genetic variation that arises by random (i.e., not adaptively directed) mutation and
recombination; that populations evolve by changes in gene frequency brought about by
random genetic drift, gene flow, and especially natural selection; that most adaptive
genetic variants have individually slight phenotypic effects so that phenotypic changes
are gradual (although some alleles with discrete effects may be advantageous, as in
certain color polymorphisms); that diversification comes about by speciation, which
normally entails the gradual evolution of reproductive isolation among populations; and
that these processes, continued for sufficiently long, give rise to changes of such great
magnitude as to warrant the designation of higher taxonomic levels (genera, families,
and so forth). (Futuyma, 1986, page 12)
Since then almost all of these tenets of the modern synthesis have been challenged as being either
inaccurate or incomplete. They include gradualism, externalism, gene centrism, and selfish com-
21

petition. For instance, the saltationists’ scholarship was revived by Gould’s theory of punctuated
equilibrium. This presents an open challenge to the gradualism of Darwin (Gould 1984). Evidence
for non-random mutations has been accumulating, as well (Shapiro, 2005). The revisionist muta-
tionists now claim that the Darwinian way of describing the introduction of novel variants merely as
“random” is inadequate. Clearly, the generation of variation by mutation-and-altered-development
is spontaneous in many ways, but it also exhibits habits or tendencies. These tendencies impose
biases on the outcome of evolution. The failure to recognize that such “internal” tendencies have
an impact on evolution is a key failure of Darwinism according to neo-mutationist scholarship
(Yampolsky and Stoltzfus, 2001). Similarly, molecular biology and epigenetics have studied cellu-
lar functioning in far more depth than was possible at the time the modern synthesis was originally
proposed. It is now confirmed that inheritance takes place through extra-genetic mechanisms as
well as through genetic processes (Jablonka and Lamb, 2008). At the very least, the emphasis in
genotype-phenotype mapping has shifted from individual genes to networks of genes (Brem and
Kruglyak, 2005; Ehrenreich et al., 2010; Pigliucci and Muller, 2010). Cellular environment, tran-
scription and regulation mechanisms are being given as much significance as was previously given
solely to genes and their associated proteins. Systems biologists, who work on dynamic multi-level
selection systems, claim that reduction of such a system to any one level, such as genes, is bound
to give an incomplete picture of reality (Noble, 2006). On another front, Lynn Margulis’s work on
symbiogenesis proves that processes other than natural selection (for instance, symbiosis) have also
acted as creative evolutionary agents, especially for the evolution of eukaryotes from prokaryotes
(Margulis, 1981; Margulis and Sagan, 2002). This symbiosis, being a rather long-term cooperative
process, provides quite some challenge to the ‘selfish’ understanding of evolution. The horizon-
tal gene transfer between organisms that ranges from the entire genome (through hybridization,
symbiosis and parasitism) to partial exchange has resulted in another significant challenge to the
strictly vertical classification in the Darwinian ‘Tree of Life’ (Boto 2010). New metaphors such
as ‘Web of Life’ are being understood and developed by biologists in light of the accumulating
evidence for lateral gene transfers (Goldenfeld and Woese, 2007). In response to these numerous
challenges to the fundamental tenets of the modern synthesis, on the one hand there are calls for
22

extensions in the neo-Darwinian modern synthesis (Jablonka and Lamb, 2005; Pigliucci and Muller,
2010), and on the other there is a growing community of evolutionists who are working towards
non-Darwinian explanations of evolution especially in light of the new biological findings (Rose and
Oakley, 2007; Shapiro, 1997). We represent the latter group!
2.3 From Evolutionary Theory to Evolutionary Computation
Darwin’s theory has taken its fair share of blows but more importantly it has withstood the test
of time. One of the reasons is that it is so easy to summarize its thesis. Everyone has their own
version; here is our rephrasing of Darwin’s argument:
Evolution by Natural Selection preserves the fitter modified descendents
w.r.t. adaptation to their respective living conditions; and this slow and
continuous accumulation of fitter characteristics gives rise to ever-divergent
variations, that over time come to be classified as different species.
One of the key ideas in Darwin’s theory is adaptation. Credit must go to John Holland for taking
this Darwinian understanding of evolution through natural adaptation seriously and converting it
into an eligible design for artifical adaptive systems (Holland, 1975). Though Holland’s initial
algorithm had little or nothing to do with optimization, his students started applying what came
to be known as Genetic Algorithms (GA) or classical evolutionary algorithm (C-EA) to problems
of optimization (Goldberg, 1989). We shall show here the recipe that maps an evolutionary theory
on to a much simplified C-EA. The ingredients include:
• An individual encoding and representing a candidate solution to the problem
• A population of individuals
• An objective function that maps each individual to its fitness value
• A selection function that can introduce bias in the population towards the survival of the
fitter individuals
23

• A binary function to introduce variation through crossing ‘selected’ individuals to (re)produce
descendents
• A unary function to introduce mutation based variations to descendents
• A stopping criteria
2.3.1 Classical Evolutionary Algorithms
To be sure, living beings evolve and grow, but evolution and growth does not necessarily solve any
problems, and even if it could be said that natural evolution solves a problem, it does so only across
the span of millions of years. These factors are in contrast with the “fast buck,” efficient engineering
mindset of our “information age.” On the other hand, evolution has sustainably designed and
produced a tremendous variety of species capable of living in and adapting to extreme environments.
If engineering algorithms can take some cues from the natural processes of evolution, perhaps many
non-trivial, computationally hard problems can be heuristically solved in a reasonable time, helped
by the ever-increasing computational capacity and speed of computer processors. So the basic idea
is to mimic natural evolution in how it has evolved complicated and better-fitted organisms from
their very rudimentary origins.
In the evolutionary algorithms of computer science, instead of going to the very origins, a basic
encoding representation of the problem to be solved is assumed to be in place, and the population
of solutions is initialized randomly (within the bounds of the problem domain). Furthermore, there
must be a function which serves as the meter for fitness. Now some of the randomly initialized
individuals will have a higher fitness value against their encoded solutions than others. The as-
sumption in classical evolutionary algorithms, inspired by Darwin’s theory, is that if we have a
selection function which can bias the next generation of individuals in the direction of the fitter
individuals in the population, then the fitter individuals will have a greater chance to participate in
generating the next batch of individuals that make up the population; and over many generations,
the population will gradually improve in its average fitness (because of fitter individuals crossing
over) and move closer to the global optima. The selection function serves as an exploitation fac-
tor, while crossover and mutation functions serve as exploration factors within the solution/search
24

space. The continual exploitation and exploration by the evolving candidate solutions heuristically
converges onto the global optima, as shown in Figure 2.1.
Figure 2.1: Flow chart for Classical Evolutionary Algorithms
2.3.2 Classic Algorithms in EC are Foundationally Darwinian
(Beyer et al., 2002) have summarized the origin and basic working principles of Evolutionary
Algorithms with brialliant brevity:
Evolutionary algorithms (EA) form a class of probabilistic optimization methods that
are inspired by some presumed principles of organic evolution. .... The general frame of
EP, GA, and ES is essentially the same and very simply summarized by a loop over par-
tially randomized variation and selection operators steering exploration and exploitation
(or chance and necessity) and, in contrast to traditional optimization procedures, acting
upon a set of search points in the decision variable space.
The above quote also highlights the dyadic formulation of EA models i.e. chance and necessity.
We have already mentioned in the introduction that Darwin’s conception of evolution primarily
revolves around two fundamentals: blind chance and mechanistic or necessitarian laws. This fact
also gets translated into its EA models. In the later chapters we will see how Peircean concep-
tion of evolution is triadic and how its translated model (Peircean-EA) also acquires this third
characteristic.
A review of literature in the field lays bare the fact that almost all the classical evolutionary
algorithms (including genetic algorithms, evolutionary programming and evolution strategies) have
25

their roots in the Darwinian theory of evolution. One does not even need to survey the literature
to establish this fact; some of the leading experts in EC explicitly acknowledge it themselves:
The GA works on the Darwinian principle of natural selection....Whether the specifica-
tions be nonlinear, constrained, discrete, multimodal, or even GA is entirely equal to
the challenge. (Reeves, 2002)
A leading textbook in the field of AI writes:
Genetic Algorithms begin with a population of candidate problem solutions. Candidate
solutions are evaluated according to their ability to solve problem solutions: only the
fittest survive and combine with each other to produce the next generation of possible
solutions. Thus, increasingly powerful solutions emerge as in a Darwinian universe.
It is oddly fitting that these approaches should seek the origins of intelligence in the
same processes that, it may be argued, gave rise to life itself.... Certainly one of the
strongest models of learning we have may be seen in the human and animal systems that
have evolved towards equilibration with the world. This approach to learning through
adaptation is reflected in genetic algorithms, genetic programming, and artificial life
research. (Luger, 2002)
We believe that there is a high level of correlation between the three techniques, as they are
based on the same principles and have the same underlying architecture. Therefore, the evidence
we found for GA being Darwinian should in principle be enough for generalizing it to the broader
spectrum of EA:
The majority of current implementations of evolutionary algorithms descend from three
strongly related but independently developed approaches: genetic algorithms, evolu-
tionary programming, and evolution strategies. (Back et al., 1997b)
Having shown the evidence that the main EA in EC are Darwinian in their foundations, we
now turn to the last leg of this chapter. The more initiated readers of EC can of course skip the
following section.
26

2.4 Instantiation of an EA with an Optimization Problem
In Digital Image Processing, image registration is the process of “match[ing] two or more pictures
taken, for example, at different times, from different sensors, or from different viewpoints” (Brown,
1992, page 325).In this process a smaller template is to be searched within a larger image. Usually
the scaling and rotation and other transformations of the template are involved in the search, but
in order to simplify this illustration for readers outside the computer science community, we limit
the search space to two dimensions and do not add those transformations.
(a) Search Image (b) Template Image (c) Template Matching
Figure 2.2: Image registration: A 2-D problem
Figure 2.2(a) shows the search image, Figure 2.2(b) shows a scaled up template image, while
in Figure 2.2(c) the white box shows the smaller template image matching the larger search image
at a specific coordinate. Hence, it is a problem of searching for that x and y (or row and column)
coordinates where the template makes a maximum correlation with the image at that location.
Correlation coefficient ranging from -1 to +1 becomes the fitness value. This is how a searching
problem has been translated into an optimization language in this case. We discuss the ingrediants
of solving this optimization problem through an EA now:
Representation
For any problem to be solved through an EA, we need to encode it in a specific representation. The
template matching, without the scaling, rotation and other linear or non-linear transformations
can be represented as a 2D problem, as mentioned above. The complete candidate solution to
the image registration through template matching problem is just a 2D vector, where one axis
27

needs to cover the maximum number of rows of pixels that the search image has, and the second
axis covers the maximum number of columns. This (row #,col #) pair can completely represent
the solution to this specific problem, as the template image can then be placed at this specific
(row #,col #) coordinate and its solution can be validated through correlation. Had scaling or
rotation been involved, more fields would sure need to be added to the representation along with
the location vector. Usually EA use binary encoding to represent individuals and populations, as
this helps in crossover and mutation operators, as it shall become obvious shortly, but there are
other representations possible as well. In the case of binary representation, to cover the search
space of 1024 x 512 rows and columns a 19 bit long binary representation would be required - 10
bits to cover the 1024 rows of the search image, and further 9 bits to cover the 512 columns of the
search image, as shown in Figure 2.3.
Figure 2.3: Representation of EA Individuals for Image Registeration Problem
Population
Instead of a point search, EA are a population based searching algorithm, where the individuals
in the population interact with each other to get to the global optima in the search space. In the
case of image registeration the population of the EA would comprise of (row #,col #) pairs, how
many such pairs, or the size of the population is a variable and is one of the design parameters,
28

but is usually fixed. The only problem is that even if the size of the population is known to the
algorithm (let us say it is 100), what are the exact values of these 100 (row #,col #)? Classically,
EA populate these using uniform random distribution, in our case the first axis ranges from 1 to
1024, and the second ranges from 1 to 512.
Fitness Evaluation
Once the EA has the initial population, it needs to pass it on to the fitness evaluation function,
which in this case simply takes the template image, places it on the specific (row #,col #) of the
search image and calculates the correlation value between the two (in simpler terms, it calculates
how much the overlapping parts of the image match each other). This correlation value becomes
the fitness value against this individual. A high fitness value of an individual means that a high
percentage of the template image matches with the underlying search image when placed at the
specific (row #,col #) represented by this individual. The same process needs to be repeated for
the entire population.
Selection
Selection of the fittest is carried out based on the fitness values. The objective of the selection
function is to introduce bias within the population, meaning that in the next generation of the
randomly-initialized population, all individuals should not have an equal chance of representation.
Those individuals that turned out to be fitter should have a higher chance of participating in the
process of generating the next batch of individuals, and this selection is done for every generation.
There are various selection functions that the community has devised and keeps on devising. We
will illustrate only tournament selection here. Tournament size is again a design choice; for instance,
if the tournament size is 4 and the population size is 100, then the tournament selection function
will have to perform 100 tournaments in each of which 4 random individuals are picked from the
population, and the fittest among them wins that tournament. Appropriate tournament size is a
tradeoff. If the tournament size is too large, there is a chance that only a few fittest individuals
would survive until the next generation, and if the tournament size is too small, then too small a
29

selection pressure might result in a random search, and this selection function might not fulfill its
objective of exploiting or introducing the needed bias for the search. Once the 100 individuals have
been selected, they are to generate the next generation by using variation operators. The two most
classical ones are crossover and mutation.
Crossover
Crossover is a binary operator that takes two parents and produces two children by crossing them
at randomly chosen crossover points. Here is where the binary representation comes in handy.
The process is extensilvely explained in Figure 2.4. It shows the power of crossover operator as an
exploratory tool, as the least significant bits of two parents (P1 & P2) across the crossover point
get swapped. This crossover produces two children (C1 & C2) or two new search points that are a
variation of parent search points. This crossover operator is applied to all parents with a certain
probability which is termed “crossover probability.”
Figure 2.4: Crossover Operator’s Procedure - Parents 1 and 2 produce Children 1 and 2
Mutation
Mutation is a unary operator that is applied to a single individual with a very small mutation
probability. All it does is to flip a bit from 0 to 1, or 1 to 0, and hence produces a variation in the
population. Together with crossover, it gives exploratory powers to an EA.
30

Stopping Criteria
Once the population of the EA is initialized, selection, fitness and variation operators keep on
acting to produce generation after generation, unless one of the following usual stopping criterion
is met:
• Desirable fitness (correlation value >= 0.95) is achieved by an individual in the population
• Maximum number of generations have been exhausted (usually 10,000)
• No significant improvement in fitness has occurred since past 500 or so generations
2.5 Non-Classical Variant EAs
We have chosen classical EA to open up our case - the objective being the understanding of Peircean
EA in contrast. There are otherwise ‘countably infinite’ varities of EA it seems (pun intended,)
and making an exhaustive survey of advance varieties would almost be a meaningless activity for
this dissertation2.
2.5.1 Non-Darwinian-type Ideas
(Deb et al., 2002) propose a non-dominated sorting genetic algorithm for multi-objective functions
that uses a specialized selection operator incorporating a mathematical cuboid function to preserve
the spread of population. The use of such mathematical cuboids to achieve selection is hard to
argue for, both in nature and in the Darwinian accounts. (Runarsson and Yao, 2000) propose a
stochastic ranking system for selection using a bubble sort like procedure. Although this is a basic
algorithm in computer science, it is nowhere to be seen in the natural or Darwinian world. (Salomon,
1997) proposed a deterministic genetic algorithm saying that it is due to random application of
variation operator that genetic algorithms perform non-optimally. Such deterministic applications
of mutations are nowhere to be seen in natural as well as Darwinian world. (Herrera and Lozano,
2000) use triangular probability distributions for fuzzy recombination, and a hypercube topology
2We have ensured that there is no related work when it comes to Peirce and an evolutionary algorithm directly.
31

for exploration and exploitation. It is hard to find any example in the natural and Darwinian world
for both these phenomena.
These were some of the variants of classical EA, examples of which are available neither in
natural nor Darwinian world. Of greater relevance to our central claim are those variants of EA,
examples of which can be found in natural accounts, but provisions for such deviations are not to
be found in a Darwinian model. Their motivation has either been some social phenomenon or, as
is with most cases, merely the empirical improvement in the accuracy or efficiency from function
optimization point of view (Mantere, 2006; Beyer, 1996; Fogel and Chellapilla, 1998; Ray and Liew,
2003; Amor and Rettinger, 2005).
2.5.2 Peirce-type Ideas
Without the pretence of a survey, here is list of a few EA variants, which carry Peirce-type ideas
within their construction. We will briefly describe them here; their details can be had through the
references. We compare and contrast them with Peircean EA at the end of chapter 4 - where the
comparison makes more sense.
IDEA 1 :: Diversity Retaining Measures
Loss of diversity within the population means, for all practical purposes, the ‘death of evolution.’
Since crossover will not produce a new variety, and mutation rates are kept very low, there cannot
be further possible exploration of the search space, once diversity has been lost. This often means
a premature convergence to a local minima for an EA. To help avoid this, different operators have
been developed which help retain diversity, or in other words, help prevent stagnation due to loss
of diversity.
Mostly this loss of diversity is triggered because of the presence of a relatively very fit individual
in the population, and if the selection function is not balanced enough, in a few generations, this
one individual replicates itself, replacing the entire population. Keeping population in niches, and
the fitter individuals in every niche sharing their fitness with the rest of the sub populations is one
way to suppress the triggering phenomenon (Sareni and Krahenbuhl, 1998) in the selection phase.
32

In some EA especially those in which the population size is to remain fixed, it has to be determined
which individuals out of the new and old lot will remain and which will be replaced. During
this replacement phase, one technique that helps avoid stagnation is that the child which is most
similar to its parent, and with a higher fitness replaces the parent (Mahfoud, 1995). Other hybrid
formulations also have been proposed in which the individuals who explicitly do not contribute
towards increasing or maintenance of diversity are replaced (Lozano et al., 2008).
IDEA 2 :: Distributed Population Dynamics
The class of EA that evolves the population distibutively and comes closer to P-EA is Island Model
EA (IMEA) and Diffusion Model EA (DMEA), also called Cellular models. This section briefly
outlines their salient features.
Island Model EA
1. Based on Eldridge and Gould’s punctuated equilibria theory (Gould, 2002).
2. Mostly implemented in parallel processes/processor, with a certain communication topology
(Segura et al., 2011; Skolicki, 2005; Srinivasa et al., 2007).
3. Each subpopulation/deme is assigned a different processor, and usually the population is
evenly distributed.
4. Each subpopulation evolves independently.
5. Migration usually takes place after regular intervals (generations).
Diffusion Model EA
1. Based on population genetics idea of local (not global) selection and crossover.
2. Genetic makeup spreads from individual to population based on diffusion model (Back et al.,
1997a; Jaimes and Coello, 2009).
3. Designed for parallel processing with one individual per processor.
33

4. Selection of parents dependent on the underlying parallel architecture, usually SIMD (Single
Instruction, Multiple Data streams).
IDEA 3 = IDEA 1 + IDEA 2
Some EAs are a hybrid, having both the distributed/hierarchical population dynamics as well as
mechanisms of introducing fresh supply of individuals within the population - to avoid stagnation.
(Hu et al., 2003) combine two ideas for multi-objective EA: maintaining a hierarchical or-
ganization of repositories of individuals according to different fitness ranges, and the continual
introduction of new genetic material at the bottom level fitness repositories. They claim that this
is a sustainable exploratory model that avoids stagnation in the long-term.
(Amor and Rettinger, 2005) propose a GA and a self-organizing maps (SOM) as a hybrid
combination that tackles stagnation. Their GA-SOM maintains search histories keepting track
of the frequencies of neurons in the lattice of the SOM. This way the population is uniformaly
divided, as the lattice covers the search space uniformly. A reseeding operator ensures diversity
by introducing new individuals around those neurons (equivalently, search regions) which have had
the least frequent activations. (Kubota et al., 2005) have also designed a SOM based reproduction
operators.
Summary
This chapter has basically tried to achieve three objectives: a) to place Darwin’s theory in its
intellectual context, highlighting some of its achievements, and also listing some shortcomings that
were “naturally” ignored at that point in time in history due to the intellectual environment, but
which perhaps need to be revisited and repaired in light of contemporary scientific understanding,
if possible; b) to establish a relation between Darwin’s complex theory and John Holland’s adaptive
system as its simpler EA model; c) to instantiate the classical EA for an interesting digital image
processing optimization problem of image registration through template matching; and d) to enlist
some of the divergent non-classical EAs. This chapter also sets the tone for the next chapter, where
Charles Sanders Peirce’s evolutionary framework is explored for its potential contributions within
34

EC.
35

36

Chapter 3
Peircean Theory of Evolution
Peirce offers an evolutionary theory that embraces the advancements in classical theories while
avoiding some of their most glaring shortcomings. The notion of evolutionary growth is at the
heart of Peirce’s philosophy of pragmaticism. Building on the insights of (Spencer, 1867), Peirce
notes that “evolutionary growth” is not a mere increase; rather, it has certain characteristics that
must be accounted for in any theory of evolution. Evolution is a two-fold passage from homogenous
forms to heterogeneous on the one hand, and from unorganized to organized systems on the other.
The first passage leads to an increase in diversity, while the second movement leads to an increase in
regularity or uniformity. Given the fact of evolutionary growth (and its defining characteristics) the
question emerges: “What is the cause of this growth? Is it pure chance or is it mechanistic law?” An
adequate theory of evolution has to account not only for the fact of diversity and uniformity in the
universe but also for the fact that there is a process of “growth” in both phenomena. Peirce notes
that neither the randomness of pure chance nor the determinism of mechanistic law can account
for the simultaneous increase in diversity and uniformity (Peirce, 1958, 1.174)1. The following
observation by Peirce identifies the three elements that are at work in evolutionary growth:
[D]iversification is the vestige of chance spontaneity; and wherever diversity is increas-
ing, there chance must be operative. On the other hand, wherever uniformity is in-
creasing, habit must be operative. But wherever actions take place under an established
1Standard way to cite Peirce is (Volume number. Paragraph number). For example (1.174) means volume 1,paragraph 174. This format will be followed wherever Peirce is cited.
37

uniformity, there, so much feeling as there may be, takes the mode of a sense of reaction.
(Peirce, 1958, 6.267)
Summarily stated, we can say:
1) Diversity is the result of chance
2) Uniformity is the result of mechanistic reaction
3) Growth is the result of habit
For Peirce, biological evolution and its mechanisms did not suddenly come into existence in
the universe some four billion years ago with the emergence of the first forms of self-replicating
RNA. It is a process that is at least fourteen billion years old – or as old as the universe itself.
Biological evolution is only a specific manifestation of a more general phenomenon: evolution at the
cosmological level. A natural problem arises regarding the terms that should be used to describe
the basic elements for such a universal evolutionary theory. If biological terms are used, they
are instantly rendered useless at the level of physics, chemistry and cosmology, and vice-versa.
Hence, Peirce uses the technical terms “Firstness,” “Secondness,” and “Thirdness” (which he calls
“categories”) to describe chance, reaction, and habit respectively. Peirce’s use of the triadic scheme
illustrates the fact that his theory of evolution is entangled with other aspect of his philosophy, such
as phenomenology, metaphysics, logic, semiotics, etc. In non-technical language he has described
different manifestations of the three categories in these terms:
Firstness: chance, spontaneity, feeling, possibility
Secondness: reaction, mechanical necessity, force, competition
Thirdness: habit, generalizing tendency, law, cooperation
For Peirce, wherever there is evolutionary growth in the universe, there is dynamic interaction
between chance, necessity, and habit. Evolutionary growth, in the most general sense, is a movement
from pure chance or spontaneity (Firstness) to necessity (Secondness) by the gradual growth of habit
(Thirdness). The following quote summarizes Peirce’s hypothesis of evolution at the cosmological
38

level:
This theory is that the evolution of the world is hyperbolic, that is, proceeds from one
state of things in the infinite past, to a different state of things in the infinite future.
The state of things in the infinite past is chaos, tohu bohu, the nothingness of which
consists in the total absence of regularity. The state of things in the infinite future is
death, the nothingness of which consists in the complete triumph of [mechanistic] law
and absence of all spontaneity. Between these, we have on our side a state of things
in which there is some absolute spontaneity counter to all law, and some degree of
conformity to law, which is constantly on the increase owing to the growth of habit.
The tendency to form habits or tendency to generalize is something which grows by its
own action, by the habit of taking habits itself growing. Its first germs arose from pure
chance. (Peirce, 1958, 8.137)
In short, the triadic relationship between Firstness, Secondness, and Thirdness (the relationship
between chance, necessity, and habit) is the basic principle underpinning evolutionary growth. By
making biological evolution a subset of his theory of cosmological evolution, not only can the
question of biological origins be stated in more precise terms, but also the principles underpinning
Peirce’s theory of evolution can become open to scientific scrutiny, which is not the case in the
classical theories.
[T]he problem of how genuine triadic relationships first arose in the world is a better,
because more definite, formulation of the problem of how life first came about; and no
explanation has ever been offered except that of pure chance, which we must suspect
to be no explanation. (Peirce, 1958, 6.322)
Perhaps owing to Darwin’s efforts, Peirce did not need to waste his energies in conversing with
the creationists’ arguments. While Darwin’s main issue was to deal with the question of creation
of fixed species, Peirce’s problem is readily and visibly different. He is intellectually engaged in
conversation with the philosophical school of nominalism on the one hand (who advocates that there
are no universals or generals in the working of our universe, only specifics, implying that laws are a
39

mere fiction) and on the other, mechanical philosophers (who advocate such a narrowing conception
of cause and effect, that these mechanistic laws render spontenaity, freedom and chance a mere
lifeless illusion). Peirce proposed pragmaticism as a simultaneous response to both nominalist and
mechanical philosophers, for according to Peirce, the natural processes of evolution as well as the
very human processes of scientific inquiry are not possible while remaining faithful to any of those
two philosophical frameworks. But without making this disseration too absorbed in philosophy, our
objective is to highlight the fact that Darwin had a different set of issues and questions to address
than Peirce. Academic experience suggests that when the questions are different, the answers
ought to be different as well. The intelligibility of answers, at times, depends on the ingenuity of
the questions posed or the issues pursued.
3.1 Understanding Peirce’s Framework through Evidence
Many of the intricate philosophical issues facing Peirce were settled by showing how his category of
Thirdness was actively operative in the natural world, and how because of its realness the presumed
definitions of both chance and laws would have to be evolved by the scientific community. This
chapter sets the following three goals for itself 1) define Peirce’s Firstness, Secondness and Third-
ness, 2) show how they are operative in the biological world, and 3) show how the movement in the
modern physics is supportive of Peircean ideas rather than the doctrines of 19thand 20thcenturies.
3.1.1 Evidence for Peirce’s Thirdness in the biological world
We now step into Darwin’s Origin of Species to highlight how the tendency to take habits (Third-
ness) plays a significant role in the biological evolution at times. Please bear with us for the long
quotes, but understanding Darwin’s findings is important for understanding Peirce’s Thirdness:
Exhibit A: From Origin of Species, Chapter 6
I will now give two or three instances both of diversified and of changed habits in the
individuals of the same species. In either case it would be easy for natural selection
to adapt the structure of the animal to its changed habits, or exclusively to one of
40

its several habits. It is, however, difficult to decide, and immaterial for us, whether
habits generally change first and structure afterwards; or whether slight modifications of
structure lead to changed habits; both probably often occurring almost simultaneously.
Of cases of changed habits it will suffice merely to allude to that of the many British
insects which now feed on exotic plants, or exclusively on artificial substances. Of
diversified habits innumerable instances could be given: I have often watched a tyrant
flycatcher (Saurophagus sulphuratus) in South America, hovering over one spot and
then proceeding to another, like a kestrel, and at other times standing stationary on
the margin of water, and then dashing into it like a kingfisher at a fish. In our own
country the larger titmouse (Parus major) may be seen climbing branches, almost like
a creeper; it sometimes, like a shrike, kills small birds by blows on the head; and I have
many times seen and heard it hammering the seeds of the yew on a branch, and thus
breaking them like a nuthatch. In North America the black bear was seen by Hearne
swimming for hours with widely open mouth, thus catching, almost like a whale, insects
in the water.
As we sometimes see individuals following habits different from those proper to their
species and to the other species of the same genus, we might expect that such individuals
would occasionally give rise to new species, having anomalous habits, and with their
structure either slightly or considerably modified from that of their type. And such
instances occur in nature. Can a more striking instance of adaptation be given than
that of a woodpecker for climbing trees and seizing insects in the chinks of the bark?
Yet in North America there are woodpeckers which feed largely on fruit, and others
with elongated wings which chase insects on the wing. On the plains of La Plata, where
hardly a tree grows, there is a woodpecker (Colaptes campestris) which has two toes
before and two behind, a long pointed tongue, pointed tail-feathers, sufficiently stiff
to support the bird in a vertical position on a post, but not so stiff as in the typical
woodpeckers, and a straight strong beak. The beak, however, is not so straight or so
strong as in the typical woodpeckers, but it is strong enough to bore into wood. Hence
41

this Colaptes in all the essential parts of its structure is a woodpecker. Even in such
trifling characters as the colouring, the harsh tone of the voice, and undulatory flight,
its close blood-relationship to our common woodpecker is plainly declared; yet, as I can
assert, not only from my own observations, but from those of the accurate Azara, in
certain large districts it does not climb trees, and it makes its nest in holes in banks! In
certain other districts, however, this same woodpecker, as Mr. Hudson states, frequents
trees, and bores holes in the trunk for its nest. I may mention as another illustration
of the varied habits of this genus, that a Mexican Colaptes has been described by De
Saussure as boring holes into hard wood in order to lay up a store of acorns.
Petrels are the most aerial and oceanic of birds, but in the quiet sounds of Tierra del
Fuego, the Puffinuria berardi, in its general habits, in its astonishing power of diving,
in its manner of swimming and of flying when made to take flight, would be mistaken
by any one for an auk or a grebe; nevertheless it is essentially a petrel, but with many
parts of its organisation profoundly modified in relation to its new habits of life; whereas
the woodpecker of La Plata has had its structure only slightly modified. In the case
of the water-ouzel, the acutest observer by examining its dead body would never have
suspected its sub-aquatic habits; yet this bird, which is allied to the thrush family,
subsists by diving – using its wings under water, and grasping stones with its feet. All
the members of the great order of Hymenopterous insects are terrestrial, excepting the
genus Proctotrupes, which Sir John Lubbock has discovered to be aquatic in its habits;
it often enters the water and dives about by the use not of its legs but of its wings, and
remains as long as four hours beneath the surface; yet it exhibits no modification in
structure in accordance with its abnormal habits.
He who believes that each being has been created as we now see it2, must occasionally
have felt surprise when he has met with an animal having habits and structure not
in agreement. What can be plainer than that the webbed feet of ducks and geese are
formed for swimming? Yet there are upland geese with webbed feet which rarely go
2Another incidence where Darwin’s main addressee being Creationists is highlighted
42

near the water; and no one except Audubon has seen the frigate-bird, which has all its
four toes webbed, alight on the surface of the ocean. On the other hand, grebes and
coots are eminently aquatic, although their toes are only bordered by membrane. What
seems plainer than that the long toes, not furnished with membrane of the Grallatores
are formed for walking over swamps and floating plants? – the water-hen and landrail
are members of this order, yet the first is nearly as aquatic as the coot, and the second
nearly as terrestrial as the quail or partridge. In such cases, and many others could be
given, habits have changed without a corresponding change of structure. The webbed
feet of the upland goose may be said to have become almost rudimentary in function,
though not in structure. In the frigate-bird, the deeply scooped membrane between the
toes shows that structure has begun to change.
He who believes in separate and innumerable acts of creation may say3, that in these
cases it has pleased the Creator to cause a being of one type to take the place of one
belonging to another type; but this seems to me only re-stating the fact in dignified
language. He who believes in the struggle for existence and in the principle of natu-
ral selection, will acknowledge that every organic being is constantly endeavouring to
increase in numbers; and that if any one being varies ever so little, either in habits or
structure, and thus gains an advantage over some other inhabitant of the same country,
it will seize on the place of that inhabitant, however different that may be from its own
place. Hence it will cause him no surprise that there should be geese and frigate-birds
with webbed feet, living on the dry land and rarely alighting on the water; that there
should be long-toed corncrakes, living in meadows instead of in swamps; that there
should be woodpeckers where hardly a tree grows; that there should be diving thrushes
and diving Hymenoptera, and petrels with the habits of auks.
These few paragraphs of Darwin exhibit that sometimes habits of individuals/species change
first and the structural/bodily changes follow. In the time that the structural changes have not
taken place, and that must be a slower process than the individual’s choice of change of habit, this
3and again...
43

changed habit could not have been passed on to the next generation through the genetic-based
laws of inheritance, but there must be other social means of learning and teaching through which
these new and individual habits are to generalize and spread within the population of a specie. As
is evident, Darwin’s struggle to settle the creationists’ case makes him miss the gaps being created
between his own arguments. Only because he is busy speaking to a creationist, he misses the point
that over here natural selection takes a back seat; what is categorically more important here is
the willful spontaneous change of habit (Firstness) and a conscious sustaining/fixation of the habit
(Thirdness), even though the habits do not have the structural support as of yet; which becomes
secondary for that individual’s evolutionary growth at the moment and can come in later through
natural selection preserving those variations in structure that are in the direction of well-sustained
and taught habits of the species (Secondness).
So for Evolution to take place in reality, sometimes evolution by habit-taking becomes
more important than the ‘mechanisms’ of natural selection.
3.1.2 Evidence for Peirce’s Firstness in the biological world
Even though Darwin had initially insisted on the absolute independence of the processes of variation
and selection, in his later editions he conceded about the “variations which seem to us in our
ignorance to arise spontaneously. It appears that I formerly underrated the frequency and value of
these latter forms of variation, as leading to permanent modifications of structure independently
of natural selection” (Darwin, 1872, page 421).
This, in Darwin, opens up multiple possibilities:
a) that of a relation between the processes of variation and selection, as indicated by recent molecu-
lar biology findings where the rates of mutation as well as genomic rearrangement for an individual
get affected by stress under ecological challenges, often resulting in positively evolving the genome
(Shapiro, 2011)
b) redefining chance as a blind uniform distribution that does not get effected by anything outside
of it, to other distributions such as Gaussian/Normal. This would mean that chance has its own
44

habits as well, and that the outcomes of events involving randomness are not necessarily distributed
in a blind and habit-less way, all equally likely and uniformly. Fortunately we do not have to explain
this part as Probability is an established and well understood domain of mathematics now.
c) legitimizing the saltation sholarship, in which large genomic changes have historically taken
place under special circumstances in quick jumps, as opposed to a strictly neo-Darwinian slow and
continuous accumulation of one advantageous change at a time, naturally selected over long raging
times.
d) legitimizing neo-mutationist scholarship which ascribes the role of creativity to processes of
spontaneity as well, previously strictly associated with natural selection.
So for Evolution to take place in reality, sometimes evolution by spontaneity becomes
more important, sometimes evolution through natural selection, and sometimes evolu-
tion through habit-taking. If that is what the empirical evidence suggests, and that is
also what Peirce forwards as his evolutionary conception, then surely Peirce can help
take out the controversy in Darwin. Same is the case with evolution only through competition
in Darwin vs. evolution also through cooperation in Peirce (see Appendix A). Having seen Peirce’s
categories working in the biological world, let us now turn to their technical definitions.
3.2 Understanding Peirce’s Evolutionary Framework through Def-
initions
This section details how Peircean theory is precisely a synthesis of the above-mentioned three
different types of phenomena, in which the inter-relations of Firstness, Secondness and Thirdness
is what defines what evolution is, not a reduction to any one of the three - universally, and under
all conditions.
45

3.2.1 Firstness
Out of the womb of indeterminacy we must say that there would have come something,
by the principle of Firstness. (Peirce, 1958, 1.412)
The classical view of the world assumes that all specifications of the universe go back to the
beginning, where “chance” played its utmost part to put in place all the perfect values to all the
respective variables that shall ever need to operate. The Peircean framework gives an explanation
in which spontaneity plays a dynamic role not only in the initial event but also in continual growth.
By thus admitting pure spontaneity or life as a character of the universe, acting always
and everywhere though restrained within narrow bounds by law, producing infinitesimal
departures from law continually, and great ones with infinite infrequency, I account for
all the variety and diversity of the universe, in the only sense in which the really sui
generis and new can be said to be accounted for. (Peirce, 1958, 6.59)
3.2.2 Secondness
Where the idea of Firstness is characterized by independence from everything else, the idea of
Secondness is precisely that which is necessitated by some First. The units of Firstness did not
spring up in isolation, but “in accidental reaction upon one another, and thus into a kind of
existence.” (Peirce, 6.199) Peircean Secondness is encapsulated by functions such as reaction,
another, compulsion, negation.
Generally speaking genuine Secondness consists in one thing acting upon another, –
brute action. I say brute, because so far as the idea of any [regularity] or reason comes
in, Thirdness comes in. When a stone falls to the ground, the law of gravitation does
not act to make it fall. The law of gravitation is the judge upon the bench who may
pronounce the law till doomsday, but unless the strong arm of the law, the brutal sheriff,
gives effect to the law, it amounts to nothing. True, the judge can create a sheriff if
need be; but he must have one. The stone’s actually falling is purely the affair of the
stone and the earth at the time. This is a case of reaction. (Peirce, 1958, 8.330)
46

Although Darwinism argues that the process of evolution reflects the balance between chance
and laws, Peirce rejected the idea that such a balance offers any adequate explanation of the physical
universe and the natural world as we know it. He demonstrates that Thirdness is a prerequisite for
evolution and eventually for life.
3.2.3 Thirdness
The mode of being which consists in the fact that future facts of Secondness will take
on a determinate general character, I call a Thirdness. (Peirce, 1958, 1.26)
Where Second is always instantaneous, Third shall always have a process involved. As soon as
the idea of relation, pattern, logic, reason or law manifests itself in any phenomenon, it is due to
the presence of Thirdness. It is precisely because of this Third or tendency to generalize or take
habits that the above mentioned ideas are possible in reality.
[The tendency to take habits] is a generalizing tendency; it causes actions in the future
to follow some generalization of past actions; and this tendency is itself something
capable of similar generalizations; and thus, it is self-generative. We have therefore
only to suppose the smallest spoor of it in the past, and that germ would have been
bound to develop into a mighty and over-ruling principle, until it supersedes itself
by strengthening habits into absolute laws regulating the action of all things in every
respect in the indefinite future. (Peirce, 1958, 1.409)
According to Peircean conception then, three elements are active in the world:
first, chance; second, reactive mechanisms; and third, habit-taking. It is with the
interrelation of these three elements that Peirce explains the evolution of the world
from a state of nothingness to the one that we experience now.
47

3.3 Understanding Peirce’s Evolutionary Framework through a
Possible Synthesis
The following observation by Peirce shows the relationship between his triadic logic of three cate-
gories, non-biological evolution and biological evolution in the universe.
Three conceptions are perpetually turning up at every point in every theory of logic,
and in the most rounded systems they occur in connection with one another. They
are conceptions so very broad and consequently indefinite that they are hard to seize
and may be easily overlooked. I call them the conceptions of First, Second, Third....
In psychology Feeling is First, Sense of reaction Second, General conception Third,
or mediation. In biology, the idea of arbitrary sporting is First, heredity is Second,
the process whereby the accidental characters become fixed is Third. Chance is First,
[Selection] is Second, the tendency to take habits is Third. Mind is First, Matter is
Second, Evolution is Third. (Peirce, 1958, 6.32)
For Peirce, the most important philosophical implication of Thirdness is that there is meaning
and intelligibility in the universe (Peirce, 1958, 1.366). If evolution is restricted to the biological
domain then both meaning and intelligibility are severely undermined, if not completely obliterated.
As is the case with evolution at the non-biological levels, evolution at the biological level cannot be
understood or explained only with the aid of two categories. This means that Peirce is critiquing
both Darwin’s theory (which considers evolution to be the product of only chance and necessity)
and Lamarck’s theory (which considers evolution to be the product of necessity and habit). Peirce
recognizes that there have been three primary characteristics for the explanation of natural process
of evolutionary growth: Neo-Darwinism has primarily relied on chance (Firstness), Spencer on
mechanical principles (Secondness), and Lamarck on habitual use and disuse of organs (Thirdness).
According to Peirce they all suffer from one acute problem: by reducing the explanation of evolution
to one, two or any combination of two categories, the theory that thus forms cannot independently
and consistently explain the process of evolution4. Cosmological, geological and biological evidence
4This argument can benefit from a discussion on Peirce’s Reduction Thesis as well, but due to limitations of spaceit is left out as a future possibility.
48

supports Peirce’s claim.
Biological development cannot be separated from geological developments on earth. The geo-
logical record shows the same pattern or habit identified by Peirce for the cosmos: progressively in-
frequent catastrophic geological changes have affected the otherwise stabilizing biological life forms.
Citing (King, 1877), a leading geologist of his day, Peirce says: “the testimony of monuments and
of rocks is that species are unmodified or scarcely modified, under ordinary circumstances, but are
rapidly altered after cataclysms or rapid geological changes. Under novel circumstances, we often
see animals and plants sporting excessively in reproduction, and sometimes even undergoing trans-
formations during individual life, phenomena no doubt due partly to the enfeeblement of vitality
from the breaking up of habitual modes of life, partly to changed food, partly to direct specific
influence of the element in which the organism is immersed.”
If evolution has been brought about in this way, not only have its single steps not been
insensible, as both Darwinians and Lamarckians suppose, but they are furthermore nei-
ther haphazard on the one hand, nor yet determined by an inward striving on the other,
but on the contrary are effects of the changed environment, and have a positive general
tendency to adapt the organism to that environment, since variation will particularly
affect organs at once enfeebled and stimulated. This mode of evolution, by external
forces and the breaking up of habits, seems to be called for by some of the broadest and
most important facts of biology and paleontology; while it certainly has been the chief
factor in the historical evolution of institutions as in that of ideas; and cannot possibly
be refused a very prominent place in the process of evolution of the universe in general.
(Peirce, 1958, 6.17)
Peirce’s theory does not portray itself as a thesis for cataclysm or as an antithesis of Darwin
or Lamarck. His triadic theory only embraces the advancements in the abovementioned classical
theories, while avoiding some of their most glaring shortcomings. Not surprisingly, Peirce’s view of
biological evolution fits well with the fossil record as well as with the latest contentions of the evolu-
tionists mentioned in the introduction. For instance, Gould’s conception of punctuated equilibrium
which is at stark contradiction with Darwinian gradualism, sits well with Peircean cataclysm. The
49

Peircean provision, that under specific conditions the mutations have a tendency to adapt as op-
posed to being pure blind chance universally, is in line with the revisionist mutationists’ claims.
Instead of reducing the entire focus to genes, this theory suggests a multiplicity of mechanisms
from environment to metabolism, which is what the molecular findings of epigentics and systems
biology demand. Generalizing tendency and relationship, which are the manifestation of Peircean
Thirdness, are precisely those terms that Lynn Margulis is looking for in an evolutionary theory
that could support the evidence of symbiogenesis (see Appendix A). Philosophically speaking, what
takes the controversy out of evolution is the fact that for Peirce real chance has patterns and real
laws have exceptions.
3.3.1 Definition :: Peircean Evolutionary Theory
One might still be wondering as to what is the equivalent Peirce’s single sentence definition for the
theory of evolution. After all, we did give our version of Darwin’s theory in the last chapter. Here
is our encapsulated Peircean synthesis:
The processes of growth and selective adaptation to its environment are
characterized by the formation of new or the modification of old habits.
This relational definition first of all uses such general language that it avoids major controversies
especially in the wake of the modern biological developments (as listed in section 2.2). But more
importantly, this has room for both freedom and consciousness as exhibited in the higher life
forms, as well as the mechanisms exhibited by the lower life forms. Lastly, it brings into relation
spontaneity, adaption/selection and habits; or Firstness, Secondness and Thirdness in Peircean
terms.
3.4 Nature of Chance and Laws in Light of Modern Physics and
Cosmology
We now turn our gaze from biology to physics - the mother of natural sciences. The world Darwin
belonged to was still coordinated by Newtonian physics, and the assumption that the universe is a
50

perfect Cartesian machine was at work in most theories of 19thcentury, including that of Darwin’s.
We have already seen his conception of chance, how it was not an active agency, but only filled the
gaps of our ignorance for the time being. The other side of the same coin then is the assumption
that the laws are immutable and eternally perfect. This section has two objectives, a) to show
the transition in scientific understanding of these two categories, chance and laws, and b) to show
how Peirce had anticipated such a universe and how his categories of Firstness, Secondness and
Thirdness fit the description provided by the 21stcentury physics. Since we are not physicists
ourselves, we will bring in three quantum physics authorities to present our case.
3.4.1 Heisenberg on the Nature of Reality
The mechanistic world view has an operational assumption, that any event can be precisely worked
out by first breaking down our understanding to the most basic and elementary particles and then
putting it all together “accurately” in a bottom-up fashion. Heisenberg exclaims that the nature
of reality is not this simple any more:
What is an elementary particle?...If one wants to give an accurate description of the
elementary particle and here the emphasis is on the word ‘accurate’ the only thing which
can be written down as description is a probability function. But then we can see that
not even the quality of being... belongs to what is described. It is a possibility for being
or a tendency for being. (Heisenberg, 2007, pg. 44)
The probability function does unlike the common procedure in Newtonian mechanics
not describe a certain event but, at least during the process of observation, a whole
ensemble of possible events. (Heisenberg, 2007, pg. 28)
3.4.2 Weyl on the Historical understanding of Reality
Hermann Weyl, being a theoretical physicist as well as a historian of mathematics, shows in his
primer how the human understanding of laws of nature has moved from the search of determinism
to that of statistical regularities:
51

The idea of functional law, to which science seems to reduce causality, is not altogether
unproblematic. Twice in its history physics believed that it had overcome in principle
the decomposition of the world into individual systems (individual events and their
elements, which after all are only approximately isolated from one another) and had
grasped the world as “a whole in which all is interwoven.” The physics of central
forces and later the pure field physics seemed for a moment to have reached that goal.
Causal law here took the following form: the derivaties with respect to time of the
state quantities at a world point are mathematical functions of the state quantities
themselves and their spatial derivatives at that point. Consequently, the state of the
world at any moment would determine the state at the immediately following moment
by means of differential laws. Thus only the world’s state at a single moment would
remain ‘arbitrary’ or ‘accidental,’ and from it the world’s whole past and future could
be computed by integration of the ‘Laplacean world formula.’ (Weyl et al., 2009, pg.
190-191)
The classical definition of quantitative probability coined by Laplace – the quotient of
the number of favorable cases over the number of all possible cases – emphasizes the
objective aspect. Yet, this definition presupposes explicitly that the different cases are
equally possible. Thus it contains as an aprioristic basis a quantitative comparison of
possibilities. ...It is because of the arbitrariness of such a measure that Laplace, from
his consistently deterministic conception of nature, is in the end unable to ascribe to
probability anything but a subjective meaning; it deals with events whose premisses
are incompletely known, and thus is “relative to this our knowledge and ignorance.”
Laplace therefore calls two events equally possible if we are equally undecided as to
their occurence. (Weyl et al., 2009, pg 195)
In the fourth part of his Ars conjectandi, Jacob Bernoulli throws the bridge from the
subjective to the objective conception of probability by means of his “law of large
numbers.” According to this objective interpretation the probability calculus serves
to establish regularities expressed in the mean values of many similar events rather
52

than in the individual event. Beside the strictly valid causal laws we thus have
regularities of a statistical nature. (Weyl et al., 2009, pg 196)
As long as one believes in strict causality, statistics must find its proper foundation in
a reduction to strict law. If, however, there should be a ‘primary probability’ for the
individual atomic events that cannot be reduced to causal laws – and such seems to
be the case according to the most recent development of physics – then we seem to be
forced to introduce into the natural laws as an original factor either that probability
itself or some quantity connected with it. (Weyl et al., 2009, pg 198)
Most of the physical concepts, especially those concerning matter with its atomic struc-
ture (e.g. the density of gas), are not exact but statistical, that is, they represent mean
values affected with a certain degree of indeterminancy. Similarly most of the usual
physical ‘laws,’ especially those concerning matter, must not be construed as strictly
valid laws of nature but as statistical regularities. (Weyl et al., 2009, pg 199)
At any rate, in the actual conduct of physical research, statistics today plays at least as
important a part as the strict law. Attempts to reduce one to the other have gradually
fallen back behind the independent building up of a statistical thermodynamics. Of
the two laws which are of universal significance for all physical phenomena, the laws
of conservation of energy and the law of continuously increasing entropy, one is the
prototype of a strict law, the other of a statistical law. ...From all that has been said
it will be clear how little contemporary physics, based as it is half on laws and half on
statistics, can pose as a champion of determinism. (Weyl et al., 2009, pg 202-211)
This historical account puts an end to the world as known by Darwin. We would let Gerald
Holton conclude - a research professor of physics as well as a historian of science at Harvard.
3.4.3 Holton Concludes
Max Born was awarded the Nobel Prize in Physics in 1954, primarily for his statistical
interpretation of quantum mechanics. The fact that the award came so long after his
53

original publication (1926) indicates that this interpretation was not at once regarded
as an indubitable, basic “discovery,” but that it gradually came to be accepted as a
fruitful viewpoint in physical theory. Insofar as many scientists do accept the notion
that natural processes are fundamentally random rather than deterministic, we have to
admit that the “Newtonian world machine,” or the mechanistic viewpoint advocated
by Descartes and Boyle in the seventeenth century, has been abandoned. (Holton and
Brush, 2001)
Summary
What becomes blindingly obvious after reading the above sections is how the quantum physics of
21stcentury and Charles Sanders Peirce are almost on the same page, and how this new conception
of reality is different from that of Darwin’s. Mainly because chance has an element of law at work,
and laws have an element of chance at work:
a) Peirce embedded in his evolutionary framework such a notion of chance that is actually an active
agency rather than the old notions where chance only represented one’s ignorance,
b) For Peirce, laws of nature are not immutable, universal and unintelligible as imagined in the
past. Instead they are characteristically fallible, just like our interpretations about them.
c) But more importantly, Peirce managed to establish a relation between the creative and spon-
taneous chance and increasingly regular laws by placing a Third active agent between the two:
habit-taking tendency of generalization.
With this triadic evolutionary framework, Peirce brings natural laws into the umbrella of evo-
lutionary inquiry as well, and becomes readily relevant for the contemporary and future human
understanding and explanation of the universe (Smolin, 2013). This explanatory power over the
other evolutionary accounts, which are divorced from any extra-biological development, not only
gives Peirce’s theory a scientific superiority in principle, but also gives us a warrant to venture
into evolutionary algorithms based on Peirce’s theory of evolution. Lastly - and we hope Schwefel
agrees by now - Peirce’s theory goes a long way to meet the conditions that Schwefel foresees for a
universal theory of evolution:
54

Current evolutionary algorithms are certainly better models of organic evolution. Nev-
ertheless, they are still far from being isomorphic mappings of what happens in nature.
In order to perform better, an appropriate model of evolution would have to comprise
the full temporal and spatial development on the earth (a real global model) if not
within the whole universe. (Schwefel, 1997)
55

56

Chapter 4
Peircean Framework for Evolutionary
Algorithms
This chapter begins with a discussion on the principles derivable from Peirce’s theory of evolution
from EC’s point of view, and ends with a description of a detailed Peircean evolutionary algorithm.
The objective of this chapter is thus, to make the transition possible; the transition from a complex
and philosophical evolutionary theory to a much simpler, pseudo-codable algorithm - just like John
Holland once managed to do for GA.
4.1 Principles derived from Peirce’s theory for Evolutionary Al-
gorithms
In basic terms, the standard EA implementation is a two-pronged strategy that appears in EA
literature under different names: “exploration-exploitation,” “variation-selection,” and “chance-
necessity” (Beyer et al., 2002). If we could rewrite that in Peircean terminology, it would roughly
translate into “Firstness-Secondness.” The foremost difference is that the Peircean EA would have
a Third (generalizing) element working simultaneously, an element that we implement in the form
of clustering. A second difference is that for Peirce the meaning of “Firstness” is not confined
to variation, and the meaning of “Secondness” is not captured entirely by selection, either. But
57

thirdly and most importantly, the relationship and interplay between the three elements of evolution
build an entirely different system, and hence those elements get translated into an entirely different
evolutionary algorithm. This section elaborates upon the different meanings that can be derived
from the Peircean concepts from the standpoint of evolutionary computation. Next section explains
the new Peircean EA.
4.1.1 Firstness as Spontaneity
Unlike the classical EA, where random population is only generated once - at the beginning, in
Peircean EA, Firstness is an ever present phenomenon which means that there has to be a provision
for the random individuals to be generated every iteration. This could raise the possibility of
population explosion in case the number of iterations increases. We could easily work our way
around that, as shall be explained in the next section (in case keeping a constant population is an
objective).
4.1.2 Secondness as Necessity
Through Secondness, Peirce gives us many different operators that can be employed to change the
population or to generate the next generation of individuals under the brute forces of reactions.
In Peircean language the operators are: Compulsion, Dependence, Negation, Another. If we were
to categorize them, we would find that some function of crossover points could be employed to
develop the operator ‘Another’. The selection operator fits Compulsion but it could also be used for
affecting the population through the external, biased input. We will discuss that shortly. Different
functions of mutation could be used to actualize operators like Independence and Negation. In
short, the standard operators of recombination, mutation and selection can be extended to devise
the Secondness operators.
4.1.3 Thirdness as Generalizing tendency
The most striking feature of Thirdness is the generalizing tendency translating into collaboration
and cooperation among organisms. This new paradigm gives us two directions. First and foremost,
58

it strongly suggests that the algorithm has an inclination towards collaboration between individuals
i.e., after Firstness has taken place and new individuals have been randomly initialized, they interact
with each other using Secondness operators but brought into relation by clustering (Thirdness).
Clustering distributes the population into smaller sub-populations, each cluster representing the
generalization of its constituent individuals. Clustering represents Thirdness in that, in Peircean
EA, intra-cluster level operators like crossover and selection (Secondness) are taking place but
only on those individuals (Firstness) that are being related with each other through the long-term
bonds of each cluster. Thirdness is that which relates Firstness and Secondness, and clustering is
doing precisely that and cannot be reduced to Secondness. Secondly, this paradigm opens up new
possibilities for survival; instead of a singleton ‘survival of the fittest,’ the Peircean framework gives
us another epitome of survival: ‘collaboration.’ The more an individual is part of a collaboration,
the greater its chances of surviving. The criterion for clustering is not fitness-based; rather, it is
spatial, binding each cluster to its own contextual environment more naturally.
4.1.4 Dynamic System
Peirce acknowledges the individual organism as actively taking part in the evolutionary process.
Hence the system is not closed like the Darwinian system. This gives us four openings in the field
of Evolutionary Computation, and further establishes the explanatory power of Peircean framework:
1. Interactive Evolutionary Computations, where the fitness or objective function is scored by
a human operator, is now justified in this dynamic framework, which had to be incorporated
in classical EA by moving outside the Darwinian model - just to achieve results.
2. Unlike in classical EA where there is no role for external bias or prior information, Peircean
EA provides a semiotic framework where, external bias is acknowledged, accounted for, and
allowed. It could be initially taken as an input, which, through the operator of Compul-
sion acts as a guide for the development of future generations. In the absence of any prior
information about the problem at hand, the algorithm should work unaffected.
59

3. Schwefel’s meliorization becomes a possibility now due to individual’s Firstness. We do not
know how to implement it into an EA for now, but it is a possibility nevertheless for the future,
where the agent can have the will and consciousness to shape the present in accordance to a
future in mind.
4. Peirce recognizes evolution taking place at multiple levels of reality in parallel instead of
reducing the entire process to the genetic or individual level. In the Peircean framework
Firstness, Secondness and Thirdness work at all levels of natural (or algorithmic) reality and
in our case at Individual, Cluster and at Global level.
4.1.5 Reality at Multiple Levels
For Peirce, evolution by Firstness, Secondness and Thirdness takes place at multiple levels of reality
in parallel instead of solely at the genetic or individual level. Figure 4.1 allows us to see a snapshot
of the search space at one hypothetical generation along with the key agents/levels involved in
the Peircian development of an Evolutionary Algorithm. In the Peircian framework Firstness,
Secondness and Thirdness have to work at all levels of natural reality and in our case at Individual,
Cluster and Global level. For a complete understanding of this new paradigm we shall like to
establish the exact roles played by Firstness, Secondness, Thirdness on all levels of our algorithmic
reality, namely: Individual, Cluster and Global levels as seen in Figure 4.1. This would eventually
turn into a 3 x 3 matrix of relationships shown in Table 4.1. This exercise will help us to extract
principles from Peirce’s theory and to give a final shape to our EA in the next section.1
1But is this whole cluster scheme not analogous to Darwin’s group selectionism? The analogy fits only at asuperficial level. If we take the analogy at face value we cannot explain the emergence of clusters to begin with.Clusters are those phenomena in empirical reality where individuals cooperate with each other and pass on acquiredhabits to the next generation. Based on the accumulated (and accumulating) evidence in biology, selection is awoefully inadequate concept to explain the reality of cooperation, habit formation, etc., in nature. It is obviously thecase that selection does not disappear once clusters emerge, and selection is important in helping us to understandthe survival of clusters once they have emerged-but selection is a blinder that obscures some of the key dynamicsthat are at work during the origin/emergence of clusters. In a way, our main contribution is the working out of therelationship among Firstness, Secondness and Thirdness at individual, cluster and global levels of our algorithmicreality. It is out of this exercise that we then devise the algorithm. Now at this level the analogy of our proposedalgorithm with a group selectionist model does not hold.
60

Figure 4.1: Hypothetical 2D search space depicting a cluster formation within the population
Firstness at Individual level
Spontaneity, chance, and complete freedom are some of the ideas associated with Firstness. At the
individual level, the mutation operator introduces this spontaneity to bring about something novel.
Firstness at Cluster level
The cluster center is the individual which represents its respective cluster and is thus distinct
from other members of that cluster. A mutated cluster center would result in the introduction of
spontaneity at the cluster level.
Firstness at Global level
Each cluster has its own fittest individual that can also be called the cluster ideal. The set of these
fittest individuals from each cluster form the global representations. Mutated cluster ideals are
responsible for the introduction of spontaneity at the global level. Another mode of spontaneity at
the global level is that of cataclysmic changes, such as those that take place at the first generation,
61

when totally new elements are introduced in the search space, and these cataclysmic new elements
could also be introduced in later generations with a low probability.
Secondness at Individual level
Keywords that correspond to Secondness are “necessity” and “compulsion”. At the individual level
Secondness manifests itself in the form of genetic heredity - the individual is compelled to receive
its genetic blueprint from its parents and then to pass it on to the next generation. Crossover will
play the role of the Secondness operator and bring two individuals to react and produce offspring.
Secondness at Cluster level
At the cluster level various elements are involved at the same time to encapsulate Secondness. First,
the selection operator is responsible for retaining the fittest individuals or cluster ideals. In turn,
these cluster representatives play an active role in crossover for the next generation. Cluster-based
survival is also an indication of Secondness at cluster level.
Secondness at Global level
Just as intra-cluster interactions are the key aspect of Secondness at the cluster level, at the global
level inter-cluster dynamics play their part. Cluster representatives or the cluster ideals exchange
information across the clusters using the Secondness operators. This information exchange, between
geographically or functionally fittest individuals, becomes the cause for making the entire cluster
move across the search-landscape. It could result in either cluster unification or diversification in
the long run. Selection towards the global ideals is also a part of Secondness at the global level.
Thirdness at Individual level
Thirdness is best described by the keywords “representation,” “relationship,” “continuity,” “habit,”
“generalization,” “pattern,” “regularity,” etc. At the individual level Thirdness appears as the
representation of a chromosome or individual’s size, and all other parameters dictated by the
problem that the EA is going to solve or the function that is being optimized using the Peircean
62

EA.
Thirdness at Cluster level
Thirdness at the cluster level gives us another technique for a mating operator. The more the
strength or objective function’s value of an individual, the more is the effective radius of that
individual, and within that radius there is a higher probability of it crossing with other individuals.
However, this has not been tested in Peircean EA in this study.
Thirdness at Global level
At the global level Thirdness takes the form of generalization. The very idea of clustering stems
from this generalization. All the parameters for cluster radii or number of clusters at any time
are specified dynamically at this level. Other than that, all parameters that help optimize the
evolutionary algorithm are dynamically set by the Thirdness factor, for example, crossover or
mutation rates, prior information or bias, efficiency improvement measures, memorization of states,
etc.
Table 4.1: Relationship of Peircian principles at different levels of algorithmic realityIndividual Cluster Global
Firstness mutation mutated cluster center mutated cluster ideals
Secondness crossover cluster based survival,selection towards clus-ter ideals
inter-cluster interac-tion, selection towardsglobal ideals
Thirdness meaningful representa-tion, problem specificparameters
radius based mating op-erators
cluster formation, priorinput bias, optimizationparameters
4.2 Peircean Evolutionary Algorithm
The preceding section has laid down the general framework for designing evolutionary algorithms.
The Thirdness principle’s most important contribution is that the population shall cooperate and
survive in cluster-based communities. The Secondness principle dictates the terms under which
individuals and clusters interact with each other through various operators of recombination and
63

selection. The Firstness principle introduces and retains novelty in the population through various
operators of variation. Figure 4.2 lays out the basic algorithm in the form of a pseudo code.
Figure 4.2: Algorithm 1: Pseudo code for Peircean Evolutionary Algorithm
A variety of algorithms could be devised based on multiple interpretations of the Peircean
principles listed in the previous section. This multiplicity is not an issue as long as the interpretation
or the algorithm does not violate the spirit of Peirce’s evolutionary framework. While stressing
the need for a synthesis within the EC community, our claim is that this general framework has
the potential to act as a theoretical foundation for many variant EAs, especially co-evolutionary
and the like. We have only devised one instance of the otherwise general Peircean framework for
carrying out the experiments.
Algorithm 1 when stated simply is this: Until the stopping criteria are met, 1) distribute the
population in clusters. 2) First each cluster internally generates its next generation. 3) Next each
cluster’s fittest individuals make an inter-cluster information exchange. 4) Finally a small number
of new individuals also get introduced into the population. However, it shall be helpful if we take
a step by step tour of this P-EA:
Line 1: Initialization In case we do not have prior knowledge, the population is initialized in
the standard way, using uniform random distribution in the first generation. If we have any prior
bias available that we want to add to the first generation, we do that by either generating population
through a prior distribution (instead of using uniform distribution) or by simply inserting a few
individuals which encode the prior information along with other randomly generated individuals.
64

Line 2: Stopping criteria There are three stopping criteria for the algorithm: 1) minimum
accepted fitness value achieved by any individual in the population, 2) maximum number of gen-
erations exhausted, which is 10,000 generations, or 3) if the minimum value of the fitness of the
population does not improve for a certain number of generations beyond the first 1000 generations.
This certain number will be referred to as stop-count from now onwards.
Line 3: Parameter tuning While the other parameters that are listed in Table III remain
constant, we have experimented with two parameters: Cluster radius and mutation rate. Very
simple heuristics are employed. Mutation rate is logarithmically decreased from 0.2 to 0.02 in
about the first 1000 generations. There on, it becomes static at 0.02. In the case of cluster
radius, it starts from a constant number. In case the number of clusters become less than 2, the
cluster radius is decreased linearly relative to the previous generation, and in case the number
of clusters exceed 6, the cluster radius is relatively increased. While these are not precisely self
adaptive parameters, and the numbers for minimum and maximum bounds might be arbitrary, but
nevertheless Peircean EA has this potential that is to be exploited for future work.
Line 4: Cluster analysis We have used subtractive clustering which takes cluster radius
for input instead of specifying the number of clusters and returns the number of clusters and
cluster center representatives from the population (Chiu, 1994). It is a single pass algorithm but
notice how adding the procedure of clustering to the iterations of an EA does not add much to its
running time complexity as clustering is a polynomial time operation with respect to population
size. Since precise clustering is not the problem we want to solve, we could use any efficient linear
time algorithm available for that matter (Han et al., 2006). In any case it does not add to the
number of function calls to the objective function.
Line 6: Intra-cluster evolution This procedure is the heart of P-EA and its pseudo-code
is expressed as algorithm 2 in Figure 4.3 as well. Its function is to take one cluster at a time and
produce the next generation while identifying each cluster’s fittest individuals. One thing to note
here is that no special selection operator is involved. Parents are chosen using a uniform random
distribution which does introduce a very small selection pressure (as a handful of cluster individuals
get selected more than once), but other than that no special selection is done. Even though we
65

did mention the possibility of experimenting with various Secondness operators, we have restricted
to the standard multi-point recombination operator and standard mutation operator. This assures
that any improvements in the results over a classical EA are attributed to the Peircean framework
and not to any complex operators. Since there is always a chance of a few individuals getting
selected more than once in line 3 of algorithm 2, the following relationship holds:
Size of (parents + non-parents) ≥ Size of (Input cluster)
Size of (parents) == Size of (children)
Size of (children + non-parents) ≥ Size of (Input cluster)
Hence to keep the cluster population size constant, in line 9 of algorithm 2, the fittest among
children and non-parents are selected to represent the next generation of the cluster. Notice this is
not elitism, since parents are completely replaced with their children, and there is always a chance
of losing the previous fittest individual. The minus 1 at the end of the above-mentioned line is to
counter the effect of line 9 of algorithm 1. It keeps the total population size in place.
Figure 4.3: Algorithm 2: Pseudo-code for intra-cluster evolution
Line 8: Inter-cluster evolution Inter-cluster evolution is in many respects similar to Intra-
cluster evolution. The difference is that instead of a single cluster, representatives from each cluster
recombine and mutate to produce the next batch and the fittest are chosen in an elitist selection.
The pseudo-code is exhibited in Figure 4.4 as algorithm 3.
66

Figure 4.4: Algorithm 3: Pseudo-code for inter-cluster evolution
Line 9: Add random individuals to population Last but not the least, through Firstness,
in every generation new individuals come into being and become part of the current population.
They are of two types. Some of the individuals are generated absolutely randomly, the way the
first generation comes about. Some random individuals are mutated versions of cluster centers
and cluster fittest. We keep a uniform distribution between the two types. To avoid population
explosion we devised a mechanism in which (x * total clusters) new individuals are added to the
population in each generation, and likewise x weakest individuals farthest from their respective
cluster centers are eliminated. This is another definition of survival of the fittest: The more an
individual is a part of a cluster, the more its chances of survival.
4.3 Comparison of Peircean EA with Related EA
This section is the counterpart of section 2.5. While in chapter 2 some alternatives to classical
Darwinian EA were presented, here some of them are to be discussed in light of the Peircean EA.
One thing to note is that the P-EA is a very minimanlist algorithm. It is not advance, in the sense
of having outrageously “unnatural” components to it. This aspect of the P-EA contrasts with the
advance (non-Classical) EA variants discussed in section 2.5, and we leave the discussion at just
that, and get to the Peirce-related EAs.
67

4.3.1 IDEA 1 :: Diversity Retaining Measures
Most diversity maintaining techniques have come to the community as an afterthought - after
the problem of stagnation had been reported from various pockets of EC community. Hence, the
techniques, when embedded into the EAs seem forceful and artificial. P-EA on the other hand,
does not go for the stagnation-avoidance per se. It is because of the persistent presence of Firstness
as a real entity in a Peircean world-view, that raw genetic material gets continually introduced
within the population. In other words, stagnation gets tackled as a result of the natural translation
of Peirce’s theory into an algorithm, not because it is a special challenge within the EC community
that needs to be dealt with.
4.3.2 IDEA 2 :: Distributed Population Dynamics
This section outlines the differences of Island Model EA (IMEA) and Diffusion Model EA (DMEA)
with P-EA.
1. All three are based on different underlying conceptions of evolution that translate into their
respective implementations.
2. Elaborate set of parameters, on top of the standard selection, recombination and mutation, are
required in IMEA such as distribution of subpopulations, communication topology, migration
scheme, number of migrants, assimilation scheme etc. As opposed to that current P-EA only
requires radius based clustering and Firstness operator parameters additionally. Hence P-EA
is really a very simple EA model in comparison.
3. As the parallel processing of EA matured it made sense to do an independent component-
wise analysis of IMEA and DMEA, for instance, by keeping everything else the same and
varying the number of subpopulations. The simple P-EA currently does not have independent
components in the same sense. For instance, we could show the difference in results by turning
off the Firstness at one level, but really the Firstness at other levels would still be operative.
4. IMEA as well as DMEA are designed to exploit the efficiency of parallel processing, hence
global selection is avoided as much as possible since it hurts the efficiency and the whole point
68

in parallelization of EA. P-EA are not designed with such bias in mind. In that sense it is
more general.
5. Localization in IMEA or DMEA are artificially constrained by the architecture whereas P-EA
promises a natural yet dynamic division of population based on clustering, within which the
local selection and recombination etc. make more natural sense.
The fact that IMEA is based on Gould’s theory and not the classical Darwinian gradualism
makes our case easier to understand, which is: A different theory or conception of natural evolution
would result in a different EA model which would have different powers. It is still possible for
someone to state that IMEA is purely Darwinian. However, as evident, that is not the case.
Similarly P-EA might have similarities with many models, but it is based on a different theory of
evolution.
4.3.3 IDEA 3 = IDEA 1 + IDEA 2
(Hu et al., 2003) have incorporated both ideas, but they have extended a hierchical fair competition
(HFC) model (as exhibited in sub-cultures and sub-populations within societies, e.g. school systems
(Hu and Goodman, 2002)) for parallel implementation of multi-objetive evolutionary algorithms
(MOEA). They explicitly say that this phenomenon of fair competition between the individuals
having a similar fitness range, not with individuals outside that range, is demonstrated in some
parts of biological and some parts of societal life; not applicable at any wider scale. The P-EA is
general in two respects: 1) It is based on Peirce’s universal theory of evolution, and 2) P-EA is a
general framework which can even provide a theoretical basis for HFC based multi-objective EAs
- this relation does not hold true the other way around.
(Amor and Rettinger, 2005) propose that a successful EA should have three characteristics:
1) good exploration early on, 2) good exploitation later on, and 3) an introduction of novelty
within the population throughout. They use a special type of artificial neural networks called
self-organizing maps (SOM) to help achieve the three tasks. Our contention is that this choice of
SOMs and explicit search histories is not backed up by any natural evolutionary theory. Peirce can
provide them that!
69

Conclusion
(Hu et al., 2003) state in their conclusion that the “current MOEAs still suffer from their convergent
nature inherited from the conventional EA framework.” We go one rational step further and say
that the conventional EA frameworks inherit the problem from the conventional dyadic theory
of evolution. This additional Thirdness and a real Firstness in Peirce brings the balance in the
exploration-exploitation equation.
70

Chapter 5
Experimental Evidence
This is a good time for a re-cap. Darwin’s biological theory of evolution has been successfully estab-
lished, to the extent that it has even found its use in computer science in the shape of evolutionary
algorithms. However, and in time, some of the scientists working in evolutionary sciences seemed
to have hit the wall, with no way out of their problems, while using a strictly Darwinian toolbox.
By now what was once called a modern synthesis has broken down into varying directions which
seem less and less Darwinian in the strictest sense of the word. These are groups with ideas such
as epigenetics, systems biology, neo-mutationists, symbiogenesis, etc. One major problem now in
evolutionary biology is how to bring a new evolutionary synthesis to hold all these varying ideas
together. We have briefly presented Peircean framework as a potential synthesis for these diverging
groups in section 3.3.
Similarly, engineers working with classical evolutionary / genetic algorithms also hit the wall
when they encountered the problem of stagnation. Some have proposed engineering solutions to
varying degree of success, but often at the cost of getting divorced from the underlying natural
theories of evolution (Lozano et al., 2008; Mahfoud, 1995; Sareni and Krahenbuhl, 1998). We
propose a route once followed at the beginning by John Holland, and quite recently reinforced by
Schwefel. We believe that the solution for the problems of EC, inlcuding stagnation, are to come
from nature. We have thus far defined Peirce’s theory of universal evolution and established how
it is more modern and more explanatory than Darwin’s theory. We have also described how a
71

Peircean EA can be carved out of Peirce’s theory of evolution. It is now time to test the strength
of our interpretation of Peircean ideas.
5.1 Experimental Setup
The objective of our experiments is to compare the results of classical Darwinian EA (D-EA)
and Peircean EA (P-EA) on a benchmark suite of problems. The criterion of fairness has been
a priority while designing the experiments for such a comparison. Although we are aware of the
consequences that ‘No free lunch’ theorem (Wolpert and Macready, 1995) would have on any
comparative matrices used between D-EA and P-EA, yet there are limitations to the application
of this theorem as well, especially in coevolutionary algorithms (Wolpert and Macready, 2005).
However, we are still not going to make any claim that our Peircean EA would out-do Darwinian
EA on any given data even though coevolution has similarities with Peircean Thirdness, but any
further development of this thought is beyond the scope of this research work. Another consequence
of this theorem is that there is no unanimously acknowledged set of functions or problems that are
an established benchmark for testing of EA (English, 1996). Literature shows many types of
benchmarks, ranging from purely mathematical functions with known or unknown global minima
(Hwang and He, 2006; Leung and Wang, 2001; Koziel and Michalewicz, 1999) to real world problems
(Hwang and He, 2006; Han and Kim, 2004). The test set used by us (Salomon, 1995) has 4
mathematical optimization functions of different characteristics shown in Table 5.1. The main
reason behind using this benchmark is that it covers a variety of functions, while still being concise.
An elaborate function set is explored in appendix ??.
Table 5.1: Benchmark mathematical functions
Function Limits Value N
f1(x) =∑n
i=1 10i−1x2i −10.0 <= xi <= 10.0 0 5
f2(x1, x2) = 100(x2 − x21)2 + (1− x1)2 −1.5 <= xi <= 1.5 0 2
f3(x) = n+∑n
i=1(x2i − cos(2πxi)) −5.12 <= xi <= 5.12 0 100
f4(x) =∑n
i=1−xi sin(√|xi|) −500 <= xi <= 500 -12569.4537 30
Function f1 is a simple 5 dimensional quadratic parabola with different eigen values along each
72

axis. For optimization the convergence speed usually depends upon the ratio between the smallest
and largest eigen values. Function f2 was proposed by Rosenbrock. The difficult part of this
function is the narrow curved valley that contains the minimum at x = (1, 1). Figure 5.1 shows the
2D function from multiple views.
Figure 5.1: Multiple views of function f2
Function f3 is generalized form of Rastrigin’s function. It is highly multimodal having its
minimum at x = 0. The probability of making progress towards the global minimum decreases as
we move closer to it because of presence of it being surrounded by multiple local minima on all
sides. This characteristic makes it a very interesting function. Figure 5.2 shows different views of
the 2D forms of f3.
Figure 5.2: Multiple views of function f3
73

Last function f4 was proposed by Schwefel, and we are using a 30-dimensional form of it instead
of 20D used in the benchmark just to introduce more rigor. The difficulty posed by this function
is that the global minimum is reached when, ∀i, xi = 420.9687. This makes it an unconventional
function for any EA. Figure 5.3 shows multiple views of 2D form of Schwefel’s function. Detailed
references about these functions are listed in (Salomon, 1995).
Figure 5.3: Multiple views of function f4
This section also illustrates the fairness in our experimental setup. If there is any bias, it is by all
means towards helping D-EA perform optimally, and not the other way around. For instance we did
extensive experiments just to find out the best parameter settings for D-EA on f4, and used that for
quoting the results. Furthermore, we then exported the same parameters to P-EA. The parameters
optimized experimentally include: Crossover probability, mutation probability, number of crossover
points, selection pressure. Final implementation of D-EA is taken from canonical algorithm of (Back
et al., 1997a) with tournament selection having selection pressure of 5, 4-point crossover operator,
and 0.02 mutation rate. Representation and parameter values are listed in Table 5.2. Although
Peircean theory does not restrict us from exploring or inventing new operators, we strictly used
74
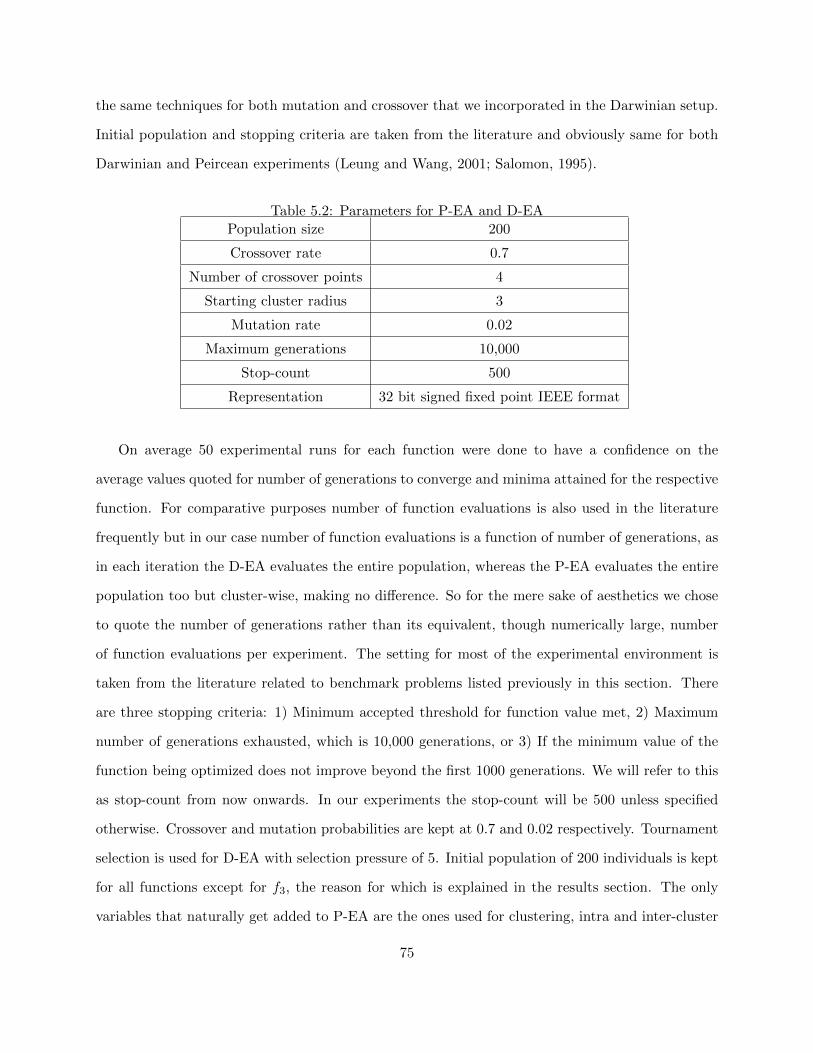
the same techniques for both mutation and crossover that we incorporated in the Darwinian setup.
Initial population and stopping criteria are taken from the literature and obviously same for both
Darwinian and Peircean experiments (Leung and Wang, 2001; Salomon, 1995).
Table 5.2: Parameters for P-EA and D-EA
Population size 200
Crossover rate 0.7
Number of crossover points 4
Starting cluster radius 3
Mutation rate 0.02
Maximum generations 10,000
Stop-count 500
Representation 32 bit signed fixed point IEEE format
On average 50 experimental runs for each function were done to have a confidence on the
average values quoted for number of generations to converge and minima attained for the respective
function. For comparative purposes number of function evaluations is also used in the literature
frequently but in our case number of function evaluations is a function of number of generations, as
in each iteration the D-EA evaluates the entire population, whereas the P-EA evaluates the entire
population too but cluster-wise, making no difference. So for the mere sake of aesthetics we chose
to quote the number of generations rather than its equivalent, though numerically large, number
of function evaluations per experiment. The setting for most of the experimental environment is
taken from the literature related to benchmark problems listed previously in this section. There
are three stopping criteria: 1) Minimum accepted threshold for function value met, 2) Maximum
number of generations exhausted, which is 10,000 generations, or 3) If the minimum value of the
function being optimized does not improve beyond the first 1000 generations. We will refer to this
as stop-count from now onwards. In our experiments the stop-count will be 500 unless specified
otherwise. Crossover and mutation probabilities are kept at 0.7 and 0.02 respectively. Tournament
selection is used for D-EA with selection pressure of 5. Initial population of 200 individuals is kept
for all functions except for f3, the reason for which is explained in the results section. The only
variables that naturally get added to P-EA are the ones used for clustering, intra and inter-cluster
75

functions. We’ve tried radius-based clustering. For intra as well as inter-clustering we have used
the same settings and implementation for crossover and mutation operators that was used in D-EA.
Table 5.2 lists the parameters used and their values, including the common representation scheme.
5.2 Results
This section presents the detailed results for multiple experimental runs against each benchmark
function listed in Table 5.1. Against a total of 50 experiments, four statistics for the following three
aspects are calculated:
value converged at,
euclidean distance of point of convergence from the global optimum point,
and number of generations it took to converge/stop.
The four statistics for 50 experiments against each of the above-mentioned aspects are: minimum
(or best), maximum (or worst), mean and standard deviation. Table 5.3 shows these 12 statistics
when D-EA is tested on the four benchmark functions while Table 5.4 shows the same for P-EA.
Function f1 is the simplest out of the lot. It’s a simple quadratic with a smooth gradient towards
the single minimum in the search space. The only complexity is perhaps added by its 5 dimensions.
Both D-EA and P-EA show the same performance in terms of the number of generations as shown
in the column titled f1 in tables 5.3 and 5.4, while both P-EA and D-EA manage to converge
sometimes at the global minimum fitness value of 0 shown by best value. However, this is the only
similarity between the two. The main difference is that P-EA achieves this global minimum value
with a very small standard deviation. The other difference is that the worst value achieved by P-EA
(0.00169373) is still several times better than the mean value achieved by D-EA (0.00723572). Both
these statistics show the consistency of P-EA.
f2 is a 2 dimensional problem but apart from the difficult and narrow contour leading to the
global minimum, literature shows a small population size of only 15 rather than 200 against this
function, which adds to the complexity. While D-EA without exception (std. generations = 0) stops
because of stop-count after 1500 generations, P-EA gets consistently close to the global optimal
value and location. The fact of consistency is captured by the difference in the standard deviation
76

for the converged values for P-EA and D-EA respectively.
Table 5.3: Compiled results for 50 runs of D-EA on f1, f2, f3 and f4, with stop-count = 500
f1 f2 f3 [100-D] f4 [30-D]
Best value 0 0.03668213 17.20363923 -11073.16103647
Worst value 0.15411377 20.54306412 36.23110552 -9289.66431338
Mean value 0.00723572 2.79967430 26.60560718 -10187.67149048
Std value 0.02220748 4.32435847 4.08085164 404.02023424
Min distance 0 0.03125 4.03112533 1883.30742526
Max distance 2.00000763 3.83051584 5.84107118 3271.54978445
Mean distance 0.04645569 2.53123548 4.98577523 2755.63172195
Std distance 0.28198878 0.86250604 0.38016095 314.59786089
Min generations 17 1500 3005 1693
Max generations 1500 1500 4751 2906
Mean generations 1329.66 1500 3955.72 2275.38
Std generations 466.39 0 418.0183 283.18868665
The complexity in function f3 is due to the fact that as we move closer to the global minimum,
there are a huge number of local minima surrounding it on all sides which makes it highly probable
for an EA to get stuck. D-EA got affected by these pockets of local minima, as shown by the mean
distance from the global minimum it stopped at, while P-EA consistently converged. The difference
between the mean values in both tables is significant for f3. At the same time, P-EA converges to
acceptable minimum value in far lesser number of generations as compared to D-EA.
Schwefel’s function or f4 in our case is perhaps the most complex of this benchmark. Likewise
the gap between the mean values converged at by D-EA and P-EA is widest in this case. While
D-EA barely crosses the -10,000 mark on average and never touches the global minima once, P-EA
shows a good mean value of -12,500, and reaches the known global minimum value of -12,569.4 at
least once in the 50 experiments, as shown in Table 5.4. However, there is a trade off, the P-EA
converges to acceptable minimum value in greater number of generations as compared to D-EA.
But on a closer inspection, the reason why D-EA stops comparatively earlier is because of meeting
the third stopping criterion: the stop-count, as it fails to meet the first two stopping criteria. Since
P-EA in its evolutionary run keeps on exploring and finding better individuals, hence it does not
77

stop because of stop-count or by reaching the maximum limit on the number of generations. It
often stops because of the fact that the minimum accepted threshold for function value had been
evolved, as evidenced by mean value (and distance) close to global minimum and in comparison to
D-EA, a very low standard deviation value.
Table 5.4: Compiled results for 50 runs of P-EA on f1, f2, f3 and f4, with stop-count = 500
f1 f2 f3 [100-D] f4 [30-D]
Best value 0 0 0.00043512 -12569.48476389
Worst value 0.00169373 0.00107193 0.00998695 -12217.85535537
Mean value 0.00024567 0.00004288 0.00691935 -12500.62990597
Std value 0.00052553 0.00021219 0.00169906 92.42908979
Min distance 0 0 0.00390625 0.12129379
Max distance 0.00676582 0.13975425 0.02194551 1384.65925883
Mean distance 0.00339717 0.01257788 0.01744841 364.52082151
Std distance 0.00280038 0.03367504 0.00546443 450.18003363
Min generations 72 6 1856 2875
Max generations 2484 5953 3087 8310
Mean generations 1352.24 1198.9 2308.22 5552.24
Std generations 571.26 1370.97 303.98 1400.34
Another series of experiments for Schwefel’s function were done keeping stop-count as variable
for P-EA. The objective was to see the trend of average minimum value found at the cost of
increasing average generations allowed. P-EA consistently reached around the desired minimum
in less than 10,000 generations (which is the max generations allowed) as shown in Table 5.5. An
interesting pattern in P-EA is the increasing accuracy in terms of mean minimum value attained
as the stop-count condition is relaxed without compromising much in terms of mean number of
generations. No such meaningful pattern was found for D-EA. The best figures for average minimum
value were attained for P-EA when bias tests were conducted. As discussed in section 4, Peircean
framework allows us to use aprior bias or information that we have regarding the problem. Assuming
we took advantage from the shape of Schwefel function, and placed a couple of individuals in the
initial random population having a bias such that those two individuals had for all xi = 400. Last
column in Table 5.5 shows the results where in less than 3000 generations on average, P-EA gets
78

closest to the globally known minima each time. Although this performance was expected with
such an initial bias, but the important factor here is that Peircean framework has a provision for
bias.
Table 5.5: P-EA on f4 with varying stop-counts
Stop-count = 500 Stop-count = 1500 Max gen Bias test
Experiments 50 50 50 10
Best value -12569.4847 -12569.4856 -12569.4858 -12569.4849
Mean value -12500.6299 -12548.1603 -12560.2230 -12568.8151
Mean distance 364.5208 106.3488 66.6988 1.4409
Mean generations 5552.24 5903.9 6382.12 2930.2
Figure 5.4 shows the convergence of fitness for the 1000 generations against all four benchmark
functions. The solid line depicts P-EA while the dotted one shows D-EA convergence graph. It
shows that as the functions become progressively difficult the gap between P-EA and D-EA widens,
and eventually P-EA converges around the global optimum, whereas D-EA gets stuck in some local
optima.
0 200 400 600 800 1000 1200−50
0
50
100
150
200
250
300
350Convergence of P−EA vs C−EA on F1 (1000 gen.)
P−EAC−EA
(a) f1
0 200 400 600 800 1000 1200−2
−1
0
1
2
3
4
5
6Convergence of P−EA vs C−EA on F2 (1000 gen.)
P−EAC−EA
(b) f2
0 200 400 600 800 1000 12000
50
100
150
200
250
300
350Convergence of P−EA vs C−EA on F3 (1000 gen.)
P−EAC−EA
(c) f3
0 200 400 600 800 1000 1200−14000
−12000
−10000
−8000
−6000
−4000
−2000
0Convergence of P−EA vs C−EA on F4 (1000 gen.)
P−EAC−EA
(d) f4
Figure 5.4: Convergence comparison: P-EA vs. D-EA
79

Finally we compare P-EA with other EA in Table 5.6 and Table 5.7 for functions f3 and f4
respectively. The list includes: Orthogonal Genetic Algorithm with Quantization (OGA), Fast
Evolution Strategy (FES), Particle Swarm Optimization (PSO), Evolutionary Optimization (EO),
two versions of Quantum-inspired Evolutionary Algorithm (QEAH and QEAR), a Fast Evolutionary
Programming (FEP), and a pair of conventional/classical Genetic Algorithms and Evolutionary
Programming (CGA and CEP). Results of OGA and CGA are referred from (Leung and Wang,
2001); FES from (Yao and Liu, 1997); PSO and EO from (Angeline, 1998); QEAH, QEAR, FEP
and CEP from (Han and Kim, 2004). The last two rows of the two tables show the results for our
implementation of D-EA and P-EA (with stop-count = 500). A point to note here is that some of
the above-mentioned EA are optimized in ways that the P-EA is not. For instance, OGA, which
apparently gives the best figures, has been optimized to explore the solution space along orthogonal
dimensions. Still, the rudimentary P-EA fares well, as it converges near the optimal location (only
OGA is better) for 30 dimensional case of f3, that too in the minimum number of generations. For
the more complicated Schwefel’s function, its performance is still comparable.
Table 5.6: P-EA compared with other EA on f3 [30-D]
Mean Value Std Value Mean Function Evaluations
OGA 0 0 224,710 feval
FES 0.16 0.33 500,030 feval
PSO 47.1354 1.8782 250,000 feval
EO 46.4689 2.4545 250,000 feval
CGA 22.967 0.78 335,993 feval
QEAwH 0.039 0.19 5000 gen
QEAwR 18.7 7.4 5000 gen
FEP 0.046 0.012 5000 gen
CEP 89 23.1 5000 gen
D-EA 5.1638 2.5315 1549.4 gen / 309,880 feval
P-EA 0.0087 0.0006 445.32 gen / 89,064 feval
80

Table 5.7: P-EA compared with other EA on f4 [30-D]
Mean Value Std Value Mean Function Evaluations
OGA -12569.4537 6.447 x 10−4 302,166 feval
CGA -8444.7583 65.7326 458,653 feval
FES -12556.4 32.53 900,030 feval
QEAwH -12569.48662 3 x 10−9 9000 gen
QEAwR -12353.447 163.8 9000 gen
FEP -12554.5 52.6 9000 gen
CEP -7917.187 634.5 9000 gen
D-EA -10187.6715 404.0202 2275.38 gen
P-EA -12500.6299 92.4291 5552.24 gen / 1,111,0448 feval
5.3 Results on an Extended Benchmark
Another set of results on an extended benchmark of 12 mathematical functions of up to 100 di-
mensions is listed in this section. Figure 5.5 lists the forms of the benchmark functions. For the
lack of space refer to Leung and Wang (2001) for detailed description of each function. Against
most of the functions the 30 dimensional form has been used, while F7 and F9 are 100-D. Table 5.8
lists the comparative results of both the Peircean (P-EA) and the classical Darwinian evolutionary
algorithm (D-EA) when they are run for 50 times against each function. For 50 independent runs,
what the table lists against each function is how P-EA and D-EA perform statistically in terms
of, 1) the average number of generations it took the EA to converge, 2) the average of the best
function values that the EA converged at, and 3) the standard deviation of those values.
The most interesting observation is that apart from one exception (F14), P-EA, by the time it
stops, is almost always very close to the global minimum value for each function, that too with a
small standard deviation value. This is not the case for D-EA. But more important than that is
the fact that for P-EA evolvability, improvement and growth seem to be vital. In terms of number
of generations (or function evaluations equivalently) P-EA seems to be more efficient or at par with
D-EA in most of the cases. In some cases where D-EA makes an early stop (e.g. F7,F9,F10,F14) it
is always the case where D-EA has stopped because of premature convergence to a local minimum
due to stagnation of its population. This fact is evident by looking at the respective mean function
81

value columns. In all of these tough specific cases, P-EA keeps on evolving its population and gets
much closer to the global minimum before stagnating. The results would make more sense when
the above-mentioned analysis is coupled with the stagnation analysis and cluster analysis, detailed
in the next chapter.
Figure 5.5: Extended Benchmark Functions
82

Table 5.8: Comparison between Peircean-EA and Darwinian-EA
F.Mean number ofgenerations
Mean functionvalue (standarddeviation)
Globalminfunc.value
P-EA D-EA P-EA D-EA
F1 2895.2 2286.48-12569.3(0.6195)
-10962.6(305.88)
-12569.4
F2 2308.22 3955.720.0069(0.0017)
26.61(4.08)
0
F3 1502.82 1513.740.2730(0.0324)
2.3272(0.2686)
0
F4 1618.92 1689.120.0198(0.0233)
0.4264(0.4000)
0
F7 8984.52 2656.7-95.08(0.8304)
-73.93(1.8858)
-99.2784
F9 8414.04 3826.02-78.32(0.0087)
-58.68(1.5735)
-78.3324
F10 7840.6 1764.0652.71(30.02)
1899.87(900.19)
0
F11 1502.74 1527.760.0570(0.0097)
0.0780(0.1436)
0
F12 1620.76 1606.70.5241(0.2964)
0.5969(0.3046)
0
F13 1500 1647.40.0146(0.0023)
0.0274(0.1416)
0
F14 10000 2770.183307.14(2946.47)
325689.5(92779.9)
0
F15 2750.94 3120.780.4978(0.5710)
29.1570(13.3290)
0
83

84

Chapter 6
Analyses
The empirical data on the benchmark functions suggests that P-EA holds promise for the EC
community even from the perspective of function optimization, as it has good exploratory powers.
The question however still remains as to what causes the P-EA to keep on exploring till it converges
near the global optima. One intuitive claim is based on Schwefel’s hypothesis, that P-EA performs
well because it mimics the natural processes of evolution better than the classical EA. In this
chapter there are three aspects that need to be analyzed for P-EA: 1) how long before the diversity
of the population is lost; 2) what is the effect of number of clusters on diversity; and, 3) the effect
of clusters on Holland’s schema theorem.
6.1 Stagnation Analysis
Stagnation has been a major problem noted in the literature with respect to population based
EA (Deb, 2001; Whitley, 1994). Allegations are leveled against selection and the fittest or best
individual taking over the entire population. Consequently, there are various stagnation avoidance
techniques employed in the literature as well (Friedrich et al., 2008; Sareni and Krahenbuhl, 1998).
It is hypothesized that the problem might be rooted in the Darwinian paradigm itself, hence any
cosmetic solution will either be artificial or not from within the Darwinian framework. Peircean
EA however, does not use any special technique to avoid stagnation. Whatever it is that causes the
delays in its population getting stagnated comes from within the Peircean framework - for instance
85

its cluster based retention of population (Thirdness) and Firstness principles of spontaneity. For
monitoring the stagnation a simple analysis was made. At the end of each generation or iteration,
the percentage difference between the fittest individual and the entire population was calculated.
There could be other metrics for calculating stagnation but the choice of this metric is justified by
the fact that in D-EA stagnation is primarily caused by the fittest individual replicating itself in
the next generation while getting selected with most probability (Deb, 2001; Friedrich et al., 2008;
Whitley, 1994).
The results in Table 6.1 show the average generations for both D-EA and P-EA against each
function up till which the population had stagnated by 99%. D-EA stagnates very early on and
this fact is translated in all the results as seen above. This behavior is consistently shown for each
function. P-EA on the other hand shows greater resilience towards stagnation and the average
number of generations before it stagnates is in quite contrast to that of D-EA. This gives the P-EA
diversity in the population for a much longer time frame and hence there is a greater chance that
P-EA does not converge at a local optima. This is one of the reasons why P-EA shows meaningful
results and patterns in the experiment.
Table 6.1: Number of generations before the population stagnates,20 experiments
Fn. EA Min Max Mean Std
F1 D-EA 25 54 38 7
P-EA 100 3696 851 1133
F2 D-EA 4 13 7 2
P-EA 82 2091 704 558
F3 D-EA 22 69 37 12
P-EA 361 1099 511 188
F4 D-EA 40 56 46 4
P-EA 335 924 530 156
6.2 Cluster Analysis
In Holland’s schema theorem formulation, disruption analysis is already well known for suggesting
how potentially good schemas can be disrupted early on by the crossover operator (Chen and Smith,
86

1999). This has implications for all EA that use crossover operators. We hypothesize that those EA
that partition the population into subpopulations minimize the chances of disruption by crossover.
Peircean-EA first divides the population into clusters and then performs the within-cluster crossover
to generate the next batch of individuals for each cluster, followed by the crossover of the fittest
individuals from across clusters. To monitor the effect of number of clusters on the continual
evolution of good schemas, the following experimental setting was devised. The population size
was fixed to 200 individuals, mutation rate to 0.02 and crossover to 0.7, while the stop-count is fixed
to 500. To make a fair trial, it was necessary to turn off Firstness completely, so that if increasingly
fit individuals keep on getting generated it cannot be because of spontaneity. Instead of uniform
random selection, tournament selection with tournament size of 2 was introduced, so that the only
thing which separates a D-EA and this constricted version of P-EA is that the individuals are
evolved in clusters rather than as one global unit. The only stopping criterion is the stop-Count,
which means that when no better fit individual is found in the last 500 generations, the EA stops
due to this stagnation. To complete the experiment Schwefel’s 30 dimensional function is chosen,
the number of clusters is increased iteratively from 1 to 25 and 10 trials against each number of
clusters are made. The mean number of generations of those 10 trials is plotted against number of
clusters in Figure 6.1.
As is obvious from the y-axis, the increase in clusters does result in prolonged diversity and
exploration up to a point, but beyond that the EA begins to stagnate earlier. Following analysis
explains the behavior observable in the above figure:
1. As the number of clusters is increased from 1, due to less global interactions the retention and
evolution of good schemas becomes more and more probable, as indicated by the prolonged
termination of EA due to stop-count. So for longer duration the better fitted schema keep
on getting explored rather than disrupted.
2. But there is not a linear relation between the number of clusters and the retention/evolution
of schemas, firstly due to a finite and static population size. For instance, against a population
size of 200 and the number of clusters being 25, only 8 individuals on average belong to any
cluster. This is too inexpressive a gene pool to help in evolution, as observable in the latter
87

0 5 10 15 20 251950
2000
2050
2100
2150
2200
2250
2300
2350
Number of Clusters
Mea
n N
umbe
r of
Gen
erat
ions
for
Exp
lora
tion
Effect of Number of Clusters on Stagnation
Figure 6.1: Cluster Analysis: Effect of clustering on stagnation using Schwefel’s function
part of the graph.
3. But mainly because as the number of clusters is further increased, inter-cluster crossover
begins to act more disruptively, since intra-cluster would now be a crossover of decreasing
number of individuals closer together in each cluster, while inter-cluster crossover would be
between an increasing number of individuals, and would become characteristic of the global
population of a D-EA.
6.3 Schema Theorem
What is a schema? A schema is John Holland’s formulation of a building block, a string comprising
of either 0s or 1s or * (asterisks). For instance if H = 1****0 is the schema, then it represents all
possible binary individuals in a 6 dimensional search space that begin with a 1 and end at 0. Given
this definition of schema H, there are four more definitions,
1) l, which is the lenght of schema H, here 6;
88

2) d(H), which is the defining length of schema H, in other words, the distance between the
outermost defined bits, here 6-1 = 5;
3) o(H), or the order of schema H, which is given by the total number of non-asterisks in the
schema, in this case 2; and
4) m(H, t), which is the number of instances of H present in the GA population at time t.
Now given all of the above, schema thoerem makes a prediction about E(m(H, t + 1)), which
is the expected number of instances in the population belonging to the schema H surviving till the
next generation. Of course, E(m(H, t+ 1)) must be a factor of m(H, t), however, in classical EA,
three operators affect it: selection, crossover, and mutation.
1) Selection
A standard fitness proportional selection affects the m(H, t) by the following ratio:
E(m(H, t+ 1)) =f(H)
f(p)∗m(H, t) (6.1)
where, f is the fitness function, f(p) is the sum fitness of the entire population, and f(H) is the
sum fitness of those instances belonging to the schema f(H) =∑∀x∈H
f(xi).
2) Crossover
For a single point crossover operator, we can see the more the value of d(H) with respect to
l, higher are its chances of getting disrupted by the crossover. Since the chance of an individual
taking part in the crossover is driven by the crossover probability (pc), hence the chances of the
individual still surviving and being a part of the schema are(
1− pc(d(H)l−1
)), updating equation
6.1 as:
E(m(H, t+ 1)) ≥ f(H)
f(p)∗m(H, t) ∗
(1− pc
(d(H)
l − 1
))(6.2)
3) Mutation
For a mutation rate of pm, the defined bits (o(H)) instances belonging to schema have a survival
chance of (1− pm)o(H), updating equation 6.2 as follows:
89

E(m(H, t+ 1)) ≥ f(H)
f(p)∗m(H, t) ∗
(1− pc
(d(H)
l − 1
))∗(
(1− pm)o(H))
(6.3)
Typically pm is kept very low, such that pm � 1 =⇒((1− pm)o(H)
)≈ 1−o(H)pm, simplifying
the equation 6.3 to:
E(m(H, t+ 1)) ≥ f(H)
f(p)∗m(H, t) ∗
(1− pc
(d(H)
l − 1
))∗ (1− o(H)pm) (6.4)
Equation 6.4 captures Holland’s schema theorem for building block hypothesis. What it suggests
is that the number of individuals belonging to the schema that survive the disruption and are still
above average in terms of fitness get increased by a factor of f(H)f(p) every generation!
6.3.1 Clusters as Schema - Effects on Disruption Analysis
Ignoring the effect of mutation in equation 6.4, and denoting the disruption probability caused by
the crossover as dc, we get the following representation:
E(m(H, t+ 1)) ≥ f(H)
f(p)∗m(H, t) ∗ (1− dc) (6.5)
“The schema theorem given in equation 6.4 applies not only to schemas but to any subset of
strings in the search space” (Mitchell, 1998). When we apply it to clusters, meaning, when a cluster
represents a schema, how does that effect the equation for a Peircean EA?
It means that the crossover operator which potentially caused disruption when the entire popula-
tion was classically treated as one big cluster, would not be a problem when it comes to intra-cluster
evolution, since the schema is now the cluster, so any two instances belonging to the cluster, when
crossed-over, would still result within the search space covered by that cluster. Any disruption that
is to come, is to come from the crossover taking place in the inter-cluster evolution. If dc is the
disruption probability for one instance of a schema, this factor gets further reduced by 1m(H,t) ; since
only one instance (fittest) of the cluster can, at max, get disrupted by the inter-cluster evolutionary
crossover now, per generation. This makes the following survival rate when a cluster is a schema:
1− dcm(H,t)
90

Within the inter-cluster evolution, the selection factor becomes a ratio of f(Hfittest) and f(C),
where f(C) is the average fitness of all the fittest individuals, one from each cluster. Putting these
two factors in equation 6.5 updates the schema theorem for clusters as follows:
E(m(H, t+ 1)) ≥f(Hfittest)
f(C)∗m(H, t) ∗
(1− dc
m(H, t)
)(6.6)
So, when H is a cluster, and if we have only one cluster, then it means that
f(Hfittest)f(C) = 1, and
(1− dc
m(H,t)
)= 1, since dc = 0 m(H, t+ 1) = m(H, t)
But for any higher number of clusters, equation 6.6 suggests that those clusters H whose best
fitness > average fitness of all clusters’ best instances, keep on surviving, evolving and thriving.
But more importantly, perhaps, those H that are not fit enough relatively speaking, at any time t,
become extinct slowly; it may take m(H, t) generations before they lose all their instances.
6.4 Summary
The previous chapter was focused on guaging the performance of the P-EA, especially in contrast
with the D-EA. This chapter focused itself on why the performance of P-EA came out as a more
robust EA of the lot. The hunch that the factors of Firstness (continual introduction of individ-
uals to the population) and Thirdness (cluster based population dynamics,) besides the standard
selection and variation operators in EA, has been evaluated in this chapter. The three investigative
questions analyzed for P-EA were: 1) how long before the diversity of the population is lost; 2) what
is the effect of number of clusters on diversity; and, 3) the effect of clusters on Holland’s schema
theorem. The finding of the first investigation is that both P-EA as well as D-EA stagnate, but
P-EA stagnation is delayed by quite a margin comparitively speaking. The finding of the second
investigation is that turning every factor off in P-EA that departs from our implementation of a
standard D-EA, apart from clustering in P-EA, still makes a difference in terms of it’s exploratory
potential. Two less a number of clusters, and too many clusters both result in a deteriorated ex-
ploration and early stagnation. Finally, the theoretical finding in the last investigation suggests
that clusters with good fitness have a greater potential for growing, exploring and exploiting, while
91

those with weaker fitnesses do not die out quickly, rather they also get a fair chance of explorative
survival before getting extinct.
92

Chapter 7
Future of EC & our Conclusions
This chapter lists the future directions as hypothesized by the pioneering figures from EC - De Jong
and Schwefel. It also then maps the Peircean framework on to those future directions. Towards
the end some conclusions are presented.
7.1 Hans-Paul Schwefel’s Future Directions for EC
Schwefel lays out a comprehensive plan of how to engage with the future directions in EC (Schwefel,
1997). Schwefel identifies that “there is a bulk of as yet unincorporated phenomena and mechanisms
of organic evolution underpinning the hope for further breakthroughs in devising ever more useful
evolutionary algorithms.” He suggests that by comparing what is deficient in current EAs with
what is empirically and observably present in organic evolution and nature, further improvements
and directions in EC development can be defined:
“Current evolutionary algorithms are certainly better models of organic evolution. Nev-
ertheless, they are still far from being isomorphic mappings of what happens in nature.
In order to perform better, an appropriate model of evolution would have to comprise
the full temporal and spatial development on the earth (a real global model) if not
within the whole universe. We must be more modest in order to understand at least a
little of what really happens – as always within natural sciences.”
93

In general we term this as Schwefel’s hypothesis, but additionally he has also pointed out
the specific aspects of organic evolution which should be modelled by an EA. We shall represent
Schwefel’s future directions point by point, using his own words:
1) Evolvability: Organic evolution certainly does not only aim at finding static optima just once
and with ultimate precision. Organic evolution happens within an ever-changing environment,
where evolvability is more important than precision.
2) Dynamic interaction of agents: The environment is not only intrinsically dynamic; it is
changed by the mutual actions of all participants in the evolutionary game. From the per-
spective of one species, the search for meliorization (a term preferred to optimization by the
author) takes place on a trampoline, deformed by (re-)actions of other species, as well.
3) Multiple selection criteria: Organic evolution always deals with a situation of multiple se-
lection criteria.
4) Spatial distribution of population: Larger populations in the real world are spatially dis-
tributed (small populations are always prone to extinction). Neither mating nor predation
thus takes place in a way that includes all individuals with equal chance. There is no global
selection in either case. Consequently modeling selection as an asynchronous and spatially
distributed process with several predators at a time among the prey, which is the normal case
in ecological systems, has not yet been done but should be considered.
5) Cooperation as important as competition: Many attacks against EAs as optimization, or,
better, meliorization, algorithms have been mounted by those who emphasize cooperative be-
havior within evolutionary processes. This controversy is not necessary. Cooperative problem
solving by sharing resources or dividing the problem into subproblems to conquer may lead
to novel approaches not yet taken into account thoroughly. At least, theoretical results are
missing.
6) Self-adaptive parameter rates: ES with self-adaptive individual mutation rates: Very low
rates as observed in temporary optimal or near-equilibrium situations may not be good for
94

starting conditions. Autoadaptation of strategy parameters, be they discrete or real valued,
can and has been handled in two different ways: these parameters vary either from individual
to individual or from subgroup to subgroup. The possibilities of on-line adaptation of a
variable epigenetic apparatus are by far not exhausted in all EAs today.
7) Social learning and epigenetic factors: Haeckel component of an evolutionary algorithm
can be taken as any kind of individual adaptation to the local – temporal environmental
conditions; but the result of this cannot be transferred genetically to the next generation.
Completely different from this mechanism is the life-long individual learning and all kinds of
social transmission of the knowledge (and prejudice) gained this way. EA individuals so far
have no kind of brain of their own but this might be added in the future.
8) Gender: [EA] do not yet handle two (or more) sexes really ... [for] a purposeful employment
in optimization.
9) Variable genome length: Genome length variation should be an important ingredient in all
cases of so-called structure optimization tasks, where the number of variables is not known
in advance, but is a variable itself.
7.1.1 Equivalence between Peircean Framework and Schwefel’s Future Direc-
tions
“The invention of broadly efficient [evolutionary] algorithms is a design challenge as stiff as the
most difficult that have been faced this century.” Here (Goldberg, 1993) effectively says that ad-
hoc design “hacks” often do not work in EA. It is not the case that we can take Schwefel’s guidelines
point by point and build a piece-meal yet efficient algorithm. This section highlights how most of
Schwefel’s demands for the future of EA are met naturally by Peircean evolutionary framework.
Their relationship is also captured in Table 7.1.
1) Evolvability: Thirdness of Peirce is a true representation of what Schwefel means by evolvabil-
ity. It is this process of evolutionary growth through generalizing tendency that converts a
possibility of getting to the global optima into an actuality. Otherwise, stressing on efficiency
95

and precision through selection has been the primary cause of premature convergence Her-
rera and Lozano (2000). In P-EA the focus on selectionism is balanced due to the dynamics
introduced by Thirdness, such as parameter tuning and clustering. This makes it possible to
stress more on the function of evolvability, as all the clusters separately evolve.
2) Dynamic interaction of agents: Through the inter-cluster operations, the fittest clusters
affect other clusters and individuals. Eventually the clusters get pulled by the fittest towards
the best explored optima. This action and reaction of different clusters addresses Schwefel’s
call for meliorization.
3) Multiple selection criteria: Unlike a global flat selection operator, P-EA has multiple pos-
sibilities:
• There are multiple levels, and hence selection can take place at individual as well as
cluster and global levels
• Due to clusters, there is a natural possibility of having multiple selection operators now
• We have experimented by introducing a cooperative selection operator as well, but this
idea has unexplored potential for future work
4) Spatial distribution of population: By evolving the populations in clusters, P-EA directly
meets this demand of Schwefel.
5) Cooperation as important as competition: There are three sides to this party. On the
one hand is the group which reduces every phenomenon to competition, and consequently all
empirical behaviors of cooperation are also explained away as epi-phenomenon, only keeping
selfish competition as real. The group at the other extreme suggests the opposite. They read
everything from a cooperative angle (Todes, 1987). Peirce on the other hand is the third.
He acknowledges reality in its multiplicity. His understanding of evolution has real place for
competition as well as cooperation, given certain conditions. This fact translates into P-EA,
as there is clustering and cooperation as well as selection of the fittest through competition.
96

6) Self-adaptive parameter rates: Peircean system does not take anything as a given, as Peirce
applies his evolutionary principles at the cosmological levels as well. For him even the physical
laws are at different evolutionary stages. The presence of exceptions against every law is a
sign for that. Any EA that also evolves the parameters would be more Peircean in spirit. We
have only emulated this self-adaption of cluster radii and mutation rate so far, as described
in chapters above.
7) Social learning and epigenetic factors: P-EA does employ the social learning especially
when the inter-cluster fittest individuals change within their lifetime, and when during the
addition of Firstness individuals to the population, some are locally affected by their cluster
representatives. At the moment our P-EA is handicapped by the fact that there is nothing
much beyond the genetic makeup, and that is also primarily due to the type of mathematical
benchmark we have used. Perhaps for another type of problem set, a brain or extra-genetic
elements could be added to the genotype and then epigenetic operators could be involved
more meaningfully.
Table 7.1: Relation between Peircean framework and Schwefel’s future challenges
Schwefel’s Research Guide-lines
Peircean EA
1 Evolvability Thirdness guarantees evolv-ability
2 Dynamic interaction of agents Inter-Cluster evolution
3 Multiple selection criteria Cooperative selection besidesnatural selection
4 Spatial distribution of popula-tion
Clustering
5 Cooperation as important ascompetition
P-EA treats cooperation asreal just like competition
6 Self-adaptive parameter rates Cluster radius, mutation rate
7 Social learning and epigeneticfactors
Firstness, inter-cluster infor-mation exchange
8 Gender N/A
9 Variable genome length N/A
97

7.2 Kenneth De Jong’s Agenda for the 21stCentury and Peirce
While presenting a history of the field of GA, Kenneth De Jong concludes with an extended “agenda
for the 21stcentury” (De Jong, 2005). This section briefly outlines them and then draws parallels
with Peircean framework, wherever possible.
1) Developing a more general EC/EA framework: Pitching the variety of design choices in
classical EAs (GA, ES, EP) and other variant EAs, there is a better chance of understanding
the implications of those design choices when a more general framework of EC is developed,
and the variant EAs are seen as instances of that general framework. Abstractly speaking,
the basic components of that general EC framework are:
• A population of individuals
• A notion of fitness
• A notion of population dynamics biased by fitness
• A notion of inheritance of properties from parent to child
Peircean framework would only add two more building blocks to complete De Jong’s proposal
- which mainly covers Secondness:
• A notion of introducing freshness (Firstness) at various levels
• A notion of Habit/Generalization (Thirdness)
2) Decentralized and Speciation models: EAs, no doubt, are naturally parallelizable. Some
of the evolutionary effects that can be covered by this parallelism include speciation, niching,
and punctuated equilibria, with subpopulations developing in equilibrium according to the
underlying parallel architectural topology, punctuated by infrequent migrations from neigh-
boring subpopulations.
While De Jong admits that the theories for traditional EAs do not help understand the
effects of design choices for these parallel implementations. We believe that the Peircean
98

formulation, both in its form of evolutionary theory, as well as coupled by its EC theory, can
be strengthened further to help the EC community.
3) Self adapting and coevolutionary systems: Natural reality of course shows more complex
phenomena, such as self-adaptive mutation, selection and other parameteric rates. It also
shows individuals and species co-evolving each other in more modes than just through com-
petition. The fact is that the traditional simple EA does not have explicit support for these
complex behaviours. The challenge is to come up with a theoretical framework for EC that
also incorporates self-adaptation and co-evolution.
The previous section already explains well how Peircean EA naturally has a place for both
these complex phenomena. It will make for a very constructive future work to come up with
a formal theoretic formulation that helps in coordination of the self-adaptive parameters.
4) Incorporating more biology into EA, especially Lamarckian ideas: Since Holland’s clas-
sical GA was inspired from Modern Synthesis’ point of view, it is not surprising to see Lamar-
ckian factors being factord out from the GA consequently. There is enough evidence that the
genetic viewpoint is a limited account of evolution, and that the epi-genetic factors are heri-
table and sustain through generations (Noble, 2013).
Although Peircean framework has scope for other-than-genetic learning, it will make for a
very interesting future work to construct intelligent agents that learn over their lifetime and
also transmit this learning to their next generation.
5) Using EA to further our evolutionary understanding: It is always easy to see how an
algorithm can and will be viewed as only a degenerated form of the actual evolutionary
processes at work in natural reality. But one positive strand within EC is to look at EAs as
tools that further our understanding of the natural evolutionary reality. This sentiment is
more vividly elaborated by (Mitchell and Taylor, 1999) and we quote:
Evolutionary computation can sometimes serve as a useful model for biological evo-
lution. It allows dissection and repetition in ways that biological evolution does not.
99

Computational evolution can be a useful tool for education and is beginning to pro-
vide new ways to view patterns in evolution, such as power laws and descriptions of
non-equilibrium systems. Evolutionary theory, as developed by biologists, typically
tries to linearize systems, for ease of analysis with differential equations, or to treat
units in isolation, as in single-locus selection. While evolutionary computation is
not inconsistent with such theory, it tends to be outside it, in that real difference
in capacity and complexity are often observed, and are not really describable by
stable equilibria or simple changes in gene frequencies, at least in ways that are
interesting. There is reason to believe that theories of evolutionary computation
might extend the language of biological evolutionary theory and contribute to new
kinds of generalizations and analyses that have not been available up to now.
We feel that with this experimental verification of Charles Peirce’s ideas, we have not only
tried to put forth a viable EC framework, but have also brought new interpretive and scientific
meaning for the Charles Peirce scholarship and community (Akhtar et al., 2013). We also
hope that this work triggers appropriate discussions within the community of evolutionary
biologists as well.
7.3 Conclusions
Science works by rigorous experimental testing of the theories that make claims or predictions about
the working principles of the natural world. In this article we have presented an interpretation of
Peirce’s theory of evolution, modeled it algorithmically, and tested the model scientifically through
lab experiments. We understand Peirce’s pragmatism to include the idea that in order to attain
more clarity with regard to a concept, one must first of all translate the concept into a set of
testable abductive hypotheses, and then test them under laboratory conditions for their practical
considerations. We feel that, remaining true to computer science, we have formulated testable
hypotheses out of Peirce’s theory and set up experiments that could test their validity.
All other things being equal (experimentally), evolutionary algorithms based on an interpre-
100

tation of Peirce’s theory maintain diversity far longer than those based on Darwin’s theory. This
issue of diversity is significant for the EC community, and any solution to this problem warrants
serious consideration. We have not introduced any ad hoc mechanism in our proposed algorithm
that seeks to redress this problem artificially. If there is an apparent improvement in the main-
tenance of diversity in the Peircean EA (as indicated by the results) we would like to attribute
it to Peirce’s theory of evolution (or our understanding of it). The experimental evidence for the
predictive accuracy of Peircean theory presented in this paper has been consistent so far, at least
in the domain of evolutionary computation.
More importantly, Schwefel’s and Peirce’s abductive hypotheses have passed our inductive tests.
Schwefel’s hypothesis-that the more an evolutionary algorithm models natural evolution, the better
it will perform-is supported by our experimental results. Peirce’s hypothesis-that in the beginning
was chaos and over long periods of time, habitually that chaos is transformed into regularity-has also
been affirmed. Consequently, it also helped explain the fact of stagnation, which has been a problem
for the EC community. Darwin’s theory predicts diversity to result from simple beginnings by using
blind chance and mechanistic laws alone. However, models of Darwin’s theory in computer science,
namely evolutionary algorithms, lead to the opposite of diversity: stagnation at a very early stage.
This unpredicted behavior leads, in optimization terms, to premature convergence, and this has
been amply documented in the literature. Darwin’s theory does not say a word about stagnation,
as it only focuses on diversity, so the issue of stagnation could not get any support from within
the Darwinian paradigm. On the other hand, Peirce’s theory anticipates stagnation (complete
regularity) in the long run. This was confirmed by the experiments as well. We feel that we have a
strong case of understanding Peirce’s theory pragmatically here, given the constraints of computer
science. Of course the real pragmatic tests would need to come from evolutionary biology.
We have also indicated how new findings in cellular biology, epigenetics and systems biology are
already unknowingly heading in the direction of Peirce and knowingly heading away from Darwinian
notions. It is concluded that our interpretation of Peirce’s work is significant enough to help solve
some of the contemporary problems in evolutionary algorithms as well. We think that the passing
of this pragmatic test, plus the movement within contemporary physics as well as biology, are
101

convergent signs that point towards the truth of Peirce’s evolutionary framework.
102

Appendix A
Cooperation vs. Competition in
Evolution
A.1 Cooperation
A recent publication conducting a 43 years survey of the theoretical and empirical literature on
cooperation by (West et al., 2007) suggests:
Explaining cooperation remains one of the greatest challenges for evolutionary biology,
irrespective of whether it is altruistic or mutually beneficial.
Darwin was aware of the problem in his lifetime, and flirted with some ideas of group selection
to work around it. In Darwin’s own words (Darwin, 1859):
I will not here enter on these several cases, but will confine myself to one special difficulty,
which at first appeared to me insuperable, and actually fatal to my whole theory. I
allude to the neuters or sterile females in insect-communities;.. This difficulty, though
appearing insuperable, is lessened, or, as I believe, disappears, when it is remembered
that selection may be applied to the family, as well as to the individual, and may thus
gain the desired end. (Chapter 7, pages 236-237)
103

Natural selection cannot possibly produce any modification in any one species exclu-
sively for the good of another species; though throughout nature one species incessantly
takes advantage of, and profits by, the structure of another. But natural selection can
and does often produce structures for the direct injury of other species, as we see in
the fang of the adder, and in the ovipositor of the ichneumon, by which its eggs are
deposited in the living bodies of other insects. If it could be proved that any part of the
structure of any one species had been formed for the exclusive good of another species,
it would annihilate my theory, for such could not have been produced through natural
selection. (Chapter 6, page 200)
Natural selection will modify the structure of the young in relation to the parent, and
of the parent in relation to the young. In social animals it will adapt the structure of
each individual for the benefit of the community; if each in consequence profits by the
selected change. What natural selection cannot do, is to modify the structure of one
species, without giving it any advantage, for the good of another species. (Chapter 4,
page 86)
Professor Stephen Stearns, an accomplished neo-Darwinist evolutionary biologist at Yale’s Ecol-
ogy department (Stearns, 2009b), has summarized Darwin’s words in this fashion: If ever it could
be shown that individuals repeatedly and reliably sacrificed their own fitness to in-
crease the fitness of others, the theory of natural selection would be refuted. The
post-Darwinian work which has been referred to as neo-Darwinism has persistently tried to ex-
plain cooperation, altruism, and symbiosis by first and foremost reducing it to various forms of
selfishness. This is one hypothesis. Another equally valid hypothesis is that cooperation is an
independent agent in the evolutionary process - at least as much a factor as mutation (chance) and
selfish struggle (competition). Cooperation is understood as being irreducible to chance mutation or
selfishness or competition - it is actually the opposite of both. Cooperation means that selfish
replicators forgo some of their reproductive potential to help one another (Taylor and
Nowak, 2009). What neo-darwinists miss is that cooperation could be understood as a deliberate,
purposive collaboration or association: a long term process rather than a cost-benefit happenstance.
104

From the perspective of our thesis the most important difference between the Darwinian theory
of evolution and the Peircean alternative is that the latter identifies various forms and levels of
cooperation as genuine factor in the process of evolution whereas the former sees cooperation as
an epiphenomenon which in reality is nothing other than selfish cost-benefit calculations. We shall
now entertain both these hypotheses one by one.
A.1.1 Hypothesis 1: Only selfishness is real, cooperation is not
The plentiful forms of selection employed by neo-Darwinism are simply variants of the first hypothe-
sis - only selfishness is real and any appearance of cooperation or altruism is an epiphenomenon that
can ultimately be explained in terms of selfishness. All recent neo-Darwinist attempts to reduce
cooperation to selfishness are themselves the result of an evolutionary process within Darwinism.
The first stage was the abstract theory of group selection, an idea explored by Darwin himself as ev-
ident from some of his work quoted above. While it solved some cases, it had a major shortcoming:
group selected altruism results in evolutionarily unstable strategy, as it favors the selfish mutant
invader over the otherwise cooperating group of un-selfish individuals. This was followed by the
theory of kin selection, which avoided the selfish invader problem through cost-benefit calculations
proposed by Bill Hamilton’s equations. Kin selection operates at the genetic level and it involves
maximizing the inclusive fitness of kin even if the individual has to bear the cost of sacrificing its
own phenotype under certain scenarios. This theory did help explain some of the altruistic and
cooperative behavior in nature, but factually not all the individuals involved in cooperation in the
natural world are kin. Furthermore, any theory using inclusive fitness makes certain mathematical
assumptions (Nowak et al., 2010):
First, for inclusive fitness theory all interactions must be additive and pairwise. This
limitation excludes most evolutionary games that have synergistic effects or where more
than two players are involved. Many tasks in an insect colony, for example, require the
simultaneous cooperation of more than two individuals, and synergistic effects are easily
demonstrated.
Second, inclusive fitness theory can only deal with very special population structures.
105

It can describe either static structures or dynamic ones, but in the latter case there
must be global updating and binary interactions. Global updating means that any two
individuals compete uniformly for reproduction regardless of their (spatial) distance.
Binary interaction means that any two individuals either interact or they do not, but
there cannot be continuously varying intensities of interaction.
These particular mathematical assumptions, which are easily violated in nature, are
needed for the formulation of inclusive fitness theory. If these assumptions do not hold,
then inclusive fitness either cannot be defined or does not give the right criterion for
what is favoured by natural selection.
The final stage was the theory of reciprocal altruism or mutualism, which obviously avoided the
problems of kin selection by the incorporation of game theoretic aspects - especially the repeated
trials of prisoner’s dilemma. The most famous and simplest win-win strategy for players seems
to be tit-for-tat so far (Axelrod, 1984) which helps retain long term self-interests of individuals,
presuming they start with cooperation. It has many variations, but for reciprocal altruism to work,
it has a very stringent set of pre-requisites that need to be met. The repeatedly interactive species
need to have good memory systems, good recognition system for distinction between individuals,
and disincentive to cheat, to begin with (Stearns, 2009a). Even though its simplistic mappings
onto natural systems have been questioned (Clutton-Brock, 2009), this reciprocity does seem to fit
in some scenarios very well, especially in some higher animals, and it also gives good results in tit-
for-tat game theoretic strategies, but these higher animals are only a negligibly recent phenomenon
when seen on the geological scale.
Evolutionary transformations such as single cells to multi-cellular specialized colonies strictly
require cooperative coordination mechanisms for interaction between different cells (Moser et al.,
2009). Not surprisingly, most of these lower and basic level phenomena do not have any of the
strong pre-requisites for reciprocal altruism to take place, and yet cooperation is ascribed to them
in one game-theoretic form or another, frequently using gene-centric view.
106

A.1.2 Hypothesis 2: Cooperation is as real as Selfishness
Microbiologists such as Lynn Margulis pursue the second hypothesis: that cooperation is as real a
phenomenon as selfishness; and not only do living organisms possess both characteristics but it is
historically necessary that, under certain circumstances, the cooperative aspect overshadowed the
selfish part for life to have developed to the extent it has till today. Through her work she has
come to the conclusion that (non-game-theoretic) symbiosis is so important that it is responsible
for the creation of new species in certain scenarios. This conception of cooperation involves two
organisms coming into a long term relationship and in the process a new entity bigger than the
two participating individuals comes into being. Some examples from (Margulis and Sagan, 2002)
(page 91) include: coral reef animals and their algae, photosynthetic clams, sulfide-oxidizing, two-
meter-long tube worms, urbanized termites; and grass munching cows. Now an established term,
symbiogenesis is considered as an explanation for the evolution of eukaryotic cells from the primitive
prokaryotic cells (Margulis, 1981; Lake, 2009). Molecular biology has come to the conclusion that
life as we see it today, could not have evolved unless there were high levels of cooperative associations
amongst the basic elements of life at all key stages of evolutionary history (Taylor and Nowak, 2009;
Smith and Szathmary, 1997):
Evolution occurs in populations of reproducing individuals. Mutation, selection, and
cooperation can be seen as the three fundamental principles of evolution. Cooperation
is needed for evolution to construct new levels of organization. The origin of life, the
emergence of the first cell, the arrival of eucarya, the rise of multi-cellular organisms,
and the advent of human language are all based on cooperation. A higher level of orga-
nization emerges, whenever the competing units on the lower level begin to cooperate.
On another front, Systems biology and epigenetics hold responsible not the genes but the
interaction of information between the intra-cellular elements and the extra-cellular environment
as an equally important factor in the evolutionary process. (Noble, 2006, 2010), (Woese, 2004), and
(Shapiro, 1997) through their work prove that reducing the entire natural system to selfish gene-eye-
view does not work anymore. In fact, they say that there is no single level that explains everything.
107

Rather their biology is going towards probabilistic and dynamically interactive systems and this
is another blow to the explicitly genetic level kin selection, mutualism and other explanations of
cooperation in neo-Darwinian literature.
It seems reasonable at this point to emphasize that most of the jargon used in evolutionary
biology to explain the natural phenomenon of symbiotic relationships is an example of misplaced
concreteness in scientific language. The following terminologies become instantly meaningless for
species not belonging to the negligible class of “higher animals”: altruism, benefit, game payoff
matrix, selfish gene, inclusive fitness, mutualism, reciprocal altruism, etc. The relation of symbiotic
entities is one of association not “benefits” or “costs” or “cooperation” or “competition” in the
game-theoretic terms (Margulis and Sagan, 2002) (pages 15-18). It is evident that this ignorance
of metabolic and physical associations in the contemporary evolutionary analyses is a legacy of
Darwin’s own writings. (Caldwell, 1999) has done a histogram analysis of frequent words used in
The Origin of Species, and while we find hundreds of repetitions for words such as destruction, kill,
exterminate, individual, race, perfect, selection, species, by contrast the following terms are missing:
association, affiliation, cooperation, community, and symbiosis. It is time that these more natural
metaphors are developed for a revision of otherwise stagnant evolutionary analyses on cooperation
(Goldenfeld and Woese, 2007). It is also time that nature be accepted along with its multifaceted
mechanisms rather than enforcing upon it simplistic conveniences.
We conclude this section by highlighting another important difference between Darwin and
Peirce. Whereas Darwin’s theory requires a negation of an important part of Peirce’s theory (re-
ducing Peircean cooperation to selfishness), there is nothing in Peircean theory that requires a
negation of Darwinian principles. Darwin pursues the first hypothesis, in which only selfishness is
real, and cooperation emerged because individuals sought to exploit the benefits offered by simu-
lations of cooperation. On the other hand, Peirce pursues the second hypothesis, where relations
such as cooperation are taken to be as real as competition in the empirical reality. It is noticed
that Peirce does not present an anti-thesis of Darwin. He does not say that only cooperation is
real and competition emerged from it and is explainable and reducible to cooperative elements.
In the Peircean framework cooperation is an important element represented by his third agency.
108

In short, Darwin’s dyadic theory of evolution requires a rejection of Peirce’s theory. But Peirce’s
triadic theory embraces and corrects Darwin’s theory.
109

110

Bibliography
Akhtar, J., Awais, M. M., and Koshul, B. B. (2013). Putting Peirce’s Theory to the Test: Peircean
Evolutionary Algorithms. Transactions of the Charles S. Peirce Society: A Quarterly Journal in
American Philosophy, 49(2):203–237.
Amor, H. B. and Rettinger, A. (2005). Intelligent exploration for genetic algorithms: using self-
organizing maps in evolutionary computation. Proceedings of GECCO, pages 1531–1538.
Angeline, P. J. (1998). Evolutionary optimization versus particle swarm optimization: Philosophy
and performance differences. In Porto, V. W., Saravanan, N., Waagen, D., and Eiben, A. E.,
editors, Proc. Evol. Prog. VII, pages 601–610. Springer-Verlag, Berlin, Germany.
Axelrod, R. (1984). The Evolution of Cooperation. Basic Books, New York.
Back, T., Fogel, D. B., and Michalewicz, Z. (1997a). Handbook of Evolutionary Computation. CRC
Press.
Back, T., Hammel, U., and Schwefel, H. P. (1997b). Evolutionary computation: Comments on the
history and current state. IEEE Transactions on Evolutionary Computation, 1(1).
Balazs, K. and Koczy, L. T. (2012). Genetic and bacterial memetic programming approaches in
hierarchical-interpolative fuzzy system construction. In 2012 IEEE International Conference on
Fuzzy Systems (FUZZ-IEEE), pages 1–8.
Beyer, H. G. (1996). Toward a theory of evolution strategies: Self-adaptation. IEEE Transactions
on Evolutionary Computation, 3(3):311–347.
111

Beyer, H.-G., Schwefel, H.-P., and Wegener, I. (2002). How to analyse evolutionary algorithms.
Theor. Comput. Sci., 287(1):101–130.
Bohm, D. (1969). Some remarks on the notion of order. In Waddington, C. H., editor, Towards a
theoretical biology, pages 18–40. Sketches, Edinburgh, United Kingdom.
Bowler, P. J. (1983). The Eclipse of Darwinism: Anti-Darwinian Evolution Theories in the Decades
around 1900, page 14. Baltimore: The Johns Hopkins University Press.
Brem, R. B. and Kruglyak, L. (2005). The landscape of genetic complexity across 5,700 gene
expression traits in yeast. Proc Natl Acad Sci USA, 102:1572–1577.
Brown, L. G. (1992). A survey of image registration techniques. ACM Computing Survey, 24(4):325–
376.
Caldwell, D. E. (1999). Post-modern ecology - is the environment the organism? Environmental
Microbiology, 1(4):279–281.
Caraffini, F., Neri, F., Iacca, G., and Mol, A. (2013). Parallel memetic structures. Information
Sciences, 227:60–82.
Chen, S. and Smith, S. (1999). Putting the “genetics” back into genetic algorithms (reconsidering
the role of crossover in hybrid operators). In Wolfgang Banzhaf, C. R., editor, Foundations of
Genetic Algorithms 5. Morgan Kaufmann.
Chiu, S. L. (1994). Fuzzy model identification based on cluster estimation. Journal of Intelligent
and Fuzzy Systems, 2:267–278.
Clutton-Brock, T. (2009). Cooperation between non-kin in animal societies. Nature, 462:51–57.
Darwin, C. (1871). The Descent of Man: And Selection in Relation to Sex. Number v. 2 in The
Descent of Man: And Selection in Relation to Sex. John Murray.
Darwin, C. R. (1859). On the origin of species by means of natural selection, or the preservation of
favoured races in the struggle for life. London: John Murray, online repository: http://darwin-
online.org.uk/, 1 edition.
112

Darwin, C. R. (1872). On the origin of species by means of natural selection, or the preservation of
favoured races in the struggle for life. London: John Murray, online repository: http://darwin-
online.org.uk/, 6 edition.
Darwin, F. (1995). The life of Charles Darwin. Senate, London.
Dawkins, R. (1976). The selfish gene. Oxford University Press, New York.
De Jong, K. (2005). Genetic algorithms: a 30 year perspective. In Booker, L., Forrest, S., Mitchell,
M., and Riolo, R., editors, Perspectives on Adaptation in Natural and Artificial Systems, pages
11–31. Oxford University Press, USA.
De Jong, K. A. (1993). Genetic algorithms are NOT function optimizers. In Whitley, D. L., editor,
Foundations of Genetic Algorithms 2, pages 5–17. San Francisco, CA: Morgan Kaufmann.
Deb, K. (2001). Multi-objective optimization using evolutionary algorithms. John Wiley and Sons.
Deb, K., Pratap, A., Agarwal, S., and Meyarivan, T. (2002). A fast and elitist multiobjective
genetic algorithm: Nsga-ii. IEEE Transactions on Evolutionary Computation, 6(2).
Ehrenreich, I. M., Torabi, N., Jia, Y., Kent, J., Martis, S., Shapiro, J. A., Gresham, D., Caudy,
A. A., and Kruglyak, L. (2010). Dissection of genetically complex traits with extremely large
pools of yeast segregants. Nature, 464:1039–1042.
English, T. M. (1996). Evaluation of evolutionary and genetic optimizers: No free lunch. Evolu-
tionary Programming V, MIT Press, pages 163–169.
Fogel, D. (1994). An introduction to simulated evolutionary optimization. Neural Networks, IEEE
Transactions on, 5(1):3–14.
Fogel, D. and Chellapilla, K. (1998). Revisiting evolutionary programming. AeroSense’98:
Aerospace/Defense Sensing and Controls.
Friedrich, T., Oliveto, P. S., Sudholt, D., and Witt, C. (2008). Theoretical analysis of diversity
mechanisms for global exploration. Proceedings of the Genetic and Evolutionary Computation
Conference (GECCO), pages 945–952.
113

Fuhrmann, J., Rurainski, A., Lenhof, H.-P., and Neumann, D. (2010). A new lamarckian genetic
algorithm for flexible ligand-receptor docking. Journal of Computational Chemistry, 31:1911–
1918.
Futuyma, D. J. (1986). Evolutionary Biology. Sinauer Associates, Sunderland, Massachusetts.
Goldberg, D. (1989). Genetic algorithms in search, optimization, and machine learning. Addison-
Wesley Professional, Upper Saddle River,NJ, USA.
Goldberg, D. E. (1993). Making genetic algorithms fly: a lesson from the Wright brothers. Advanced
Technology for Developers. Annals of Mathematics and Artificial Intelligence, 2(5).
Goldenfeld, N. and Woese, C. (2007). Biology’s next revolution. Nature, 445:369.
Gould, S. (2002). The Structure of Evolutionary Theory. Belknap Press of Harvard University
Press.
Han, J., Kamber, M., and Pei, J. (2006). Data Mining: Concepts and Techniques. Morgan Kauf-
mann Publishers, 2nd edition.
Han, K. H. and Kim, J. H. (2004). Quantum-inspired evolutionary algorithms with a new termina-
tion criterion, h gate, and two-phase scheme. IEEE Transactions on Evolutionary Computation,
8(2):156–169.
Heisenberg, W. (2007). Physics and Philosophy: The Revolution in Modern Science. Harper
Perennial modern classics. HarperCollins.
Herrera, F. and Lozano, M. (2000). Gradual distributed real-coded genetic algorithms. IEEE
Transactions on Evolutionary Computation, 4(1).
Holland, J. H. (1975). Adaptation in Natural and Artificial Systems. University of Michigan Press,
Ann Arbor, MI, USA.
Holton, G. and Brush, S. (2001). Physics, the Human Adventure: From Copernicus to Einstein
and Beyond. Rutgers University Press.
114

Hu, J., Seo, K., Fan, Z., Rosenberg, R. C., and Goodman, E. D. (2003). Hemo: A sustainable
multi-objective evolutionary optimization framework. In Cant-Paz, E., Foster, J. A., Deb, K.,
Davis, L., Roy, R., O’Reilly, U.-M., Beyer, H.-G., Standish, R. K., Kendall, G., Wilson, S. W.,
Harman, M., Wegener, J., Dasgupta, D., Potter, M. A., Schultz, A. C., Dowsland, K. A., Jonoska,
N., and Miller, J. F., editors, GECCO, volume 2723 of Lecture Notes in Computer Science, pages
1029–1040. Springer.
Hu, J. J. and Goodman, E. (2002). The hierarchical fair competition (hfc) model for parallel
evolutionary algorithms. In Evolutionary Computation, 2002. CEC ’02. Proceedings of the 2002
Congress on, volume 1, pages 49–54.
Huxley, T. (2010). On the Reception of the ’Origin of Species’. Kessinger Publishing.
Hwang, S. F. and He, R. S. (2006). A hybrid real-parameter genetic algorithm for function opti-
mization. Advanced Engineering Informatics, 20(1):7–21.
Jablonka, E. and Lamb, M. J. (2005). Evolution in Four Dimensions: Genetic, Epigenetic, Behav-
ioral, and Symbolic Variation in the History of Life. Bradford Books, MIT Press.
Jablonka, E. and Lamb, M. J. (2008). The epigenome in evolution: beyond the modern synthesis.
VOGis Herald, 12:242–254.
Jaimes, A. L. and Coello, C. A. C. (2009). Applications of parallel platforms and models in
evolutionary multi-objective optimization. In Lewis, A., Mostaghim, S., and Randall, M., editors,
Biologically-Inspired Optimisation Methods, volume 210 of Studies in Computational Intelligence,
pages 23–49. Springer Berlin Heidelberg.
King, C. (Aug., 1877). Catastrophism and evolution. The American Naturalist, 11(8):449–470.
Koziel, S. and Michalewicz, Z. (1999). Evolutionary algorithms, homomorphous mappings, and
constrained parameter optimization. IEEE Transactions on Evolutionary Computation, 7.
Kubota, R., Horio, K., and Yamakawa, T. (2005). Reproduction strategy based on self-organizing
115

map for genetic algorithms. International Journal of Innovative Computing, Information and
Control, 1(4):595–607.
Lake, J. A. (2009). Evidence for an early prokaryotic endosymbiosis. Nature, 460(7258):967–971.
Leung, Y. W. and Wang, Y. P. (2001). An orthogonal genetic algorithm with quantization for
global numerical optimization. IEEE Transactions on Evolutionary Computation, 5(1):41–53.
Lozano, M., Herrera, F., and Cano, J. R. (2008). Replacement strategies to preserve useful diversity
in steady-state genetic algorithms. Information Sciences, 178(23):4421–4433.
Luger, G. F. (2002). Artificial Intelligence - Structures and Strategies for Complex Problem Solving.
Pearson Education Publishers.
Mahfoud, S. W. (1995). Niching Methods for Genetic Algorithms. PhD thesis, University of Illinois
at Urbana-Champaign.
Mantere, T. (2006). A min-max genetic algorithm with alternating multiple sorting for solving
constrained problems. Finnish Artificial Intelligence Society: FAIS, pages 61–57.
Margulis, L. (1981). Symbiosis in Cell Evolution. Freeman, San Francisco.
Margulis, L. and Sagan, D. (2002). Acquiring Genomes: The Theory of the Origins of the Species.
Basic Books.
Mayr, E. (1993). What was the evolutionary synthesis? Trends in Ecology and Evolution, 8:31.
Mayr, E. (2004). What Makes Biology Unique?: Considerations on the Autonomy of a Scientific
Discipline. Cambridge University Press.
Menand, L. (2001). The Metaphysical Club: A Story of Ideas in America. Farrar, Straus and
Giroux.
Mitchell, M. (1998). Chapter 1: Genetic algorithms: An overview. In An Introduction to Genetic
Algorithms, A Bradford book.
116

Mitchell, M. and Taylor, C. E. (1999). Evolutionary computation: An overview. Annual Review of
Ecology and Systematics, 30:593–616.
Monod, J. (1971). Chance and Necessity: An Essay on the Natural Philosophy of Modern Biology.
Alfred A. Knopf, New York.
Moscato, P. (1989). On evolution, search, optimization, genetic algorithms and martial arts: To-
wards memetic algorithms. Technical Report C3P Report 826, California Institute of Technology.
Moser, M., Legate, K. R., Zent, R., and Fassler, R. (2009). The tail of integrins, talin, and kindlins.
Science, 324(5929):895–899.
Nawa, N. E. and Furuhashi, T. (1998). A study on the effect of transfer of genes for the bacterial
evolutionary algorithm. In Jain, L. C. and Jain, R. K., editors, KES (3), pages 585–590. IEEE.
Nietzsche, F. (1974). The Gay Science: With a Prelude in Rhymes and an Appendix of Songs.
Knopf Doubleday Publishing Group.
Noble, D. (2006). The Music of Life: Biology Beyond Genes. Oxford University Press.
Noble, D. (2010). Biophysics and systems biology. Philosophical Transactions of The Royal Society,
368:1125–1139.
Noble, D. (2013). Physiology is rocking the foundations of evolutionary biology. Experimental
Physiology.
Nowak, M. A., Tarnita, C. E., and Wilson, E. O. (2010). The evolution of eusociality. Nature,
466(7310):1057–1062.
Ochs, P. W. (1993). Charles sanders peirce. In Griffin, D. R., Cobb, J. B., Ford, M. P., Gunter, P.
A. Y., and Ochs, P. W., editors, Founders of Constructive Postmodern Philosophy, pages 68,85.
State University of New York Press.
Peirce, C. S. (1931–1958). In Hartshorne, Weiss, P., and Burks, A. W., editors, The Collected
Papers of Charles Sanders Peirce, volume I–VIII. Cambridge, MA: Harvard University Press.
117

Pigliucci, M. and Muller, G. B. (2010). Evolution, the extended synthesis. Cambridge, Mass. London
: MIT Press.
Ray, T. and Liew, K. M. (2003). Society and civilization: An optimization algorithm based on the
simulation of social behavior. IEEE Transactions on Evolutionary Computation, 7(4).
Reeves, C. R. (2002). Genetic Algorithms - Principles and Perspectives: A Guide to GA Theory.
Secaucus, NJ, USA: Kluwer Academic Publishers.
Rose, M. R. and Oakley, T. H. (2007). The new biology: beyond the modern synthesis. Biology
Direct, 2(30).
Runarsson, T. P. and Yao, X. (2000). Stochastic ranking for constrained evolutionary optimization.
IEEE Transactions on Evolutionary Computation, 4(3):284–294.
Salomon, R. (1995). Reevaluating genetic algorithm performance under coordinate rotation of
benchmark functions - a survey of some theoretical and practical aspects of genetic algorithms.
BioSystems, 39:263–278.
Salomon, R. (1997). Derandomization of genetic algorithms. Proceedings of The Fifth Congress on
Intelligent Techniques and Soft Computing EUFIT’97, pages 645–649.
Sareni, B. and Krahenbuhl, L. (1998). Fitness sharing and niching methods revisited. IEEE
Transactions on Evolutionary Computation, 2(3):97–106.
Schwefel, H.-P. (1997). Challenges to and future developments of evolutionary algorithms. In Back,
T., Fogel, D. B., and Michalewicz, Z., editors, Handbook of Evolutionary Computation, chapter H.
CRC Press.
Segura, C., Segredo, E., and Leon, C. (2011). Parallel island-based multiobjectivised memetic
algorithms for a 2d packing problem. In Proceedings of the 13th annual conference on Genetic
and evolutionary computation, GECCO ’11, pages 1611–1618, New York, NY, USA. ACM.
Shapiro, J. (2011). Evolution: A View from the 21st Century. FT Press Science Series. FT Press
Science.
118

Shapiro, J. A. (1997). A third way: alternative to darwinism and creationism. Boston Review,
22(1):32–33.
Shapiro, J. A. (2005). A 21st century view of evolution: genome system architecture, repetitive
dna, and natural genetic engineering. Gene, 345:91–100.
Skolicki, Z. (2005). An analysis of island models in evolutionary computation. In Proceedings of
the 2005 workshops on Genetic and evolutionary computation, GECCO ’05, pages 386–389, New
York, NY, USA. ACM.
Smith, M. J. and Szathmary, E. (1997). The Major Transitions in Evolution. Section 1.5, New
York: Oxford University Press.
Smolin, L. (2013). Time Reborn: From the Crisis in Physics to the Future of the Universe. Houghton
Mifflin Harcourt.
Spencer, H. (1867). First Principles. London: Williams and Norgate, 2nd, part 2, ch. 18 edition.
Srinivasa, K. G., Venugopal, K. R., and Patnaik, L. M. (2007). A self-adaptive migration model
genetic algorithm for data mining applications. Information Sciences, 177(20):4295–4313.
Stearns, S. (2009a). Lecture 17: Key events in evolution. In Principles of Evolution, Ecology and
Behavior. YALE EEB 122, http://oyc.yale.edu/ecology-and-evolutionary-biology/principles-of-
evolution-ecology-and-behavior/content/sessions/lecture17.html.
Stearns, S. (2009b). Lecture 36: Selfishness and altruism. In Principles of Evolution, Ecology and
Behavior. YALE EEB 122, http://oyc.yale.edu/ecology-and-evolutionary-biology/principles-of-
evolution-ecology-and-behavior/content/sessions/lecture36.html.
Taylor, C. and Nowak, M. A. (2009). How to evolve cooperation, Games, Groups, and the Global
Good. New York: Springer.
Todes, D. (1987). Darwin’s malthusian metaphor and russian evolutionary thought, 1859-1917.
ISIS, 78(294):537–551.
119

West, S. A., Griffin, A. S., and Gardiner, A. (2007). Social semantics: altruism, cooperation,
mutualism, strong reciprocity and group selection. Journal of Evolutionary Biology, 20.
Weyl, H., Helmer, O., and Wilczek, F. (2009). Philosophy of Mathematics and Natural Science.
Princeton University Press.
Whitley, D. (1994). A genetic algorithm tutorial. Statistics and Computing, 4(2):65–85.
Woese, C. R. (2004). A new biology for a new century. Microbiology and Molecular Biology Reviews,
68(2):173–186.
Wolpert, D. H. and Macready, W. G. (1995). No free lunch theorems for search. Technical Report
SFI-TR-02-010, Santa Fe Institute.
Wolpert, D. H. and Macready, W. G. (2005). Coevolutionary free lunches. IEEE Transactions on
Evolutionary Computation, 9(6):721–735.
Yampolsky, L. Y. and Stoltzfus, A. (2001). Bias in the introduction of variation as an orienting
factor in evolution. Evolution and Development, 3(2):73–83.
Yao, X. and Liu, Y. (1997). Fast evolution strategies. In Angeline, P. J., Reynolds, R., McDonnell,
J., and Eberhart, R., editors, Evolutionary Programming VI, pages 151–161. Springer-Verlag,
Berlin, Germany.
Yuan, Q., Qian, F., and Du, W. (2010). A hybrid genetic algorithm with the baldwin effect.
Information Sciences, 180(5):640–652.
120