evidence for tyrosine peptide homologies in the hla antigen system
TRANSCRIPT

Evidence for Tyrosine Peptide Homologies in the HLA Antigen SystemAuthor(s): Charlotte Cunningham-Rundles, Casper Jersild, Arne Svejgaard, Bo Dupont andRobert A. GoodSource: Proceedings of the National Academy of Sciences of the United States of America,Vol. 72, No. 12 (Dec., 1975), pp. 5081-5085Published by: National Academy of SciencesStable URL: http://www.jstor.org/stable/65351 .
Accessed: 03/05/2014 11:30
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
National Academy of Sciences is collaborating with JSTOR to digitize, preserve and extend access toProceedings of the National Academy of Sciences of the United States of America.
http://www.jstor.org
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions

Proc. Nat. Acad. Sci. USA Vol. 72, No. 12, pp. 5081-5085, December 1975 Immunology
Evidence for tyrosine peptide homologies in the HLA antigen system (radiolabeling/histocompatibility/surface antigens/evolutionary relationships)
CHARLOTTE CUNNINGHAM-RUNDLES*, CASPER JERSILDt, ARNE SVEJGAARDt, Bo DUPONT*, AND ROBERT A. GOOD*
* Memorial Sloan-Kettering Cancer Center, New York, N.Y. 10021; and t Tissue Typing Laboratory, University Hospital, Copenhagen, Denmark
Contributed by Robert A. Good, September 4, 1975
ABSTRACT The tetrameric HLA antigens are composed of two heavier chains which carry the alloantigenic determi- nants and two lighter chains identified as ,B2-microglobulin. Although at least 40 different antisera are required to define the varying HLA specificities, it appears that these antigens may be closely related to each other and to the immunoglob- ulins. Through the use of a new electrophoretic technique, which is able'to compare simultaneously the tyrosine pep- tides produced from radioiodinated cell surface proteins, this report gives evidence that HLA antigens of the three chromo- somal loci may have similar amino-acid sequences. Since the retention of homologous tyrosine residues and a tendency for sequence preservation surrounding these residues are fea- tures of immunoglobulin structure, this may indlicate that similarly conservative evolutionary mechanisms have been operative in the HLA allelelic proteins or that immunoglobu- lins and HLA antigens may indeed have a commnon evolu- tionary origin.
The serologically determined histocompatibility antigens are tetrameric structures composed of two polypeptide chains of molecular weight 45,000, and two light chains of molecular weight 11,500, now identified as 32-microglobulin (32-m) (1-4). The larger component carries the serologically identi- fied alloantigenic determinants (5). Although at least 40 dif- ferent antisera are required to define the varying specifici- ties of the HLA antigens, accumulating evidence from sev- eral sources has suggested that a rather limited degree of chemical heterogeneity may actually be present (6--8). This investigation was undertaken to compare HLA antigens of different specificities, using a newly devised method for the chemical analysis of microgram amounts of lactoperoxidase radioiodinated cell surface antigens. Since lactoperoxidase binds ['251]iodine to tyrosine residues (9), the electrophoretic technique described here permits direct comparison of ra- dioiodinated cell surface proteins, based upon the number and mobility of tyrosine peptides. In particular, this report presents evidence that antigenic products of the first, sec- ond, and third HLA chromosomal locit may have substantial homology of primary amino-acid sequence (10).
MATERIALS AND METHODS
lodination and Immune Complex Formation. Lympho- cytes (107) were isolated from heparinized blood of HLA typed normal donors using the Ficoll-Hypaque density gra- dient technique (11), and viability was determined by try- pan blue exclusion. Suspensions of lymphocytes were enzy-
Abbreviations: fl2-m, f32-microglobulin; NP-40, Nonidet P-40; Dnp-, dinitrophenyl-.
Sixth International Histocompatibility Conference nomenclature has been revised to HLA-A, HLA-B, and HLA-C antigens, re- spectively. These names will be used here.
matically iodinated in phosphate-buffered saline with 2-4 mCi of 1251 (New England Nuclear), lysed with Nonidet P-40 (NP-40), and cleared of IgG and TgM by immune com- plexing (12). HLA antigens were then precipitated by monospecific antisera and an excess of rabbit anti-human IgG (Pentex) (13, 14). Controls were AB sera obtained from normal male donors and were precipitated by rabbit anti- human IgG.
Antisera. Monospecific antisera used were: HLA-A3 (Menth), HLA-A9 (Kisch), HLA-B7 (94C Hansen or CLB- B), HLA-B14 (D1971/73), and HLA-CW3 (UPS). In addi- tion, antisera to HLA-BW35 (Wesa), which also crossreacts with HLA-BW15, was used on cells from an individual who does not have HLA-BW15.
High Voltage Electrophoresis. The radiolabeled HLA immune complexes were washed three times in phosphate- buffered saline, dissolved in 5 M urea-0.1 M mercaptoetha- nol, and incubated at 370 for 30 min. After dialysis against 5% formic acid overnight (using viscose dialysis tubing which retains proteins exceeding 10,000 daltons), the com- plexes were digested with pepsin (EC 3.4.23.1), 2% by weight (twice crystallized, Worthington) at 37? for 16 hr, and then dried under reduced pressure over NaOH. After reconstitution in 15% NH4HCO3, L-(l-tosylamido-2-phenyl) ethyl chloromethyl ketone trypsin (EC 3.4.21.4), 2% by weight (Worthington), was added, and digestion was contin- ued for 4 hr at 37?. The digested proteins were then dried under reduced pressure over P205. Radioactivity was deter- mined in a Packard Auto Gamma Spectrometer (model 5285). Samples were then dissolved in 0.1 ml of pH 3.5 pyri- dine-acetic acid-water (33:1:300, vol/vol) and applied to a 1-2 cm origin of a 46 X 57 cm sheet of Whatman no. 1 or no. 3 chromatography paper. In general, 10,000 to 15,000 cpm were applied per cm of origin. The peptides were then subjected to high voltage electrophoresis in the same buffer for 60 min at 3 kV in a Savant LT48-A high voltage electro- phoresis tank cooled by Varsol. Markers used were acid fuchsin, cyanol-blue, E-dinitrophenyl-lysine(E-Dnp-lysine), glycyl-alanine, and a mixture of acidic, basic, and neutral amino acids. After the paper was dried, autoradiographs were made using Kodak R/P medical x-ray film. Markers were developed by staining with cadmium ninhydrin stain (15).
In addition to HLA antigens, two samples of pure f2-m (kindly provided by Dr. M. Poulik and Dr. T. Plesner) were iodinated, reduced, alkylated, enzymatically digested, and electrophoresed in parallel to the HLA antigens. These io- dinated proteins were checked for purity on 0.1% sodium dodecyl sulfate-0.5 M urea-5% polyacrylamide gels and were found to present one sharp peak at a molecular weight corresponding to 11,500 (16).
5081
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions

5082 Immunology: Cunningham-Rundles et al. Proc. Nat. Acad. Sci. USA 72(1975)
Gby-Aa *
Val _
e-Dnp-Lys * _
Glu-~~~~~~~~~-
FIG. 1. An autoradiograph of a peptic-tryptic digest of ra- dioiodinated HLA antigens HLA-A9 and HLA-B14. Markers are included at the left. Free 1251 applied as a marker produces two bands, one at the position of E-Dnp-lysine and one at the position of Glu.
RESULTS
The resulting radioactive tyrosine peptides derived from HLA antigens of the first, second, and third HLA loci ap- pear on autoradiography as a number of vertically stacked, distinct narrow bands which can be readily compared from one sample to the next. It has been found practical to run up to eight samples on one sheet of paper. In general, samples run simultaneously are best compared.
Fig. 1 shows a representative autoradiograph obtained from lymphocyte antigens HLA-A9 and HLA-B14. The ra- dioactive peptides appear to have similar mobilities in each sample. In Fig. 2, antigens HLA-A3, HLA-B7, and HLA- CW3 were used. Similar peptide banding is present in all ex- periments. For Fig. 3, trypsin digestion of HLA antigens has been allowed to proceed for 12 hr instead of the usual 4 hr. The number of bands at the anode end is greatly diminished although two distinct, similar bands remain in each sample.
Gly-Ala P 0 _
c-Dnp-LysP
~iI GII.
FIG. 2. This is similar to Fig. 1 except that HLA-A3, HLA-B7, and HLA-CW3 are compared.
Evidently, HLA antigens are quite sensitive to trypsin, and the timing of this step is important if digests are to be com- pared from different experiments. In spite of the new pat- tern of radioactive peptides observed in this figure, the two digested antigens, HLA-A3 and HLA-B7, have retained very similar peptide mobilities. This observation reinforces the possibility of homology between HLA antigens of different specificity.
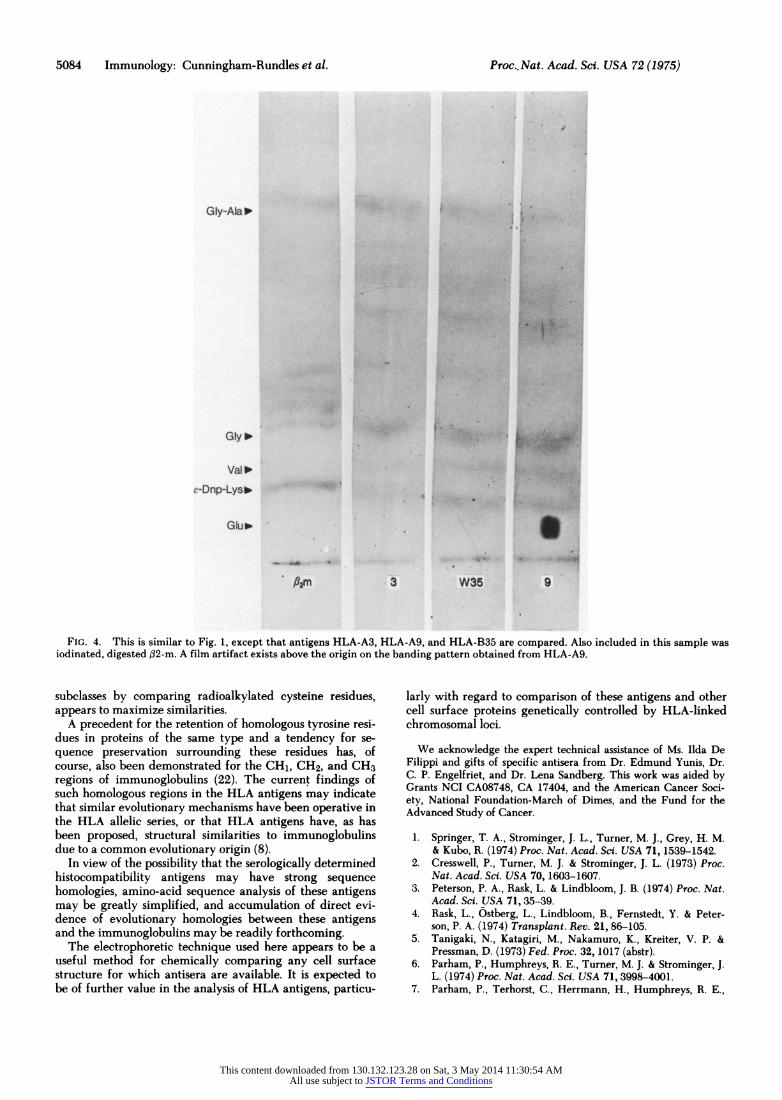
Fig. 4 illustrates the banding patterns observed for digest- ed antigens HLA-A3, HLA-B19, and HLA-BW35; similarly digested radioiodinated urinary f32-m is included in this fig- ure in order to compare peptides produced from free #32-m and the HLA aggregate which incorporates f32-m. A differ- ence in peptide mobilities is observed although several /32-in peptides appear to have mobilities similar to those of pep- tides of the HLA antigens. Again, the HLA antigens them- selves have a banding pattern similar to one another.
DISCUSSION
Although about 40 antisera are known to define antigenic
specificities of the HLA system, the current study has been~~ undertaken in rder to provid evidence for he possibilit
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions

Immunology: Cunningham-Rundles et al. Proc. Nat. Acad. Sc. USA 72(1975) 5083
GhJb
I+ onghi_
FIG. 3. Trypsin digestion has been allowed to proceed for 12 hr. HLA-A3 and HLA-A7 are compared.
that a rather uniform molecular structure underlies this ap- parent heterogeneity. HLA antigens digested with pepsin and trypsin have eight to ten tyrosine-containing peptides, the range of mobilities of which are either similar or identi- cal in different samples tested. This remained true even when the trypsin digestion was lengthened and the peptide patterns obtained were different. Since in other experiments using these antisera no molecular species other than those expected from detergent isolation of HLA antigens were found on polyacrylamide gels, it is very unlikely that extra- neous (non-HLA) antigens are present and responsible for this appearance.
The theoretical number of tyrosine peptides that could be produced by enzyme digestion of HLA antigens is not cer- tain, but a reported four to five tyrosine residues in the larg- er (papain-derived) peptide (17) and six tyrosines from ,82-m (18) would produce a total of 12 to 13 radioactive peptides if cleavage between each tyrosine were possible. In fact, only five major and one minor radioactive peptide are actually produced from enzyme-digested urinary ,82-in (Fig. 4). The reason for this may be evident from amino-acid sequence
analysis, as residues 63, 66, and 68 of ,32-m are tyrosine (19). Particularly between the latter two residues (where there is an intervening serine) cleavage would be hindered. If the HLA aggregate (as demonstrated here) has eight to ten ra- dioactive peptides and six of these arise from ,82-m, the re- maining two to four peptides could originate from the larger polypeptide, which would correlate with the number of ty- rosines reported in this component, although as noted, this value was based upon a molecular weight of 33,000 for the papain-derived component (17).
Definite correlation of the tyrosyl peptides produced from pure /32-m to peptides produced from the HLA antigen ag- gregate is not readily possible (Fig. 4). This may be due to differences in availability of tyrosine residues in free /32-m as opposed to /32-m incorporated in the HLA aggregate. This was especially probable since the HLA aggregate was iodin- ated in a native state while on the surface of the lympho- cyte. This difference in peptides between free and bound /32-m could indicate that /2-m is a relatively unexposed polypeptide in the HLA complex.
Regardless of the exact origin of each peptide, the striking similarity of tyrosine peptides produced by enzyme cleavage of HLA proteins provides direct evidence that the primary amino-acid sequences of the serologically determined histo- compatibility antigens have strong homologies.
This finding would be in agreement with recent sugges- tions from other laboratories. Variation in number of sialic acid residues is evidently a primary reason for charge heter- ogeneity of HLA antigens of different specificities (6), and three HLA preparations (each containing several specifici- ties) had only glycine as a detectable NH2-terminus in addi- tion to isoleucine, which could be assigned to the NH2-ter- minus of /32-m (19). Additionally, amino-acid analyses of two HLA specificities, HLA-A2 and HLA-B7, have recently been shown to be quite similar (17). Moreover, it appears that a general structural feature of the 45,000-dalton compo- nent is a series of three compact domains connected by sev- eral stretches of polypeptide chain, each domain containing a single disulfide bridge (8) and enclosing 60 to 70 amino acids. This would certainly indicate the preservation of ho- mologous features in HLA antigens of diverse specificities.
HLA-C (third series) antigens have the same molecular weight as HLA-A and HLA-B antigens, but appear to be more vulnerable to proteolysis, since the 45,000-dalton com- ponent isolated by detergents is often reduced to 28,000- 30,000 daltons during preparation (P. Wernet, C. Cunning- ham-Rundles, C. Jersild, A. Svejgaard, B. Dupont, R. A. Good, and H. Kunkel, submitted for publication). Although this vulnerability may be due to primary sequence differ- ences among the HLA-C and HLA-A and HLA-B antigens, the underlying homology of tyrosine peptides demonstrated here emphasizes the similarity of HLA-C antigens to those of the other two chromosomal loci.
Based upon ion exchange chromatography of tryptic pep- tides of several H2 specificities labeled with [3H]- and [14C]lysine and arginine, Brown and coworkers have con- cluded that a divergent range of peptides may be encoun- tered for murine antigens of different specificities (20). Rec- onciliation between these findings and those of the current report may be possible if there is a preservation of peptide sequence in the region surrounding tyrosine residues, while the intervening stretches may have less similar sequences. As Brown et al. point out, their technique maximizes peptide differences. The technique used here, in principle similar to that of Frangione et al. (21) in the chemical typing of IgG
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions
5084 Immunology: Cunningham-Rundles et al. Proc. Nat. Acad. Sci. USA 72(1975)
"TT
1,
Gly-Aial
Gly lo
Val '0
e-Dnp-Lys*'
FIG. 4. This is similar to Fig. 1, except that antigens HLA-A3, HLA-A9, and HLA-B35 are compared. Also included in this sample was iodinated, digested ,B2-m. A film artifact exists above the origin on the banding pattern obtained from HLA-A9.
subclasses by comparing radioalkylated cysteine residues, appears to maximize similarities.
A precedent for the retention of homologous tyrosine resi- dues in proteins of the same type and a tendency for se- quence preservation surrounding these residues has, of course, also been demonstrated for the CH1, CH2, and CH3 regions of immunoglobulins (22). The current findings of such homologous regions in the HLA antigens may indicate that similar evolutionary mechanisms have been operative in the HLA allelic series, or that HLA antigens have, as has been proposed, structural similarities to immunoglobulins due to a common evolutionary origin (8).
In view of the possibility that the serologically determined histocompatibility antigens may have strong sequence homologies, amino-acid sequence analysis of these antigens may be greatly simplified, and accumulation of direct evi- dence of evolutionary homologies between these antigens and the immunoglobulins may be readily forthcoming.
The electrophoretic technique used here appears to be a useful method for chemically comparing any cell surface structure for which antisera are available. It is expected to be of further value in the analysis of HLA antigens, particu-
larly with regard to comparison of these antigens and other cell surface proteins genetically controlled by HLA-linked chromosomal loci.
We acknowledge the expert technical assistance of Ms. Ilda De Filippi and gifts of specific antisera from Dr. Edmund Yunis, Dr. C. P. Engelfriet, and Dr. Lena Sandberg. This work was aided by Grants NCI CA08748, CA 17404, and the American Cancer Soci- ety, National Foundation-March of Dimes, and the Fund for the Advanced Study of Cancer.
1. Springer, T. A., Strominger, J. L., Turner, M. J., Grey, H. M. & Kubo, R. (1974) Proc. Nat. Acad. Sc. USA 71,1539-1542.
2. Cresswell, P., Turner, M. J. & Strominger, J. L. (1973) Proc. Nat. Acad. Sci. USA 70,1603-1607.
3. Peterson, P. A., Rask, L. & Lindbloom, J. B. (1974) Proc. Nat. Acad. Sci. USA 71,35-39.
4. Rask, L., Ostberg, L., Lindbloom, B., Fernstedt, Y. & Peter- son, P. A. (1974) Transplant. Rev. 21, 86-105.
5. Tanigaki, N., Katagiri, M., Nakamuro, K., Kreiter, V. P. & Pressman, D. (1973) Fed. Proc. 32, 1017 (abstr).
6. Parham, P., Humphreys, R. E., Turner, M. J. & Strominger, J. L. (1974) Proc. Nat. Acad. Sci. USA 71, 3998-4001.
7. Parham, P., Terhorst, C., Herrmann, H., Humphreys, R. E.,
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions

Immunology: Cunningham-Rundles et al. Proc. Nat. Acad. Sci. USA 72(1975) 5085
Waterfield, M. D. & Strominger, J. L. (1975) Proc. Nat. Acad. Sci. USA 72, 1594-1598.
8. Peterson, P. A., Rask, L., Sege, K., Klareskog, L., Anudi, H. & Ostberg, L. (1975) Proc. Nat. Acad. Sci. USA 72, 1612-1616.
9. Marchalonis, J. J. (1969) Biochem. J. 113, 299-305. 10. Cunningham-Rundles, C., Jersild, C., Dupont, B. & Good, R.
A. (1975) Tenth Leukocyte Culture Conference Abstract, Amsterdam.
11. Boyum, A. (1967) Scan. J. Clin. Lab. Invest. 21, suppl. 97, 77-89.
12. Vitetta, E. S., Baur, S. & Uhr, J. W. (1971) J. Exp. Med. 134, 242-264.
13. Schwartz, B. D & Nathenson, S. G. (1971) J. Immunol. 107, 1363-1367.
14. Wernet, P., Jersild, C., Dupont, B., Hansen, J. A., Svejgaard, A., Good, R. A. & Kunkel, H. G. (1974) Brook Lodge Confer- ence, "Immunological and Chemical Approaches to the Cell
Surface," June 17-21, eds Kahan, B. & Reisfeld, R. (Plenum Press, New York).
15. Heilman, J., Barrollier, J. & Watzke, E. (1957) Hoppe-Seyler's Z. Physiol. Chem. 309,219-221.
16. Shapiro, A. L., Vifiuela, E. & Maizel, J. V. (1967) Biochem. Biophys. Res. Commun. 28,815-819.
17. Tanigaki, N. & Pressman, D. (1974) Transplant. Rev. 21, 15- 34.
18. Berggard, I. & Bearn, A. G. (1960) J. Biol. Chem. 243, 4095- 4103.
19. Cunningham, B. A., Wang, J. L., Berggaird, I. & Peterson, P. A. (1973) Biochemistry 12, 4811-4822.
20. Brown, L. J., Kato, K., Silver, J. & Nathenson, S. G. (1974) Biochemistry 13, 3174-3178.
21. Frangione, B., Milstein, C. & Franklin, E. C. (1969) Nature 221 149-151.
22. Edelman, G. M. (1970) Biochemistry 9,3197-3205.
This content downloaded from 130.132.123.28 on Sat, 3 May 2014 11:30:54 AMAll use subject to JSTOR Terms and Conditions