eukaryotic tree of life · petersenet al., 2014, genom. biol. evol. eukaryotic tree of life....
TRANSCRIPT
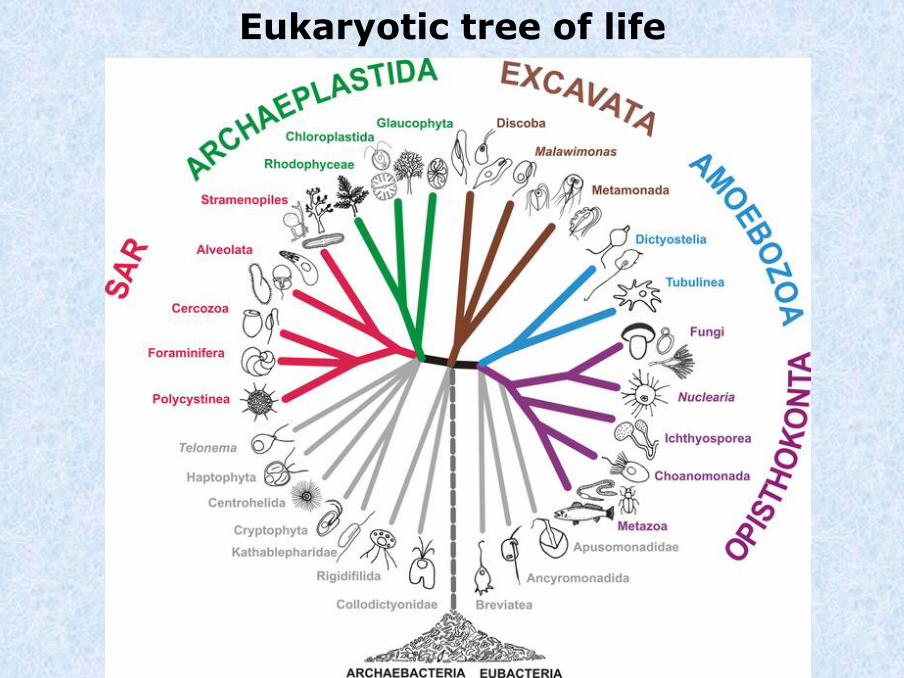
Eukaryotic tree of life
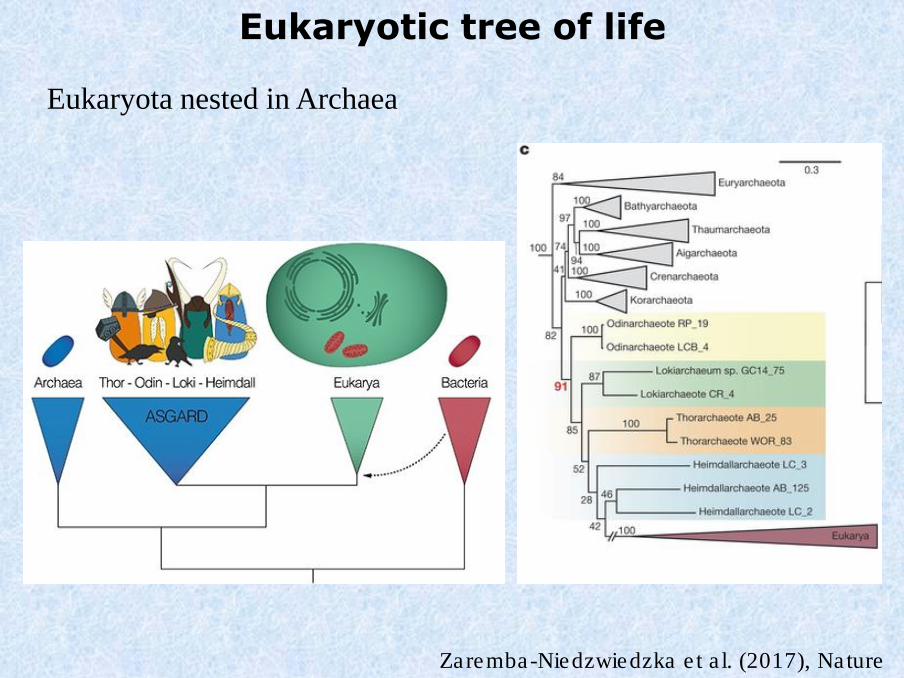
Eukaryotic tree of life
Eukaryota nested in Archaea
Zaremba-Niedzwiedzka et al. (2017), Nature
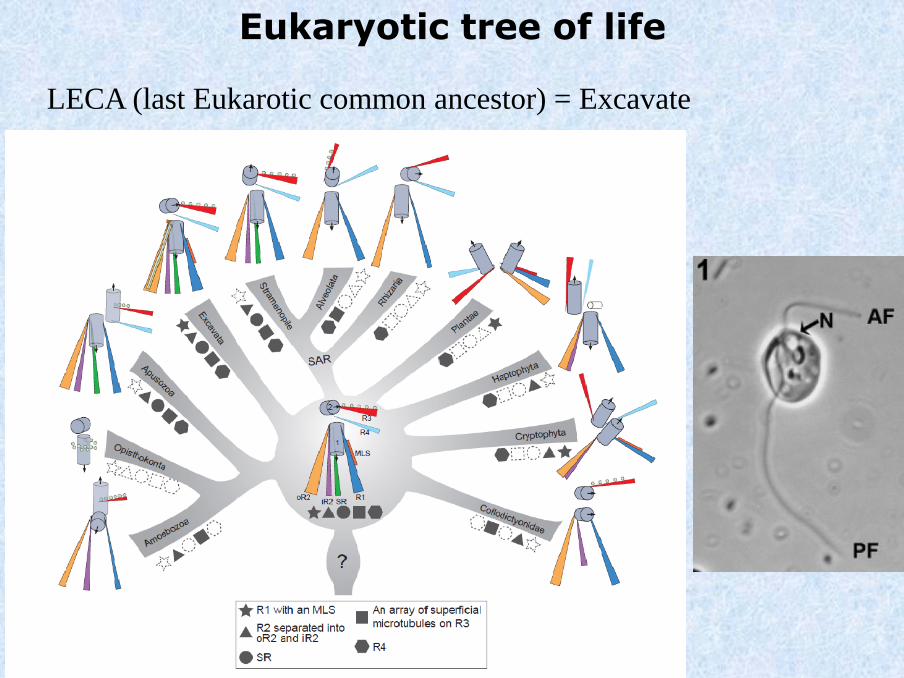
Eukaryotic tree of life
LECA (last Eukarotic common ancestor) = Excavate
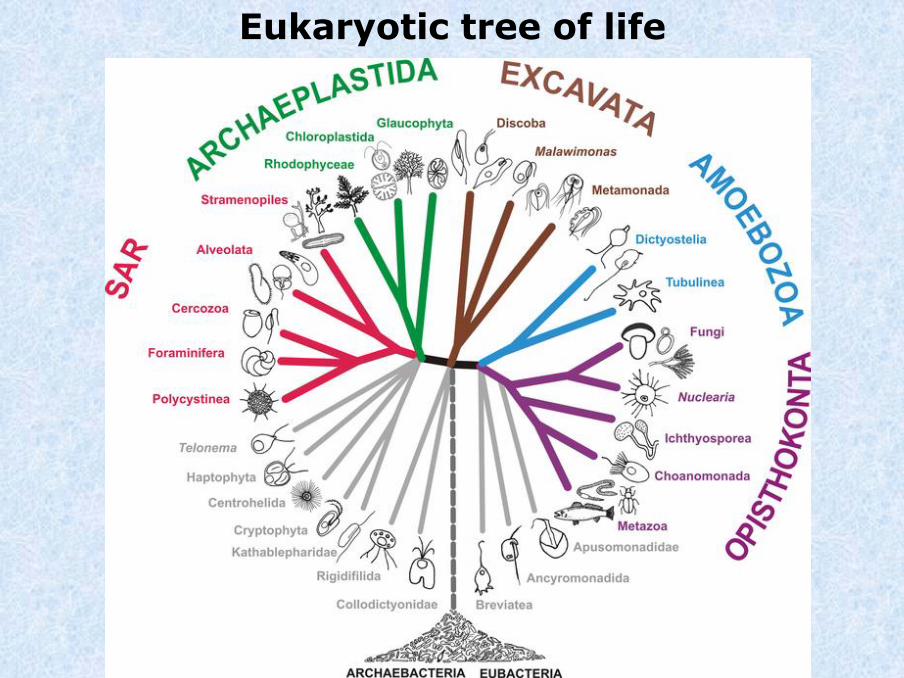
Eukaryotic tree of life
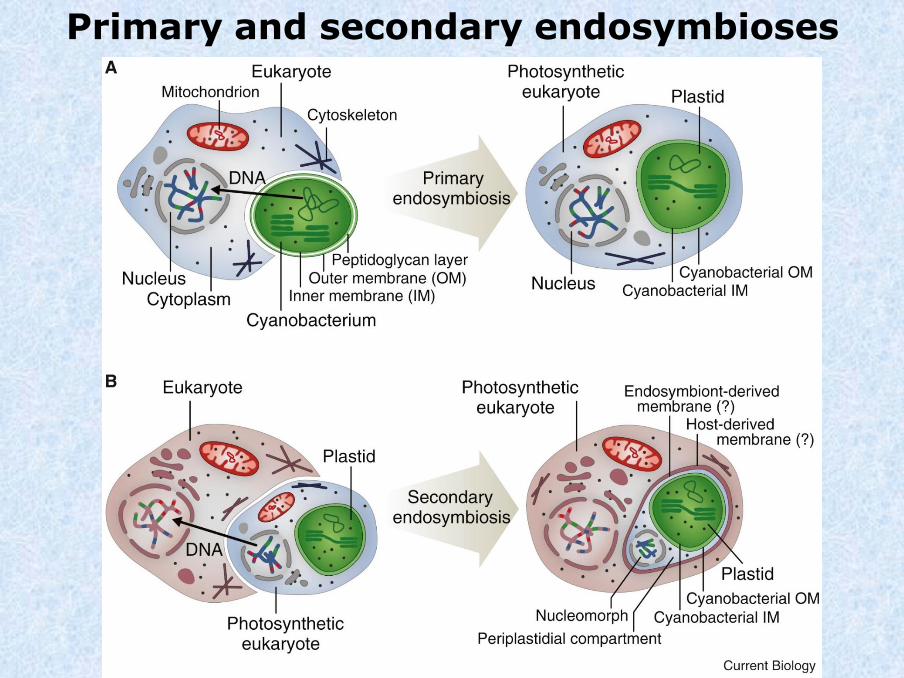
Primary and secondary endosymbioses
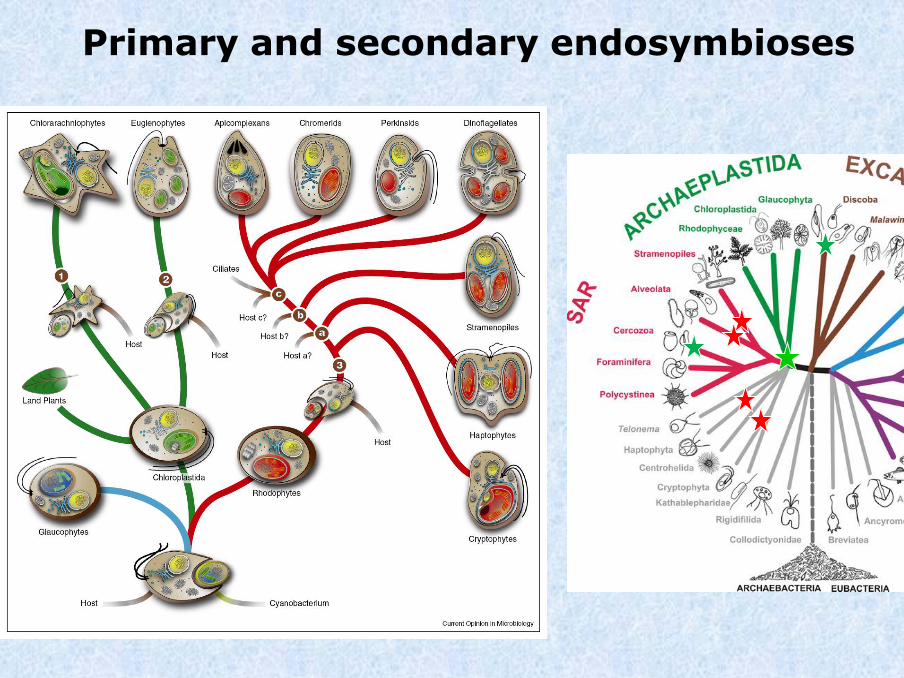
Primary and secondary endosymbioses

Primary and secondary endosymbioses
Stiller et al., 2014
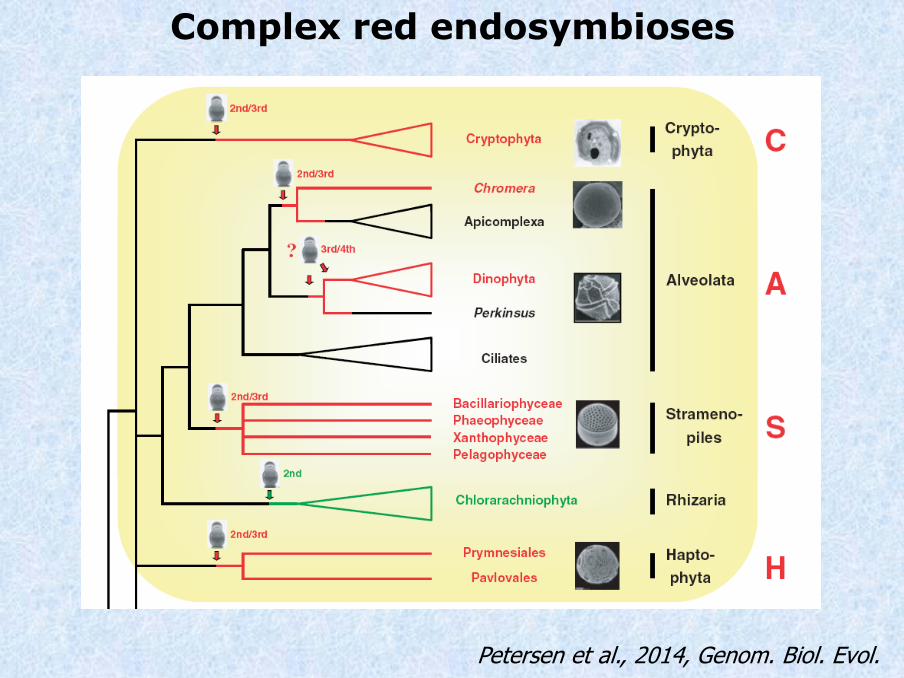
Complex red endosymbioses
Petersen et al., 2014, Genom. Biol. Evol.
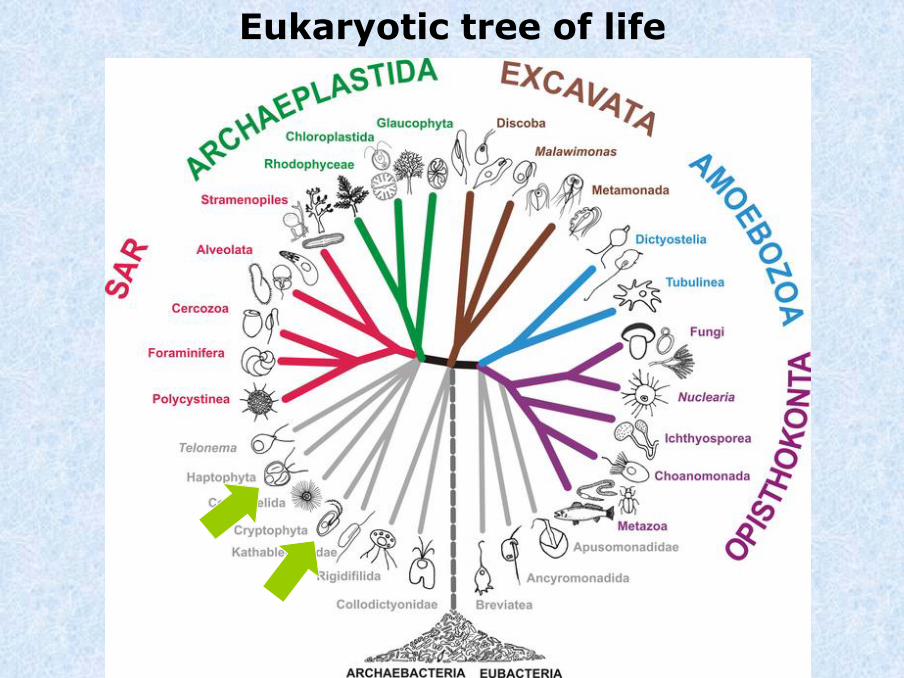
Eukaryotic tree of life
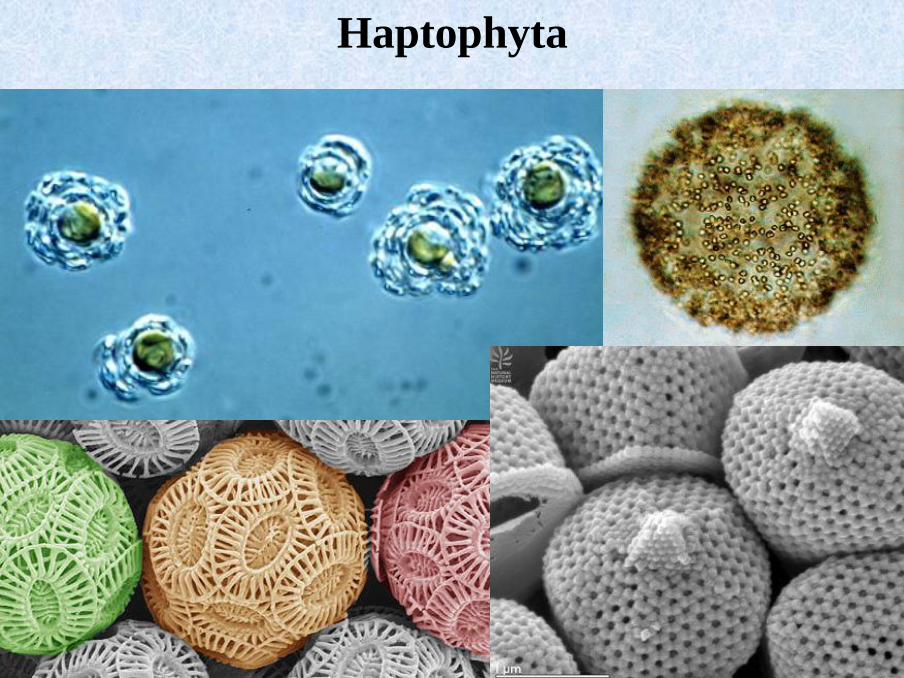

Haptophyta
▪ predominantly marine flagellates
▪ two flagella without mastigonemata
▪ haptonema – 6-7 MT in a cross-section
▪ plastids with lamellae comprised of 3 thylakoids, no stigma, no girdle lamella
▪ production of inorganic cellulosic or calcareous scales (coccoliths).

Satellite image of a bloom in the English Channel off the coast of Cornwall, 24 July 1999.
▪ significantly affecting the global clima on the Earth (biogeochemical cycle of carbon and sulphur)
Haptophyta
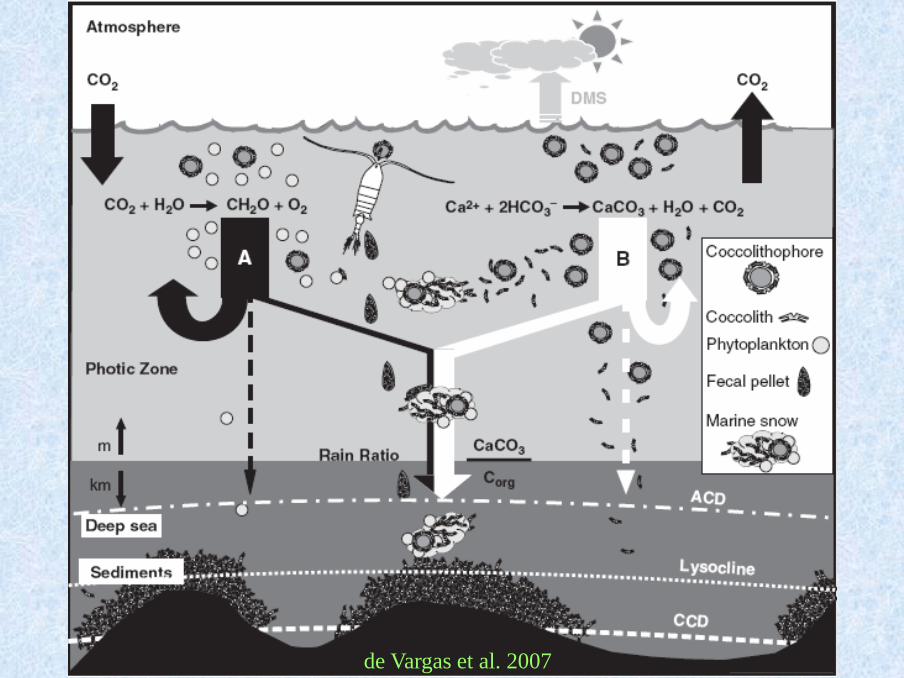
de Vargas et al. 2007
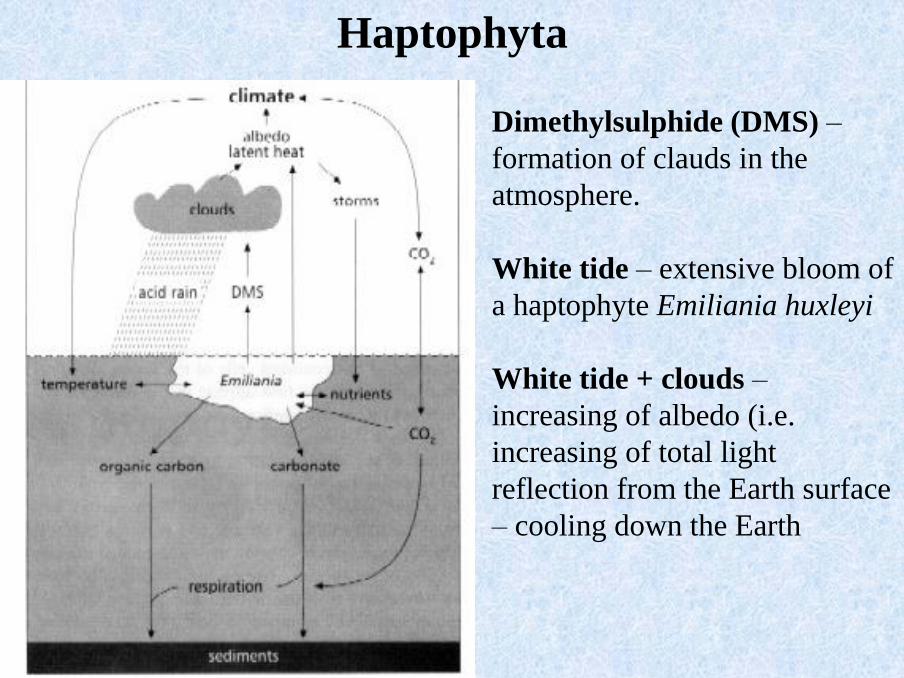
Dimethylsulphide (DMS) –
formation of clauds in the
atmosphere.
White tide – extensive bloom of
a haptophyte Emiliania huxleyi
White tide + clouds –
increasing of albedo (i.e.
increasing of total light
reflection from the Earth surface
– cooling down the Earth
Haptophyta
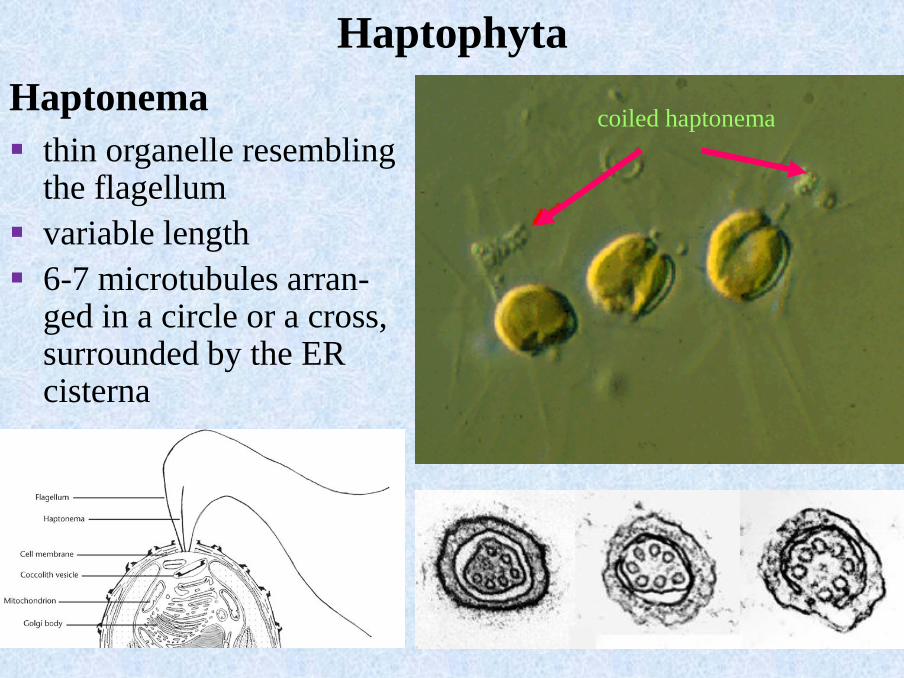
coiled haptonemaHaptonema
Haptophyta
▪ thin organelle resembling the flagellum
▪ variable length
▪ 6-7 microtubules arran-ged in a circle or a cross, surrounded by the ER cisterna
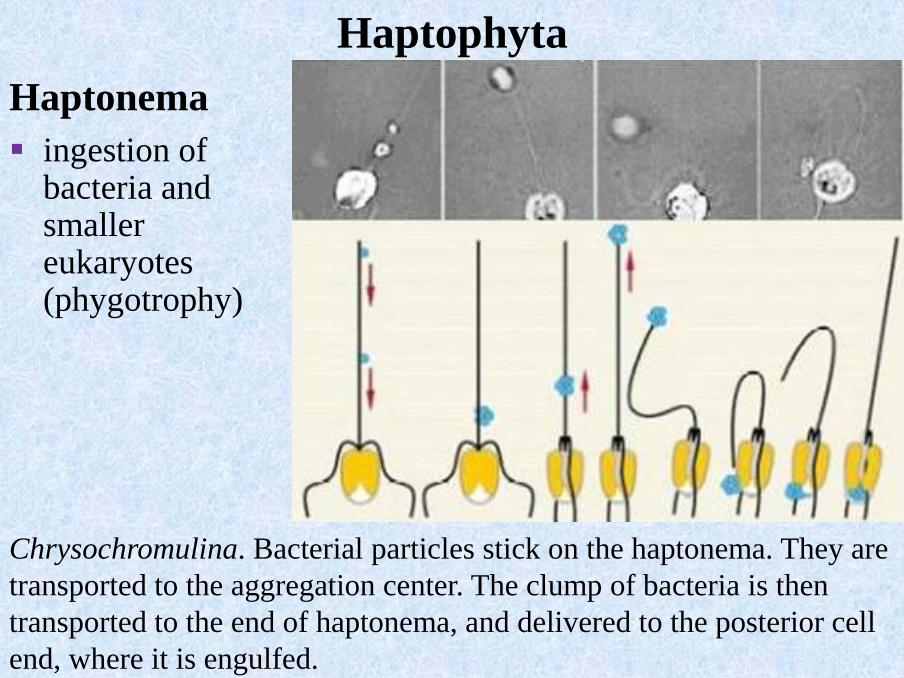
Chrysochromulina. Bacterial particles stick on the haptonema. They are
transported to the aggregation center. The clump of bacteria is then
transported to the end of haptonema, and delivered to the posterior cell
end, where it is engulfed.
Haptophyta
Haptonema
▪ ingestion ofbacteria and smaller eukaryotes(phygotrophy)
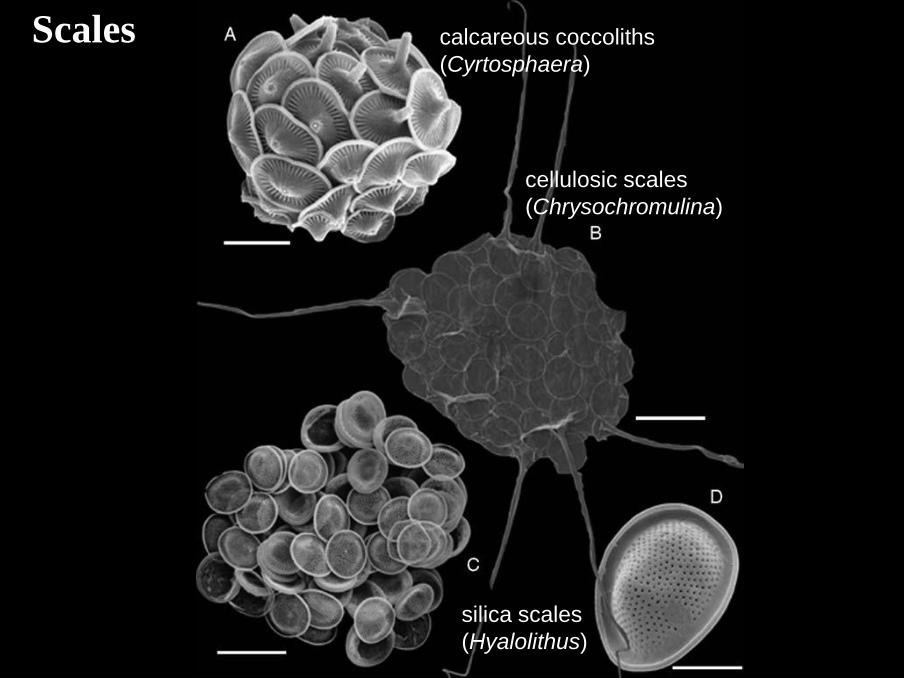
cellulosic scales
(Chrysochromulina)
calcareous coccoliths
(Cyrtosphaera)
silica scales
(Hyalolithus)
Scales

Prymnesium
Haptophyta - polysaccharide scales (cellulose)
Chrysochromulina sp.Ch. vexillifera
Tiny cellulosic scales formed in many
Haptophyte species
Chrysochromulina – only cellulosic
scales
Pleurochrysis carterae
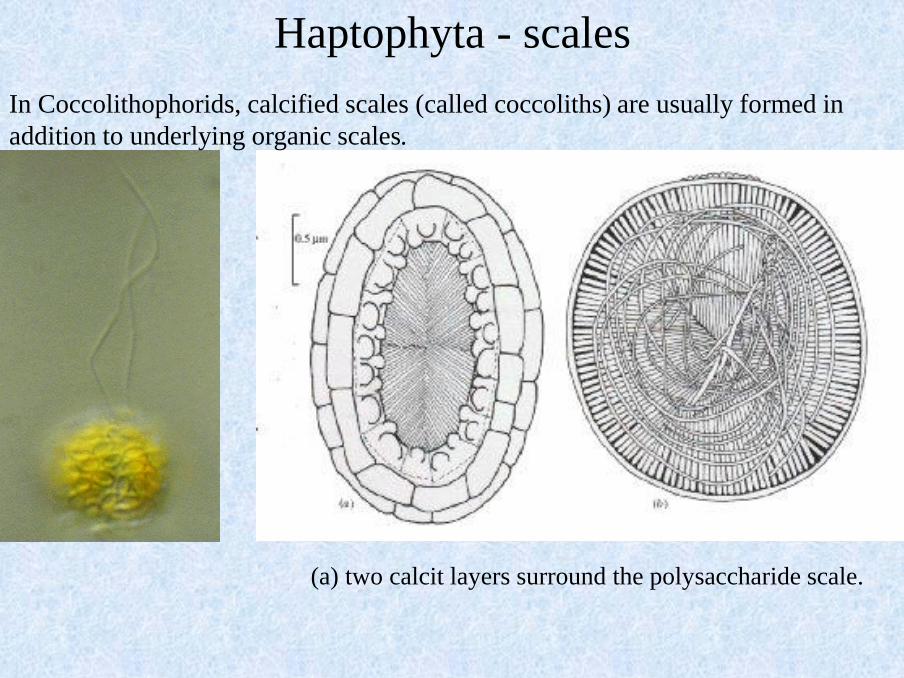
Haptophyta - scales
In Coccolithophorids, calcified scales (called coccoliths) are usually formed in
addition to underlying organic scales.
(a) two calcit layers surround the polysaccharide scale.
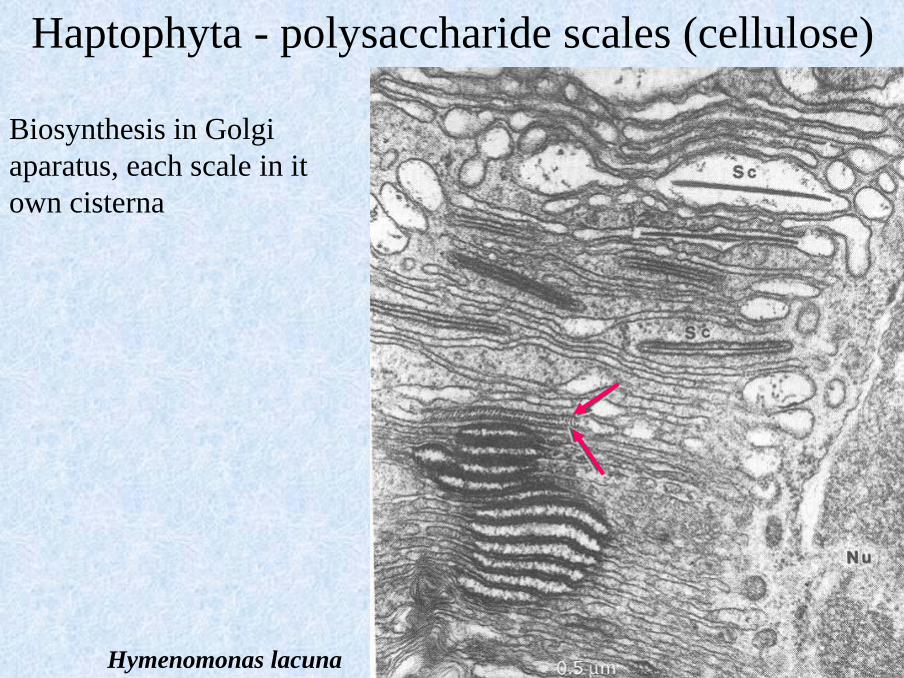
Hymenomonas lacuna
Biosynthesis in Golgi
aparatus, each scale in it
own cisterna
Haptophyta - polysaccharide scales (cellulose)
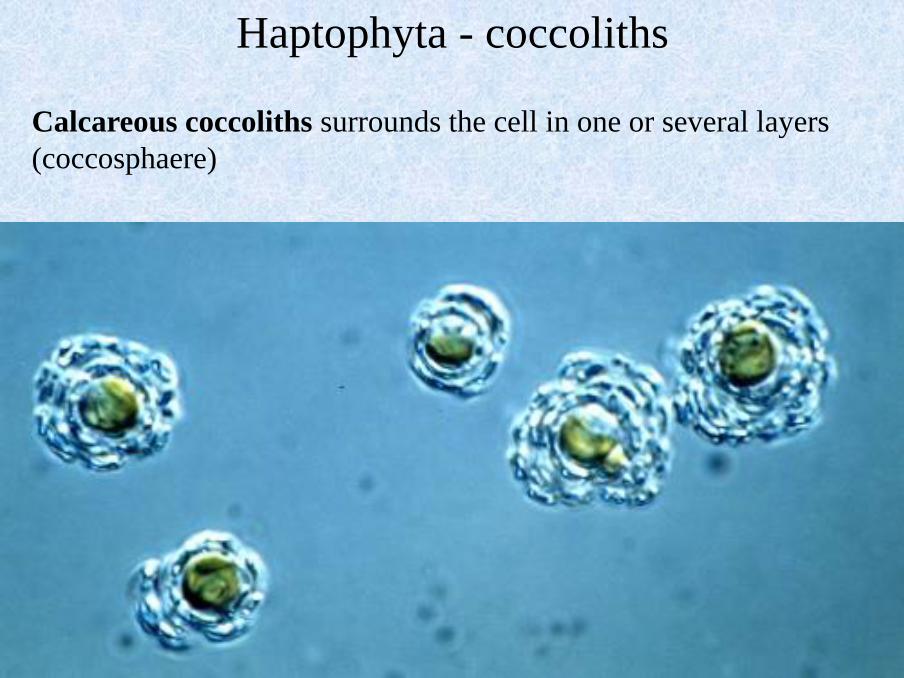
Calcareous coccoliths surrounds the cell in one or several layers
(coccosphaere)
Haptophyta - coccoliths
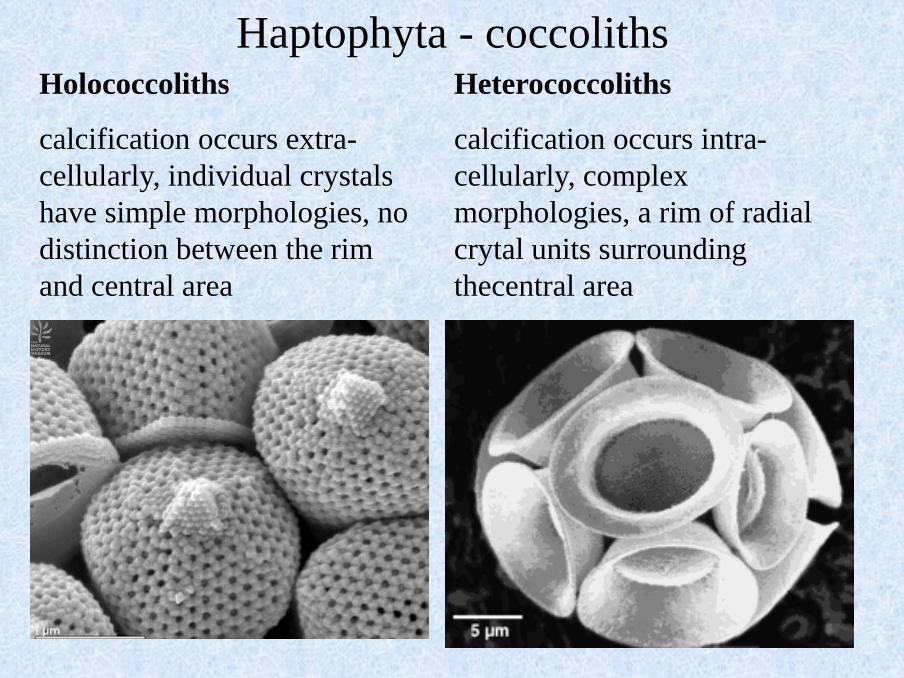
Holococcoliths
calcification occurs extra-
cellularly, individual crystals
have simple morphologies, no
distinction between the rim
and central area
Heterococcoliths
calcification occurs intra-
cellularly, complex
morphologies, a rim of radial
crytal units surrounding
thecentral area
Haptophyta - coccoliths
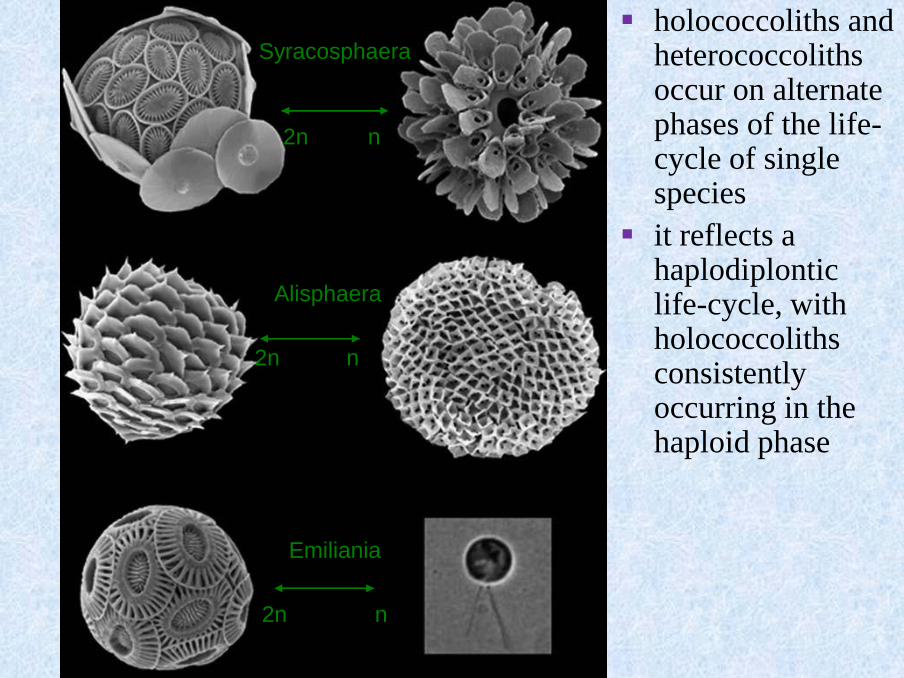
Syracosphaera
n2n
n2n
n2n
Alisphaera
Emiliania
▪ holococcoliths and heterococcoliths occur on alternate phases of the life-cycle of single species
▪ it reflects a haplodiplontic life-cycle, with holococcoliths consistently occurring in the haploid phase
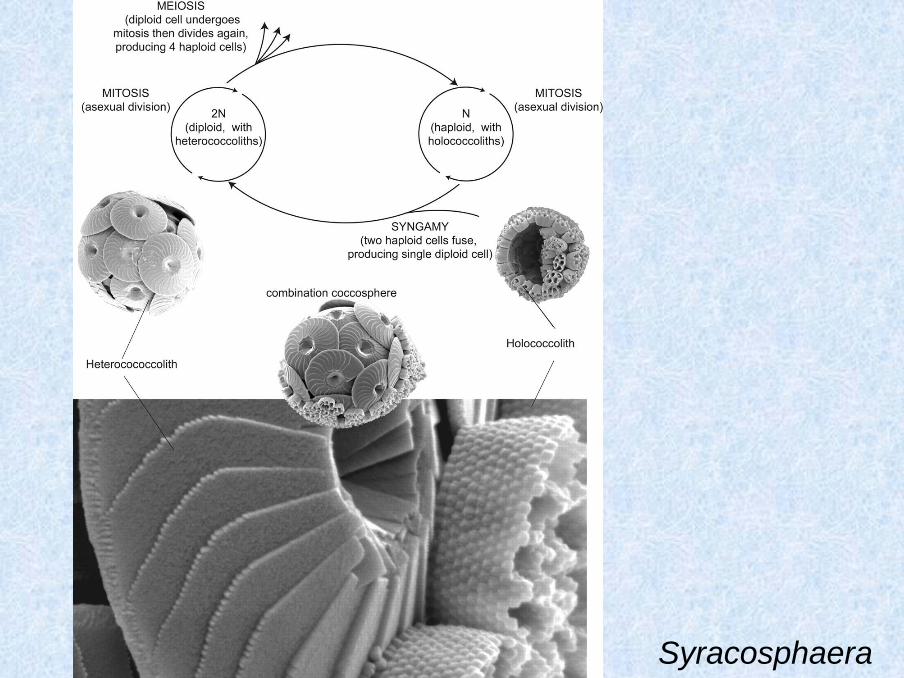
Syracosphaera
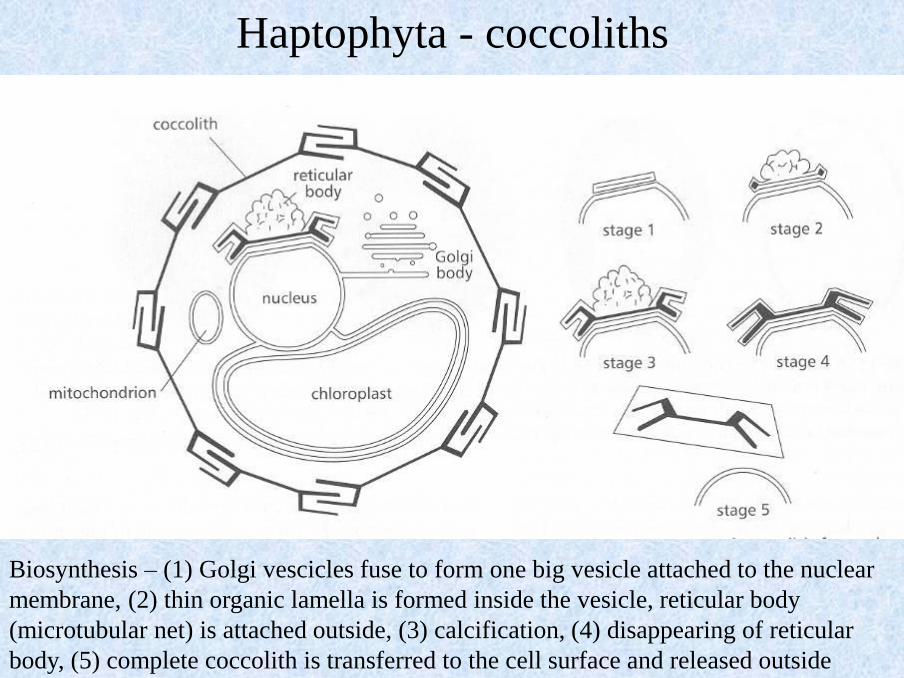
Biosynthesis – (1) Golgi vescicles fuse to form one big vesicle attached to the nuclear
membrane, (2) thin organic lamella is formed inside the vesicle, reticular body
(microtubular net) is attached outside, (3) calcification, (4) disappearing of reticular
body, (5) complete coccolith is transferred to the cell surface and released outside
Haptophyta - coccoliths
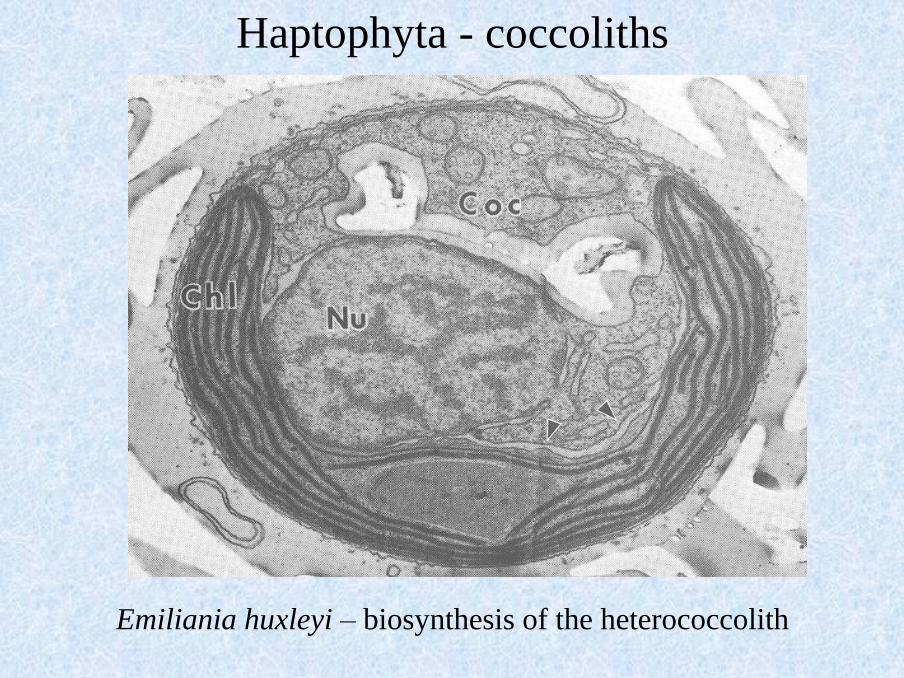
Emiliania huxleyi – biosynthesis of the heterococcolith
Haptophyta - coccoliths

Scyphosphaera

Biosynthesis (E. huxleyi)
Haptophyta - coccoliths
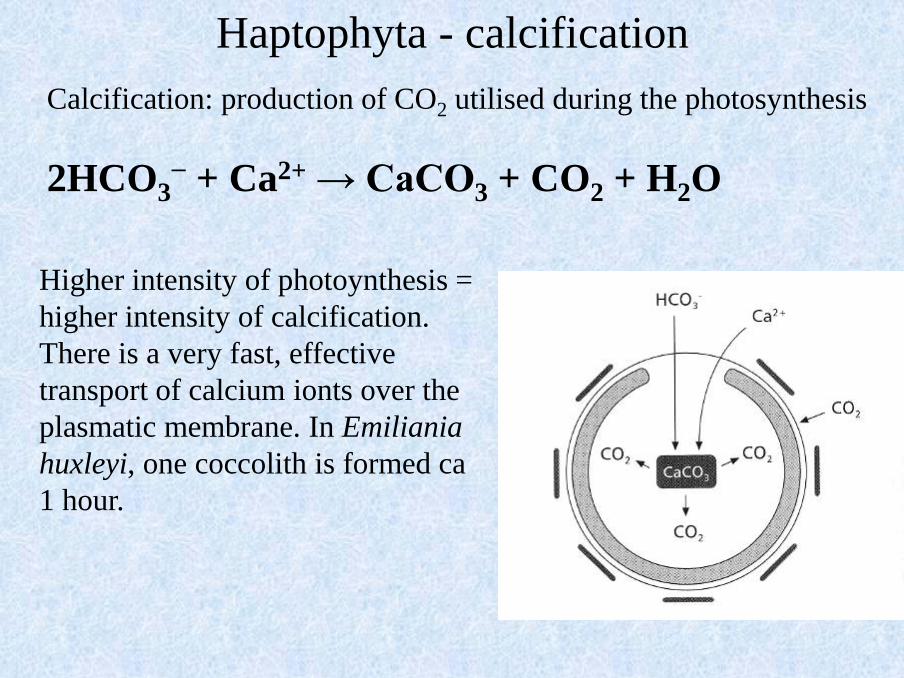
Calcification: production of CO2 utilised during the photosynthesis
2HCO3− + Ca2+ → CaCO3 + CO2 + H2O
Higher intensity of photoynthesis =
higher intensity of calcification.
There is a very fast, effective
transport of calcium ionts over the
plasmatic membrane. In Emiliania
huxleyi, one coccolith is formed ca
1 hour.
Haptophyta - calcification

Haptophyta – value of coccoliths
▪ utilisation of CO2 for the photosynthesis
▪ defence against the predators (higher cell volume) and pathogens (bacteria, viruses)
▪ regulation of floating (production of heavy coccoliths)
▪ diffraction of sunlight into the cell center (photosynthesis in deap-sea species)
Chrysochromulina
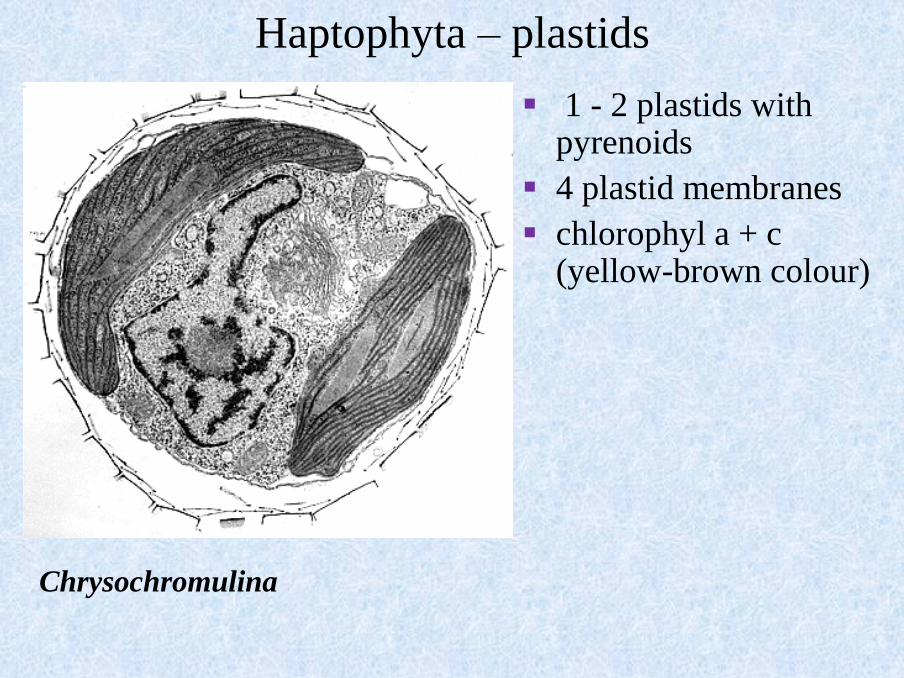
Haptophyta – plastids
▪ 1 - 2 plastids with pyrenoids
▪ 4 plastid membranes
▪ chlorophyl a + c (yellow-brown colour)

Calyptrosphaera
sphaeroidea - pyrenoid
Haptophyta – plastids

Chrysochromulina polylepis ichtyotoxins
Haptophyta – production of toxins
White Cliffs of Dover
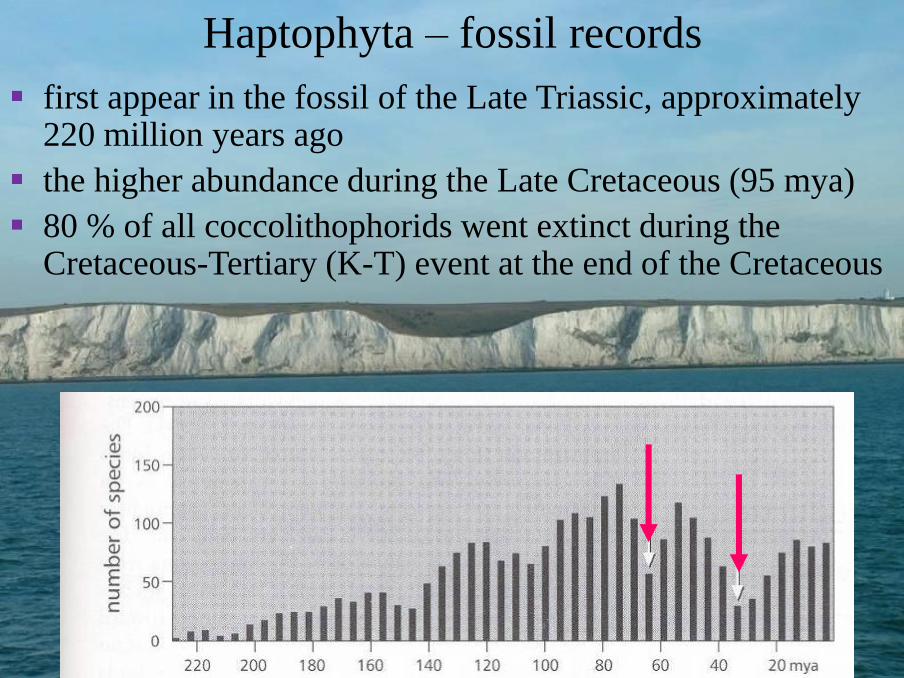
Haptophyta – fossil records
▪ first appear in the fossil of the Late Triassic, approximately 220 million years ago
▪ the higher abundance during the Late Cretaceous (95 mya)
▪ 80 % of all coccolithophorids went extinct during the Cretaceous-Tertiary (K-T) event at the end of the Cretaceous
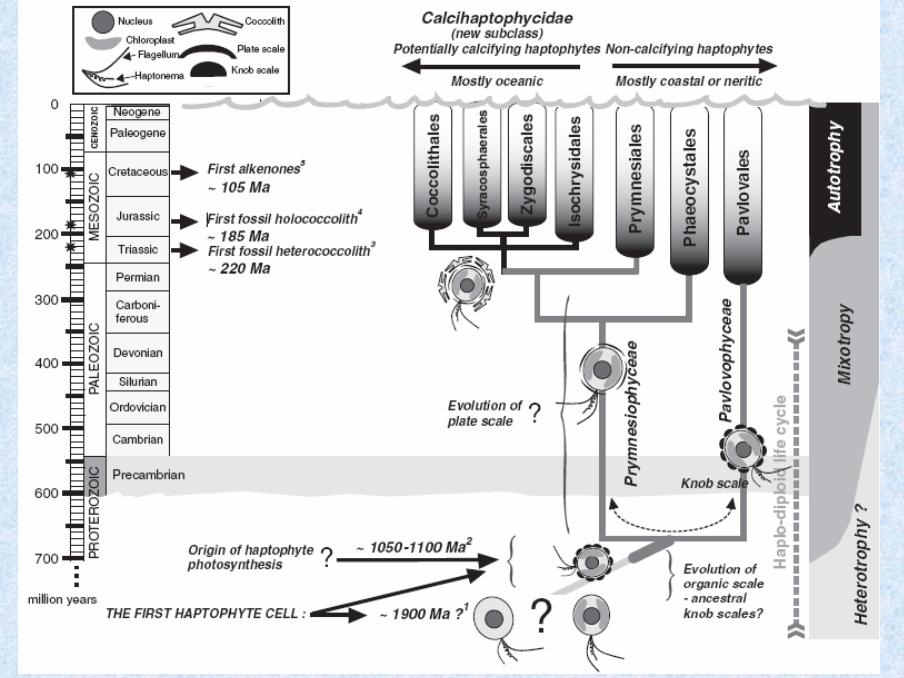
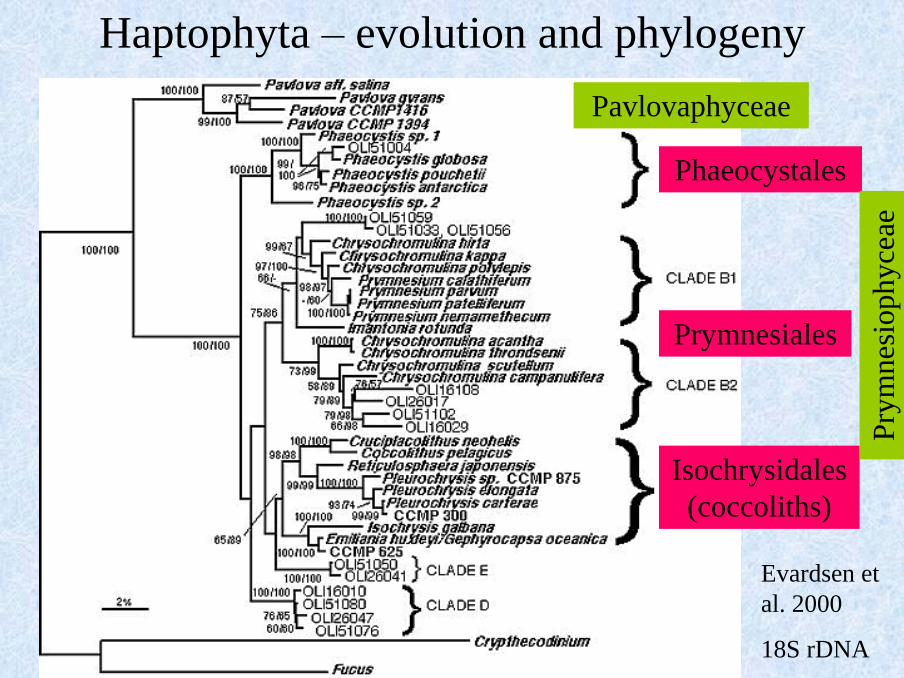
Evardsen et
al. 2000
18S rDNA
Pavlovaphyceae
Phaeocystales
Prymnesiales
Isochrysidales
(coccoliths)
Haptophyta – evolution and phylogeny
Pry
mn
esio
phy
ceae
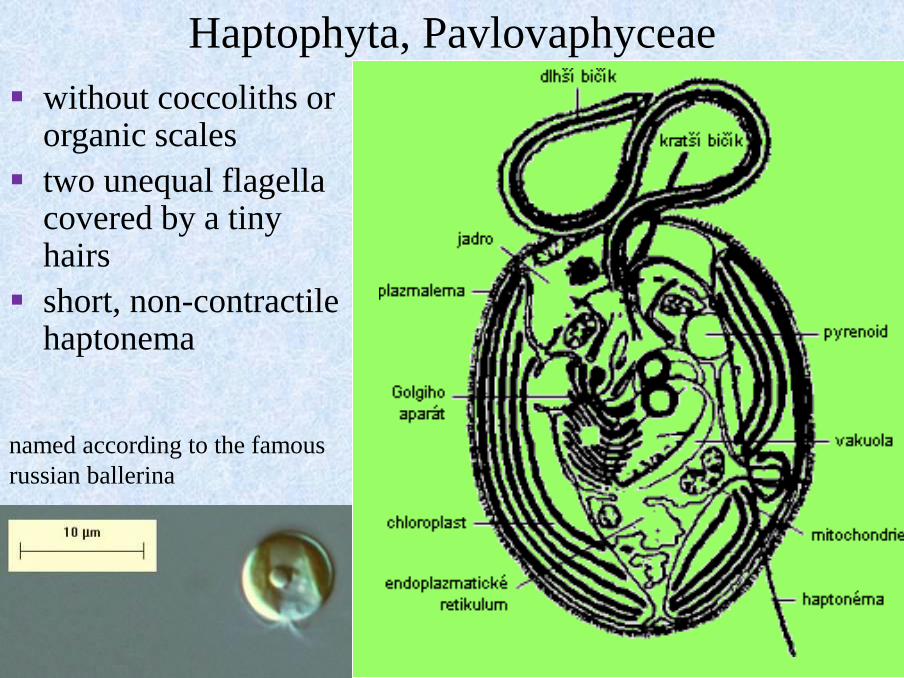
Haptophyta, Pavlovaphyceae
▪ without coccoliths or organic scales
▪ two unequal flagella covered by a tiny hairs
▪ short, non-contractile haptonema
nákres Pavlova
named according to the famous
russian ballerina
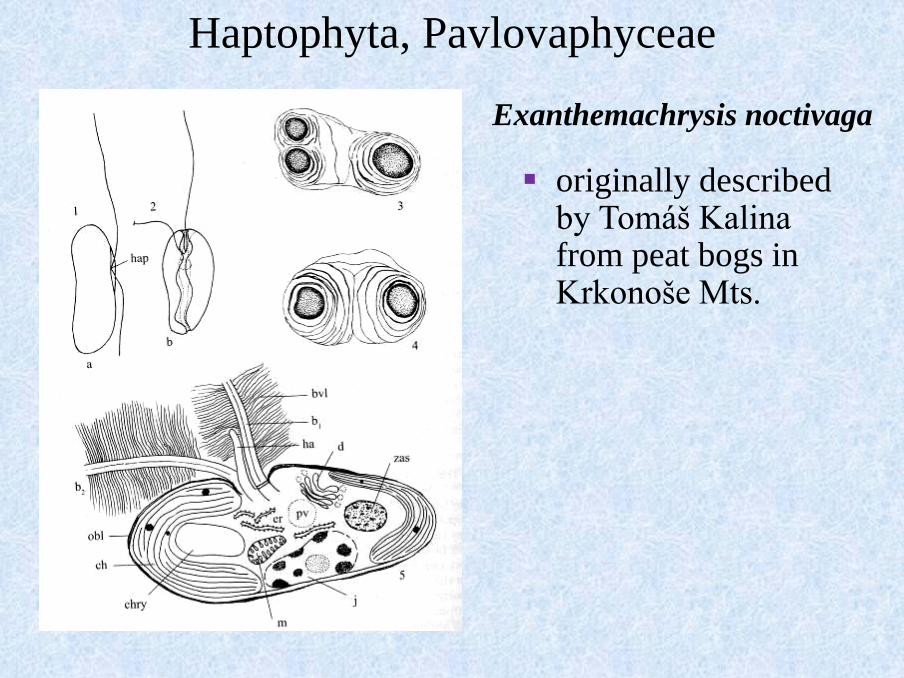
Exanthemachrysis noctivaga
Haptophyta, Pavlovaphyceae
▪ originally described by Tomáš Kalina from peat bogs in Krkonoše Mts.
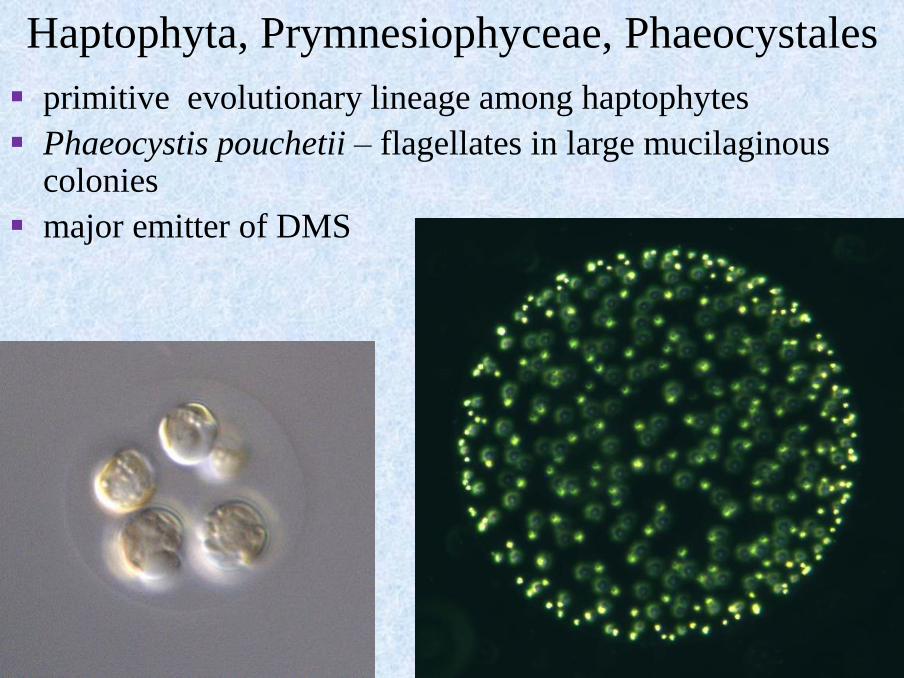
Haptophyta, Prymnesiophyceae, Phaeocystales
▪ primitive evolutionary lineage among haptophytes
▪ Phaeocystis pouchetii – flagellates in large mucilaginous colonies
▪ major emitter of DMS

Phaeocystis bloom
▪ foam accumulated on beaches
Chrysochromulina
Prymnesium
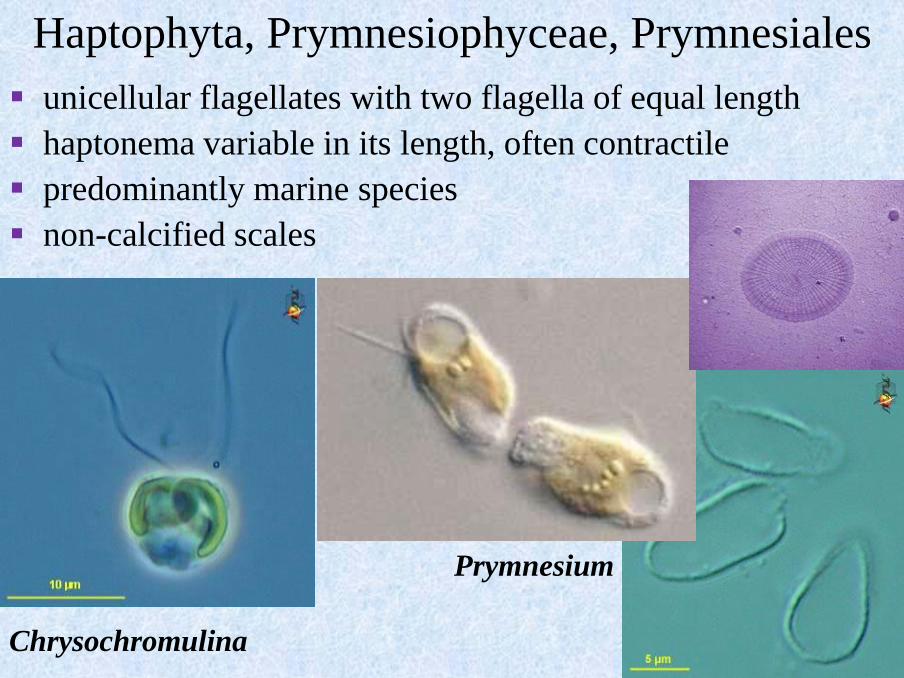
Haptophyta, Prymnesiophyceae, Prymnesiales
▪ unicellular flagellates with two flagella of equal length
▪ haptonema variable in its length, often contractile
▪ predominantly marine species
▪ non-calcified scales
Pleurochrysis carterae
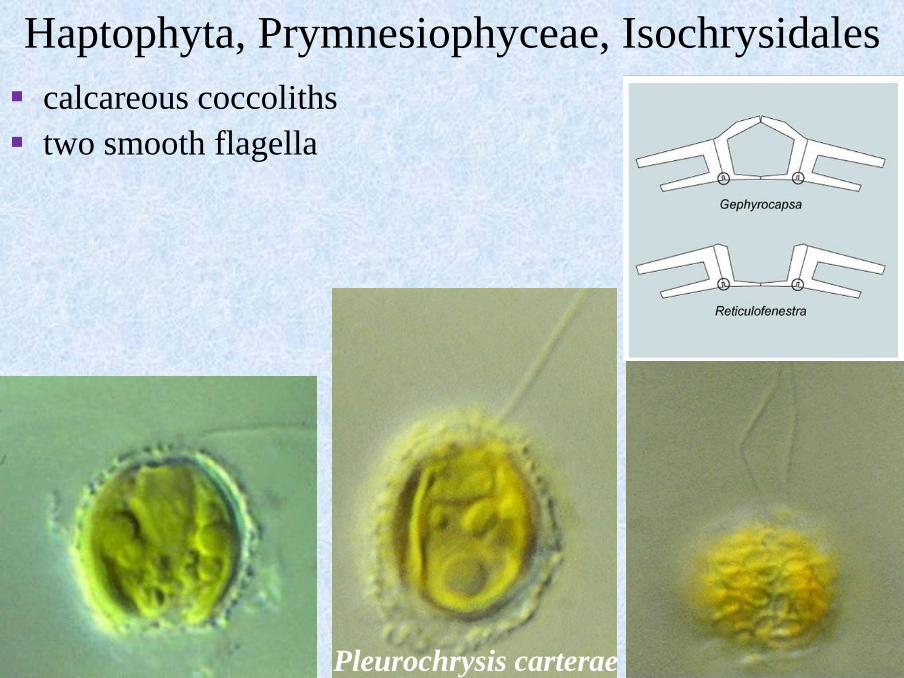
Haptophyta, Prymnesiophyceae, Isochrysidales
▪ calcareous coccoliths
▪ two smooth flagella

Pleurochrysis
▪ simple coccoliths
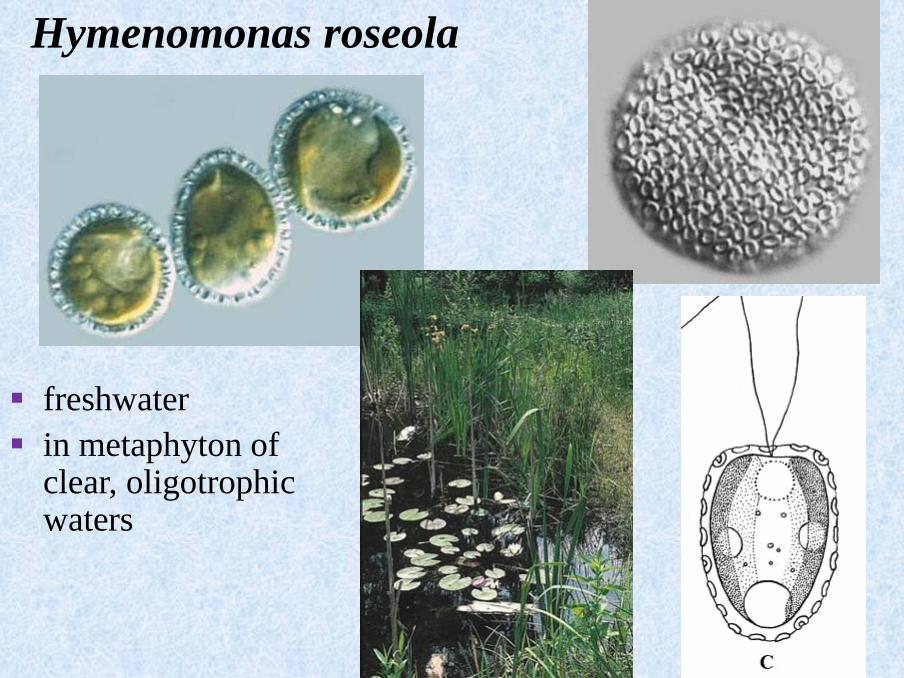
Hymenomonas roseola
▪ freshwater
▪ in metaphyton of clear, oligotrophic waters
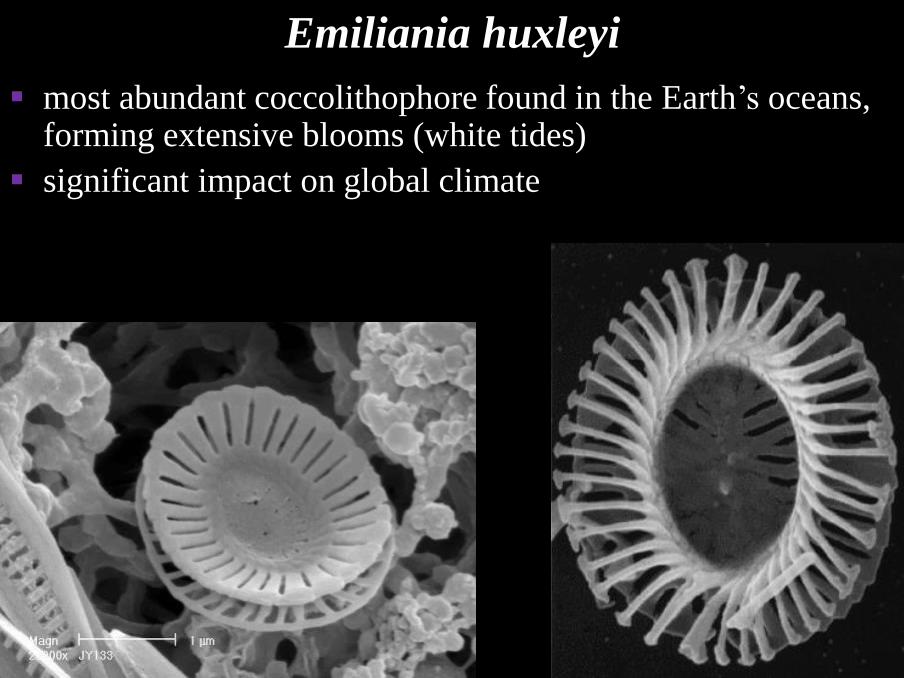
Emiliania huxleyi
▪ most abundant coccolithophore found in the Earth’s oceans, forming extensive blooms (white tides)
▪ significant impact on global climate
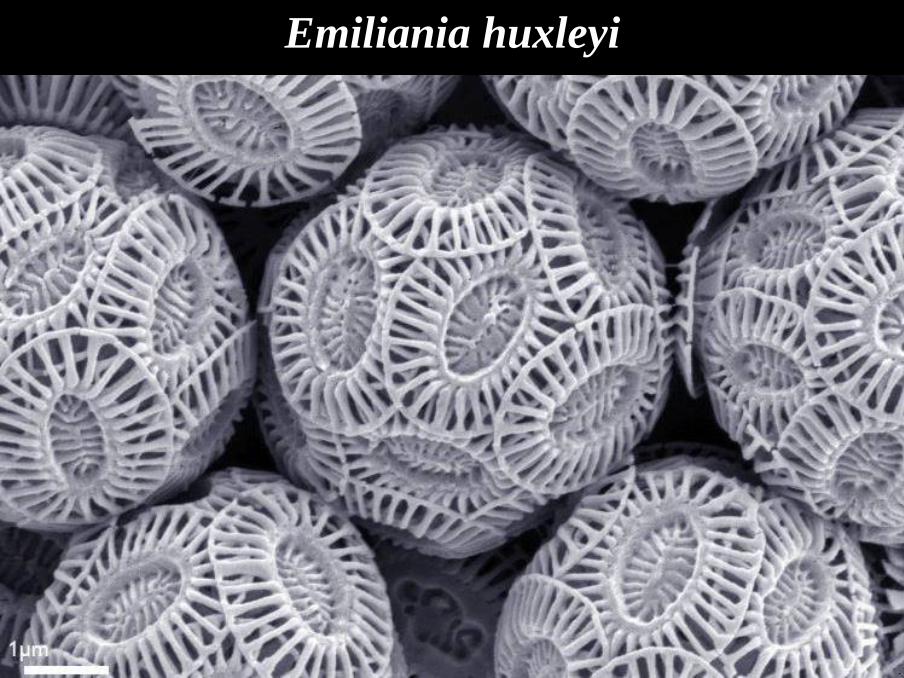
Emiliania huxleyi

Emiliania huxleyi

http://ina.tmsoc.org/Nannotax3/index.ph
p?dir=Coccolithophores&top=34&base=
300

Algirosphaera robusta
Thorosphaera
flabellata
Pontosphaera
syracusana
Isochrysidales - coccoliths

Florisphaera profunda
Braarudosphaera
bigelowi
Rhabdosphaera
clavigera
Isochrysidales - coccolithsF. profunda


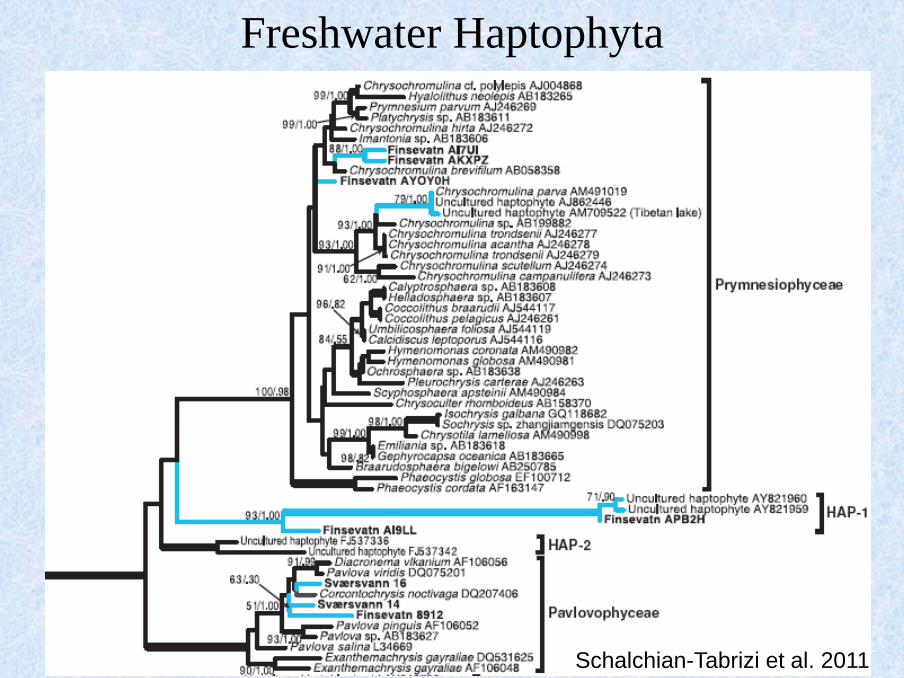
Freshwater Haptophyta
Schalchian-Tabrizi et al. 2011
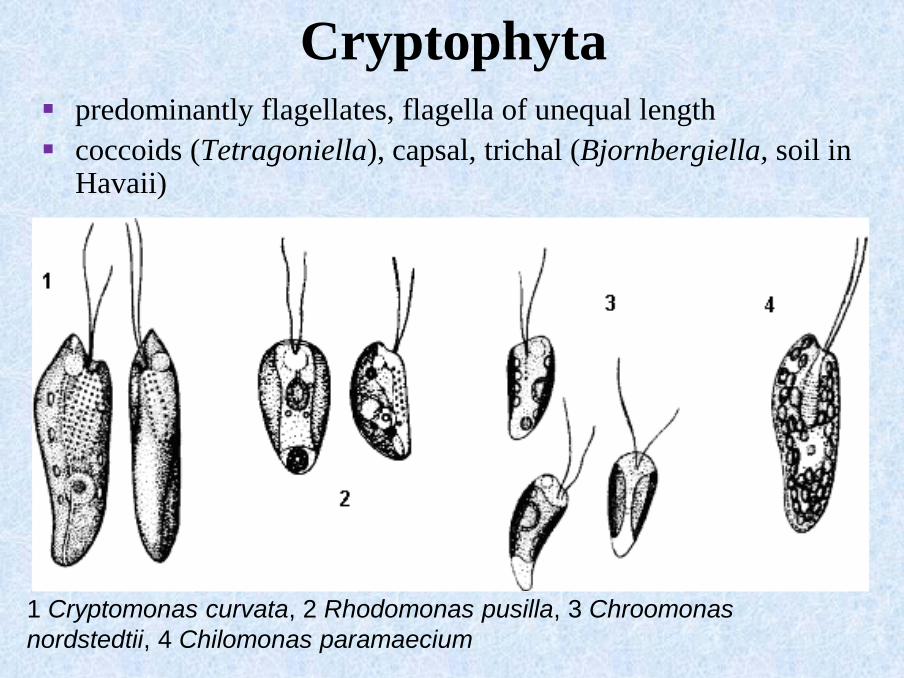
Cryptophyta▪ predominantly flagellates, flagella of unequal length
▪ coccoids (Tetragoniella), capsal, trichal (Bjornbergiella, soil in Havaii)
1 Cryptomonas curvata, 2 Rhodomonas pusilla, 3 Chroomonas
nordstedtii, 4 Chilomonas paramaecium
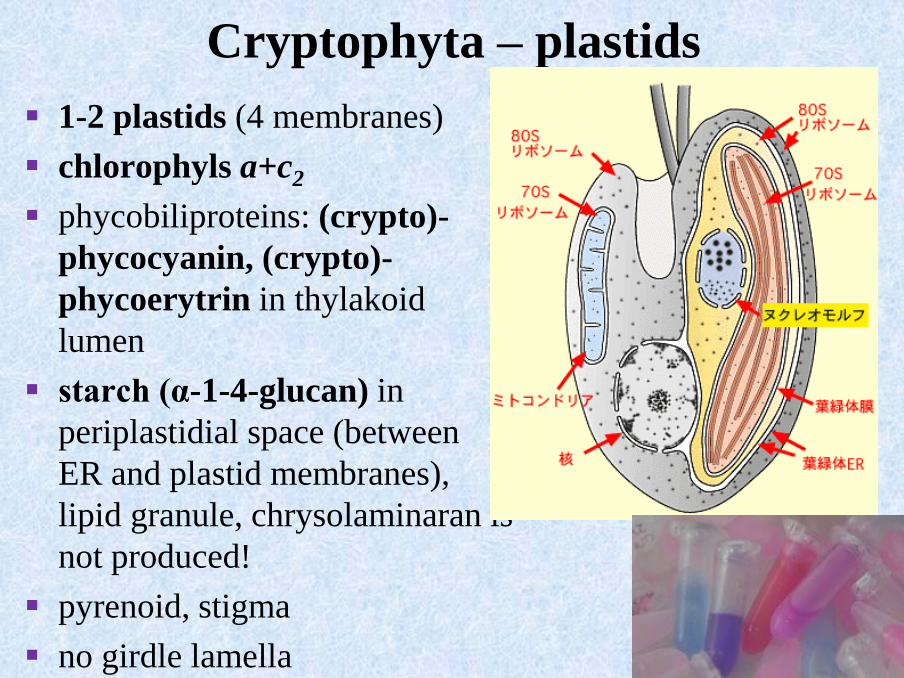
Cryptophyta – plastids
▪ 1-2 plastids (4 membranes)
▪ chlorophyls a+c2
▪ phycobiliproteins: (crypto)-
phycocyanin, (crypto)-
phycoerytrin in thylakoid
lumen
▪ starch (α-1-4-glucan) in
periplastidial space (between
ER and plastid membranes),
lipid granule, chrysolaminaran is
not produced!
▪ pyrenoid, stigma
▪ no girdle lamella
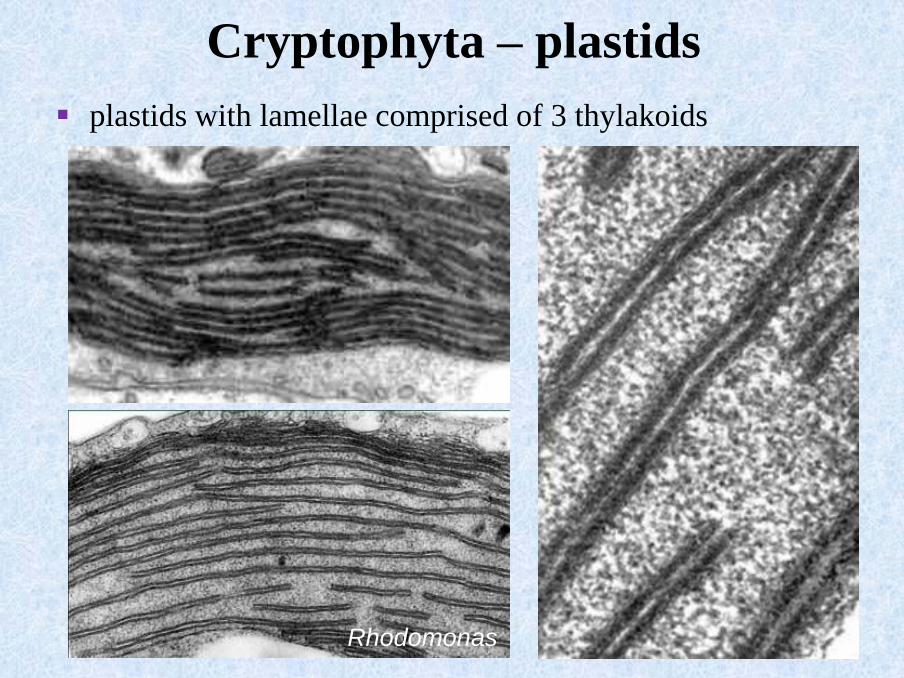
Rhodomonas
▪ plastids with lamellae comprised of 3 thylakoids
Cryptophyta – plastids
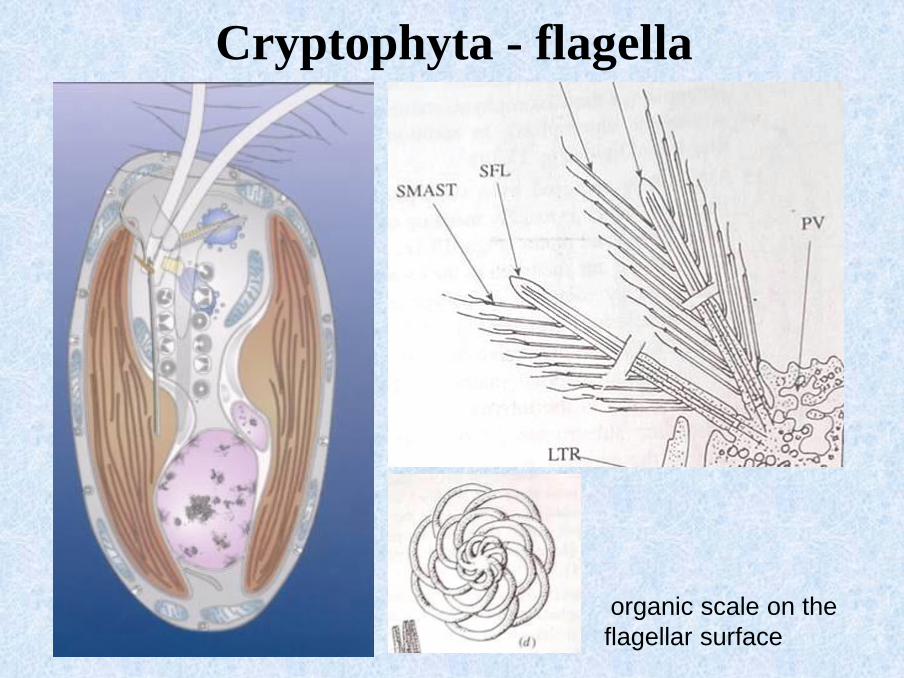
Cryptophyta - flagella
organic scale on the
flagellar surface
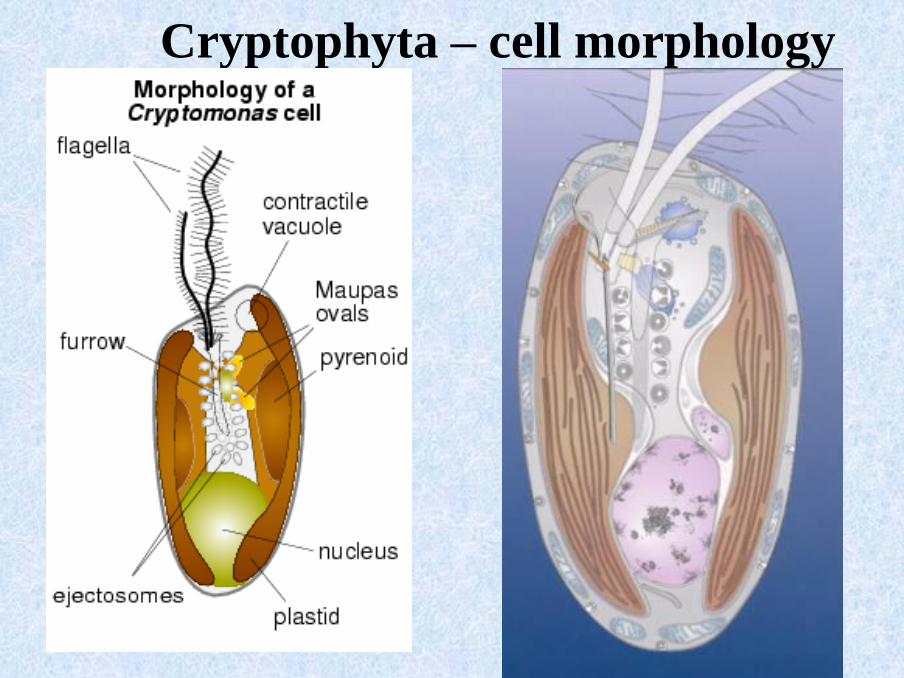
Cryptophyta – cell morphology

Cryptophyta - ejectosomes
▪ explosive structures surrounding the furrow, and under theplasmatic membrane
▪ two connected ribbons of different sizes, that are rolled up and under tension
▪ discharged upon mechanical or chemical stress irritating the cells

Cryptophyta – nucleomorph
▪ often attached to pyrenoids
▪ two membranes
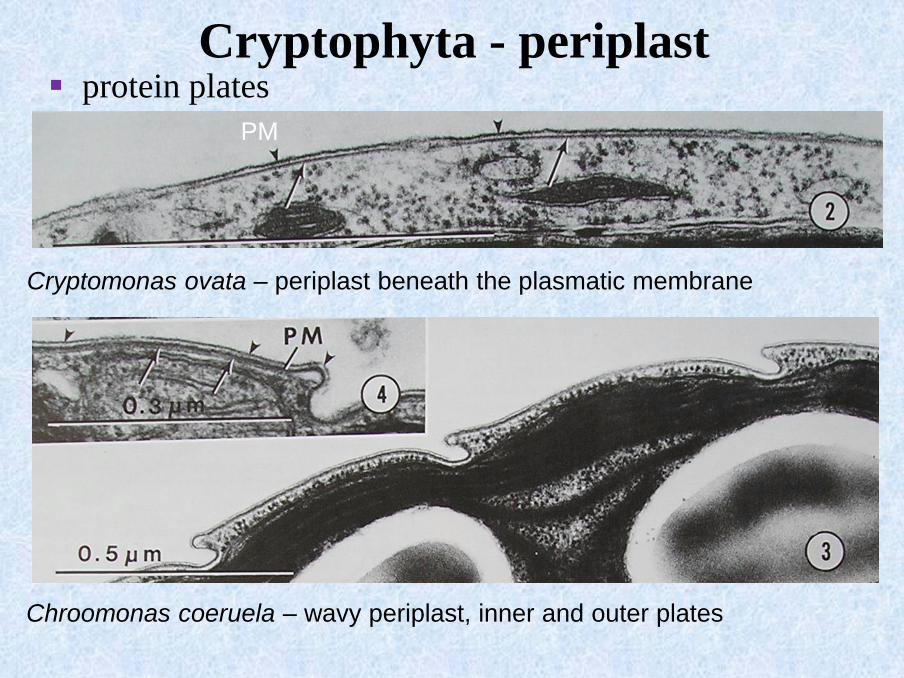
Chroomonas coeruela – wavy periplast, inner and outer plates
Cryptophyta - periplast
Cryptomonas ovata – periplast beneath the plasmatic membrane
PM
▪ protein plates
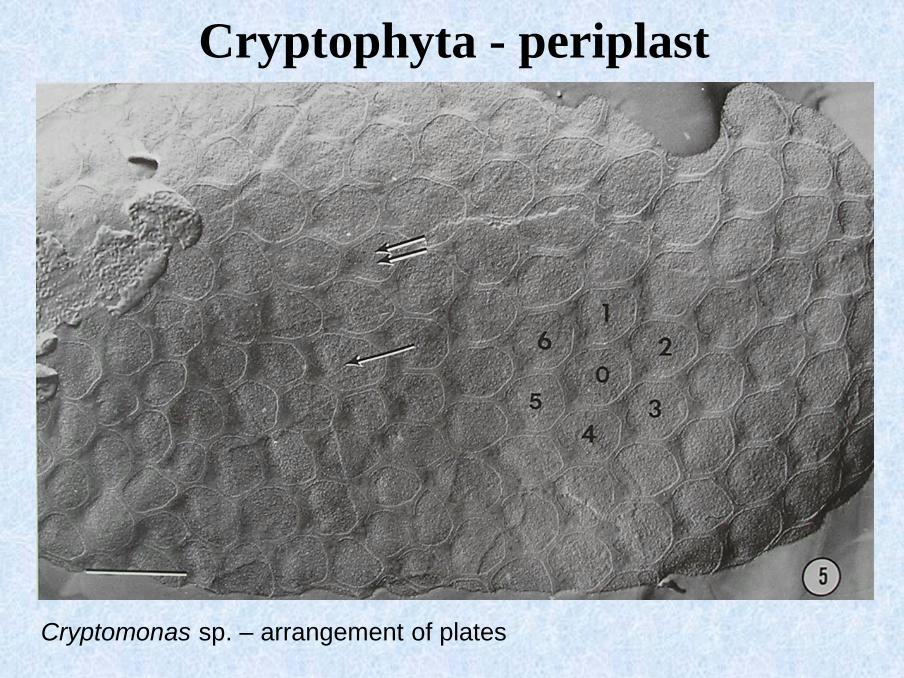
Cryptomonas sp. – arrangement of plates
Cryptophyta - periplast
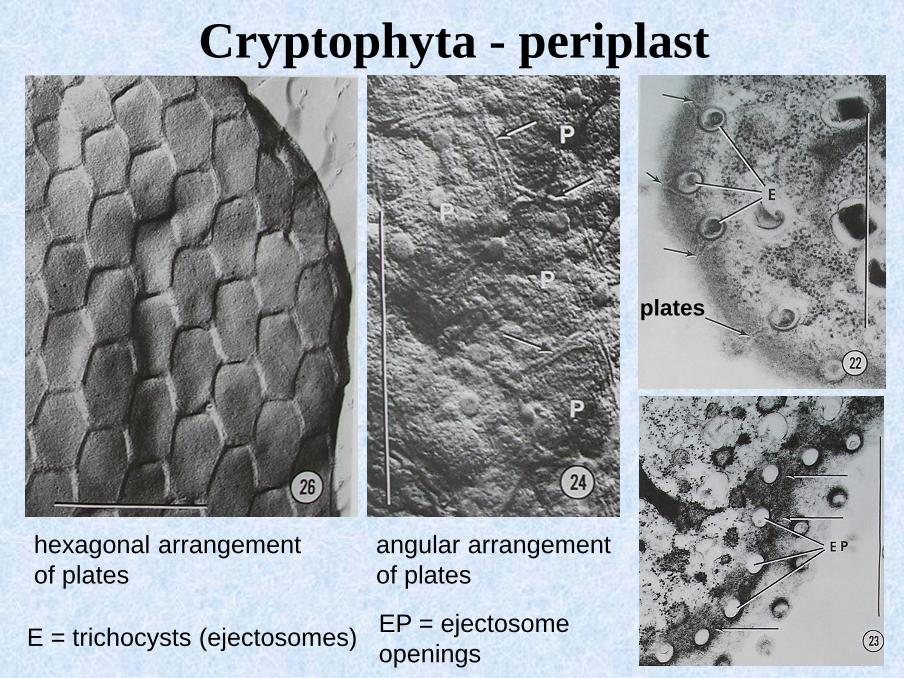
angular arrangement
of plates
plates
E = trichocysts (ejectosomes)EP = ejectosome
openings
hexagonal arrangement
of plates
Cryptophyta - periplast

Cryptophyta - reproduction
▪ asexual – schizotomy of flagellates
▪ sexual – isogamy, formation of planozygote (4 flagella)
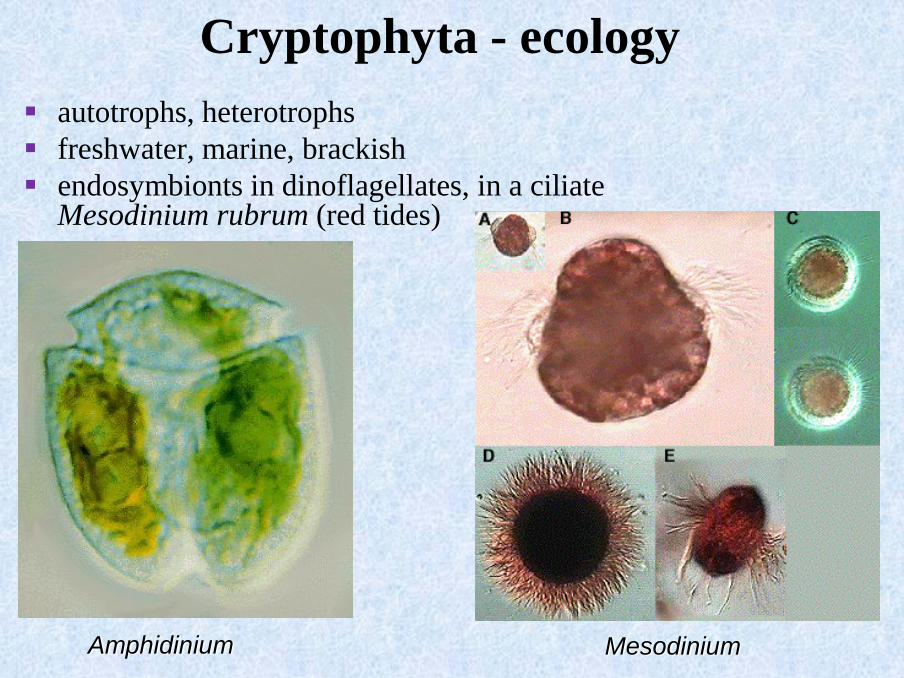
Cryptophyta - ecology
▪ autotrophs, heterotrophs
▪ freshwater, marine, brackish
▪ endosymbionts in dinoflagellates, in a ciliate Mesodinium rubrum (red tides)
Amphidinium Mesodinium

Cryptophyta - systematics
▪ ca 20 genera, half of them in freshwater
▪ molecular data still unknown for many genera = traditional
systematics
▪ Cryptomonadales – free-living flagellates, genera delimited
by the colour of plastids
▪ olive-brown: Cryptomonas, Campylomonas
▪ blue-green: Chroomonas
▪ red-green: Rhodomonas
▪ colourless (loss of plastids): Chilomonas

CryptomonasCryptomonas reflexa Cryptomonas rostratiformis
Cryptomonas sp.
Cryptomonas acuta
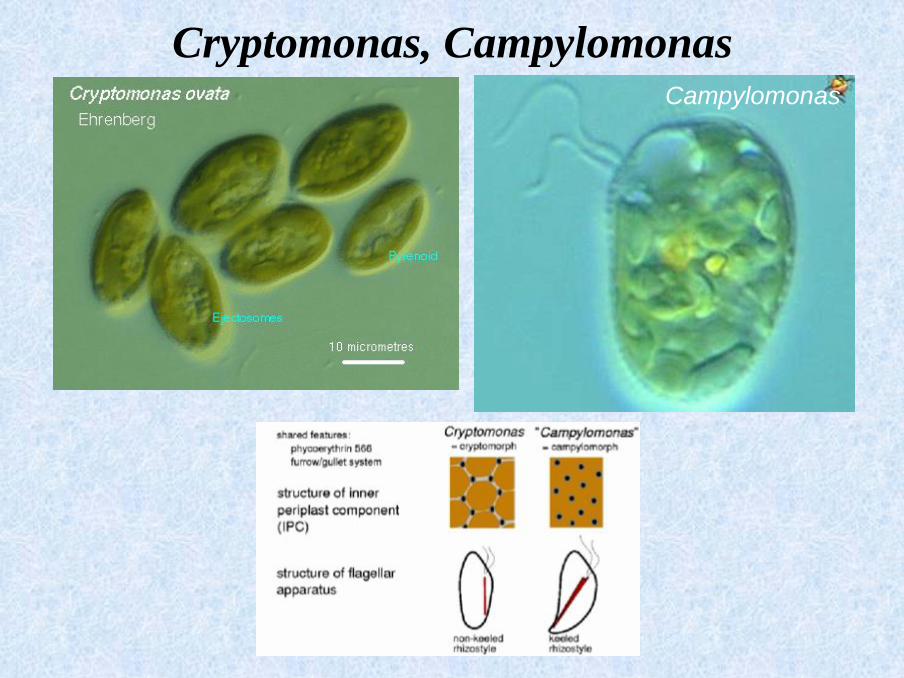
Cryptomonas, CampylomonasCampylomonas
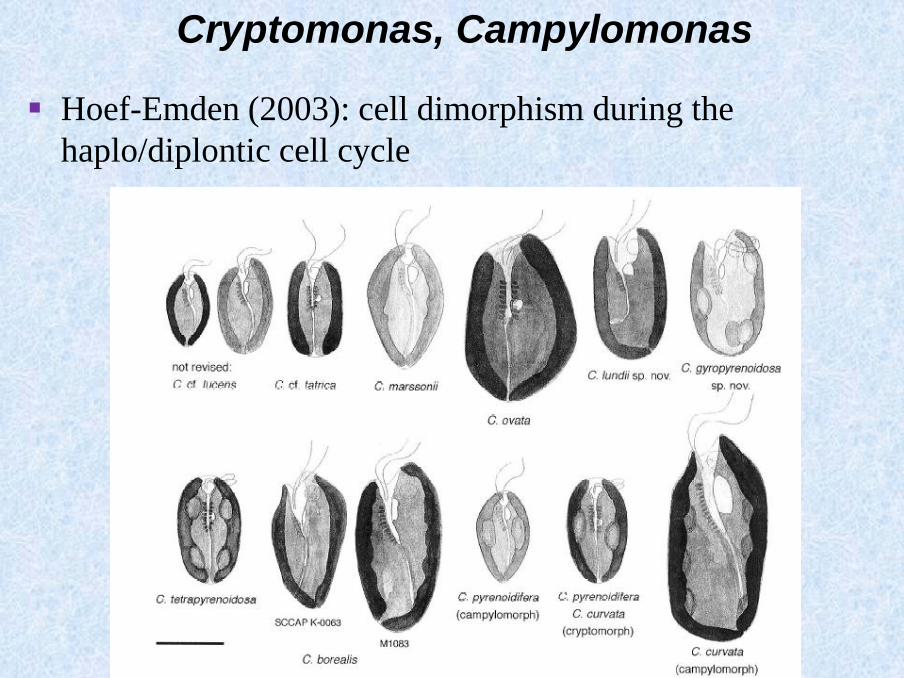
Cryptomonas, Campylomonas
▪ Hoef-Emden (2003): cell dimorphism during the
haplo/diplontic cell cycle
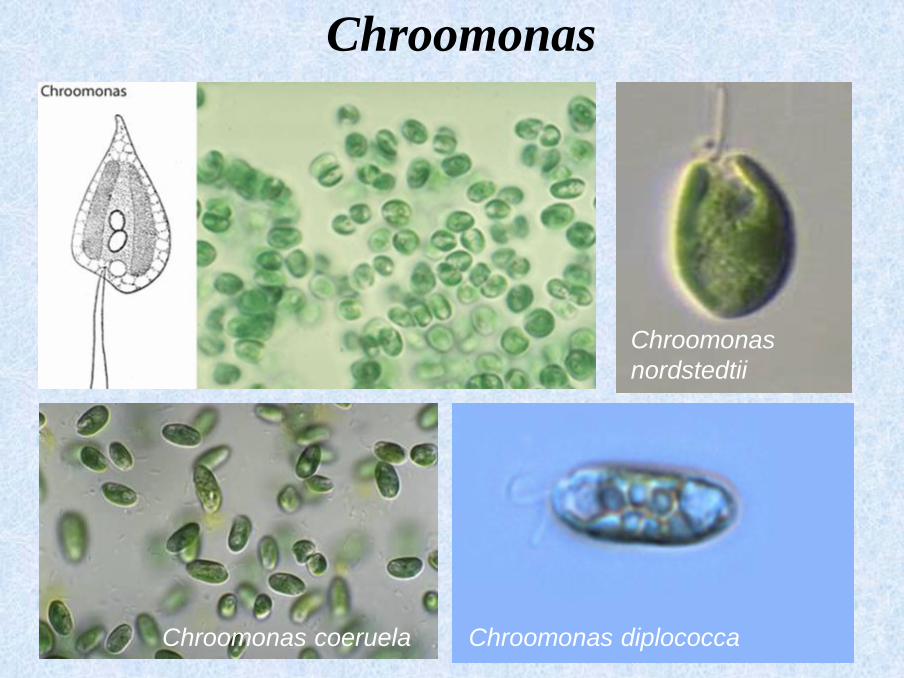
Chroomonas coeruela
Chroomonas
nordstedtii
Chroomonas diplococca
Chroomonas

RhodomonasRhodomonas baltica
Rhodomonas duplex
Rhodomonas cf. marina (Balt)

Chilomonas – heterotrophic
Chilomonas paramecium

Hemiselmis – marine

Cryptophyta - molecular data
▪ phylogenetic data do not correspond with the traditional
morfological features - needed revision of the genus
Cryptomonas
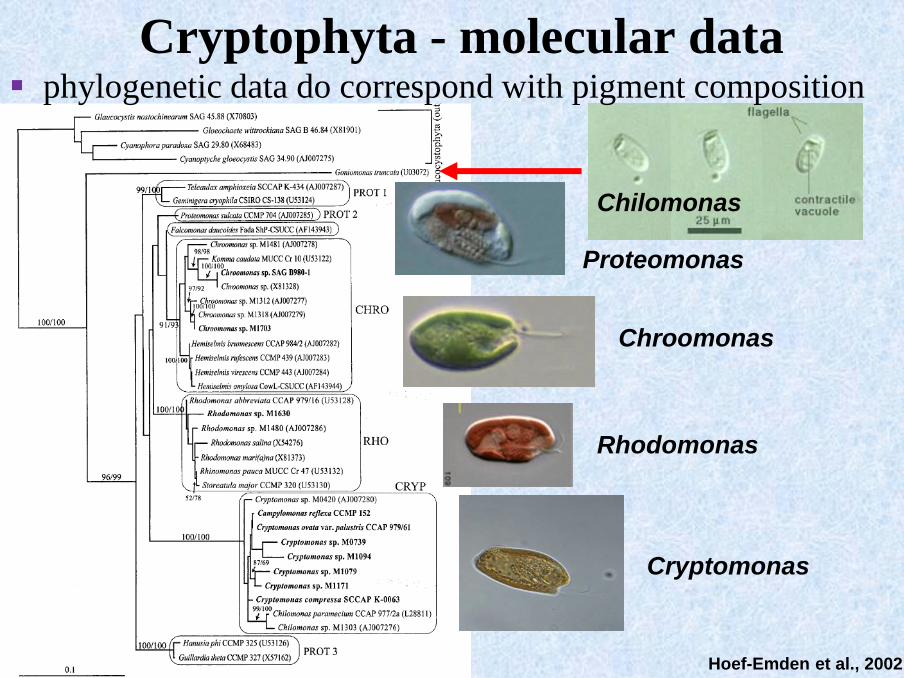
Hoef-Emden et al., 2002
Proteomonas
Chroomonas
Rhodomonas
Cryptomonas
Chilomonas
Cryptophyta - molecular data▪ phylogenetic data do correspond with pigment composition
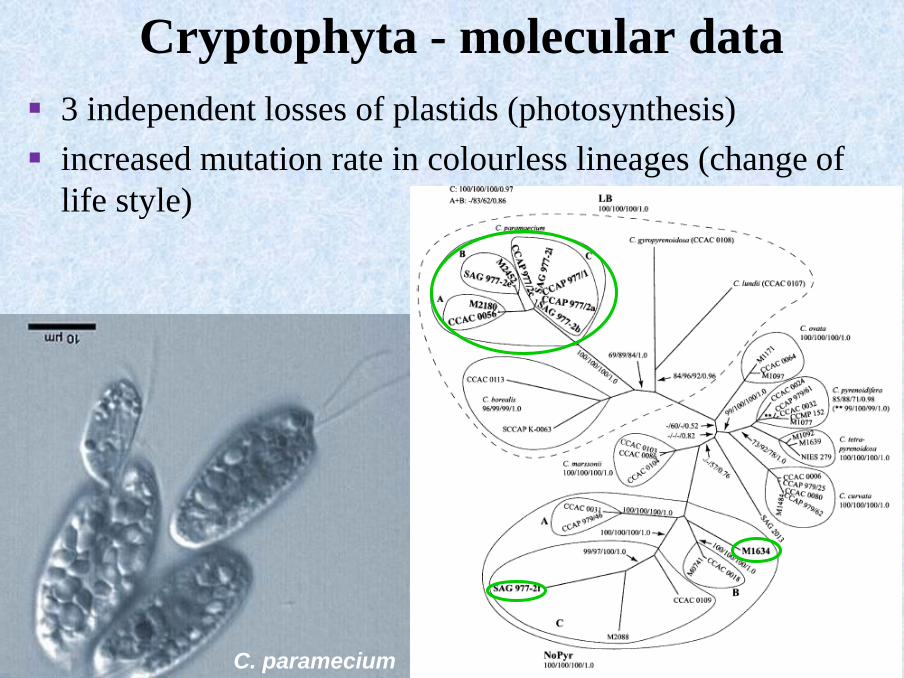
C. paramecium
Cryptophyta - molecular data
▪ 3 independent losses of plastids (photosynthesis)
▪ increased mutation rate in colourless lineages (change of
life style)