evaluation of plaque assay for the maedi-progressive pneumonia
TRANSCRIPT

APPLIED MICROBIOLOGY, Sept. 1974, p. 366-373Copyright 0 1974 American Society for Microbiology
Vol. 28, No. 3Printed in U.S.A.
Evaluation of a Plaque Assay for the Maedi-ProgressivePneumonia-Visna Viruses
R. S. TROWBRIDGE
New York State Institute for Research in Mental Retardation, Staten Island, New York 10314
Received for publication 8 May 1974
A simple and direct plaque assay for maedi virus, two strains of progressivepneumonia virus, and two strains of visna virus has been developed andevaluated. The technique allows the plaques formed by these viruses to belocalized without disturbing the host-cell substrate of sheep choroid plexus cellsor the gelled maintenance medium over the host-cell monolayer. Diethylamino-ethyl-dextran supplementation of the medium used to overlay strain K796 visnavirus-infected cultures decreases the time required for maximum plaque develop-ment from 12 to 10 days, enhances the contrast of the plaques, increases the titerof plaque-forming units, and permits a plaque size heterogeneity to be realized.Both large and small plaques occur in cultures infected with the visna viruses,one strain of progressive pneumonia virus, or maedi virus. In contrast, theplaques observed in cultures infected with the second strain of progressivepneumonia virus are relatively homogeneous in size.
The viruses of visna, maedi, and progressivepneumonia are the cause of disease in sheep,characterized by paresis and/or dypsnea whichultimately and irreversibly progresses to thenatural host's death (5, 6, 8-11, 20; N. G. Rogerset al., Bacteriol. Proc., p. 181, 1971). The stud-ies with these viruses have been dependentupon an end-point assay which determined theamount of virus capable of producing a cyto-pathic effect in most of the cells within 14 to 21days after infection of the host-cell population.The end points obtained depend not only uponthe amount of virus present but also have beenstated to depend on both the strain and the pas-sage level of the cells (6, 16, 19). These factscomplicate studies of the host-virus interaction.Additionally, these viruses have not been sub-jected to procedures which would allow genet-ically pure lines of virus to be obtained.
Plaque formation by visna virus has beenreported by Harter (3) and by Harter andChoppin (4) to require 11 to 14 days. Visualiza-tion of the visna virus-induced plaques involveseither fixing and staining the host-cell substrateor adding a secondary cellular overlay to thevirus-infected host cell monolayer. The numberof visna virus-specific plaque-forming units ob-tained by the former method was found to be2.5-fold greater than the number of tissue cul-ture infective units obtained by end-point titra-tion (3). In contrast, Lopez, Eklund, and Had-low found the titer of progressive pneumoniavirus obtained by this plaque assay to be
361
one-tenth the titer obtained by end-point titra-tion (6). The use of a secondary cellular overlaycomposed of cells which are fused and killed byvisna virus, but which do not support virussynthesis, will not detect a virus incapable ofcausing cell fusion or plaques containing rela-tively little progeny virus.
Subsequent reports note the use of plaqueassays for both visna virus and progressivepneumonia virus, and the plaque purification ofthese viruses (2, 12, 13, 14). These reports do notdescribe the assay or demonstrate that it satis-fies the criteria enumerated by P. D. Cooper forthe assay to be useful (1). Additionally, plaqueformation by a second strain of progressivepneumonia virus and by maedi virus has notbeen reported to occur.This report describes and evaluates a rela-
tively simple and efficient plaque assay for fivemembers of the maedi-progressive pneumonia-visna virus group.
MATERIALS AND METHODSCells. The choroid plexus was aseptically removed
from the brains of recently slaughtered lambs andtransferred to and minced in a petri dish containingseveral milliliters of growth medium (GM). The GMconsisted of 86 parts of Eagle basal medium withEarle salts containing 100 units of penicillin per ml,100 lgg of streptomycin per ml, 0.375 g of sodium bi-carbonate per liter, 200 mM L-glutamine, and 14 partsof lamb serum (Colorado Serum Company, Denver,Colo.). The medium without the serum is referred toas BME. The minced choroid plexus was rinsed with
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

MAEDI-PROGRESSIVE PNEUMONIA-VISNA VIRUSES
several changes of GM to remove the adherent blood,suspended in 2 ml of GM, transferred to a 25-cm2plastic flask, and incubated at 37 C until the tissueattached to the vessel's surface (2 days). An addi-tional 3.0 ml ofGM was then added to each vessel andthe cultures were reincubated at 37 C. A monolayer ofconfluent fibroblastic sheep choroid plexus (SCP)cells was present within 10 days after the tissue wasexplanted. These cells were dispersed with and sus-pended in a 0.1% solution of trypsin in BME. Theresulting cell suspension was transferred to a 250-mlflask containing 25 ml ofGM and incubated for 5 to 6days at 37 C. This transfer represented passage num-ber one. The confluent cells were then trypsinized,transferred to, and routinely propagated in 32-ozprescription bottles. Each bottle was seeded with 2 x106 to 3 x 106 SCP cells, and the cells were fed 50 mlof GM and harvested 3 or 4 days later when the num-ber of cells was 1.5 x 107 to 3.0 x 107 per 32-oz bottle.SCP cells from different choroid plexi were considereddifferent strains.Low passage-level cells of the individual strains
were frozen in 1.0-ml ampules at a cell density of 106to 2 x 106 cells per ml ofGM supplemented with 10%(vol/vol) glycerol and stored in liquid nitrogen. Thecells in these ampules were rapidly thawed in a 37 Cwater bath, transferred to a 250-ml plastic flask, fed25 ml ofGM, and incubated at 37 C. The medium wascompletely changed 24 h later. The cells from suchampules became confluent within 10 days after beingthawed and cultured, and were then transferred to32-oz bottles for batch cultivation.
Viruses. Strain K485 and strain K796 visna wereoriginally obtained from the brains of Icelandic sheepexperimentally infected with visna virus (17, 18). Twostocks of strain K485 visna virus, one in its eighth invitro passage and the second in its forty-first in vitropassage, were used throughout this study. These twostocks of visna virus will be referred to as K485 p8 andas K485 p41, respectively. Both virus stocks werepropagated in SCP cultures maintained in mainte-nance medium (MM) which consisted of BME sup-plemented with 0.2% bovine serum albumin. Theculture medium was harvested when these culturesexhibited maximal cytopathic effect.The eighteenth tissue culture passage of strain
K796 visna virus was propagated in confluent SCPcell cultures maintained in GM. The virus washarvested and the cultures were refed fresh GM 72 hafter the cultures were infected with one mean tissueculture infective dose (TCOD50) of seventeenth tissueculture passage visna virus (referred to as virus stock1) per cell. This harvest represented one virus stockand is referred to as stock 2. A third virus stock wasobtained by removal of the medium from thesecultures 24 h after they were refed and is referred to asvirus stock 3.The M-88 strain of maedi virus (18) was propagated
in confluent SCP cell cultures maintained in MM.The virus was harvested 10 days after the cultureswere infected. This harvest represented the sixteenthserial passage of this maedi virus strain.
Progressive pneumonia virus was generously sup-plied by C. J. Gibbs, Jr. and D. C. Gajdusek of the
National Institute of Neurological Diseases andStroke, Bethesda, Md. It originated from the brain ofa sheep affected with Montana sheep disease (N. G.Rogers et al., Bacteriol. Proc., p. 181, 1971). Virus washarvested 11 days after SCP cultures were infected.This harvest represents the fifth passage of this virusin SCP cell cultures. This strain of progressive pneu-monia virus will be referred to as G-PPV.
K. K. Takemoto (National Institute of Allergy andInfectious Diseases, Bethesda, Md.) graciously pro-vided the second strain of progressive pneumoniavirus, referred to as T-PPV. It originated from thelungs of a sheep affected with progressive pneumoniain Montana (5, 6) and has been serially passed eighttimes in cultures of sheep testicle cells.
Assay of infectious virus. Cultures of confluentSCP cells in 21-cm2 plastic petri dishes were preparedfor the plaque assay by being initiated with 3.5 x 101to 4.0 x 105 SCP cells in 5.0 ml of GM. The cells inthese cultures were confluent and were routinely usedfor the plaque assay 3 days after being initiated andincubated at 37 C in a humidified atmosphere con-taining 5% C02. The GM was removed from and 1.0ml of serial dilutions of virus was added to eachculture. Virus was diluted in cold MM. Adsorption ofvirus was allowed to occur for 2 h at 37 C. The virusinoculum was redistributed over the monolayer athalf-hour intervals during the 2-h viral-adsorptionperiod.The cell sheet was then overlaid with 5.0 ml of
plaque medium (PM). The 1.0 ml of diluted virusinoculum and the PM were thoroughly mixed as thePM was being added to each culture. This PMconsisted of double-strength BME without phenol redsupplemented with 0.4% bovine serum albumin, my-costatin (200 units/ml), and an additional 1.125 g ofsodium bicarbonate per liter mixed with an equalvolume of 1% agarose (Fisher Scientific Co., Cat.#J-24040-S). The agarose was suspended in steriledeionized water and sterilized in a boiling water bathfor 1 h prior to being used in the plaque assay. Thesterilized agarose was stored at 4 C and was not usedfor this initial overlay if it had been remelted morethan once. After the PM in the plate cultures hadsolidified, the cultures were reincubated at 37 C inhumidified air containing 5% C02. A 2.5-ml feederlayer of PM was added to the initial PM overlay onthe tenth postinfection day in cultures infected withG-PPV or maedi virus.A 2-ml amount of PM containing 1: 10,000 (wt/vol)
neutral red was added to the cultures at various timesafter they were infected. The viable cells incorporateda sufficient amount of neutral red for the plaques to beobserved within 4 to 6 h of incubation at 37 C.
Visualization of the plaques was enhanced byviewing the stained monolayer against an evenlyilluminated background covered with a transparentmedium blue-green filter (Edmund Scientific Co.,Barrington, N.J., Cat. #82,042).
Infective strain K796 visna virus titers were alsodetermined by the 50% end-point method describedby Thormar (15). Five confluent test-tube cultures ofSCP cells were used for each 10-fold dilution of virus.
Antiserum and viral neutralization. Visna virus-
367VOL. 28, 1974
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

TROWBRIDGE
specific antiserum no. 8909 was obtained from a sheep(no. K814) 4 years after it had been intracerebrallyinjected with visna virus (18, 20). Both this antiserumand normal lamb serum were inactivated at 56 C for0.5 h and then were diluted and added to PM so thatthe final concentration of serum was 0.4%. Thenumber of plaques in those cultures infected withapproximately 50 plaque-forming units (PFU) andoverlaid with PM containing 0.4% antiserum was
compared to the number of plaques in cultures thatwere infected with the same amount of virus butoverlaid with PM containing 0.4% inactivated normallamb serum.
Chemicals. The stock solution of DEAE-dextran(molecular weight 2 x 106l) and the stock solution ofdextran sulfate (molecular weight 5 x 10') (Phar-macia Fine Chemicals, Inc., Piscataway, N.J.) con-
tained 5 mg of polyion per ml of deionized water.These were autoclaved for 15 min at 15 lb/in2.The stock solution of neutral red (Fisher Scientific
Co., Springfield, N.J.) contained 1 g of neutral red per100 ml of deionized water.
RESULTS
Influence of polyions on plaque formation.Preliminary studies, performed to determine ifSCP cell monolayers could concentrate neutralred, indicated that these cells remained viablefor at least 24 days without further additions ofPM. Subsequent studies showed that visnavirus formed plaques on SCP cell monolayersand that the plaques could be visualized by theaddition of neutral red to the cultures on thetwelfth day after infection. The plaques were
difficult to enumerate at this time due to thepoor contrast between the unstained plaquearea and the surrounding neutral red-stainedcells (Fig. 1). Substitution of either lamb serum
or of fetal calf serum for bovine serum albuminin the PM only resulted in a decrease in thecontrast between the plaques and their periph-eral viable cells with a concomitant decrease inthe number of plaques.
Since polyions have been shown to dramati-cally alter both the efficiency of plaque forma-tion (eop) and the morphology of the plaquesproduced by viruses (1, 7), we proceeded todetermine if these chemicals would increase theeop of visna virus on SCP cell monolayers. Theaddition of either diethylaminoethyl (DEAE)-dextran or dextran sulfate to the medium usedboth for virus dilution and for viral adsorptionresulted in a significant decrease in the plaque-forming titer (Table 1). Dextran sulfate was alsofound to dramatically decrease the plaque-forming titer when it was added to the PM usedto overlay the virus-infected SCP cell mono-
layer (Table 1). In contrast, DEAE-dextranincreased the titer of visna virus and improvedthe contrast between the plaque and its sur-
rounding cells when this polycation was addedto the PM (Table 1 and Fig. 1). There was also amore noticeable heterogeneity of plaque size inmonolayers overlaid with PM containingDEAE-dextran than in monolayers overlaidwith PM not containing DEAE-dextran.The addition of DEAE-dextran to the PM
was also found to decrease the time required formaximum plaque development (Table 2). Theplaque number in monolayers infected withapproximately 60 PFU and overlaid with PMwas less than that observed in similar culturesoverlaid with PM containing DEAE-dextranwhen these cultures were stained and theplaques enumerated on the 9th, 10th, and 11thdays after infection. The number of plaques inthose cultures overlaid with PM containingeither no DEAE-dextran or 5 pg of this polycat-ion per ml was one-fifth to one-half of thenumber of plaques observed in those culturesoverlaid with PM containing 10 or 15 ,ug of thispolycation per ml of PM. The number ofplaques in the cultures overlaid with PM notsupplemented with DEAE-dextran increasedfivefold from the 9th to the 12th day afterinfection. In contrast, the number of plaques inthe cultures overlaid with PM containing 5 ,ug ofthis polycation increased only twofold duringthis interval. Similarly, the number of plaquesobserved on the 11th day in cultures overlaidwith PM containing 10 or 15 ,g of DEAE-dex-tran was only 1.1-fold greater than the 9-dayvalue.These results show that DEAE-dextran, at a
concentration of 10 or 15 ug of PM per ml,increased the eop by visna virus. Additionally,the presence of this polycation decreased thetime required for maximum plaque develop-ment from greater than 12 days to 10 days. Theplaques in those cultures overlaid with PMcontaining DEAE-dextran began to coalesce onthe 11th day after infection. The decrease in thenumber of plaques observed in these cultures onthe 12th day was therefore due to plaqueoverlap. The diameter of the plaques observedon the 10th day postinfection in visna virus-infected cultures ranged from 0.9 to 5.4 mmwith an average diameter of 3.1 mm. The resultsto be reported hereafter were obtained fromplate cultures overlaid with PM containing 10to 20 Aig of DEAE-dextran per ml.Plaque formation by maedi virus, T-PPV,
G-PPV, and strain K485 visna virus. Theresults with strain K796 visna virus led to ourdetermining how long cultures infected withmaedi virus, two strains of progressive pneumo-nia virus, or two different stocks of strain K485visna virus and overlaid with PM supplemented
APPL. MICROBIOL.368
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

MAEDI-PROGRESSIVE PNEUMONIA-VISNA VIRUSES
FIG. 1. Plaques produced by strain K796 of visna virus. Cultures were stained with neutral red for 4 to 6 h onday 10 after infection. Culture overlaid with plaque medium (left) and culture overlaid with plaque mediumsupplemented with 15 ,g of DEAE-dextran per ml (right).
TABLE 1. Effect of polyions on visna virusplaque-forming titer
Titer (PFU x 105/ml) whenPolyion added polyion added to
(Og/ml) Adsorption Plaquemedium medium
None 180 220DEAE-dextran (10) 60 290DEAE-dextran (20) 55 310Dextran sulfate (10) 0.4 <0.1Dextran sulfate (20) 0.15 <0.1
with 15 ,gg of DEAE-dextran per ml required toattain a maximum plaque number (Table 3). Amaximum plaque number in cultures infectedwith T-PPV was present on the 10th day afterinfection. Cultures infected with K485 p41 re-quired 12 days to attain a maximum plaquenumber, whereas cultures infected with K485p8 required 13 days to attain their maximumplaque number. In contrast, maedi virus-infected cultures and G-PPV-infected culturescontained a maximum number of plaques on
the 16th and the 18th day after infection,respectively.
TABLE 2. Increase of visna virus plaque number onSCP cell monolayers overlaid with PM and PMsupplemented with various concentrations of
DEAE-dextran
Mean plaque no.a onDEAE-dextran concn postinfection day
in PM (ug/ml)9 10 11 12
0 10 28 38 495 24 33 44 4010 54 53 58 5415 53 56 60 51
a Values in the table are the mean number ofplaques per plate obtained from three plates for eachpostinfection day.
A plaque size heterogeneity was observed incultures infected with strain K485 visna virus,strain K796 visna virus, T-PPV, and maedivirus (Fig. 1 and 2), whereas the size of plaquesin G-PPV-infected cultures was relatively morehomogeneous (Fig. 2).
Evaluation of the plaque assay. This plaqueassay's utility was demonstrated by the exist-ence of a linear relationship between relativevirus concentration and plaque number (Table
VOL. 28, 1974 369
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

TABLE 3. Time of maximum plaque formation in cultures infected with strain K485 visna virus, maedi virus,T-PPV, or G-PPV and overlaid with PM supplemented with DEAE-dextran
Mean plaque no. on postinfection dayaVirus - _ _ _ _ _ _ _ _ _ _ _ _ _
7 8 9 10 11 12 13 14 16 18 20
T-PPV 28 41 75 96 96K485 p8 11 27 56 62 84 75K485 p41 13 18 23 35 34 34Maedi 45 52 50 52G-PPV 6 37 72 60
a Values are the mean number of plaques per plate obtained from three plates for each postinfection day.
4), by the absence of plaques in uninfectedcultures, by the presence of 1,000 to 2,000 PFUin the plaque's center and the absence ofdetectable virus (less than 40 PFU) 8 mm fromthe plaque's center, by plaque-formation notoccurring when strain K796 visna virus-infectedcultures were overlaid with.,PM containing anti-serum, and by the cell passage level not affect-ing the titer of plaque-forming units in a pre-served visna virus stock (Table 5) (1).The sensitivity and the reproducibility of the
plaque assay and of the end-point assay werecompared. One stock of strain K796 visna viruswas assayed by plaque technique on one strainof SCP cells over a 5-month period. This samestrain of cells at the 11th and at the 12thpassage level was used to determine the TCID5Otiter. Twenty tube cultures were used for each10-fold dilution of virus. Tissue culture infec-tious units (TCIU) were obtained by multiply-ing the TCID5O titers by 0.69. The mean numberof PFU per milliliter was 1.5 x 107 and thestandard deviation was 2.7 x 106 (Table 5). Themean number of TCIU per milliliter was 2.8 x106 and the standard deviation was 1.4 x 106.The PFU: TCIU ratio was 5.4. These resultsshow that the plaque assay is a more sensitiveand a more reproducible indicator of visna virusinfectivity than is the end-point assay.Three different strains of SCP cells were used
to assay one strain K796 visna virus stock todetermine the differences in susceptibility ofthese SCP cell strains to visna virus-inducedplaque formation (Table 6). Two of the threetested cell strains yielded equivalent PFU titerswhich were 10 times the titer obtained on thethird cell strain.
DISCUSSIONThe present work shows that SCP cells incor-
porate neutral red and that the plaques pro-duced on SCP cell monolayers by five membersof the maedi-progressive pneumonia-visna viruscomplex can be detected with the aid of this
vital stain. Incorporation of the polycation,DEAE-dextran, into the plaque medium en-hances the contrast between the plaques andtheir peripheral viable cells, increases the ap-parent efficiency of plaque formation, decreasesthe time required for plaque formation by visnavirus, and results in a plaque size heterogeneitybeing observed in cultures infected with maedivirus, visna virus, and one strain of PPV. Theenhancing effect of DEAE-dextran and theinhibitory effect of dextran sulfate (Table 1)when these polyions are present in the plaquemedium indicates that the agarose containssulfated polysaccharides which are coun-teracted in their effects by the polycation.These results are in accord with the observationby Harter (3) of visna virus having a higher eopon monolayers maintained under medium con-taining carboxymethylcellulose than in cellsoverlaid with medium containing either agar oragarose.The finding that the eop of visna virus may
vary from one SCP cell strain to another (Table6) demonstrates that all strains of SCP cells arenot equally suitable for use in the plaque assayand dictates that a comparison of the eop ofvisna virus on several cell strains must be madebefore any one cell strain is used for the assay.The visna virus titer obtained on high passage
level cells, on cells which had been subjected tostorage in liquid nitrogen, and on low passagelevel cells was found to not be significantlydifferent (Table 5). Therefore, cells of thosestrains which serve as a suitable host substratein the plaque assay, as evidenced by yielding acomparatively high visna virus eop, may befrozen and stored in liquid nitrogen for futureuse in the plaque assay.The finding that the plaque-forming titer of a
given visna virus sample is fivefold greater thanthe titer of TCIU indicates that infection of aculture requires at least five infectious particlesfor the culture to be scored as infected 14 daysafter infection. This indicates that clones of
370 TROWBRIDGE APPL. MICROBIOL.
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

MAEDI-PROGRESSIVE PNEUMONIA-VISNA VIRUSES
FIG. 2. Plaques in SCP cell cultures infected with T-PPV for 10 days (top, left), with maedi virus for 18 days(top, right), and with G-PPV for 18 days (bottom). Magnification x1.4.
371VOL. 28, 1974
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from
TROWBRIDGE
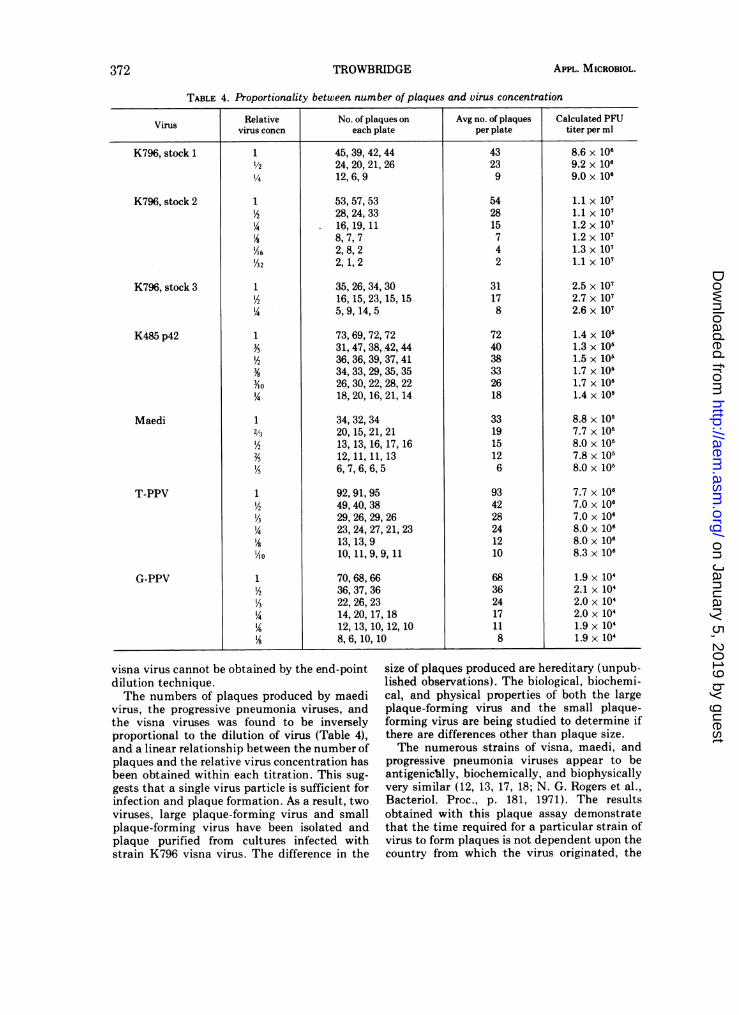
TABLE 4. Proportionality between number of plaques and virus concentration
Virus Relative No. of plaques on Avg no. of plaques Calculated PFUvirus concn each plate per plate titer per ml
45,39,42,4424,20,21,2612, 6, 9
53, 57, 5328, 24, 3316, 19, 118,7,72,8,22, 1,2
35,26,34,3016,15,23,15,155, 9, 14, 5
73,69,72,7231,47,38,42,4436,36,39,37,4134,33,29,35,3526,30,22,28,2218, 20, 16, 21, 14
34,32,3420,15,21,2113, 13, 16, 17, 1612, 11, 11, 136, 7, 6, 6, 5
92,91,9549,40,3829,26,29,2623,24,27,21,2313, 13, 910, 11, 9, 9, 11
70,68,6636,37,3622,26,2314,20,17,1812, 13, 10, 12, 108, 6, 10, 10
43239
542815742
31178
724038332618
331915126
934228241210
68362417118
8.6 x 1069.2 x 1069.0 x 106
1.1 x 1071.1 x 1071.2 x 1071.2 x 1071.3 x 1071.1 X 107
2.5 x 1072.7 x 1072.6 x 107
1.4 x 1051.3 x 1051.5 x 1051.7 x 1051.7 x 1051.4 x 105
8.8 x 1057.7 x 1058.0 x 1057.8 x 1058.0 x 105
7.7 x 1067.0 x 1067.0 x 1068.0 x 1068.0 x 1068.3 x 106
1.9 x 104
2.1 x 1042.0 x 1042.0 x 1041.9 X 1041.9 X 104
visna virus cannot be obtained by the end-pointdilution technique.The numbers of plaques produced by maedi
virus, the progressive pneumonia viruses, andthe visna viruses was found to be inverselyproportional to the dilution of virus (Table 4),and a linear relationship between the number ofplaques and the relative virus concentration hasbeen obtained within each titration. This sug-
gests that a single virus particle is sufficient forinfection and plaque formation. As a result, twoviruses, large plaque-forming virus and smallplaque-forming virus have been isolated andplaque purified from cultures infected withstrain K796 visna virus. The difference in the
size of plaques produced are hereditary (unpub-lished observations). The biological, biochemi-cal, and physical properties of both the largeplaque-forming virus and the small plaque-forming virus are being studied to determine ifthere are differences other than plaque size.The numerous strains of visna, maedi, and
progressive pneumonia viruses appear to beantigenic~ally, biochemically, and biophysicallyvery similar (12, 13, 17, 18; N. G. Rogers et al.,Bacteriol. Proc., p. 181, 1971). The resultsobtained with this plaque assay demonstratethat the time required for a particular strain ofvirus to form plaques is not dependent upon thecountry from which the virus originated, the
K796, stock 1
K796, stock 2
K796, stock 3
K485 p42
Maedi
T-PPV
G-PPV
1
'/2'A1
'6/32
1
l/4
1
2/3
Y5
Yl o
12/.3
I/5
1I/2
I/3
VI o
1I/2
I/3
'4'/6
1S
APPL. MICROBIOL.372
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from

MAEDI-PROGRESSIVE PNEUMONIA-VISNA VIRUSES
TABLE 5. Influence ofSCP cell passage level on strainK796 PFU titer
Date Mean _ Coefficient ofplates Passage PFUJ sb variation forseeded v titera individual assay (%)
12/4 7 1.8 x 10' 3.5 2012/8 10 1.3 x 10' 1.8 141/2 16 1.4 x 107 1.8 13
1/11 17 1.4 x 10' 3.0 211/18 9 1.8 x 10' 1.2 61/18 19 1.3 x 10' 1.4 111/25 10 1.7 x 10' 1.9 111/29 22 1.1 x 10' 1.4 125/15 13 1.8 x 10' 1.2 6
aPlaque-forming units per milliliter obtained from themean of the number of plaques from three plates.
h Standard deviation. Mean titer for 9 assays = 1.5 x 10'PFU/ml; standard deviation = 2.7 x 106; coefficient ofvariation = 17.6%.
TABLE 6. Plaque-forming titer of visna virus on threeSCP cell strains
Cell straina Mean PFU + S./ml
H 1.8 X 0.3 x107V 2.0 0.2 x 107Va 2.4 0.5 x 106
a Each individual cell strain originated from thechoroid plexus of a different sheep's brain. The cellscomposing all cultures had a fibroblastic morphology.organ of virus origin, or its in vitro passage level.Additionally, the virus' organ and country oforigin do not determine the morphology of theplaques produced by a particular virus. Thisplaque assay should allow investigators to de-termine if these viruses are biologically differ-ent.
ACKNOWLEDGMENTSThis work was partially supported by Public Health
Research grant #1R01 CA12678-01 from the National CancerInstitute.
I gratefully acknowledge Carl Torchio for his skillful anddedicated technical expertise and Halldor Thormar for sup-plying me with the viruses and antiserum.
LITERATURE CITED1. Cooper, P. D. 1967. The plaque assay of animal viruses, p.
243-311. In K. Maramorosch and H. Koprowsky (ed.),Methods in virology, vol. 3. Academic Press Inc., NewYork.
2. Haase, A. T., and W. Levinson. 1973. Inhibition of RNAslow viruses by thiosemicarbazones. Biochem. Biophys.Res. Commun. 51:875-880.
3. Harter, D. H. 1969. Observations on the plaque assay ofvisna virus. J. Gen. Virol. 5:157-160.
4. Harter, D. H., and P. W. Choppin. 1967. Plaque assay ofvisna virus using a secondary cellular overlay as anindicator. Virology 31:176-178.
5. Kennedy, R. C., C. M. Eklund, C. Lopez, and W. J.Hadlow. 1968. Isolation of a virus from the lungs ofMontana sheep affected with progressive pneumonia.Virology 35:483-484.
6. Lopez, C., C. M. Eklund, and W. J. Hadlow. 1971. Tissueculture studies of the virus of progressive pneumonia, aslow infectious disease of sheep. Proc. Soc. Exp. Biol.Med. 138:1035-1040.
7. Pattyn, S. R., and L. DeVleesschauwer. 1969. Plaqueproduction by group A arboviruses. II. Enhancingeffect of DEAE-dextran on plaque numbers in chickembryo cells. Acta Virol. 13:26-33.
8. Sigurdardottir, B., and H. Thormar. 1964. Isolation of a
viral agent from the lungs of sheep affected with maedi.J. Infect. Dis. 114:55-60.
9. Sigurdsson, B. 1954. Maedi, a slow progressive pneumo-nia of sheep: an epizoological and a pathological study.Brit. Vet. J. 110:255-270.
10. Sigurdsson, B., P. A. Palsson, and H. Grimsson. 1957.Visna, a demyelinating transmissible disease of sheep.J. Neuropathol. Exp. Neurol. 16:389-403.
11. Sigurdsson, B., H. Thormar, and P. A. Palsson. 1960.Cultivation of visna virus in tissue culture. Arch.Gesamte Virusforsch. 10:368-381.
12. Stone, L. B., K. K. Takemoto, and M. M. Martin. 1971.Physical and biochemical properties of progressivepneumonia virus. J. Virol. 8:573-578.
13. Stone, L. B., E. Scolnick, K. K. Takemoto, and S. A.Aaronson. 1971. Visna virus: a slow virus with an RNAdependent DNA polymerase. Nature (London)229:257-258.
14. Takemoto, K., and L. B. Stone. 1971. Transformation ofmurine cells by two "Slow Viruses", visna virus andprogressive pneumonia virus. J. Virol. 7:770-775.
15. Thormar, H. 1963. The growth cycle of visna virus inmonolayer cultures of sheep cells. Virology 19:273-278.
16. Thormar, H. 1963. Neutralization of visna virus byantisera from sheep. J. Immunol. 90:185-192.
17. Thormar, H. 1965. A comparison of visna and maediviruses. I. Physical, chemical and biological properties.Res. Vet. Sci. 6:117-129.
18. Thormar, H., and H. Helgadottir. 1965. A comparison ofvisna and maedi viruses. II. Serological relationship.Res. Vet. Sci. 6:456-465.
19. Thormar, H., and B. Sigurdardottir. 1962. Growth ofvisna virus in primary tissue cultures from variousanimal species. Acta Pathol. Microbiol. Scand.55:180-186.
20. Thormar, H., and P. A. Palsson. 1967. Visna and ma-
edi-two slow infections of sheep and their etiologicalagents, p. 291-308. In M. Pollard (ed.), Perspectives invirology, vol. 5. Academic Press Inc., New York.
VOL. 28, 1974 373
on January 5, 2019 by guesthttp://aem
.asm.org/
Dow
nloaded from