erythropoietin and brain developmenterythropoietin acts to stimulate neural progenitor cells and to...
TRANSCRIPT

INTRODUCTION
Erythropoietin is a member of the haematopoietic cytokinesuperfamily that includes thrombopoietin, interleukin 3 (IL3),IL6, granulocyte-macrophage colony-stimulating factor (GM-CSF), prolactin, leukaemia inhibitory factor (LIF), ciliaryneurotrophic factor (CNTF) and cardiotrophin 1 (CT1),cytokines known to stimulate haematopoietic, neural or cardiacdevelopment. Erythropoietin receptor (EpoR) and othermembers of the corresponding cytokine receptor superfamilyare characterized by polypeptides with a single transmembranedomain and an extracellular domain with a WSXWSmotif (Youssoufian et al., 1993). During erythropoiesis,erythropoietin acts by binding to its receptor to stimulate theproliferation, differentiation and maturation of erythroidprogenitor cells. Erythropoietin activates phosphorylation ofJak2/STAT5 proteins as well as other signal transduction
pathways. Receptor homodimerization occurs uponerythropoietin binding, increasing the affinity of Jak2 for themembrane proximal region of the receptor, phosphorylatingJak2 and the cytoplasmic region of the receptor (Watowich etal., 1999).
Erythropoietin is required for the production of matureerythrocytes, the yolk sac being the first site of erythropoiesis(embryonic days 7-11 (E7-11)). In the embryo proper atE10, the aorta-gonad-mesonephros region gives rise tohaematopoietic stem cells responsible for definitivehaematopoiesis (Medvinsky and Dzierzak, 1996). By E12,the major site of erythropoiesis shifts to the liver. Theeffects of targeted deletion of erythropoietin receptor aredramatic when haematopoiesis shifts from the yolk sac tothe foetal liver and definitive erythropoiesis resulting incombined failure of definitive erythroid progenitors toproliferate and differentiate into mature erythrocytes. Death
505Development 129, 505-516 (2002)Printed in Great Britain © The Company of Biologists Limited 2002DEV3540
Erythropoietin, known for its role in erythroiddifferentiation, has been shown to be neuroprotectiveduring brain ischaemia in adult animal models. Althoughhigh levels of erythropoietin receptor are produced inembryonic brain, the role of erythropoietin during braindevelopment is uncertain. We now provide evidence thaterythropoietin acts to stimulate neural progenitor cells andto prevent apoptosis in the embryonic brain. Mice lackingthe erythropoietin receptor exhibit severe anaemia anddefective cardiac development, and die at embryonic day13.5 (E13.5). By E12.5, in addition to apoptosis in foetalliver, endocardium and myocardium, the erythropoietinreceptor null mouse shows extensive apoptosis in foetalbrain. Lack of erythropoietin receptor affects braindevelopment as early as E10.5, resulting in a reduction inthe number of neural progenitor cells and increasedapoptosis. Corresponding in vitro cultures of cortical cellsfrom Epor–/– mice also exhibited decreases in neuron
generation compared with normal controls and increasedsensitivity to low oxygen tension with no surviving neuronsin Epor–/– cortical cultures after 24 hour exposure tohypoxia. The viability of primary Epor+/+ rodentembryonic cortical neurons was further increased byerythropoietin stimulation. Exposure of these cultures tohypoxia induced erythropoietin expression and a tenfoldincrease in erythropoietin receptor expression, increasedcell survival and decreased apoptosis. Cultures of neuronalprogenitor cells also exhibited a proliferative response toerythropoietin stimulation. These data demonstrate thatthe neuroprotective activity of erythropoietin is observed asearly as E10.5 in the developing brain, and that inductionof erythropoietin and its receptor by hypoxia maycontribute to selective cell survival in the brain.
Key words: Erythropoietin, Receptor, Brain, Heart, Development,Neural Progenitor Cells, Mouse
SUMMARY
Erythropoietin receptor signalling is required for normal brain development
Xiaobing Yu 1, John J. Shacka 3, Jeffrey B. Eells 2, Carlos Suarez-Quian 4, Ronald M. Przygodzki 5, BojanaBeleslin-Cokic 1, Chyuan-Sheng Lin 6, Vera M. Nikodem 2, Barbara Hempstead 7, Kathleen C. Flanders 3,Frank Costantini 6 and Constance Tom Noguchi 1,*1Laboratory of Chemical Biology, NIDDK, National Institutes of Health, Bethesda, MD 20892, USA2Genetics and Biochemistry Branch, NIDDK, National Institutes of Health, Bethesda, MD 20892, USA3Laboratory of Cell Regulation and Carcinogenesis, NCI, National Institutes of Health, Bethesda, MD 20892, USA4Department of Cell Biology, Georgetown University Medical School, Washington, DC 20007, USA5Armed Forces Institute of Pathology, Washington, DC 20306, USA6Department of Genetics and Development, Columbia University, New York, NY 10032, USA7Cornel Medical College, New York, NY 10021, USA*Author for correspondence (e-mail: [email protected])
Accepted 23 October 2001

506
occurs at around E13.5 and is believed to result primarilyfrom severe anaemia that ensues in the foetuses (Lin et al.,1996; Wu et al., 1995).
The ability of thrombopoietin, prolactin/prolactin receptoror IL6/soluble IL6 receptor to replace erythropoietin duringerythropoiesis (Baiocchi et al., 2000; Kieran et al., 1996;Socolovsky et al., 1998) shows that stimulation by thehaematopoietic cytokines can be similar and may be morecrucial for proliferative than for instructive signalling. This isparticularly relevant to the production of EpoR in non-erythroid cells such as haematopoietic stem cells (Orlic et al.,1995), endothelium (Anagnostou et al., 1994; Ribatti et al.,1999), neuronal cells (Masuda et al., 1993; Morishita et al.,1997) and muscle (Ogilvie et al., 2000). EpoR is also requiredfor normal embryonic heart development and vascularization.In addition to compromised erythropoiesis, Epor–/– embryosexhibit ventricular hypoplasia and disrupted vasculature in theheart prior to death (Wu et al., 1999). Myoblasts and primarysatellite cells produce EpoR and exhibit a proliferativeresponse to erythropoietin that can interrupt differentiation intomyotubes in culture (Ogilvie et al., 2000). Administration oferythropoietin in vivo also stimulates neovascularizationand/or blood vessel formation in endothelial tissues such as thechick embryo chorioallantoic membrane (Ribatti et al., 1999)and murine uterine endometrium (Yasuda et al., 1998).Furthermore, in adult brain, erythropoietin administration isneuroprotective against ischaemic damage in vivo (Bernaudinet al., 1999; Brines et al., 2000; Sakanaka et al., 1998). Theseobservations provide evidence that erythropoietin activity isnot restricted to the erythroid lineage.
We have previously indicated that the expression oferythropoietin receptor transcripts in mouse brain peaks at mid-gestation, then subsequently decreases to modest levels in theadult (Liu et al., 1994). Endogenous erythropoietin receptorexpressed in cultured hippocampal and cerebral corticalneurons is functional and protects against glutamate-induceddamage (Masuda et al., 1993; Morishita et al., 1997).Erythropoietin also stimulates the proliferation of neuronalNT2 cells exposed to hypoxia and increases the number ofdopaminergic neurons in cultures of neuronal stem cells (Chinet al., 2000; Studer et al., 2000). In brain, production oferythropoietin has been detected in astrocytes and neurons(Juul et al., 1999; Masuda et al., 1994a), suggesting possibleautocrine or paracrine regulation by erythropoietin. Theinduction of erythropoietin production by hypoxia suggeststhat erythropoietin may be particularly relevant inneuroprotection during hypoxia (Digicaylioglu et al., 1995;Juul et al., 1998; Masuda et al., 1994a). Animal studies of brainischaemia show that erythropoietin administration providesprotection against hippocampal CA1 neuronal damage andmemory loss in gerbils (Sakanaka et al., 1998). In rats,erythropoietin has been shown to be neuroprotective in thecerebral cortex and to alleviate navigation disability caused bypermanent occlusion of the cerebral artery (Sadamoto et al.,1998).
The current study evaluates the function of EpoR in thedeveloping brain using mice with an Epor targeted deletion(Epor–/–) (Lin et al., 1996). We find that lack of EpoRsignalling affects brain development as early as E10.5,resulting in a reduction of neural progenitor cells and increasedapoptosis, and in a marked reduction in the survival of neuronal
cell cultures exposed to hypoxia. The antiapoptotic effect oferythropoietin on embryonic neuron cultures is also observed.These studies provide evidence that EpoR signalling extendsbeyond erythropoiesis and haematopoietic activity andprovides in vivo evidence that erythropoietin plays a role inneural progenitor cell survival and neuronal cell generation inthe developing brain.
MATERIALS AND METHODS
Production of Epor –/– miceHemizygous mice for the targeted deletion of the Epor gene (Epor–/–)(Lin et al., 1996) were mated and embryos harvested from pregnantfemales at various stages of gestation. Embryos were genotyped byPCR analysis of DNA isolated from yolk sac. The PCR primers usedto detect the null EpoR were: P1, GCC CCC TCT GTC TCC TACTT; P2, CGC CTC AAA ACC AGA AAC AG; P3, GAA GAG CTTGGC GGC GAA TG (Lin et al., 1996).
Production of Epor -lacZ transgenic miceA hybrid reporter gene was constructed consisting of theEpor 5′proximal promoter fragment extending from –1778 or –150 bp 5′ tothe first codon linked to the lacZ coding region, and transgenic micegenerated as previously described (Liu et al., 1997). Embryos wereharvested and fixed in 1% formaldehyde, 0.2% glutaraldehyde, 2 mMMgCl2, 5 mM EGTA and 0.02% NP-40 in PBS for 1 hour at 4°C.After three washes in PBS plus 0.02% NP-40 for 30 minutes each,embryos were stained overnight at 37°C in solution with 5 mMK3Fe(CN)6, 5 mM K4Fe(CN)6, 2 mM MgCl2, 0.01% sodiumdeoxylcholate, 0.02% NP-40, and 1 mg ml–1 X-gal. Stained embryoswere fixed and embedded in paraffin for sectioning. Nuclear fast redwas used as a counter stain.
ImmunohistochemistryEmbryos were fixed in 10% formalin, embedded in paraffin andsectioned. Sections were deparaffinized in xylene. Immunostainingwas carried out using polyclonal anti-EpoR (Santa CruzBiotechnology, Santa Cruz, CA) at a dilution of 1:400 at 4°Covernight. Tenfold excess blocking peptide was used to determine thespecificity of the primary antibody. Antibody binding was visualizedusing a biotinylated secondary antibody and an avidin/biotinylatedperoxidase complex (ABC) (Vector Laboratories, Burlingame, CA).Sections were counter stained with haematoxylin for visualization.Polyclonal antibody for von Willebrand (Factor VIII) (Dako,Carpinteria, CA) was used at a 1:1600 dilution. Forimmunocytochemistry of murine cortical cells, antibodies and dilutionwere as follows: MAP2 1:500 (Sigma, St. Louis, MO), β-tubulin typeIII (TuJ1) monoclonal 1:500 (Babco, Richmond, CA), nestin (Rat401) 1:1000 (BD PharMingen, San Diego, CA) and fluorescentlabelled secondary antibody. Cell nuclei were counterstained with0.001% DAPI (Sigma, St Louis, MO).
TUNEL assayFor terminal-deoxynucleotidyl transferase (TdT)-mediated dUTP-end-labelling (TUNEL) analysis of apoptosis (Gavrieli et al., 1992),sections were treated with proteinase K and processed in a reactionmixture buffer containing digoxigenin-labelled dUTP and TdT(Roche Molecular Biochemicals, Indianapolis, IN), incubated withanti-digoxigenin antibody conjugated to alkaline phosphatase (AP)(Dako, Carpinteria, CA), visualized with a New Fuchsin substratereaction and counter stained with haematoxylin. Sections incubatedwith the TUNEL reaction mixture but without TdT were served asnegative controls. For neuronal cells, cultures were fixed in cold,freshly prepared 4% paraformaldehyde solution for 1 hour at 25°Cand washed with PBS three times. After a 2 minute incubation in
X. Yu and others

507Erythropoietin and brain development
0.1% Triton X-100 and 0.1% sodium citrate, the cultures werewashed three times with PBS and then incubated in the TUNELreaction mixture for 2 hours at 37°C. Samples were analyzedunder a fluorescence microscope. The proportion of apoptoticprimary cortical neurons was quantified by counting the number ofneurons with and without fragmented DNA in premarkedmicroscope fields.
Cell cultureHuman neuronal NT2 cells (Pleasure et al., 1992) were maintainedin Opti-MEM I (Life Technologies, Gaithersburg, MD) with 10% or2% foetal bovine serum with 5% CO2 and 20% or 2% oxygentension. For isolation of cortical cells, the cortex from E10.5embryos was dissected out and placed into neurobasal dissectionmedia (1× Hanks’ balanced salt solution (HBSS) with 10 mMHEPES, 0.3% glucose, 0.72% sucrose, pH 7.2). The tissue wasdigested with papain (7 units ml–1 with 2.2 mM cysteine and 1 mMEDTA in dissection media) for 15 minutes at 37°C. The tissue waswashed to inactivate the papain and triturated with a glass Pasteurpipette. The cells were counted using trypan blue and were platedat a density of 2×105 cells per well in neurobasal medium with B27supplement (Life Technologies, Gaithersburg, MD) onto poly-L-ornithine/fibronectin-coated glass coverslips in 24 well tissueculture plate. Four days after plating, cells were fixed and stainedwith MAP2 or β-tubulin type III, and TUNEL.Primary cultures ofrat embryonic cortical neurons were prepared from E18 rat brains(Hampson et al., 1998; Priestley et al., 1990). The cortices weredissected and incubated for 10 minutes in a solution of 0.05% DNaseI and 0.25% trypsin (Life Technologies, Gaithersburg, MD). Thesample was treated with 0.5% soybean trypsin inhibitor (Sigma, StLouis, MO) in HBSS (Life Technologies, Gaithersburg, MD) to stopdigestion. Cells were dissociated by gentle trituration using Pasteurpipettes and resuspended in neurobasal medium (NBM) with the B-27 supplement (Life Technologies, Gaithersburg, MD), glutamine,and glutamate. Cells were initially plated at a density of 3.2×106 perwell in polylysine-coated 6-well plates. The neuronal content of theprimary cultures was verified by intense immunohistochemicalstaining of the cultures with anti-neurofilament protein (a neuronalmarker) and little staining with anti-glial fibrillary acidic protein (amarker for astroglial cells). Antibodies were obtained from RocheMolecular Biochemicals (Indianapolis, IN). On day 8, cells weresubjected to supplement starvation upon exchange of the media forLocke’s solution (154 mM NaCl, 5.6 mM KCl, 2.3 mM CaCl2, 1.0mM MgCl2, 3.6 mM NaHCO3, 5 mM glucose and 5 mM HEPESbuffer, pH 7.2) (Chan et al., 1999) and cultured with 5% CO2 and20% O2 or 2% O2 for 24-48 hours. Cell survival was quantified bycounting the total number of viable cells using trypan blue or bycounting the number of undamaged neurons in premarkedmicroscopic fields. TUNEL assay was used for the detection ofapoptotic nuclei.
RNA isolation and quantitative RT-PCR analysisTotal RNA was isolated from neuronal cultures using STAT-60 (Tel-Test, Friendswood, TX) treated with RNase-free DNase (Promega,Madison, WI) at 5 units per 100 µg of RNA at 37°C for 30 minutes,followed by phenol-chloroform extraction and ethanol precipitation.First-strand cDNA was synthesized using MuLV reverse transcriptaseand oligo-d(T)16 (PE Applied Biosystems, Foster City, CA).Quantitative real-time RT-PCR analyses were carried out todetermine the level of expression with the use of gene-specificprimers and fluorescent labelled Taqman probes or SYBR green dye(Molecular Probes, Eugene, OR) in a 7700 Sequence Detector (PEApplied Biosystems, Foster City, CA) (Ogilvie et al., 2000). Theprobe was designed to span exon junctions in order to prevent theamplification of any contaminating genomic DNA. Serial dilutionsof plasmid containing the cDNA of interests were used as templateto determine a standard curve. All results were normalized with
mouse S16 (mS16), rat S16 or human β-actin. The human EPORprimer sequences used were as follows: forward primer, 5′-GCT CCCTTT GTC TCC TGC T-3′; reverse primer, 5′-CTC CCA GAA ACACAC CAA GTC CT-3′; probe, 5′-AGC GGC CTT GCT GGC GG-3′. The rat Epor primer sequences were as follows: forward primer,5′-GAG AAT GAG TTT GAG GGT CTC TTC A-3′; reverse primer,5′-CCT CTA GGT GGG CAG GTG G-3′; probe, 5′-GGG TAA CTTCCA GCT ATG GCT GTT GCA AC-3′. The humanGATA3primersequences were as follows: forward primer, 5′-CGG CTT CGG ATGCAA GTC-3′; reverse primer, 5′-GTC GAG GTT GCC CCA CAG-3′; probe, 5′-AGG CCC GGT CCA GCA CAG AAG G-3′. Thehuman β-actin primers were as follows: forward primer, 5′-CCTGGC ACC CAG CAC AAT-3′; reverse primer, 5′-GCC GAT CCACAC GGA GTA CT-3′; probe, 5′-TCA AGA TCA TTG CTC CTCCTG AGC GC-3′. The mouse nestin primers were as follows:forward primer, 5′- GGC TAC ATA CAG GAT TCT GCT GG-3′;reverse primer, 5′-CAG GAA AGC CAA GAG AAG CCT-3′. RatBcl-xL: forward primer, 5′-ATC CAG GAG AAC GGC GG-3′;reverse primer, 5′-GGC TCT CGG GTG CTG TAT TG-3′; probe, 5′-TGG GAC ACT TTT GTG GAT CTC TAC GGG A-3′. Rat Epo:forward primer, 5′-GAG GTA CAT CTT GGA GGC CAA GGA-3′;reverse, 5′-TTC TCA CTC AGT CTG GGA CCT TC-3′;probe, 5′-GCA GAA AAT GTC ACA ATG GGC TGT GC-3′. The sequencesof mS16 and GATA3 primers and probes have been describedpreviously (Ogilvie et al., 2000).
Immunoprecipitation and western blottingFor immunoprecipitation experiments, primary cortical cellscultured for 8 days were changed to Locke’s solution for 16 hoursunder 20% or 2% oxygen tension, then were exposed to 5 U ml–1
of erythropoietin (Amgen, Thousand Oaks, CA) for 30 minutes.Cells were washed with cold PBS twice and then lysed in modifiedRIPA buffer (50 mM Tris-HCL, 150 mM NaCl, 1 mM EDTA, 1%NP-40, 0.25% sodium deoxycholate and protease inhibitors). Wholecell lysates (1 mg) were incubated with 4 µg of anti-phosphotyrosine4G10 antibody (Upstate Biotechnology, Lake Placid, NY) overnightat 4°C. Protein A agarose-captured immunocomplexes wereseparated on 4-12% Novex Bis-Tris NuPAGE Gels (Invitrogen,Carlsbad, CA) and transferred to PVDF membrane (Invitrogen,Carlsbad, CA) using a Novex Xcell II Mini-Cell and Blot Module(Invitrogen, Carlsbad, CA). Membranes were probed with anti-Jak2at 1:1000 dilution (Upstate Biotechnology, Lake Placid, NY) andanti-STAT5 at 1:250 dilution (BD Transduction Laboratories,Lexington, KY) followed by horseradish-peroxidase (HRP)-coupled secondary antibodies and developed by enhancedchemiluminescence (ECL) (Amersham Pharmacia Biotech,Piscataway, NJ).
Transfection into NT2 cellsThe Epor proximal promoter with or without GATA binding siteslinked to a luciferase reporter (Chin et al., 1995) was transfected intoNT2 cells using FuGENE 6 (Roche Molecular Biochemicals,Indianapolis, IN) in the presence or absence of a human GATA3expression vector. Luciferase activity was determined 48 hours aftertransfection. For stable overexpression ofGATA3, human GATA3cDNA was cloned into the pIRES2-EGFP expression vector(Clonetech Laboratories, Palo Alto, CA). The GATA3construct wastransfected into NT2 cells using FuGENE 6 (Roche MolecularBiochemicals, Indianapolis, IN). Transfected cells were grown inselection medium containing G418 (Life Technologies, Gaithersburg,MD) for 2 weeks and harvested for analysis. Cells transfected withpIRES2-EGFP without insert were used for control.
StatisticsData are expressed as means plus or minus standard error of the mean(SEM). Significance of differences was examined using the Student’st test. P values <0.05 were considered to be significant.
508
RESULTS
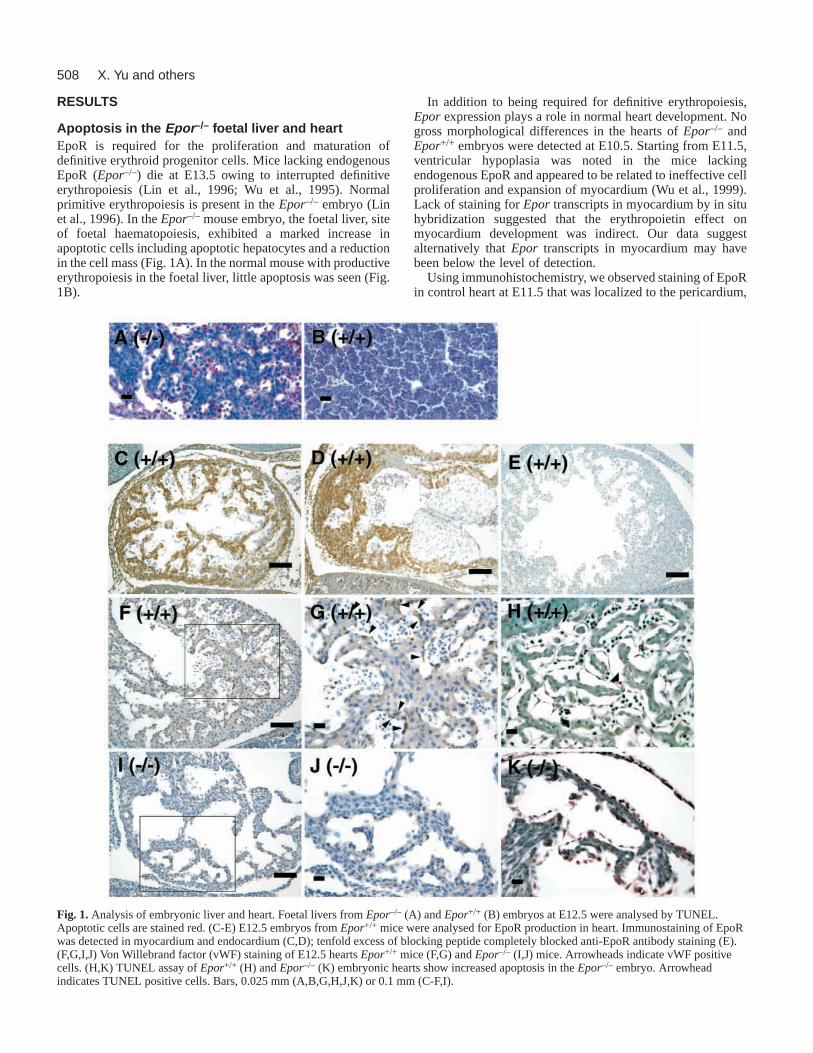
Apoptosis in the Epor –/– foetal liver and heartEpoR is required for the proliferation and maturation ofdefinitive erythroid progenitor cells. Mice lacking endogenousEpoR (Epor–/–) die at E13.5 owing to interrupted definitiveerythropoiesis (Lin et al., 1996; Wu et al., 1995). Normalprimitive erythropoiesis is present in the Epor–/– embryo (Linet al., 1996). In theEpor–/– mouse embryo, the foetal liver, siteof foetal haematopoiesis, exhibited a marked increase inapoptotic cells including apoptotic hepatocytes and a reductionin the cell mass (Fig. 1A). In the normal mouse with productiveerythropoiesis in the foetal liver, little apoptosis was seen (Fig.1B).
In addition to being required for definitive erythropoiesis,Epor expression plays a role in normal heart development. Nogross morphological differences in the hearts of Epor–/– andEpor+/+ embryos were detected at E10.5. Starting from E11.5,ventricular hypoplasia was noted in the mice lackingendogenous EpoR and appeared to be related to ineffective cellproliferation and expansion of myocardium (Wu et al., 1999).Lack of staining for Epor transcripts in myocardium by in situhybridization suggested that the erythropoietin effect onmyocardium development was indirect. Our data suggestalternatively that Epor transcripts in myocardium may havebeen below the level of detection.
Using immunohistochemistry, we observed staining of EpoRin control heart at E11.5 that was localized to the pericardium,
X. Yu and others
Fig. 1. Analysis of embryonic liver and heart. Foetal livers from Epor–/– (A) and Epor+/+ (B) embryos at E12.5 were analysed by TUNEL.Apoptotic cells are stained red. (C-E) E12.5 embryos from Epor+/+ mice were analysed for EpoR production in heart. Immunostaining of EpoRwas detected in myocardium and endocardium (C,D); tenfold excess of blocking peptide completely blocked anti-EpoR antibody staining (E).(F,G,I,J) Von Willebrand factor (vWF) staining of E12.5 hearts Epor+/+ mice (F,G) and Epor–/– (I,J) mice. Arrowheads indicate vWF positivecells. (H,K) TUNEL assay of Epor+/+ (H) and Epor–/– (K) embryonic hearts show increased apoptosis in the Epor–/– embryo. Arrowheadindicates TUNEL positive cells. Bars, 0.025 mm (A,B,G,H,J,K) or 0.1 mm (C-F,I).

509Erythropoietin and brain development
myocardium, and endocardium (Fig. 1C,D). A markedreduction in von Willebrand factor positive endothelial cellswas observed in the heart at E12.5 ofEpor–/– mice (Fig. 1I,J)compared with the normal control (Fig. 1F,G). We measuredTUNEL staining in the hearts of control and Epor–/– mice inorder to determine whether the decreased cell proliferationcaused by the lack of EpoR is related to loss of anti-apoptoticeffect of EpoR signalling, resulting in an increase in apoptosis.A TUNEL procedure was used to identify the apoptosis in situ(Fig. 1H,K). In the heart ofEpor–/– mice at E11.5, therewas significant increase of TUNEL-positive nuclei in themyocardium and endocardium (Fig. 1K). By comparison, onlyoccasional TUNEL-labelled nuclei were observed in the heartof control animals (Fig. 1H). The defects in the embryonicheart appear in the formation of both the compact zone(epicardial region) and the trabeculae. Thus, cardiac myocytesin both regions may requireerythropoietin signalling forsurvival/proliferation. This isconsistent with EpoR localization inthe normal embryonic heart (Fig.1C,D). EpoR production in normalembryos and increased apoptosisin Epor–/– embryonic heart arespecifically localized. Lack of
EpoR staining in the endocardial cushion (Fig. 1D) isconsistent with lack of cushion defects in the null mutant.
EpoR production in embryonic brainWe and others have shown production of EpoR in brain(Digicaylioglu et al., 1995; Juul et al., 1999; Liu et al., 1997;Liu et al., 1994; Marti et al., 1996; Yasuda et al., 1993) withparticularly high levels in the embryonic brain during mid-gestation (Liu et al., 1997; Liu et al., 1994). Immunostainingfor EpoR was performed in sections of wild-type embryos atdifferent stages. At E10.5, EpoR staining was broadly presentin the all layers of neuroepithelia in forebrain, including thecortex, midbrain and hindbrain (Fig. 2A). By E11.5, EpoR wasexpressed most prominently in neuroepithelia of midbrain andhindbrain (Fig. 2B-F). In midbrain, EpoR production waslocalized to the intermediate zone cells (Fig. 2C). In hindbrain,
Fig. 2.EpoR production in developingbrain. (A-G) Immunostaining ofEpor+/+ embryos for EpoR shows theproduction pattern of EpoR in thebrain at E10.5 (A), E11.5 (B-F) andE12.5 (G). The inset shows transgeneexpression of an Epor-lacZ promoter-reporter gene construct that providestissue-specific expression in regionscorresponding to endogenous EpoRproduction at E10.5 (A) and E12.5(G). (C-F) Increased magnifications ofB. (H,I) Hypoplasia ofneuroepithelium of the fourth ventriclewas observed in the E12.5 Epor–/–
embryo (I) compared with Epor+/+
(H). (J-Q) TUNEL analysis ofembryonic brain of Epor–/– (J-M) andEpor+/+ (N-Q) mice at E10.5(J,K,N,O) and E12.5 (L,M,P,Q).Increased apoptosis was observed forEpor–/– as early as E10.5. Increasedapoptotic cells are shown inneuroepithelium of E10.5 midbrain(coronal section) (J,K) and E12.5hindbrain (sagittal section) (L,M). Redblood cells (indicated by arrowheadand enlarged in the inset) wereobserved in the E10.5 neuroepithelium(K). (K,M,O,Q) Increasedmagnifications of J,L,N and P,respectively. Abbreviations: GE,ganglionic eminence; IV, fourthventricle; VZ, ventricular zone; IZ,intermediate zone. Bars, 1 mm (B,G),0.4 mm (A, inset A, inset G,H-J,L,N,P), 0.2 mm (K,O) and 0.05mm (C-F,M,O,Q).

510
EpoR was produced in intermediate zone and ventricular zone(Fig. 2F). EpoR production in neuroepothelium of the forebrainwas still present, although it is reduced. It was present in thesubventricular and ventricular zones of cortex (Fig. 2D). EpoRimmunoreactive cells were distributed through the intermediatezone of septum and ganglionic eminence (GE) (Fig. 2E). ByE12.5, strong staining was mainly localized to the intermediatezone of pons and medulla of the hindbrain region (Fig. 2G).During development, Epor–/– mice exhibited neuroepithelialtissue hypoplasia in the region of the pontine fixture adjacentto the fourth ventricle by E11.5, and this became furtherexaggerated at E12.5 (Fig. 2I) compared with the normalcontrol (Fig. 2H). The degenerative changes in the Epor–/–
nervous system were confirmed by TUNEL assay. As early asE10.5, Epor–/– mice showed increased apoptotic cells in thecerebral cortex (data not shown) and neuroepithelium of themidbrain (Fig. 2J,K) that was absent in the normal control (Fig.2N,O). Marked increases in TUNEL-positive cells withincreasing age was observed in regions of neuroepithelialtissues associated with EpoR production inEpor+/+ embryos(Fig. 2L,M). The ganglionic eminence that exhibited EpoRstaining by E11.5 in Epor+/+ embryos also showed increasedTUNEL-positive nuclei in Epor–/– mice (data not shown).
We have previously observed the highest level ofendogenous EpoR production and humanEPOR transgene
expression in the brain at E10.5 (Liu et al., 1997). To studythe function of EpoR signalling during neurogenesis, weexamined primary cell cultures ofEpor–/–, Epor+/– and
X. Yu and others
Fig. 3.Neuronal progenitor cells from Epor–/– cortex. (A) At E9.5,nestin expression is comparable in cortices of the Epor+/+ andEpor+/– (n=5), and Epor–/– embryos (n=3). (B) Single-cellsuspension isolated from E10.5 Epor–/– cerebral cortices (n=6)showed marked reduction in total cell numbers compared withEpor+/+ and Epor+/– cortices (n=13). (C) By E10.5, nestin mRNAlevel was downregulated by four times in the Epor–/– cortex (n=4)compared with Epor+/+ and Epor+/– (n=6). (D,E) A marked decreasein the numbers and proportion of nestin positive progenitor cellsacutely isolated from the E10.5 Epor–/– cortex (n=4) was alsoobserved compared with Epor+/+ and Epor+/– cortices (n=12).(F,J,K) Fewer neurons were produced from in vitro cultures ofEpor–/– cortical cells. Cells were stained for MAP2 (F) or β-tubulinIII (red) (J,K) and with DAPI (blue) after 4 days of culture in NBM.The proportions of cells with MAP2 positive staining are indicated in(F) for Epor–/– (n=5) and for Epor+/+ and Epor+/– (n=6).Representative fields for Epor+/+ cultures with 41% of 200 cells (J)and for Epor–/– cultures with 23% of 199 cells (K) with β-tubulin IIIpositive staining are shown. (G-I,L,M) EpoR production improvedneuronal cell survival under hypoxia. Six and 24 hours after cellswere switched to Locke’s solution and cultured under the hypoxiccondition of 2% oxygen tension, cells were stained with MAP2 (red),TUNEL (green) and DAPI (blue). After 6 hours, the number ofsurviving neurons was markedly decreased (G) and apoptoticneurons were significantly increased (H) in Epor–/– cultures (n=3).No surviving Epor–/– neurons were observed after 24 hours exposure(n=3) (G). About 10% of the surviving cells were neurons in theEpor+/+ cultures. Erythropoietin addition increased the survival ofEpor+/+ neurons producing EpoR (I); no erythropoietin response wasobserved in Epor–/– cultures. In all experiments (E-M), cells wereinitially plated at the same density. For cell enumeration, thepercentage of nestin (E) and MAP2 (F-I) positive cells isolated foreach individual embryo were determined by an investigator blind tothe genotypes by quantifying 20 microscopic fields documented bydigital camera images. A total of 2000-4000 cells were counted.Scale bars: 0.025 mm in L,M; 0.05 mm in J,K. (B-F) P < 0.01. Openbars represent Epor+/+ or Epor+/+ and Epor+/– cultures, as indicated,and solid bars represent Epor–/– cultures.

511Erythropoietin and brain development
Epor+/+ E10.5 cortical cells. Single-cell suspensions werecollected from E10.5 cerebral cortices. The total number ofcortical cells harvested fromEpor–/– embryos was half thatfrom Epor+/+ embryos, whereas the number fromEpor+/–
embryos was comparable to that from Epor+/+ (Fig. 3B). AtE10, the murine cerebral cortex is a single layer of germinalneuroepithelium composed predominantly of cells producingnestin, an intermediate filament protein normally found inneural precursor cells (Frederiksen and McKay, 1988; Lendahlet al., 1990). At E9.5, we found no differences in nestinproduction in the cortex of the Epor–/–, Epor+/+ or Epor+/–
embryos (Fig. 3A). By contrast, by E10.5, nestin productionin Epor–/– cortical cells fell by four times compared with thatof Epor+/+ cells (Fig. 3C). The levels in theEpor+/– corticalcells were comparable to those ofEpor+/+ cells. These datasuggest that, between E9.5 and E10.5, lack of EpoR signallingresults in a marked decrease in the proportion of neuralprogenitor cells. This was further supported by directexamination of nestin positive cells acutely harvested fromE10.5 cortex (Fig. 3D,E). TheEpor+/+ and Epor+/– embryosexhibited 71±1% nestin positive cells compared with 57±2%for Epor–/– embryos. Additional evidence for reduction in thecapacity for neuronal cell production in the Epor–/– cortex wasobtained by cultures of cortical cells harvested at E10.5. Cellsisolated fromEpor+/+, Epor+/– and Epor–/– embryos wereplated at the same density. After 4 days of culture, the numbersof total cells and MAP2 or β-tubulin-III immunoreactiveneurons were determined. The Epor–/– cultures contained 27%MAP2 immunoreactive neurons compared with 39.6% forEpor+/+ and 33.8% for Epor+/– cultures (Fig. 3F). Undernormoxic culture conditions, the proportion of TUNELpositive cells was similar in the three groups. During thehypoxia, erythropoietin production can be induced up to 100-fold in neurons. In erythroid cells EpoR signalling increasesnot only progenitor cell proliferation but also survival anddifferentiation, especially under hypoxia. To study the survivaleffect of EpoR signalling onEpor–/– cortical neurons, Epor–/–
cortical cells were grown for 4 days in culture and thenswitched into trophic-factor-free Locke’s solution andcultured at 2% O2 for an additional 6 or 24 hours. After 6-hour culture, the proportion of apoptotic Epor–/– neurons wassignificantly higher than that ofEpor+/+ neurons (Fig. 3H).The percentage of survivingEpor+/+ neurons was decreasedfrom 39.6±1.7% to 28.5±2.4%, whereas the percentage ofsurviving Epor–/– neurons was decreased from 26.5±0.9% to14.3±1.3% (Fig. 3G). After 24 hours, most of theEpor–/–
cortical cells became apoptotic and no MAP2 positive neuronswere observed (Fig. 3G,M). However, in the correspondingEpor+/+ cell culture, fewer apoptotic cells were observed and10.1±0.9% of surviving cells were neurons (Fig. 3G,L). TheEpor–/– neurons exhibited greater sensitivity and markedlylower survival to hypoxia treatment thanEpor+/+ neurons. Theerythropoietin effect onEpor+/+ and Epor–/– cortical neuronsunder hypoxia was also studied. Erythropoietin additionincreased Epor+/+ neurons survival after 24 hours of cultureunder hypoxia (Fig. 3I). In the presence of erythropoietin,14.2±1.2% of surviving cells were neurons, compared with10.1±0.9% in the absence of erythropoietin. However, noerythropoietin neuroprotective effect was observed in theEpor–/– cell culture, and no neurons survived. These dataindicate that EpoR signalling is important in neuronal cell
survival and the relative effects are even greater under hypoxicchallenge.
Erythropoietin stimulation of neuronal cellsReduction in neural progenitor cells in the Epor–/– brain isconsistent with the reduction in neural cell survival andincreased apoptosis owing to loss of viability associated witherythropoietin signalling. To examine the direct effect oferythropoietin on neural progenitor cell proliferation, weexamined cultured human NT2 neuronal cells that produceEpoR and are erythropoietin responsive. NT2 cultures wereexposed to reduced oxygen tension (2% O2) for three days andcell growth was monitored. Compared with control cultures,there was a modest decrease in cell growth by hypoxia. Bycontrast, cells cultured with erythropoietin (5 U ml–1) andhypoxia showed an increase in cell number 1.4 times that ofcultures without erythropoietin (Fig. 4A). We also observedthat low oxygen tension induced EpoR production more thantwofold (Fig. 4B). In erythroid progenitor cells, erythropoietininduces expression of the transcription factor GATA1, a zincfinger containing protein that activates several erythroidspecific genes including Epor(Chin et al., 1995). Other GATA-like proteins such as GATA3, which is required for normalbrain development (Pandolfi et al., 1995), show some structuralsimilarity to GATA1 and bind to similar DNA motifs, butexhibit different temporal and tissue specificity (Ko and Engel,1993; Merika and Orkin, 1993). We observed that NT2 cellsproduce GATA3 and that erythropoietin induces GATA3production (Fig. 4C). We also found that stable transfection ofan expression vector for GATA3 into NT2 cells resulted inincreased production of EpoR (Fig. 4D). Although we detectedGATA2 production in NT2 cells, no increase in EpoRproduction was observed when an expression vector forGATA2 was used (data not shown). The Epor proximalpromoter has a GATA consensus binding site that is requiredfor active Eporpromoter activity in erythroid cells. We usedtransfection assays of reporter gene constructs to show thatdeletion of this GATA binding site significantly decreased Eporpromoter activity in NT2 cells (Fig. 4E). Increased GATA3production inducedEpor promoter activity twofold. We alsofound that absence of EpoR signalling downregulated GATA3in E10.5 Epor–/– cortex (Fig. 4F).
To studyEpor promoter activity in vivo, we used a hybridtransgene consisting of a human Epor long (1778) or short(150) promoter with an intact GATA binding site fused toa lacZ reporter gene. As early as E10, three of 1778 linesshowed transgene expression broadly distributed in theneuroepithelium of midbrain and hindbrain, where endogenousEpor expression was present (Fig. 2A). At E12, transgeneexpression was observed in the intermediate zone of theneuroepithelium of pons and medulla around the fourthventricle in two of the 150 lines with the short Epor promoter(Fig. 2G). These regions corresponded to endogenous Eporexpression. The transgene expression was also active in thehaematopoietic tissues of these 150 lines at this stage. Thesedata provide evidence that theEpor promoter can drive Eportissue-specific expression.
Neuroprotective effect of erythropoietin inembryonic neuronal culturesExamination of primary embryonic cortical cell cultures

512
demonstrated that EpoR production was required for neuronalsurvival under hypoxia. When it binds to EpoR, erythropoietinstimulates neuron survival, but cortical cells from the mouseembryo were not enough for large-scale studies. To studyfurther the effect of erythropoietin on differentiated embryonicneuron survival, cortical neurons were isolated from ratembryos at E18 and cultured in neurobasal medium (NBM).
After 8 days in culture, cells were subjected to trophic factorwithdrawal, the media changed to Locke’s solution and cellscultured for an additional 24 hours. Addition of erythropoietin(5 U ml–1) with the change into Locke’s solution resulted in anincrease in cell viability by ~1.5 times (Fig. 5A). Whenincubation was carried out at reduced oxygen tension (2%oxygen), the effect was even greater (1.75 times increase). The
X. Yu and others
C.
00
GAT
A-3
/β-a
ctin
(F
old
chan
ge)
-Epo +Epo
A.
00
0.5
1.0
1.5C
ell c
ount
(F
old
chan
ge)
-Epo +Epo2% O2
D.
00Epo
R/β
-act
in (
Fol
d ch
ange
)
-GATA-3 +GATA-3
00
1
2
3
Epo
R/β
-act
in (
Fol
d ch
ange
)
20% O2 2% O2
B.
E.
Rel
ativ
e lu
cife
rase
act
ivity
00
1.0
EpoR ∆EpoR
0.5
1.5
2.0
2.5
EpoR
1
2
3
1
2
3
F.
GAT
A-3
/mS
16 (
Fol
d ch
ange
)
0EpoR: (+/+) (-/-)
0.5
1.0
1.5
LuciferaseSp1GATA
EpoR promoter
GATA-3: - +-
E10.5
NT2
NT2NT2
NT2NT2
Fig. 4. Erythropoietin stimulation of neuronal cells.(A) Erythropoietin (5 U ml–1) stimulated NT2 cell proliferation whencultured under 2% oxygen tension for 3 days. (B) Eporexpressionwas induced in NT2 cells cultured with 5% CO2 and 2% O2compared with 20% O2, determined by quantitative RT-PCR usingTaqman probes. (C) GATA3 production was induced byerythropoietin stimulation. (D) Eporexpression was induced in NT2cells overexpressing GATA3. (E) In transfection assays, the humanEporpromoter was linked to a luciferase reporter gene to assessEporpromoter activity in NT2 cells. GATA3 production inducedEporpromoter activity (EpoR). Mutation of the GATA motif(∆EpoR) markedly decreases promoter activity. (F) GATA3expression was downregulated in E10.5 Epor–/– cortex (solid bar)compared with expression in the Epor+/+ cortex (open bar).
Fig. 5. Effect of erythropoietin on the viability of embryonic neurons.(A) Cell viability was determined for primary embryonic rat corticalneurons supplement starved for 24 hours in Locke’s solution with orwithout erythropoietin (5 U ml–1) under normoxia (20% O2) andhypoxia (2% O2). (B) The percentage of cells undergoing apoptosiswas determined by TUNEL analysis. (C-E) Real-time RT-PCRquantitation was used to determine the levels of Epor (C),erythropoietin (Epo) (D) and Bcl-xL (E) gene expression in the cellsafter 24 hours of culture in Locke’s solution. Expression is normalizedto ribosomal protein S16 levels as control. (F) Induction ofphosphorylation of Jak2 and STAT5 in cortical neurons treated witherythropoietin cultured at 20% or 2% oxygen tension was examined.(A-C) Cultures with and without supplemental erythropoietin(5 U ml–1) are represented by solid and open bars, respectively.

513Erythropoietin and brain development
protective effect of erythropoietin on these primary embryonicneurons was also seen as a reduction in percentage of TUNELpositive cells (Fig. 5B). Addition of erythropoietin decreasedthe TUNEL positive cells incubated in Locke’s solution by afactor of 0.80 and 0.70 when cells were cultured at 20% and2% oxygen, respectively. The greater differences (P<0.05)observed at low oxygen tension provided evidence thaterythropoietin signalling can be protective even in local regionsof hypoxia or ischaemia. The anti-apoptotic protein Bcl-xL isa Bcl2 family member associated with the protective activityof erythropoietin (Silva et al., 1996). Although the alternateshort splicing form, Bcl-xS, inhibits the ability of Bcl2 toenhance survival (Boise et al., 1993), Bcl-xL, is the major formin the nervous system (Gonzalez-Garcia et al., 1995). We foundthat, like the induction of Bcl-xL during erythropoietinstimulation of erythropoiesis, erythropoietin increasedproduction of Bcl-xL in these primary neuronal cell cultures(Fig. 5E). Incubation of these embryonic cortical neurons at2% oxygen induced EpoR production tenfold (Fig. 5C),markedly greater than the induction observed for the NT2neuronal cell line. Erythropoietin was produced in thesecultures and incubation at low oxygen tension further increasedexpression fourfold (Fig. 5D), providing an example of theresponse of erythropoietin production to hypoxia in neuronalcells. To examine potential signalling pathways forerythropoietin-induced cortical neuron survival, lysates ofuntreated or erythropoietin-treated cortical cell cultures wereimmunoprecipitated with an anti-phosphotyrosine antibodyand immunoblotted with anti-Jak2 or anti-STAT5 antibody(Fig. 5F). As observed in erythroid progenitor cells, we foundthat erythropoietin induced Jak2 and STAT5 tyrosinephosphorylation in cortical neurons under normoxic or hypoxiccondition.
Initial cultures of embryonic cortical neurons in the presenceof NBM were not sensitive to erythropoietin. No change wasobserved on the viability of cells with trophic factors presentwith and without erythropoietin added during the initial phaseof culture (Fig. 6A). However, erythropoietin supplementationof NBM for 4 days prior to the switch to supplement starvationwas neuroprotective. Pretreatment for 4 days witherythropoietin while cells were grown in NBM was sufficientto reduce apoptosis when cells were supplement starved in theabsence of erythropoietin (Fig. 6B,C). The differences wereparticularly significant at low oxygen tension resulting in a30% increase in numbers of neurons in each field witherythropoietin treatment compared with the control (P<0.05)after 24 hours. The Bcl-xL protein level was also induced inthe cortical neuronal cultures by erythropoietin pretreatment(data not shown).
DISCUSSION
During erythropoiesis, erythropoietin binding to its receptorstimulates proliferation, survival, and differentiation oferythroid progenitor cells. Absence of erythropoietin or itsreceptor interrupts definitive erythropoiesis in the foetal liver(Lin et al., 1996; Wu et al., 1995) as well as development ofother specific organ systems (Wu et al., 1999), and tissuedefects extend beyond erythroid cell maturation. We describehere corresponding defects with Epor–/– embryos, specific
production of EpoR in the pericardium, endocardium andmyocardium of developing heart but not the endocardialcushion, possibly reflecting their distinct developmentalorigins (Fig. 1). We show explicitly that there is extensivethinning and apoptosis of the ventricular compact zone andtrabeculae zone, indicating a reduction in proliferation of themyocardium as well as endocardium by E12.5 in the Epor–/–
heart, corresponding to regions of EpoR staining in the Epor+/+
embryo. These data provide evidence that erythropoietin actsas a viability factor during cardiac development and isnecessary to prevent apoptosis and expansion or proliferationof myocardial and endocardial progenitor cells. The ability oferythropoietin to stimulate endocardium is consistent withearlier observations of EpoR production in endothelial cellsof vascular or aortic origin and of stimulation ofneovascularization (Anagnostou et al., 1990; Anagnostou et al.,1994). In addition to the effect on myocardium, erythropoietinalso stimulates myoblasts of skeletal muscle origin (Ogilvie etal., 2000). Erythropoietin promotes the proliferation of primarymuscle satellite cells and can interrupt myoblast differentiationto myotubes. The suggested role for erythropoietin in thesevaried tissues provides support for the hypothesis that, ratherthan being restricted to the erythroid lineage, erythropoietinacts more generally to stimulate select progenitor cells toproliferate and expand, and to prevent apoptosis duringdifferentiation. These observations add new light to thedetection of high levels of erythropoietin expression duringembryonic brain development.
75
50
25
0
Neu
rons
per
fiel
dA.
+Epo-Epo
20
10
0
C.
Neu
rons
per
fiel
d 2%O2
+Epo-Epo
+Epo-Epo
30
20
10
0
Neu
rons
per
fiel
dB. 20%O2
Fig. 6. Effect oferythropoietin pretreatmenton viability of embryonicneurons. (A) Primaryembryonic rat corticalneurons were treated with(closed bars) and without(open bars) erythropoietin (5U ml–1) on day 4 of cultureand cell viability determinedon day 8. (B,C) On day 8,cells were cultured inLocke’s solution withoutsupplemental erythropoietinunder normoxia (20% O2)(B) and hypoxia (2% O2)(C). After 24 hours,undamaged neurons werecounted in premarkedmicroscopic fields. At leastsix fields were observed foreach well, at least three wellswere studied in threeseparate experiments. A totalof 300-900 neurons in eachcondition were counted.

514
Increasing evidence suggests that erythropoietin signallingmay play a role in stimulating the development of multipleorgans. The production of EpoR in neuronal cells and theirresponse to erythropoietin (Fig. 4, Fig. 5, Fig. 6) (Chin et al.,2000; Juul et al., 1998; Masuda et al., 1993; Morishita et al.,1997; Studer et al., 2000), as well as the production oferythropoietin in astrocytes and neuronal cells (Juul et al.,1999; Masuda et al., 1994a), provide evidence forerythropoietin activity in the brain. During development, EpoRproduction in the brain peaks at mid-gestation. We found that,in the absence of EpoR production in Epor–/– mice, increasedapoptotic cells in the brain were first seen at E10.5 (Fig. 2).Neuroepithelium hypoplasia became apparent by E11.5. Theregions affected corresponded to high endogenous EpoRactivity in Epor+/+ embryos. To demonstrate that this is not dueto general hypoxia caused by severe anaemia and heart defects,we examined Epor–/– embryos at E9.5 and E10.5, prior tosignificant anaemia or heart defects in the embryo proper. Asearly as E10.5, reduction of neural progenitor cells in Epor–/–
embryonic cortex was observed. Fewer neurons were generatedfrom these cortical cells harvested from Epor–/– embryos andcultured in vitro compared to Epor+/+ neural progenitor cells(Fig. 3). By contrast, no differences were detected in corticalcells at E9.5 (Fig. 3A). These data suggest that erythropoietinreceptor signalling plays a role in the establishing neuralprogenitor cell population during neurogenesis, particularlyafter E9.5. Loss of EpoR does not affect progenitor cells ingeneral, but instead only those from select lineages or tissuetypes. For example, although the Epor–/– embryo is severelyanaemic by E12.5, the numbers of colony forming units(CFU)-granulocyte/macrophage and CFU-megakaryocyte inculture from E12.5 Epor–/– foetal livers were normal (Lin etal., 1996).
Erythropoietin production on both sides of the blood-brainbarrier is inducible by hypoxia (Digicaylioglu et al., 1995;Masuda et al., 1994b). During anaemic stress, in addition toincreased erythropoietin production, we observed inductionof EpoR in both haematopoietic tissue and brain (Liu et al.,1997). Increase of erythropoietin and induction of EpoR inhaematopoietic tissue is consistent with the increasederythropoietic response to hypoxia or anaemic stress, and raisethe possibility that, in the brain, erythropoietin may provide, inpart, an analogous neuroprotective effect. Animal studiesdemonstrated that erythropoietin can be protective in the adultbrain to ischaemic challenge, particularly when increased bydirect administration (Brines et al., 2000; Sakanaka et al.,1998). We now show that low oxygen has a specific effect onneuronal cells to induce both erythropoietin and EpoRexpression, and in the absence of EpoR expression, neuronalcell survival is markedly affected by hypoxic treatment.This suggests a corresponding neuroprotective role forerythropoietin receptor signalling during brain development.
By E12.5, as with embryonic heart and erythroiddevelopment, the Epor–/– genotype is associated withsignificant apoptotic activity in the brain corresponding toregions of EpoR expression in Epor+/+ embryos (Fig. 2). Theneuroprotective effect of EpoR signalling was already evidentin cultures of E10.5 cortical cells. When E10.5 cortical cellswere cultured under normal differentiation medium, there wasno significant difference between the apoptosis observed inEpor–/– and Epor+/+ surviving neurons.
Cortical neurons were cultured under hypoxia and in theabsence of trophic factors to study the survival of neuronsdifferentiated from cortical cells. We observed a significantincrease in apoptosis of ~40% in Epor–/– neurons comparedwith <10% in Epor+/+ neurons after 6 hours. The proportionof surviving Epor+/+ neurons decreased to 0.7 after 6 hoursand to 0.25 after 24 hours incubation. Corresponding culturesisolated from Epor–/– embryos were markedly more affected,with the proportion of surviving neurons decreasing to 0.5after 6 hours and no neurons surviving after 24 hours ofincubation (Fig. 3). Erythropoietin addition further increasedthe survival of Epor+/+ neurons but not the Epor–/– neurons.The neuroprotective effect of EpoR signalling was clearlyevident in these E10.5 cortical cell cultures. EpoR productionprovides for neuron survival under hypoxia challenge. It alsosuggests that the neuroprotective effect of erythropoietin ismediated by its unique binding with EpoR. A specific survivalrole for erythropoietin and induction of EpoR by hypoxia werealso demonstrated in cultures of cortical neurons from normalanimals. These cultures showed increased viability andreduced apoptosis when cultured with supplementalerythropoietin and in the absence of trophic factors (Fig. 5,Fig. 6). In addition to erythropoietin expression in brain byastrocytes and neurons, the induction of EpoR production inthe neuronal cultures when exposed to low oxygen tensionsuggests that the erythropoietin response during hypoxiachallenge to the brain may involve both an upregulation oferythropoietin production and an increase in neuronalsensitivity to erythropoietin mediated by increasing EpoRproduction. These data support the hypothesis thaterythropoietin can act as a survival factor for neurons and canplay a role in stress response to hypoxia or ischaemia.
A principal function of erythropoietin is to rescue committederythroid progenitor cells from apoptosis (Kelley et al., 1993;Koury and Bondurant, 1990). Erythropoietin binding to itsreceptor in differentiating haematopoietic cells activatesJak/STAT and other signal transduction pathways to controlcellular proliferation, survival and specific gene expression.Erythropoietin also stimulates Jak2 phosphorylation inendothelial and muscle cells (Ogilvie et al., 2000; Ribatti et al.,1999), and we observed stimulation of Jak2 phosphorylationin cultured cortical neurons (Fig. 5F). Bcl-x prevents massivecell death in neurons and foetal liver and death in utero(Motoyama et al., 1995), and Bcl-xL has been implicated inantiapoptotic effects mediated by erythropoietin on erythroidprogenitor cells (Silva et al., 1996). Erythropoietin wassufficient to induce Bcl-xL in primary neurons when culturedwithout trophic factors under low oxygen tension (Fig. 5),suggesting that the ability of erythropoietin to induce Bcl-xLexpression may be among the critical factors in neuronal cellsurvival.
Analysis of transgenic embryos showed that EpoRpromoter drives specific transgene expression in braincorresponding to regions of endogenous EpoR productionduring development (Fig. 2). GATA1 and SP1 binding sitesare primary features of minimal promoter activity in erythroidcells (Chin et al., 1995). During erythropoiesis,erythropoietin stimulation of erythroid progenitor cellsinduces the production of the transcription factor GATA1,which is required for erythropoiesis and transactivation of amany erythroid specific genes including those for globin and
X. Yu and others

515Erythropoietin and brain development
EpoR itself (Chiba et al., 1991; Chin et al., 1995; Zon et al.,1991). The GATA family of zinc-finger transcription factorsbinds to similar consensus sequences (A/TGATAA/G) (Koand Engel, 1993; Merika and Orkin, 1993). In the brain, EpoRexpression is related in part to the role of GATA3. Targetingof GATA3 results in severe deformities of the brain (Pandolfiet al., 1995). We observed that erythropoietin could induceGATA3 production in NT2 cells (Fig. 4) and in other non-haematopoietic cells (Ogilvie et al., 2000). In addition,overproduction of GATA3 in NT2 cells increases EpoRproduction. These effects are analogous to the interactionbetween GATA1 and EpoR in erythroid progenitor cells.Reporter gene transfection assays showed that, in neuronalcells, the GATA consensus sequence in the Eporpromoterwas required for its expression and that GATA3 induced Eporpromoter activity (Fig. 4). Mutating the GATA consensussequence in the Eporpromoter diminished transactivity ofGATA3. Furthermore, an Eporpromoter containing theGATA binding site was active in embryonic brain in regionscorresponding to EpoR production (Fig. 2). Interestingly,during early neurogenesis at E10.5, GATA3 production wasdownregulated in the Epor–/– cortical cells. These data raisethe possibility that the effects of erythropoietin reported herefor developing brain and in other progenitor cells may begenerally mediated in part via induced expression of theGATA-like transcription factors.
We present here evidence that erythropoietin plays a rolein brain and neuronal development and that production ofEpoR in neuronal tissues is required for erythropoietin tostimulate the survival and provide for proliferation ofcorresponding progenitor cells. The induction oferythropoietin and its receptor by hypoxia suggests that forerythropoietin administration has therapeutic potential fortissue damage from ischaemia or hypoxic challenge in thecentral nervous system (Bernaudin et al., 1999; Brines et al.,2000; Sakanaka et al., 1998) and demonstrate that theprotective role of erythropoietin is more general and extendsbeyond haematopoiesis. Furthermore, the induction of EpoRby hypoxia in select progenitor cells allows for specificity ofcell response to increases in erythropoietin levels. Forexample, in cultures of embryonic neural stem cells, hypoxiaas well as erythropoietin stimulates the selective increase indopaminergic neurons (Studer et al., 2000). Neuronal cellsappear to share common regulatory elements withhaematopoietic cells in the production of EpoR. Themultiple-organ response to erythropoietin that includeshaematopoietic, neuronal, endothelial and muscle cells maybe a consequence of the similarities of respectivedifferentiating stem cells (Bjornson et al., 1999; Gussoni etal., 1999; Jackson et al., 1999). The common effect oferythropoietin to promote cell viability and to provide for cellproliferation may relate to the plasticity of tissue derived stemcells, particularly to that of the neural stem cells, which canalso contribute to haematopoietic and muscle tissue whentransplanted in vivo (Bjornson et al., 1999; Galli et al., 2000).Therefore, insights obtained from haematopoietic progenitorcell differentiation may provide a window on neuronal celldifferentiation.
We thank M. Yamamoto for human GATA3 cDNA and A. N.Schechter for helpful discussion.
REFERENCES
Anagnostou, A., Lee, E. S., Kessimian, N., Levinson, R. and Steiner, M.(1990). Erythropoietin has a mitogenic and positive chemotactic effect onendothelial cells. Proc. Natl. Acad. Sci. USA87, 5978-5982.
Anagnostou, A., Liu, Z., Steiner, M., Chin, K., Lee, E. S., Kessimian, N.and Noguchi, C. T.(1994). Erythropoietin receptor mRNA expression inhuman endothelial cells. Proc. Natl. Acad. Sci. USA91, 3974-3978.
Baiocchi, M., Marcucci, I., Rose-John, S., Serlupi-Crescenzi, O. andBiffoni, M. (2000). An Il-6/Il-6 soluble receptor (Il-6r) hybrid protein (H-Il-6) induces Epo-independent erythroid differentiation in human Cd34(+)cells. Cytokine12, 1395-1399.
Bernaudin, M., Marti, H. H., Roussel, S., Divoux, D., Nouvelot, A.,MacKenzie, E. T. and Petit, E. (1999). A potential role for erythropoietinin focal permanent cerebral ischemia in mice. J. Cereb. Blood Flow Metab.19, 643-651.
Bjornson, C. R., Rietze, R. L., Reynolds, B. A., Magli, M. C. and Vescovi,A. L. (1999). Turning brain into blood: a hematopoietic fate adopted byadult neural stem cells in vivo. Science283, 534-537.
Boise, L. H., Gonzalez-Garcia, M., Postema, C. E., Ding, L., Lindsten, T.,Turka, L. A., Mao, X., Nunez, G. and Thompson, C. B. (1993). bcl-x, abcl-2-related gene that functions as a dominant regulator of apoptotic celldeath. Cell 74, 597-608.
Brines, M. L., Ghezzi, P., Keenan, S., Agnello, D., de Lanerolle, N. C.,Cerami, C., Itri, L. M. and Cerami, A. (2000). Erythropoietin crosses theblood-brain barrier to protect against experimental brain injury. Proc. Natl.Acad. Sci. USA97, 10526-10531.
Chan, S. L., Tammariello, S. P., Estus, S. and Mattson, M. P. (1999).Prostate apoptosis response-4 mediates trophic factor withdrawal-inducedapoptosis of hippocampal neurons: actions prior to mitochondrialdysfunction and caspase activation. J. Neurochem. 73, 502-512.
Chiba, T., Ikawa, Y. and Todokoro, K. (1991). GATA-1 transactivateserythropoietin receptor gene and erythropoietin receptor-mediatedsignals enhance GATA-1gene expression. Nucleic Acids Res. 19, 3843-3848.
Chin, K., Oda, N., Shen, K. and Noguchi, C. T. (1995). Regulation oftranscription of the human erythropoietin receptor gene by proteins bindingto GATA-1 and Sp1 motifs. Nucleic Acids Res. 23, 3041-3049.
Chin, K., Yu, X., Beleslin-Cokic, B., Liu, C., Shen, K., Mohrenweiser, H.W. and Noguchi, C. T. (2000). Production and processing of erythropoietinreceptor transcripts in brain. Brain Res. Mol. Brain Res. 81, 29-42.
Digicaylioglu, M., Bichet, S., Marti, H. H., Wenger, R. H., Rivas, L. A.,Bauer, C. and Gassmann, M. (1995). Localization of specificerythropoietin binding sites in defined areas of the mouse brain. Proc. Natl.Acad. Sci. USA92, 3717-3720.
Frederiksen, K. and McKay, R. D. (1988). Proliferation and differentiationof rat neuroepithelial precursor cells in vivo. J. Neurosci. 8, 1144-1151.
Galli, R., Borello, U., Gritti, A., Minasi, M. G., Bjornson, C., Coletta, M.,Mora, M., De Angelis, M. G., Fiocco, R., Cossu, G. et al. (2000). Skeletalmyogenic potential of human and mouse neural stem cells. Nat. Neurosci.3, 986-991.
Gavrieli, Y., Sherman, Y. and Ben-Sasson, S. A. (1992). Identification ofprogrammed cell death in situ via specific labeling of nuclear DNAfragmentation. J. Cell Biol.119, 493-501.
Gonzalez-Garcia, M., Garcia, I., Ding, L., O’Shea, S., Boise, L. H.,Thompson, C. B. and Nunez, G. (1995). bcl-xis expressed in embryonicand postnatal neural tissues and functions to prevent neuronal cell death.Proc. Natl. Acad. Sci. USA92, 4304-4308.
Gussoni, E., Soneoka, Y., Strickland, C. D., Buzney, E. A., Khan, M. K.,Flint, A. F., Kunkel, L. M. and Mulligan, R. C. (1999). Dystrophinexpression in the mdxmouse restored by stem cell transplantation. Nature401, 390-394.
Hampson, A. J., Grimaldi, M., Axelrod, J. and Wink, D. (1998).Cannabidiol and (–)Delta9-tetrahydrocannabinol are neuroprotectiveantioxidants. Proc. Natl. Acad. Sci. USA95, 8268-8273.
Jackson, K. A., Mi, T. and Goodell, M. A. (1999). Hematopoietic potentialof stem cells isolated from murine skeletal muscle. Proc. Natl. Acad. Sci.USA96, 14482-14486.
Juul, S. E., Anderson, D. K., Li, Y. and Christensen, R. D. (1998).Erythropoietin and erythropoietin receptor in the developing human centralnervous system. Pediatr. Res. 43, 40-49.
Juul, S. E., Yachnis, A. T., Rojiani, A. M. and Christensen, R. D. (1999).Immunohistochemical localization of erythropoietin and its receptor in thedeveloping human brain. Pediatr. Dev. Pathol. 2, 148-158.

516
Kelley, L. L., Koury, M. J., Bondurant, M. C., Koury, S. T., Sawyer, S. T.and Wickrema, A. (1993). Survival or death of individual proerythroblastsresults from differing erythropoietin sensitivities: a mechanism forcontrolled rates of erythrocyte production. Blood82, 2340-2352.
Kieran, M. W., Perkins, A. C., Orkin, S. H. and Zon, L. I. (1996).Thrombopoietin rescues in vitro erythroid colony formation from mouseembryos lacking the erythropoietin receptor. Proc. Natl. Acad. Sci. USA93,9126-9131.
Ko, L. J. and Engel, J. D. (1993). DNA-binding specificities of the GATAtranscription factor family. Mol. Cell. Biol.13, 4011-4022.
Koury, M. J. and Bondurant, M. C. (1990). Erythropoietin retards DNAbreakdown and prevents programmed death in erythroid progenitor cells.Science248, 378-381.
Lendahl, U., Zimmerman, L. B. and McKay, R. D. (1990). CNS stem cellsexpress a new class of intermediate filament protein. Cell60, 585-595.
Lin, C. S., Lim, S. K., D’Agati, V. and Costantini, F. (1996). Differentialeffects of an erythropoietin receptor gene disruption on primitive anddefinitive erythropoiesis. Genes Dev.10, 154-164.
Liu, Z. Y., Chin, K. and Noguchi, C. T. (1994). Tissue specific expressionof human erythropoietin receptor in transgenic mice. Dev. Biol. 166, 159-169.
Liu, C., Shen, K., Liu, Z. and Noguchi, C. T. (1997). Regulated humanerythropoietin receptor expression in mouse brain. J. Biol. Chem. 272,32395-32400.
Marti, H. H., Wenger, R. H., Rivas, L. A., Straumann, U., Digicaylioglu,M., Henn, V., Yonekawa, Y., Bauer, C. and Gassmann, M. (1996).Erythropoietin gene expression in human, monkey and murine brain. Eur.J. Neurosci. 8, 666-676.
Masuda, S., Nagao, M., Takahata, K., Konishi, Y., Gallyas, F., Jr, Tabira,T. and Sasaki, R.(1993). Functional erythropoietin receptor of the cellswith neural characteristics. Comparison with receptor properties of erythroidcells. J. Biol. Chem. 268, 11208-11216.
Masuda, S., Okano, M., Yamagishi, K., Nagao, M., Ueda, M. and Sasaki,R. (1994a). A novel site of erythropoietin production. Oxygen-dependentproduction in cultured rat astrocytes. J. Biol. Chem. 269, 19488-19493.
Masuda, S., Okano, M., Yamagishi, K., Nagao, M., Ueda, M. and Saski,R. (1994b). A novel site of erythropoietin production: oxygen-dependentproduction in cultured rat astrocytes. J. Biol. Chem.269, 19488-19493.
Medvinsky, A. and Dzierzak, E. (1996). Definitive hematopoiesis isautonomously initiated by the AGM region. Cell 86, 897-906.
Merika, M. and Orkin, S. H. (1993). DNA-binding specificity of GATAfamily transcription factors. Mol. Cell. Biol.13, 3999-4010.
Morishita, E., Masuda, S., Nagao, M., Yasuda, Y. and Sasaki, R. (1997).Erythropoietin receptor is expressed in rat hippocampal and cerebral corticalneurons and erythropoietin prevents in vitro glutamate-induced neuronaldeath. Neuroscience76, 105-116.
Motoyama, N., Wang, F., Roth, K. A., Sawa, H., Nakayama, K., Negishi,I., Senju, S., Zhang, Q., Fujii, S. and Loh, D. Y. (1995). Massive cell deathof immature hematopoietic cells and neurons in Bcl-x-deficient mice.Science267, 1506-1510.
Ogilvie, M., Yu, X., Nicolas-Metral, V., Pulido, S. M., Liu, C., Ruegg, U.T. and Noguchi, C. T. (2000). Erythropoietin stimulates proliferation andinterferes with differentiation of myoblasts. J. Biol. Chem. 275, 39754-39761.
Orlic, D., Anderson, S., Biesecker, L. G., Sorrentino, B. P. and Bodine,D. M. (1995). Pluripotent hematopoietic stem cells contain high levelsof mRNA for c-kit, GATA-2, p45 NF-E2 and c-myb and low levelsor no mRNA for c-fms and the receptors for granulocyte colony-
stimulating factor and interleukins 5 and 7. Proc. Natl. Acad. Sci. USA92,4601-4605.
Pandolfi, P. P., Roth, M. E., Karis, A., Leonard, M. W., Dzierzak, E.,Grosveld, F. G., Engel, J. D. and Lindenbaum, M. H. (1995). Targeteddisruption of the GATA3 gene causes severe abnormalities in the nervoussystem and in fetal liver haematopoiesis. Nat. Genet. 11, 40-44.
Pleasure, S. J., Page, C. and Lee, V. M. (1992). Pure, postmitotic, polarizedhuman neurons derived from NTera 2 cells provide a system for expressingexogenous proteins in terminally differentiated neurons. J. Neurosci. 12,1802-1815.
Priestley, T., Horne, A. L., McKernan, R. M. and Kemp, J. A. (1990). Theeffect of NMDA receptor glycine site antagonists on hypoxia-inducedneurodegeneration of rat cortical cell cultures. Brain Res.531, 183-188.
Ribatti, D., Presta, M., Vacca, A., Ria, R., Giuliani, R., Dell’Era, P., Nico,B., Roncali, L. and Dammacco, F. (1999). Human erythropoietin inducesa pro-angiogenic phenotype in cultured endothelial cells and stimulatesneovascularization in vivo. Blood93, 2627-2636.
Sadamoto, Y., Igase, K., Sakanaka, M., Sato, K., Otsuka, H., Sakaki, S.,Masuda, S. and Sasaki, R. (1998). Erythropoietin prevents placenavigation disability and cortical infarction in rats with permanentocclusion of the middle cerebral artery. Biochem. Biophys. Res. Commun.253, 26-32.
Sakanaka, M., Wen, T. C., Matsuda, S., Masuda, S., Morishita, E., Nagao,M. and Sasaki, R.(1998). In vivo evidence that erythropoietin protectsneurons from ischemic damage. Proc. Natl. Acad. Sci. USA95, 4635-4640.
Silva, M., Grillot, D., Benito, A., Richard, C., Nunez, G. and Fernandez-Luna, J. L. (1996). Erythropoietin can promote erythroid progenitorsurvival by repressing apoptosis through Bcl-XL and Bcl-2. Blood88, 1576-1582.
Socolovsky, M., Fallon, A. E. and Lodish, H. F. (1998). The prolactinreceptor rescues EpoR–/– erythroid progenitors and replaces EpoR in asynergistic interaction with c-kit. Blood92, 1491-1496.
Studer, L., Csete, M., Lee, S. H., Kabbani, N., Walikonis, J., Wold, B. andMcKay, R. (2000). Enhanced proliferation, survival and dopaminergicdifferentiation of CNS precursors in lowered oxygen. J. Neurosci. 20, 7377-7383.
Watowich, S. S., Liu, K. D., Xie, X., Lai, S. Y., Mikami, A., Longmore, G.D. and Goldsmith, M. A. (1999). Oligomerization and scaffoldingfunctions of the erythropoietin receptor cytoplasmic tail. J. Biol. Chem. 274,5415-5421.
Wu, H., Liu, X., Jaenisch, R. and Lodish, H. F. (1995). Generation ofcommitted erythroid BFU-E and CFU-E progenitors does not requireerythropoietin or the erythropoietin receptor. Cell83, 59-67.
Wu, H., Lee, S. H., Gao, J., Liu, X. and Iruela-Arispe, M. L. (1999).Inactivation of erythropoietin leads to defects in cardiac morphogenesis.Development126, 3597-3605.
Yasuda, Y., Masuda, S., Chikuma, M., Inoue, K., Nagao, M. and Sasaki,R. (1998). Estrogen-dependent production of erythropoietin in uterus andits implication in uterine angiogenesis. J. Biol. Chem. 273, 25381-25387.
Yasuda, Y., Nagao, M., Okano, M., Masuda, S., Sasaki, R. and Konishi,H. (1993). Localization of erythropoietin and erythropoietin-receptor inpostimplantation mouse embryos. Dev. Growth Differ.35, 711-722.
Youssoufian, H., Longmore, G., Neumann, D., Yoshimura, A. and Lodish,H. F. (1993). Structure, function and activation of the erythropoietinreceptor. Blood81, 2223-2236.
Zon, L. I., Youssoufian, H., Mather, C., Lodish, H. F. and Orkin, S. H.(1991). Activation of the erythropoietin receptor promoter by transcriptionfactor GATA-1. Proc. Natl. Acad. Sci. USA88, 10638-10641.
X. Yu and others