estudio paleopalinolÓgico de rubielos de mora … · 2006-2007 pp. 27-53 issn 0210-3524 ... mapa...
TRANSCRIPT

ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL)
Y SUS IMPLICACIONES PALEOCLIMÁTICAS
Uxue Villanueva Amadoz


ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS*
Uxue Villanueva Amadoz**
RESUMEN
Se ha realizado un estudio taxonómico de la palinoflora encontrada en un nuevo yacimiento denominadoFuente del Rull, de edad Rambliense-Aragoniense Inferior. Está localizado en las inmediaciones de Rubielos deMora, donde se localizan algunos de los yacimientos más importantes y de conservación excepcional deEuropa referentes al Mioceno Inferior, no sólo por su contenido en fauna sino también por su paleoflora. Lasfacies lacustres anóxicas someras de grano fino han determinado su Konservat-Lagerstätten, revelando unapredominancia de las gimnospermas de la familia Taxodiaceae y Pinaceae, y en una menor proporción unagran variedad de angiospermas de características mesófilas.
Palabras clave: Paleopalinología, Mioceno Inferior, Rubielos de Mora, Taxonomía.
ABSTRACT
Palaeopalynological study of Rubielos de Mora (Teruel) and its palaeoclimatic implications.
A taxonomic study has been realized of the Ramblian-basal Aragonian palinoflora found in a new sitecalled Fuente del Rull. It is located in the surrounding areas of Rubielos de Mora, where there are some of the
91[I
]■20
06-2
007
■PP
. 27-
53■
ISSN
0210
-352
4
29
* Resumen del trabajo realizado con una ayuda concedida por el Instituto de Estudios Turolenses en su XXIConcurso de Ayudas a la Investigación de 2003.
** Área de Paleontología, Departamento de Ciencias de la Tierra, C/ Pedro Cerbuna, Universidad de Zaragoza.50012 Zaragoza.

most important deposits with exceptional conservation of Europe relating to the Lower Miocene, not only forits fauna content but also for its palaeoflora. The shallow anoxic fine-grained lacustrine facies has determinedits Konservat-Lagerstätten, revealing a predominance of the gimnosperms of the family Taxodiaceae andPinaceae, and in a minor proportion a great variety of angiosperms of mesophytic characteristics.
Key words: Palaeopalynology, Lower Miocene, Rubielos de Mora, Taxonomy.
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
30
INTRODUCCIÓN
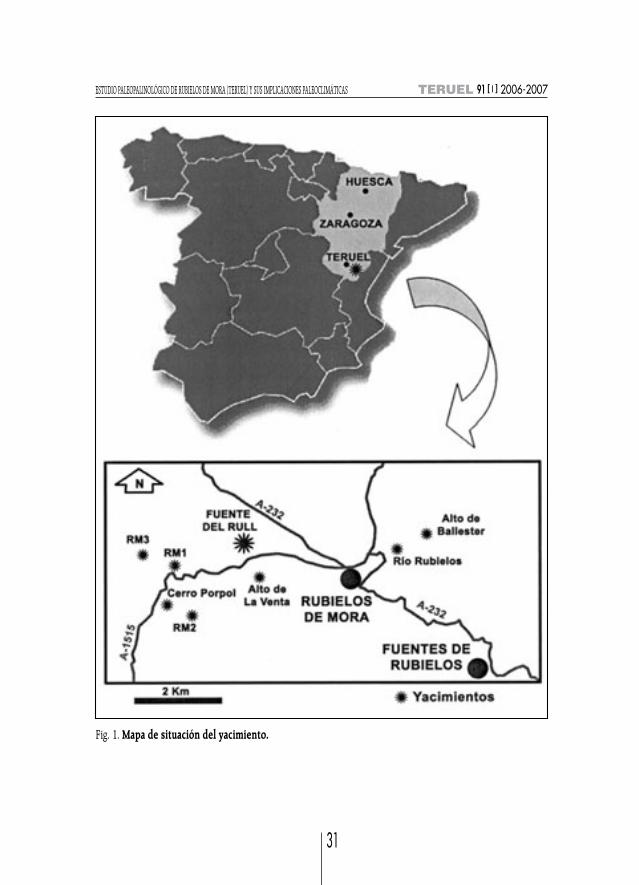
En el presente proyecto aparecen nuevos datos obtenidos de un nuevo yacimiento, denomina-do Fuente del Rull debido a su localización junto a la fuente conocida bajo este nombre. El yaci-miento se localiza a unos 50 km al suroeste de Teruel capital (fig. 1) y a unos 6 km al noroeste de lapoblación de Rubielos de Mora, con coord. U.T.M., x= 855.50, y= 626.00, (Hoja 591, 28-23 delMapa Geológico 1:50.000; GODOY y ANADÓN, 1986).
Además, se conocen otros yacimientos situados en el extremo occidental de la cuenca (Rubielosde Mora 1, 2 y 3 y cerro del Porpol) y otros situados en la parte central y oriental de la cuenca (RíoRubielos, Alto de Ballester 1 y 2 y Alto de la Venta).
Crusafont et al. (1966) dató el yacimiento RM1 mediante roedores, atribuyéndoles una edadVidaboniense basal (Mioceno Inferior), coincidiendo en edad con el yacimiento RM2; para una capasuprayacente a la anterior, estudiada por Bruijn y Moltzer (1974) se atribuyó al Orleaniense, basedel Aragoniense (zona MN4, es decir, zona ibericus con Cordylodon intercedens). Posteriormente,López Martínez (1989) localizó restos de roedores en la zona MN3 (Rambliense Superior), coinci-diendo con Montoya et al. (1996) en su colocación dentro del límite entre las zonas MN3 y MN4)para el yacimiento de Alto de Ballester. Por tanto, se deduce que los restos vegetales (incluyendolos palinomorfos) pertenecen a la Unidad Superior descrita por Anadón et al. (1988a y 1988b) y quese integran dentro de los pisos continentales Rambliense Superior-Aragoniense Inferior (fig. 2).Esta edad coincide, a su vez, con el piso marino Burdigaliense.
El yacimiento paleontológico existente en Rubielos de Mora es considerado como uno de losmás importantes de Europa de edad Rambliense-Aragoniense Inferior (Mioceno Inferior), el cual hasido datado gracias al estudio de mamíferos (MONTOYA et al., 1996). Representa uno de los lugaresprivilegiados en el que se ha encontrado, hasta el momento, tanto micro como macroflora delNeógeno en la Península Ibérica, la cual está escasamente estudiada. Esta importancia reside nosolo por su contenido en paleoflora sino también por su riqueza en otros grupos fósiles comomamíferos, aves, roedores, anfibios, insectos, arácnidos, crustáceos y moluscos. Por tanto, la excep-cional conservación de los restos fósiles hace que se considere un yacimiento de tipo Konservat-Lagerstätten (SEILACHER, 1970; PEÑALVER y SEILACHER, 1995).
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
31
Fig. 1. Mapa de situación del yacimiento.

91 [ I ] 2006-2007 Uxue Villanueva Amadoz
32
Fig. 2. Columna estratigráfica del yacimiento Fuente del Rull (Rubielos de Mora, Teruel).

ANTECEDENTES
Los primeros estudios geológicos realizados en la cuenca de Rubielos de Mora están relacionadoscon aspectos de minería, centrados en la extracción de petróleo (FERNÁNDEZ-NAVARRO, 1914), ligni-tos y pizarras bituminosas del Oligoceno en el Cerro Porpol (GAVALA, 1921; REYES y FEIXAS, 1984).Los estudios estratigráficos y estructurales se deben a Anadón (1983), Godoy y Anadón (1986),Anadón et al. (1988a y 1988b), los cuales remarcan que las secuencias cíclicas registran condicionesalternantes de fondo óxico-anóxico en las zonas marginales de un lago meromíctico debido a varia-ciones cíclicas en el volumen de agua del lago. A su vez, se registran cambios mineralógicos cíclicos yse concluye que la fuerte subsidencia y el paleoclima determinaron la formación de este lago; Pradoet al. (1988) y Anadón et al. (1989 y 1991) demostraron la disposición asimétrica de los depósitoslacustres como consecuencia de la estructura de half-graben y recalcan la importancia de considerarla morfometría de este paleolago para posibles explotaciones de pizarras bituminosas.
Los primeros datos paleontológicos se refieren a restos vegetales mal conservados de los génerosSalix (Salicaceae), Corylus y Ostrya (Betulaceae), que fueron relacionados con los hallados en la cuen-ca de Ribesalbes (Castellón) de edad similar (HERNÁNDEZ-SAMPELAYO y CINCÚNEGUI, 1926).Posteriormente se realizó un estudio palinológico (FERNÁNDEZ-MARRÓN y ÁLVAREZ-RAMIS, 1988;ÁLVAREZ-RAMIS y FERNÁNDEZ-MARRÓN, 1994) deduciendo el paleoclima, describiendo la dominanciade Inaperturopollenites fspp., Triporopollenites fspp., Trivestibulopollenites fsp., Tricolpopollenitesfspp., Subtriporopollenites fsp., Caryapollenites sp. y Aceripollenites sp. Baltuille et al. (1992) contri-buyeron a los primeros trabajos exclusivamente palinológicos sin indicar el lugar donde se extrajeronlas muestras. Posteriormente Roiron (1992) recopiló los restos de macroflora del Neógeno del Sur deFrancia y del Noreste de España y Barrón (1997) realizó un estudio paleobotánico de la cuenca mio-cena inferior de Rubielos de Mora. Los porcentajes de polen de los afloramientos Alto de la Venta yRío Rubielos los obtuvo Alcalá (1997). Aparecen otras síntesis del contenido paleobotánico deRubielos de Mora en Barrón et al. (1996) y en Ferrer et al. (1998). Posteriormente, Roiron et al.(1999) describieron dos asociaciones polínicas junto con sus respectivas reconstrucciones paleoam-bientales, y Barrón y Santisteban (1999) correlacionaron facies deposicionales de la cuenca median-te análisis palinológicos deduciendo la evolución paleoclimática. Siguen las nuevas aportaciones demacroflora por parte de Rubio et al. (2000a y 2000b), Rubio (2001) y Barrón y Diéguez (2001).
Existe una síntesis de la riqueza de mamíferos, aves, roedores, anfibios, insectos, arácnidos,crustáceos y moluscos en Montoya et al. (1996).
En cuanto a vertebrados, el primer yacimiento dentro de la cuenca fue localizado por Crusafont etal. (1966). Aguirre y Moissenet (1972) describieron los restos de un Rhinoceróntido de gran tamaño,así como otros fósiles: Insectivora (Galeria sp., Sorex sp., Talpidae gen.), Rodentia (Pseudodyromys
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
33

sp.), Lagomorpha (Lagopsis pena) y Perissodactyla (Equidae, Anchitherium aurelianense, Rhino-cerotidae, Brachypotherium sp., Dicerohinus, Palaeomerycidae kaupi, Lagomerycidae, Procervulusdichotomus). Bruijn y Moltzer (1974) concluyeron que Cordylodon intercedens es el mejor biomar-cador estratigráfico dentro de Insectivora y debería situarse en la parte inferior de la Zona ibericus,asimismo sugirieron que la presencia de Blackia, Neocometes, Cordylodon, Talpidae y ranas, asícomo la ausencia de ardillas y Cainotherium, indicaban un biotopo húmedo.
Sanchiz (1977) presentó dentro de un catálogo de todos los anfibios fósiles de España presenteshasta la fecha el yacimiento de Rubielos II con la biozonación MN4a de MEIN (1975). En este yaci-miento se encontró Chelotriton paradoxus (Salamandradidae) y Rana pueyoi (Ranidae).
Álvarez Sierra y Daams (1987) describieron la distribución de Pseudotheridomys fejfari(Eomyidae) restringida al Aragoniense Inferior. López-Martínez (1989) estudió los lagomorfos pro-cedentes de la cuenca.
Los insectos han sido estudiados en primer lugar por Martínez-Delclós et al. (1991); Fernández-Rubio et al. (1991) describieron una nueva especie que denominaron zygaena? turolensis en las rit-mitas lacustres; Peñalver y Seilacher (1995) y Peñalver y Martínez-Delclós (1996) describieron lasacumulaciones de chironómidos (Diptera Nematocera: Chironomidae), insectos que precisan deagua principalmente dulce, al quedarse atrapados en las redes microbiales en la superficie del agua.También recalcaron la característica de los chironómidos en la formación de enjambres en los már-genes de los lagos durante dicho período reproductivo. Pertenecen al yacimiento de Río Rubielos II.Peñalver et al. (1996) sugirieron que la presencia del subgénero Ctenodahpnia indicaba la existenciade aguas temporales, o al menos en algunas zonas, con corrientes de baja energía durante elMioceno en el ambiente lacustre de Rubielos de Mora. Esta deducción se obtuvo del hecho de queestos efipios se producían por estrés hídrico. Peñalver (1997) mostró que orugas de Lepidoptera sealimentaban de Zelkova, Salix o Myrica en los alrededores del lago. Siguiendo la misma línea deinvestigación, Peñalver y Martínez-Delclós (1997) observaron tres tipos de interacción insecto-plan-ta: a) hojas parcialmente comidas por orugas lepidópteras, b) galerías en las hojas realizadas por lar-vas minadoras, y c) ovoposiciones en las hojas. Se describió en Peñalver et al. (1997) el segundomiriápodo encontrado en todo el mundo perteneciente al género Lithobius (Familia Lithobiidae,Clase Chilopoda). Fue hallado también en el yacimiento Río Rubielos. En Peñalver (1998) se hizohincapié en la tafonomía y paleoecología de los insectos encontrados en esta zona.
En Heie y Peñalver (1999) se describió un nuevo género (Palaeophylloxera nov. gen.) pertene-ciente a la familia Phylloxeridae (Hemiptera, Phylloxeroidea) y en Peñalver y Baena (2000) se recal-có la presencia del género Dicyphus.
Herzeck et al. (2000) presentaron el primer registro fósil de Aragocylapus miocaenicus n. gen.,n. sp. (Heteroptera:Miridae:Cylapinae).
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
34

MARCO GEOLÓGICO
La cuenca de Rubielos de Mora, situada en la Cadena Ibérica, consiste en el relleno de una cube-ta alargada por materiales del Mioceno inferior-medio según una dirección ENE-OSO, con unasdimensiones máximas de 12 km de largo por 3 km de ancho. Esta cubeta se desarrolló en un con-texto extensivo (SIMÓN, 1984), localizándose entre la fosa de Teruel y las fosas costero valencianas(de orientación N030E, al igual que la orientación de las fallas normales). La orientación de lasfallas en esta zona es de NW-SE, con una menor proporción en fallas de orientación E-W. La inter-sección entre fallas de diferente dirección provoca la estructura en horst y graben, así como un esti-ramiento de la cobertera mesozoica (GUIMERÀ, 1990).
La cuenca presenta una estructura sinclinal asimétrica, con los estratos más verticalizados en suflanco sur. Estos materiales miocenos aparecen discordantes sobre materiales mesozoicos. La forma-ción de la cubeta sinclinal se debe a la aparición de dos fallas normales kilométricas de orientaciónENE-WSW. En este contexto se forman flexiones en los materiales dentro de la cuenca subsidente aladaptarse al cambio de buzamiento de la superficie de la falla, con el resultado de la formación de lacubeta sinclinal asimétrica (GUIMERÀ, 1990). Además en la zona oriental de la fosa se presentanslumps y diversos olistolitos estratiformes de calizas cretácicas y facies asociadas (ANADÓN, 1983).
ESTRATIGRAFÍA
Las muestras pertenecen a la Unidad Superior, situada sobre la Unidad Media. Según Godoy yAnadón (1986) la Unidad Media está constituida por lutitas versicolores, calizas, arenas y lignitos.Esta Unidad es un cambio lateral de facies de la Unidad Inferior. Existe un primer tramo calizomargoso con intercalaciones de arcillas, margas grises y, en ocasiones, lignitos. Sobre estas calizasaparece otro tramo formado por lutitas grises, pardas y rojizas con intercalaciones de areniscas gri-ses y amarillas y calizas arenosas. Esta Unidad correspondería con un área de depósito lacustresomero de edad Aragoniense con importantes episodios de aportes terrígenos tractivos.
La Unidad Superior es denominada “Serie bituminosa” y se considera de edad Aragoniense.Está constituida por margas, arcillas laminadas y ritmitas. En la zona de Río Estrecho se distingueestá Unidad por la presencia de lutitas y margas blancas, verdosas y pardas con intercalaciones depelitas laminadas grises y niveles bituminosos, mientras que en la parte oriental de la Cuenca exis-te una sucesión lutítico-margosa, de gris a pardo oscura, a menudo laminadas conteniendo restosvegetales bien conservados y moluscos lacustres. En esta última sucesión existen esporádicas inter-calaciones de calizas y areniscas de escasa potencia. El emplazamiento de olistolitos y facies asocia-das (slumps, etc.) es el registro perteneciente a la actividad tectónica ocurrida en los márgenes de lacuenca. Correspondería con un depósito lacustre de cierta profundidad con una preservación de rit-
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
35

mitas, restos vegetales delicados y ausencia de bioturbación que indicaría anoxia en el fondo dellago.
En la parte oriental de la cuenca aflora una Unidad que resulta ser un cambio lateral de faciesde las dos últimas unidades de edad Aragoniense. Está constituida por arenas y areniscas amari-llentas o pardas, más raramente grises, con abundantes nódulos ferruginosos y con intercalacionesde lutitas y margas grises.
Centrándonos en el yacimiento de Fuente del Rull, se observa una clara dominancia de lutitaslaminadas con intercalaciones de calizas y margocalizas fundamentalmente (fig. 2).
METODOLOGÍA
Las muestras se procesaron según los protocolos paleopalinológicos clásicos, adaptándolos alentorno de trabajo de los laboratorios del Departamento de Ciencias de la Tierra de la Universidadde Zaragoza.
En el trabajo de gabinete se han observado los granos de polen al microscopio óptico para cono-cer tanto su estructura como la ornamentación de la exina. En la descripción de los granos de poleny esporas se ha utilizado la terminología de Punt et al. (1994) en su intento de unificar todos losconceptos aparecidos hasta el momento con el único objeto de facilitar la comprensión del signifi-cado de cada término. Se ha optado por aplicar términos en castellano basándonos en trabajos pre-vios realizados en español (SÁENZ, 1978). Asimismo, se han fotografiado (aumento x 1000) y mejo-rado infográficamente.
DESCRIPCIÓN SISTEMÁTICA
Nos hemos basado fundamentalmente en la monografía publicada por Thomson y Pflug(1953), en la cual se propone una nomenclatura para la palinología terciaria de Europa.Complementariamente hemos utilizado otras obras como Kedves, 1969; Seyítoglu y Benda, 1998,y Akgün y Akyol, 1990.
Abreviaturas utilizadas:
anch. Anchura ex. Exina p. Polarcresc. Crescente gs. Grosor supf. Superficieec. Ecuatorial long. Longitud v. Vistaengr. Engrosamiento orn. Ornamentación Ø Diámetro
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
36

ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
37
PTERIDOFITAS
EQUISETACEAE
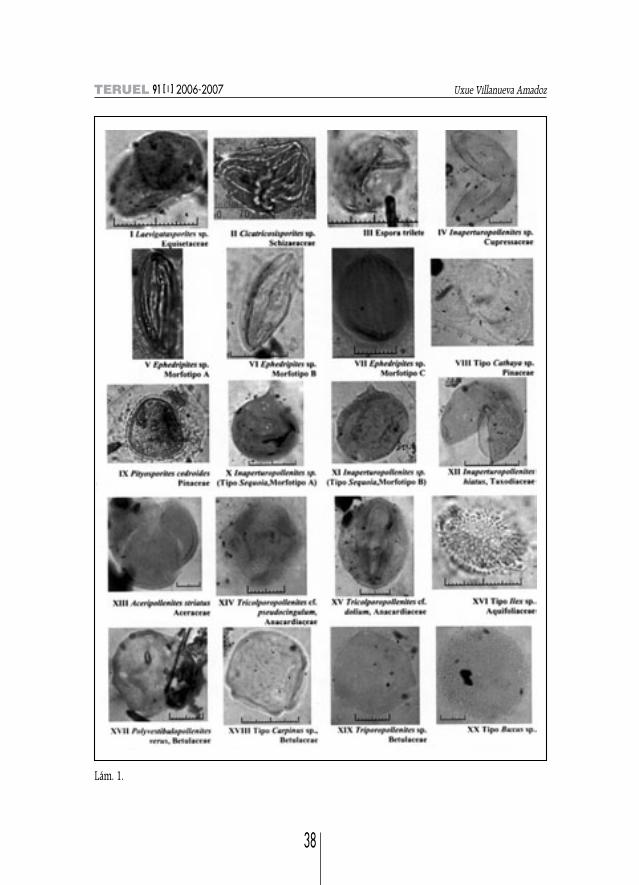
Equisetum sp. (=Laevigatasporites sp.) (lám. 1, fig. I)Descripción: Subesférico; Ø medio de 24 µm; monocolpado; ex. 2 µm; psilado.
SCHIZAEACEAECicatricosisporites sp. (lám. 1, fig. II)
Descripción: Subtriangular en v. ec.; eje de mayor long. 33 µm; espora trilete con numerosasbandas concéntricas de 3-4 µm de gs.
INDETERMINADO: Espora trilete (lám. 1, fig. III)Descripción: Heteropolar, subtriangular en v. ec., bordes convexos; long. 15 y anch. 12 µm; tri-
lete, anch. de la laesura de 1,8 µm, con brazos que se extienden hasta los bordes; ex. de 1 µm.
GIMNOSPERMAS
CUPRESSACEAE
Inaperturopollenites sp. MORFOTIPO A. Descripción: Isopolar, subesférico, laesura muy marcada; long. 28-30 µm; ina-
perturado; psilado con gemas dispersas (lám. 1, fig. IV).MORFOTIPO B. Descripción: Apolar, subesférico; 35-40 µm de Ø medio; inaperturado; psilado.
EPHEDRACEAETipo Ephedra (=Ephedripites sp.) (lám. 1, figs. V, VI, VII)
MORFOTIPO A. Descripción: Isopolar, de prolato a perprolato, en v. ec. elíptico; long. 42-44 µmy anch. 20-24 µm; poliplicado, colpos y surcos rectilíneos, con una anch. del colpo de 13, 3 µm; ex.de 0,7 µm de gs. (lám. 1, fig. V).
MORFOTIPO B. Descripción: Isopolar, perprolato, en v. ec. elíptico; long. 37 µm y anch. 17 µm;poliplicado, colpos y surcos anastomosados; ex. de 1,7 µm de gs. (lám. 1, fig. VI).
MORFOTIPO C. Descripción: Isopolar, prolato, en v. ec. elíptico; long. 22,5 µm y anch. 17 µm;poliplicado, colpos y surcos rectilíneos, con una anch. del colpo de 2 µm (lám. 1, fig. VII).
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
38
Lám. 1.

PINACEAE
1) Tipo Cathaya sp. (=Pinus tipo haploxylon (Rudolph 1935), =Abietineapollenites microalatus,=Pityosporites microalatus (R. Pot. 1931b) Th. y Pf. 1953 (lám. 1, fig. VIII)
Descripción: Heteropolar, bilateral, bisacado, en v. p.r bordes ligeramente elípticos con un cuer-po romboidal, sacos originados en el margen del cuerpo, engr. (4 µm) en el área adyacente a launión entre el cuerpo y los sacos; long. total 60-75 µm, altura total 45 µm, sacos de long. mayores35 x 25 µm, cuerpo con ejes máximos 50 x 35; cappa de 2,5 µm de gs.; verrugoso, con un reticula-do amplio en los sacos.
2) Pinus tipo sylvestris Rudolph 1935 (=Pityosporites labdacus, =Pinus tipo diploxylon)Descripción: Heteropolar, bilateral, bisacado, en v. ec. cuerpo subesférico y sacos circulares, en
v. p. cuerpo esferoidal y sacos elípticos, con una constricción aparente entre el cuerpo y los sacos;long. total de 70 µm; analeptomado o inaperturado; gs. de la parte dorsal de la ex. del cuerpo igualo mayor a 5 µm, semitectado; sacos microverrugados con orn. no muy marcada; cuerpo muy verru-goso, con un engr. claro del cuerpo cerca de la unión con los sacos.
3) Tipo Cedrus (=Pityosporites cedroides Th. y Pfl.) (lám. 1, fig. IX)Descripción: Heteropolar, bilateral, bisacado, en v. ec. cuerpo subesférico aplanado y sacos cir-
culares, en v. p. cuerpo trapezoidal y sacos elípticos, con una constricción aparente entre el cuerpoy los sacos; en v. p. cuerpo de ejes mayores de 50 x 40 µm, sacos más o menos del mismo tamañoque el cuerpo; exina de 4 µm de gs., verrugoso, con un reticulado amplio en los sacos aéreos.
4) Tipo PiceaDescripción: Heteropolar, bilateral, bisacado; long. total de 70 µm; analeptomado o inapertura-
do con los sacos fusionados al cuerpo; reticulado fino.
TAXODIACEAE
1) Tipo Sequoia, Metasequoia, Cryptomeria (=Inapertupollenites sp.) (lám. 1, figs. X, XI)MORFOTIPO A. Descripción: Heteropolar, bilateral, subprolato o subesférico; altura 30 µm y
anch. 28 µm; inaperturado con una papilla prominente de 2 µm de altura y 2 µm de anch.; exinadelgada de 0,8 µm de gs.; baculado (lám. 1, fig. X).
MORFOTIPO B. Descripción: Heteropolar, bilateral, suboblato; altura 27 µm y anch. 33-37 µm;inaperturado con una papilla prominente de 2 µm de altura y 2 µm de anch.; exina delgada de 0,8µm de gs.; baculado. (lám. 1, fig. XI).
2) Tipo Taxodium o Glyptostrobus (=Inapertupollenites sp.; = Taxodiaceaepollenites hiatus;Inaperturopollenites hiatus (R. Pot.) Th. y Pf.) (lám. 1, fig. XII)
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
39

Descripción: Apolar, bilateral, subesférico con una fisura triangular; 26 µm de Ø medio; inaper-turado; exina muy delgada de 0,5 µm de gs.; psilado con alguna gema dispersa.
ANGIOSPERMAS
ACERACEAE
Tipo Acer (=Aceripollenites striatus) (lám. 1, fig. XIII)Descripción: Isopolar, trilobulado en v. p., subprolato; long. 25-28 y anch. 21-22 µm; tricolpado,
con anch. del colpo 2,5 µm; ex. 1-1,5 µm; estriado-rugulado.
ANACARDIACEAE
1) Tipo Rhus sp. (=Tricolporopollenites cf. pseudocingulum) (R. Pot. 1931) Th y Pf. 1953 (lám. 1,fig. XIV)
Descripción: Isopolar, subprolato, apiculado; long. 28 y anch. 25 µm; tricolporado; ex. 1,5 µm;estriado-rugulado.
2) Tricolporopollenites cf. dolium (R. Pot. 1931) Th y Pf. 1953 (lám. 1, fig. XV)Descripción: Isopolar, prolato; long. 30 y anch. 22 µm; ex. 1,5 µm; tricolporado con poros de Ø
medio de 3 µm; verrugoso.
AQUIFOLIACEAE
Tipo Ilex (lám. 1, fig. XVI)Descripción: Isopolar, prolato, en v. ec. elíptico y en v. p. subtriangular; long. 25 y anch. 19 µm;
tricolporado; ex. 2,5 µm; clavado, insulado.
BETULACEAE
1) Tipo Alnus (=Polyvestibulopollenites verus (R. Pot. 1931b) Th. y Pf. 1953) (lám. 1, fig. XVII)Descripción: Subisopolar, oblato, en v. ec. elíptico y en v. p. subcuadrangular o subpentagonal;
Ø medio de 24-25 µm, pentaporado, angularperturado, con poros 2 x 1 µm conectadas entre símediante arci de gs. 2 µm y con marcados onci; exina de gs. 1,5 µm; psilado.
2) Tipo Carpinus (lám. 1, fig. XVIII)Descripción: Subisopolar, oblato, en v. ec. elíptico y en v. p. subcuadrangular, con uno de los
poros no equisdistante al resto; ejes mayores de 25 x 28 µm; tetraporado, angularperturado, con 2µm de Ø medio del poro; exina delgada de 0,7 µm de grosor; rugulado.
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
40
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
41
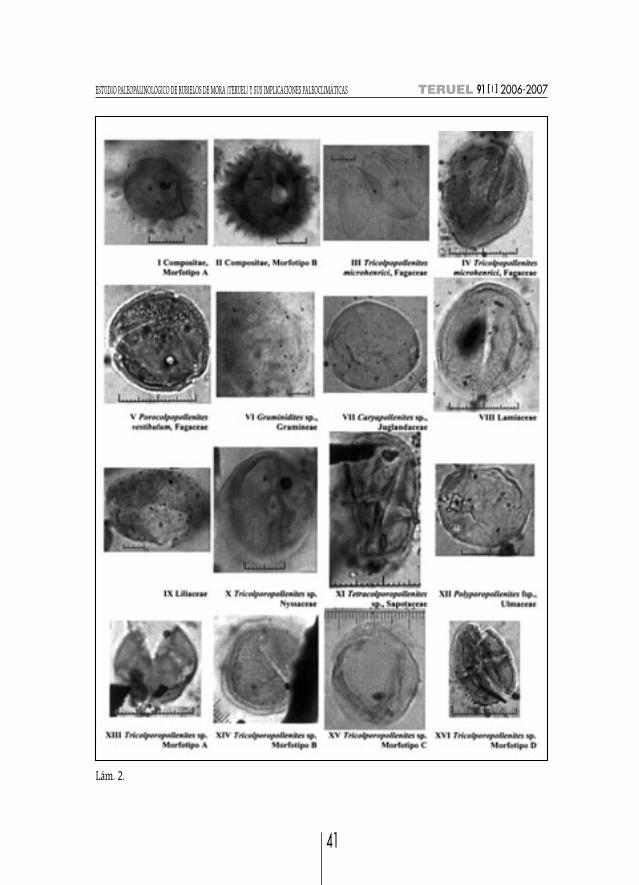
Lám. 2.

3) Tipo Corylus (=Triporopollenites sp.) (lám. 1, fig. XIX)Descripción: Isopolar, de suboblato a oblato, en v. ec. elíptico y en v. p. subtriangular; ejes
mayores de 22 x 22 µm; triporado, angularperturado, con poros de Ø medio de 2 µm rodeados porannuli; exina de gs. 0,8 µm, rugulado.
BUXACEAE
Tipo Buxus (lám. 1, fig. XX)Descripción: Apolar, radial, esférico; 28-35 µm de Ø; periporado con más de 50 poros, aperturas
simples tipo poro sin annuli de 1,5 µm de Ø, poros más grandes que los brochi; ex. ondulada de gs.1,2 µm, semitectado; reticulado o vermiculado-rugulado, con gemas dispersas de 0,7 µm de h; colu-melas delgadas distinguibles.
COMPOSITAEMORFOTIPO A. Descripción: Isopolar, subesférico; 28 µm de Ø medio sin contar con las espi-
nas; fenestrado, con amplias lagunas; espinoso con espinas de 3 µm de altura (lám. 2, fig. I).MORFOTIPO B. Descripción: Isopolar, subprolato; long. 29 y anch. 28 µm; tricolporado con
poro elongado meridionalmente (7 x 5 µm); ex. de 7 µm de gs. contando las espinas; espinoso conespinas de 4 µm de altura (lám. 2, fig. II).
ERICACEAE (=Tetradopollenites callidus (Potonié 1931a) Pf. y Th. 1953)Descripción: Heteropolar, tétrada tetraédrica, subtriangulares; tétrada de dimensiones mayores
35 x 29 µm; granos tricolporados, colpos largos y estrechos (de anch. menor a 0,5 µm) con poroselongados transversalmente; rugulado.
FAGACEAE
1) Tipo Quercus ilex-coccifera (=Tricolpopollenites microhenrici Th. y Pf.) (lám. 2, figs. III, IV)Descripción: Isopolar, prolato; en v. ec. elíptico, bordes redondeados; long. 35 y anch. 25 µm;
tricolpado con colpos engrosados irregulares; exina de gs. 1 µm; verrugoso.
2) Porocolpopollenites vestibulum (R. Pot.) Th. y Pf. (lám. 2, fig. V)Descripción: Isopolar, subtriangular en v. p., bordes redondeados; long. 26 y anch. 24 µm; tri-
colporado con colpos de 2 µm de gs.; exina de gs. 1,5 µm; granular.
GRAMINEAE (=Monoporopollenites sp.; =Graminidites gramineoides; Graminidites sp. Cooksonex Potonié emend. Krutzsh, 1970) (lám. 2, fig. VI)
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
42

Descripción: Isopolar, subesférico; grano de gran tamaño (35 µm de Ø); monoporado con unporo de pequeño tamaño (3,3 µm de Ø) con un annulus marcado; ex. tectada delgada de 0,8 µm degs.; escábrido.
JUGLANDACEAE1) Carya sp. (=Caryapollenites sp.; =Caryapollenites fsp.; =Subtriporopollenites simplex) (lám. 2,fig. VII)
Descripción: Suboblato, en vista polar triangular con bordes redondeados y con una depresióncircular en el centro del grano; ejes mayores de 31 x 35 µm; triporado con poros ligeramente elon-gados transversalmente (3 x 2,5 µm) donde la mayoría aparecen en posición subecuatorial; exinatectada de 2 µm de gs., endexina más delgada que la ectexina, cerca de la apertura la endexinadesaparece; psilado.
2) Platycarya sp. Descripción: Isopolar, suboblato, en vista polar triangular con bordes redondeados; long. 13 y
anch. 14 µm; triporado, angulaperturado, poros de 1,5 µm de Ø sin annuli; ex. tectada de 1 µm degs., endexina más delgada que la ectexina; psilado.
3) Engelhardia sp. (=Triatripollenites fsp.)Descripción: Isopolar, subprolato; en vista polar subtriangular; altura 22 y anch. 19 µm; tripora-
do, con poros sin annuli de 1,5 µm de Ø; ex. tectada de 1 µm de gs., endexina más delgada que laectexina y desapareciendo cerca de las aperturas; ornamentación basada en nanoespículas regular-mente distribuidas.
LAMIACEAE (lám. 2, fig. VIII)Descripción: Isopolar, subprolato, subesférico en v. p.; grano de long. 33,5 y anch. 28,5 µm;
hexacolpado, con colpos muy estrechos de anch. 1,5 µm; gs. de la ex. (3 µm) en los intercolpi uni-forme, adelgazándose hacia los colpos, doble capa de la ex.; reticulado.
LILIACEAE (lám. 2, fig. IX)Descripción: Heteropolar, bilateral, prolato de bordes redondeados; long. 40 y anch. 36 µm;
monocolpado; reticulado con brochi > 2 µm.
NYSSACEAETipo Nyssa (=Tricolporopollenites sp.; =Nyssapollenites (Potonié 1931) Nagy 1969) (lám. 2, fig. X)
Descripción: Isopolar, prolato; long. 27 y anch. 23,5 µm; tricolporado; exina reticulada.
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
43
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
44
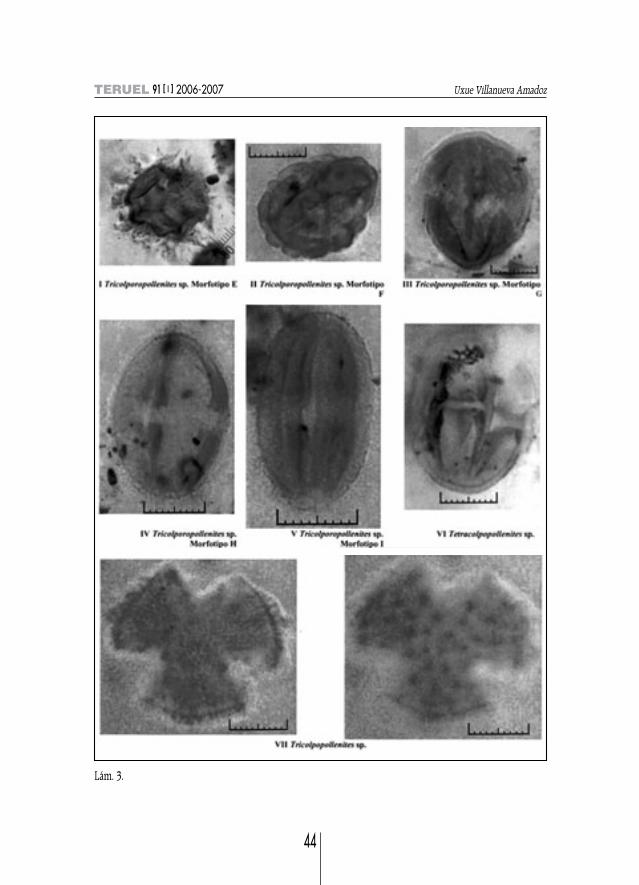
Lám. 3.

SALICACEAE
Tipo Salix (=Tricolpopollenites retiformis Th. & Pf.) Descripción: Isopolar, de subesférico a subprolato, en v. p. circular; long. 23 y anch. 22 µm; tri-
colpado, con anch. del colpo de 1,5 µm; per-reticulado con brochi > 2 µm, muri formado por unafila de columelas.
SAPOTACEAE (=Tetracolporopollenites sp.) (lám. 2, fig. XI)Descripción: Isopolar, prolato; long. 26 y anch. 16 µm; tetracolporado con poros elongados
ecuatorialmente (2 x 3 µm); ex. 1,5 µm, infratectum columelar; escábrido.
ULMACEAE
Tipo Ulmus / Zelkova (=Polyporopollenites fsp.) (lám. 2, fig. XII)Descripción: Isopolar, oblato, en v. p. pentagonal; Ø medio de 35-40 µm generalmente pentapo-
rado con poros ovales de 1,5 µm de Ø; exina de 1 µm de gs.; rugulado.
INDETERMINADOS (Descritos según su morfología)
Tricolporopollenites sp.MORFOTIPO A. Descripción: Isopolar, suboblato, en v. ec. subrómbico, subesférico en v. p.,
ligeramente apiculado; long. 21 y anch. 23 µm; tricolporado, con poros elípticos elongados trans-versalmente de 2,5 x 6,5 µm; ex. 1,3 µm, per-reticulado con brochi < 1 µm (lám. 2, fig. XIII).
MORFOTIPO B. Descripción: Isopolar, suboblato, subesférico; Ø medio de 27-28 µm; tricolpora-do, con anch. del colpo 2,8 µm adelgazándose hacia los polos con terminación apuntada; ex. de 3,5µm de gs., infratectum columelar muy marcado; reticulado-estriado (lám. 2, fig. XIV).
MORFOTIPO C. Descripción: Apolar, subesférico; Ø medio de 25 µm; tricolporado con Ø mediodel poro de 3 µm; ex. de gs. /1 µm; reticulado con brochi < 1 µm (lám. 2, fig. XV).
MORFOTIPO D. Descripción: Isopolar, prolato con polos redondeados; long. 28 y anch. 18 µm;tricolporado, con un anillo transversal elíptico; ex. 1 µm; per-reticulado con brochi < 1 µm (lám. 2,fig. XVI).
MORFOTIPO E. Descripción: Isopolar, subtriangular; eje de long. mayor de 24 µm; tricolporado;ex. 1,2 µm; espinoso con espinas resolutas de 6 µm de altura (lám. 3, fig. I).
MORFOTIPO F. Descripción: Isopolar, prolato; long. 22 y anch. 16,5 µm; tricolporado; ex.2 µm; verrugoso de altura 1 µm (lám. 3, fig. II).
MORFOTIPO G. Descripción: Isopolar, subprolato, apiculado; long. 29 y anch. 26 µm tricolpo-rado; ex. 1,5 µm verrugoso de altura 1 µm rugulado (lám. 3, fig. III).
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
45

MORFOTIPO H. Descripción: Isopolar, prolato; long. 29 y anch. 20 µm; tricolporado; ex. 1 µmen el ecuador y 2 µm en los polos; verrugoso (lám. 3, fig. IV).
MORFOTIPO I. Descripción: Isopolar, prolato; long. 24,5 y anch. 15 µm; tricolporado con un Ømedio del poro de 3 µm; ex. 1 µm verrugoso (lám. 3, fig. V).
Tetracolpopollenites sp. (lám. 3, fig. VI)Descripción: Isopolar, prolato, bordes redondeados; long. 29,5 y anch. 22 µm; tetracolpado con
una anch. del colpo de 1,5 µm; ex. de 1 µm de gs., infratectum columelar; psilado.
Tricolpopollenites sp. (lám. 3, fig. VII)Descripción: Isopolar, prolato, en v. p. subesférico; eje de long. máx. de 30 µm; tricolporado; ex.
0,5 µm cerca del colpo y 2 µm en los intercolpi, cresc.; espinoso con ex. estriada entre las espinas.
DEDUCCIONES PALEOCLIMÁTICAS
En un contexto general, el Neógeno comienza hace unos 23 millones de años con el Miocenobajo un clima subtropical. A finales del Neógeno es característica la expansión de taxones caducoscomo árboles, herbazales y compuestas, así como de la limitación de las coníferas a altas altitudes ylatitudes. Hace unos 10 millones de años prácticamente todos los restos de angiospermas tropica-les se extinguieron (TRAVERSE, 1988) coincidiendo con la formación de hielo en la Antártida y elinicio de la expansión de una vegetación tipo estepa en América y Eurasia, dominada por matorra-les y herbazales y constituida por compuestas y chenopodiales.
En Rubielos de Mora, tanto el tipo de fauna como el tipo de flora existente en el MiocenoInferior indican un clima subtropical con taxones mesófilos adaptados a una estación de carácterseco. Este hecho ha sido constatado por muchos autores. En el registro paleopalinológico dominanlas gimnospermas (Cathaya, Ephedra, Glyptostrobus o Taxodium, Picea, Pinus, y Sequoia) sobre lasangiospermas (Acer, Alnus, Buxus, Carya, Engelhardia, Fagus, Ilex, Platycarya, Quercus, Salix,Compositae, Gramineae, Ulmáceas). Dentro de este último grupo dominaban las gramíneas,Juglandaceae (Carya y Platycarya) y Ulmaceae. Existen taxones de ribera adaptados a condicioneslocales de aguas estacionales (Salix, Alnus, Ephedra, Liliaceae, Nyssa), así como taxa adaptados amayor altitud (Picea, Cathaya).
A continuación se enumeran las cantidades de cada taxón en el yacimiento Fuente del Rull:
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
46

ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
47
Equi
setu
msp
.1
Trico
lpor
opoll
enite
scf.
doliu
m1
Tipo
Nyss
a3
Cica
trico
sispo
rites
sp.
1Ti
poIle
x1
Tipo
Salix
1Pt
erid
ofita
Inde
term
inad
o4
Tipo
Aln
us1
Tetra
colp
orop
ollen
itess
p.2
Inap
ertu
ropo
lleni
tess
p. M
orfo
tipo A
175
Tipo
Car
pinu
s1
Tipo
Ulm
us/Z
elkov
a15
Inap
ertu
ropo
lleni
tess
p. M
orfo
tipo B
120
Tipo
Cory
lus
9Tr
icolp
orop
ollen
itess
p. A
1Ti
po E
phed
ra, M
orfo
tipo A
1Ti
po B
uxus
1Tr
icolp
orop
ollen
itess
p. B
1Ti
po E
phed
ra, M
orfo
tipo B
2CO
MPO
SITA
E, M
orfo
tipo A
1Tr
icolp
orop
ollen
itess
p. C
1Ti
po E
phed
ra, M
orfo
tipo C
1CO
MPO
SITA
E, M
orfo
tipo B
1Tr
icolp
orop
ollen
itess
p. D
1Ti
po C
atha
ya39
Tetra
dopo
lleni
tes c
allid
us1
Trico
lpor
opoll
enite
ssp.
E1
Tipo
Pinu
sdip
loxy
lon
46Ti
po Q
uercu
s ilex
-cocc
ifera
10Tr
icolp
orop
ollen
itess
p. F
1Ti
po C
edru
s5
Poro
colp
opoll
enite
s ves
tibul
um1
Trico
lpor
opoll
enite
ssp.
G1
Tipo
Pice
a12
Gram
inid
itess
p.17
Trico
lpor
opoll
enite
ssp.
H1
Tipo
Sequ
oia, M
orfo
tipo A
75Ti
po C
arya
15Tr
icolp
orop
ollen
itess
p. I
1Ti
po Se
quoia
, Mor
fotip
o B41
Tipo
Plat
ycar
ya1
Tetra
colp
opoll
enite
ssp.
1Ti
po T
axod
ium
115
Tipo
Eng
elhar
dtia
1Tr
icolp
opoll
enite
ssp.
1Ti
po A
cer
2LA
MIA
CEAE
1Ti
po R
hus
2LI
LIAC
EAE
1Pa
linom
orfo
s ind
eter
min
ados
:20
Tota
l pali
nom
orfo
s: 75
8

En los márgenes de este paleolago meromíctico se encuentran insectos fósiles articulados adap-tados a medios acuáticos mixolimnion. Debido a que aparecen articulados, se deduce que las aguassuperficiales debieron ser tranquilas y el hundimiento debió de ser rápido. La profusión de dípterosindicaría una abundacia de materia orgánica en descomposición y aguas estancadas. Éstos, a su vez,eran parasitados por himenópteros y este hecho correspondía con períodos de pérdida de agua deescasa envergadura en el lago (MARTÍNEZ-DELCLÓS et al., 1991).
DISCUSIÓN
Existen problemas de nomenclatura y clasificación de los granos de polen y esporas para elTerciario, ya que las Angiospermas comienzan su gran expansión en detrimento de las coníferascomo el Pinus. Traverse (1988) apunta tres razones por las cuales se acentúa esta dificultad de asig-nar una nomenclatura a los taxones cenozoicos:
1) Problemática de identificación, ya que granos de polen de angiospermas extinguidos general-mente son sólo identificables a nivel de género (a veces sólo a nivel de familia).
2) Escasez de flores de angiospermas fósiles, que provoca que los escasos ejemplares encontra-dos se comparen con material de referencia actual.
3) Evolución en mosaico y problemas de rango. Se ha observado que varios órganos de lasangiospermas han evolucionado a diferentes pulsos (evolución en mosaico). A veces asignar un tipode polen a un género de planta extinguido es erróneo (POTONIÉ, 1956) ya que podría provenir deotra planta un tanto diferente a las plantas actuales.
Potonié propuso tres tipos de sistemas de clasificación y nomenclatura:
a) Nomenclatura natural, en donde su referencia a un taxón extinguido es cierto y probado.Ejemplo: Pinus sylvestris.
b) Nomenclatura semi-natural, en donde la referencia de un taxón extinguido se sospecha perono está probado. Ejemplo: Betulaceapollenites.
c) Nomenclatura artificial, donde no se sabe bien la afinidad del polen por lo que su nombre sebasa en las características morfológicas. Ejemplo: Tetracolporites.
Erdtman, al igual que Potonié, aconsejaba no usar nombres genéricos para pólenes fósiles pre-pleistocenos. Sin embargo, todos los nombres de taxones fósiles o actuales están regidos por elCódigo Internacional de Nomenclatura Botánica (ICBN-2000). El problema deriva en si es lícito eluso de nombres genéricos actuales para clasificar polen y esporas fósiles o si es necesario usarnombres genéricos en base a su morfología. Actualmente existen dos escuelas diferentes para laclasificación de los granos de polen: 1) clasificación morfológica; 2) clasificación según un sistemanatural (sistema botánico).
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
48

Los trabajos realizados hasta el momento (ROIRON et al., 1999; BARRÓN y SANTISTEBAN, 1999)aplican una asignación actualista, es decir, suponen que las especies eran las mismas en el MiocenoInferior que en la actualidad. Álvarez-Ramis y Fernández-Ramón (1994) realizaron un intento de man-tener la nomenclatura parataxonómica, asignando a los granos de polen su planta productora. Estetipo de clasificaciones difieren de las tendencias actuales, ya que la mayoría de los palinólogos que tra-bajan en instituciones geológicas relacionados más directamente con la Estratigrafía usan el sistemamorfológico (morfotaxa). Sin embargo, aquellos palinólogos que trabajan en instituciones botánicas,más relacionados con el estudio de la evolución de la flora y la vegetación, usan el sistema natural(géneros, familias y, más raramente, también especies). La mejor manera de superar esta controversia(Leon Stuchlik, comunicación personal) es usar las dos clasificaciones simultáneamente. Cada tipo depolen debería relacionarse con cualquier morfotaxón (si existiera) y debería proponerse su afinidadbotánica a especies, géneros o incluso familias actuales si no se conociera su afinidad directa.
CONCLUSIONES
Anadón (1983) habló de una evolución de ambiente lacustre somero a litoral hacia ambienteslacustres más profundos, con estratificación de las aguas y con largos períodos de anoxia en elfondo con la formación de los niveles de pizarras bituminosas. Este estado permanente de falta deoxígeno en el fondo lacustre permitió la acumulación de materia orgánica y la conservación de tipoKonservat-Lagerstätte (SEILACHER, 1970; PEÑALVER y SEILACHER, 1995) de estos yacimientos.
Una problemática existente en la elaboración de este trabajo ha residido en la inexistencia deuna clasificación unificada para palinomorfos terciarios, ya que podría aplicarse tanto una sistemá-tica basada en caracteres morfológicos, siguiendo la literatura clásica, como una basada en su afini-dad biológica siguiendo las pautas biologistas. Esta última sistemática ofrece la ventaja de su com-paración con especies actuales, sin embargo, en muchas ocasiones es imposible asignarles una, portanto, una buena solución es combinar ambas clasificaciones.
En Fuente del Rull se han identificado géneros de granos de esporas y de polen tanto de climatemplado como tropicales, aunque claramente existe una dominancia de taxa de clima templado.Esta dominancia indicaría un clima cálido templado para el Mioceno Inferior en el área de estudio.Dentro del conjunto de cálidos-templados eran dominantes las coníferas (Cathaya, Ephedra,Glyptostrobus o Taxodium, Picea, Pinus, y Sequoia) junto con angiospermas (Acer, Alnus, Buxus,Carya, Engelhardia, Fagus, Ilex, Salix, Quercus, Compositae, Gramineae y Ulmáceas). Por tanto,existen algunos taxones exclusivos para este ambiente local de lago meromíctico de aguas somerasestacionales con corrientes de baja energía, situado en un entorno compuesto por taxones relativosa bosques caducos mesófilos y herbazales bajo un clima subtropical con una clara estacionalidad,junto con otros taxa adaptados a mayor altitud.
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
49

Respecto a la paleoecología en este entorno, junto al lago había numerosas interacciones de laflora, mamíferos, aves, roedores, anfibios, insectos, arácnidos, crustáceos y moluscos, tanto con elmedio como entre sí. Datos tales como los descritos por Peñalver (1997 y 1998) y Peñalver yMartínez-Delclós (1997) respecto a la existencia de indicios de herviborismo por parte de insectospermiten realizar una reconstrucción de los paleoambientes existentes en esta zona, así como ofre-cer una valiosa información acerca de las cadenas tróficas.
Agradecimientos
En este proyecto han colaborado el Dr. José Javier Ferrer Plou del Departamento de Ciencias de laTierra de la Universidad de Zaragoza y el Dr. José Bienvenido Díez del Departamento de GeocienciasMarinas de la Universidad de Vigo, a los cuales se agradecen sus aportaciones y dedicación.
BIBLIOGRAFÍA
AGUIRRE, E. y MOISSENET, E. (1972), «Précisions sur le gisement miocène de Rubielos de Mora (Province deTeruel, Espagne)», Mélanges de la Casa de Velázquez, 8, pp. 561-564.
AKGÜN, F. y AKYOL, E. (1990), «Palynostratigraphy of the coal-bearing Neogene deposits in Büyük Menderes gra-ben, Western Anatolia», Geobios, 32 (3), pp. 367-383.
ALCALÁ, B. (1997), «Prospección palinológica en el Neógeno de Teruel», Teruel, 85(I), pp. 9-20.
ÁLVAREZ-RAMIS, C. y FERNÁNDEZ-MARRÓN, T. (1994), «Conexiones establecidas entre los palinomorfos y losmacrorrestos vegetales del Mioceno medio de Rubielos de Mora (Teruel)», en Polen y esporas: contribu-ción a su conocimiento, VIII Simposio de Palinología (A.P.L.E.) (De Irene La-Serna), pp. 323-331.
ÁLVAREZ-SIERRA, M.A. y DAAMS, R. (1987), «Pseudtheridomys fejfari, a new species of Eomyidae (Rodentia) fromthe Lower Mioceneof North teruel (Spain)», Scripta Geologica, 83, pp. 19-26.
ANADÓN, P. (1983), «Características generales de diversas cuencas lacustres terciarias con pizarras bituminosas delNE de la Península Ibérica», Comunicaciones del X Congreso Nacional de Sedimentología, 1, pp. 9-12.
ANADÓN, P.; CABRERA, L.; INGLÉS, M.; JULIÀ, R. y MARZO, M. (1988a), «The Miocene lacustrine basin of Rubielosde Mora, Excursion Guidebook», International workshop-field seminar on lacustrine facies models inrift systems and related natural resources, p. 32.
ANADÓN, P.; CABRERA, L. y JULIÀ, R. (1988b), «Anoxic-oxic cyclical lacustrine sedimentation in the MioceneRubielos de Mora Basin, Spain», en A.J. FLEET, K. KELTS y M.R. TALBOT (eds.), Lacustrine PetroleumSource Rocks, Geological Society London special Publication, 40, pp. 353-367.
ANADÓN, P.; CABRERA, L.; JULIÀ, R. y MARZO, M. (1991), «Sequential arrangement and asymmetrical fill in theMiocene Rubielos de Mora Basin (northeast Spain)», en P. ANADÓN, L. CABRERA y K. KELTS (eds.),Lacustrine Facies Analysis, Spec. Publs. Int. Ass. Sediment., 13, pp. 257-275.
ANADÓN, P.; CABRERA, L.; JULIÀ, R.; ROCA, E. y ROSSELL, L. (1989), «Lacustrine oilshale basins in tertiary grabens fromNE Spain (western european rist system)», Palaeogeography, Palaeoclimatology, Palaeoecology, 70, pp. 7-28.
BALTUILLE, J.M.; BECKER-PLANTEN, J.D.; BENDA, L. e IVANOVIC CALZAGA, Y. (1992), «A contribution to the subdi-vision of the Neogene in Spain using Palynology», Newsletter on Stratigraphy, 27, pp. 41-57.
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
50

BARRÓN, E. (1997), «Estudio paleobotánico de la cuenca miocena inferior de Rubielos de Mora (Teruel, España)»,en A. GRANDAL D’ANGLADE, J.C. GUTIÉRREZ-MARCO y L. SANTOS FIDALGO (eds.), Libro de Resúmenes yExcursiones de XIII Jornadas de Paleontología, pp. 149-151.
BARRÓN, E. y DIÉGUEZ, C. (2001), «Estudio macroflorístico del Mioceno Inferior lacustre de la Cuenca de Rubielosde Mora (Teruel, España)», Boletín Geológico y Minero, 112, pp. 13-56.
BARRÓN, E.; RIVAS CARBALLO, M.R. y VALLE, M.F. (1996), «Síntesis bibliográfica de la vegetación y clima de laPenínsula Ibérica durante el Neógeno», Revista Española de Paleontología, nº extraordinario, pp. 225-236.
BARRÓN, E. y SANTISTEBAN, C. (1999), «Estudio palinológico de la cuenca miocena de Rubielos de Mora (Teruel,España). Aspectos paleoecológicos y paleobiogeográficos», Boletín de la Real Sociedad Española deHistoria Natural. (Sec. Geología), 95 (1-4), pp. 67-82.
BRUIJN, H. y MOLTZER, J.G. (1974), «The rodents from Rubielos de Mora: the first evidence of the existence of dif-ferent biotopes in the Early Miocene of eastern Spain», Proceedings Koninkle Nederlanden AkademieVan Wetenschappen, B, 77, pp. 129-145.
CRUSAFONT-PAIRÓ, M.; GAUTIER, F. y GINSBURG, L. (1966), «Mise en évidence du Vindobonien inférieur continen-tal dans l’Est de la province de Teruel (Espagne)», Comptes Rendus Sommaires des Séances de laSocieété Géologique de France, 1, pp. 31-32.
FERNÁNDEZ-MARRÓN, T. y ÁLVAREZ-RAMIS, C. (1988), «Note preliminaire sur l’étude paléobotanique du gisementde Rubielos de Mora (Teruel, Espagne)», Résumés Seminaire de Paléobotanique. Organisation Francaisede Paléobotanique, 7.
FERNÁNDEZ-NAVARRO, L. (1914), «La cuenca petrolífera de Rubielos de Mora», Revista de la Academia deCiencias, 13, pp. 237-255.
FERNÁNDEZ-RUBIO, F.; PEÑALVER, E. y MARTÍNEZ-DELCLÒS, X. (1991), «Zygaena? turolensis, una nueva especie deLepidoptera Zygaenidae del Mioceno de Rubielos de Mora (Teruel). Descripción y filogenia», Estudiosdel Museo de Ciencias Naturales de Álava, 6, pp. 77-93.
FERRER, J.; ROIRON, P.; LIÑÁN, E.; DÍEZ, J.B. y MUÑOZ-BARRAGÁN, P. (1998), «La riqueza paleobotánica de Rubielosde Mora», Resúmenes de Comunicaciones de las I Jornadas sobre Patrimonio de Rubielos de Mora(Teruel).
GAVALA, J. (1921), «Nota acerca de los yacimientos de lignitos y pizarras bituminosas de Rubielos de Mora(Teruel)», Boletín del Instituto Geológico y Minero de España, 42, 2.ª ser., pp. 263-302.
GODOY, A. y ANADÓN, P. (1986), Memoria y mapa Geológico de España E. 1:50.000, 2ª ed. Hoja 591 (Mora deRubielos), Instituto Geológico y Minero de España, 52 pp.
GUIMERÀ, J. (1990), «Formación de una cubeta sinclinal en un contexto extensivo: la cuenca miocena de Rubielosde Mora (Teruel)», Geogaceta, 8, pp. 33-35.
HEIE, O.E. y PEÑALVER, E. (1999), «Palaeophylloxera nov. Gen., the first fossil specimen of the familyPhylloxeridae (Hemiptera: Phylloxeroidea); Lower Miocene of Spain», Geobios, 32 (4), pp. 593-597.
HERCZEK, A.; POPOV, Y.A. y PEÑALVER, E. (2000), «The first record of cylapinous plant bugs from the LowerMiocene of Spain: Aragocylapus miocaenicus n. gen., n.sp. ( Heteroptera: Miridae: Cylapinae)», ActaGeológica Hispánica, 35 (1-2), pp. 183-188.
HERNÁNDEZ-SAMPELAYO, P. y CINCÚNEGUI, M. (1926), «Cuenca de esquistos bituminosos de Ribesalbes(Castellón)», Boletín del Instituto Geológico y Minero de España, 6, 3.ª ser., pp. 3-86.
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
51

KEDVES, M. (1969), Palynological studies on Hungarian Early Tertiary deposits, Budapest, Akadémiai Kiadó.
LÓPEZ-MARTÍNEZ, N. (1989), Revisión sistemática y biogeográfica de los Lagomorfa (Mammalia) del Terciario yCuaternario de España, Memorias del Museo Paleontológico de la Universidad de Zaragoza.
MARTÍNEZ-DELCLÒS, X.; PEÑALVER, E. y BELINCHÓN, M. (1991), «Primeras aportaciones al estudio de los insectosdel Mioceno de Rubielos de Mora, Teruel (España)», Revista Española de Paleontología, nº extraordina-rio, pp. 125-137.
MEIN, P. (1975), «Biozonation du Neogène Mediterranéen à partir des Mamiferes», Proceedings of the VIthCongress. R.C.M.N.S. Bratislava, September 4-7, 1975, 2: 18.
MONTOYA, P.; PEÑALVER, E.; RUIZ-SÁNCHEZ, F.J.; SANTISTEBAN, C.; ALCALÁ, L.; BELINCHÓN, M. y LACOMBA, J.I.(1996), «Los yacimientos paleontológicos de la cuenca terciarias continental de Rubielos de Mora(Aragón)», Revista Española de Paleontología, nº extraordinario, pp. 215-224.
PEÑALVER, E. (1997), «Hojas fósiles del Terciario de Teruel con marcas de herbivorismo debidas a orugas», Boletínde la Sociedad Entomológica Aragonesa, 19, pp. 29-33.
• (1998), Estudio tafonómico y paleoecológico de los insectos del Mioceno de Rubielos de Mora (Teruel),Teruel, Instituto de Estudios Turolenses.
PEÑALVER, E. y BAENA, M. (2000), «Primer registro fósil del género Dicyphus (Insecta: Hemiptera:Miridae);Mioceno Inferior de Teruel (España)», Revista Española de Paleontología, 15 (1), pp. 49-56.
PEÑALVER, E. y MARTÍNEZ-DELCLÓS, X. (1996), «Niveles con concentraciones de insectos chironómidos en elMioceno de Rubielos de Mora (Teruel) y Ribesalbes (Castellón): Consideraciones tafonómicas y paleoeco-lógicas», Reunión de Tafonomía y Fosilización, pp. 305-310.
PEÑALVER, E. y MARTÍNEZ-DELCLÒS, X. (1997), «Evidencias de interacción entre insectos y plantas durante elMioceno (cuencas lacustres de Rubielos de Mora, Teruel y Ribelsabes-Alcora, Castellón)», en J.P. CALVO yJ. MORALES (eds.), Avances en el conocimiento del Terciario Ibérico, pp. 153-155.
PEÑALVER, E.; MARTÍNEZ-DELCLÒS, X. y DE RENZI, M. (1996), «Registro de pulgas de agua [Cladocera: Daphniidae:Daphnia (Ctenodaphnia)] en el Mioceno de Rubielos de Mora (Teruel, España)», II Reunión de Tafonomíay Fosilización, pp. 311-317.
PEÑALVER, E.; MARTÍNEZ-DELCLÒS, X. y SERRA, A. (1997), «Hallazgo del género Lithobius (Chilopoda,Lithobiomorpha) en el registro fósil del Mioceno de Rubielos de Mora», en J.P. CALVO y J. MORALES(eds.), Avances en el conocimiento del Terciario Ibérico, pp. 149-152.
PEÑALVER, E.; DE SANTISTEBAN, C. y BARRÓN, E. (1999), «Fossil insects and palaeobotany of the Rubielos de MoraBasin (Teruel)», en G. MELÉNDEZ y C. SORIA-LLOP (eds.), The geological and paleontological heritage ofCentral and Eastern Iberia (Iberian Range, Spain), Publicaciones del Seminario de Paleontología deZaragoza, 4, pp. 95-116.
PEÑALVER, E. y SEILACHER, A. (1995), «Rubielos de Mora. Eine untermiozäne Fossil-Lagerstätte», Fossilien, 4, pp.211-216.
POTONIÉ, R. (1956), «Synopsis der Gattungen der Sporae dispersae», Sporites. Beih. Geol. Jahrb., 23, pp. 1-103.
PRADO, J.G.; SUÁREZ-RUIZ, I.; BORREGO, M.A. y GARCÍA, A.M. (1988), «The nature of organic matter in oil shalesfrom Rubielos de Mora», Inter. Workshop, Lacustrine facies models in rift system and related naturalresources, Barcelona-Rubielos de Mora.
91 [ I ] 2006-2007 Uxue Villanueva Amadoz
52

PUNT, W.; BLACKMORE, S.; NILSSON, S. y LE THOMAS, A. (1994), Glossary of pollen and spore terminology, LPPContribution series No. 1, Utrecht, LPP Foundation.
REYES, J.L. y FEIXAS, J.C. (1984), «Las pizarras bituminosas: definición, composición y clasificación», I CongresoEspañol de Geología, vol. 2, pp. 817-827.
ROIRON, P. (1992), Flores, végetation et climats du Neogène Mediterranéen: apports de macroflores du Sud de laFrance et du Nord-Est de l’Espagne, Université de Montpellier II, Sciences et Techniques du Languedoc.
ROIRON, P.; FERRER, J.; LIÑÁN, E.; RUBIO, C.; DÍEZ, J.B.; POPESCU, S. y SUC, J.P. (1999), «Les flores du bassin lacustrede Rubielos de Mora. Nouvelles données sur les conditions climatiques au Miocène inférieur dans larégion de Teruel (Espagne)», en Comptes rendus de l’Académie des Sciences : sciences de la terre et desplanètes, 329(12a), pp. 897-904.
RUBIO, C. (2001), Geología y paleobotánica del Mioceno lacustre del sector Este de la localidad turolense deRubielos de Mora (Barranco de Alto Ballester), Tesis de Licenciatura (inédita), Universidad de Zaragoza.
RUBIO, C.; DÍEZ, J.B. y FERRER, J. (2000a), «Asociaciones fósiles representadas en los materiales miocenos deRubielos de Mora, Teruel. Un buen ejemplo del rico patrimonio paleontológico aragonés», Resúmenesdel I Congreso Ibérico de Paleontología/ XVI Jornadas de la Sociedad Española de Paleontología, pp. 185-186.
RUBIO, C.; ROIRON, P.; FERRER, J.; LIÑÁN, E.; DÍEZ, J.B.; POPESCU, S. y SUC, J.P. (2000b), «Nouvelles données sur lastratigraphie et les flores du bassin miocène de Rubielos de Mora (Teruel, Espagne)», en J. FERRER, J.B.DÍEZ, C. RUBIO y M.E. DIES (eds.), 10éme Colloque de l’Organisation FranVaise de Paléobotanique,Monografías de la Academia de Ciencias Exactas, Físicas, Químicas y Naturales de Zaragoza, 16, pp. 36-46.
SÁENZ, C. (1978), Introducción a la Palinología y vocabulario palinológico, Madrid, Blume.
SANCHIZ, F.B. (1977), «Catálogo de los anfibios fósiles de España (Noviembre de 1977)», Acta Geológica Hispánica,12, pp. 103-107.
SEILACHER, A. (1970), «Begriff und Bedeutung der Fossil-Lagertätten», Neves Jarhburg für Geologie und PaläleogieAbhandlungen, pp. 34-39.
SEYÍTOGLU, G. y BENDA, L. (1998), «Neogene palynological and isotopic age data from Selendi and UVak-Gürebasins, western Turkey: A contribution to the upper limit of eskihisar sporomorph association»,Newsletter on Stratigraphy, 36 (2), pp. 105-115.
SIMÓN, J.L. (1984), Compresión y distensión alpinas de la Cadena Ibérica oriental, Teruel, Instituto de EstudiosTurolenses.
THOMSON, P.W. y PFLUG, H. (1953), «Pollen und Sporen der mittel-europäischen Tertiärs», Palaeontographica,Abt. B., 94, pp. 1-138.
TRAVERSE, A. (1988), Paleopalynology, Allen and Unwin.
Recibido el 23 de junio de 2004Aceptado el 14 de septiembre de 2004
ESTUDIO PALEOPALINOLÓGICO DE RUBIELOS DE MORA (TERUEL) Y SUS IMPLICACIONES PALEOCLIMÁTICAS 91 [ I ] 2006-2007
53