evolution 1itc.gsw.edu › faculty › bcarter › isci › evolution1.pdfthe tumblebugs are my...
TRANSCRIPT

Evolution 1
“Evidence”
Unless otherwise noted* the artwork and photographs in this slide show are original and © by Burt Carter. Permission is granted to use them for non-commercial, non-profit educational purposes provided that credit is given for their origin. Permission is not granted for any commercial or for-profit use, including use at for-profit educational facilities. Other copyrighted material is used under the fair use clause of the copyright law of the United States. *Scanned images are from course textbook: Hewitt et al., Integrated Science and are used under fair use clause of copyright law. (“Hewitt” on images.)

The treatment of evolution in your textbook begins with the presumed driving mechanism (natural selection) and ends with the “evidence” for evolution. It is easy for me to see why it is approached this way. The authors, like most other textbook authors, begin with the assumption that there is a cohesive theory of evolution with natural selection at its base. this is not correct, and I think it is counter-productive in many ways to pretend it is. Much better, I think, is to recognize that hypothetical driving mechanisms are independent hypotheses from what they are supposed to drive. Remember that the quite reasonable hypothesis of continental drift, well founded in observation, suffered a great deal of criticism because the driving mechanism originally proposed for it was completely wrong. Similarly, rejecting the idea of evolution because natural selection is not up to the task of causing it (or, worse, because you fine it distasteful) is as nonsensical as rejecting the movements of continents because they can’t possible “flee the poles” by moving east or west. So, we will begin by examining the various observations upon which, since a long time before Darwin, the hypotheses that species have evolved have been based. Then we will see why Darwin’s version of the hypothesis was far better than any predecessor. Then we will end with his hypothetical driving mechanism and see some of the reasons I think it doesn’t work.

The discussion of “evidence” in your book begins on page 503. I will follow the order in the book, with just a couple of exceptions. The first two points (#’s 1 & 2) haven’t anything to do with whether evolution has occurred or not. They are, quite obviously, really about whether natural selection has happened or not. We will leave these two for later and start with #3. I will also add a couple of observations to the list, beginning with an obvious one the authors have left out.

0. Biodiversity. – These are millions of species on Earth. Estimates range from a couple of million to fifty or so million, but there are plenty of them. Why? It gets even more interesting. Lots and lots of these things do exactly, or almost exactly, the same thing. In North America alone there are a couple of hundred species of dung beetles. They either feed on or lay their eggs in (or both) manure. Flip over a cow patty and you’ll probably find more than one species of beetle feeding on it. The tumblebugs are my favorites. Individuals or mating pairs will chew off a piece of a patty, roll it into a ball, roll it for some distance – often quite a long distance for their size, then dig a hole to bury it in. Once it’s in the hole the female lays eggs in it before it’s covered. There are two different genera of tumblebugs in the US. The sacred scarabs of ancient Egypt did this too. Maybe they were considered sacred because their wide distribution of the fertility in animal dung helped the Egyptians grow food and thrive.

3. Similarities in body structures. – These are the homologies that we talked about in lecture. Remember that the definition of homology has changed over time. A. In the earliest versions it simply referred to the similarity of many things in terms of the component parts: vertebrate limbs, mammalian necks, and so on. Some homologies are very “deep” – your head is homologous to a fly’s head on a very basic level. We now understand a lot of the genetics of this process. B. By the middle 1800’s the underlying mechanical cause of the similarities was apparent. Homologous structures develop during growth from the same parts of an embryo in which those parts are not yet differentiated. The initial nub that would eventually grow into your arm would have been structurally identical to the nub that grew into a whale’s front flipper. How the DNA instructed the limb to develop is what is different, not the basic idea behind a hand or a flipper. C. After Darwin the idea was expanded to include a historical component. Structures in related species are homologous because they have evolved from a common ancestor who used the basic plan to do one or the other (or neither) of the things that it was modified to do later. We now understand that the modification(s) come about because of a change in the genetic instructions for growing the structure.

Hewitt

4. Vestigial organs. – Parts of an organism that are not functional. To the example of snake legs (actually only the pelvis is there) add that some living whales also have a pelvis, even though they do not have hind legs. Consider, however, this diagram which shows both a living and some fossil whales: (I would have ordered the pictures with the oldest fossil at the bottom. Why?)
In many cases the vestigial organ is known to be descended from a species in which it was not vestigial.
Hewitt

Displays at the Smithsonian in Washington, DC show the pelvic bones of both whales and snakes.
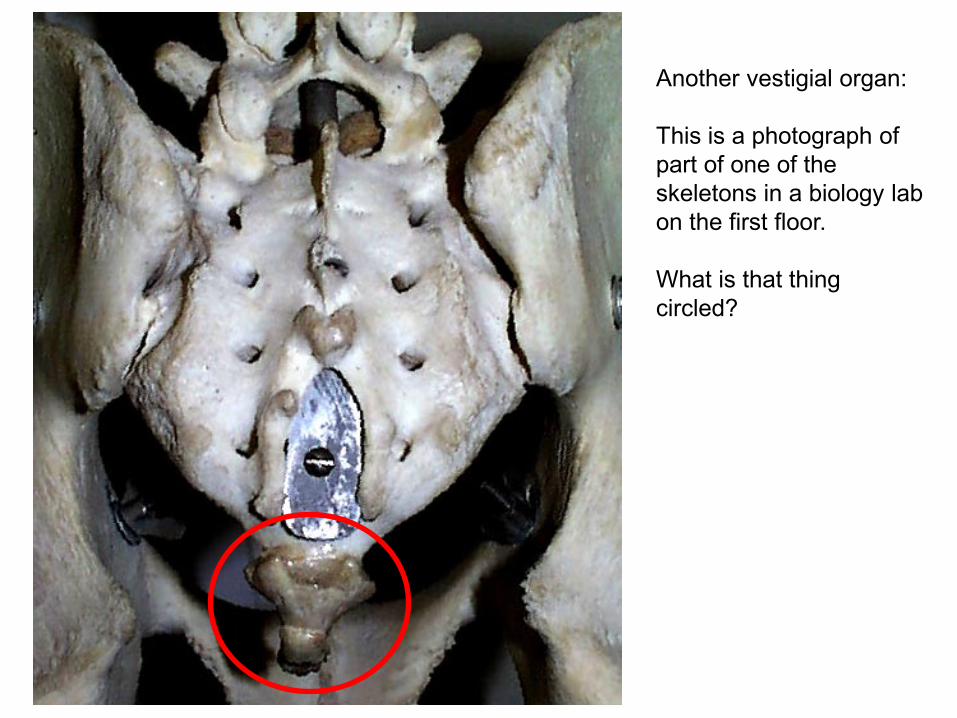
Another vestigial organ: This is a photograph of part of one of the skeletons in a biology lab on the first floor. What is that thing circled?

(4b) There is another category of homologies that are even more interesting. They are perfectly functional – not vestigial in that sense – but are vestigial in the way they function – the arrangement of parts in particular, is vestigial and, notably, often very bizarre. My favorite example, just because of how weird it is, is well covered in Richard Dawkins’ book “The Greatest Show on Earth”. Adult mammals don’t have gills (though embryonic mammals do – see a later slide) but they have larynxes (“voice boxes”). Sharks don’t have larynxes but they have gills. As it turns out, the structural supports for fish’s gills are homologous to a number of things in mammalian bodies, stretching from neck to chest. One of them is the larynx. When a lion roars, or when you whisper sweet nothings in someone’s ear, the roaring or the cooing are being done with fish gills. Or, at least, something that would have grown up to be fish gills rather than a voice box, had it been under the control of fish genes. The nerves that operate and sense activity in those various derived organs are not like most of the nerves in your body. Rather than branching from your nerve chord at the vertebrae, these come from the brain directly. They are called cranial nerves. There are two cranial nerves that enervate the larynx in mammals. One is the anterior laryngeal nerve and the other the recurrent laryngeal nerve. (Don’t memorize these names. The awesome observation about them that’s coming up soon is the point.)

First let’s see how the nerves and the gills work in a fish – a shark. The vagus nerve is common to all vertebrates – it is the principle cranial nerve that extends back into the body cavity. (It is not the nerve inside the spine.) As it passes the gill arches it branches to enervate them. Gills function to exchange gasses between the body and the surrounding water. This is how fish “breathe”. In gills the gasses simply pass across the cell membranes of blood vessels in the gills – oxygen in/CO2 out of the blood. The arrangement of nerves and blood vessels is very orderly and sensible here. Each gill gets a nerve and some blood vessels.
(from Dawkins, 2009, The Greatest Show on Earth, p. 361)

(from Dawkins, 2009, The Greatest Show on Earth, p. 361)
In an embryonic mammal, when the vestigial gills are still obvious, the arrangement is similar. However, as the mammal matures and the various parts are developed into their adult positions the orderliness of it goes all to hell in a handbasket. The anterior pharyngeal nerve makes sense, but what the recurrent one does is remarkable. It runs backward past the thing it is to enervate, loops around a blood vessel in the anterior part of the chest, and then comes back, running parallel to itself in order to get back to the pharynx – right beside where it started! In a human this is a matter of a few inches of detour, but in an adult giraffe the total detour can be 15 feet or more long.

As an engineering achievement the recurrent pharangeal nerve leaves a lot to be desired, as this circuit would in a new house.
But as a vestige of evolutionary history we can understand it’s quirkiness. Suppose that the homeowner had changed his mind 3 times about where he wanted the outlet? (Where would the junction boxes be on the diagram?)

Homeowner: “I’d like an outlet on a dedicated circuit just under the panel.” Done.

Homeowner: “I’ve changed my mind. I want the outlet at the right end of the wall, at desk level.” Done.

Homeowner: “Well, that looks silly. Move the outlet down to the ordinary level.” Done.

Homeowner: “No, that looks silly too. Put it back under the panel.” Done.

Your body is crawling with similar things – things that make no engineering sense whatsoever. When male testes descend from their position in the abdomen to the scrotum during development (pre-birth), the vas deferens, already quite long to connect to the penis, descends on the “wrong” side of the ureter connecting the kidney to the bladder – and then it elongates even more, roughly doubling its length. It does that because, like the recurrent pharyngeal nerve from the brain, it comes up out of the scrotum, loops over the ureter, and then goes back down parallel to itself to get back to where it needs to be – right beside where it originated. There cannot be an engineering explanation for this, there must be a historical one, relating to developmental changes during evolution. Numerous other nerves, blood vessels, and tubes of various sorts do similar things. The list of things awaiting you as you grow old that result from your upright stance is scary -- back troubles because gravity pulls on things that should be slung under your spine rather than pulling it obliquely forward; things that should have stresses oriented on them differently (your knees, female uteruses), or that have drain holes in the wrong place (your sinuses): it’s an illogical mess in there. It’s still a good life, but I wish I didn’t have to put up with all this nonsense. Four legged animals do not. (And neither would you if you were a quadruped.) And on, and on … The bizarre arrangements of these things are vestiges of former conditions, tweaked by evolution to work in some other context. As Dawkins says, “history is written all over us.”

5. DNA and other molecular similarities. – The pattern of nucleotides in the DNA of related species is very similar. The same can be said of the proteins coded for by that DNA, of course. And the more closely related two species are, the more similar their DNA
and proteins. Even those portions of the DNA that apparently do not code for proteins (introns, for example) are similar in close relatives. we carry around the
same baggage – useful and useless – that our cousins do.
Like vestigial structures and the “imperfect” adaptations we’ve just seen, these suggest historical development (with modification) from some common ancestor.

6. Development. – We have already alluded to this while talking about homologies, but the real story that development tells us about evolution lies in the obvious similarities of early developmental stages, even in distantly related organisms. In some cases the larvae of invertebrates that are virtually indistinguishable from each other develop into large clam-like animals on the one hand (brachiopods), and colonies of almost microscopic clones on the other (bryozoans). The head of an octopus, the crawling “foot” of a snail, and the burrowing appendage of a clam all derived from the same embryonic cells. And remember those gill slits that developed into your larynx. This picture from your book shows the general form of some vertebrate embryos. See if you can spot “your” tail.
(post birth) (functional external)

Is this destined to be an arm or a pig’s foot?
(Can you tell without looking back at the previous page?)
The great diversity of vertebrate limbs, from fish fins to pigs’ feet to human hands to Flipper’s flippers to bats’ wings – all develop along
different pathways from a thing that initially looks pretty much like this.

If you take this back far enough, every animal on the planet begins life structurally the same as a protist. Initially we were all a single diploid cell, created by fertilization. What we are now is just a matter of the instructions we got for the development and integration of those cells. Close relatives will generally have instructions that are more alike than they are similar to more distant relatives. My brothers look more alike than either of them looks like our second cousins (who we just saw at a family reunion). For the same reason, but writ a lot larger, we look more like chimps than we do like tigers.

7. Hierarchical organization of living things. – The taxonomic hierarchy, in other words. The obvious similarities among “related” things, like ‘cats’ (in the broad sense), or ‘dogs’, or ‘primates’,
or ‘reptiles’ all makes the best sense if they are truly related, by descent, as similar-looking humans are. I include a photograph from an earlier slide show to remind you of the idea
Photo by Yonnie Williams

8. Biogeography. – The patterns of geographic distribution of organisms, particularly those that are not able to move freely wherever they want like humans are, make good historical
sense. We mentioned in talking about plate tectonics that early Mesozoic fossils on the Gondwanaland continents are very similar. That is not the case in the present – they all have
highly distinct biotas now. Some good studies of molluscs that live on the continental shelf indicate that the distinctiveness began to appear soon after the continents rifted and has gotten
progressively more pronounced with time. The difference, in other words, has a history.
This was apparent to Europeans pretty soon after their navies and merchants began bringing back bizarre plants and animals from faraway places. Curious folks began to wonder how the oddness of places like Australia came about. How did all the kangaroo and wallaby species on
the planet know to hop off of Noah’s ark and hop eastward, not stopping until they got to Australia? (And the wombats would have had to waddle to Australia). And how did they get
from mainland Asia to Australia anyway?

9. Fossil Succession. (Not in the book.) – Remember from our discussion of geologic time that fossil species (and higher taxa as well) are found in an invariable vertical order. we
examined the pattern in a very broad sense (below), but remember that it applies at any level to any group of species.
CENOZOIC – (“whole life”) MESOZOIC – (“middle life”) PALEOZOIC – (“ancient life”)

Darwin’s hypothetical genealogy of organisms (his “family tree”) will help us understand all of these observations. Notice as we go that all of them are related to the agency of shared common ancestors.

0. Biodiversity.
These six species: are all descended from this one ancestor! (As are all the extinct species represented by the terminal branches between them. ~70 in all.)

3. Similarities in body structures. (homologies)
*
The descendants (red circles) of a single ancestor (red asterisk) will all have modified that ancestor’s basic parts into their own versions, to do whatever needs to be done for them to function as needed. Even more distant descendants (blue circles) of an ancestor (blue asterisk) will retain the same basic parts, but will have modified them much more extensively. *
Remember that some of the modifications might have left a structure either without a function (vestigial -- which certainly meets the definition of changed structure) or arranged in a bizarre manner (“poorly designed”).

A little modification of the DNA
5. DNA and other molecular similarities. The authors of your book should have left this one until after they mentioned the taxonomic hierarchy since they are two faces of the same thing. The DNA (and the products it codes for) are homologous in the genotypes, while the taxa whose growth and form are controlled by those various versions of DNA are homologous in the phenotypes. Remember the idea of “clumpiness”. It works in both the molecules and the structures they build.
A lot more modification of the DNA
Way more modification of the DNA

6. Development. Part 1. We’ll do two examples out of millions we could choose. (Remember that development is just the mechanism for generating homologous structures.)
Original fish-like vertebrate.
All its fish descendants go through similar developmental stages to those of the ancestor (orchestrated by their inherited DNA), but tweak it in their own ways.
All the other descendants – amphibians, reptiles, birds, mammals – also go through similar developmental stages, but tweaked in even more profound ways.
FISH TETRAPODS

6. Development. Part 2. How the cells arrange themselves in the early stages of development is one of the deepest homologies in animals. You did it the same way a sand dollar does. Snails do it the same way that spiders do.
Original animal.
Earliest cells (after two divisions)
spirally arranged.
Earliest cells (after two divisions) radially arranged.
Remember, we could do this over and over. Bird feathers and reptilian scales are homologous. We know because they develop similarly up to a point and then go different ways ……..

Species (letters)
Genera
Families
Orders
Classes
etc.
Taxa are simply recognized by the
homologies they’ve inherited from their
ancestors. The “deeper” the
common ancestor, the more distant the
relationship.
7. Hierarchical organization of living things.

8 – Biogeography. Continental scale: rifting occurs at the times indicated by red arrows (on left), splitting a population of some ancestral species. Descendant populations evolve differently in the separated continents (outlined).
B C A
B&C rift and 2 more biotas evolve (green and purple).
A rifts from B/C and 2 distinct biotas evolve (red and blue).
Island scale: a population is established in an island chain. As subpopulations colonize different islands in the chain they evolve new traits.

9. Fossil Succession. E
ach
time
inte
rval
has
a u
niqu
e, n
on-r
epea
ting
asse
mbl
age
of s
peci
es
The species in these 2 intervals …
… are m
ore closely related than the species in these 2 intervals

So Darwin’s brilliant idea of branching lineages, of a true family tree for all of life, explains all that we know about the patterns we see in and associated with species: why there are so many, why they are so amenable to being
grouped into taxa, why they occur where they occur (and not other places), why they seem to have changed over time …
None of Darwin’s predecessors had this insight.
There is no reason to doubt that species have evolved. The question is, how
did it happen?