evolution of the axial system in craniates: morphology and function
TRANSCRIPT

REVIEW Open Access
Evolution of the axial system in craniates:morphology and function of the perivertebralmusculatureNadja Schilling
Abstract
The axial musculoskeletal system represents the plesiomorphic locomotor engine of the vertebrate body, playing acentral role in locomotion. In craniates, the evolution of the postcranial skeleton is characterized by two majortransformations. First, the axial skeleton became increasingly functionally and morphologically regionalized. Second,the axial-based locomotion plesiomorphic for craniates became progressively appendage-based with the evolutionof extremities in tetrapods. These changes, together with the transition to land, caused increased complexity in theplanes in which axial movements occur and moments act on the body and were accompanied by profoundchanges in axial muscle function. To increase our understanding of the evolutionary transformations of thestructure and function of the perivertebral musculature, this review integrates recent anatomical and physiologicaldata (e.g., muscle fiber types, activation patterns) with gross-anatomical and kinematic findings for pivotal craniatetaxa. This information is mapped onto a phylogenetic hypothesis to infer the putative character set of the lastcommon ancestor of the respective taxa and to conjecture patterns of locomotor and muscular evolution. Theincreasing anatomical and functional complexity in the muscular arrangement during craniate evolution isassociated with changes in fiber angulation and fiber-type distribution, i.e., increasing obliqueness in fiberorientation and segregation of fatigue-resistant fibers in deeper muscle regions. The loss of superficial fatigue-resistant fibers may be related to the profound gross anatomical reorganization of the axial musculature during thetetrapod evolution. The plesiomorphic function of the axial musculature -mobilization- is retained in all craniates.Along with the evolution of limbs and the subsequent transition to land, axial muscles additionally function toglobally stabilize the trunk against inertial and extrinsic limb muscle forces as well as gravitational forces.Associated with the evolution of sagittal mobility and a parasagittal limb posture, axial muscles in mammals alsostabilize the trunk against sagittal components of extrinsic limb muscle action as well as the inertia of the body’scenter of mass. Thus, the axial system is central to the static and dynamic control of the body posture in allcraniates and, in gnathostomes, additionally provides the foundation for the mechanical work of the appendicularsystem.
IntroductionThe axial musculoskeletal system represents the plesio-morphic propulsive engine of the vertebrate body andmaintains a central role in locomotion in all craniates.Considering its evolutionary antecedence to the appen-dicular system and its importance for locomotion, ourunderstanding of the axial system is surprisingly limitedcompared to our understanding of the limbs.
The evolution of the axial system is marked by pro-found changes in its morphology and function. Theincreasing differentiation of its muscular, neural, andskeletal elements is certainly partly responsible for thediversity of locomotor mechanics among craniates. Thearrangements of the axial musculature among verte-brates show at least as much diversity as any other mus-cle system. Understanding the adaptive value of thevarious muscular arrangements is an undertaking towhich this review attempts to contribute. To develop aplausible scenario of the evolutionary transitions in thestructure and function of the perivertebral musculature,
Correspondence: [email protected] of Systematic Zoology and Evolutionary Biology, Friedrich-Schiller-University Jena, Germany
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
© 2011 Schilling; licensee BioMed Central Ltd. This is an Open Access article distributed under the terms of the Creative CommonsAttribution License (http://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction inany medium, provided the original work is properly cited.

the functional, anatomical, and physiological charactersof representatives of pivotal taxa were mapped onto aphylogenetic hypothesis. Such an approach allows infer-ence of the most likely character set of the last commonancestor of the respective taxa as well as informed specu-lations concerning the patterns of locomotor and muscu-lar evolution. The function of a muscle can be deducedfrom morphological and physiological variables such asits topography, fiber architecture, fiber-type composition,in-vivo muscle strain and ex-vivo work loops. The inte-gration of these data with other physiological data suchas the muscle’s activity as well as with biomechanicaldata such as the associated locomotor kinematics allowsone to test functional hypotheses and to infer a muscle’spossible functions. Because only some of these variableshave been studied in axial muscles of a number of crani-ates, inference of the muscle function will be based on asubset of this ideally available information (i.e., muscletopography, fiber architecture, fiber-type composition,activation patterns, kinematics).Parts of the proposed scenario cannot be tested directly
because some kinds of information, such as data aboutsoft tissues, are either inadequately preserved in the fossilrecord or are missing altogether. An indirect method, the‘extant phylogenetic bracket’ often allows reconstructionof soft tissue characters of fossils [1]. Hypotheses arethereby formulated by evaluating osteological characterstates causally related with the tested characters in atleast the first two extant outgroups of the fossil taxon ofinterest [outgroup rule, [2]]. Regarding the axial system,simple inference from extant sister taxa fails in somecases because of the fundamental anatomical differencesamong the groups and the absence of the critical osteolo-gical traits in the respective sister taxa. Additionally, thedata available on soft tissue characters such as fiber com-position are currently too incomplete for many extantcraniates to allow a strict phylogenetic reconstruction ofthe evolution of their axial system. Assuming that thesame biomechanical laws operate now as have in thepast, the inferred intramuscular transformations thataccompanied gross-anatomical and functional changesduring craniate evolution were inferred from studyingspecies that resemble the hypothetical last commonancestor of the particular taxon of interest. For that rea-son, this review focuses on specific craniate taxa only.Groups highly derived in their postcranial anatomy andlocomotor style such as snakes, birds, or monotremeswere not included in the proposed scenario; although, ofcourse, they would be potentially interesting and relevantto some of the major themes discussed below.Axial muscles may serve a number of different locomo-
tor functions. They may produce movements of the axialskeleton that generate positive or negative external work(referred to as mobilization). They also may counteract,
control, or restrict movements that are either passivelyinduced by gravitational or inertial forces, actively pro-duced by antagonists, or transmitted to the trunk byextrinsic limb muscles, i.e. they stabilize the trunk. Suchstabilizing role may involve long periods of activation, forexample to ensure such as the structural linking of theskeletal elements (called tonic, local stabilization), but alsofaster, briefer muscle action for quick responses for exam-ple required to stabilize the trunk against rapid loading(dynamic, global stabilization). Accordingly, local stabili-zers can be expected to contain high proportions of fati-gue-resistant fibers and are likely in close proximity to thejoint they stabilize, while global stabilizers should containprimarily fast contracting fibers and be well effectivelypositioned relative to the axis of motion. Mobilization, forexample to produce body propulsion, may involve slow orfast fibers depending on locomotor speed. As is the casefor global stabilizers, mobilizers are expected to be wellsituated for the production of locomotor work. This classi-fication, first proposed as human-specific trait based ontheir back muscle topography and activity [3,4], wasadopted and further developed by research on other mam-malian species [e.g., [5,6]], and revealed as generallyapplicable to the trunk musculature of tetrapods [e.g., [7]].Although too strict categorization risks oversimplification,because muscles likely fulfill different functions during dif-ferent behaviors or even the course of one behavior, suchclassification of the perivertebral muscles into local andglobal stabilizers as well as global mobilizers has heuristicvalue and provides a framework for the formulation of tes-table hypotheses [8]. Because the evolution of the axialmuscle function and morphology is tightly linked to theevolution of the postcranial skeleton, a few relevantaspects of the evolutionary transformations in the postcra-nial skeleton will first be summarized before the evolutionof the perivertebral musculature is discussed.
Evolution of the postcranial skeletonThe evolution in the postcranial system in craniatesfrom the agnathan fish ancestors to mammals is charac-terized by two major transformations. First, the axialskeleton became more and more regionalized. Second,the ancestrally axial-based locomotion became increas-ingly appendage-based with the evolution of extremitiesand their reorganization within tetrapods. Both eventswere associated with fundamental changes in the bodyplanes in which movements occur and moments act onthe body. Furthermore, the moments acting on thetrunk changed substantially during tetrapod evolutionwith the transition to land.In petromyzontids, the axial skeleton consists of more
or less similar, arch shaped elements situated dorsally tothe notochord (arcualia) (Figure 1). In gnathostomefishes, the vertebral column is regionalized into trunk
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 2 of 19

Figure 1 Hypothesized evolutionary transformations of the morphology and function of the axial system in craniates. Data werecompiled from various sources (see text) and mapped onto a simplified phylogenetic hypothesis based on [71]. Character states plesiomorphicfor craniates are indicated by arrows. – Axial skeleton (rectangles): Notochordates (i.e., Cephalochordata + Craniata) ancestrally possess anotochord, eponymous for the group. In early vertebrates, cranio-caudally uniform vertebral elements evolved (VE). In gnathostomes, the axialskeleton is regionalized. A trunk (= dorsal, D) and tail region (caudal, CD) are distinguished in gnathostome fishes, while a cervical (C), truncal,sacral (S), and caudal region are present in early tetrapods. In mammals, the truncal region is further subdivided into a thoracic (T) and a lumbar(L) region. – Axial musculature (circles): Gross anatomy and fiber orientation: Transformations in the arrangement of the perivertebralmusculature are illustrated by schematic cross-sections showing the gross-anatomical changes (left) and cartoons of a few body segments in
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 3 of 19

and tail by the presence of ribs and large neural andhemal spines, whereas cervical, truncal, sacral, and tailregions are distinguished in tetrapods. In mammals, thetruncal series was further subdivided into a thoracic anda lumbar region, resulting in altogether five morphologi-cally and functionally different divisions of the vertebralcolumn (Figure 1).Subdivisions of the axial skeleton allow particular body
regions to be morphologically and physiologically spe-cialized for certain functions such as body propulsion.For example, the primary function of the tail in gnathos-tome fishes is propelling the body by lateral undulations[e.g., carangiform swimming, [9]] and therefore it mustallow lateral flexion but resist longitudinal compression.In adaptation to this locomotor function, the tail regionhas no ribs and large hemal arches to provide attach-ment sites and leverage for the axial muscles. This cra-nio-caudal regionalization of the body is augmented bysoft tissue traits such as differences in fiber population[10,11], fiber contractile properties [12,13] or thearrangement of the connective tissue [14-16]. Thereduction of the role of trunk bending in locomotion incarangiform swimmers compared to anguilliform loco-motion, as for example in agnathans, may reduce pres-sure peaks in the body cavity, and thereby interferencewith inner organ function, but first and foremost itreduces the internal work of locomotion because onlypart of the body undergoes bending. Similarly, the for-mation of functional regions of the mammalian trunkfacilitated specialization of the vertebral series. Thethoracic region allows movements in the horizontal andtransverse planes, reflected by more or less horizontallyoriented zygapophyses, and the presence of ribs formingthe rib cage provides rigidity for the thorax to ensure
lung function [17-20]. In contrast, intense motions inthe sagittal plane are facilitated in the rib-free lumbarregion due to vertical zygapophyses [19-21].In contrast to the primarily axial-based locomotion of
aquatic craniates, body propulsion results from inte-grated action of trunk and limbs in tetrapods. Therefore,in addition to the plesiomorphic function of contribut-ing to the work of locomotion, the body axis providesthe foundation for the production of mechanical workby the limbs, and thus is central to the static anddynamic control of body posture and the integration ofcoordinated actions of the limbs in all tetrapods. In thelineage leading from the hypothesized ancestor of tetra-pods to therian mammals, body propulsion becameincreasingly dependent on limb action. In salamandersand lizards, the fore- and hindlimbs are composed ofthree serially homologous elements that functionroughly in the same manner regarding their range ofexcursion and positioning during locomotion [22]. Theevolutionary transformation from the ancestral (tetra-pod) sprawled limb posture to the derived parasagittalposition in therian mammals entailed a dissociation ofserial and functional homologues [23,24]. With thereduction of the coracoid, the scapula lost its rigid con-nection to the trunk in therian mammals and gainedmobility unique among tetrapods. In the hindlimb, theproximal part of the autopodium was elongated to forma new functional segment. As a result, the typical ther-ian limb consists of three functionally equivalent ele-ments plus a contact segment [i.e., scapula-femur,humerus-shank, lower arm-metatarsus, hand-toes[23,25]]. Associated with the evolution of a parasagittallimb posture was a fundamental change in the momentsthat act on the trunk. While extrinsic pro- and retractor
lateral perspective illustrating the changes in muscle and/or fiber arrangement (right). Dorsal and ventral parts of the myomeres are innervatedby separate rami of the ventral root in agnathan fishes (light and dark brown). In each segment, muscle fibers span longitudinally betweenadjacent myosepta. In gnathostomes, the dorsal and ventral myomere parts are morphologically separated by the horizontal septum (pink)resulting in epaxial (ep) and hypaxial (hy) muscles. Likely associated with the evolutionarily new requirements to stabilize the body against long-axis torsion, deeper muscle fibers are obliquely oriented. In non-amniote tetrapods, the epaxial musculature retained its segmental organizationin contrast to the hypaxial musculature, which comprises the polysegmental subvertebral (sv) and the abdominal wall muscles (the latter are notshown here). The majority of the epaxial fibers connects adjacent myosepta longitudinally, while deeper fibers run at different angles. Inamniotes, the epaxial musculature is reorganized into three longitudinal and polysegmental muscle tracts (tr: transversospinal, lo: longissimus, ilc:iliocostalis). In mammals, the transversospinal muscle is subdivided into several entities forming the transversospinal system (trs). The mammalianventrovertebral musculature is strengthened by the psoas major (ps). – Axial muscle function (diamonds): The plesiomorphic function of theaxial musculature is to mobilize the body in the horizontal plane. The horizontal and torsional moments that result from the evolution of finsand a heterocercal tail, which tend to laterally bend the trunk and cause long-axis torsion, respectively, have to be counteracted by the axialmuscles in gnathostome fishes. In tetrapods, as a consequence of the evolution of supporting limbs and transition to land, the axial musclesadditionally function to globally stabilize the trunk against inertial and extrinsic limb muscle forces as well as against gravitational forces. Notethat the evolution of limbs preceded the transition to land. In tetrapods with a sprawled limb posture, extrinsic limb muscle forces in thehorizontal plane are relatively large. The greater agility and maneuverability as well as an increased importance of limb action for bodypropulsion, likely requires the axial muscles to dynamically stabilize the trunk to a greater extent in amniotes than in non-amniote tetrapods.Associated with the evolution of sagittal mobility and a parasagittal limb posture in mammals, the axial muscles additionally function to globallystabilize the trunk against sagittal components of extrinsic limb muscle action as well as against inertia. Furthermore, the axial musculaturemobilizes the trunk in the sagittal plane during asymmetrical gaits.
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 4 of 19

muscle activity can be expected to act primarily in thehorizontal plane and thus cause lateral bending in asprawled limb posture, swinging the legs back and forthin a parasagittal plane results in the limb pro- andretractors acting on the trunk in the sagittal plane andthus causing sagittal bending [26]. Furthermore, the lat-eral components of the propulsive forces, that tend tolaterally bend the trunk and exert rotational torque onthe girdles, are larger in an animal with a sprawled limbposture compared to one with parasagittal limb motion[27]vs. [28].
Evolution of axial muscle function andmorphologyAgnathansThe organization of the axial musculature into serialunits (i.e., myomeres) by a complex myoseptal system isplesiomorphic for craniates (Figures. 1, 2). Each myo-mere is composed of a superficial layer of tonic fibersand a central stack of twitch fibers, all fibers spanninglongitudinally between adjacent myosepta [29-35]. Thedorsal part of the myomere is innervated by a dorsalbranch of the ventral root, while the ventral portion isinnervated by a ventral branch [36]. These two myomereportions are innervated by different motoneurons [37],intermingled in the ventral portion of the gray matter ofthe spinal chord [38]. Each motoneuron innervates mus-cle fibers in two or three myomeres, resulting in con-tractions that extend beyond a given segment [36].Observations on swimming lampreys show a rhythmic,
alternating, and posteriorly propagating activation of theaxial musculature suitable for producing a travelingwave of lateral bending [39,40]. In both, hagfish andlampreys, the whole body is involved in the undulatorymovements with little longitudinal variation in eitherthe burst duration as a percentage of cycle duration orin the lateral displacement [40,41] (i.e., anguilliformswimming), which likely accounts for the anterior-pos-teriorly undifferentiated musculoskeletal system; thebody segments are a repetition of virtually identical sub-units. The generated force is primarily transmitted tothe notochord by the myoseptal system. The notochordoccupies a position near the neutral axis of lateral bend-ing and has been shown to 1) dominate the viscoelasticproperties of the body, 2) provide dynamic passive stabi-lity, and 3) act as a power amplifier in hagfish [42,43]. Ithas been suggested that the muscular system activelytunes the body’s stiffness in order to match its resonantfrequency to undulatory frequency during locomotion[42-44]. Particularly the superficial, tonic fibers are wellsuited to modulate the stiffness of the body over longperiods; possibly directly via the myoseptal system andindirectly via the skin, onto which the myosepta attach[45]. The parietal, tonic fibers could also be involved in
slow frequency swimming, as has been shown for var-ious gnathostome fishes (see below), but unfortunately,no separate recordings from the parietal vs. the centralfibers exist.Given the great similarities in myotome organization
between lancelets and agnathan craniates [29,38,46,47],the morphology and the function of the axial muscula-ture of agnathan craniates to 1) produce lateral bending,and thus to mobilize the trunk, and 2) to modulate thebody’s stiffness are most likely plesiomorphic for crani-ates (Figure 1).
Gnathostome fishesIn contrast to agnathan fishes and lancelets, a transverseseptum (Septum horizontale) separates the myomeresinto epaxial and hypaxial parts in gnathostomes, whichare innervated by separate rami of the ventral root ofthe spinal nerve. This general separation into epaxialand hypaxial muscles is retained in all gnathostomes,regardless of how profoundly the axial musculature wasreorganized in the different taxa. The traditional view ofepaxial and hypaxial muscles with their respective inner-vation is challenged however by the fact that dorsal andventral parts of the myomeres are also innervated byseparate rami in the hagfish [Peters, 1963, cited in [38]]and the lamprey [37]. Therefore, the horizontal septummorphologically separates two previously neurologicallydistinct units in gnathostomes [48]. Further, in actinop-terygian and lungfishes three rami emerging from theventral root innervate the dorsal, medial, and ventralparts of a myomere, respectively [48-50]. Most likelyassociated with that, the extreme dorsal and ventral por-tions show distinct activation patterns that are notnecessarily correlated with the activity of the centralfibers near the horizontal septum [51]. Nevertheless, thehorizontal septum represents the major transmitter ofmuscle force to the axial skeleton [52], and thereforerepresents an important locomotor adaptation apo-morphic for gnathostome fishes (Figure 1).Gnathostome fishes have complexly folded, W-shaped
myomeres [45,48,53], which are primarily composed oftwitch fibers. Tonic fibers are segregated superficiallyand laterally in a wedge-shaped area close to the hori-zontal septum (Figure 2), providing good leverage forthe production of lateral bending [e.g., chondrichthyans:[54,55]; actinopterygians: [47,56]; lungfish: [10]]. Consid-erable variation in the amount of tonic fibers and therelative proportion of tonic to twitch fibers may occuralong the body or interspecifically and depending onlifestyle [e.g., [10,57-59]], but the general arrangement isvery similar among gnathostome fishes. In chondrichth-yan fishes, one spinal nerve innervates muscle fibers intwo adjacent myomeres [38]. Similar to agnathans, theaxial muscles of gnathostome fishes are activated
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 5 of 19
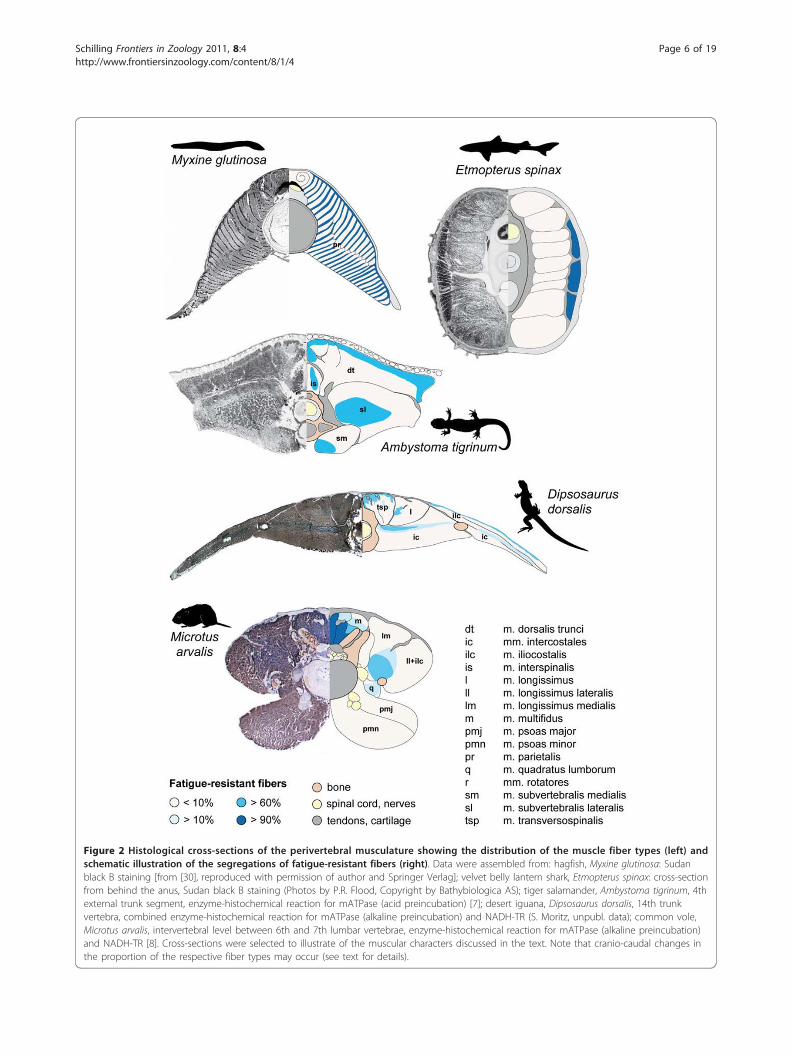
Figure 2 Histological cross-sections of the perivertebral musculature showing the distribution of the muscle fiber types (left) andschematic illustration of the segregations of fatigue-resistant fibers (right). Data were assembled from: hagfish, Myxine glutinosa: Sudanblack B staining [from [30], reproduced with permission of author and Springer Verlag]; velvet belly lantern shark, Etmopterus spinax: cross-sectionfrom behind the anus, Sudan black B staining (Photos by P.R. Flood, Copyright by Bathybiologica AS); tiger salamander, Ambystoma tigrinum, 4thexternal trunk segment, enzyme-histochemical reaction for mATPase (acid preincubation) [7]; desert iguana, Dipsosaurus dorsalis, 14th trunkvertebra, combined enzyme-histochemical reaction for mATPase (alkaline preincubation) and NADH-TR (S. Moritz, unpubl. data); common vole,Microtus arvalis, intervertebral level between 6th and 7th lumbar vertebrae, enzyme-histochemical reaction for mATPase (alkaline preincubation)and NADH-TR [8]. Cross-sections were selected to illustrate of the muscular characters discussed in the text. Note that cranio-caudal changes inthe proportion of the respective fiber types may occur (see text for details).
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 6 of 19

alternating and sequentially consistent with the produc-tion of a traveling wave of trunk bending [e.g., chon-drichthyan: [60]; actinopterygians: [59,61-63]; lungfish:[64]]. Red, tonic fibers are active during low-tailbeat-fre-quency, sustained swimming, while white, twitch fibersare additionally recruited during fast bursts and high-tailbeat-frequency swimming [e.g., [65-70]].The evolution of paired extremities increased the
maneuverability in gnathostomes [71]. The associatedextrinsic muscles apply forces to the body that inducetorsional and bending moments on the trunk. Becausemany gnathostome fishes are neutrally buoyant, primar-ily the horizontal (fore/aft) and the lateral componentsof the propulsive forces produced by the fins play a rolein locomotion. The horizontal components cause rota-tional torque on the girdles and thus lateral bending,requiring preferably longitudinal fiber orientation forstabilization, while the lateral components induce long-axis torsion and require an oblique fiber orientation[26,72]. Because early representatives of gnathostomessuch as placoderms typically had a heterocercal tail fin,additional torque about the long-axis of the body likelyresulted from tail beating. Compared to agnathan fishes,in which the muscle fibers are oriented longitudinally[29], the evolutionarily new requirements to stabilize thebody against long-axis torsion are reflected by the apo-morphic oblique fiber orientation found in mostgnathostome fishes. For example, the fibers are parallelto the long axis of the body in the superficial portion ofthe epaxial myomeres, while deeper fibers run at anglesbetween 10° and 35° relative to the body axis [73]. Inthe lateral hypaxial musculature, muscle fibers of thetwo oblique layers have opposing radial orientations[45], well suited to stabilize the body against long-axistorsion (Figure 1). In addition, oblique fiber orientationprovides an advantage for shortening velocity due to thegreater architectural gear ratio, that is, a greater short-ening distance resulting from fiber rotation as a conse-quence of the constant volume of the segment [74].Hence, the axial musculature of non-tetrapod gnathos-
tomes retained its plesiomorphic function of mobilizingthe body and producing locomotor work. Associatedwith the evolution of fins and a heterocercal tail, theaxial musculature also stabilizes the body against thelocomotor forces produced by the extrinsic fin musclesand torsional moments resulting from tail beating(Figure 1). These new functions are reflected by anoblique fiber orientation hypothesized to be apomorphicfor gnathostomes.
TetrapodsThe plesiomorphic segmental organization of the axialmusculature underwent stepwise reorganization duringthe evolution of tetrapods. In salamanders, the only
available postural model for early representatives of thetetrapods, the epaxial musculature retained its plesio-morphic segmental arrangement in contrast to thehypaxial muscles. The hypaxial musculature consists ofthe abdominal wall muscles and a subvertebral musclemass, which is associated with the ventral aspect of thevertebrae and ribs. Additional to the rectus system,the abdominal wall generally comprises three layers: theexternal and the internal oblique muscles as well asthe transversus muscle. The latter is an apomorphic fea-ture of tetrapods [53] and involved in ventilation [75].In most urodeles, the lateral hypaxial musculature issecondarily segmentally organized by tendinous inscrip-tions [76,77] and displays different fiber angles depend-ing on the layer [78,79]. Associated with the evolutionof polysegmental hypaxial muscles was likely a changein muscle fiber type distribution from a superficial posi-tion of fatigue-resistant fibers in fishes to a deep locali-zation in tetrapods such as salamanders [7]. As ingnathostome fishes and thus plesiomorphic for tetra-pods, the majority of the fibers connect adjacent myo-septa longitudinally; only deeper fibers associated withthe vertebrae run at different angles within the epaxialmyomeres [80-82]. The segregation of the muscle fibertypes in the epaxial musculature of urodeles resemblesthe pattern plesiomorphic for craniates [47,83]. That is,tonic and slow-twitch fibers are co-localized superfi-cially, while fast-twitch fibers form the bulk of the deepmuscle [7,84] (Figure 2). In the only two salamanderspecies for which data exist so far, this pattern is moreor less unchanged along the trunk [7].Similar to fishes, when salamanders swim, their main
epaxial and all hypaxial muscles are active synchro-nously and alternating. Activation propagates along thebody, consistent in timing with the production of a tra-veling wave of lateral undulation [85-90]. Thus, in sala-manders, most axial muscles mobilize the body duringswimming, i.e. their plesiomorphic function is retained.In accordance with its poor mechanical advantage fortrunk bending and high percentage of tonic red andtwitch intermediate muscle fibers [7], the biphasic activ-ity of the interspinalis muscle suggests that this musclefunctions in vertebral stabilization rather than lateralbending [90]. Active modulation of the body’s stiffnesswas suggested as one of the adaptations to swimming insalamanders [85], and the superficial segregation of fati-gue-resistant fibers in the dorsalis trunci muscle couldmodulate the body stiffness via the myoseptal systemand the skin [7]. Unfortunately, no study has investi-gated the recruitment patterns of the different fiberpopulations in this muscle, but the striking resemblanceof myomere organization to non-tetrapod craniatesinvites such speculation. Nevertheless, when salaman-ders swim, most of their axial muscles produce lateral
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 7 of 19

bending, some likely also modulate the body’s stiffness,and others provide local stabilization.The evolution of limbs predated the transition to land
as has been argued based on the analysis of early repre-sentatives of tetrapods such as Acanthostega [91] andmembers of the sister-group of tetrapods such as Tik-taalik [92]. Because aquatic stepping was likely the pri-mitive locomotor function of the tetrapod limb [93],trunk stabilization against locomotor forces produced byextrinsic limb muscles is evolutionarily older than stabi-lization against gravitational forces. Thus, the evolution-ary transition to land, basically a transition from high tolow viscosity and density and from low to high gravita-tional loads, was primarily associated with decreasedinertia and drag during the limb’s swing phase andincreased gravitational loading of the body resulting inincreased postural work for limb and trunk muscles[94]. Furthermore, the vertical components of the forcesproduced by the limbs, that are partially compensatedby buoyancy during aquatic stepping, induce long-axistorsion of the body during terrestrial stepping [26].A comparison of axial muscle activity during aquatic
and terrestrial stepping showed that muscle recruitment(i.e., intensity) increased in all trunk muscles, despitesimilar temporal patterns of muscle activation [90]. Thissuggests that the trunk is stiffened during terrestrial loco-motion, whereas the basic functions of the muscles areconserved across environments. Consistent with this, theperivertebral musculature contains an overall higher pro-portion of red tonic and intermediate twitch fibers in sal-amanders when compared to other sarcopterygians suchas lungfish. Comparisons of the fiber type composition invarious ecotypes, for example of predominantly terrestrialvs. aquatic species would allow testing this hypothesis. Inaddition, fatigue-resistant fibers are segregated in a cen-tral region of the lateral part and in ventral proximity tothe vertebral column in the medial part of the subverteb-ral muscle, allowing them to provide stability against tor-sion and sagging, respectively [7].During both aquatic and terrestrial stepping, body pro-
pulsion is achieved by concerted trunk and limb muscleaction in salamanders. Lateral bending was suggested tobe actively produced by the trunk muscles to facilitatethe placement of the feet, which serve as anchors andcontribute to stride length [95,96]. But lateral bendingmay also result passively from extrinsic limb muscleaction acting on the trunk via the limb girdles [97,98].Consistent with the production of a standing wave of lat-eral bending, uniphasic and cranio-caudally synchronizedactivity of the majority of the trunk muscles has beenobserved [85-90] (Figure 3). Additional bursts close tolimb girdles indicate that the dorsalis trunci muscle alsostabilizes the trunk against limb muscle action [88]. Thisadditional activity likely serves to dynamically stabilize
the trunk in the horizontal plane. Accordingly, the mus-cle primarily contains white twitch fibers [7,99], whichare arranged parallel to the long-axis of the body[80,81,100] and the fore/aft and lateral components ofextrinsic limb muscle action can be expected to begreater than the vertical ones given the sprawled limbposture. Consistent with their oblique fiber orientation[77,80], activity of the lateral hypaxial muscles resistslong-axis torsion [86,89,90]. The biphasic activity of thefatigue-resistant interspinalis muscle suggests that itfunctions as a local stabilizer during stepping, similar toits function during swimming [90].In summary, the axial musculature of basal tetrapods
such as salamanders mobilizes the trunk by producinglateral bending, modulates body stiffness (both putativeplesiomorphic) and provides local stability to ensure theintegrity of the axial skeleton during swimming (putativeapomorphic for tetrapods). During aquatic stepping, itadditionally resists extrinsic limb muscle forces causinglateral bending and long-axis torsion of the trunk; func-tions likely plesiomorphic for the group. During terres-trial locomotion, the axial musculature also stabilizesthe body against gravitational forces (Figure 1); an apo-morphic function for terrestrial tetrapods.
AmniotesA notable difference between anamniote and amniotetetrapods is the greater terrestrial agility in amniotes.Early amniotes were gracile, small animals with a snout-vent length of up to 24 cm [e.g., Paleothyris or Hylono-mus, [101]], and thus comparable to extant small lizardssuch as desert iguanas. Analyses of the axial skeletonand reconstructions of the associated musculature invarious fossils indicate great similarity between theseearly amniotes and generalized extant lizards and there-fore imply similar trunk motions [102,103]. Their dietand associated with that their lifestyle was presumablyalso similar to extant small lizards, i.e. mainly preyingon arthropods, mixed with some plant material[104,105]. Therefore, both burst and slow locomotionmust have constituted the locomotor repertoire of earlyamniotes. Associated with a higher aerobic capacity[106] and relatively higher body temperatures duringactivity [107], amniotes such as lizards are characterizedby greater swiftness and maneuverability compared toanamniote tetrapods such as salamanders. Swifter move-ments and increased performance are connected withfaster accelerations and decelerations of the limbs andthe center of mass of the body (CoM), and thus higherpeak loading of the limbs and trunk. Consequently,amniotes have an increased need for dynamic stabiliza-tion of the body compared to anamniote tetrapods.Similar to lissamphibians, amniotes such as lizards
exhibit a sprawling limb posture in which the feet are
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 8 of 19

positioned far laterally from the body axis. Compared toa parasagittal limb posture, a sprawling posture is asso-ciated with greater lateral components of the propulsiveforces see [[27]vs. [28]] and greater horizontal compo-nents of extrinsic limb muscle forces due to pro- and
retraction of the stylopods in the horizontal plane[22,26,108,109]. Both aspects result in moments that lat-erally bend the trunk. Compared to salamanders, limbaction can be expected to play a greater role in the pro-duction of locomotor work of lizards because of their
Figure 3 Activity patterns and hypothesized functions of the epaxial muscles in tetrapods during locomotion [modified from [118]].Data for the epaxial muscle activity were assembled from: spotted salamander, Ambystoma maculatum, m. dorsalis trunci, 8th external trunksegment, mean and standard error [90]; desert iguana, Dipsosaurus dorsalis, m. longissimus dorsi, 14th trunk vertebra, mean and standarddeviation (S. Moritz, unpubl. data); dog, Canis familiaris, m. longissimus thoracis et lumborum, 6th lumbar vertebra, median and upper and lowerquartiles [118]. The x-axis represents the stride cycle beginning with the touch down of the ipsilateral hindlimb. The footfall patterns of the bothhindlimbs are illustrated on the bottom of each graph (walk, trot: black: ipsilateral limb (iHL), gray: contralateral limb (cHL); gallop: black: trailinglimb (tHL), gray: leading limb (lHL). Note that for the galloping dog, the EMG trace associated with the trailing hindlimb is black, the oneassociated with the leading hindlimb is gray. Bending traces above the electromyograms indicate the unimodal lateral flexion and extension onthe body side ipsilateral to the recorded muscle activity (salamander, lizard) and the bimodal flexion and extension in the sagittal plane(mammal). Body planes in which moments and/or movements are suggested to occur are illustrated in the right top corner of each graph (fordetails see Figure 1). Note that the unilateral and monophasic epaxial activity in the walking salamander and lizard associated with the ipsilateralstance phase corresponds to the main activity observed in mammals. In mammals, the increased need for sagittal stability is met by bilateralactivity resulting from a second burst during ipsilateral swing phase.
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 9 of 19

relatively stronger limbs and greater limb excursions.Therefore, lateral bending in lizards may be a conse-quence of limb posture and limb muscle action, in addi-tion to being actively produced for example tocontribute to the production of locomotor work [72]and to facilitate limb positioning [109,110]. It ishypothesized that during the evolution of amniotes ashift in trunk muscle function occurred from primarilyproducing lateral bending (anamniote tetrapod mode) toincreasingly controlling and counteracting momentscaused by limb action and greater peak loading.In amniotes, the epaxial muscle mass was reorganized
into longitudinal and polysegmental tracts, forming thetransversospinal, the longissimus, and the iliocostalisgroups (Figure 2). The complexity and the arrangementof these tracts vary greatly among amniotes due to dif-ferentiation into smaller muscle units and/or variationsin their relative sizes [20,100]. The hypaxial musculatureshows a wide range of variation in ectothermic amniotessuch as lizards primarily due to splitting and delamina-tion of the main layers [111,112]. This anatomicallymore complex arrangement compared to other tetra-pods such as salamanders is likely partially related totheir enhanced locomotor performance but likely alsobecause axial muscles fulfill other functions such as ven-tilation in addition to their plesiomorphic role in loco-motion [113]. As in salamanders, the muscle fibers inthe various layers of the lateral hypaxial musculature areoriented obliquely at different angles [72,111]. In theepaxial musculature, the most medial tract shows anoblique fiber orientation in lizards, while the fibers inthe two lateral tracts are more or less parallel to thelong-axis of the body [100,114,115]. In contrast to ana-mniotes, in which the motoneuron pools of the epaxialand hypaxial muscles overlap in the medial column,motoneurons are spatially segregated in amniotes [116].Motoneurons innervating epaxial muscles are located inthe ventromedial portion of the ventral horn, while thehypaxial motoneurons reside dorsolaterally. Therefore,discrete pools serve individual muscles, resulting in atopographic map of motor pool organization that likelyfacilitates proper control of the anatomically and, moreimportantly, functionally diverse muscles originatingfrom the same myotome [38].It remains controversial whether or not epaxial and
hypaxial muscles are involved in the production or thecounteraction of lateral bending in lizards as they are insalamanders [117,118]. A functional division betweenepaxial and lateral hypaxial muscles was proposed as abasal feature of amniotes [117]; the former serving tostabilize the trunk against torsional forces [119], whilethe latter function to laterally bend the trunk and pro-vide stabilization against long-axis torsion [72]. For theepaxial muscles, Ritter concluded that they are not
involved in bending based on the timing of the activityas well as denervation experiments [117]. Several obser-vations question this hypothesis: 1) Recent recordingsfrom walking lizards do suggest that the timing of theactivity of the epaxial muscles is consistent with theproduction of lateral bending [120] (Figure 3) andthereby confirm previous recordings [110]. These recentdata imply speed dependency in the epaxial musclefunction, and thus may reconcile the controversy obser-vations [120]. 2) The denervation experiment, whichprovided the main evidence against lateral stabilization,was carried out around the mid-trunk, where the impactof the extrinsic limb muscles is likely to be small. Also,possible compensatory actions of other muscles such asthe hypaxial muscles were not tested. Furthermore, thetiming of epaxial muscle activity in lizards is similar tothat in salamanders and mammals, for which a stabiliz-ing function against lateral bending was shown, at leastnear the limb girdles, by simultaneous recordings ofextrinsic limb and back muscles [88,121]. 3) The impor-tance of lateral trunk bending, its production or coun-teraction, is reflected in the anatomy of the epaxialmuscles. The two lateral tracts, well positioned to actlaterally on the vertebral column, are relatively large inlizards [122], and their muscle fibers are oriented longi-tudinally, a fiber orientation well suited to laterallymobilize and stabilize the trunk [100,114,115]. Thus, amobilizing and/or stabilizing role in lateral bending can-not be ruled out for the epaxial muscles in lizards andfurther experiments, for example manipulating the loco-motor forces, are necessary to clarify the function of theepaxial muscles in lizards.In addition to the plesiomorphic side-to-side move-
ments, rotations about the long-axis of the body are animportant component of amniote locomotion and parti-cularly the transversospinalis muscle was thought toprovide torsional stabilization based on its activity[117,119] and the morphology of the neural spines[103]. Its oblique fiber orientation [100,114] is consistentwith a stabilizing function against long-axis torsion anddistinguishes amniote from anamniote tetrapods. Aspointed out above, compared to salamanders, extrinsiclimb muscle and inertial forces can be expected to begreater in lizards with their greater agility and locomo-tor speed. Therefore, torsional stabilization is addition-ally provided by the epaxial musculature of lizards [117],but solely accomplished by the lateral hypaxial musclesin salamanders [86,89].The evolutionary disintegration of the plesiomorphic
segmental organization of the epaxial musculature oftetrapods resulted in longitudinal, polysegmental muscletracts in amniotes and, likely more importantly, in anoverlapping muscle arrangement. Although this segmen-tal disintegration may be connected with a slightly
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 10 of 19

increased number of sarcomeres in series and thereby asmall increase in contraction speed, one advantage of apolysegmental over a segmental arrangement may bethat it allows for stabilization or mobilization of a wholeregion of the trunk by activating a single motor unit. Incontrast, simultaneous activation of several adjacent seg-ments is required in a myomeric organization to affect alarger body region (e.g., to produce a standing wave).Simultaneous action on a body region may be advanta-geous if the primary mode of trunk bending duringlocomotion is a standing wave, rather than a travelingwave, during which adjacent segments undergo lateralexcursion sequentially. On the other hand, an overlap-ping arrangement with attachment sites on each verte-bra also allows the production of a traveling wave, as forexample in snakes [123,124]. But, more importantly, thepossession of muscle fibers of different lengths orga-nized in an overlapping arrangement may increase theanimal’s maneuverability because it allows for activationand control of specific and varying body regions andthus for greater versatility. Associated with the reductionof the myoseptal system, the muscle fibers also actdirectly on the vertebrae in amniotes rather than indir-ectly via the myosepta. Direct muscle action on the ver-tebral column was associated with a greater degree ofvertebral structuring, i.e., relatively longer processes andlarger protuberances, which provide increased leverarms and attachment sites for the muscles [103,112]. Insummary, possibly greater contraction speed and dis-tance, more precise and selective activation and controlof a specific body region due to an overlapping musclearrangement, and improved muscle lever arms may havefacilitated more rapid mobilization and stabilization ofthe body and are likely connected with the greater agi-lity and versatility of amniotes.Preliminary results on the perivertebral musculature of
lizards indicate, when compared with results on mam-mals [8], similarities in the overall fiber type distributionamong these amniotes [125]. Fatigue-resistant fibers aresegregated in deeper muscle areas, close to tendons andbones, while the majority of the muscles comprises pri-marily fast twitch fibers (Figure 2). Consistent with thesuperficial and polysegmental muscles functioning inmobilization and global stabilization in lizards, they con-tain primarily fast-twitch glycolytic muscle fibers [125].To allow these polysegmental muscles to act on a givendivision of the vertebral column without causing verteb-ral dislocation, monosegmental muscle fibers arehypothesized to ensure spinal integrity. The demandsfor local stabilization can be expected to be greater inlizards compared to salamanders due to their greatertrunk loading and their polysegmental structure of bothepaxial and hypaxial muscles. Given their topographyand fatigue-resistant properties [125], local stabilization
is probably accomplished by the deeper fibers of thetransversospinalis muscle (Figure 2). Unfortunately, noEMG recordings exist of this muscle region to test thishypothesis. In contrast to anamniote tetrapods, in whichtonic and slow-twitch fibers are segregated superficially,likely to modulate the body stiffness via the myoseptalsystem and the skin, the fatigue-resistant fibers ofamniotes are regionalized in the depth of the musclesclose to the bones and intramuscular tendons inamniotes [8] (Figure 2). This intramuscular reorganiza-tion has been suggested to be related to the completeindependence from water [35]. Independence fromwater required changes in skin anatomy to reduce eva-poration and may have simultaneously decreased theskin’s ability to participate in force transmission.Furthermore, the intimate connection between the myo-septal system and the skin was dissolved with the evolu-tion of longitudinal muscle tracts. Thus, axial muscleforces are directly transmitted to the vertebrae inamniotes [112] and the body does not function as ahydrostatic system in body support as in anamniotes.Thus, the loss of the superficial fatigue-resistant fibersmay be associated with the substantial reorganization ofthe epaxial musculature and the high degree of amnioteterrestriality.In summary, the axial musculature of lizards appears
to fulfill similar functions as to those in salamanders,allowing tentative inference that these functions are ple-siomorphic for amniotes. But, compared to salamanders,the need for local and especially global stabilization ofthe trunk is increased in lizards due to their greater agi-lity and locomotor speed, and this need is reflected inthe detailed muscle morphology.
MammalsOne of the most striking apomorphic characteristics ofmammalian locomotion is sagittal bending [126-128].The ability to dorsoventrally flex and extend the bodyaxis enabled the evolution of asymmetrical gaits inmammals such as gallop or half-bound [19] [note theconvergent evolution of galloping in crocodilians[129-131]]. Several vertebral characteristics have beenproposed to be prerequisite for sagittal bending and,thus, to have predictive value for the trunk regioninvolved: 1) reduction of ribs in the posterior trunk andthus the formation of a lumbar region; 2) orientationand width of the spinous processes and thus the posi-tion of the anticlinal vertebra in the vertebral series; 3)orientation of the zygapophyseal facets and thus thelocation of the diaphragmatic vertebra(e) along the ver-tebral column [18-21,132]. A comparative analysis ofintervertebral movements in small therians during fastlocomotion showed however that these skeletal charac-ters were not simply related to the trunk region involved
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 11 of 19

in bending during locomotion, questioning their predic-tive value for the trunk region involved in sagittal bend-ing [133,134]. It has been suggested, therefore, thatbehaviors other than those directly related to locomo-tion may have driven the evolution of sagittal mobility,which was subsequently incorporated into the locomo-tor repertoire [133,135].During the evolution of mammals, extensive fusion
and reorganization of the epaxial tracts was associatedwith the reduction of the posterior ribs and the evolu-tion of a rib-free lumbar region. In many mammals, thetwo lateral tracts are inseparable in the lumbar regionand therefore referred to as the sacrospinalis muscle[100]. Associated with the evolution of asymmetricalgaits and the corresponding intense dorsoventral flexionof the posterior trunk were major reorganizations in themuscular system. For example, in mammals the size ofthe two medial epaxial tracts is increased compared tolizards [20]. Accordingly, both the multifidus and thelongissimus muscles exhibit recruitment patterns consis-tent with the mobilization of the axial skeleton in gal-loping mammals [118,136,137]. Additionally, comparedto non-mammalian amniotes, the subvertebral muscula-ture was strengthened in mammals and assists theabdominal wall muscles as an antagonist of the epaxialmusculature. Parts of the hindlimb musculature shiftedonto the trunk (i.e., puboischiofemoralis as iliopsoasmuscle) and an axial slip of the subvertebralis musclebecame independent as the psoas minor muscle[112,138,139]. Both act as hindlimb protractors and flex-ors of the vertebral column in mammals [140-143].These muscular changes in the epaxial and the hypaxialmusculature augmented the fast, glycolytic muscle massaround the vertebral column and therefore were likelyassociated with the evolution of vigorous sagittal spinemovements, i.e. the evolution of asymmetrical gaits [8].Consistent with the caudally increasing importance ofsagittal bending in body propulsion [133], the propor-tion of glycolytic muscle mass relative to the total anato-mical cross-sectional area of the axial musculatureincreases caudally [5,6,8].The increased mobility in the posterior trunk and its
vigorous mobilization during fast locomotor activitieswas hypothesized to be associated with an increasedneed for local stabilization [8]. The evolutionary subdivi-sion of the transversospinalis muscle into several mus-cular entities in mammals (i.e., the transversospinalsystem) is probably related to this greater demand forintervertebral stabilization because it was accompaniedby the functional specialization of its subunits [8]. Sev-eral deep, mono- and polysegmental muscles evolved (e.g., rotatores, intermammillares, mammilloaccessoriimuscles) and are predominantly composed of fatigue-resistant, slow fibers and thus well suited to provide
sustained intervertebral stability [5,6,144]. In contrast,the superficial, multisegmental division of the transver-sospinal complex (i.e., the multifidus muscle) contains ahigh proportion of fast fibers [e.g., [8,145,146]; Figure 2]that can mobilize as well as dynamically stabilize thetrunk [118,121,137,136].Another consequence of the greater mobility in all
body planes, particularly in the posterior trunk, is anincreased need for postural feedback. Mammals differfrom other amniotes in that they possess a central, slowregion in the in the lateral longissimus muscle, whichextends between the iliac blades and the 4th to 2nd pre-sacral vertebrae [8]. This region contains a large numberof muscles spindles [147,148] and is activated tonicallyand independently from the rest of the muscle belly[149,150]. Its responsiveness is modulated by the verteb-ral position [151]. It was suggested to function as a pro-prioceptive system monitoring the position of the pelvisrelative to the vertebral column [147,149]. Because nosuch region has been found in lizards (S. Moritz, pers.commun.) and salamanders [7], it is hypothesized torepresent an apomorphic character of mammals and tobe correlated with the evolution of a mobile lumbarregion [8].During the evolution of mammals, truncal motions in
the sagittal plane were added to the plesiomorphicmovements in the horizontal and transverse planes.Both, lateral bending and long-axis torsion occur duringsymmetrical gaits [e.g., [134,152-155]]. They are, how-ever, less pronounced in mammals than in other tetra-pods. The functional roles of the axial muscles duringsymmetrical gaits have been investigated in more detailin mammals than in any other tetrapod group, but stillseem poorly understood compared to the understandingof the limb musculature. Whereas the functional rolesof the lateral hypaxial muscles were clarified in a seriesof experiments [156-158], the function of the epaxialmuscles have become more clear only recently. Becausetheir activity was not directly correlated with the pro-duction of lateral bending or tilting, the epaxials weresuggested to stabilize the trunk [137,159-163]; thereby,only two studies tested the specific locomotor forcesand moments that may require stabilization [’sagittalrebound’, [164,165]]. Their primary function, at leastnear the hindlimb girdle, is to provide global stabiliza-tion against the vertical components of retractor mus-cles and the horizontal components of pro- andretractor muscles [121]. Furthermore, epaxial musclesprobably assist in the production of lateral bending dur-ing symmetrical gaits because the observed cranio-cau-dal activation patterns during walking and trottingaccord in timing with both the traveling and the stand-ing wave of trunk bending observed in these gaits,respectively [118]. Consistent with a function as
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 12 of 19

dynamic stabilizers as well as mobilizers, the largestepaxial muscles (i.e., the multifidus and the sacrospinalismuscles) consist predominantly of fast, glycolytic fibers[see [8] and references therein] (Figure 2).Compared to the sprawled limb posture of lower tet-
rapods, the parasagittal limb posture of mammals canbe expected to result in relatively smaller lateral butgreater sagittal components of the propulsive forces pro-duced by the limbs [97]. Furthermore, although the ver-tical moments acting on the trunk due to inertia aresimilar in lizards and mammals with the same size andlocomotor speed, they are most likely largely passivelystabilized in lizards by their horizontally oriented zyga-pophyseal facets, but would bend and extend the trunksagittally in mammals due to their more vertical facets.Both, the locomotor forces produced by the limbs andthe inertia of the body, result in an increased need fordynamic muscular stabilization in the sagittal plane.This increased need is reflected by changes in musclemorphology and function in mammals compared tolizards. For example, the two medial epaxials, best suitedto provide sagittal stability and mobility due to theirmore dorsal position relative to the neutral axis of thevertebral column, are increased in size in mammals [20].Furthermore, all epaxial muscles have a distinct obliquefascicle orientation [100], which allows for mobilizationand stabilization in all planes of the body simultaneouslyand thus better meets the complex needs for trunkmobility and stability in mammals. This oblique fiberorientation likely provides an advantage in the shorten-ing velocity of the entire muscle [74]. Furthermore, allmammals investigated so far display a biphasic andbilateral activity in their epaxial muscles during symme-trical gaits [137,159-166]. Of these two bursts duringeach locomotor cycle, only the main burst occurringduring ipsilateral hindlimb stance corresponds to theepaxial activity observed in other tetrapods (Figure 3),while the second burst, associated with the hindlimbswing phase, distinguishes mammals from other tetra-pods [118] and thus appears to be an apomorphic fea-ture of mammals. Based on recruitment symmetry (i.e.,bilateral activity) or asymmetry (i.e., unilateral activity)between both body sides a net extensor or net lateralbending/torsional moment can be inferred [167]. A netextensor moment is expected if sagittal forces dominate(e.g., due to the vertical oscillations of the CoM or verti-cal components of the extrinsic limb muscles), and themain function of the muscle is to stabilize the trunk inthe sagittal plane. The fact that mammals consistentlyshow biphasic, bilateral activity in their epaxial musclescorroborates the interpretation that there is an increasedneed for sagittal stability [118].Among amniotes, only birds and mammals are able to
locomote and ventilate their lungs at the same time
[113], except secondarily derived solutions for examplein varanid lizards [168]. In mammals, the evolution of adiaphragm freed most axial muscles from a ventilatoryfunction during locomotion [158,169]. Because the dia-phragm attaches to the posterior ribs, action of the dia-phragm results in anterior tilting of the ribs. To providea firm base for the action of the diaphragm, the ribsneed to be stabilized (e.g., pulled caudally). The abdom-inal wall muscles, namely the oblique muscles, are wellpositioned to retract the ribs and counteract rib protrac-tion. However, both the internal and the external obli-ques are locomotor muscles [158], stabilizing the trunkagainst sagittal shear during locomotion [157], andtherefore cannot provide costal stabilization. Especiallyduring asymmetrical gaits, inhalation is coupled withtrunk extension [133,170], thus the oblique abdominalwall muscles would have to stabilize the ribs duringsagittal extension. Such activity of the oblique hypaxialmuscles would cause sagittal flexion and thus interferewith the extension of the trunk. Rather, the obliqueabdominal wall muscles are in a good position to assistthe rectus abdominis muscle, which is the most impor-tant spinal flexor and active at the appropriate time[137]. EMG recordings of the external oblique muscle ingalloping dogs are consistent with such locomotor func-tion [Deban, Schilling, Carrier, unpubl. data]. However,neither during symmetrical nor during asymmetricalgaits can rib stabilization be provided by the abdominalwall muscles.Rib stabilization and possibly widening of the pleural
cavity during inhalation may be provided by the quadra-tus lumborum muscle based on its activation pattern ashas been shown in rabbits [171]. The homology of thismuscle has been subject of controversy [i.e., partiallysubvertebralis, [172], intercostalis system: intertransver-sarii muscles, [112], levatores costarum muscles, [173]].Its innervation either from the dorsal or the ventralrami [174] and the location of its motoneurons in theventromedial and the lateromedial motor pools [175]implies a mixed origin. However, its anatomical positionon the ventral aspects of the centra and insertion ontothe most posterior ribs allows the quadratus lumborummuscle to provide costal stabilization without interferingwith locomotor events. Its proximity to the vertebralcolumn gives it poor leverage for sagittal flexion andtherefore its contribution to sagittal bending can beexpected to be low. Consistent with its function in ribstabilization, the quadratus lumborum muscle showed astriking central accumulation of slow fatigue-resistantfibers, particularly in its anterior part in various therians[5,6,8,176]. This central region was hypothesized to actindependently from the rest of the muscle belly [8],similar to deep slow regions in anti-gravity muscles[177,178]. In accordance with a function in ventilation,
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 13 of 19

its muscle fiber type composition did not show the phy-siological adaptations found in other perivertebral mus-cles with changes in body shape [e.g., in ferrets [179]] orbody size [176].In summary, the evolution of sagittal mobility in
mammals added a new body plane, in which movementscan be produced but also have to be controlled andcounteracted. Thus, the axial muscles in mammalsmobilize the trunk in the sagittal plane (apomorphic formammals), in addition to their plesiomorphic role inbending and twisting (Figure 1). In the epaxial muscula-ture, the increased need for dynamic sagittal stabiliza-tion due to the parasagittal limb posture and the verticalzygapophyses was met 1) locally by the evolution ofnumerous deep, short, fatigue-resistant muscles and 2)globally by a biphasic activity of superficial, polysegmen-tal, fast muscles.
Concluding remarksIntramuscular (re)organization in craniatesMuscular properties such as the distribution of musclefiber types are primarily determined by a muscle’s func-tion and less by phylogeny. During the evolution ofcraniates, the composition and distribution of fiber typeschanged profoundly with a general tendency to segre-gate fatigue-resistant fibers in deeper muscle regions.Various factors have been discussed to account for acertain, ‘preferred’ location of a given fiber type within amuscle or a muscle group such as heat loss or thermalbalance [reviewed in [180]]. Briefly, it is argued thatbecause red muscle tissue has better circulation at restthan white one, a superficial position of red fibers wouldcause greater heat loss [181], assuming that the environ-ment is cooler than the animal. The thermal balanceargument is based on the observation that muscle fibersincrease their shortening speed and power as theybecome warmer, which would be advantageous for dee-ply located, more insulated fibers. The temperaturedependence of these characteristics is essentially similarbetween red and white fibers [e.g., [182-186]], and there-fore would support either fiber type distribution. Thecomparison of the intramuscular organization amongcraniates illustrates that red or white fibers may be clo-ser to the core of the body indicating that other factorsin addition to heat conservation are relevant to intra-muscular organization.Muscle fibers of different types are either segregated
from each other within a muscle or a muscle group orthey are intermingled (’salt-and-pepper pattern’). Gath-ering one fiber type may be advantageous because itunites similar metabolic needs, neural control, and bio-mechanical properties. For example, red and white fibersdiffer in their blood supply, in both the course and thebranching pattern of the capillary network [e.g.,
[187-189]] as well as in their capillary to fiber ratio [e.g.,[190-195]]. Whether the higher capillary content, andthus a relative higher collagen proportion per musclearea due to the vessel walls accounts for the differentbiomechanical properties reported for red and whitemuscle tissue [e.g., [189,196]] or differences in the con-nective tissue itself, for example in the structure of theendomysial collagen [197-200], is controversial, but agreater potential for elastic energy storage and a higherstiffness was found in red compared to white muscle tis-sue [199,201]. Thus, congregating fibers of similar meta-bolic needs may reduce the costs of the formation andmaintenance of the supply network and concentratingfibers with similar mechanical properties may reduceintramuscular shear [188,202]. Furthermore, segregationof a specific fiber type allows a muscle region to specia-lize for a specific function, because the properties of thevarious fiber types are optimized for different motortasks [203]. Thus, an accumulation of a specific fibertype indicates that this muscle or muscle region fulfillsfirst and foremost the same function in the same man-ner. In contrast, a mixed composition of a muscle ormuscle region places fibers with different contractileproperties in the biomechanically advantageous position.Such arrangement allows the muscle to fulfill the samefunction in different ways, i.e. by using different fibertypes and thus different motor units, for example toaccomplish the function with various force, speed, orfrequency [204,205].In addition, the reorganization of the myoseptal sys-
tem into polysegmental muscle tracts resulted in anarchitectural problem in amniotes. The evolution ofpolysegmental muscle tracts likely increased the impor-tance of local stabilization of the intervertebral joints toallow the polysegmental muscles to act on larger butvariable units of the vertebral column without causingintervertebral instabilities. To provide local stabilizationand prevent vertebral dislocation, short muscle bundlescontaining fatigue-resistant fibers and interlinking thevertebrae (i.e., monosegmental muscles) must be posi-tioned close to the vertebral column, while the polyseg-mental muscles are necessarily layered above. Thus,simple architectural constraints additionally influencemuscle-fiber-type distribution. Further research is neces-sary to increase our understanding of why muscle fibersof a given type are localized in particular muscle areasand how the observed patterns of muscular organizationevolved.
Methodological caveatsMuscle is one of the most plastic tissues, which allowsthe study of adaptations to changing functional demandson the one hand, but requires a thorough selection ofthe individuals and species studied on the other hand,
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 14 of 19

because interindividual or interspecific variability maymask the investigated traits. Hence, observed differencesmay represent phylogenetic divergence, functional diver-gence, and/or effects of environmental factors that dif-fered among the individuals studied (phenotypicplasticity). The species discussed herein were selectedbased on their resemblance (particularly body size andproportions as well as locomotor mode) to early repre-sentatives of higher taxa pivotal for the reconstructionof the evolution of the craniate axial system. The mus-cular differences observed among them are assumed tocorrelate with evolutionary changes in function andmorphology and that these differences are greater thaninter-individual variation.This approach bears several caveats limiting inference
of character states. 1) Depending on the fossil recordand the availability of extant species resembling earlyrepresentatives of a given group in the critical traits, theconclusions are better supported in some groups thanothers. For example, extant small mammals such asmice, rats, or tree-shrews highly resemble Mesozoicmammals such as Morganucodon in their postcranialanatomy [reviewed in [8]] and therefore are well-suitedto infer soft tissue characters for early mammals. Incontrast, salamanders differ in several essential postcra-nial characters from early representatives of tetrapodssuch as Acanthostega or Ichthyostega [91,206] such asthe reduction of ribs and the relatively small body size.However, salamanders are the only available posturalmodel for early tetrapods among extant taxa [207,208]and were therefore considered herein despite these post-cranial differences. 2) All species represent a mosaic ofplesiomorphic and apomorphic features [groundplan;[2]]. For example, extant agnathans resemble early crani-ates such as the conodonts in their myomeric organiza-tion of their axial muscles or the possession of anotochord as the main axial skeleton [69]. However,they are highly specialized relicts of a multifacetedgroup of jawless craniates that possessed for exampledermal armor to a varying extent [i.e., ostracoderms,[69]]. Therefore, inference of plesiomorphic axial musclecharacteristics for craniates is potentially confounded byderived character states in extant hagfish and lampreys.3) The depth to which we know intra-taxon variationand the confidence with which we can infer the set ofcharacter states in the common ancestor of the respec-tive groups varies greatly. Groups such as actinoptery-gians or mammals have been investigated intensively.Therefore, their interspecific variability and the adaptivevalue of the various muscular arrangements are fairlywell-understood. In such groups, we can start sortingout character states that represent phylogenetic historyfrom those that are more likely the immediate result ofadaptation. Only very few species have been studied so
far in other groups such as salamanders or lizards andthe ground-plan set of character states may not beunequivocal yet. Some caution is required when speciesfrom such groups are used to infer character states inancestors as the full extend of within-group muscularvariation has not been established yet. However, thisconsidered, inclusion of the currently known evidencein hypotheses as stated herein provides a clear frame-work for future hypothesis driven research with optionsfor falsification.
AcknowledgementsI am very grateful to M.S. Fischer for fostering my interest in vertebratefunctional morphology and thank him for numerous stimulating discussions,insightful criticism, and providing a productive environment. I am indebtedto D.R. Carrier, S.M. Deban, C. Anders, F. Goller, and M. Schmidt for valuablediscussions that were instrumental in developing many of the presentedideas. Much of this work has benefited from discussions with students andreflects their effort and enthusiasm for the axial system. Particularly, I thankS. Moritz, B. Hesse, and H. Stark for fruitful discussions. The technical supportof I. Weiß, M. Krüger, R. Petersohn, K. Felbel, E. Gretscher, and M. Roser isgreatly appreciated. Thanks to P.R. Flood and S. Moritz for providing imagesand data for Figures 2 and 3. Three reviewers provided constructive criticismand helpful suggestions, which greatly improved the manuscript.The many years of financial support by the Center of InterdisciplinaryPrevention of Diseases related to Professional Activities (KIP) founded andfunded by the Friedrich-Schiller-University, Jena and theBerufsgenossenschaft Nahrungsmittel und Gastgewerbe, Erfurt made thisresearch possible. Specifically, I thank R. Grieshaber, H.-C. Scholle, and I. Bradl.Financial support was furthermore provided by the German ResearchCouncil (DFG), the Erwin-Riesch-Foundation, the German Exchange Program(DAAD), and the Thuringian Ministry of Science, Research, and Art (all to NS).
Authors’ contributions and informationPart of this manuscript is based on the author’s Habilitation-Thesis. Thanks tothe organizers of the 103. Annual Meeting of the German Zoological Societyas well as to M. Nickel and C.S. Wirkner for their invitation to the symposiumand the opportunity to publish this review.
Competing interestsThe authors declare that they have no competing interests.
Received: 13 September 2010 Accepted: 10 February 2011Published: 10 February 2011
References1. Witmer LM: The Extant Phylogenetic Bracket and the importance of
reconstructing soft tissues in fossils. In Functional morphology in vertebratepaleontology. Edited by: Thomason, JJ. Cambridge University Press, NewYork; 1995:19-33.
2. Hennig W: Phylogenetic systematics. University of Illinois Press, Urbana, IL,USA; 1966, 1-280.
3. Bergmark A: Stability of the lumbar spine. Acta Ortho Scand 1989,230:1-54.
4. Mottram SL, Comerford MJ: Stability dysfunction and low back pain.J Orthop Med 1998, 20:13-18.
5. Schilling N: Characteristics of paravertebral muscles - fibre typedistribution pattern in Ochotona rufescens (Mammalia: Lagomorpha).J Zool Syst Evol Res 2005, 43:38-48.
6. Schilling N, Arnold D, Wagner H, Fischer MS: Evolutionary aspects andmuscular properties of the trunk - Implications for human low backpain. Pathophysiology 2005, 12:233-242.
7. Schilling N, Deban SM: Fiber-type distribution of the perivertebralmusculature in Ambystoma. J Morph 2010, 271:200-214.
8. Schilling N: Metabolic profile of the perivertebral muscles of smalltherian mammals: Implications for the evolution of the mammaliantrunk musculature. Zoology 2009, 112:279-304.
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 15 of 19

9. Breder CM: The locomotion of fishes. Zoologica 1926, 4:159-257.10. Dunn EA, Davison W, Maloiy GMO, Hochachka PW, Guppy M: An
ultrastructural and histochemical study of the axial musculature in theAfrican lungfish. Cell Tissue Res 1981, 220:599-609.
11. Gill HS, Weatherley AH, Lee R, Legere D: Histochemical characterization ofmyotomal muscle of five teleost species. J Fish Biol 1989, 34:375-386.
12. Altringham JD, Wardle CS, Smith CI: Myotomal muscle function atdifferent locations in the body of a swimming fish. J Exp Biol 1993,182:191-206.
13. Davies MF, Johnston IA, van de Wal JW: Muscle fibers in rostral andcaudal myotomes of the Atlantic Cod (Gadus morhua L.) have differentmechanical properties. Physiol Zool 1995, 68:673-697.
14. Willemse JJ: Functional anatomy of the myosepta in fishes. Proc Kon NedAkad Wet (C) 1966, 69:58-63.
15. Long JH, Nipper KS: The importance of body stiffness in undulatorypropulsion. Amer Zool 1996, 36:678-694.
16. Gemballa S: Myoseptenarchitektur und Rumpfmuskulatur derActinopterygii - Ein vergleichend-anatomischer Ansatz zum Verständnisder undulatorischen Lokomotion. Verh Ges Ichthyol 1998, 1:29-58.
17. Virchow H: Die Wirbelsäule des Löwen nach der Form zusammengestellt.SB Ges naturf Freunde Berlin 1907, 1907:1-43.
18. Fick R: Handbuch der Anatomie und Mechanik der Gelenke unterBerücksichtigung der bewegenden Muskeln. 3. Teil. Spezielle Gelenk-und Muskelmechanik. Gustav Fischer Verlag, Jena 1911, 1-688.
19. Rockwell H, Evans FG, Pheasant HC: The comparative morphology of thevertebrate spinal column. Its form as related to function. J Morph 1938,63:87-117.
20. Slijper EJ: Comparative biological-anatomical investigations on thevertebral column and spinal musculature of mammals. Verh Kon AkadWetenschappen Amsterdam 1946, 45:1-128.
21. Washburn SL, Buettner-Janusch J: The definition of thoracic and lumbarvertebrae. Am J Phys Anthrop 1952, 10:251-252.
22. Rewcastle SC: Stance and gait in tetrapods: an evolutionary scenario.Symp Zool Soc Lond 1981, 48:239-267.
23. Fischer MS: Crouched posture and high fulcrum, a principle in thelocomotion of small mammals: The example of the rock hyrax (Procaviacapensis) (Mammalia: Hyracoidea). J Hum Evol 1994, 26:501-524.
24. Schmidt M, Fischer MS: Morphological integration in mammalian limbproportions: Dissociation between function and development. Evolution2009, 63:749-766.
25. Fischer MS, Schilling N, Schmidt M, Haarhaus D, Witte HF: Basic limbkinematics of small therian mammals. J Exp Biol 2002, 205:1315-1338.
26. Gray J: Animal locomotion. Norton, New York; 1968, 1-479.27. Farley CT, Ko TC: Mechanics of locomotion in lizards. J Exp Biol 1997,
200:2177-2188.28. Budsberg SC, Verstraete MC, Soutas-Little RW: Force plate analysis of the
walking gait in healthy dogs. Am J Vet Res 1987, 48:915-918.29. Maurer F: Die Elemente der Rumpfmuskulatur bei Cyclostomen und
höheren Wirbeltieren. Morphol Jahrb 1894, 21:473.30. Flood PR, Storm Mathisen J: A third type of muscle fibre in the parietal
muscle of the Atlantic hagfish Myxine glutinosa. Z Zellforsch 1962,58:638-640.
31. Mellgren SI, Storm Mathisen J: Oxidative enzymes, glycogen, and lipid instriated muscle. A histochemical study in the Atlantic hagfish (Myxineglutinosa L.). Z Zellforsch 1966, 71:169-188.
32. Teräväinen H: Anatomical and physiological studies on muscles oflamprey. J Neurophysiol 1971, 34:954-973.
33. Teräväinen H, Rovainen CM: Fast and slow motoneurons to body muscleof the sea lamprey. J Neurophysiol 1971, 34:990-998.
34. Dahl HA, Nicolaysen KR: Actomyosin ATPase activity in Atlantic hagfishmuscles. Histochemie 1971, 28:205-210.
35. Lie HR: A quantitative identification of three muscle fiber types in thebody muscles of Lampetra fluviatilis and their relation to bloodcapillaries. Cell Tissue Res 1974, 154:109-119.
36. Peters S, Mackay B: The structure and innervation of the myotomes ofthe lamprey. J Anat 1961, 95:575-585.
37. Wallén P, Grillner S, Feldman JC, Bergelt S: Dorsal and ventral myotomemotoneurons and their input during fictive locomotion in the lamprey.J Neurosci 1985, 5:654-661.
38. Fetcho JR: A review of the organization and evolution of motoneuronsinnervating the axial musculature of vertebrates. Brain Res 1987, 434:243-280.
39. Grillner S, Wallen P, Dale N, Brodin L, Buchanan J, Hill R: Transmitters,membrane properties and network circuity in the control of locomotionin lamprey. TINS 1987, 10.
40. Williams TL, Grillner S, Smoljaninov VV, Wallen P, Kashin S, Rossignol S:Locomotion in lamprey and trout: the relative timing of activation andmovement. J Exp Biol 1989, 143:559-566.
41. Grillner S, Wallen P, Brodin L: Neuronal network generating locomotorbehavior in lamprey: circuity, transmitters, membrane properties, andsimulation. Annual Review in Neurosciences 1991, 14:169-199.
42. Long JH, Koob-Emunds M, Sinwell B, Koob TJ: The notochord of hagfishMyxine glutinosa: visco-elastic properties and mechanical functionsduring steady swimming. J Exp Biol 2002, 205:3819-3831.
43. Koob TJ, Long JH: The vertebrate body axis: Evolution and mechanicalfunction. Amer Zool 2000, 40:1-18.
44. Blight AR: The muscular control of vertebrate swimming movements. BiolRev 1977, 52:181-218.
45. Maurer F: Die ventrale Rumpfmuskulatur der Fische (Selachier, Ganoiden,Teleostier, Crossopterygier, Dipnoer). Jen Zeitschr 1913, 49:1-118.
46. Flood PR: Structure of the segmental trunk muscle in Amphioxus. ZZellforsch 1968, 84:389-416.
47. Bone Q: Evolutionary patterns of axial muscle systems in someinvertebrates and fish. Amer Zool 1989, 29:5-18.
48. Wikström DA: Ueber die Innervation und den Bau der Myomeren derRumpfmuskulatur einiger Fische. Anat Anz 1897, 13:401-408.
49. Marcus H: Über Myotome, Horizontalseptum und Rippen beiHypogeophis und Urodelen. Z Anat Entw Gesch 1937, 107:531-552.
50. Fetcho JR: The organization of the motoneurons innervating the axialmusculature of vertebrates. I. Goldfish (Carassius aurats) andmudpuppies (Necturus maculosus). J Comp Neur 1986, 249:521-550.
51. Jayne BC, Lauder GV: Are muscle fibers within fish myotomes activatedsynchronously ? Patterns of recruitment within deep myomericmusculature during swimming in the Largemouth bass. J Exp Biol 1995,198:805-815.
52. Westneat MW, Hoese WH, Pell CA, Wainwright SA: The horizontal septum:mechanisms of force transfer in locomotion in scombrid fishes(Scombidae, Pericformes). J Morph 1993, 217:183-201.
53. Nishi S: Muskeln des Rumpfes.Edited by: Bolk L. Handbuch dervergleichenenden Anatomie der Wirbeltiere Urban 1938:5:351-446.
54. Kryvi H: Ultrastructure of the different fibre types in axial muscles of thesharks Etmopterus spinax and Galeus melastomus. Cell Tissue Res 1977,184:287-300.
55. Kryvi H, Totland GK: Fibre types in locomotory muscles of thecartilaginous fish Chimaera monstrosa. J Fish Biol 1978, 12:257-265.
56. Johnston IA, Ward PS, Goldspink G: Studies on the swimming musculatureof the rainbow trout. I. Fibre types. J Fish Biol 1975, 7:451-458.
57. Boddeke R, Slijper EJ, van der Stelt A: Histological characteristics of thebody musculature of fishes in connection with their mode of life. Proc KNed Akad Wet Ser C Biol Med Sci 1959, 62:576-588.
58. Johnston IA, Patterson S, Ward P, Goldspink G: The histochemicaldemonstration of myofibrillar adenosine triphosphatase activity in fishmuscle. Can J Zool 1974, 52:871-877.
59. Coughlin DJ, Rome LC: Muscle activity in steady swimming scup,Stenotomus chrysops, varies with fiber type and body position. Biol Bull1999, 196:145-152.
60. Donley JM, Shadwick RE: Steady swimming muscle dynamics in theleopard shark Triakis semifasciata. J Exp Biol 2003, 206:1117-1126.
61. Jayne B, Lauder G: Red muscle motor patterns during steady swimmingin largemouth bass: effects of speed and correlations with axialkinematics. J Exp Biol 1995, 198:1575-1587.
62. Gillis GB: Neuromuscular control of anguilliform locomotion: patterns ofred and white muscle activity during swimming in the american eelAnguilla rostrata. J Exp Biol 1998, 201:3245-3256.
63. Altringham J, Ellerby D: Fish swimming: patterns in muscle function. J ExpBiol 1999, 202:3397-3403.
64. Horner AM, Jayne BC: The effects of viscosity on the axial motor patternand kinematics of the African lungfish (Protopterus annectens) duringlateral undulatory swimming. J Exp Biol 2008, 211:1612-1622.
65. Bone Q: On the function of the two types of myotomal muscle fibre inelasmobranch fish. J marine biol Ass 1966, 46:321-349.
66. Rayner MD, Keenan MJ: Role of red and white muscles in the swimmingof the skipjack tuna. Nature 1967, 214:392-393.
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 16 of 19

67. Johnston IA, Davison W, Goldspink G: Energy metabolism of carpswimming muscles. J Comp Physiol B 1977, 114:203-216.
68. Hudson RCL: On the function of the white muscles in teleosts atintermediate swimming speeds. J Exp Biol 1973, 58:509-522.
69. Ellerby DJ, Altringham JD, Williams T, Block BA: Slow muscle function ofPacific bonito (Sarda chiliensis) during steady swimming. J Exp Biol 2000,203:2001-2013.
70. Coughlin DJ: Aerobic muscle function during steady swimming in fish.Fish Fish 2002, 3:63-78.
71. Westheide W, Rieger R: Spezielle Zoologie. II. Wirbel- oder Schädeltiere.Spektrum Akademischer Verlag Heidelberg, Berlin 2009, 1-712.
72. Carrier DR: Activity of the hypaxial muscles during walking in the lizardIguana iguana. J Exp Biol 1990, 152:453-470.
73. Alexander RM: The orientation of muscle fibers in the myomeres offishes. J Mar Biol Ass UK 1969, 49:263-290.
74. Brainerd EL, Azizi E: Muscle fiber angle, segment bulging and architecturalgear ratio in segmented musculature. J Exp Biol 2005, 208:3249-3261.
75. Brainerd EL, Owerkowicz T: Functional morphology and evolution ofaspiration breathing in tetrapods. Respir Phys Neurobiol 2006, 154:73-88.
76. Maurer F: Der Aufbau und die Entwicklung der ventralenRumpfmuskulatur bei den urodelen Amphibien und deren Beziehungenzu den gleichen Muskeln der Selachier und Teleostier. Morphol Jahrb1892, 18:76-179.
77. Maurer F: Die ventrale Rumpfmuskulatur von Menobranchus, Menopomaund Amphiuma, verglichen mit den gleichen Muskeln anderer Urodelen.Jen Zeitschr 1911, 47:1-40.
78. Simons RS, Brainerd EL: Morphological variation of hypaxial musculaturein salamanders (Lissamphibia: Caudata). J Morph 1999, 241:153-164.
79. Brainerd EL, Simons RS: Morphology and function of lateral hypaxialmusculature in salamanders. Amer Zool 2000, 40:77-86.
80. Francis ETB: The anatomy of the salamander. Oxford University Press,London; 1934, 1-381.
81. Auffenberg W: The epaxial musculature of Siren, Amphiuma, and Necturus(Amphibia). Bull Florida State Mus 1959, 4:251-265.
82. Willemse JJ: Arrangements of connective tissue fibres and muscle fibresin the tail musculature of adult newts (Triturus cristatus, T. alpestris andT. vulgaris) (Amphibia, Urodela). Zoomorphology 1974, 77:255-269.
83. Flood PR, Kryvi H, Totland GK: Onto-phylogenetic aspects of muscle fibertypes in the segmental trunk muscles of lower chordates. Folia Morphol1977, 25:64-67.
84. Totland GK: Three muscle fibre types in the axial muscle of axolotl(Ambystoma mexicanum Shaw): a quantitative light- and electronmicroscopic study. Cell Tissue Res 1976, 168:65-78.
85. Frolich L, Biewener AA: Kinematic and electromyographic analysis of thefunctional role of the body axis during terrestrial and aquaticlocomotion in the salamander Ambystoma trigrinum. J Exp Biol 1992,162:107-130.
86. Carrier DR: Action of the hypaxial muscles during walking and swimmingin the salamander Dicamptodon ensatus. J Exp Biol 1993, 180:75-83.
87. D’Aout KD, Aerts P, De Vree F: The timing of muscle strain and activationduring steady swimming in a salamander, Ambystoma mexicanum. NethJ Zool 1996, 46:263-271.
88. Delvolve I, Bem T, Cabelguen JM: Epaxial and limb muscle activity duringswimming and terrestrial stepping in the adult newt, Pleurodeles waltl.J Neurophysiol 1997, 78:638-650.
89. Bennett WO, Simons RS, Brainerd EL: Twisting and Bending: Thefunctional role of salamander lateral hypaxial musculature duringlocomotion. J Exp Biol 2001, 204:1979-1989.
90. Deban SM, Schilling N: Activity of trunk muscles during aquatic andterrestrial locomotion in Ambystoma maculatum. J Exp Biol 2009,212:2949-2959.
91. Coates MI: The Devonian tetrapod Acanthostega gunnari Jarvik:postcranial anatomy, basal tetrapod interrelationships and patterns ofskeletal evolution. Trans R Soc Edinb Earth Sci 1996, 87:363-421.
92. Shubin NH, Daeschler EB, Jenkins FAJ: The pectoral fin of Tiktaalik roseaeand the origin of the tetrapod limb. Nature 2006, 440:764-771.
93. Clack JA: Gaining ground: the origin and evolution of tetrapods. IndianaUniversity Press 2002, 1-400.
94. Gillis GB, Biewener AA: Hindlimb muscle function in relation to speedand gait: in vivo patterns of strain and activation in a hip and kneeextensor of the rat (Rattus norvegicus). J Exp Biol 2001, 204:2717-2731.
95. Evans FG: The anatomy and function of the foreleg in salamanderlocomotion. Anat Rec 1946, 95:257-281.
96. Roos PJ: Lateral bending in newt locomotion. Proc Kon Ned Akad Wet (C)1964, 67:223-232.
97. Gray J: Studies on the mechanics of the tetrapod skeleton. J Exp Biol1944, 20:88-116.
98. Barclay OR: The mechanics of amphibian locomotion. J Exp Biol 1946,23:177-203.
99. Totland GK: Histological and histochemical studies of segmental musclein axolotl Ambystoma mexicanum. Norw J Zool 1976, 24:73-84.
100. Nishi S: Zur vergleichenden Anatomie der eigentlichen (genuinen)Rückenmuskeln. Gegenbaurs Morphol Jb 1916, 50:167-318.
101. Laurin M: The evolution of body size, Cope’s rule and the origin ofamniotes. Syst Biol 2004, 53:594-622.
102. Olson EC: The dorsal axial musculature of certain primitive Permiantetrapods. J Morph 1936, 59:265-311.
103. Sumida SS, Modesto S: A phylogenetic perspective on locomotorystrategies in early amniotes. Amer Zool 2001, 41:586-597.
104. Carroll RL: The ancestry of reptiles. Phil Trans Roy Soc Lond 1970,257:267-306.
105. Sues HD, Reisz RR: Origin and early evolution of herbivory in terrestrialtetrapods. TREE 1998, 18:141-145.
106. Janis CM, Keller JC: Modes of ventilation in early tetrapods: Costalaspiration as a key feature of amniotes. Acta Palaeontol Pol 2001,42:137-170.
107. Schmidt-Nielsen K: Animal physiology. Cambridge University Press,Cambridge; 1979, 1-560.
108. Brinkman D: The hind limb step cycle of Iguana and primitive reptiles.J Zool (Lond) 1981, 181:91-103.
109. Jenkins FAJ, Goslow GE: The functional anatomy of the shoulder of theSavannah monitor lizard (Varanus exanthematicus). J Morph 1983,175:195-216.
110. Reilly SM: Quantitative electromyography and muscle function of thehindlimb during quadrupedal running in the lizard Sceloporus clarkii.Zoology 1995, 98:263-277.
111. Maurer F: Die ventrale Rumpfmuskulatur einiger Reptilien. Festschrift zumsiebzigsten Geburtstage Carl Gegenbauers 1896, 1-252.
112. Starck D: Vergleichende Anatomie der Wirbeltiere aufevolutionsbiologischer Grundlage. Springer Verlag, Berlin, Heidelberg, NewYork 1978, 1-3.
113. Carrier DR: Conflict in the hypaxial musculo-skeletal system:documenting an evolutionary constraint. Amer Zool 1991, 31:644-654.
114. Tsuihiji T: Homologies of the transversospinalis muscles in the anteriorpresacral region of Sauria (crown Diapsida). J Morph 2005, 263:151-178.
115. Tsuihiji T: Homologies of the Longissimus, Iliocostalis, and hypaxialmuscles in the anterior presacral region of extant Diapsida. J Morph2007, 268:986-1020.
116. Fetcho JR, Reich NT: Axial motor organization in postmetamorphic tigersalamanders (Ambystoma tigrinum): a segregation of epaxial andhypaxial motor pools is not necessarily associated with terrestriallocomotion. Brain Behav Evol 1992, 39:219-228.
117. Ritter D: Epaxial muscle function during locomotion in a lizard (Varanussalvator) and the proposal of a key innovation in the vertebrate axialmusculoskeletal system. J Exp Biol 1995, 198:2477-2490.
118. Schilling N, Carrier DR: Function of the epaxial muscles in walking,trotting, and galloping dogs: Implications for the evolution of epaxialmuscle funtion in tetrapods. J Exp Biol 2010, 213:1490-1502.
119. Ritter D: Axial muscle function during lizard locomotion. J Exp Biol 1996,199:2499-2510.
120. Moritz S, Fischer MS, Schilling N: Aktivierung der epaxialenRumpfmuskulatur beim Wüstenleguan. In Kongressband 15 Erfurter Tage“Prävention von arbeitsbedingten Gesundheitsgefahren und Erkrankungen”(Verlag Bussert & Stadeler) Edited by: Grieshaber R, Stadler M, Scholle H-Ch2009, 493-497.
121. Schilling N, Carrier DR: Function of the epaxial muscles during trotting.J Exp Biol 2009, 212:1053-1063.
122. Vallois HV: Les transformations de la musculature de l’épisome chez lesvertébrés. Arch Morph Gén Exp 1922, 13:1-538.
123. Gans C: Terrestrial locomotion without limbs. Amer Zool 1962, 2:167-182.124. Gasc JP: Axial musculature.Edited by: Gans, C. Biology of the Reptilia New
York, Academic Press; 1981:11:355-435.
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 17 of 19

125. Moritz S, Fischer MS, Schilling N: Gemeinsamkeiten und Unterschiede inder Fasertypenverteilung bei Echsen und Säugetieren. In Kongressband16 Erfurter Tage “Prävention von arbeitsbedingten Gesundheitsgefahren undErkrankungen” (Verlag Bussert & Stadeler) Edited by: Grieshaber R, Stadler M,Scholle H-Ch 2010, 431-434.
126. Howell AB: Speed in animals. Univ Chicago Press, Chicago 1944, 1-270.127. Hildebrand M: Motions of the running cheetah and horse. J Mamm 1959,
40:481-495.128. Gambaryan PP: How mammals run. John Wiley and Sons, New York 1974,
1-367.129. Cott HB: Scientific results of an inquiry into the ecology and economic
status of the Nile crocodile. Trans Zool Soc Lond 1961, 29:211-356.130. Zug GR: Crocodilian galloping: a unique gait for reptiles. Copeia 1974,
1974:550-552.131. Webb GJW, Gans C: Galloping in Crocodylus johnstoni - a reflection of
terrestrial activity ? Rec Aust Mus 1982, 34:607-618.132. Virchow H: Die Wirbelsäule des Löwen nach der Form zusammengestellt.
SB Ges naturf Freunde Berlin 1907, 1907:43.133. Schilling N, Hackert R: Sagittal spine movements of small therian
mammals during asymmetrical gaits. J Exp Biol 2006, 209:3925-3939.134. Schilling N, Fischer MS: Kinematic analysis of treadmill locomotion of tree
shrews, Tupaia glis (Scandentia: Tupaiidae). Mamm Biol 1999, 64:129-153.135. Fischer MS: Die Lokomotion von Procavia capensis (Mammalia:
Hyracoidea): Zur Evolution des Bewegungssystems bei Säugetieren. AbhNaturwiss Verein 1998, 33:1-188.
136. Tokuriki M: Electromyographic and joint-mechanical studies inquadrupedal locomotion III. Gallop. Jap J vet Sci 1974, 36:121-132.
137. English AW: The functions of the lumbar spine during stepping in thecat. J Morph 1980, 165:55-66.
138. Howell AB: Morphogenesis of the architecture of hip and thigh. J Morph1938, 62:177-218.
139. Jones CL: The morphogenesis of the thigh of the mouse Mus musculuswith special reference to tetrapod muscle homologies. J Morph 1979,162:275-310.
140. Engberg I, Lundberg A: An electromyographic analysis of muscularactivity in the hindlimb of the cat during unrestrained locomotion. ActaPhysiol Scand 1969, 75:614-630.
141. Rasmussen SA, Chan AK, Goslow GEJ: The cat step cycle:electromyographic patterns for hindlimb muscles during posture andunrestrained locomotion. J Morph 1978, 155:253-270.
142. Carlson-Kuhta P, Trank TV, Smith JL: Forms of forward quadrupedallocomotion. II. A comparison of posture, hindlimb kinematics, and motorpatterns for upslope and level walking. J Neurophysiol 1998, 79:1687-1701.
143. Smith JL, Carlson-Kuhta P, Trank TV: Forms of forward quadrupedallocomotion. III. A comparison of posture, hindlimb kinematics, andmotor patterns for upslope and level walking. J Neurophysiol 1998,79:1702-1716.
144. Kojima R, Okada M: Distribution of muscle fibre types in thoracic andlumbar epaxial muscles of Japanese macaques (Macaca fuscata). FoliaPrimatol 1996, 66:38-43.
145. Yokoyama I: Analyses of the fibre composition of the lumbar backmuscles in mammals. Nippon Seikeigeka Gakkai Zasshi 1982, 56:579-594.
146. McFadden KD, Bagnall KM, Mahon M, Ford DM: Histochemical fibercomposition of lumbar back muscles in the rabbit. Acta Anat 1984,120:146-150.
147. Carlson H: Histochemical fiber composition of lumbar back muscles inthe cat. Acta Physiol Scand 1978, 103:198-209.
148. Schwartz-Giblin S, Rosello L, Pfaff DW: A histochemical study of laterallongissimus muscle in rat. Exp Neurol 1983, 79:497-518.
149. Carlson H: Morphology and contraction properties of cat lumbar backmuscles. Acta Physiol Scand 1978, 103:180-197.
150. Carlson H: Observations on stretch reflexes in lumbar back muscles ofthe cat. Acta Physiol Scand 1978, 103:427-445.
151. Ge W, Long CR, Pickar JG: Vertebral position alters paraspinal musclespindle responsiveness in the feline spine: effect of positioning duration.J Physiol 2005, 569:655-665.
152. Jenkins FAJ, Camazine SM: Hip structure and locomotion in ambulatoryand cursorial carnivores. J Zool (Lond) 1977, 181:351-370.
153. Graaff vdKM, Harper J, Goslow GEJ: Analysis of posture and gait selectionduring locomotion in the striped skunk (Mephitis mephitis). J Mamm1982, 63:582-590.
154. Pridmore PA: Trunk movements during locomotion in the marsupialMonodelphis domestica (Didelphidae). J Morph 1992, 211:137-146.
155. Haussler KK, Bertram JE, Gellman K, Hermanson JW: Segmental in vivovertebral kinematics at the walk, trot and canter: a preliminary study.Equine Vet J Suppl 2001, 33:160-164.
156. Carrier DR: Ventilatory action of the hypaxial muscles of Iguana iguana: afunction of slow muscle. J Exp Biol 1989, 143:435-457.
157. Fife MM, Bailey CL, Lee DV, Carrier DR: Function of the oblique hypaxialmuscles in trotting dogs. J Exp Biol 2001, 204:2371-2381.
158. Deban SM, Carrier DR: Hypaxial muscle activity during running andbreathing in dogs. J Exp Biol 2002, 205:1953-1967.
159. Carlson H, Halbertsma J, Zomlefer M: Control of the trunk during walkingin the cat. Acta Physiol Scand 1979, 105:251-253.
160. Shapiro LJ, Jungers WL: Electromyography of back muscles duringquadrupedal and bipedal walking in primates. Am J Phys Anthrop 1994,93:491-504.
161. Licka TF, Peham C, Frey A: Electromyographic activity of the longissimusdorsi muscles in horses during trotting on treadmill. Am J Vet Res 2004,65:155-158.
162. Wada N, Akatani J, Miyajima N, Shimojo K, Kanda K: The role of vertebralcolumn muscles in level versus upslope treadmill walking - Anelectromyographic and kinematic study. Brain Res 2006, 1090:99-109.
163. Wada N, Miyajima N, Akatani J, Shimojo K, Kanda K: Electromyographicactivity of m. longissimus and the kinematics of the vertebral columnduring level and downslope treadmill walking in cats. Brain Res 2006,1103:140-144.
164. Ritter DA, Nassar PN, Fife MM, Carrier DR: Epaxial muscle function introtting dogs. J Exp Biol 2001, 204:3053-3064.
165. Robert C, Valette JP, Denoix JM: The effects of treadmill inclination andspeed on the activity of three trunk muscles in the trotting horse. EquineVet J 2001, 33:466-472.
166. Zomlefer M, Provencher J, Blanchette G, Rossignol S: Electromyographicstudy of lumbar back muscles during locomotion in acute highdecerebrate and low spinal cats. Brain Res 1984, 290:249-260.
167. Wakeling JM, Ritruechai P, Dalton S, Nankervis K: Segmental variation inthe activity and function of the equine longissimus dorsi muscle duringwalk and trot. Equine Comp Exerc Phys 2007, 4:95-103.
168. Owerkowicz T, Farmer CG, Hicks JW, Brainerd EL: Contribution of gularpumping to lung ventilation in monitor lizards. Science 1999,284:1661-1663.
169. Carrier DR: Function of the intercostal muscles in trotting dogs:Ventilation or locomotion? J Exp Biol 1996, 199:1455-1465.
170. Bramble DM, Carrier DR: Running and breathing in mammals. Science1983, 219:251-256.
171. Boyd W, Blincoe H, Hayner JC: Sequence of action of the diaphragm andquadratus lumborum during quite breathing. Anat Rec 1965, 151:579-582.
172. Kardong KV: Vertebrates: Comparative anatomy, function, evolution.McGraw-Hill Company, Boston;, 2 1998, 1-747.
173. Langenberg W, Jüschke S: Morphologie und Innervation der Mm.levatores costarum und ihre Beziehung zu den Mm. intertransversariilaterales lumborum an Mensch und Känguruh. Z Anat Entw Gesch 1970,130:255-264.
174. Bogduk N: Clinical anatomy of the lumbar spine and sacrum. ChurchillLivingstone, New York; 1997, 1-261.
175. Takahashi Y, Ohtori S, Takahashi K: Somatotopic organization of lumbarmuscle-innervating neurons in the ventral horn of the rat spinal cord.J Anat 2010, 216:489-495.
176. Hesse B, Fischer MS, Schilling N: Distribution pattern of muscle fibre typesin the perivertebral musculature of two different sized species of mice.Anat Rec 2010, 293:446-463.
177. Chanaud CM, Pratt CA, Loeb GE: Functionally complex muscles of the cathindlimb V. The roles of histochemical fiber-type regionalization andmechanical heterogeneity in differential muscle activation. Exp Brain Res1991, 85:300-313.
178. Scholle HC, Schumann NP, Biedermann FHW, Stegeman DF, Grassme R,Roeleveld K, Schilling N, Fischer MS: Spatiotemporal surface EMGcharacteristics from rat triceps brachii muscle during treadmilllocomotion indicate selective recruitment of functionally distinct muscleregions. Exp Brain Res 2001, 138:26-36.
179. Moritz S, Fischer MS, Schilling N: 3D-Fibre type distribution in theparavertebral muscles of the domestic ferret (Mustela putorius f. furo)
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 18 of 19

related to trunk movements during locomotion. Zoology 2007,110:197-211.
180. Kernell D: Muscle regionalization. Can J Appl Physiol 1998, 23:1-22.181. Loeb GE: Hard lessons in motor control from the mammalian spinal
cord. TINS 1987, 10:108-113.182. Hajdu S: Behaviour of frog and rat muscle at higher temperatures.
Enzymologia 1950, 14:187-193.183. Ranatunga KW, Wylie SR: Temperature dependent transitions in isometric
contractions of rat muscle. J Physiol Lond 1983, 339:87-95.184. Ranatunga KW: Temperature-dependence of shortening velocity and rate
of isometric tension development in rat skeletal muscles. J Physiol 1982,329:465-483.
185. Ranatunga KW: The force-velocity relation of rat fast- and slow-twitchmuscles examined at different temperatures. J Physiol Lond 1984,351:517-529.
186. Ranatunga KW: Endothermic force generation in fast and slowmammalian (rabbit) muscle fibers. Biophys J 1996, 71:1905-1913.
187. Lee JC: Vascular patterns in the red and white muscles of the rabbit.Anat Rec 1958, 132:597-611.
188. Asmussen G, Kiessling A: Charakterisierung von besonderenMuskelfasergruppen in der Skelettmuskulatur des Frosches durch ihreInnervation und ihre Gefäßversorgung. Acta Anat 1974, 90:226-242.
189. Andersen P, Kroese AJ: Capillary supply in soleus and gastrocnemiusmuscles of man. Pflügers Arch 1978, 375:245-249.
190. Lie HR: A quantitative identification of three muscle fiber types in thebody muscles of Lampetra fluviatilis and their relation to bloodcapillaries. Cell Tissue Res 1974, 154:109-119.
191. Plyley MJ, Groom AC: Geometrical distribution of capillaries inmammalian striated muscle. Am J Physiol 1975, 228:1376-1383.
192. Walesby NJ, Johnston IA: Fibre types in the locomotory muscles of anantarctic teleost, Notothenia rossii - A histochemical ultrastructural andbiochemical study. Cell Tissue Res 1980, 208:143-164.
193. Hoppeler H, Mathieu O, Weibel ER, Krauer R, Lindtstedt SL, Taylor CR:Design of the mammalian respiratory system VIII. Capillaries in skeletalmuscles. Respir Physiol 1981, 44:129-150.
194. Gleeson TT, Nicol CJM, Johnston IA: Capillarization, mitochondrialdensities, oxygen diffusion distances and innervation of red and whitemuscle of the lizard Dipsosaurus dorsalis. Cell Tissue Res 1984, 237:253-258.
195. Totland GK: Capillary distribution in the lateral muscle of axolotlAmbystoma mexicanum. Acta Zoologica 1984, 65:221-226.
196. Hudlicka O: Growth of capillaries in skeletal and cardiac muscle. Circ Res1982, 50:451-461.
197. Kovanen V, Suominen H, Heikkinen E: Connective tissue of “fast” and“slow” skeletal muscle in rats - effects of endurance training. Acta PhysiolScand 1980, 108:173-180.
198. Kovanen V, Suominen H, Heikkinen E: Collagen of slow twitch and fasttwitch muscle fibres in different types of rat skeletal muscle. Eur J ApplPhysiol 1984, 52:235-242.
199. Kovanen V, Suominen H, Heikkinen E: Mechanical properties of fast andslow skeletal muscle with special reference to collagen and endurancetraining. J Biomech 1984, 17:725-735.
200. Laurent GH, Sparrow MP, Bates PC, Millward DJ: Turnover of muscleprotein in the fowl. Collagen content and turnover in cardiac andskeletal muscles of the adult fowl and the changes during stretch-induced growth. Biochem J 1978, 176:419-427.
201. Wells JB: Comparison of mechanical properties between slow and fastmammalian muscles. J Physiol 1965, 178:252-269.
202. Gorb SN, Fischer MS: Three-dimensional analysis of the arrangement andlength distribution of fascicles in the triceps muscle of Galea musteloides(Rodentia, Caviomorpha). Zoomorphology 2000, 120:91-97.
203. Rome LC, Funke RP, Alexander RM, Lutz GJ, Aldridge H, Scott F,Freadman M: Why animals have different muscle fibre types. Nature 1988,335:824-827.
204. Henneman E, Olson CB: Relations between structure and function in thedesign of skeletal muscles. J Neurophysiol 1965, 28:581-598.
205. Desmedt JE, Godaux E: Fast motor units are not preferentially activatedin rapid voluntary contractions in man. Nature 1977, 267:717-719.
206. Ahlberg PE, Clack JA, Blom H: The axial skeleton of the Devoniantetrapod Ichthyostega. Nature 2005, 437:137-140.
207. Edwards JL: Two perspectives on the evolution of the tetrapod limb.Amer Zool 1989, 29:235-254.
208. Carroll RL, Irwin J, Green DM: Thermal physiology and the origin ofterrestriality in vertebrates. Zool J Linn Soc 2005, 143:345-358.
doi:10.1186/1742-9994-8-4Cite this article as: Schilling: Evolution of the axial system in craniates:morphology and function of the perivertebral musculature. Frontiers inZoology 2011 8:4.
Submit your next manuscript to BioMed Centraland take full advantage of:
• Convenient online submission
• Thorough peer review
• No space constraints or color figure charges
• Immediate publication on acceptance
• Inclusion in PubMed, CAS, Scopus and Google Scholar
• Research which is freely available for redistribution
Submit your manuscript at www.biomedcentral.com/submit
Schilling Frontiers in Zoology 2011, 8:4http://www.frontiersinzoology.com/content/8/1/4
Page 19 of 19