erhan's taxol project-2007
DESCRIPTION
Taxol-lipid interactionsTRANSCRIPT

97
PACLITAXEL (TAXOL’un) ANTİ-KANSER ETKİ MEKANİZMASININ MODEL LANGMUİR-BLODGETT BİYOMEMBRAN SİSTEMİ İLE
MOLEKÜLER DÜZEYDE İNCELENMESİ
Dr. Erhan Süleymanoğlu
Gazi Üniversitesi,
Eczacılık Fakültesi,
Merkez Laboratuvarı ve Farmasötik Kimya Anabilim Dalı,
Hipodrom,
06330-Ankara
ANKARA
2007

98
İÇİNDEKİLER
Proje Özet Bilgi Formu…………………………………………………………………………………….1 Proje ile ilgili Genel Hususlar……………………………………………………………………………3 Motivasyon ve Ankara’da Bulunan Araştırma Olanakları…………………………………….4
1. Giriş……………………………………………………………………………………………………..7 1.1. Kanser……………………………………………………………………………………….…….7 1.2. Kanserin Tedavi Şemaları……………………………………………………………………………8 1.2.1. Sistemik Tedavinin Temel Prensipleri…………………………………………………………..8 1.2.2. Kemoterapinin Prensipleri…………………………………………………………………………..9 1.2.3. Sistemik Moleküler Hedefli Terapilerin Temel Prensipleri…………………………….19 1.2.4. Gen Aktarımı ve Kanser Tedavilerindeki Rolü………………………………………………22 1.2.5. Gen Tedavisi ve Gen Nakli………………………………………………………………………….24 1.2.6. DNA Kompaktizasyonu ve Nanoparçacık Oluşumunu
Meydana Getiren Bileşenler………………………………………………………………………..33 1.3. Taxol® (Paclitaxel)…………………………………………………….……………………………….43 1.3.1. Kanser Tedavisinde ve Kanserin Önlenmesinde Taxol®‘un Rolü…………………….43 1.3.2. Taxol®’un ve Taxotere™’in Klinik Kullanımı……………………………………………..…..46 1.3.4. Taxol®’un Etki Mekanizması……………………………………………………………………….54 1.3.5. Projemin Bilimsel Dayanağı………………………………………………………………………..58 1.3.6. Literatür Özeti…………………………………………………………………………………………...60 2. Gereç ve Yöntem..………………………………………………………………………………………….61 2.1. Hücre Membranın Yapısı…………………………………………………………………………………61 2.1.1. Biyomembranların İçeriği……………………………………………………………………………..61 2.2. Biyolojik Membranların Sıvı Kristal Özellikleri………………………………………………….66 2.2.1. Lipid Çift Katman…………………………………………………………………………………………66 2.2.2. Membranın Fizikokmyasal Özelliklerini Etkileyen Faktörler…………………………….67 2.2.3. Lipid Fazlarını Etkileyen Faktörler ve Biyosistemlerdeki Önemi……………………….70 2.2.4. İlâçların Biyomembran Akışkanlığı Üzerine Etkisi…………………………………………..71 2.2.5. Biyomembran Akışkanlığının Anlamı……………………………………………………………..73 2.3. Biyomembran ve Model Membran Araştırmalarında
Kullanılan Başlıca Yöntemler……………………………………………………………………………..80 2.3.1. Biyofiziksel Yöntemler……………………………………………………………………………………81 2.3.1.1. Spektroskopik Yöntemler……………………………………………………………………………..81 2.3.1.2. Difference Spectroscopy………………………………………………………………………………84 2.3.1.3. Fourier Transform Kızılötesi (FTIR) Spektroskopisi………………………………………84 2.3.1.4. Raman Spektroskopisi………………………………………………………………………………..85 2.4. Langmuir-Blodgett Monolayer Yöntemi…………………………………………………………….85 2.4.1. Yüzeysel Gerilimin Wilhelmy Plate Yöntemiyle Ölçülmesi…………………………………87 2.4.2. Lipid Tek Katmanların Özelliklerinin İzoterm Yöntemiyle İncelenmesi……………..88 2.4.3. Sub-fazda Madde-Lipid Etkileşmelerinin İncelenmesi ve Lipid Moleküllerinin İşgal Etileri Alanın Ölçülmesi……………………………………………..88 2.4.4. Tek Katmanda Yer Alan Maddeler Arasındaki Etkileşmelerinin İncelenmesi…………………………………………………………………………….89 2.4.5. Tek Katmanlarda Maddelerin Karışım Oluşturma Özelliklerinin İncelenmesi…………………………………………………………………………………90 2.4.6. Tek Katmanlarda Bulunan Moleküllerin Yapısal Değişikliklerinin İncelenmesi……………………………………………………………………………..91 2.4.7. Tek Katmanın Sabitlilik ve Deformasyon Özelliklerinin Hesaplanması……………….92 2.4.8. İzoterm Şekillerinin Analizi ve Yorumlanması…………………………………………………92 2.4.9. Projemdeki Deneysel Tasarım………………………………………………………………………..93 2.4.10. Projemde Geliştirilen Yeni Deneysel Protokol………………………………………………..93 2.4.10.1. Kimyasallar………………………………………………………………………………………………93

99
2.4.10.2. Langmuir-Blodgett Tek Katmanın Oluşturulması………………………………………..94 2.4.10.3. Yüzey Basınç-alan (π-A) İzotermler…………………………………………………………….97 2.4.10.4. Diferansiyel Taramalı Kalorimetrik (Differential Scanning Calorimetry) Ölçümleri…………………………………………………….95 2.4.10.5. Fourier Transform Infrared (FTIR) Spektrometresi……………………………………..95 3. Bulgular……………………………………………………………………………………………………………..97 3.1. FTIR Ölçümleri………………………………………………………………………………………………..98 3.2. Diferansiyel Taramalı Kalorimetre (Differential Scanning Calorimetry) Ölçümleri……………………………………………………112 3.3. Langmuir-Blodgett Tek Katman Ölçümleri………………………………………………………..119 4. Tartışma…………………………………………………………………………………………………………..163 4.1. FTIR Ölçümleri………………………………………………………………………………………………163 4.2. DSC Ölçümleri……………………………………………………………………………………………….173 4.3. Langmuir-Blodgett Tek Katman Ölçümleri……………………………………………………….175 4.3.1. Bu Projede Yapılan Langmuir-Blodgett Tek Katman Ölçümlerinin Daha Önce Başka Bir Cihazla Yaptığım Ölçümlerle Kıyaslanması…………………………177 4.4. Taxol®-Fosfolipid Membran Arasında Oluşan Yapı……………………………………………186 4.5. Biyoteknolojik Boyut……………………………………………………………………………………….187 4.6. Hücresel ve Moleküler Biyolojik Boyut……………………………………………………………..192 4.7. Projemde Geliştirilen Tasarımın Diğer Analitik Uygulamalardaki Yeri………………..202 4.7.1. İlâç Ligandlarıın Ayrışımı ve Analizi için İmobilize Lipozom Kromatografisinin Geliştirilmesi………………………………………….202 4.7.2. Projenin Devamı Olarak Yapılması Öngörülen Ortak Çalışmalar……………………..207 4.7.3. Öncü Hipotez………………………………………………………………………………………………209 4.7.4. Fakültemizde Bundan Sonraki Proje Hedeflerim…………………………………………….213 5. Sonuç……………………………………………………………………………………………………………….216 Eklerα…………………………………………………………………………………………………………………217
α Birçok Ek belge niteliğinde olduğundan bunlara sayfa numarası verilmemiştir.

100
PROJE ÖZET BİLGİ FORMU
Proje Kodu: 02/2005-15
Proje Başlığı: PACLITAXEL (TAXOL’un) ANTİ-KANSER ETKİ
MEKANİZMASININ MODEL LANGMUİR-BLODGETT
BİYOMEMBRAN SİSTEMİ İLE MOLEKÜLER DÜZEYDE
İNCELENMESİ
Proje Yürütücüsü ve Yardımcı Araştırmacılar:
Dr. Erhan Süleymanoğlu
Projenin Yürütüldüğü Kuruluş ve Adresi: Gazi Üniversitesi, Eczacılık
Fakültesi, Merkez Laboratuvarı ve Farmasötik Kimya Anabilim Dalı,
Hipodrom, 06330-Ankara
Destekleyen Kuruluş(ların) Adı ve Adresi: Gazi Üniversitesi, Bilimsel
Araştırmalar Projeleri Daire Başkanlığı
Projenin Başlangıç ve Bitiş Tarihleri: Ekim-2005-Ekim-2007
Türkçe Özet: Bu çalışmada, kemoterapide yoğun olarak kullanılan Taxol® (Paclitaxel) ile farklı
zincir yapılarına ve elektrostatik özelliklerine sahip fosfolipidler arasında oluşan moleküler
etkileşmeler Langmuir-Blodgett tek katman, FTIR ve DSC ölçümleriyle incelenmiştir. Sınır
yüzeylerde oluşturulan lipid tek katman ve lipid veziküllerinde (lipozom) yer alan lipid çift
katmanlar model hücre zarları olarak uygulanmıştır. Neticeler, Taxol® konsantrasyonuna, pH,
T°C, tampon sistemine, hidrofobik ve elektrostatik ortama bağlı olarak açıklanmıştır. İlâç ile farklı
fosfolipid molekülleri arasındaki etkileşmelerinin sonucu olarak meydana gelen “self-assembly”
agregatlarının sekonder yapısal değişiklikleri FTIR spektroskopisi ile incelenmiştir. Elde edilen
veriler, karbonil, fosfat, kolin ve CH2 gruplar seviyesinde değerlendirilmiştir. Tek katmanda
meydana gelen ilâç-fosfolipid etkileşmeleri ara yüzeydeki lipidlerin moleküler alanlarına
bağımlılık göstermektedir. Taxol®’un, fosfatidilkolin türü zwitteriyonik lipidler ile termodinamik
olarak sabit yapılar oluşturduğu DSC ölçümleri ile ortaya çıkarılmıştır. Tek katmanda oluşan söz
konusu yapıları Brewster Angle (BAM), atomik kuvvet (AFM), floresan ve Raman mikroskopi
yöntemleriyle de görüntülemeyi ümit ediyoruz. Bu farmasötik nanoformülasyonlarının in vivo

101
ortamda hücre zarı akışkanlığı üzerindeki muhtemel etki mekanizması ile ilgili yeni bir hipotez de
sunulmuştur.
Bu çalışma Gazi Üniversitesi Bilimsel Araştırma Projeleri Daire Başkanlığı tarafından
İngilizce Özet:
Abstract: Introduction: The current study reports our last results of recent project on the role of biomembranes in cancer therapeutics.
Aim: The objective is to emphasize their relevant physicochemical features in depicting molecular mechanisms of action of antineoplastic agents with further implications in drug design.
Methods: Langmuir-Blodgett monolayers were used to simulate lipid surface of cell membranes of both normal and cancer cells trying to approach surface forces governing molecular interactions between various phospholipids and Paclitaxel (Taxol®)-a widely used anticancer drug. Fourier Transform Infrared (FTIR) Spectroscopy was applied for following subsequent changes in lipids after drug recognition trying to relate this to its fluidizing effect. This was further studied by Differential Scanning Calorimetric (DSC) measurements, which showed further thermodynamic characteristics of drug-lipid interaction.
Results and Discussion: Biophysical analysis showed that miscible drug-lipid complexes form at air/water interface, followed by instability of the monolayer due to their interaction. Spectroscopic and thermodynaic measurements showed the phase separation, the extend of liquid-condensed phases, as well as microdomain formation in monolayers. We are also trying to apply Atomic Force Microscopy (AFM), Brewster-Angle Microscopy (BAM) and Raman Microscopy to extend our previously proposed model of the role of phospholipid specificity of Taxol® and its cellular effects. Relevant physicochemical reasoning and models will be presented in more detail.
Acknowledgements: This work was supported by Gazi University Research Fund (Project
No: 02/2005-15).
Anahtar Kelimeler: Paclitaxel (Taxol®), Model ve Biyomembranlar, Tümör Hücre
Zarı Akışkanlığı, Langmuir-Blodgett Tekkatman, İlâç-fosfolipid etkileşmeleri.
Projeden Kaynaklanan Yayınlar: 2 makale ve 4 poster sunumunun
yapılması plânlanmıştır. Tüm bunlar hazırlanma aşamasındadır.
Bilim Dalı: İnterdisipliner
Doçentlik B. Dalı Kodu: Yok G. Ü. Akademik Yükseltilme
ve Atanma Ölçütlerine göre puan durumum: 835

102
Proje ile ilgili genel hususlar.
İlâçlarla organizma arasındaki oluşan karmaşık tepkimeler ve yolaklar
hücresel ve moleküler düzeyde yapılan incelemelerinin önemini artırmıştır. Hücre ile
ilâçlar arasında ilk temas noktası biyolojik lipid zarlardır. Bu bağlamda, çeşitli ilâç-
hücre etkileşmelerindeki lipid zarların rolü küçümsenemez ve bunlarla ilgili elde
edilen veriler son derece önemlidir. Bu verilere dayanarak çeşitli ilâçların etki
mekanizmalarının daha iyi anlaşılması sağlanmaktadır. Günümüzde kullanılan
birçok farmasötik formülasyon amfifilik veya katamfifiliktir ve bu özellikler sayesinde
biyolojik zarlarla tepkimelere girmektedirler.
Çeşitli antibiyotiklererin, anti-enflamatuarların ve anestetiklerin model ve
biyolojik membranlar üzerinde etkileri uzun yıllar önce araştırmacıların dikkatini
çekmiş ve ayrıntılı olarak farklı düzeneklerde incelenmiştir. Bu araştırmalar
sayesinde, örneğin anestetiklerin etki mekanizması ile ilgili, halâ geçerliliğini
yitirmemiş lipid teoriler ortaya atılmıştır1 ve birçok farmakodinamik modeller bu
teorilerine dayanmaktadır. Söz konusu modeller farklı ilâçların etki mekanizmalarına
da uyarlanmaya çalıştırmıştır. Diğer yandan, normal hücre ile kanser hücrenin
akışkanlığında önemli farklılıklar ortaya çıkarılmıştır ve bu buluşlar anti-kanser
ilâçların membran üzerinden çalışan bileşenler olarak algılanması gerektiği
konusunu da gündeme getirmiştir.
Sunduğum bu projede de, kemoterapilerde yoğun olarak kullanılan Taxol
(Paclitaxel) ile kanser hücreyi2 modelleyen çeşitli membran modelleri arasında
oluşan etkileşmeler ele alınmıştır. Projemin iki boyutu vurgulanmıştır. Bunlardan ilki
biyoteknolojik boyut olup, çeşitli lipidlere dayalı kontrollü salınım sistemlerdeki
Taxol-fosfolipid komplekslerinin oluşumu üzerine durmaktadır. Günümüzde
kullanılan birçok çok halkalı (heterosiklik) farmasötik formülasyonların hidrofobik
özelliklerinden dolayı çözünürlülük sorunu ortaya çıkmıştır ve bunların biyolojik
1 Bu öncü teoriler için Bk.: Trudell JR. A unitary theory of anesthesia based on lateral phase separations in nerve membranes. Anesthesiology. 1977 Jan;46(1):5-10. Seelig A, Allegrini PR, Seelig J. Partitioning of local anesthetics into membranes: surface charge effects monitored by the phospholipid head-group.Biochim Biophys Acta. 1988 Apr 7;939(2):267-76. Suezaki Y, Tatara T, Kaminoh Y, Kamaya H, Ueda I. A solid-solution theory of anesthetic interaction with lipid membranes: temperature span of the main phase transition. Biochim Biophys Acta. 1990 Nov 2;1029(1):143-8; Jibu M. Theory of cell membrane organizers and pressure reversal of anesthesia. Med Hypotheses. 2001, Jan;56(1):26-32. 2 Kanser hücre yüzeyini (cancer cell surface) olarak ifade edilmesi çok daha doğru olacaktır.

103
sıvılarda uygulama etkinliğini olumsuz etkilemektedir. Bu yüzden, çözünürlülük
sorununu bazı kontrollü salınım sistemleri ile çözmeye çalışılmaktadır. Ancak, bunun
için ilâç-lipid kompleks oluşumunun ön incelemesi gerekmektedir ve bu önemli konu
projemde de ele alınıp değerlendirilmiştir. Bu tür fizikokimyasal verilere dayanarak
daha sonraki ilâç geliştirme aşamalarında yeni, tedavi potansiyeli ve özellikleri
iyileştirilmiş anti-kanser formülasyonlarının geliştirilmesini mümkün kılmaktadır.
Projemin ikinci boyutu ise bu anti-neoplastik ajanın hücre seviyesindeki etki
mekanizması ile ilgilenmekte ve böylece daha fundamental moleküler biyolojik
vurguları içermektedir. Örneğin, günümüzde geçerli olan depolimerizasyona karşı
hücre mikrotübüllerin stabilizasyonunu sağladığı mekanizması3, Taxol‘un tüm
moleküler düzeyde gösterdiği etkileri açıklamak için yeterli değildir. Örneğin, bir
membran proteini olan ve kemoterapiye direnç mekanizmalarının ortaya çıkmasında
sorunlu olan P-glikoprotein (P-gp) tam olarak nasıl çalıştığını ve etki ettiğini
anlayabilmek için, çeşitli anti-neoplastik ilâçların bu gibi ortamda nasıl davrandıkları
konusunun önemini artırmaktadır. Başka bir deyişle, moleküler düzeyde herhangi bir
anti-kanser terapinin geliştirilmesi için bu ilâçlarla kanser hücre membranları
arasında oluşan etkileşmelerinin ortaya çıkarılmasını gerektirmektedir. Bu bağlamda,
Taxol ile fosfolipidler arasındaki moleküler bağlar, güçler ve etkileşmeler
vurgulanmıştır. Bu olayların daha iyi anlaşılması amacıyla ilk önce bu projemin kesin
raporunda geniş bir giriş bölümüne yer verilmiştir. Söz konusu bu bölüm, projemi
inceleyecek olanlara yukarıda bahsedilen bu 2 ayrı boyutu da daha iyi
algılayabilmelerini sağlayacaktır.
Motivasyon ve Ankara’da bulunan araştırma olanakları.
Sunduğum projede, özellikle nanoteknolojik boyut vurgulanmıştır. Bunun için
de çok sağlam dayanaklarım ve ciddi sebeplerim vardı. Her şeyden önce, arzu ettiğim
cihazların yerine Ankara’da mevcut olan imkânları ve üniversitemizin çok kısıtlı proje
bütçesini göz önünde bulundurarak proje konusunu kararlaştırdım. Bu bağlamda,
Fakültemizde çok yetersiz bir altyapı ortamında ve tek başıma yürütebileceğim bir
projeyi tasarladım4. Sonuç olarak, bu proje sayesinde nanoteknolojinin temellerini
3 Taxol‘un etki mekanizmasına aşağıdaki bölümlerde ayrıntılı olarak yer verilmiştir. 4 Fakültemizde göreve başlar başlamaz daha 04 Şubat 2004 tarihinde yapılan bir Fakülte toplantısında yönetim tarafından benden ve diğer 2 uzman arkadaşımızdan tek başımıza proje çalışması çıkarmamız istenmiştir. Bu toplantıda ayrıca Üniversitemizin herhangi bir uluslararası boyutu olmadığı ve ulusal sıralamada bile yeri

104
oluşturan “self-assembly” olarak meydana getirilen biyomakromolekül komplekslerin
araştırıldığı ve fizikte, kimyada, malzeme biliminde, tıpta ve biyonanoteknolojide
uygulama potansiyeli olan orijinal bir cihaz Fakültemize alınmıştır. Bunun için de
ulusal yükseköğretim stratejileri (Bk.:
http://www.yok.gov.tr/duyuru/strateji_gorus.htm) ve üniversitemizin strateji plan
geliştirme programı (http://www.strateji.gazi.edu.tr/) esas alınmıştır. Projemdeki bu
vurgu uluslararası proje destek programlarına katılmamız bakımından da önemlidir,
çünkü Avrupa Birliği’nin gerek 6cı, gerekse 7ci çerçeve programlarında öncelikli konu
başlıklarına önem verileceği ayrıca vurgulanmaktadır (Bk.: www.fp7.org.tr). Söz
konusu başlıklar arasında genom bilimler ve nanoteknolojiler başı çekmektedir ( Bk.:
cordis.europa.eu/fp7/home_en.html;http://www.innovation.public.lu/servlet/front).
Ancak bu proje destek programlarına Türkiye’den çok az sayıda proje gönderilmiştir.
Bu arada, ülkemizde de ulusal nanoteknoloji stratejileri geliştirilmiştir5. Bu tür
nanoteknoloji çalışmaları da en ileri düzeyde Ankara’da yapılmaktadır. Bunun için de
gereken altyapıyı ve AR-GE olanakları kurulmuştur. Örneğin, O. D. T. Ü.’inde birkaç
seneden bu yana Nanoteknoloji Merkezi faaliyet göstermektedir. Ayrıca, aynı
üniversite Avrupa Birliği kaynaklarından 1.5 milyon EU’luk subvansiyon sayesinde
Merkez Laboratuvaru kurmuştur (Bk.: www.centrallab.metu.edu.tr). Bu Merkez
Laboratuvarın Ocak-2006 tarihinde kurulmuş olup, asıl amacı üniversitelere ve
sanayi kuruluşlarına bilimsel destek ve yürütülmekte olan projelerde öçlüm
olanakları sağlamaktır. Avrupa Birliğinden akreditasyon alan tek kuruluş O. D. T. Ü.
Merkez Laboratuvarıdır. Bu projemin ön ölçüm kısmını oluşturan FTIR ve DSC
ölçümlerini bu laboratuvarda yaptım.
Bunun dışında, Bilkent Üniversitesinde Devlet Plânlama Teşkilâtından alınan
25 Milyon Dolarlık proje ile büyük bir alana yayılmış Ulusal Nanoteknoloji Merkezi
(Bk.: www.nano.bilkent.edu.tr) kurulmuştur ve bunun da 2 yıla kadar tamamlanması
beklenilmektedir. Hacettepe Üniversitesi Beytepe kampüsünde de birçok
nanoteknolojik projenin yürütüldüğü BİYOMEDTEK Merkezi kurulmuştur
(www.biyomedtek.com). Yakın bir tarihte de, Teknopark açılacaktır
(www.teknopark.hacettepe.edu.tr). Buna paralel olarak, Ankara Üniversitesi 5 sene
önce Devlet Plânlama Teşkilâtından almış olduğu 8 Milyon Dolarlık proje desteği ile
istenilen düzeyde olmadığı (bunun için bk.: Ek-1 ve Ek-2) ve Fakültemizde cihazların son derece demode ve eski olduğu gerekçe gösterilerek tarafımdan orijinal proje çıkarmam rica edilmiştir. 5 Bk.: Ek-3.

105
Biyoteknoloji Enstitüsü kurmuştur. Daha sonra, bu enstitüye bağlı Merkez
Laboratuarı da kurulmuştur (Bk.: http://www.biotek.ankara.edu.tr/).
Görüldüğü gibi, Ankara’da bulunan diğer üniversitelerde bu tür büyük
yatırımlar yapılırken üniversitemizde bu ölçekte henüz daha herhangi bir girişim
olmamıştır. Ancak, son yıllarda üniversitemizde de büyük gelişmeler kaydedilmiştir.
Şöyle ki, Yarıiletken Araştırma Merkezine bağlı bir Nano-Club faaliyete başlamıştır.
Ancak, bu birimin biyonanoteknolojiye hitap etmemektedir, daha ziyade mühendislik
bilimleriyle ilgilenmektedir. Ancak, bunun dışında, yakında çok önemli olan Ulusal
Nanotıp Konsorsiyumu üniversitemizde kurulmuştur (Yarıntılı bilgi için Bk: Ek-4) ki,
bu çok isabetli bir gelişmedir. Üniversitemizin6 Teknoparkı da tamamlandığında
uluslararası düzeyde nanoteknolojik araştırmaların yapılabileceği ümit edilmektedir.
Ancak, bu tür merkezleri diğer araştırma merkezlerle yarışabilir düzeyde
tutabilmek için büyük ölçekli, sürekli ve uzun vadeli projeler çıkarmak
gerekmektedir. Projemi de bu amaçla sundum. Şimdiye kadar Fakültemizde herhangi
bir nanoteknoloji başlıklı proje çıkarılmadığı gibi laboratuvar donanımı da son derece
yetersizdir. Projemi de bu iki hususu göz önünde bulundurarak yazdım. Bu
doğrultuda, cihaz alımı ile sonuçlanabilecek bir proje teklifi sunulmuştur. Tüm
zorluklara rağmen, projemi tamamlamış bulunmaktayım. Ancak, bu aşamadan sonra
bunun devamı da gelmelidir. Bu yüzden daha şimdiden ulusal ve uluslararası proje
destekleme kuruşlarına çok daha kapsamlı bir projemi daha sunmak üzereyim (Bk.:
Ek-16).
6 Üniversitemizin büyük çabaları ve girişimleri sayesinde ayrıca Ulusal Kızılötesi ve Sinhrotron Araştırma Merkezi de kurulma aşamasındadır.

106
1. Giriş.
1.1. Kanser7.
Kanser terimi ile, bu projemin kesin raporunun kapsamına ve amacına uygun
olarak hücresel büyümedeki düzenleme mekanizmalarında meydana gelen önemli
hasarlar gösteren birçok hastalık kastedilmektedir. Bu konu son derece hızlı
geliştiğinden ve çok geniş olduğundan, kanser ile ilgili daha ayrıntılı bilgi için
aşağıdaki dipnotta vermiş olduğum geniş ve güncel kaynakçaya başvurulmalıdır.
Tarihsel olarak, kanser araştırmaları bir hayli deskriptiv olmuştur. Başka bir
deyişle, birçok araştırmacı normal hücrelerle onkogenik hücreler arasında
gözledikleri farklılıkları kataloglaştırmışlardır. Uzun yıllar boyunca hastalığın etkileri
nedenlerinden çok daha iyi incelenmiştir. Son yıllarda, birkaç kanser türü üzerinde
moleküler düzeyde yapılan araştırmalar sayesinde metastaz oluşum mekanizmaları,
tümörlerin bağışıklık sisteminden tanınmaları ve yok edilmeleri gibi çok önemli olan
konular bağlamında, tümör hücrelerinin bazı özelliklerinin anlaşılmasını mümkün
kılmıştır.
Tümör hücreleri intraselüler (hücre içi) haberleşme (intracellular
communication) mekanizmalarında birtakım hasarları içerdiklerinden, bunların
patolojik davranışlarındaki plazma membranının8 da muhtemel rol oynayabileceği
tahmin edilmektedir (Petty, H. R., 1993). Gerçekten de, projemde anti-neoplastik
ilâçların biyoteknolojik uygulamaları dışında, tümör hücre membran akışkanlığı
7 Bu bölümle ilgili daha ayrıntılı bilgiler için özellikle bk.:
1. Advances in Molecular Oncology (Advances in Experimental Medicine and Biology) (Advances in Experimental Medicine and Biology), by Fabrizio d'Adda di Fagagna (Editor), Susanna Chiocca (Editor), Fraser McBlane (Editor), Ugo Cavallaro (Editor); Springer; 1 edition (July 12, 2007).
2. Targeted Molecular Imaging in Oncology, by E. Edmund Kim and David J. Yang (Editors), Springer; 1 edition (January 15, 2001).
3. Principles of Molecular Oncology, by Miguel H. Bronchud, MaryAnn Foote, Giuseppe Giaccone, Olufunmilayo I. Olopade, Paul Workman (Editors), Humana Press; 3rd ed. edition (November 30, 2007).
4. The Molecular Biology of Cancer, by Michael Khan, Stella Pelengaris (Editors), Wiley-Blackwell; 1 edition (February 28, 2006).
5. Molecular Biology of Cancer: Mechanisms, Targets, and Therapeutics,by Lauren Pecorino, Oxford University Press, USA; 1 edition (May 21, 2005).
6. F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor and Francis, New York.
7. The Biology of Cancer, by Robert A. Weinberg, Garland Science; 1 edition (June 9, 2006). 8 Projemin de hücresel boyutu söz konusu model normal ve tümör plazma membran (zar) akışkanlığı üzerinde odaklanmaktadır. Bunun için özgün bir de öncü hipotez geliştirilmiştir. Daha ayrıntılı bilgiler için bu proje kesin raporumun yorumlar bölümünde yer almaktadır.

107
üzerindeki muhtemel etkilerini vurgulamaktadır. Onkogenler ve kansere yol açan
genlerin ürünleri ligandlar, reseptörler ve sitozolik sinyalleşme aygıtlar gibi normal
büyümeyi sağlayan transmembran sinyal iletim elemanlarının farklı şekillerinden
ibarettir. Bu gen ürünleri, sinyal iletiminin şu ana kadar geliştirilmiş en iyi, ancak
henüz daha yetersiz olan modelini teşkil etmektedir. Bazı tümör hücrelerin membran
taşıyıcıları bunları kemoterapötik ilâçlara karşı korumaktadır. Adherence-promoting
membran proteinlerinin yetersiz ekspresyonu metastaza yol açmaktadır (Petty, H. R.,
1993). Tümör antijenlerinin intraselüler aktarımları ve bunu takiben plazma
membranlardaki ekspresyonları immün sistem trafından indüklenen destrüksyonu
ortaya çıkarmaktadır (Petty, H. R., 1993; Cereijido M, et al., 2007; Chidgey M,
Dawson C., 2007; Ma DW., 2007; Song RX., 2007; Baritaki S. et al., 2007)9. Bu konu,
projemin yorum bölümünde ölçümlerden elde edilen neticeler doğrultusunda tekrar
ele alınmıştır.
1.2. Kanserin Tedavi Şemaları.
1.2.1. Sistemik Tedavinin Temel Prensipleri
Büyüme mekanizmalarının da kontrolden çıkmasını takiben muhtemel olarak
tek hücreden veya birkaç hücreden ortaya çıkan malign tümör genel olarak heterojen
tümör popülasyonundan ibarettir. Bu hücreler daha sonra komşu dokulara göç
ederek bunların da malign tarnsformasyonuna neden olup, kendi vaskülatürünü
geliştirip lenfatik ve hematolojik dağılım göstermek suretiyle metastazlara neden
olmaktadır. Metastaz olan hastalarda kanserin bir sistemik hastalık olarak ele
alınması gerektiği ve cerrahi veya radyoterapi gibi konvansyonel yaklaşımların
yetersiz olacağı açıktır. Bu bağlamda, günümüzde kanser hastaların bakımı cerrahi,
radyoterapi ve sistemik tedavilerini de içeren multidisipliner bir yaklaşımı gerektirir
(Sliifer S ve Stoter G. , 2005).
9 Bk. özellikle: Chapter 9: Membranes in Cancer, In: Howard R. Petty, Molecular Biology of Membranes-Structure and Function, Plenum Pres, New York and London, 1993, pp. 353-377; Cereijido M, Contreras RG, Flores-Benítez D, Flores-Maldonado C, Larre I, Ruiz A, Shoshani L. New diseases derived or associated with the tight junction. Arch Med Res. 2007 Jul;38(5):465-78; Chidgey M, Dawson C. Desmosomes: a role in cancer? Br J Cancer. 2007 Jun 18;96(12):1783-7; Ma DW. Lipid mediators in membrane rafts are important determinants of human health and disease. Appl Physiol Nutr Metab. 2007, Jun;32(3):341-50; Baritaki S, Apostolakis S, Kanellou P, Dimanche-Boitrel MT, Spandidos DA, Bonavida B. Reversal of tumor resistance to apoptotic stimuli by alteration of membrane fluidity: therapeutic implications. Adv Cancer Res. 2007;98:149-90 ve Song RX. Membrane-initiated steroid signaling action of estrogen and breast cancer. Semin Reprod Med. 2007 May;25(3):187-97.

108
Kanser hastalarının 60%’dan fazlasında metastazların görüldüğünden, çoğu
kez sistemik olarak etki eden ajanlarla tedavilere başvurulmaktadır. Bu şekilde
sistemik etki mekanizmasına sahip anti-tümör ajanları 3 ayrı gruba dahil
edilmektedir: sitotoksik ilâçlar, hormonal ajanlar, ve biological responce modifiers
adlı bileşenler. Bu gruplarda yer alan çeşitli ajanlar yıllarca uygulanmışlardır (Sliifer
S ve Stoter G., 2005)10. Daha spesifik olarak temel kanser hücre biyolojisi,
farmakoloji, endokrinoloji konulu araştırmalar, ve yeni bileşenlerin ve yeni tedavi
stratejilerin geliştirilmesinin temelini oluşturmaktadır. Bunlara ilâve olarak, başarılı
kanser tedavilerin neticesinde uzun süreli olarak sağlığına kavuşan kişilerin
sayısındaki artış göz önünde bulundurulduğunda, daha sonraları görülen bazı
toksisitelerinin de önemi artmıştır.
1.2.2. Kemoterapinin Prensipleri.
Kemoterapötik ilâçların önemli bir özelliği hızlı bölünen hücrelere hasar
vererek hücre ölümüne yol açmalarıdır. Hücre hasarı farklı etki mekanizmalarına
sahip birkaç tür sitotoksik ilâçlar tarafından meydana getirilir (Bk.: Tablo-1 ve Şekil
1.1)11.
10 Tarihçeleri için bk.: Skipper H. E. Historic milestones in cancer biology: a few that are important to cancer treatment. Semin. Oncol., 1979, 6: 506-514. 11 Projem güncel olan ve çok yoğun olarak kullanılan Taxol® ile ilgilidir. Ancak kemoterapilerde kullanılan birçok antikanser ilâcı daha mevcuttur. Tüm bunlarla ilgili okuyuculara daha ayrıntılı bilgi vermek amacıyla bu kesin raporumun sonunda Ek-14 verilmiştir.

109
Tablo-1: Bazı sitotoksik ilâçlar.
(Kaynak: Sliifer S ve Stoter G. Chapter 2: Principles of Systemic Therapy. In: F.
Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor and
Francis, New York, pp. 35-49).

110
Şekil 1.1. Bazı popüler anti-kanser ilâçların etki mekanizmaları (Bk.:
www.chemotherapy.com; Ayrıca bk.: Danimarka Kraliyet Kanser Araştırmaları
Enstitüsü- www.cancer.dk)12.
12 Kemoterapinin hücresel boyutu ile ilgili aşağıdaki kaynaklara başvurulmalıdır:
1. Gossage L, Madhusudan S., Cancer pharmacogenomics: role of DNA repair genetic polymorphisms in individualizing cancer therapy. Mol Diagn Ther. 2007;11(6):361-80.
2. Mandal S, Varma K, Jain S., Cutaneous manifestations in non-Hodgkin's lymphoma. Acta Cytol. 2007 Nov-Dec;51(6):853-9.
3. Hamm C, Verma S, Petrella T, Bak K, Charette M; On behalf of the Melanoma Disease Site Group of Cancer Care Ontario’s Program in Evidence-based Care. Biochemotherapy for the treatment of metastatic malignant melanoma: A systematic review. Cancer Treat Rev. 2007.
4. Cordes N, Park CC., β1 integrin as a molecular therapeutic target. Int J Radiat Biol. 2007;83(11):753-760.
5. Nicolson GL, Conklin KA. Reversing mitochondrial dysfunction, fatigue and the adverse effects of chemotherapy of metastatic disease by molecular replacement therapy. Clin Exp Metastasis. 2007 Dec 5.
6. Korbelik J, Cardeno M, Matisic JP, Carraro AC, MacAulay C., Cytology microarrays. Cell Oncol. 2007;29(5):435-42.
7. Cellular Oncology, Mol Diagn Ther. ve Cancer dergileri son derece faydalı bilgiler içermektedir.
8. Vattemi E, Claudio PP. Tumor suppressor genes as cancer therapeutics. Drug News Perspect. 2007 Oct;20(8):511-20.
9. Xi Y, Edwards JR, Ju J. Investigation of miRNA Biology by Bioinformatic Tools and Impact of miRNAs in Colorectal Cancer-Regulatory Relationship of c-Myc and p53 with miRNAs. Cancer Inform. 2007;3:245-253.
10. Hoskin DW, Ramamoorthy A., Studies on anticancer activities of antimicrobial peptides. Biochim Biophys Acta. 2007 Nov 22.
11. Cuzick J, Cafferty FH, Edwards R, Møller H, Duffy SW. Surrogate endpoints for cancer screening trials: general principles and an illustration using the UK Flexible Sigmoidoscopy Screening Trial. J Med Screen. 2007;14(4):178-85.
12. Singh VK, Jia Z. Targeting synuclein-gamma to counteract drug resistance in cancer. Expert Opin Ther Targets. 2008 Jan;12(1):59-68.

111
Söz konusu sitotoksil ajanların bu hücre hasarını hangi boyutta ve şiddette
meydana getirdikleri ise genel olarak hücresel ve farmakolojik özelliklere bağlıdır.
Sitotoksik tedaviye karşı bazı tümörlerin duyarlılığını etkileyen faktörler arasında
şunlar sıralanabilir:
- Hücre döngüsü zamanı ve “doubling time”.
- Büyüme fraksiyonu
- Sitotoksik ilâçlara karşı duyarlılık
- Tümör heterojenliği.
Bir ilâcın anti-tümör etkinliğini tayin eden farmakolojik özellikler ise
şunlardır:
- İlâç konsantrasyonu
- İlâca maruziyet zamanı.
Başlangıçta, kemoterapinin tümör hücrelerini nasıl etkilediği konusunda
lösemi L1210 deneysel sıçan (murine) modeli kullanılarak önemli veriler elde
edilmiştir (Skipper H. E., 1979). Bu modele göre, bir tümör eksponensiyel olarak sabit
bir “doubling time” ile ilerler ve yarı logaritmik kağıt üzerine büyüme hızı çizildiğinde
Şekil 1.2-(A)’daki eğriyi oluşturur.
13. Johansson Swartling F. Identifying candidate genes involved in brain tumor formation. Ups J Med Sci.
2008;113(1):32-71. 14. Cheng HY, Obrietan K. Revealing a Role of MicroRNAs in the Regulation of the Biological Clock.
Cell Cycle. 2007 Oct 1;6(24). 15. Lo AS, Zhu Q, Marasco WA. Intracellular antibodies (intrabodies) and their therapeutic potential.
Handb Exp Pharmacol. 2008;(181):343-73. 16. Namiki M, Ueno S, Kitagawa Y, Konaka H, Mizokami A, Koh E, Fukagai T. Hormonal therapy. Int J
Clin Oncol. 2007 Dec;12(6):427-32. 17. Keeble JA, Gilmore AP. Apoptosis commitment--translating survival signals into decisions on
mitochondria. Cell Res. 2007 Dec;17(12):976-84. 18. Tejpar S. The use of molecular markers in the diagnosis and treatment of colorectal cancer. Best Pract
Res Clin Gastroenterol. 2007;21(6):1071-87. 19. McGrogan BT, Gilmartin B, Carney DN, McCann A. Taxanes, microtubules and chemoresistant breast
cancer. Biochim Biophys Acta. 2007 Nov 12. 20. Jacob K, Sollier C, Jabado N. Circulating tumor cells: detection, molecular profiling and future
prospects. Expert Rev Proteomics. 2007 Dec;4(6):741-56. 21. Kischel P, Waltregny D, Castronovo V. Identification of accessible human cancer biomarkers using ex
vivo chemical proteomic strategies. Expert Rev Proteomics. 2007 Dec;4(6):727-39.

112
Şekil 1.2. (A): Yarı logaritmik kağıt üzerine çizilmiş eksponensiyel tümör
büyüme kinetiği. (B): Bu şekilde büyüyen bir tümörün antineoplastik ilâçla
muamelesi. Ok işaretleri ilâcın uygulanma sıklığını göstermektedir (Bk.: Sliifer
S ve Stoter G. Chapter 2: Principles of Systemic Therapy. In: F. Cavalli H. H.
Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor and Francis,
New York, pp. 35-49).
Şekil 1.2.’de, tümörlerin beli bir yüzdeliğinin öldürüldüğünü göstermektedir ve
bu olay “fractional cell killing” olarak adlandırılır. Bu şekilde, ilâç konsantrasyonunun
yükseltilmesi öldürülen tümörlerin yüzdeliğini de etkilemektedir. Bu modele göre,
öldürülen hücrelerin oranı tümör hücrelerin toplam sayısına bağımlılık
göstermektedir. Bu modele göre, ilâç uygulaması yeterli miktarda ve sıklıkta yapıldığı
takdirde, bir tümörü tamamen yok etmek mümkündür.
Ancak, daha sonra, bu model, hastalar için pek uygun olmadığı ortaya
çıkmıştır (Sliifer S ve Stoter G. Chapter 2: Principles of Systemic Therapy. In: F.
Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor and
Francis, New York, pp. 35-49). Sitostatik ilâçların genelde hızla bölünen hücreler
fraksiyonuna karşı çok etkili olduklarından, tümör hücrelerinin önemli bir bölümü
etkilenmeyecektir. Prensip olarak, tümör hücre büyüme faktörlerinin uygulanması

113
tümör hücre bölünmesini de artıracaktır ki, bu olaya “recruitment” veya “priming of
tumor cells” adı verilmektedir.
L1210 modeli ile katı tümörler arasındaki ikinci önemli fark ise büyüme
fraksyonu ile “doubling time” arasındaki ilişki sabit olmadığı ve tümör büyüklüğüne
göre farklılaştığı gözleminden ileri gelmektedir.
İlk başta, bir tümör L1210 modelinde olduğu gibi eksponensiyel büyüme hızı
gösterir. Ancak, daha sonra, tümörün büyüklüğü artığında henüz daha
açıklanamayan nedenlerden dolayı büyüme hızı durur. Büyüme hızı ile zaman
arasındaki bu tür ilişki, Gompertz tipi olarak da adlandırırılan sigmoidal eğri ile ifade
edilmektedir (Şekil 1.3-A). Büyüme hızı ile tümör büyülüğü arasındaki bu ilişki
kemoterapi şemalarının geliştirilmesinde önemli bir motivasyon sebebini
oluşturmuştur.
Şekil 1.3. (A): Sigmoidal tümör büyüme hızı. (B) Sigmoidal büyüme hızına
sahip bir tümörün belli bir ilâç dozuyla uyarlanması. Kemoterapiden kaynaklanan
tümör büyüklüğündeki azalma, büyüme fraksyonu artırmakta ve geriye kalan
tümörlerin büyüme hızındaki artış meydana gelmektedir (Bk.: Sliifer S ve Stoter G.
Chapter 2: Principles of Systemic Therapy. In: F. Cavalli H. H. Hauser ve S. B. Kaye
(Ed.): Textbook of Medical Oncology, Taylor and Francis, New York, pp. 35-49).
Kemoterapinin etkinliğini etkileyen başka bir faktör de tümör hücrelerinin
konsantrasyonudur (Bk.: Sliifer S ve Stoter G. Chapter 2: Principles of Systemic

114
Therapy. In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical
Oncology, Taylor and Francis, New York, pp. 35-49). İlâçlara karşı direncin meydana
gelmesi birkaç mekanizmayla açıklanmaktadır. Tümör hücrelerinde P-glikoprotein ve
multidrug-related protein (MRP) gibi ilâç pompalarının ekspresyonu kemoterapiye
karşı direncin gelişmesindeki başlıca nedenidir. Diğer direnç mekanizmaları tümör
hücrelerince alınan ilâç miktarındaki düşüş, ilâç detoksifikasyonu veya tümör
hücrelerinin kemoterapiden kaynaklanan DNA hasarını onarım kapasitesini
artırmaları sayesinde meydana gelmektedir (Bk.: Tablo-2).
Tablo-2. Hücresel İlâç Direncinin ve etkilenen Sitotoksik İlâçların Etki
Mekanizmaları (Bk.: Sliifer S ve Stoter G. Chapter 2: Principles of Systemic Therapy.
In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor
and Francis, New York, pp. 35-49 ).
Ancak, şimdiye kadar sıralanan mekanizmalardan, en önemlisi apoptoza13 yol
açan hücresel yolaklardaki patolojilerdir. Son yıllarda kemoterapiden kaynaklanan
programlanmış hücre ölümünü14 indükleyen mekanizmalar hakkında önemli bilgiler
13 Programlanmış hücre ölümü. 14 Programlanmış hücre ölümü kavramı 1989li yıllarda Almanya’da Heidelberg Üniversitesinde Catastrophy
Theory’nin bir matematiksel modeline dayanmaktadır. İlk önce normal hücre döngüsünde scenescence
doğrultusunda ilerleyen hücre bölünme olaylarını açıklamak için ele alınmıştır. Daha sonra İskoçya’da Glasgow Üniversitesinde Dr. Renato Baserga’nın ve diğer birçok araştırmacının geliştirdiği kanser hücre kültürü düzenekleri sayesinde bu hücresel olay kanser gibi patolojik olaykara da aktarılmıştır. Bu konuda yığınlarca yayın bulunmaktadır. Ancak bu kesin raporun amacı için şunlara başvurulması tavsiye edilebilir:

115
elde edilmiştir. Kemoterapiden kaynaklanan ilk olay çoğu kez tümör hücresinin DNA
hasarıdır (Gossage, L. ve Madhusudan, S., 2007), ancak mikrotübüller gibi diğer
hücresel makromoleküller de hasarın hedefi olabilmektedir (McGrogan et al., 2007).
Tümör hücresi tarafından bu hasar tespit edildiğinde, ancak hasarın
oranılmadığında, kaspazlar, Bcl-2 grubu proteinleri veya p53 onkogenlerin önemli rol
oynadığı birkaç yolak aktive edilir (Vattemi, E. ve Claudio, PP., 2007 ). Bu olaylarda
mitokondrinin de rol oynadığı ortaya çıkarılmıştır (Nicolson, GL ve Conklin, KA.,
2007; Xi, Y. Ve Edwards, JR, 2007; Keeble, JA ve Gilmor, AP, 2007). DNA’yı
parçalayan endonükleazların aktivasyonundan sonra (Gossage, L. ve Madhusudan,
S., 2007), hücrede kromatin kondenzasyonu ve hücresel fragmentasyon gibi olaylar
meydana gelmektedir°.
Bu projemin de temelini oluşturan kemoterapi ile bağlantılı olarak gelişen
apoptoza özgü hücresel membran bleb15 oluşumları son yıllarda araştırmacılarının
1. Eberle J, Kurbanov BM, Hossini AM, Trefzer U, Fecker LF. Overcoming apoptosis deficiency of melanoma-Hope for new therapeutic approaches. Drug Resist Updat. 2007 Dec 1. 2. Kooistra K, Zhang YH, Noteborn MH. Viral elements sense tumorigenic processes: approaching selective cancer therapy. Mini Rev Med Chem. 2007 Nov;7(11):1155-65. Bu derleme özellikle yeni anti-kanser tedavilerinin geliştirilmesi ile ilgilidir. 3. Ohnishi T. The role of the p53 molecule in cancer therapies with radiation and/or hyperthermia. J Cancer Res
Ther. 2005 Jul-Sep;1(3):147-50. 4. McConkey DJ. Therapy-induced apoptosis in primary tumors. Adv Exp Med Biol. 2007;608:31-51. 5. Seynhaeve AL, Eggermont AM, ten Hagen TL. TNF and manipulation of the tumor cell-stromal interface: "ways to make chemotherapy effective". Front Biosci. 2008 Jan 1;13:3034-45. 6. Klampfer L. The role of signal transducers and activators of transcription in colon cancer. Front Biosci. 2008 Jan 1;13:2888-99. 7. Bertazza L, Mocellin S. Tumor necrosis factor (TNF) biology and cell death. Front Biosci. 2008 Jan. 1;13:2736-43. 8. Havelka AM, Berndtsson M, Olofsson MH, Shoshan MC, Linder S. Mechanisms of action of DNA-damaging anticancer drugs in treatment of carcinomas: is acute apoptosis an "off-target" effect? Mini Rev Med Chem. 2007 Oct;7(10):1035-9. Bu derleme özellikle yeni anti-kanser ilâç formülasyonlarının geliştirilmesi ile ilgilidir. 9. Van Brocklyn JR. Sphingolipid signaling pathways as potential therapeutic targets in gliomas. Mini Rev Med
Chem. 2007 Oct;7(10):984-90. Bu derleme özellikle projeminin de temelini oluşturduğu kanserde fosfolipidlerin rolüüzerine odaklanmaktadır. Projemde yoğun olarak ben de glikosfingolipidleri uyguladım. Bunun için Bk.: bu proje metninin Neticeler bölümünü. 10. Moran E, Nencioni A. The role of proteasome in malignant diseases. J BUON. 2007 Sep;12 Suppl 1:S95-9. ° Bu konu ile ilgili bk.: Sjakste N, Sjakste T. Possible involvement of DNA strand breaks in regulation of cell differentiation. Eur J Histochem. 2007 Apr-Jun;51(2):81-94.; Evenson DP, Kasperson K, Wixon RL. Analysis of sperm DNA fragmentation using flow cytometry and other techniques. Soc Reprod Fertil Suppl. 2007;65:93-113; Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495-516; Yoshida A, Pommier Y, Ueda T. Endonuclease activation and chromosomal DNA fragmentation during apoptosis in leukemia cells. Int J Hematol. 2006 Jul;84(1):31-7; Hewitson TD, Bisucci T, Darby IA. Histochemical localization of apoptosis with in situ labeling of fragmented DNA. Methods Mol Biol. 2006;326:227-34. 15 Apoptoza bağlı olarak gelişen membran bleb’lerle ilgili bk.:
1. Larsen AK, Lametsch R, Elce JS, Larsen JK, Thomsen B, Larsen MR, Lawson MA, Greer PA, Ertbjerg P. Genetic disruption of calpain correlates with loss of membrane blebbing and differential expression of RhoGDI-1, cofilin and tropomyosin. Biochem J. 2007 Dec 12.

116
dikkatini çekmektedir. Önümüzdeki yıllarda da apoptotik membran bleb oluşumları
moleküler onkolojik araştırmalarının önemli bir bölümünü teşkil edeceği açıktır.
Tümörlerin kemosensitivesini (kemoterapiye karşı duyarlılığını) etkileyen
diğer faktör ise genomik instabilitedirϒ. Muhtemelen bir tümörün tek bir hücreden
ortaya çıktığı bilinse de, bir tümörün genomik instabiliteden kaynaklanan
sebeplerden dolayı, birkaç hücre döngüsünden sonra, çok heterojen bir tümör hücre
popülasyonunu oluşturduğu ortaya çıkmıştır. Bu gibi çeşitli hücrelerden oluşan
popülasyonuna kemoterapi uygulandığında, duyarlı olan tümör hücreleri apoptoza
zorlanacaklardır, daha az duyarlı hücreler ise büyüme hızını devam ettireceklerdir.
Kemoterapi birkaç kez tekrar edildiğinde, tedavi etkinliği sıfırlanacak, çünkü tümör
popülasyonu tamamen ilâca-dirençli hücrelerden oluşacaktır. Bu gerçek de çok iyi
çalışan matematiksel modellerle kanıtlanmıştır (Bk.: Sliifer S ve Stoter G. Chapter 2:
Principles of Systemic Therapy. In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.):
Textbook of Medical Oncology, Taylor and Francis, New York, pp. 35-49).
2. Tournaviti S, Hannemann S, Terjung S, Kitzing TM, Stegmayer C, Ritzerfeld J, Walther P, Grosse R,
Nickel W, Fackler OT. SH4-domain-induced plasma membrane dynamization promotes bleb-associated cell motility. J Cell Sci. 2007 Nov 1;120(Pt 21):3820-9.
3. Özellikle bk.: Charras GT, Coughlin M, Mitchison TJ, Mahadevan L. Life and Times of a Cellular Bleb. Biophys J. 2007 Oct 5.
4. Schultz H, Hume J, Zhang de S, Gioannini TL, Weiss JP. A novel role for the bactericidal/permeability increasing protein in interactions of gram-negative bacterial outer membrane blebs with dendritic cells. J
Immunol. 2007 Aug 15;179(4):2477-84. 5. Sheetz MP, Sable JE, Döbereiner HG. Continuous membrane-cytoskeleton adhesion requires
continuous accommodation to lipid and cytoskeleton dynamics. Annu Rev Biophys Biomol Struct. 2006;35:417-34.
6. Nusbaum P, Lainé C, Seveau S, Lesavre P, Halbwachs-Mecarelli L. Early membrane events in polymorphonuclear cell (PMN) apoptosis: membrane blebbing and vesicle release, CD43 and CD16 down-regulation and phosphatidylserine externalization. Biochem Soc Trans. 2004 Jun;32(Pt3):477-9.
7. Martínez MC, Kunzelmann C, Freyssinet JM. Plasma membrane remodelling and cell stimulation. Med
Sci (Paris). 2004 Feb;20(2):189-95. 8. Hamill OP, McBride DW Jr. Induced membrane hypo/hyper-mechanosensitivity: a limitation of patch-
clamp recording. Annu Rev Physiol. 1997;59:621-31. 9. Uchiyama Y. Apoptosis: The history and trends of its studies. Arch Histol Cytol. 1995 Jun;58(2):127-
37. 10. Verkleij AJ, Post JA. Physico-chemical properties and organization of lipids in membranes: their
possible role in myocardial injury. Basic Res Cardiol. 1987;82 Suppl 1:85-91. 11.
ϒ Bu konu ile ilgili olarak daha ayrıntılı bilgi için bk.: Calcagnile O, Gisselsson D. Telomere dysfunction and telomerase activation in cancer--a pathological paradox? Cytogenet Genome Res. 2007;118(2-4):270-6; Wang Y. Chromosome instability in yeast and its implications to the study of human cancer. Front Biosci. 2008 Jan 1;13:2091-102; Cheung AL, Deng W. Telomere dysfunction, genome instability and cancer. Front Biosci. 2008 Jan 1;13:2075-90; Gasparini P, Sozzi G, Pierotti MA. The role of chromosomal alterations in human cancer development. J Cell Biochem. 2007 Oct 1;102(2):320-31; Oberdoerffer P, Sinclair DA. The role of nuclear architecture in genomic instability and ageing. Nat Rev Mol Cell Biol. 2007 Sep;8(9):692-702. Venkitaraman AR. Chromosomal instability in cancer: causality and interdependence. Cell Cycle. 2007 Aug;6(19):2341-3. 2007 Jul 18.

117
Kemosensitivite, anti-tümör aktivitesinin prognostik gösterge olduğundan, bu
aktiviteyi önceden belirlemek amacıyla birçok yönteminin geliştirilmesi günümüzde
büyük bir önem taşımaktadır. Bu konuda sitopatolojik, moleküler genetik ve
moleküler onkolojik, mikroskopik vs. moleküler tıp konularına giren yığınlarca
yöntem mevcuttur16. Tümörlerin kemosensitivitesini etkileyen diğer önemli faktörler
arasında farmakolojik özellikler başı çekmektedir. Bunlar arasında, göğüs ve prostat
16 Ancak, projem bunlarla ilgilenmemektedir. Sunduğum proje, biyoteknolojik boyutu
dışında, daha fundamental hücresel etki mekanizmalarının modellemesi üzerine odaklanmaktadır.
Böyle bir yaklaşımın çok daha rasyonel olduğu açıktır. Bunun da sebepleri şunlardır. Her şeyden
önce yukarıda adı geçen moleküler yöntemleri Fakültemizde uygulayabilmek için uzun vadeli proje
desteğine, özellikle buradaki araştırmacı statümün özerkliğine ve en önemlisi uluslararası
ortaklarımın katılımlarına ve katkılarına ihtiyaç vardır. Ülkemizde her ne kadar da moleküler
genetik yöntemlerinin gerçekleştirilmesi için gereken birtakım cihazlar alınmışsa da, bunların çoğu
çalıştırılamamaktadır. TÜBİTAK çalıştaylarında bu gerçekler defalarca dile getirilmiştir.
Fakültemizdeki durum da pek farklı değildir. Fakültemizde maddî olanaksızlardan dolayı bu olmasa
da, diğer üniversitelerde bu yüzden yurt dışından çok büyük paralar ödenerek pratik seminerlerde
bu yöntemleri göstermek amacıyla firma temsilcileri getirilmektedir. Bu konuda eğitilmiş ve bu
konularda bağımsız olarak proje yürütebilecek akademisyenlerin sayısı da bir hayli yetersizdir.
Avrupa Birliği destekli FP7 çerçeveye veya TÜBİTAK proje destekleme birimlerine gönderilen
moleküler genetik konulu proje sayısına bakacak olursak bu gerçek de ortaya çıkacaktır. En önemli
engel ise, bu yöntemlerin dayanağı olan tümör hücre kültürü düzenekleri ise Fakültemizde mevcut
olmamasıdır. Mevcut olsaydı bile, son birkaç sene yaşanan bitmek tükenmek bilmeyen elektrik
kesintileri ve özel hücre kültürünün gerektirdiği basınçlı kapılarla donatılmış aseptik ortamın
bulunduğu laboratuvarlarının olmaması yüzünden bunları çalıştırmak zaten imkânsızdır. Bunların
Fakültemizde kurulabilmesi için şu anda tam olarak nasıl olacağı kestirilemeyen uzun vadeli parasal
ve teknik desteğe ihtiyaç vardır. Fakültemizde hücre kültürü düzeneğini kurulması için, sadece bir
düzeneğin kurulması yeterli değil, bir de söz konusu hücrenin hangi hücre döngüsü evresinde
bulunduğunu tespit etmek için henüz daha ne flow sitometre (akım sitometresi), ne fluorescently
activated cell sorter (FACS), ne de başka güvenilir bir cihaz vardır. Bu bağlamda, bu yöntem ve
yaklaşımlarının seçiminde büyük zorlukların yaşanacağı aşikârdır, çünkü bu yaklaşımların birçoğu
biopsiden alınan ve buradan üretilen tümör hücrelerinin kullanımına dayanmaktadır. Ayıca da,
Fakültemizdeki sıfatım gereği şu anda bu projeyi bu tür uluslararası boyuta hedeflendirmeyi
mümkün kılmıyor. Bu bağlamda, tarafıma sadece yazılı olarak görev verildiği ve projeme müdahale
edilmeyeceği sözü yazılı olarak verildiği takdirde bu proje konusunu da moleküler genetik
boyutunda gerçekleştirmeyi uygun görüyorum. Çünkü bu konuda yurt dışında ihtisas yaptım. Yurt
dışından tarafıma doktora bursu verilirken de mezun olduktan sonra Türkiye’de bu yeni teknolojiyi
ancak oturttuğum kaydıyla verilmiştir.

118
kanser vakalarda olduğu gibi endokrin terapilerin rolünü vurgulamak gerekir. Bunun
dışında, immünoterapi, adjuvant systemic treatment (kemoradyoterapi ve
kemoimmünoterapi, kemoterapi ile paralel hipertermi) vs. terapi programları
uygulanmaktadır. Bu çok önemli konulara bu kesin raporumun bu giriş bölümünü
daha fazla uzatmamak amacıyla burada yer verilmemiştir. Ancak, bu konularla ilgili
daha ayrıntılı bilgi www.chemotherapy.com sitesinden ve ilgili güncel kitaplardan
edinilebilir (Bk. örneğin: Sliifer S ve Stoter G. Chapter 2: Principles of Systemic
Therapy. In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical
Oncology, Taylor and Francis, New York, pp. 35-49).
1.2.3. Sistemik Moleküler Hedefli Terapilerin Temel Prensipleri.
Son yıllarda sistemik kanser tedavileri17 sitotoksik kemoterapinin kullanımına
dayandırılmıştır. Bu tedavilerin prensiplerine bir önceki bölümde yer verilmiştir.
Seçiciliğin (selectivity) olmadığından, kemoterapi sadece kanser hücrelerini değil,
normal hücreleri ve dokuları da öldürüp, geniş spektrumlu yan etkilere yol
açmaktadır.
Biyolojik ve moleküler araştırmalar hücre çekirdeğinin dışında bulunan ve
karsinojenezde ve kanser oluşumlarında çok önemli rol oynayan birçok olayı ortaya
17 Bu konu ile ilgili daha ayrıntılı bilgi için bk.:
1. Sharma SV, Settleman J. Oncogene addiction: setting the stage for molecularly targeted cancer therapy. Genes Dev. 2007 Dec 15;21(24):3214-31.
2. Bianco R, Damiano V, Gelardi T, Daniele G, Ciardiello F, Tortora G. Rational combination of targeted therapies as a strategy to overcome the mechanisms of resistance to inhibitors of EGFR signaling. Curr Pharm Des. 2007;13(33):3358-67.
3. Hideshima T, Anderson KC. Preclinical studies of novel targeted therapies. Hematol Oncol Clin North Am. 2007 Dec;21(6):1071-91.
4. Rojo F, Dalmases A, Corominas JM, Albanell J. Pharmacodynamics: biological activity of targeted therapies in clinical trials. Clin Transl Oncol. 2007 Oct;9(10):634-44.
5. Bhatti M, Yahioglu G, Milgrom LR, Garcia-Maya M, Chester KA, Deonarain MP. Targeted photodynamic therapy with multiply-loaded recombinant antibody fragments. Int J Cancer. 2007 Oct 31.
6. Voltz E, Gronemeyer H. A new era of cancer therapy: Cancer cell targeted therapies are coming of age. Int J Biochem Cell Biol. 2008;40(1):1-8.
7. Goldenberg DM. "Targeted therapy with monoclonal antibodies: the new generation of pharmaceuticals". Conf Proc IEEE Eng Med Biol Soc. 2006;1:nil21-2.
8. Garman KS, Nevins JR, Potti A. Genomic strategies for personalized cancer therapy. Hum Mol Genet. 2007 Oct 15;16 Spec No. 2:R226-32.
9. Dittmer DP, Krown SE. Targeted therapy for Kaposi's sarcoma and Kaposi's sarcoma-associated herpesvirus. Curr Opin Oncol. 2007 Sep;19(5):452-7.
10. Korfel A, Thiel E. Targeted therapy and blood-brain barrier. Recent Results Cancer Res. 2007;176:123-33.
11. Kalyn R. Overview of targeted therapies in Oncology. J Oncol Pharm Pract. 2007 Dec;13(4):199-205.

119
çıkarmıştır. Bu olaylar hücre sitoplazmasında, hücre membranında18 veya
ekstraselüler ortamda meydana gelmektedir. Şu ana kadar gösterildiği üzere, bu
olayların normal fizyolojik şartlar altında fazla görülmemeleri veya aktivitelerinin çok
düşük düzeylerde olmasına karşın, kanserde önemli rol oynadıkları tespit edilmiş, bu
olayları spesifik olarak baskılayan ilâç formülasyonlarının geliştirilmesi ise bir
sonraki anti-kanser ilâç jenerasyonun (neslinin) tasarımlarında büyük önem
kazanacaktır (Bk.: Şekil 1.4).
Şekil 1.4. Hedeflendirilmiş Tedavilerinin genel gösterimi (Bk.: Ferry A. I. M.
Eskens and Jaap Verweij, Chapter 3: Principles and Examples of Systemic Molecular
Targeted Therapies. In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of
Medical Oncology, Taylor and Francis, New York, pg. 51).
Eğer moleküler hedeflendirilmiş terapiler seçici ve spesifik olarak bu kansere
özgü olaylara müdahale edebilseydi ve böylece spesifik olmayan sitotoksik
18Söz konusu projemin hücresel boyutu da membranlarla ilgilidir, ancak apoptozdan çok antineoplastik ilâçların membran akışkanlığı üzerindeki muhtemel etkilerini vurgulamaktadır. Bu konuda da ilgili öncü hipotez ve model sunulmuştur. Bu konu ile ilgili daha ayrıntılı bilgi için Yorum bölümüne başvurulmalıdır.

120
kemoterapilerin yan etkiler engellenebilseydi, anti-kanser tedavilerinin çok daha
başarılı yapılabileceği açıktır.
Bu konu çok geniş olduğundan buna bu kesin raporumda daha fazla yer
verilmeyecektir. Ancak bu çok önemli konu ile ilgili son derece güncel bilgiler için
Bk.: Ferry A. I. M. Eskens and Jaap Verweij, Chapter 3: Principles and Examples of
Systemic Molecular Targeted Therapies. In: F. Cavalli H. H. Hauser ve S. B. Kaye
(Ed.): Textbook of Medical Oncology, Taylor and Francis, New York, pg. 51.
Kanser hastalarının 60%’dan fazlasında metastazların görüldüğünden, çoğu
kez sistemik olarak etki eden ajanlarla tedavilere başvurulmaktadır. Bu şekilde
sistemik etki mekanizmasına sahip anti-tümör ajanları 3 ayrı gruba dahil
edilmektedir: sitotoksik ilâçlar, hormonal ajanlar, ve biological responce modifiers
adlı bileşenler. Bu gruplarda yer alan çeşitli ajanlar yıllarca uygulanmışlardır (Sliifer
S ve Stoter G., 2005)19. Daha spesifik olarak temel kanser hücre biyolojisi,
farmakoloji, endokrinoloji konulu araştırmalar, ve yeni bileşenlerin ve yeni tedavi
stratejilerin geliştirilmesinin temelini oluşturmaktadır. Bunlara ilâve olarak, başarılı
kanser tedavilerin neticesinde uzun süreli olarak sağlığına kavuşan kişilerin
sayısındaki artış göz önünde bulundurulduğunda, daha sonraları görülen bazı
toksisitelerinin de önemi artmıştır.
Sitotoksik terapiden farklı olarak, kansere özgü olayların işlevsel ve yapısal
özelliklerinin ortaya çıkarılması, bu olayları spesifik olarak baskılayan ajanların
rasyonel olarak geliştirilmesiyle sonuçlandırılacak hedefe-bağlı (a target-based
approach) bir yaklaşımı mümkün kılmaktadır.
Şimdiye kadar ortaya çıkarılan hedefler arasında varlıkları sayesine kontrolden
çıkan hücre bölünmesine (proliferasyona) yol açan genelde proteinler veya işlevi
hasar gören reseptörler veya sinyal proteinleri başı çekmektedir. Moleküler
hedeflendirilmiş tedaviler kanser hücreye özgü bir hedef ile buna bağlı olarak ve
rasyonel olarak tasarlanan ilâçlar arasında spesifik tepkimenin ortaya çıkmasına
19 Tarihçeleri için bk.: Skipper H. E. Historic milestones in cancer biology: a few that are important to cancer treatment. Semin. Oncol., 1979, 6: 506-514.

121
bağlıdır ve bu şekilde normal hücreleri görmeyip, sadece kanser hücrelerini seçici
olarak etkilemeleri amacıyla gündeme getirilmiştir20.
Moleküler hedeflendirilmiş tedaviler RNA ve DNA replikasyonu
etkilemediğinden, aküt hücre ölümü olmamaktadır, ancak hücrelerin kontrol dışı
proliferativ aktiviteleri baskılanmaktadır ve bunlar “quiescence”21 denilen duruma
geçmektedir. Bu hedefe-bağlı kavram seçici olmayan ve tersinir olmayan DNA
hasarının meydana geldiği ve aküt hücre ölümünün gerçekleştiği normal ve kanser
hücrelerinin eşit olarak etkilendiği (Bk.: bir önceki bölüm) konvansiyonel sitotoksik
anti-kanser yaklaşımından tamamen farklıdır.
1.2.4. Gen Aktarımı ve Kanser Tedavilerindeki Rolü
İnsan gen tedavisi, birçok otoimmün ve kalıtsal hastalıkların ve kanser
patolojilerinin tedavisinde şimdiye kadar uygulanan konvansyonel yaklaşımlar
dışında bu hastalıkların tedavilerinde yeni bir umut olarak ortaya çıkmıştır
(GREGORIADIS, 1993; MOLDAWER, 1999; KIRBY, 1999, STONE, et. al., 2000;
FLOTTE, 2001; TEMPLETON, 2003; EL-ANEED, 2004; KOBAYASHİ, et. al., 2005;
MEIDAN, 2006; PELISEK, et. al. 2006) . Gen tedavisi, son yıllarda moleküler biyoloji
ve biyoteknolojideki gelişmeler sayesinde mümkün olmuştur. Olağanüstü öneme
sahip ve uluslararası boyutta olan insan genomun gen dizilişinin ortaya çıkarılması
projesi (Human Genome Sequencing Project) bu yeni tedavi yöntemini, birçok
hastalığın moleküler düzeyde incelenmesini gerçekleştirerek bu doğrultuda yeni
veriler ortaya çıkararak desteklemektedir. Bu gibi çok değerli genetik bilgilerin elde
edilmesiyle, gen tedavisinin yakın gelecekte moleküler tıp alanında önemli
yöntemlerden birini oluşturacağı inanılmaktadır. Gen tedavisinin amacı hastalığa
neden olan genetik bozukluklarının tedavi edilmesidir (TEMPLETON, 2003).
20 Bk.: Chapter 3. Ferry , A. L. M,, Eskens and Jaap Verweij, Principles and examples of systemic molecular targeted therapies, In: F. Cavalli H. H. Hauser ve S. B. Kaye (Ed.): Textbook of Medical Oncology, Taylor and Francis, New York, 2005). 21 Apoptoz olaylarının önemli bir bölümünü teşkil eden “scenescence” olayı doğrultusunda ortaya çıkmaktadır.

122
DNA dizilişindeki nokta mutasyonları teorik olarak tedavisi mümkün olan
birçok hastalığa neden olmaktadır. Genomlarda, nokta mutasyonlarını ortadan
kaldırmak için çok sayıda yaklaşım uygulanmaktadır. Bunlardan hibrid (veya
chimeric) RNA-DNA oligonükleotid veya söz konusu DNA dizilişinin kesilmesi
(deletion of DNA sequences) her ne kadar gelecek vaat eden yöntemler olarak
sunulmuşsa da frekans olarak meydana gelme olasılıkları çok düşük olduğundan,
henüz daha uygulamaları söz konusu değildir. Sonuç olarak, gen tedavisinin amacı
genetik düzenlemelerinin gerçekleştirilmesi olduğundan ve bu da klinik ortamlarda
yapılması bir hayli zor olduğundan, günümüzde gen tedavi yaklaşımların birçoğu
işlevsel genlerin ortama katılmasına dayanmaktadır. Bu yaklaşımda, terapötik genler
farmasötik bileşenler olarak tasarlanırlar ve ortama verilirler. Söz konusu genlerin
hedeflendirildikleri hücre içi bölgeye ulaşma yollarındaki olağanüstü sitoplazmik
engellerin bulunduğundan, in vivo ortamda, yeni ve kontrolü mümkün olan özellikler
taşıyan, güvenilir ve tekrarlanabilir gen tedavi deneysel protokollerin geliştirilmesi
gerekmektedir. Gen yer değişimi tedavinin (Gene Replacement Therapy) dışında, in
vivo gen naklindeki başarı, antijenik proteinin optimal imün cevaba neden olacak
şeklinde naklinin gerçekleştirildiği, DNA aşıların geliştirilmesini de mümkün
yapacaktır GREGORIADIS, 1993; KIRBY,1999; TEMPLETON 2003; KOBAYASHI,
et. al., 2005).
Gen, ilk önce haberci RNA (mRNA)’ya nükleuste transkribe olmakta, daha
sonra ribozomlarda peptide translasyonu gerçekleştirilmektedir; bu peptid daha
sonra katlanma ve glikozilasyon aracılıyla multimerik şekillere dönüşüp işlevsel
protein olarak gen ekspresyonuna katılmaktadır. Sadece DNA nükleotid dizilişindeki
kodları bulunan genlerin tedavi işlevlerini gerçekliştirmeden önce
transkripsyonlarına ihtiyaç duyulur. Ancak, bunlar dışında genelde gen tedavi
olaylarında yer alan tüm diğer durumlarda, kodlanan genlerin transgen
ekspresyununun (rekombinant gen ifadesi) gerçekleşmesi gerekmektedir. Böylece,
nakledilen genin ulaştırılması hedeflenen nükleusa ulaşıncaya kadar herhangi bir
işlevsel biyolojik önemi yoktur. Enjekte edildiği bölgeden nükleusa ulaşıncaya kadar
tedavi amacıyla sentezlenen ve istenilen nükleotidleri içeren yeni kodlayan DNA,
birkaç biyolojik engelle karşılaşır ki bunlar çoğu kez başarılı gen naklini
engellemektedir (Şekil 1.5). Bu engelleri geçmek için, etkili DNA taşıyıcıların
tasarlanması için çok sayıda yaklaşım uygulanmaktadır.

123
1.2.5. Gen Tedavisi ve Gen Nakli
Günümüzde, gen nakli viral ve viral olmayan sistemlerle gerçekleştirilmektedir
(DE LAPORTE, et. al.,2006) (Şekil 1.5-1.8). Taşıyıcılar genel olarak polimerler,
lipidler veya polisakkaridler22 içerip hücresel nükleazlar tarafından muhtemel
degrdasyonu engelleyecek ve daha küçük boyutlu ve daha az miktarda negativ yük
taşıyan ve böylece hücreler tarafından alımı kolaylaştırılmış olan ve intraselüler
haberleşmeyi gerçekleştirebilecek şekilde olmalıdır. Bu doğrultuda, taşıyıcıya
ekstraselüler ve intraselüler engelleri aşmak için uygun fonksiyonel gruplar da
eklenebilir. Bunlar, taşıyıcının serum bileşenleriyle tepkimeye girmesini engeller ve
hücre veya doku türüne özgü bağlanmayı kolylaştıran gruplardır. Söz konusu gruplar
DNA’nın çekirdeğe girmesini endozomlardan kaçışı veya çekirdeğin porlarından
geçişi sağlayarak da kolaylaştırmaktadır. Belirli patojenin veya tümörün antijenini
kodlayan DNA vektörleri önleyici veya terapötik kullanım imkânı sağlayarak bu
patolojilere karşı DNA aşısı geliştirilmesinde önemli rol oynayabilirler
(GREGORIADIS, 1993; KIRBY, 1999). Ex vivo gen tedavisi hastadan söz konusu
hücrelerin ayrıştırılmasını ve bunları daha sonra in vitro olarak terapötik gen ile
değişime uğratılmasını kapsamaktadır (TEMPLETON, 2003; KOBAYASHI, et. al.
2005). Bu şekilde manipüle edilen edilen hücreler daha sonra tekrar hastaya
nakledilmektedir (Şekil 1.6). Diğer yandan, in vivo gen tedavisi (Şekil 1.5 ve 1.6),
direkt olarak önceden tasarlanmış genin hastanın dokularına nakledilmesinden
ibarettir. Her iki yöntemde de, henüz daha başarılmış olmayan ilgili nükleotid
dizilişinin istenilen ekspresyonunu gerçekleştirecek spesifik hedef hücrelere yeterli
miktarda aktiv ve işlevsel DNA’nın nakli konusu çözümünü beklemektedir.
Günümüzde, ilgili hedef hücrelere DNA transferi için birkaç yöntem geliştirilmiştir.
Bunlar fiziksel ve kimyasal (sentetik) yöntemler adı altında iki ayrı gruba ayrılmakta.
22
Ancak çözünürlükleri çok düşük olduğundan, polisakkaridlerin gen taşıyıcı tasarımlarında kullanımları çok kısıtlıdır.

124
Şekil 1.5. Viral ve viral olmayan gen nakli tasarımları. Kaynak:
http://homepages.strath.ac.uk/.../ BGT/BGT12/BGT12.html
Şekil 1.6. Doğrudan ve hücresel gen nakli tasarımları. Kaynak: http://
http://stemcells.nih.gov/info/scireport/chapter11.asp

125
Şekil 1.7. Viral gen taşıyıcı tasarımlarının genel şeması. Yeni gen adenovirüs
vektörüne enjekte edilmekte ve bu modifiye edilen DNA’nın daha sonra ilgili insan
hücresine hedeflendirilmesi için kullanılmaktadır. Tedavinin başarılı olması
durumda, bu şekilde nakledilen genin ürünü olan işlevsel proteinin sentezi
gerçekleşecektir.

126
Şekil 1.8. Çeşitli lipozomal veya polimere dayalı gen taşıyıcılarının hücre içi
(intraselüler) yolu. Burada (a–c) ile gösterilmiş yol, daha sonra böylece tasarlanan
gen taşıyıcının yapı-aktivite özelliklerini de değişterebilen DNA ile komplex
oluşturma yoludur. (d) Taşıyıcı kesitinden gözüken DNA. (e) Taşıyıcılar endositoz ile
hücre içine alınmakta ve daha sonra engel olarka işlev yapan endosomları aşarak
nükleusa girme özelliklerine sahip olmalılar. Bunu takiben tüm kompleks disosiye
olup nuklus içindeki transkripsiyona katılırlar. Kaynak: Laura De Laporte, Jennifer
Cruz Rea, Lonnie D. Shea, Design of modular non-viral gene therapy vectors,
Biomaterials, 27 (2006) 947–954.

127
Şekil 1.9: İntravasküler (sol) veya doku (sağ) enjeksyonundan sonra plazmid
DNA’nın in vivo akıbeti. Uygulandıktan sonra, plazmid DNA makrofajlar gibi tek
hücreli, olan mononükleer fagositlerin de aralarında bulunduğu çeşitli hücreler
tarafından alınmaktadır. Ayrıca, plazmid DNA plazma proteinleri ve ekstraselüler
matriks bileşenleri ile de etkileşmelere girmektedir. Bazı durumlarda, plazmid
DNA’nın ulaşması gereken hedef hücreyi tanıması içinintravasküler yol
(extravasation) veya doku enjeksyonu (difüzyon)’un gerçekleştirilmesi
gerekemektedir. Gen nakli için kullanılan araç ve nakil protokolüne bağlı olarak,
DNA’nın hücrelerce alınması endositotik yol ile (kalın oklar) ve/veya endositotik
olmayan yol (kesintili oklar) ile gerçekleşir. Endositozun gerçekleştiği durumlarda, bu
plazmid DNA enzimatik olarak parçalanacağınan, bu endositozu engellemek
gerekmektedir. Sadece nükleusa ulaşan plazmid DNA’nın terapötik protein
sentezleme şansı vardır (KOBAYASHI, et. al., 2005).

128
Şekil 1.10: Polipleksler ve Lipoplekslerin transfeksyon esnasında hücresel
farklı akıbetlerinin özeti. Lipopleksler ve polipleksler, kırmızı renkli ( lipid veya
polimeri gösteren) ve yeşil (plazmid DNA’yı gösteren) disk veya elips olarak
gösterilmiştir. (1) Serumun yokluğunda her iki vektör türünün de hücreye
bağlanmaları birçok elektrostatik etkileşmelere dayanır. Serum varlığında, vektörlere
bağlanan proteinler kendilerine özgü hücre yüzeyinde bulunan reseptörlerce
tanınmaktadır (bu moleküler tanınma olayı gösterilmemiştir). (2) Bazı lipozomların
katmanıyla plazma membran arasında füzyon meydana gelebilir, ancak bu DNA’nın
hücre içi geçmesiyle her zaman sonuçlanmayabilir. (3) Poliplekslerin veya
lipoplekslerin hücre içi geçiş mekanizması olarak en genel yolun endositozis
üzerinden gerçekleştirildiği bilinmektedir. (4) Polipleks veya lipoplekslerde
enkapsüle olarak bulunan terapötik DNA plazma membranının yakınında bulunan
endozomlardan çok erken salınıma uğramaktadır ve bu olay neticesinde
hedeflendirildikleri çekirdekten (nükleustan) çok uzakta kalmaları yüzünden
intraselüler nükleazlar tarafından degradasyona uğratılmaktadır. 5) Endozomların
sitoplazmik taşınımları rekombinant ve transfektant nükleik asitlerin perinükleer
bölgeye söz konusu DNA’nın daha başarılı bir şekilde çekirdeğe ulaştırılmasında
önemli rol oynamaktadır. Genel olarak buradaki amaç DNA’nın nükleusa ulaşmadan
önce söz konusu endozomlardan kaçışı sağlanmasından ibarettir. Bu kaçışı polipleks
ve lipopleks gen taşıyıcı tasarımları farklı yollar takip ederek meydana getirmektedir.
Lipoplekslerde, örneğin bu füzyon gibi (6) ve (6V ) lipid taşınımı ve membran
destabilizasyonu sayesinde gerçekleştirilmektedeir. Polipleks tasarınmları ise (en
azından polietilenimin (PEI) gibi amin içeren polikatyonlarda veya dendrimerlerde)
tamamen hidrofilik olduklarından (8) endozomların osmotik boşaltımı sayesinde

129
bunu gerçekleştirmektedir. Vektörle DNA’nın birbirinden ayrılmasıyla sonuçlanan
lipopleks disosyasyonu (7) endozomlardan kaçış prosedürü esnasında veya (7V )
sitoplazmada salınıma uğratılan lipoplekslerin Endoplazmik Retikulum gibi
sitoplazmik membran ağı ile füzyonu sonrası meydana gelmektedir. (9) Poliplekslerin
sitoplazmada disosyasyonları henüz daha bilinmeyen başka bir mekanizmaya
gerçekleşir. (10) Endozomlardan kaçışı başaramayan lipopleksler ve polipleksler
lizozomlarca dagrade edilmektedir. Lipoplesklerden veya poliplekslerden ayrılan
transfektant DNA hücre döngüsünün mitozis evresi esnasında nükleer membranın
degradasyonunun olduğu zamanda nükleusa ulaşabilir (şemada gösterilmemiştir).
Bu pasiv nükleer lokalizasyon olayı hücre döngüsünün aktivitesine ve sitoplazmada
plazmidin aktiv kalma süresine bağlıdır. (11) Aktiv, enerji harcanarak gerçekleşen
nükleusa geçiş mekanizması da mevcuttur. Bu da nükleusa geçişi kodlayan,
transkripsyon faktörleri veya DNA-protein kompleksleri gibi makromoleküler
bileşenlerin plazmidte bulunan genleri sayesinde meydana gelmektedir. Ayrıca
lipozomlarla dolu endozomların membranları direkt olarak nükleus membranı ile
füzyon yapabilmektedir (şemada gösterilmemiştir). Kaynak: Abdelatif Elouahabi and
Jean-Marie Ruysschaert Formation and Intracellular Trafficking of Lipoplexes and
Polyplexes, Mol. Therapy, Vol. 11, No. 3, March 2005, 336-347.
Şekil 1.11. Lipozom-DNA veya lipozom-plazmid DNA ile hedef hücre arasında
oluşan elektrostatik etkileşmelerinin şeması. Kaynak: : www.ph.nagasaki-u.ac.jp/.../
doc/photo/main3e.htm

130
Fiziksel gen nakli yöntemlerinden elektroporasyon uygulanmaktadır (MOHR,
et. al., 2002, TEMPLETON, 2003). Ancak bu yaklaşımda kullanılan elektrik
alanlarının DNA’yı parçalayabileceklerinden bu yöntem çok problematiktir. Diğer bir
yaklaşım ise Particle-mediated Gene Transfer (PMGT) yöntemidir ve burada “gene-
gun” olarak bilinen yöntem kullanılmaktadır. Her iki yöntemde de yüksek frekanslı
elektrik akımlarının kullanıldığı son derece ustalık gerektiren yaklaşımlardır. Son
yıllarda fiziksel yöntemler arasında direkt DNA enjeksyonu yöntemi de yerini
almıştır. Ancak bu yaklaşım daha başlangıc aşamısndadır. Bu problemler yüzünden,
günümüzde kimyasal (sentetik) gen taşıyıcı tasarımları tercih edilmektedir
(TEMPLETON, 2003; KOBAYASHI, et. al., 2005).
Hayvan hücresini hedef alarak tasarlanan böyle bir gen nakli genelde şu yolu
takip etmektedir. Küçük DNA içeren nanoparçacıkların oluşumu; bu parçacıkların
hücreler tarafından alınması; parçacıkların hücre membranından geçip sitoplazmaya
girmeleri; DNA’nın çekirdeğe girmesi ve son olarak nakledilen genlerin çekirdekte
gerçekleşen gen ekspresyonu23 (Bkz.: Şekil 1.5-1.11).
Gen nakli görüldüğü gibi etkili DNA veya oligonükleotidlerin hücre zarı
üzerinden geçişine bağlı olarak gerçekleştirilmektedir. Bu olay son derece etkisizdir
ve temelini oluşturan mekanizmalar henüz daha açıklanamamıştır. Ancak, virüsler
evrim boyunca enfekte ettikleri çeşitli hücrelerle beraberce (simbiyotik) bir yaşam
sürdürdüklerinden nanopartiküler yapılarından dolayı ve viral zarının yapısında
bulunan proteinlerce gerçekleştirilen hücre membranı üzerinden geçişi son derece
etkili bir şekilde meydana getirebilirler (STRAYER, 1999; ROBBINS, 1998;
WALTHER, et. al., 2000; SILMAN, 2000; WILSON,et. al., 2002; RITTER, et. al.,
2002; PASSİNİ, et. al., 2004; YOUNG, et. al., 2006; ZHANG, et. al., 2006). Bu
bakımdan, gen nakli için retrovirüsler, adenovirüsler, adeno-associated virüsler
(AAV) gibi birçok virüs türü gen nakli için birçok araştırmacı tarafından klinik
uygulamaya konulmuştur (Şekil 1.7). Gen naklini çok etkili bir şekilde
23
Son iki şemada gösterilen DNA genelde plazmid DNA, oligonükleotid, poliribonükleotid veya diğer model DNA’lardan biridir. Bu nükleik asit türleri arasında her ne kadar moleküler ağırlık ve konformasyon bakımından farklılıklar bulunsa da diğer bileşenlerle kompleks oluşumundaki fizikokimyasal etkenler genelde aynıdır. Bu şemalar, bu yüzden herhangi bir nükleik asit (bunlara antisense ribonukleotid tedavisi veya small interference RNA (siRNA)’ya dayalı tedavilerde olduğu gibi, RNA da dahil olmak üzere) için geçerlidir.

131
başarabildiklerine karşın, viral partiküller bağışıklık sistemini tetiklemekte ve antikor
ve diğer önemli proteinlerin sentezlenmesine neden olmaktadırlar (KAPLITT, et. al.,
1997; BUELER, 1999; HOLT, et. al., 1999; KOOTSTRA, et. al., 2003; FALKNER, et.
al., 2004; HENDRIE, et. al., 2005; HENDRIKS; et. al., 2004). Bu bağlamda,
istenilmeyen proteinlerin sentezlenmesi, klinik uygulamalarında ölümle sonuçlanan
birçok yan etkilere neden olmuşlardır. Bu yüzden, birçok üniversite ve araştırma
merkezi viral olmayan gen taşıyıcıların sentezlenmesine yönelmişlerdir. Viral
olmayan gen taşıyıcılarının geliştirilmesi fizyolojik iyonik ve pH ortamında negativ
yüklü DNA fisfat grupları ile birkaç positiv yüklü gruplar veya polimerik zincirler
arasında oluşan elektrostatik etkileşmelere ve makromoleküllerin yapısal
özelliklerine bağlı olarak hidrofobik etkileşmelere dayanmaktadır. DNA paketlenmesi
veya kondanzasyonu olarak da bilinen bu olay yoğun olarak araştırılmaktadır. Birçok
DNA kondanzasyon olayında, doğal poliamin putresin (H2N(CH2)4NH2), spermidin
(H2N(CH2)3NH(CH2)4NH2) ve spermin’in (H2N(CH2)3NH(CH2)4NH(CH2)3NH2)
türevleri olan bazı katyonik moleküller ile gerçekleştirilmektedir. Genel olarak,
nükleik asit kondanzasyonu morfolojik olarak özgün DNA nanoparçacıkların
oluşmasıyla ortaya çıkarılır. Bu dynamic light scattering (DLS) yöntemi dişinda,
elektron ve floresan mikroskopik ve son yıllarda atomik kuvvet mikroskopik
incelemeleriyle teyit edilmiştir. Ancak, bu küçük katyonik molekülleriyle DNA
arasında oluşan bileşenler çoğu zaman sabit değildir ve serum gibi fizyolojik iyonik
ortamlarda bu gibi kompleks oluşumları disosiasyonla sonuçlanmaktadır
(TEMPLETON, 2003; KOBAYASI, et. al., 2005). Bu yüzden, gen taşıyıcıları olarak,
daha yüksek valanslı poliaminler ve spermidin ve sperminin türevleri sentezlenmiştir.
Bu gibi sentetik polikatyonlar DNA’yı nanopartiküllere dönüştürüp hücre içi geçişi
kolaylaştırmaktadır. Katyonik poliaminler ve polimerler büyük hacimli DNA
moleküllerini işlev düzenleyici gen bölgesini de içerecek şeklinde daha küçük boyutlu
parçacıklara dönüştürebilmektedirler. Sonuç olarak, viral olmayan sentetik taşıyıcı
tasarımlarından birtanesi DNA’yı elektrostatik etkileşmeler sayesinde kondanze eden
ve böylece hücre içi geçişini kolaylaştıran polikatyonik polimerlerin kullanımına
dayanmaktadır (JULIANO, 1991; DRITSCHILO, et. al., 1992; ROMANCZUK, et. al.,
1999; GEBHART, et. al., 2001; ANWER, et. al., 2003; JANG, et. al. 2004). Bir diğer
yaklaşım ise H-bağlar ile DNA’ya bağlanabilen ancak nükleik asit kondanzasyonu ile
sonuçlanmayan, nötr amfifilik polimerleri kullanmakta (GEBHART, et. al., 2001;
ANWER, et. al., 2003). Günümüzde, viral sistemlere nazaran, her iki yöntemin de

132
dezavantajı çok düşük transfeksyon oranlarının olmasıdır (JANG, et. al. 2004).
Yukarıda adı geçen sentetik polimer yapılardan başka, üç ayrı polimer türü
kullanılmaktadır: çeşitli poliamin türevleri, poliamidler ve polivinil türü polimerler.
Bu polimerlerin transfeksyon kabiliyetleri bir birinden çok farklıdır. Ayrıca, yapıları
itibariyle sentezlenmeleri kolay olmakla birlikte, kullanılmakta olan sentetik
polimerler doğal bileşenler olmadıklarından, elektrostatik ve hidrofobik özelliklerinin
kontrolü bir hayli güç olup, bağışıklık sistemini aktive etmektedirler. Viral olmayan
taşıyıcılar ayrıca işlevsel organik grupları da içerebilirler ki bu hücre türüne özgü
hedeflendirmeyi (cell-specific targeting) ve nükleer localizasyonu da
kolylaştırmaktadır. Bu tür yaklaşımlarda en önemli husus, DNA’yı enzimatik
degradasyondan koruyacak şekilde toksik olmayan ve biyolojik olarak yok
edilebilinen (biyodegradable) gen taşıyıcılarının tasarlanmasıdır. Ayrıca, bu
taşıyıcılar membran reseptörlerce meydana getirilen hücresel alımı kolaylaştırıp,
DNA’nın endozomal salınımını da gerçekleştirebilmektedirler (KOBAYASHI, et. al.,
2005).
1.2.6. DNA Kompaktizasyonu ve Nanoparçacık Oluşumunu
Meydana Getiren Bileşenler
Poliaminler.
Poliaminlerden spermidin ve spermin ve bunların sentetik türevleri DNA
kompaktizasyonunu meydana getiren moleküller arasında en etkili olanlar arasında
gösterilmektedir (Şekil 1.12). Poliamin ve analoglarının DNA kondazasyon ve
kompaktizasyon reaksyonlarının >89%’nda bunu yük nötralizasyonu olarak meydana
getirmektedir. Diğer yaklaşımlarda, katyonik poliaminlerden olan cobalt heksamin3+
kullanılmaktadır. EtBr displacement assay kullanarak poliamin homologlarının
DNA’ya farklı afinite gösterdikleri ortaya çıkarılmıştır. Tetravalent poliaminlerin
DNA’yı hücre içi geçiş olayında çok etkili olmamalarına rağmen, heksaminler gibi
daha yüksek valanslı poliaminler bu geçişi daha başarılı bir şekilde
gerçekleştirmektedirler.

133
Şekil 1.12: Doğal poliamin olan sperminin (3−4-3) ve çeşitli türevlerinin
kimyasal yapıları. Poliamin türevleri primer ve sekonder amino gruplarını birleştiren
metilen gruplarının sayısını gösteren numalandırma sistemi ile kısaltılarak
gösterilmiştir (SANTHAKUMARAN et. al., 2005).
Nötr Lipidler
Nötr lipidler (Şekil 1.13) genelde önemli yardımcı bileşen rolü üstlendikleri
katyonik lipid formülasyonlarının bir parçasıdır. Bu bileşiklerde sıkça kullanılan üç
nötr lipid dioleil fosphatidiletanolamin (DOPE), kolesterol ve dioleoil fosfatidil
kolin’dir (DOPC). Birçok durumda, DOPE ile katyonik lipid’in 1:1 oranında
kullanılması en etkili transfeksyon neticelerini vermiştir (PITARD, 2002; ZHANGA,
et. al. 2004). DOPE lipid katmanlarını destabilize ettiği ve endozomal degradasyonda
rol oynadığı bilinmektedir. Katyonik lipid-DNA kompleksleri hücreye endozomal yolu
takip ederek girmektedir ve DOPE’nin işlevi endozomal membranı destabilize
etmekten ibarettir. Kolesterol gibi nötr lipidlerin kullanımı sayesinde, in vivo
transfeksyon düzeyleri yükseltilebilmiştir. Diğer lipozomlara kıyasla, kolesterol içeren
lipozomların uygulandığında birçok organda gen ekspresyon profilleri yükseltilmiştir.
İnsan hepatoma hücrelerinden biri olan, HepG2 ile yapılan deneylerde, DOPE içeren
lipozomlar ve galaktozil türevleri ile konjüge edilmiş kolesterol içeren lipozomlar
daha düşük toksisite ve daha yüksek transfeksyon etkinliğine sahip oldukarı
görülmüştür (PITARD, 2002; ZHANGA, et. al. 2004; SANTHAKUMARAN et. al.,
2005).

134
Transfeksyon etkinliğindeki bu artış karaciğerin parenhimal hücrelerine
spesifik asyaloglikoprotein reseptörlerine karşı yüksek afiniteleriyle açıklanmaktadır
(SANTHAKUMARAN et. al., 2005). Bazı araştırmacılar, kolesterolün in vivo
uygulandığında katyonik lipopoliamin pcTG90 ile nötr lipid olarak konjüge edilmiş
DOPE’ye nazaran, DOPE’nin türevi olan, kısmen florine edilmiş
gliserofosfoetanolamin (F-PE) in vitro ve in vivo gen nakil potansiyeli göstermiştir.
Bu, d-florine edimliş T lipoplekslerin in vivo uygulamalar için DOPE’nin etkin bir
alternatifi olduğunu göstermektedir.
Şekil 1.13. Nötr lipidlerden gen taşıyıcı tasarımlarında bazı sıkça kullanılan
örnekler: dioleyl phosphatidylethanolamine
(DOPE), cholesterol and dioleoyl phosphatidyl choline (DOPC); A partially
fluorinatedglycerophosphoethanolamine (F-PE),.
Katyonik Lipidler
Gen tedavisinin klinik öncesi araştırmalarda DNA naklini başarılı bir şekilde
gerçekleştiren moleküller arasında katyonik lipidler dikkati çekmektedir (AKHTAR,
et. al., 1991; ZHANG, 2004; AZZAM, et. al., 2004; HART, 2005; IGARASHI, et. al.,
2006). Bu moleküller hidrofobik ve polar grup olmak üzere 2 ayrı kısımdan
oluşmaktadır. DNA transfeksyonu için çok sayıda sentetik katyonik lipid
geliştirilmiştir (Şekil 1.14). Bunlar DNA transfeksyon ajanları adı altında ticari olarak
piyasadan da edinilebilir. Bu moleküller lipidin pozitiv yüklü polar baş grubu ile
DNA’nın negativ yük taşıyan fosfat grubu arasında elektrostatik etkileşmeye neden
olup DNA nanoparçacıklarının oluşmasına neden olmaktadır. DNA nanoparçacıklarla

135
lipidler arasında bu şekilde oluşan kompleks sayesinde, DNA’nın hücresel engelleri
daha kolay aştığı ve hücre kültürü ortamında enzimatik degradasyona daha dirençli
olduğu tespit edilmiştir (ZHANGA, 2004). Bu nanopartiküllerin boyutları 50 ile 1000
nm arasında değişmektedir. DNA kompaktizasyonu çok etkili bir şekilde
gerçekleştirmelerine rağmen, piyasada ticari ürün olarak bulunan birçok katyonik
lipid toksiktir. Sıçan ve makaklarla yapılmış deneylerden elde edilmiş veriler yüksek
dozlarda birkaç defa üst üste uygulandıklarında katyonik lipidlerin akciğer
patolojilerine neden oldukları göstermektedir (AKHTAR, et. al., 1991; ZHANGA,
2004; AZZAM, et. al., 2004; HART, 2005; IGARASHI, et. al., 2006).
Şekil 1.14: Katyonik lipidlerden bazı örnekler: lipopoliamin RPR120535,
dioctadesildiamonium bromid
(DODAB), dioleoiltrimetilammonium propan (DOTAP), ve bisguanidinium
tren kolesterol
(BGTC), (N-[1-(2,3-dioleyloxy) propyl]-N,N,N-tri-methylammoniumchloride
(DOTMA). (Bruno Pitard, Supramolecular Assemblies of DNA
Delivery Systems, Somatic Cell and Molecular Genetics, Vol. 27, Nos.1/6,
November 2002; Shubiao Zhanga, Yingmei Xua, Bing Wanga, Weihong Qiaob,
Dongliang Liub,
Zongshi Lib, Cationic compounds used in lipoplexes
and polyplexes for gene delivery, Journal of Controlled Release 100 (2004)
165–180).

136
Polivalent Katyonik Lipidler
DOTAP ve DOTMA’nın gen naklindeki başarısını takiben, birçok başka lipid
hazırlanmıştır. Aralarında, dioktadesilamidoglisilspermin (DOGS veya
ddtransfectamTT) gibi yapıların da örnek olarak gösterilebileceği, polivalent katyonik
lipidler yoğun ilgi çekmişlerdir. Diğer popüler örnekler DOSPA, DPPES, (C8)2Gly
Sper3+ ve (C18)2Sper3+’dir (Şekil 1.15). DOGS, DOTMA’dan DNA ile bağlanma
mekanizması bakımından farklılık göstermektedir. DOGS’un baş grupları poliamin
grubu aracılıyla DNA’nın minor groove ile etkileşmeye girmektedir. DOGS
fazlalığında, DNA’nın paketlendiği, nucleozom benzeri yapı oluşmaktadır. Bundan
farklı olarak, DOTMA sıvı çözeltide lipozomların meydana gelmesine neden
olmaktadır. Bunlar kendiliğinden negativ yüklü DNA molekülleriyle agregatlar
oluşturmaktadır (KOBAYASI, et. al., 2005).
Şekil 1.15: Polivalent Lipidler ve Kolesterol Türevi.

137
Polietilenimin
Polietilenimin (PEI) farklı hücre kültürlerinde ve hayvan modellerinde
oligonükleotidlerin ve plazmid DNA’nın hücre sitozolüne geçişinde etkili rol oynayan
sentetik polimerler grubunun bir bölümünü teşkil etmektedir (Şekil 1.16). Lineer ve
zincirli şeklindeki PEI DNA’yı nanoparçacıklara dönüştürmektedir. DNA
kondanzasyonunda ve transfeksyon etkinliğinde PEI’nin kullanılan şekli ve moleküler
ağırlığı rol oynamaktadır. Fizyolojk pH’da PEI/DNA bileşiklerinin endozomal
degradasyonu daha kolayca aşabildikleri gösterilmiştir. Ancak, bu nanopartiküllerin
çok düşük olan çözünürlülüğü gen naklinde kullanımlarını kısıtlamıştır.
Şekil 1.16: Lineer, zincirli ve polietilen glikol ile konjuge edilmiş
polietileniminler (SANTHAKUMARAN et. al., 2005).
Dendrimerler
Poliamidoamin (PAMAM) ve polipropilenimin (PPI) dendrimerleri (Şekil 1.17)
DNA nanopartiküllerini etkili bir şekilde oluşturma ve DNA naklini kolaylaştırma
kaabiliyetleri dikkati çekmiştir. Lineer ve zincirli polimerlerle kıyaslandıklarında
dendrimerlerin monodispers olmaları ve kontrol edilebilen işlevsel yüzeye sahip

138
olmaları bu molekülleri gen taşıyıcı tasarımlarında kullanılmaları konusu gündeme
getirilmiştir.
Şekil 1.17: Polipropilenimin dendrimerlerinin beş ayrı çeşidinin kimyasal
yapıları. Dendrimerlerin artan karmaşıklığı G1-G5 numaralandırma sistemi ile
gösterilmiştir (SANTHAKUMARAN, et. al., 2005).
Proteinler ve Polipeptidler
Hücrede DNA nanoparçacıkların oluşumu DNA’nın histonlar ve protaminler
gibi katyonik proteinler veya peptidler ile etkileşimi neticesinde
gerçekleştirilmektedir (Şekil 1.18 ve 1.19). Spermde ve bazı virüslerde bulunan DNA
nükleik asitin fosfodiester kısmıyla bağlanabilen Arginine (Arg) zengin bölgeler
içeren protamin tarafından kondanze edilmektedir.

139
Şekil 1.18: (a) Protaminde amino asit dizilişinin şematik çizimi. (b) Argininin
yan gruplarını ayıran alternativ bir yapının gösterimi. Bu şeliyle protaminin DNA ile
etkileşmelere girdiği düşünülmektedir. (c) Protaminin bir diğer olası helikal yapısı.
(Kaynak: F. P. OTTENSMEYER, R. F. WHITING, AND A. P. KORN, Three-
dimensional structure of herring sperm protamine Y-I with the aid of dark field
electron microscopy, Proc. Nat. Acad. Sci. USA, Vol. 72, No. 12, pp. 4953-4955,
December 1975).
Protamin aracılıyla gerçekleştirilen DNA kondanzasyonu arginin grupların
DNA sarmalın her iki zincirini de kilitleyerek nükleik asiti transkripsyonel olarak
aktiv olmayan konformasyona dönüştürmektedir.
Bir katyonik polipeptid olan, polilizin (poly-L-lysine, PLL), DNA ile
etkileşmelere girip nükleik asit nanoparçacıklarınınoluşumuna neden olan ek proton
içeren amin gruplarını yapısında bulundurmaktadır (Şekil 1.20). Bu şekilde
oluşturulan nanoparçacıkların büyüklüğü PLL yapısal özelliklerine (lineer veya
zincirli) ve ortamın iyonik özelliklerine bağımlılık göstermektedir. DNA naklinde çok
etkili olmamasına rağmen, reseptörlere bağlanma kaabiliyeti gen taşıyıcı
potansiyelini artırmaktadır. Ayrıca, PLL-DNA kompleks oluşumu, DNA’yı hücresel
ortamda enzimatik degradasyondan koruduğuna dait raporlar bulunmaktadır
(SANTHAKUMARAN, et. al., 2005).
140
Şekil 1.19: Hering protaminin (protamine (clupeine) Y-I.) yapısı.
Şekil 1.20: Lineer ve zincirli poli-L-lizinin kimyasal yapıları
(SANTHAKUMARAN, et. al., 2005).
Polimerler.
Kitosan (Chitosan) doğal bir polimer olup glikozidik bağlarla birbirleriyle
bağlanan D-glükozamin ve N-asetil-D-glükozamin moleküllerini içeren iki alt

141
biriminden oluşmaktadır (Şekil 1.21). Nispeten daha az toksiktir ve DNA
nanopartikül oluşmasını takiben transfeksyon etkinliği oldukça yüksektir. Böylece,
kitosan ve çeşitli türevleri gen nakli konusunda gelecek vaat eden bileşikler olarak
görülmektedir.
Şekil 1.21: Kitosan (Chitosan)’ın kimyasal yapısı (SANTHAKUMARAN, et. al.,
2005).
Poli(etilen glikol) (PEG) ve poli(etilen oksid) (PEO) gibi nötr polimerler de
yüksek pI’de DNA kondanzasyonuna neden olmaktadır. Bu yük taşımayan ve esnek
polimerler “excluded volume mechanism” adıyla bilinen mekanizma ile etki
gösterirler. DNA kondanzasyonunun satbilitesini ve transfeksyon etkinliğini artırmak
amacıyla, çoğu zaman PEG PEI, folat reseptörleri ve poli-L-lizin ile konjuge edeilir
(SANTHAKUMARAN, et. al., 2005).
Sonuç olarak, üniversitelerde ve sanayii kuruluşlarının araştırma-geliştirme
(AR-GE) merkezlerinde, DNA ile reaksyona giren ve DNA nanoparçacıkların
oluşumuna neden olan çok sayıda sentetik veya doğal bileşenin viral olmayan gen
taşıyıcıların tasarlanmasındaki kullanım olasılıkları yoğun olarak araştırılmaktadır.

142
1.3. Taxol® (Paclitaxel).
1.3.1. Kanser Tedavisinde ve Kanserin Önlenmesinde Taxol®’un
Rolü
Konvansiyonel ilâç geliştirmesi genelde deneysel hayvanların ve hücre kültürü
düzeneklerinin kullanıldığı faz-1, faz-2 ve faz-3 olarak adlandırılan aşamaları içeren
klinik öncesi araştırmaları takip etmektedir. Genel olarak da, faz-3 çok sayıda
gönüllünün iştirak ettiği klinik deneylerinden ibarettir. Söz konusu aşamalı prosedür
çoğu zaman komplementer veya alternativ tedavilerle sonuçlanmamaktadır. Buna
ilâve olarak, etkili konsantrasyonları önceden ayarlanabilen total olarak sentezlenmiş
bileşenler dışında, doğal bitkisel kökenli malzemeden izole edilen aktiv olan etken
madde miktarları genelde çok düşüktür. Aynı şekilde, komplementer ve alternativ
terapilerle sonuçlanabilecek klinik öncesi araştırmalar çok nadir olup, double-blind
klinik araştırmalar da son derece nadir olarak uygulanmaktadır (Stohs, S.J., 2005).
Bu tür komplementer ve alternativ terapilerin izlediği aşama ve gelişmelerden farklı
olarak porsuk ağacından (Pacific Yew Tree, Taxus brevifolia, bk. Şekil No: 1.22) izole
edilen ve günümüzde antikanser tedavierinde çok başarılı bir şekilde yoğun olarak
kullanılan Taxol® (Paclitaxel24)’in buluşu ve geliştirilmesi olmuştur.
24 Jenerik ismi (Bk.: Stohs, S.J., 2005). Bu raporumda her iki (ticari ve jenerik) isim de kullanılmıştır.

143
Şekil 1.22.: Porsuk ağacının farklı türleri, Taxus brevifolia, Taxus baccata vs. (www.turkforum.net; www.conifers.co.nz; home.scarlet.be ve çok prestijli Max-Plank
Enstitüsü-caliban.mpiz-koeln.mpg.de sitelerinden alınmıştır).
Taxol®‘un saflaştırılması, karakterizasyonu, klinik öncesi ve klinik araştırma
süreçleri Food and Drug Administration (FDA)’nın konvansiyonel ilâç geliştirme ve
piyasaya sunma stratejilierini takip etmiştir (Wall, M.E ve Wani, M.C., 1995; Harlan,
Jr., W.R., 2001; Stohs, S.J., 2005)25.
1960’larda, National Cancer Institute (NCI) U. S. Department of Agriculture
(USDA) tarafından rastgele bitkisel kökenli örneklerin toplandığı ve bunların
antineoplastik aktivitelerinin tarandığı bir program başlatmıştır. Toplanan bitkiler
arasında, Washington Eyaletinden getirilen T. brevifolia’nın gövdesinden kısımlar,
yapraklar ve meyveler de bulunmaktaydı (Stohs, S.J., 2005). Bunlarla yapılan ilk
taramalar çeşitli neoplastik hücrelere karşı potansiyel aktivite endeksi olarak da
25 Bk.: Harlan, Jr. W. R., New opportunities and proven approaches in complementary and altenative medicine research at the National Institutes of Health. J. Altern. Compl. Med., 7, S53, 2001; Wall, ME.. and Wani, M.C., Camptothecin and taxol: discovery to clinic (13th Bruce F. Caine Memorial Award Lecture), Cancer Res., 55, 753, 1995; Wall, ME.. and Wani, M.C., Camptothecin and taxol: from discovery to clinic. J. Ethopharmacol., 51, 239, 1995- Taxol® ‘un geliştirme tarihçesini ve klinik kullanımını özetleyen bir derlemedir.

144
kabul edilen, T. brevifolia ekstraktlarının 9KB hücrelere karşı yüksek bir
sitotoksisiteye sahip olduğu saptanmıştır (Stohs, S.J., 2005)26.
1964 yılında, Research Triangle Institute, Research Triangle Park, N. C.,’da
bulunan Wall’un laboratuvarı, National Cancer Institute tarafından gönderilen ilk T.
brevifolia bitkisel malzemeleri incelemek üzere kabul etmiştir. Saflaştırma ve
sitotoksisite prensipleri Walker WM katı tümör inhibisyonunu içeren biyoaktivite
göstergeleri kullanarak yapılmıştır (Stohs, S.J., 2005). Kasım 1966’dan itibaren, Wall
ve Wani (Wall, M.E ve Wani, M.C., 1995) bu bitkiden çok yüksek sitotoksik aktiviteye
sahip bir fraksyon izole etmişlerdir. 2 yıllık bir saflaştırma sürecini takiben, standart
etanol ekstraksyonu, H2O ile kloroform arasında oluşan etanol kısmının faz ayrımını
sonucu counter current dağlımını içeren aktiv bileşen izole edilmiştir. Taxol ismi, söz
konusu molekülün hidroksil grupları içerdiği (-ol) ve taxane çekirdeğinin bulunduğu
yapıya dayanarak (Bk.: Şekil 1.26.), T. brevifolia’dan izole edilmiş bileşene verilen bir
isimdir.
Taxol®‘un yapısal analizi, ultraviyole (UV; morötesi), kızılötesi (IR) ve kütle
spektrometresi ölçümleriyle yapılmıştır ve 1971 yılında (Wani et al., 1971) bu yapı
ayrıntılı olarak aydınlatılmıştır27. Taxol®‘un (Paclitaxel’in) yapısı Şekil 1.26’da
verilmiştir. 1960’lı yıllarda yapılan araştırmalarda Taxol®‘un L1210 ve 1534 türü
lösemi hücrelere karşı aktivitesi bulunduğu gösterilmiştir. Taxol®‘un ayrıca birçok
katı tümöre karşı aktiv olduğu gösterilmiştir (Wall, M.E ve Wani, M.C., 1995; Stohs,
S.J., 2005).
Taxol®‘un etki mekanizmasının aydınlatılması, bu sitotoksik ajanın klinik
araştırmalarda kullanılmasında önem taşımıştır. İlk araştırmalar, bir mitotik
inhibitör olarak etki ettiği ve böylece yeni ve daha başka örneği bulunmayan bir
mekanizma söz konusu olduğunu kanıtlamıştır28. Taxol®‘un yapısı ve fizikokimyasal
özellikleri Ek-7 verilmiştir. Daha ayrıntılı bilgi için Ek-7’e başvurulmalıdır.
26 Bk.: Stohs, S.J., Taxol in CCncer Treatment and Chemoprevention. In: Phytopharmaceuticals in Cancer Chemoprevention, Debasis Bagchi and harry G. Preuss (Eds.), CRC Pres, Boca Raton, London, New York, Washington D. C., 2005, pp. 519-524. 27 Bk.: Wani, M.E., Taylor, H.L. Wall, M.E., Coggan, P. and Mc Phail, A.T., Plant antitumor agents VI: the isolation and structure of taxol, a novel antileukemic and antitumor agent from Taxus brevifolia, J. Am. Chem. Soc. 93, 2325, 1971. 28 Bk.: Fuchs, D. A ve Johnson, R. K. Cytologic evidence that taxol, an antineoplastic from Taxus brevifolia, acts as a mitotic spindle poison, Cancer Treat. Rep., 62, 1219, 1978.

145
Ancak, daha sonraki araştırmalar söz konusu mekanizmanın mikrotübülleri
stabilize ettiğinden ve bunların tekrar tübüline geri depolimerizasyonunu
engellediğinden, yani kolisin, vinkristin, vinblastin ve podofilotoksin gibi diğer
antimitotik ajanların etki mekanizmalarının aksine çok farklı bir yol takip ettiğini
göstermiştir (Schiff, P. B., et al. 1979; Horwitz, S. B., 1992)29.
Hayvanlar üzerinde deneyler 1982 yılında yapılmış olup, faz-1 klinik aşaması
1983-84, faz-2 ise 1983-86 yıllarında tamamlanmıştır (Wall, M. E. ve Wani, M. C.,
1995; Stohs, S. J., 2005). ABD Ulusal Kanser Enstitüsü (NCI) 1991 yılında yeni ilâç
uygulama şemasını elde ettiğinden dolayı Cooperative Research and Development
Award (CRADA) adlı ödülü Bristol-Myers Squibb şirketine vermiştir.
Taxol® ilk olarak son derece yavaş büyüyen Taxus brevifolia’nın kabuğundan
izole edilmiştir. Taxol® çok düşük miktarlarda bu ağacın kabuğunda bulunmaktadır
ve bundan dolayı uzun süre klinik kullanımı sorun olmuştur. Bu sorun, başka taxus
türlerinde, örneğin T. baccata’nın yapraklarında çok daha yüksek miktarlarda
bulunan baccatin III veya 10-deacetylbaccatin III’ten Taxol®’un yarısentezini
gerçekleştiren Bristol-Myers Squibb şirketince çözülmüştür (Bk.: Şekil 1.24).
Taxol®’un total sentezi ile ilgili bk.: Ek-8. Taxol®’un total sentezi iki ayrı grup
tarafından yapılmıştır. Yapısından da anlaşılacağı üzere (Şekil 1.26), çok sayıdaki
asimetrik C-atomu total sentezi son derece güçleştirmektedir. Taxol®’un söz konusu
total sentezi bu ilâcın pratik olarak stoklanması anlamına gelmemesine rağmen, o
yıllarda Taxol®’un kimyası hakkında önemli bilgiler vermiştir. Bunun dışında,
Taxol®’un analoğu olan Taxotere™ (Docataxel) (Şekil 1.27) hazırlanmıştır, test
edilmiştir ve her ikisinin de klinik olarak kullanılabilecekleri kararına varılmıştır
(Stohs, S. J., 2005).
1.3.2. Taxol®’un ve Taxotere™’in Klinik Kullanımı.
Rahim karsinom ve non-small-cell lung kanser vakalarında radyoterapi veya
cerrahi müdahalelerin uygun görülmediği hastalarda cisplatin ile birlikte Taxol® ilk
başvurulan çok etkili bir ilâçtır (Bk.: Şekil 1.25). Aynı şekilde, refraktor metastatik
29 Bk.: Schiff, P. B., Fant, J. ve Horwitz, S. B., Promotion of microtubule assembly in vitro by taxol, Nature, 22, 665, 1979: Horwitz, S. B., Mechanism of action of taxol, Trends Pharmacol. Sci., 13, 134, 1992.

146
rahim karsinom vakalarında ve kombinasyon terapinin başarısızlıkla
sonuçlandığında metastatik hastalık için veya adjuvant kemoterapi sonrası dönemler
için bu ilâç birinci derecede önem kazanmıştır. Bunun dışında, AIDS’e bağlı Kaposi
sarkomlarda ikinci derecede önemlidir (Stohs, S. J., 2005).
Ayrıca, standard tedavi yöntemlerden şimdiye kadar üstünlüğü kanıtlanmış olsa
bile, head and neck karsinomlarda Taxol®’un yine etkili bir şekilde kullanılmaktadır.
4-hidroksitamoxifen ile birlikte kullanıldığında, estrojen-reseptör-negativ barsak
kanserine ve akciğer kanser hücre kültürlerinde etkili olduğu gösterilmiştir (Stohs, S.
J., 2005). Bu kombinasyon, tek başına kullanıldığı zaman gösterdiği etkiden çok daha
yüksek bir potansiyeli olduğu gösterilmiştir (Stohs, S. J., 2005).
Taxol®’un çok sitotoksik olduğundan, bu ilâçla tedavi edilen hastalarda yan
etkilerinin de görülmesi beklenmektedir. Tek başına Taxol® ile tedavi edilen katı
tümörlü hastalaın 90%’nında nötropeni ve lökopeni gibi omur iliği baskılanmaları
görülmüştür. Bu ilâçla tedavi edilen hastaların 75%’inde ise anemi görülmektedir.
Sonuç olarak, Taxol®’un fizikokimyasal özelliklerini optimize etmek suretiyle bu yan
etkileri azaltmak mümkün olacaktır. Bu amaçla, bu ilâçın çeşitli kontrollü salınım
sistemleriyle denenmesi gerekmektedir. Projemde yer alan Taxol®-fosfolipid
kompleks oluşumunun incelenmesi de bu hususu vurgulamaktadır. Bu tür lipide
(lipozomal) bağlı formülasyonlarının geliştirebilmesi kuşkusuz Taxol® ‘un yan etkileri
ve çözünürlülük problemleri çok daha azaltılmış olarak uygulama fırsatı yaratacaktır.

147
Şekil 1.23. Taxol’un popülaritesini gösteren bir şekil. Dünya üniversitelerin 80%’inde kullanılan Genel Kimya ders kitabının kapağında yer almaktadır (Bk.: General, Organic, and Biochemistry (Hardcover); by Katherine J Denniston, Joseph J
Topping, Robert L Caret, McGraw-Hill Science/Engineering/Math; 4 edition (Mar 4 2003).

148
1.24. Bristol-Myers Squibb tarafından yayımlanan monografi (Kaynak: www.ja-text.de/Print.php).

149
(Kaynak: www.brandinstitute.com/NEWS/MEDAD_11_04.HTM)

150
1.25. Üst kısım: Kuzey Amerika’da klinik kullanımda olan en popüler ilâçları gösteren bir kıyaslama tablosu. Alt kısım: Taxol® tedavisinin etkinliği (Kaynak: www.rmj.ru/articles_3834.htm)31. Bu şekil daha önce verilen Şekil 1.2 ve 1.3 ile
kıyaslanmalıdır.
30 Bu sonuçlar aşağıdaki yayınlara dayanmaktadır: 1. Pazdur R., Kudelka A.B., Kavanagh J.J., et al. // The taxoids: paclitaxel (Taxol) and docetaxel (Taxotere). // Cancer Treat. Rev. 1993, 19:351–386. 2. Gelmon K. // The taxoids: paclitaxel and docetexel. // Lancet, 1994, 344:1267–1272. 3. Bissery M.C., Nohynec G., Sanderink G.J., Lavelle F. // Docetaxel (Taxotere): a review of preclinical and clinical experience. Part 1: preclinical experience. // Anticancer Drugs. 1995, 6:339–368. 4. Hortobagyi G.N. // Recent progress in the clinical development of docetaxel. // Sem. Oncol. 1999, 26 (3 Suppl.9):32–36. 5. Bissery M.C., Guenard D., Gueritte–Volgelein F., Lavelle F. // Experimental antitumor activity of Taxotere (RP 56976, NSC 628503), a taxol anologue. // Canc. Res. 1991; 51:4845–4852. 6. Harrison S.D., Dykes D.J., Shepherd R.V., Bissery M.C. // Response of human xenografts to taxotere. // Proc. AACR 1992, 33:526. 7. Bruno R., Hille D., Thomas L., et al. // Population pharmacokinetics/ pharmacodinamics of docetaxel (Taxotere) in phase II studies. // Proc. ASCO 1995, ab. 147. 8. Fumoleau P., Chevallier B., Kerbrat P., et al. // Current status of taxotere as a new treatment in breast cancer. // Br. Canc. Res. Treat. 1994; 33:39–46. 9. Cortes J.E., Pazdur R. // Docetaxel. // J. Clin. Oncol., 1995:2643 10. Vikeljia S.J., Baker W.J., Burris H.A. III, et al. // Peridoxine therapy for palmarplantar erythrodysesthesia associated with taxotere. // N.Nat. Canc. Inst. 1993; 85:1432. 11. New P.Z., Jackson S.E., Rinaldi D., et al. // Peripherial neurotoxicity secondary to docetaxel. // Neurology 1996; 46:108. 12. Semb K.A., Aamdal S., Oian P. // Capillary protein leak syndrome appears to explain fluid retention in cancer patients who receive docetaxel treatment. // J. Clin. Oncol. 1998; 16:3426. 13. Lembersky S., Anderson R., Smith A. et al. “Phase II Trial of Doxorubicin and Docetaxel for Locally Advanced and Metastatic Breast Cancer: Preliminary Results from NSABP BP–57.” // Pr. ASCO.– 2000.– Vol. 19.– abstr. 403. 14. Raab G., Wilke H., Eidtmann H. et al. “Phase II Stady of Docetaxel and Epirubicin as First Line Chemotherapy in Metastatic Breast Cancer.” // Pr. ASCO.– 2000.– Vol. 19.– abstr. 442.

151
15. Tubiana–Hulin M., Bonneterre J. et al. “Better survival with epirubicin–docetaxel (ET) combination as first–line chemotherapy in patients with metastatic breast cancer (MBC): final results of a phase II randomized study.” // Pr. ASCO.– 2003.– Vol. 23.– P182. 16. Горбунова В.А. «Таксаны в новых лекарственных комбинациях». // Мат. Конф. «Новые противоопухолевые препараты в лечении рака». – Москва.– 28–30 сентября 1999г. 17. Nabholtz J.M., et al. // A phase III trial comparing doxorubicin and docetaxel (AT) to doxorubicin and cyclophosphamide (AC) as first line therapy for MBC. // Proc. ASCO, 1999; 18:1279. 18. Marty M., et al. // Randomized Phase II trial of the Efficacy and Safety of Transtuzumab Combined with Docetaxel in Patients with Human Epidermal Growth Factor Receptor Z–Positive Metastatic Breast Cancer Administered As First–Line Treatment. The M77001 Study Group. // J. Clin. Oncol. 23:4265–4274, 2005. 19. Sato et al. // High pathological response with the combination of docetaxel and transtuzumab as preoperative systemic treatment in breast cancer patients overexpressing Her2. A multicenter phase II study of JECBC. // Abctract ESMO 067, 152 p. 20. Nabholtz J.M., Plenkowski T., Mackey J., et al. // Phase III trial comparing TAC (docetaxel, doxorubicin, cyclophosphamide) with FAC (5–fluororacil, doxorubicin, cyclophosphamide) in the adjuvant treatment of node positive breast cancer patients: interim analysis of the BCIRG 001 Study. // Proc. ASCO 2002, ab. 141. 21. Aura A Erazo Valle–Solis et al. // Cost–effectiveness analysis of adjuvant docetaxel, doxorubicin and cyclophosphamide regimen (TAC) versus 5–fluorouracil, doxorubicin and cyclophosphamide (FAC) in early breast cancer at the Mexican social security inctitut (IMSS) Mexico. // Abstract ESMO 062, 294 p., 2006. 22. Martin M., Pienkowski T., Mackey J., et al. // Adjuvant docetaxel for node–positive breast cancer. // N. Engl. J. Med., 2005; 352:2302–13. 31 Bu sonuçlar aşağıdaki yayınlara dayanmaktadır: 1. Pazdur R., Kudelka A.B., Kavanagh J.J., et al. // The taxoids: paclitaxel (Taxol) and docetaxel (Taxotere). // Cancer Treat. Rev. 1993, 19:351–386. 2. Gelmon K. // The taxoids: paclitaxel and docetexel. // Lancet, 1994, 344:1267–1272. 3. Bissery M.C., Nohynec G., Sanderink G.J., Lavelle F. // Docetaxel (Taxotere): a review of preclinical and clinical experience. Part 1: preclinical experience. // Anticancer Drugs. 1995, 6:339–368. 4. Hortobagyi G.N. // Recent progress in the clinical development of docetaxel. // Sem. Oncol. 1999, 26 (3 Suppl.9):32–36. 5. Bissery M.C., Guenard D., Gueritte–Volgelein F., Lavelle F. // Experimental antitumor activity of Taxotere (RP 56976, NSC 628503), a taxol anologue. // Canc. Res. 1991; 51:4845–4852. 6. Harrison S.D., Dykes D.J., Shepherd R.V., Bissery M.C. // Response of human xenografts to taxotere. // Proc. AACR 1992, 33:526. 7. Bruno R., Hille D., Thomas L., et al. // Population pharmacokinetics/ pharmacodinamics of docetaxel (Taxotere) in phase II studies. // Proc. ASCO 1995, ab. 147. 8. Fumoleau P., Chevallier B., Kerbrat P., et al. // Current status of taxotere as a new treatment in breast cancer. // Br. Canc. Res. Treat. 1994; 33:39–46. 9. Cortes J.E., Pazdur R. // Docetaxel. // J. Clin. Oncol., 1995:2643 10. Vikeljia S.J., Baker W.J., Burris H.A. III, et al. // Peridoxine therapy for palmarplantar erythrodysesthesia associated with taxotere. // N.Nat. Canc. Inst. 1993; 85:1432. 11. New P.Z., Jackson S.E., Rinaldi D., et al. // Peripherial neurotoxicity secondary to docetaxel. // Neurology 1996; 46:108. 12. Semb K.A., Aamdal S., Oian P. // Capillary protein leak syndrome appears to explain fluid retention in cancer patients who receive docetaxel treatment. // J. Clin. Oncol. 1998; 16:3426. 13. Lembersky S., Anderson R., Smith A. et al. “Phase II Trial of Doxorubicin and Docetaxel for Locally Advanced and Metastatic Breast Cancer: Preliminary Results from NSABP BP–57.” // Pr. ASCO.– 2000.– Vol. 19.– abstr. 403. 14. Raab G., Wilke H., Eidtmann H. et al. “Phase II Stady of Docetaxel and Epirubicin as First Line Chemotherapy in Metastatic Breast Cancer.” // Pr. ASCO.– 2000.– Vol. 19.– abstr. 442. 15. Tubiana–Hulin M., Bonneterre J. et al. “Better survival with epirubicin–docetaxel (ET) combination as first–line chemotherapy in patients with metastatic breast cancer (MBC): final results of a phase II randomized study.” // Pr. ASCO.– 2003.– Vol. 23.– P182. 16. Горбунова В.А. «Таксаны в новых лекарственных комбинациях». // Мат. Конф. «Новые противоопухолевые препараты в лечении рака». – Москва.– 28–30 сентября 1999г. 17. Nabholtz J.M., et al. // A phase III trial comparing doxorubicin and docetaxel (AT) to doxorubicin and cyclophosphamide (AC) as first line therapy for MBC. // Proc. ASCO, 1999; 18:1279.

152
Şekil: 1.26. Taxol®‘un kimyasal yapısı. Molekülün diterpen kısmı sarı renkle gösterilmiştir. Daha iyi renkli bir görünüm için bu proje raporumla
birlikte sunduğun CD’ye bakınız. Bu diğer şekiller için de geçerlidir.
Şekil 2.27. Docataxel veya Taxotere™ (Kaynak: chimie.scola.ac-paris.fr/Sitedechimie/chi_org...).
18. Marty M., et al. // Randomized Phase II trial of the Efficacy and Safety of Transtuzumab Combined with Docetaxel in Patients with Human Epidermal Growth Factor Receptor Z–Positive Metastatic Breast Cancer Administered As First–Line Treatment. The M77001 Study Group. // J. Clin. Oncol. 23:4265–4274, 2005. 19. Sato et al. // High pathological response with the combination of docetaxel and transtuzumab as preoperative systemic treatment in breast cancer patients overexpressing Her2. A multicenter phase II study of JECBC. // Abctract ESMO 067, 152 p. 20. Nabholtz J.M., Plenkowski T., Mackey J., et al. // Phase III trial comparing TAC (docetaxel, doxorubicin, cyclophosphamide) with FAC (5–fluororacil, doxorubicin, cyclophosphamide) in the adjuvant treatment of node positive breast cancer patients: interim analysis of the BCIRG 001 Study. // Proc. ASCO 2002, ab. 141. 21. Aura A Erazo Valle–Solis et al. // Cost–effectiveness analysis of adjuvant docetaxel, doxorubicin and cyclophosphamide regimen (TAC) versus 5–fluorouracil, doxorubicin and cyclophosphamide (FAC) in early breast cancer at the Mexican social security inctitut (IMSS) Mexico. // Abstract ESMO 062, 294 p., 2006. 22. Martin M., Pienkowski T., Mackey J., et al. // Adjuvant docetaxel for node–positive breast cancer. // N. Engl. J. Med., 2005; 352:2302–13.

153
1.3.4. Taxol®’un Etki Mekanizması.
Taxol®’un hücresel mikrotübüllerdeki32 bağlanma bölgesi hücresel işlevi
bilinmeyen bir farmakolojik hedeftir. Mikobakteri kaynaklı epotilonlar, deniz yosunu
kaynaklı diskodermolidler ve yumuşak mercanlardan elde edilen elenterobinler ibi
doğal bileşenler, mikrotübüllerdeki bağlanma bölgesine yerleşerek Taxol®’un
sitotoksik etkilerine benzer etki göstermektedirler. Mikrotübülleri stabilize eden
bileşenler adı verilen bu moleküller, mikrotübüllere bağlanarak dinamiklerini bloke
etmekte ve böylece hücre bölünmesini sonlandırmaktadırlar (Bk.: Şekil 1.28-1.31).
Taxol® ve türevleri yumurtalık kanseri, metastatik göğüs kanseri, head and neck
kanseri ve böbrek kanseri tedavilerinde başvurulan başlıca ilâçtır (Bk. özellikle Şekil
1.25. Ayrıca bk.: Şekil 1.2 ve 1.3). GTP aracılıyla Taxol®’un mikrotübüllere bağlanma
noktası olarak mikrotübül lumeninin (boşluğunun) β – tubulin altbirimi
belirlenmiştir (Bk.: Şekil 1.31).
Taxol®’un bağlanma kinetiğini ve ligandlara karşı afininitesini incelemek için
en duyarlı yöntem olarak floresan işaretli taxoid Flutax-2’in kullanılması olmuştur
(Şekil 1.32). Görüldüğü üzere, floresan mikroskopik inceleme taxoidlerin çözeltideki
mikrotübüllere doğrudan bağlandığını göstermektedir. Cyclostreptin hücre
mikrotübülleri stabilize eden ilk moleküldür ve bunlara kovalent olarak bağlanarak
tersinir olmayan bir etki yapmaktadır. Bu ligandı kullanarak, Taxol®’un
mikrotübüllere bağlanma mekanizması ve ayrıca da biden fazla bağlanma noktası
bulunduğunu ortaya çıkarmıştır. Bu kovalent bağlanma, P-glikoproteini eksprese
eden hücre kültürlerinde bu ilâcı aktiv hale getirmektedir.
32 Hücre mikrotübülleri hakkında daha ayrıntılı bilgi için Moleküler Hücre Biyolojisi ders kitaplarına başvurulmalıdır.

154
Şekil. 1.28. Taxol®’un hücre bölünmesi üzerine etkisi (Kaynak: pubs.rsc.org/ej/CC/2001/b100070p/).
Şekil 1. 29. Taxol®’un mikrotübül dinamiği üzerindeki etkisi (Kaynak: pubs.rsc.org/ej/CC/2001/b100070p/).

155
Şekil 1. 30. Taxol®’un hücre mikrotübüller üzerinde ve programlanmış hücre ölümüne (apoptoza) yol açan çeşitli etkileri. (Kaynak: biologyofcells.blogspot.com/2007/12/microtubu...).

156
Şekil 1.31. Taxol®’un mikrotübül dimerlerine bağlanma şekli (Kaynak: pubs.rsc.org/ej/CC/2001/b100070p/).

157
Şekil 1.32. Mikrotübül stabilize edici moleküllerin hüceresl mekanizmaları. PtK2 hücrelri during 7 saat boyunca cyclostreptin yokluğunda (A) veya 5 µM (B) ve 10 µM paclitaxel (C) varlığında. Hücreler daha sonra yıkanıp, bu hücre kültüründe 16 saat boyunca bu şekilde bırakılmışlardır, kontrol (D), 5 µM cyclostreptin (E) ve 10 µM
paclitaxel(F). (Kaynak: biologyofcells.blogspot.com/2007/12/microtubu...).
1.3.5. Projemin Bilimsel Dayanağı.
Yukarıda ayrıntılı olarak verilen Taxol® etki mekanizması uyarınca,
mikrotübüllerin iç yüzeyinde meydana gelen bu tür üç boyutlu bağlanma şekli,
prensip olarak uzay geometrisi engelleri bakımından bağlanmayı çok zor bir hale
getirmektedir. Bu yüzden Taxol®‘un bağlanma afinitesi, kinetiğini ve üç boyutlu
şekilleri hakkında ek bilgilere ihtiyaç vardır. Çünkü bu tür bilgiler daha ileriki
aşamalarda Taxol®‘un hem daha etkili farmasötik terapötik formülasyonlarının
geliştirilmesini, hem de hücre seviyesindeki Taxol®‘un etki mekanizmasını çok daha
iyi bir şekilde anlaşılmasını mümkün kılacaktır. Bu her iki nokta da projemde
özellikle vurgulanmıştır. Bu bağlamda da, şu ana kadar teklif edilen etki
mekanizmasına muhtemel alternativler sunulmalıdır. Tarihsel olarak da zaten, aynı
mantık ve düşünce tarzı anestetiklerin etki mekanizmaları için de izlenmiştir.
Örneğin, anestetiklerin33 etkileri sadece Meyer veya Overton yasalarına dayanarak
Octanol-Water Partitioning sabitlerini hesaplayarak açıklanamamıştır. Çok daha
33 Aynı gerekçeler diğer ilâçlar için de geçerlidir.

158
sonraları bunların etki mekanizmalarına projemde de olduğu gibi lipid teorileri dahil
edilmiştir34.
Bu projemde de zaten Taxol®‘un bu etki mekanizmalarını aydınlatacak niteliği
taşıyan ek bir öncü hipotez sunulmuştur. Çünkü söz konusu etki mekanizması,
kullanılan tümör hücre kültürü türüne göre de farklılıklar göstermektedir. Projemde
öncü hipotez olarak Taxol®‘un hücre zarının akışkanlığını düzenleyerek ve dolayısıyla
da buradaki ilâç difüzyonlarını ve reseptörlerin dinamiğini kontrol ederek etki
mekanizması teklif edilmiştir. Literatür taraması da gösteriyor ki35, şu ana kadar
böyle bir bağlantı çalışılmamıştır. Bu bakımdan projemin orijinalliği tartışılamaz hale
getirilmiştir. Bu hususları Bilimsel Araştırma Proje Birimine daha projemin ilk sunuş
metninde açıkça ifade edilmiştir. Daha çok ayrıntı için bu proje teklifim
incelenmelidir. Söz konusu hipotezim de normal hücrelerle tümör hücreleri
arasındaki membran akışkanlığında gözlenen farklılıklarına dayanmaktadır36. Bu
konuya ayrıca Yorum bölümünde de ayrıntılı olarak değenilmiştir.
Multidrug Resistance (MDR) adı verilen antineoplastik ajanlara karşı gelişen
direnç mekanizmalarının ortaya çıkması sonucu Taxol® ile gerçekleştirilen
kemoterapilerde her zaman tatmin edici sonuçlar alınmamaktadır. Söz konusu MDR
olayları P-glikoprotein (P-gp) gibi membran pompapalarının aşırı ekspresyonu
sonucu meydana gelmektedir ve kemoterapötik ilâçları sitoplazmadan hücre dışına
pompalayarak bunların intraselüler konsantrasyonunu ve böylece de kemoterapilerin
etkinliğini düşürmektedir. P-gp de membran glikoproteini olduğundan Taxol® ile
hücre zarları arasında oluşan moleküleretkileşmelerinin önemini vurgulamaktadır.
Projemde de bu etkileşmeler çeşitli fosfolipid molekülleri kullanarak model hücre zarı
yüzeyleri ile modellenmiştir. MDR hücre türlerini öldürebilen Taxol® ligandlarını
bulmak için bu projemde elde edilen verilere dayanarak daha sonraki aşamalarda bu
modelleri dirençli ve dirençsiz hücre kültürleri üzerinde denenebilir. Bu amaçla, bu
projemde geliştirilen etkileşme modellerine dayanarak Taxol®’un bazı tümör hücre
kültürlerinin hücre mikrotübülleri üzerinde, hücre bölünmesi ve hücre döngüleri
üzerindeki etkileri incelenebilir. Ancak bu şekilde bu ligandların afinitelerinden yola
34 Bu konularla ilgili özellikle bk.: Drug-Membrane Interactions. Analysis, Drug Distribution, Modeling. J. K.
Seydel and . Wiese (Eds.)., Methods and Principles in Medicinal Chemistry, Vol. 15, Wiley-VCH Verlag GmbH, Weinheim, Germany, 2002, 35 Bk.: Ek-5 ve Ek-6. 36 Bu çok önemli konu için bk.: Chapter 9: Membranes in Cancer. In: Howard R. Petty, Molecular Biology of
Membranes. Structure and Function, Plenum Press, New York and London, 1993, pp. 353-374.

159
çıkarak araştırılan maddelerin Taxol® ve türevlerinin bağlanma noktasındaki
sitotoksisitelerini ortaya çıkaracaktır ve böylece yüksek afiniteli bileşenlerinin
tasarımını ve sentezini mümkün kılacaktır. Bu hususlar projemin moleküler hücresel
boyutunu teşkil etmektedir.
Bunun dışında projemin bir de daha pratik biyoteknolojik boyutu da vardır.
Sunduğum projenin nanofarmasötik biyoteknolojik boyutu olarak, model hücre
yüzeyi - Taxol® nanometrik bileşenlerin fizikokimyasal özelliklerinin yeni bir
laboratuvar protokolü geliştirerek şu ana kadar çok az uygulanan Langmuir-Blodgett
tek katman (lipid monolayer) düzeneği ile incelenmesi ile ilgilidir. Şimdiye kadar
yayımlanan deneysel tasarımlardan farklıdır ve kanımca orijinal bir yaklaşımdır. Şu
ana kadar yayımlanmış çalışmalarda, Taxol® - hücre membran lipid modellerini
teşkil eden antineoplastik ajan veya bir başka model terapötik formülasyon ve
katiyonik lipidlerle girdikleri tepkimeler neticesinde elde edilen bileşenlerin daha
sonraki intraselüler penetrasyon deneyler esnasında hücre kültüründe terapötik
bileşenlerin uygun bir şekilde taşınma kapasiteleri ile ilgili olarak, ön biyofiziksel
ölçümlere ve bunu takip eden morfolojik karakterizasyonlardan ibarettir. Ancak, söz
konusu katiyonik lipidler sitotoksik olduklarından, in vivo ortamda serum
stabiliteleri son derece düşüktür. Sunduğum projede ise, orijinal bir alternativ olarak
katiyonik lipidler yerine zwitteriyonik (nötr) fosfolipidlerin kullanılmasını
önermekteyim. Bunlar arasında aynı amaçla tasarlanan nükleik asitlere dayalı
formülasyonlarının fizikokimyasal sabitlilikleri ve termodinamik özellikleri
yayımlanmıştır. Ancak, aynı modelin daha küçük hacimli antineoplastik ajanlar ile de
geçerli olup olmadığı henüz daha araştırılmamıştır. Ayrıca, sekonder yapıları ve bu
dinamik yapılardaki meydana gelen değişikliklerin hücre içi geçiş yaparak hücre
çekirdeğindeki DNA’ya bağlanma potansiyellerini nasıl etkilediği konusu henüz daha
bilinmemektedir.
1.3.6. Literatür Özeti
Taxol®’un etki mekanizmasıyla ve tedavi potansiyeli ile bir hayli fazla yayın
bulunmaktadır. Bunların en güncel olanları Ek-5’te verilmiştir. Bunun dışında ilâç
tedavilerine paralel olarak son yıllarda çok popüler olan gen aktarım yöntemleriyle
kanser tedavi profilleri de Ek-6’da verilmiştir.

160
2. GEREÇ VE YÖNTEM
Bu projemde Taxol® (Paclitaxel) ile model hücre yüzeyleri arasında oluşan
moleküler etkileşmelerin simüle edilmesini mümkün kılan bir düzeneğin
tasarlanması amaçlanmıştır. Söz konusu düzeneği anlayabilmek için yüzey kimya ve
kolloid kimya kavramlarını çok iyi bilmek lâzım. Özellikle hücre membranın yapısı ve
fizikokimyasal özellikleri ile yüzeysel aktivite kavramları çok önemlidir. Bu amaçla,
bu proje raporumu inceleyecek olanlara yardımcı olmak amacıyla aşağıdaki bölümde
ilk önce hücre membran yapısı ile ilgili güncel bilgiler verilmiştir. Daha sonra da
projemde geliştirilen deneysel protokole ayrıntılı olarak yer verilmiştir. Bu protokole
dayanarak, başka ilâçların da membran afiniteleri incelenebilir.
2.1. HÜCRE MEMBRAN YAPISI
2.1.1. Biyomembranların İçeriği
Membranlar hücre işlevlerinin yerine getirilmesinde hayati bir rol oynar.
Plazma membranı hücreyi çevrileyerek iç hacminde bulunan sitozol ile dış çevre
arasındaki önemli farklılıklarını muhafaza eder. Hücre içi ortamda farklı bileşen, yapı
ve işleve sahip birçok organel ve cisimcik bulunur. Organeller yerine getirdikleri
işlevlerine göre farklı olarak isimlendirilirler ve böylece eukaryot hücrelerde
endoplazmik retikulum, Golgi cismi, mitokondri, kloroplastlar ve membrana-bağlı
organeller ayırt edilmektedir (Şekil 2.1).

161
Şekil 2.1. Hücre membranı (www.microscopy-uk.org.uk/.../membranes.html
sitesinden alınmıştır).
Membranlar her organelin iç hacmindeki içeriği ile sitozol arasında bulunan
karakteristik farklılıkları muhafaza eder. Hücre membran yapısının kavramı Singer
ve Nicholson tarafından (Şeki 2.2) teklif edilen ve sıvı-akışkan veya “fluid-mozaic”
membran modeli olarak da adlandırılan lipid çift katman (lipid bilayer) yapıya
dayanmaktadır. Günümüzde hücre membranı kavramı karbonhidratlar, pigmentler,
antikorlar, reseptörler ve diğer proteinleri de içerecek şeklinde geliştirilmesine
rağmen (Şekil 2.3), hala geçerliliğini koruyan fluid-mozaic membran modeline göre,
hücre membranlarını oluşturan fosfolipidler amfipatik olup suya afinitesi yüksek olan
(hidrofilik) ve sudan uzaklaştırılan (hidrofobik) kısımları içerir.

162
Şekil 2.2. Biyomembranların günümüzde geçerli olan Singer ve Nicholson modeli
(www.umass.edu/microbio/rasmol/cutlips.htm).

163
Şekil 2.3. Biyolojik membranların yapısı
(www.umass.edu/microbio/rasmol/cutlips.htm).
Şekil 2.4. Biyomembran yapısında yer alan fosfolipidlerden tipik bir görünüm (Bk.:
www.umass.edu/microbio/rasmol/cutlips.htm ve
www.agen.ufl.edu/.../lect/lect_06/lect_06.).
Bu makromoleküller birçok hayvan hücre membran hacminin yarısını
oluşturup, geriye kalanı ise proteinlerden oluştuğu bilinmektedir. Fosfolipidlerin
çift katmanlar (bilayers, Şekil 2.4) söz konusu lipidlerin bu amfipatik doğasından ve
şekillerinden kaynaklanmaktadır. Böylece, lipid moleküllerin su tarafından
çevrildiklerinde hidrofobik kısımlarının iç hacminde ve hidrofilik başlar da suya
doğru yer alacak şekilde çeşitli agregatlar ve toplaşımlar (self-assemblies) meydana
getirirler.

164
Hücre membranların lipid çift katmanları (Şekiller 2.2-2.4) fosfolipidler,
kolesterol ve glikolipidler gibi üç ayrı çeşit membran lipid molekülü içermektedir.
Hücre membranının iç ve dış tek katmanları (monolayers) lipid içeriği bakımından
birbirinden farklıdır. Buna benzer farklı lipid karışımları eukaryotik hücrelerin
çeşitli organel membranlarında da mevcuttur. Böyle bir asimetrik membran lipid
çift katman modeli söz konusu lipidlerin iki boyutlu lipid molekülleri içinde
yerleşmiş membran proteinlerin bulunduğu yapısal tasarımına dayanmaktadır.
Böylece, bazı membran proteinleri işlevlerini yerine getirmek için spesifik baş
gruplarına ihtiyaç duyar. Aynı şekilde, suyun proteinler için bir üç-boyutlu çözücü
rölü oynar ki böylece birçok enzim bu çözeltilerde aktiviteleri için seçici olarak bir
metal iyona gereksinim duyar.
Biyolojik membranların temel yapısı lipid çift katmana dayanmasına rağmen,
spesifik işlevlerin çoğu proteinlerce gerçekleştirilmektedir. Bu doğrultuda,
membranda yer alan proteinlerin miktarı ve türü geniş sınırlar içinde
değişmektedir. Sinir hücresi aksonlarda elektrik yalıtkanlığı görevi yapan proteinler
membran kütlesinin 25%’den daha düşük bir miktarı oluştururken, mitokondrinin
ve kloroplastlarda iç membranların enerji iletimi görevi üstlenen zarda ise ~75%’i
proteindir. Genel olarak, plazma membranın içeriğinin 50% proteinlere düşer.
Protein moleküllere kıyasla lipid molekülleri daha küçük olduklarından her zaman
hücresel ortamlarda proteinlerden daha fazla oranda lipid molekülleri bulunur. Bu
oranı genelde membranlarda her proteine karşılık olarak 50 lipid molekülün
bulunduğu gerçeğiyle de açıklanır.
Membran proteinleri birçok işlev görür. Plazma membranında dış uyarılara
sensör cevabını gerçekleştiren proteinler bulunur. Bu şekilde bu protein sensörleri
veya reseptörleri bilgi taşınımında da görev yapar. Diğer membran proteinleri
zarlar arasındaki iyon gradyantlarını meydana getirir. Bu gradyantlar ATP
sentezinde, önceden seçilmiş maddelerin transmembran taşınımında, veya kas ve
sinir hücrelerinde elektrik sinyallerin iletiminde kullanılır. Değişik işlevlerine
rağmen, tüm biyolojik membranlar genel bir yapıya sahiptir-her biri lipid ve protein
moleküllerince oluşturulan ince film tabakasıdır (Şekiller 2.2-2.4).

165
Biyolojik membranların iki boyutlu akışkanlar olduğu kavramı membranın
yapı-aktivite ilişkisinin ortaya çıkarılmasında büyük bir rol oynamıştır. Ancak,
membranı sadece lipid denizinde serbestçe yüzen proteinlere dayanan model bir
hayli basitleştirilmiş olduğu ortaya çıkmıştır. Birçok hücre meydana getireceği
işlevin kolaylaştırılması doğrultusunda birçok lipid ve protein molekülü lipid çift
katmanı üzerindeki belli bir yüzeyde yoğunlaştırır. Böylece membran domenler ve
membran dinamiği kavramları geliştirilmiştir. Bu kavramlar, şimdiye kadar
vurgulanan membran proteinlerin öneminin yanı sıra membran lipid türlerinin
oluşturduğu domenlere (lipid rafts) dikkati çekmektedir. Örneğin, birçok lipid
molekülü membran proteinlerden farklı olarak çift katmanı oluşturan iki ayrı tek
katmanlar arasında yer değiştirmektedir. Membran proteinleri ise membran içinde
lateral olarak hareket eder. Sonuç olarak, hem lipidlerin hem de proteinlerin bu
hareketliliği membranın çok dinamik bir yapı olduğu ve hücre membran
akışkanlığının biyolojik önemini ortaya koymuştur. Örneğin, çift katman
viskozitesini laboratuvar deneylerinde suni olarak belli bir seviyenin üzerine
çıkartarak tamamen ortadan kalktığı gözlenmiştir. Lipid çift katmanın akışkanlığı
lipidlerin içeriğine ve ortam ısısına bağlıdır. Böylece, çift katmanın sıvı fazdan jel
faza geçişini gösteren karakteristik freeze noktasında daha kısa hidrokarbon
zincirlerinin veya çift bağ içerildiğinde bu nokta daha düşüktür.
2.2. Biyolojik Membranların Sıvı Kristal Özellikleri
2.2.1. Lipid Çift Katman
Lipid-protein çift katman hidrofobik etkileşmeler sayesinde meydana gelen bir
supramoleküler yapıdır. Bu etkileşmeler molekülleri imobilize etmek için yeterince
güçlü olmadıklarından çift katman fizyolojik ortamda sıvı özellikleri gösteren, aynı
zamanda da yapısal olarak kristal şekilleri alabilmektedir. Söz konusu sıvı kristal
fazı tüm molekülleri kapsamamaktadır. Sıvı kristal fazını oluşturan moleküller
genelde uzun zincirli olup moleküler ağırlıkları MW=200-500 Da’dan birkaç bin Da
arasında değişmektedir. ESR spektroskopisini kullanarak ilk defa Hubbel ve
McConnel37 çift katmanın üst ve alt kısmında bulunan lipid hidrokarbon zincirinin
hareketliliğinin yüksekliğini gözlemişlerdir. Bu olay daha sonra Levine ve Seelig
tarafından 2H- ve 13C-NMR spektroskopisi kullanarak teyit edilmiştir. Bu
37 Hubbel, W. L. and McConnel, H. M. (1971): Molecular Motion in Spin-Labelled Phospholipids and Membranes, J. Amer. Chem. Soc., 93: 314-326.

166
araştırmalar neticesinde yağ asitlerinin son kısmında bulunan metil gruplarının
hareketli olduğunu ortaya çıkarılmıştır.
2.2.2. Membranın Fizikokimyasal Özelliklerini Etkileyen Faktörler
Fosfolipidler, glikolipidler ve diğer basit amfifiller faz değişimi ısısı (phase
transition temperature veya Tm) adı verilen bir ısı seviyesinde endotermik sıvı
kristal↔katı (jel) faz değişimine uğrarlar (Şekiller 2.5-2.6 ve Tablo-4).

167
Şekil 2.5. (A) Fosfolipid faz değişim geçişleri (phase transitions) ve bunlara
kolesterolün etkisni gösteren bir diferansiyel taramalı kalorimetrik (DSC)
termogram; (B) Fosfolipid konfigürasyonlarının katı ve sıvı fazları.
Lipidlerin katı fazda düzenli, sıvı-kristal fazda ise nispeten düzensiz
olduklarından, bu ısı noktasına ayrıca düzen-düzensizlik geçiş noktası (order-
disorder transition temperature) adı da verilmektedir. Her lipidin kendine özgü bir
Tm değeri vardır (Bk. Tablo-2.1).
Şekil 2.6. Bir maddenin faz değişimleri.
168
Tablo-2.1. Biyomembranlarda Bulunan Bazı Fosfolipidlerin Faz Değişim Isıları.

169
Lipidlerin sıvı-kristal şekilleri Lα fazını oluştururlar. Katı fazı ise Lβ şekli olarak
adlandırılır. Lβ şeklinde moleküller çift katman eksenine doğru hidrokarbon
zincirlerin tam olarak uzunluğunu muhafaza edip all-trans konfigürasyonu ve
quasi-heksagonal toplaşımlar oluşturur (Şekil 2.5-B).
Lipidlerin faz değişim mekanizmaları çeşitli fizikokimyasal yöntemler
kullanarak katı fazdaki çift tabakada ve lipid moleküllerin paketlemelerinde
(elektron mikroskopla görüntülenen “grain borders” in freeze frakture replicas)
düzensizliklerin ve defektlerin olduğunu ortaya çıkararak tespit edilir. Söz konusu
defektlerin doğal entropinin (düzensizliğin) ifadesi olduğundan, meydana gelme
olasılığı ısının yükselmesiyle artar. Faz değişimi noktasında, defektler lipidin ilk
erime noktaları olarak kabul edilirler ve bunlar akışkan lipid moleküllerinde küçük
domenler oluştururlar. Bu yüzden, faz değişimi noktasında hem akışkan hem de
katı domenler aynı anda (eşzamanlı) meydana gelir. Bu ısının Tm değeri üzerine
yükseltildiğinde geride kalan katı faz lipidleri hızlı bir şekilde eriyip akışkan faza
geçerler. Faz değişimi noktasında üç ayrı faz aynı anda meydana gelmektedir: katı
faz, sıvı-kristal faz ve interfacial lipid phase adı verilen ara faz. Interfacial lipid
phase katı ve sıvı-kristal fazları arasında yer alır ve faz değişimin meydana
gelmesinde en önemli rolü üstlenir.
Bu üç fazda yer alan lipidler hidrokarbon zincirleri arasında eşzamanlı
olmayan paketlenme olasılıkları yüzünden faz değişimi noktasında lipid çift
tabakanın geçirgenliğinin artmasına neden olur. Faz değişim noktasının üzerinde
bulunan lipidler sıvı-kristal fazında bulunur.
2.2.3.Lipid Fazlarını Etkileyen Faktörler ve Biyosistemlerdeki
Önemi
Lipid faz geçişleri ortaya çıkarıldıktan sonra bu konu ayrı bir bilim dalı
olarakgelişmiştir. Söz konusu geçişler fizikte “lyotropic phase transitions” olarak
adlandırılan teori ile açıklanmaktadır. Bu konuyla ilgili çok iyi matematiksel
ifadeler de geliştirilmiştir. Burada bunlardan projemizin amacına uygun bir şekilde

170
ve sadece lipidlerde görünen olgulardan çok kısa bahsedilecektir38. Lipid fazlarını
etkileyen faktörler kısaca şunlardır.
Çift bağların varlığı, özellikle cis-çift bağlar, CH-zincirlerinin all-trans
paketlemelerine engel olmaktadır ve Tm’i düşürmektedir. Çift bağların sayıları ve
CH-zincirlerinin pozisyonları da ayrıca Tm’i etkilemektedir. Genel olarak, Tm
hidrokarbon zincirin uzunluğuna bağlı olarak değişim gösterir. Diğer etmenler
arasında fosfolipid baş grubunun yükü ve konformasyonu, hidrokarbon zincirinde
yer alan C-atomlarının sayısı, iyonik ortam ve yüzeysel pH fosfolipid çift tabaka
akışkanlığını etkileyen faktörlerdir.
2.2.4. İlâçların Biyomembran Akışkanlığı Üzerine Etkisi
İlâç – hücre membranı etkileşmeleri uzun yıllardan beri araştırma gruplarının
ilgisini çekmektedir. Bu ilginin başlıca nedenleri kısaca şöyle özetlenebilir. Biyolojik
veya farmakolojik aktivitesi olan herhangi bir molekül kendine özgü
lipidçözünürlülüğüne göre hücrelerin hidrofobik bölgelerine yerleşip, ve buralarda
etkili konsantrasyonlarda bulunuyorsa, lipid çift katmanın yapısında
deformasyonlara neden olacaktır. Bazı ilâçlar, sıvı ortamında çeşitli ligandlarla bağ
yapacaklarından hiç bir zaman bu etkili konsantrasyonlara ulaşamayacaklardır. Söz
konusu ilâçlar membranı çok daha düşük konsantrasyonlarda etkileyebilmektedir
ve bu yüzden bu etkileri sadece in vitro olarak gözlenebilmektedir. Basit kimyasal
yapılar olarak ilâçlar sıvı ortamında pek etkili olmadıklarından ve yeterli
miktarlarda membrana bağlanmayınca zararlı etki gösteremeyeceklerinden büyük
bir ihtimalle membranların parçalanmasına neden olabilmektedir.
Bu ilâçların bazıları çok yüksek lipid çözünürlülüğüne sahiptir, ancak yüksek
lipid çözünürlülüğü membran üzerine etki göstermek için yeterli olmayabilir. Bu
olayda tek gereken şey söz konusu membranın daha düşük dozlarda bulunan
ilâçların etkilenmemesidir. Bu konu ile ilgili elde edilen veriler ve bunlara
dayanarak geliştirilen modeller anestetikler ve alkoller gibi ve bunlara benzerlik
gösteren ilâçlara dayanmaktadır. Bu ilâçlarla ilgili uzun yıllardan beri lipid yapısı ve
lipid faz değişimleri üzerine etkileri araştırılmıştır ve birçok veri elde edilmiştir. Bu
38 Bu konu ile ilgili daha ayrıntılı bilgi için Fizikokimya ders kitaplarına başvurulmalıdır.

171
sınıfa giren ilâçlardan anestetikler etki mekanizmaları bilinen tek grup olmamakla
birlikte en iyi bilinen bileşenlerdir.
İlâç – hücre membranı etkileşmeleri ile ilgili olarak geliştirilen teoriler ve
modeller ilâçların membran lipidleriyle veya membran proteinleriyle
gerçekleştiğine dair Overton dağılım kurallarına uygun olarak iki ana grupta
toplanmaktadır. İlâçların membran lipidlerine mi yoksa membran proteinlerine mi
etki ettikleri konusu uzun süredir bilimsel tartışma konusudur. Bu konu ile ilgili,
herhangi bir hidrofobik fazda meydana gelen ve ölçülebilen fiziksel değişim ilgi
çekicidir. Yüksek lipid çözünürlülüğüne sahip ilâçların membran lipidlerin ve
proteinlerin her ikisinin de hidrofobik kısımlarıyla etkileşmeye girecekleri açıktır.
Ancak, bu durumda bile hidrofobik etkileşmelerinin hücre işlevlerini nasıl bir
şekilde ve hangi yönde etkilediklerini açıklamak son derece güçtür. Bu hususları
açıklığa kavuşturmak için ilâç-hücre etkileşmeleriyle ilgili daha fazla
araştırmalarının yapılmasına ihtiyaç duyulmaktadır. Bu beklenti araştırılmakta olan
ilâcın hem hücre membran lipidleriyle hem de membran proteinleriyle etkileşmeye
gireceği varsayımına dayanmaktadır. Ancak, bu durumda çok muhtemel olarak bazı
lipidler ve proteinler diğerlerine kıyasla daha çok etkilenecektir. Bu yüzden, bunları
tespit etmek ve söz konusu ilâcın bu moleküllere göre neden daha yüksek afinite ve
seçicilik gösterdiğini ortaya çıkarmak en önemli hedefler arasındadır. Çünkü bunu
başarılı bir şekilde ortaya çıkarmak ve her durumda çalışan bir model ve teoriyle
destekleyerek, hem genel bir etki mekanizmasının keşfedilmesine, hem de daha
etkili ilâç tedavi yöntemlerinin geliştirilmesine yardımcı olacaktır.
Bazı ilâçlar hücre üzerinde kendilerine özgü reseptörleri bulunsa bile
hidrofobik etkileşmeye girebilmektedir. Örneğin, bazı benzodiazepinler ve
nöroleptikler yüksek lipid çözünürlülüğüne sahip olup bölgeye özgü mekanizmaları
doğrultusunda etki ederler. GABA veya dopamin gibi bazı ajanların doğal
ligandlarının çok yüksek hidrofilisiteye sahip olduklarından mutlaka sıvı ortamda
etkileşmeye girdikleri bilinmesine rağmen, gerçek etki bölgesi sıvı ortamı mı yoksa
membranın mı olduğu henüz daha tam olarak bilinmemektedir.
Lipofilik ilâçların membran içi konsantrasyonları sıvı ortamdaki
konsantrasyonlarından birkaç bin defa daha yüksektir ve bu gerçek afinite ligand

172
bağlanma deneylerinde her zaman göz önünde bulundurulmaktadır. Bu ilâçların
bağlanma afiniteleriyle ilgili farmakokinetik denklemler ve matematik modeller
ortaya konulmuş ve uygulanmakta olmasına rağmen bu konu hala yeterince
çalışılmış değildir.
2.2.5. Biyomembran Akışkanlığının Anlamı
Genel olarak, membran akışkanlığı membran bileşenlerinin çeşitli hareket
biçimlerine verilen isimdir. Matematiksel olarak bazı varsayımlardan yola çıkarak
1/viskozite olarak da ifade edilir. Bu hareketler kısaca şunlardır: açil zincirlerinin
fleksibilitesi ve esnekliği, membran ekseni istikametinde moleküllerin lateral
difüzyonu, bir tek katmandan diğer tek katmana moleküllerin tersine difüzyonu ve
lateral faz farklılaşmasıyla sonuçlanan faz değişimleri (phase transitions). Tüm bu
olaylar anizotropik hareketler olup ve akışa mukavemet ve izotropik direnç gücü
olarak bilinen viskozite ile ifade edilemez. Bu moleküler hareketlerin her biri çeşitli
ilâçlar tarafından etkilenmektedir.
Daha genel anlamda, bir hayli fazla farmasötik ajanı membranları daha
akışkan veya daha düzensiz hale getirmektedir. Bu özellikle lipofilisitesi yüksek olan
fenol halkaları içeren heterosiklik yapılarda görülmektedir. Biyomembranın daha
akışkan veya daha düzensiz hale gelmesi kuşkusuz membran lipidlerin ve membran
proteinlerin işlevleri üzerine tersinir olmayan önemli etkileri meydana
getirmektedir.

173
Şekil 2.7. Langmuir-Blodgett düzeneklerden genel bir görünüm.

174
Şekil 2.8. DMPE (1,2-Dimyristoyl-sn-glycero-phosphoethanol- amine)
fosfolipidin 20°C’deki izotermi. Air-water ara yüzeyinde DMPE
monomolecular film (monolayer) olarak ortaya çıkan farklı fazlar gösterir.
Görülen plato non-ordered liquid-expanded (LE) fazdan ordered liquid-
condensed (LC) faza geçişten ibarettir. Hidrofilik silikon substratına yavaşça
transferi esnasında DMPE tek katmanın kondanze olduğu noktada basınç
değişimi kırmızı çizgi ile gösterilmiştir.

175
Şekil 2.9. Langmuir-Blodgett tek katmanlardaki bir fosfolipidin uğradığı faz
değişimleri. Tipik bir izoterm gösterilmiştir. Yüzey üzerindeki fosfolipid
moleküllerin hareketliliği açıkça gözükmektedir.
Şekil 2.10. Langmuir-Blodgett tek katmanlarda Wihelmy plate ile ölçülen
yüzey basınç yönteminin bir görünümü.

176
Şekil 2.11. Farklı Langmuir-Blodgett tek katman ve çok katman düzenekleri.

177
Tek katman ( Bk.: Şekiller 2.7-2.11)39, birtakım organik moleküllerin sıvı ara
fazına uygulanmasıyla oluşturulur. Projemde uygulanan fosfolipidler genelde
Şekil 2.10 ve Şekil 2.13 gösterildiği şekilde hidrofilik baş kısımları su fazına
doğru, hidrofobik kuyruk kısımları ise gaz (hava) faza doğru yerleşir ve tek
katman bu şekilde oluşturulur. Söz konusu moleküller su yüzeyine40 ilk olarak
uygulandıklarında çok fazla serbestliğe sahip olarak yüksek bir hareketlilik
gösterirler. Bu su üzerinde her moleküle düşen alan çok fazladır ve yüzey
basınç düşüktür. Bu yüzey basınç genelde sensör sistemine bağlı Wilhelmy
palte (Bk.: Şekil 2.10 ve Şekil 2.11) adı verilen düzenekle ölçülür. Yüzeysel
basınç hesaplamalarının matematiksel ifadeleri için özellikle bk.:
www.kibron.com, www.ksvltd.com, www.nima.co.uk43,44. Bu yüzeysel basınç birbirine
karşı hareket ettirilen 2 bariyer sayesinde yükseltilmektedir (Şekiller 2.7-2.11).
Yüzey basıncın çok fazla yükselmeye başladığı bir noktaya varılır ki bu lipidin
sıvı faza geçişini göstermektedir (Bk.: Şekil 2.8 ve Şekil 2.9). Bariyerlerin
birbirine karşı istikamette daha ileri hareket ettirildiklerinde, katı fazın
değişikliğe uğradığı, yüzey basıncın daha hızlı bir şekildeki artışından da
gözlenmektedir (Şekil 2.8).
39 Bu bölümde, bu proje raporumu inceleyenlere kolaylık sağlanması amacıyla, fosfolipidlerle oluşturulan tek katmanlarla ilgili çok kısa ön bilgiler verilmiştir. Ancak, fosfolipid yüzey kimya ve kolloid teorileri matematiksel olarak da karmaşık olduğundan, bu konu ile ilgili çok daha ayrıntılı bilgilere ulaşmak için bk.: Ek-13. 40 Projemde elde edilen neticelerin tekrarlanabilirliğini test edilmesi bakımından suya ek olarak 2 ayrı tampon da denenmiştir. 41 Ayrıca Bk.: Ek-13. 42 Ancak şu anda piyasada satılan modern Langmuir-Blodgett tek katman cihazlarında son derece
uygun software programları yüklenmiş olup söz konusu hesaplamalar interaktiv olarak bilgisayar
üzerinde gösterilmektedir. Bunun için bu proje raporumun Bulgular bölümünde yer alan neticeler ve
buradaki izoterm eğrilerinin bulunduğu çıktılar incelenmelidir. 43 Ayrıca Bk.: Ek-13. 44 Ancak şu anda piyasada satılan modern Langmuir-Blodgett tek katman cihazlarında son derece
uygun software programları yüklenmiş olup söz konusu hesaplamalar interaktiv olarak bilgisayar
üzerinde gösterilmektedir. Bunun için bu proje raporumun Bulgular bölümünde yer alan neticeler ve
buradaki izoterm eğrilerinin bulunduğu çıktılar incelenmelidir.

178
Şekil 2.12. Bir reseptörün Langmuir-Blodgett tek katman üzerinde
imobilizasyonu.
Langmuir-Blodgett tek katmanların farklı düzenekleri yukarıdaki şekillerde
gösterilmiştir. Şekil 2.12 ise Langmuir-Blodgett tek katmanların farmakolojik ve
biyolojik anlamı çok daha yüksek olan bir düzeneği-bir reseptörün oluşturulan
fosfolipid tek katman üzerinde imobilizasyonunu göstermektedir. Bu şekilde
imobilize edilen reseptörlerin bunlarla bağlanan ilâç ve diğer ligandlarının bağlanma
kinetiğinin ölçülmesinde çok orijinal bir ölçüm imkânı vermektedir. Bu projemde bu
boyut amaçlanmamıştır. Çünkü böyle bir düzeneğinin kurulması için ek cihazlara
ihtiyaç vardır. Ancak fakültemizdeki bundan sonraki projelerim böyle bir düzeneğin
kurulmasını kapsamaktadır. Bu hususlara ayrıntılı olarak Ek-16’da yer verilmiştir.

179
2.3. Biyomembran ve Model Membran Araştırmalarında Kullanılan
Başlıca Yöntemler45
Günümüzde, biyomembran ve yapay membran modelleri araştırmalarında
birçok fizikokimyasal, moleküler biyolojik ve biyokimyasal yaklaşımlar
uygulanmaktadır. Bunlardan fizikokimyasal yöntemler çeşitli spektroskopik
ölçümlerden ibarettir. Bunlar hem biyomembran, hem model membranlarla
gerçekleştirilmektedir. Moleküler biyolojik yöntemler genelde biyomembranların ve
membran kanal proteinlerinin cDNA dizilişini ortaya çıkararak voltage-gated iyon
kanal ölçümleri veya tek kanal üzerinden geçişi takip eden patch-clamp tekniği gibi
yöntemlerin de yardımıyla membran gözeneklerinden gerçekleşen çeşitli iyon
geçişlerini hücre kültürleri üzerinde göstererek ortaya koyan yaklaşımlara dayanır.
Biyokimyasal yaklaşımlar da buna benzer şekilde membran yapısında yer alan ve
araştırılması istenilen makromoleküllerin saflaştırılıp tekrar yapay membran
modeline rekonstitüe edilmesine dayanır. Böyle bir in vitro rekonstitüsyon yöntemi
biyolojik sistemin in vitro ortamlarda da çalıştırıldığı takdirde incelenen biyosistemin
gerçek etki mekanizmasını ortaya çıkarmaktadır. Örneğin, birçok reseptör-ligand
bağlanma kinetiğinin ortaya çıkarılması, söz konusu reseptörlerin biyolojik hücreden
saflaştırılıp yapay membranlara rekonstitüe edilip ligand bağlanma afinitelerini
takibine dayanır. Yapay membran sistemleri genelde üç gruba ayrılır. Bunlardan
biyomembranlara ilk yaklaşım olarak lipid monolayer (Langmuir-Blodgett film-bk.:
Şekil 2.13-üst kısım) düzeneği geliştirilmiştir. Bu iki boyutlu membran yaklaşımı
birçok biyolojik ve farmakolojik etkisi olan moleküllerin model hücre üzerinden
intraselüler penetrasyon mekanizması incelenmektedir. Diğer bir iki boyutlu model
sistemi ise Black Lipid Membrane (BLM) sistemidir. Bu düzenekte dikey Pt veya
teflon plâkaları arasında bulunan boşlukta çift katman oluşturulmaktadır (Bk.: Şekil
2.13-orta kısım ve özellikle Şekil 2.14). Bu yöntem tarihsel olarak büyük önem
taşımıştır. Örneğin, ilk membran elektrofizyolojik olayları bu yaklaşımla ortaya
çıkarılmıştır. Ancak, bu yöntem çok karmaşık mikroskopik incelemelerini de
gerektirdiğinden, son derece deneyimli personelin çalışmasına bağlıdır. Üçüncü
yaklaşım ise lipid veziküllerin (lipozomların) kullanımına dayanır (Bk.: Şekil 2.13 alt
45 Bu bölümde projemi inceleyecek olanlara sadece çok kısa ön bilgilerin verilmesi amaçlanmıştır. Biyomembran düzenekleri ile daha ayrıntılı bilgi edinmek için Moleküler Hücre Biyolojisi, Hücre Fizyolojisi ve Biyofizik ders kitaplarına başvurulmalıdır.

180
kısım ve Şekil 2.15)46. Bu veziküller bir üç boyut kazandırdığından, yine üç boyutlu
olan doğal hücreye diğer iki model membran sisteminden çok daha iyi bir yaklaşım
teşkil etmektedir. Biyomembran incelemelerine geçmeden önce, araştırılmakta olan
makromoleküllerin ilk önce saflaştırılmış şekillerinin fizikokimyasal özelliklerinin
takip edilmesi gerekmektedir. Bu da çeşitli fizikokimyasal ve biyofiziksel
yaklaşımlarının önemini vurgulamaktadır. Bunlara aşağıda kısaca değinilmiştir.
2.3.1. Biyofiziksel Yöntemler
2.3.1.1. Spektroskopik Yöntemler
Aşağıda adı geçen spektroskopik yöntemler biyomembran araştırmalarda
membran bileşenlerinin yapı ve aktiviteleri ile ilgili bilgilerin elde edilmesinde çok
önemli rol oynamıştır. Bu yöntemler hızlı ve invaziv veya destrüktiv değildir. Bu
bakımdan bunların tümü fizyolojik ortamlara yaklaşan ve bu sistemleri simüle eden
koşullarda uygulanabilmektedir. Ayrıca, bunlarla membranlarda meydana gelen çok
hızlı tepkimelerin ve konformasyon değişikliklerinin kinetiğini de incelemek
mümkündür.
46 Lipozomlarla ilgili çok ayrıntılı ve güncel bilgilere Ek-9’da yer verilmiştir.

181
Şekil 2.13. Kolloid ve yüzey kimyasal araştırmalarda bir hücre yüzeyini
modellemek için kullanılan düzenekler. Sırasıyla Langmuir-Blodgett tek
katman (monolayer), Black Lipid Films (BLMs) ve lipozom sistmleri
gösterlmiştir.

182
Şekil 2.14. Biyomembran araştırmalarda bir hücre yüzeyini modellemek için
kullanılan düzenekler. Çeşitli tek katmanlı ve çok katmanlı fosfolipid
düzenekleri gösterilmiştir. Burada özellikle farklı BLM formasyonları
vurgulanmıştır.

183
Şekil 2.15. Lipozomların genel yapısı.
2.3.1.2. Difference Spectroscopy
Bazı durumlarda belli bir membran kromoforun ve çevresindeki
moleküllerin absorbsyon spektrumunda meydana gelen spektral değişikliklerinin söz
konusu olduğunda kromoforun tek başına gösterdiği konformasyon değişikliğinden
ziyade, tüm toplam spektrumdaki değişikliklerin ölçülmesi daha uygundur. Bu işlem
çift-ışınlı spektrofotometre ile yapılmaktadır. Bu yöntemle meydana gelen
tepkimelerin kinetiğinin ölçülmesi elektron taşıyıcılarının indirgenme-oksitlenme
olaylarına bağlı olduğu gibi, NMR, ESR ve termodinamik yöntemlerinden farklı
olarak moleküllerin konformasyonu hakkında çok kısıtlı bilgiler vermektedir. Ancak,
bu yöntemi kullanarak makromolekül-ligand bağlanma afinitesi ve kinetiği hakkında
çok önemli bilgiler edinebilinir.
2.3.1.3. Fourier Transform Kızılötesi (FTIR) Spektroskopisi
Bir titreşim spektroskopisi olan FTIR spektrometresi kompleks
oluşumu esnasında tepkimeye giren biyomakromoleküllerin sekonder yapıları
hakkında çok önemli bilgiler vermektedir. Tepkimeye giren fonksiyonel gruplar

184
hakkında çok önemli bilgiler de vermektedir. Nuclear Magnetic Resonance (NMR),
Electron Spin Resonance (ESR) veya Floresans Spektroskopisi vb. gibi yöntemlerin
aksine çok fazla örneğe gereksinim duyulmadığı bu ölçüm yöntemi ayrıca da çok
hacimli etiket moleküllerinin de katılmasını gerektirmemektedir ve “scattering”’ten
kaynaklanan artefaktlar da burada engellenmektedir. Ayrıca, FTIR spektrometresi
katı ve sıvı fazda bulunan örneklerle çalışma fırsatı da sunmaktadır.
2.3.1.4. Raman Spektroskopisi
Bir titreşim spektroskopik yaklaşımı olarak bu spektroskopik yöntem
moleküllerin iradyasyonunu takiben gözlenen moleküler titreşimlerin ölçülmesine
dayanmaktadır. Bu yöntem, örneğin kolesterolün fosfolipid jel-sıvı kristal faz geçişi
üzerindeki etkisinin araştırılmalarında uygulanmaktadır. Raman spektroskopisi
ayrıca Fe-S taşıyan proteinlerde (örneğin transferinde) Fe-S karşılıklı etkileşmeleri
veya hem proteinlerdeki porfirin-protein etkileşmeleri, veya çeşitli organik
moleküllerinde (örneğin, karotenoidler, vitaminler, antibiyotikler, steroidler ve
polisakkaridler) C-C titreşimleri ile ilgili çok önemli bilgiler vermiştir.
2.4. Langmuir-Blodgett Monolayer Yöntemi
Monomoleküler katmanların (tek katmanların veya monolayer’ların)
incelenmesi, membranda yer alan bileşenlerin hakkında çok önemli yapı-aktivite
bilgilerini verdiği gibi, monolayer yüzeysel basıncı, incelenmekte olan molekülün
lipid tek katmanda işgal ettiği moleküler alanı, bu moleküler alanın yüzeysel basınç
üzerindeki etkisi hakkında da bilgiler vermektedir. Bu düzenek böylece molekülün
biyomembran yüzeyinde ve hacminde nasıl paketlendiği konusunda da bilgiler
vermektedir. Bu yaklaşım böylece, yeni farmakolojik ajan-biyomembran
etkileşmelerinin yeni modellerinin de geliştirilmesinde katkıda bulunmaktadır.
Membran ortamında başka maddelerle karışım içinde bulunan bileşiklerin
karışımdaki tek tek özelliklerini, stereokimyasını ve etkileşme enerjilerini, suda
çözünen bileşiklerin lipidlere karşı gösterdikleri afinitelerini de ortaya çıkarmak
mümkündür. Monolayer yöntemi çok katmanlı oriente edilmiş (oriented multilayers)
sistemlerinin ve içeriği ve topolojisi önceden belirlenen membran sistemlerinin
oluşumunun temelini oluşturmaktadır. Monolayer yöntemini ayrıca daha pratik
amaçlarla, örneğin ortam saflılığının tayininde ve kontrolünde, ekolojik
uygulamalarda ve son yıllarda kemosensör ve biyosensör teknolojisinde

185
mikroçiplerin orientasyonu ve toplaşımı için mikroelektronik sanayide de
kullanılmaktadır.
Gaz-sıvı sınırında oluşan yüzeysel basınç veya pozitiv serbest enerji yüzeyler
arasında bu sınırın oluşturulmasının temelini teşkil etmektedir. Bu gibi yüzey
sınırlarının yüzeysel basıncı γ; γ=∆F/∆S olarak ifade edilir. Burada ∆F-serbest
enerjinin değişimi, ∆S ise – yüzey alanının değişimidir.
Yüzeyler arasında oluşan serbest enerjinin ortaya çıkmasını sıvı moleküllerinin
hacimsel olarak (sub-phase) fazlar arasındaki sınıra taşınmaları için yapılan işe
benzetilebilir. Bu taşınımın gereksinimi, ara fazda (subfazda) belli bir molekül
üzerinde diğer moleküller tarafından uygulanan ve etkili olan güçlerin kompanse
edilmeleri, yüzeyler sınırında bulunan bir moleküle ise sadece subfaz tarafından
çekim güçlerinin etkili olduğundan ortaya çıkmaktadır.
Yüzeyler arasındaki kompanse olmayan güçlerin katmandaki derinlikleri su
moleküllerin etkileşme özelliklerine göre tayin edilir. Çeşitli iyonların yokluğunda,
söz konusu güçler Van der Waals etkileşme güçleridir ve bunların intansitesi
moleküller arasında mesafenin x7 oranında propoprsiyonel olarak azalır. Böylece,
kompanse edilmeyen etkileşmelerin alanı veya bölgesi birkaç molekül çapından fazla
değildir.
Gibbs denklemine göre, faz ayrımındaki adsorpsyonu için yüzeysel dansite
parametresi şu şekilde ifade edilir:
Γ=-[a/(RT)].(δγ/δa).
Burada Γ – maddenin yüzeysel dansitesi, a –subfazda maddenin aktivitesidir.
Eğer maddenin yüzeyler arasında birikirse, o zaman δγ/δa negativtir, çünkü
örneğin lipidler için yüzey arasındaki adsorpsyon ölçülebilen yüzeysel basıncın
düşmesine neden olmaktadır. Bu durumda γ0-γ farkı (burada γ0-“saf” su yüzeyinde
oluşan yüzeysel basınçtır; γ-örneğin lipid için yüzeyler arasında adsorbsyonundan

186
sonra oluşan yüzeysel basınçtır) adsorbe edilmiş plâkanın yüzeysel basıncı olarak
tayin edilir.
Faz sınırlarında oluşan tek katmanın yüzeyinde ortaya çıkan basıncın yüzeysel
basınca eşit olduğu bir plâka olarak da tasavvur edilebilinir:
YB= γSu-γçözelti
Burada YB-yüzeysel basınç; γSu ve γçözelti – suyun ve çözeltinin yüzeysel
gerilimleridir.
2.4.1. Yüzeysel Gerilimin Wilhelmy Plate Yöntemiyle Ölçülmesi
Yüzeysel gerilimin ölçümlerinde su veya sıvı çözeltilerinin yüzeyine dik
dörtgen şeklinde küçük bir plâka temas ettirilmektedir. Bu plâka, organik çözeltilerin
önceden belli bir muameleyle uzaklaştırıldığı plâtin malzemesinden üretilir. Ancak,
bunun yanı sıra, cam ve diğer malzemeler de ölçüm amacına göre kullanılmaktadır.
Faz ayrımının sınırında bu plâkaya yüzeysel gerilim kuvvetleri etkilidir (Şekil 2.16).
Şekil 2.16. Langmuir-Blodgett tek katmanlarda Wihelmy plate ile ölçülen yüzey
basınç yönteminin bir görünümü. Bu şematik çizim daha önce verilen Şekil
2.10.’a dayanmaktadır.

187
Bu kuvvetler sayesinde plâkanın sıvı yüzeyine teması sağlanmaktadır ve bu
kuvvetin şiddetini mekanik kuvveti elektrik sinyaline dönüştürebilen ve mekanotron
adı verilen bir cihazla ölçülmektedir (Şekil 2.19). Plâkanın ölçülen kütlesi şu şekilde
tayin edilir:
wtoplam = wplâka+γ.cosθp,
Burada γ-çözeltinin yüzeysel gerilimi; p-plâkanın boyutu; wplâka – plâkanın
gerçek kütlesi.
Yüzeyler sınırında çok az miktarda maddenin uygulandığında, γ çözelti veya su
subfazının yüzeysel gerilimine eşittir. Yüzeyler arası ayrımının alanının azaldığında,
wtoplam düşmektedir.
2.4.2. Lipid Tek Katmanların Özelliklerinin İzoterm Yöntemiyle
İncelenmesi
2.4.3. Sub-fazda madde-lipid etkileşmelerinin incelenmesi ve lipid
moleküllerinin işgal ettikleri alanın ölçülmesi
Tek katmanın en önemli parametresi lipidin moleküler alanıdır (A.). Diğer bir
değişle bu bir molekülün işgal ettiği alandır. Kondanze olan katmanlardaki A değerin
tayini çift koordinatlı potansiyometre ile ölçülen yüzeysel basınç – tek katman alanı
ilişkisini x-eksenine yerleştirilmesiyle ekstrapolasyonuna dayanmaktadır. Böylece,
monolayer yöntemi ile bir tek katmanda lipidin işgal ettiği alanı ve kesitini
hesaplamak mümkündür. Bu çok önemli bir bilgidir, çünkü doğrudan doğruya doğal
biyolojik hücre membranında çeşitli makromoleküllerinin nasıl paketlendiğini
göstermektedir.
Fosfolipidler için söz konusu kesit lipidin baş kısmının türü ve içerdiği doymuş
veya doymamış yağ asitlerin oranına bağlı olarak değişmektedir. Tek katmanın

188
bileşenleri arasındaki etkileşmelerinin spesifikliği ve ayrıca da moleküllerin hidrofil
kısmı ile subfazın A değerini etkilemektedir. İyonlar ve subfaz moleküllerinin suya
etki ettikleri ve sınır bölgesindeki katmanda lipid baş gruplarının paketlenmelerini de
etkiledikleri bilinmektedir.
2.4.4. Tek Katmanda Yer Alan Maddeler Arasındaki Etkileşmelerinin
İncelenmesi
Bu yaklaşımda, tek katmanın oluşturulmasında kullanılan ve incelenmekte
olan maddeden çeşitli oranlarda hazırlanan madde-lipid örnekleriyle seri şeklinde
birkaç izoterm ölçümü yapılır. Elde edilen neticelerden, karışımda yer alan A-
değerinin konsantrasyonu Mol (M) olarak ifade edilen madde içeriği arasındaki ilişki
diyagram çizilir. Bu diyagramın bir örneği Şekil 2.17 gösterilmiştir. Bu diyagramlar
kolesterol ve diheksadesilfosfatidilkolin ile oluşturulan tek katmandaki kompleks
oluşturmaları hakkında bilgiler içermektedir. Ancak bu şekildeki eğriler başka
biyolojik veya farmakolojik aktiviteye sahip bir makromolekülün fosfolipid
membranla oluşturduğu bir kompleks için de geçerlidir.

189
Şekil 2.17. Polikremnik asidin varlığında diheksadesilfosfatifilkolin ile kolesterol
arasında etkileşmeler: I-Kolesterol-diheksadesilfosfatifilkolin tek katmanın izoterm’i:
1-kolesterol; 10-diheksadesilfosfatifilkolin; 5-kolesterol-diheksadesilfosfatifilkolin 1:1
oranında karışımı; 8-kolesterol-diheksadesilfosfatifilkolin 2:8 oranında karışımları.
Diğer oranda hazırlanan karışımlar arada izoterm eğirleri olarak gösterilmiştir. II.
Tek katman içeriğine bağlı olarak karışımlarının tek katmandaki moleküler alanın
değişmesi: 1-SiO2 grupların varlığına dayanarak 5 mM konsantrasyonundaki
polikremnik asidin varlığında; 2-polikremnik asidin yokluğunda; 3 ve 4-karışım
olarak oluşturulan tek katmanların aditiv moleküler alanlar (additive molecular
areas).
1:1 ve 4:1 stokiometriye sahip kompleksler adı geçen maddelerin varlığında
meydana gelir, ancak subfaz’a polikremnik asidin katıldığında sadece 1:1
stokiometriye sahip kompleksler oluşur.
2.4.5. Tek Katmanlarda Maddelerin Karışım Oluşturma Özelliklerinin
İncelenmesi
Bir tek katmanda iki bileşiğin karışım oluşturma özelliklerinin tayininde faz
kuralı (the phase rule) uygulanmaktadır. Basıncın ve sıcaklığın veya ortam ısısının
sabit olması koşuluyla izotermler çizilir. Ancak, bu yöntem daha çok malzeme
bilimcileri tarafından uygulandığından ve biyolojik önemi pek olmadığından, bu

190
yaklaşım projemizde öngörülmemiştir. Söz konusu yaklaşım, çözünürlükleri
hakkında bilgi bulunmayan iki ayrı maddenin biyolojik ortamda tek katmandaki
karışım oluşturma özelliklerinin tayininde uygulanmaktadır. Birçok düzenekte
olduğu gibi, projemde hem Taxol®’un, hem de kullandığımız fosfolipidlerin karışım
oranları önceden tasarlanmıştır.
2.4.6. Tek Katmanlarda Bulunan Moleküllerin Yapısal Değişikliklerinin
İncelenmesi
Lipid tek katmanlarının izoterm eğrilerinde yüzey sınırında yer alan
moleküllerinin oriyentasyonlarındaki değişikliklerinin göstergesi olan yapısal
değişikliklerinin de görmek mümkündür. Uzun hidrokarbon zincirin ucunda iki
hidrofil grubu içeren moleküller için örneğin floresan işareti olarak yoğun kullnılan
16 (9-antroiloksi)-palmitik asidin durumunda olduğu gibi, bu lipid ile oluşturulan tek
katmanın kondanze olmasıyla, ölçülen izoterm eğrisinin türevinin (slope) de değiştiği
bilinmektedir. Söz konusu değişiklik, oluşan yüzey sınırında bu işaretin hidrofil
gruplarından biri olan COOH grubunun yer değiştirmesi ve tüm molekülün tek
katmandaki konformasyonunun hareketliliği ile açıklanmaktadır. Yüzey basıncın
düşük olduğu durumlarda, bu molekül yüzey sınırında su ile en uçtaki COOH-
grubunun ve antranil döngüsüne komşu olan CO-grubunun temas yapacak şekilde
yerleşir. Bu yaklaşım, özellikle Langmuir-Blodgett tek katman ve floresan
mikroskopik araştırmalarında yoğun olarak uygulanmaktadır (Şekil 2.18).

191
Şekil 2.18. Çeşitli lipidlerin tipik π-A izoterm eğrileri.
2.4.7. Tek Katmanın Sabitlilik ve Deformasyon Özelliklerinin
Hesaplanması
Tek katmanların lateral kompaktizasyonu parametresi c=-(1/A)(δA/δπ)T
olarak ifade edilir. Söz konusu hesaplar için bu sabitin resiprokal veya tek katmanın
Gibbs deformasyonu da kullanılır:
E=-A(δπ/δA)T.
2.4.8. İzoterm Şekillerinin Analizi ve Yorumlanması
Farklı lipidlerin tek katmanları için ortam ısısını belli sınırlar içinde değiştiği
koşullarda elde edilen izotermlerde bir plato’nun oluştuğu görülmektedir: Alan (A)
değerinin azalmasıyla yüzeysel basınç değeri hafifçe yükselmektedir. Elde edilen
izoterm eğrilerden biyolojik anlamı çok yüksek olan, bir molekülün modellenen bir

192
hücre yüzeyinde nasıl paketlendiğini gösterir. Bu da, ilâç-hücre etkileşmelerin
araştırmalarında çok önemli bilgiler vermektedir.
Şekil 2.19. Bir Langmuir-Blodgett tek katman cihazının şematik şeması. Bu cihazlarla
ve çalışma prensibiyle ilgili olarak daha detaylı bilgi için bk.: www.langmuir.com veya
www.nima.co.uk. Ayrıca bk.: Ek-12 ve Ek-13.
2.4.9. Projemdeki Deneysel Tasarım
2.4.10. Projemde Geliştirilen Yeni Deneysel Protokol
2.4.10.1. Kimyasallar.
Tüm fosfolipidler Sigma Chem. Co., MI (A. B. D.) veya alt birimlerinden veya
Avanti Polar Lipids (Alabaster, AL, A. B. D.)’den satın alınmıştır. Özellikleri çok
farklılık gösterdiğinden, bu özellikleri test etmek amacıyla Paclitaxel™ çeşitli
kaynaklardan alınmıştır47.
Tüm kimyasallar en yüksek kalitede ve saflıkta alınmıştır. Kloroform (≥99.8%;
HPLC grade) ve etanol (≥99.9%; HPLC grade) kullanılmıştır. Tüm deneylerde
kullanıma hazır olarak satılan ve rezistansı 18.2 MΩ olan Milli-Q saf su kullanılmıştır.
47 Bk.: Ek-18

193
2.4.10.2. Langmuir-Blodgett Tek Katmanın Oluşturulması.
Langmuir-Blodgett cihazı NIMA Technology Ltd (The Science Park, Coventry
CV4 7EZ, İngiltere; Model 112D) ürünüdür48. Bu cihazı üreten diğer firmalara49
nazaran NIMA Technology Ltd.’in piyasaya sürdüğü son derece popüler ve çok amaçlı
modeller50 ve özellikle de bu firma temsilcilerinin tarafımıza yapmış olduğu çok
düşük fiyat teklifleri bu firma ürününün alınmasında rol oynamıştır. Söz konusu
cihazın teknik özellikleri Ek-12‘de verilmiştir. Bu cihaza diferansiyel taramalı
mikrokalorimetre (DSC), Fourier Transform Infrared (FTIR) Spektpmetresi ve
Floresan Mikroskopi sistemlerinin de uygulanıyor olabilmesi son derece önemlidir ve
çok yönlü ölçümlerin yapılmasını mümkün kılmaktadır. Söz konusu cihaz 40 cm2
esas alanı, 2 elektromekanik olarak hareket ettirilen bariyer, yüzey basınç sensörü,
inverted mikroskopisi için sapphire pencere sistemi, dipper mekanizması ve
bilgisayara bağlı software versyon 5.16 içermektedir.
Tek katmanların yüzeysel temizliliği kriteri olarak bariyer testi uygulayarak 3
ayrı tekrardan elde edilen izotermler arasındaki farkın ±0.1 mN/m’den daha fazla
olmamasına dayanmaktadır.
2.4.10.3. Yüzey basınç-alan (π-A) izotermler.
Tek katmanlar air-water (hava-su ara sınır) ara yüzeye kloroformu çözücü
olarak kullanarak uygulanmıştır. Fosfolipidler ve Paclitaxel™ stock çözeltileri ilk önce
ekvimolar (1:1) 0.2 mM’ lık konsantrasyonlarda hazırlanmıştır. Daha sonra,
bunlardan istenilen oranlarda karışımlar hazırlanmıştır. Her uygulamadan önce
Hamilton şırıngası 3 kez kloroform ile yıkanmıştır. Bunu takiben ara yüzeye birkaç
damla örnek uygulanmıştır. 10 dakika solvent evaporasyonu için beklendikten sonra,
yüzey basınç-alan (π-A) izotermleri elde edilmiştir ve kaydedilmiştir. Neticelerin
tekrarlanabilirliği bakımından güvenilir olup olmadıkları en az 3 tekrar yaparak
değerlendirilmiştir.
48 Bk.: Ek-11 ve www.nima.co.uk. 49 Kibron Ltd (www.kibron.com) ve KSV Ltd. (www.ksvltd.com)-Helsinki, Finlandiya. 50 Bk.: Ek-11.

194
2.4.10.4. Diferansiyel Taramalı Kalorimetrik (Differential Scanning
Calorimetry) Ölçümleri.
Lipid vezikülleri ilk önce geleneksel ince tabaka film (thin-film) yöntemine
göre hazırlanmıştır. Bunun için, lipid ve Paclitaxel stock çözeltileri kloroform ile
hazırlanıp, bunlardan istenilen miktarlar balon jojelere aktarılmıştır. İyi bir karışım
elde etmek için bunlar vortekslenip, gaz N2 ile solvent evaporasyonu uygulanmıştır.
Böylece balon jojenin iç cam yüzeyine ince tabaka lipid film oluşturulmuştur. Bu film
gece boyunca vakum desikatör ile muamele edilmiştir ve böylece son kloroform
kalıntı miktarları da uzaklaştırılmıştır. 1ml saf su veya adı geçen tampon ile balon
jojeye aktarılıp 1 dakika boyunca vortekslenmiştir. Balon joje daha sonra söz konusu
lipidin faz değişim ısı noktasından daha yüksek bir sıcaklığa ayarlanan sonikatörde
tutulmuştur. 5-10 dakikalık bir süreyle sonikasyon uygulanmıştır. Daha sonra bu
balon joje shaking su banyosuna 37°C’de 1 saat boyunca bırakılmış ve böylece iyi bir
ilâç-lipid karışımı hazırlanmıştır. DSC ölçümleri Perkin-Elmer Pyris Marka DSC
cihazı ile Orta Doğu Teknik Üniversitesi Merkez Laboratuvarında yapılmıştır.
Referans olarak hermetik kapanabilinen boş alüminyum kaplar kullanılmıştır. Lipid
konsantrasyonu olarak 20 mg/ml kullanılmıştır ve bundan 10 µl lipozom
süspansyonu hermetik alüminyum kaplara konulmuştur ve bunlar böylece
kapatılmıştır. Termogramlar gösterilen sıcaklık aralığında elde edilmiştir.
2.4.10.5. Fourier Transform Infrared (FTIR) Spektrometresi.
Bu ölçümler Orta Doğu Teknik Üniversitesi Merkez Laboratuvarında
yapılmıştır. Bu ölçümlerde tüm spektrum çekimleri ve direkt ölçüm yöntemi
uygulayarak gerçekleştirilmiştir.
Esas Langmuir-Blodgett tek katmanla gerçekleştirilen yüzey basınç
ölçümlerine geçilmeden önce FTIR ve DSC ölçümleri yapılmıştır. Bunun için
uluslararası standartlara uyulmuştur. Bu yaklaşımlar uyarınca ilk önce bir
formülasyonunun ön fizikokimyasal özellikleri iyi bir şekilde incelenmesi
gerekmektedir. Bununla ilgili deneysel tasarımlar aşağıdaki Tablo-2.2’de verilmiştir.
Projemde de bu hususlar aynen uygulanmıştır. Bu doğrultuda, ilk önce Paclitaxel™-
fosfolipid kompleks oluşumunun ön termal ve spektrometrik incelenmesi yapılıp tek
katman ölçümlere geçilmiştir.

195
Tablo-2.2. Ön formülasyonlarda öngörülen fizikokimyasal incelemeler*,51.
*(Kaynak: Pharmaceutical Preformulation and Formulation. A Practical Guide from Candidate Drug Selection
to Commercial Dosage Form. Mark Gibson (Ed.), Interpharm/CRC, Boca Raton Fl, 2004, pp. 23).
51 Önformülasyonlar hakkında özellikle bk.: Pharmaceutical Preformulation and Formulation. A Practical
Guide from Candidate Drug Selection to Commercial Dosage Form. Mark Gibson (Ed.), Interpharm/CRC, Boca Raton Fl, 2004.

196
3. BULGULAR

197
3.1. FTIR Ölçümleri

198
Şekil 3.1. L-α-Fosfatidilkolin’in toplam kızılötesi (IR) spektrumu.
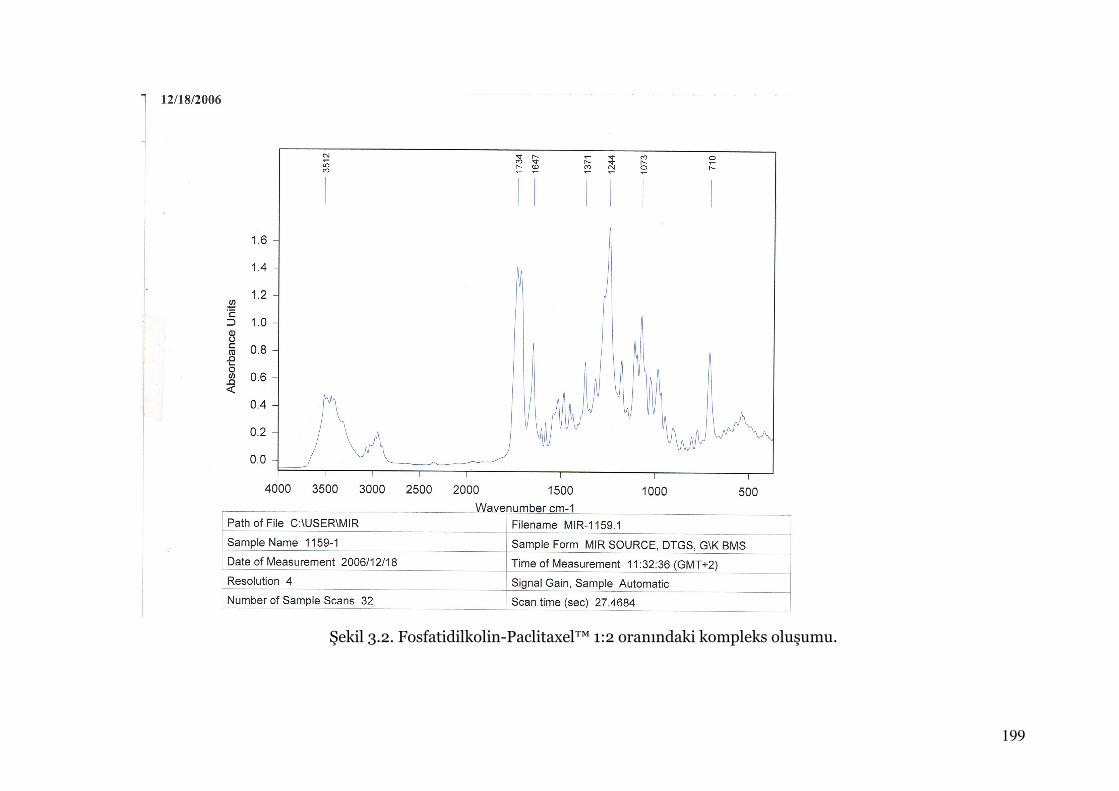
199
Şekil 3.2. Fosfatidilkolin-Paclitaxel™ 1:2 oranındaki kompleks oluşumu.

200
Şekil 3.3. Fosfatidilkolin-Paclitaxel™ 1:3 oranındaki kompleks oluşumu.

201
Şekil 3.4. Fosfatidilkolin-Paclitaxel™ 1:5 oranındaki kompleks oluşumu.

202
Şekil 3.5. Fosfatidilkolin-Paclitaxel™ 1:7 oranındaki kompleks oluşumu.

203
Şekil 3.6. Fosfatidilkolin-Paclitaxel™ 1:10 oranındaki kompleks oluşumu.
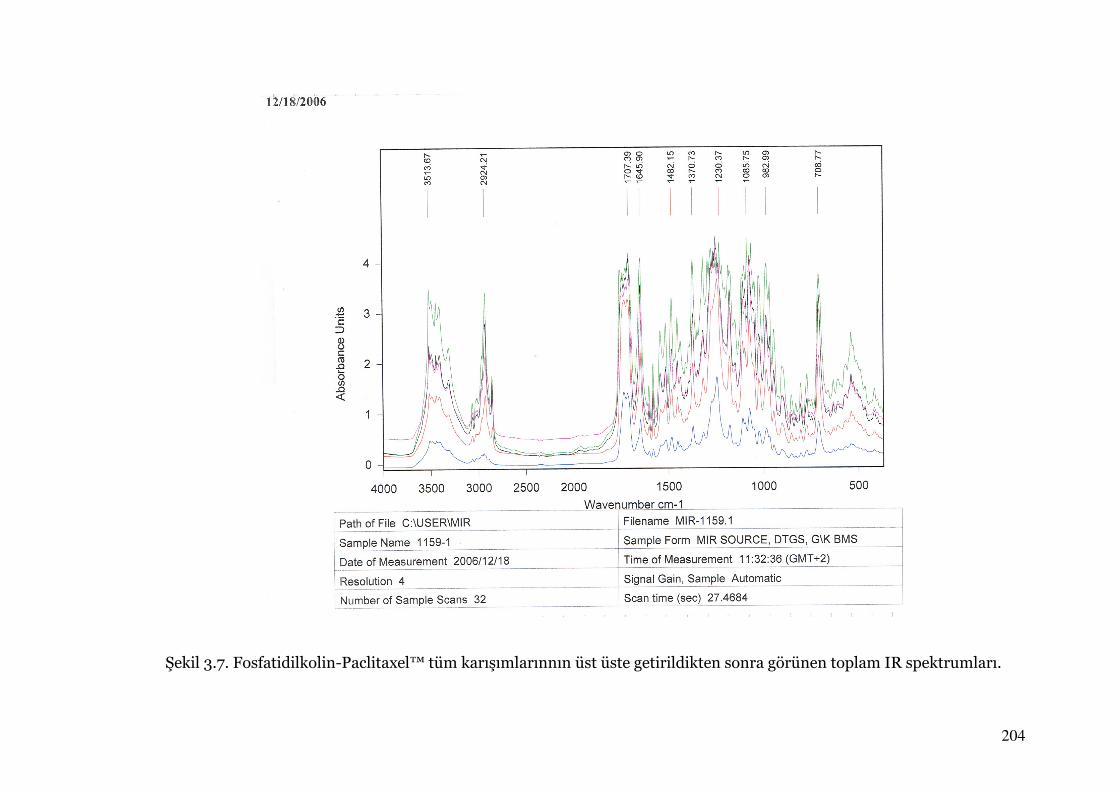
204
Şekil 3.7. Fosfatidilkolin-Paclitaxel™ tüm karışımlarınnın üst üste getirildikten sonra görünen toplam IR spektrumları.

205
Şekil 3.8. Fosfatidilkolin-Paclitaxel™ ekvimolar kompleksi. Toplam IR spektrumu verilmiştir.

206
Şekil 3.9. Fosfatidilkolin-Paclitaxel™ ekvimolar kompleksi. 700-1500 cm-1 IR aralığı verilmiştir.

207
Şekil 3.10. Fosfatidilkolin’in toplam IR spektrumu. Farklı cihaz kullanıldığından Şekil 3.1. ile kıyaslanmalıdır.

208
Şekil 3.11. Fosfatidilkolin-Paclitaxel™ ekvimolar (1:1) karışımı. Üstte fosfatidilkolinin tek başına IR spektrumu verilmiştir.
Taxol® ile 1:1 kompleksi ise aşağıdaki IR spektrumudur.

209
Şekil 3.12. Fosfatidilkolin-Paclitaxel™ ekvimolar (1:1) karışımının toplam IR spektrumu.
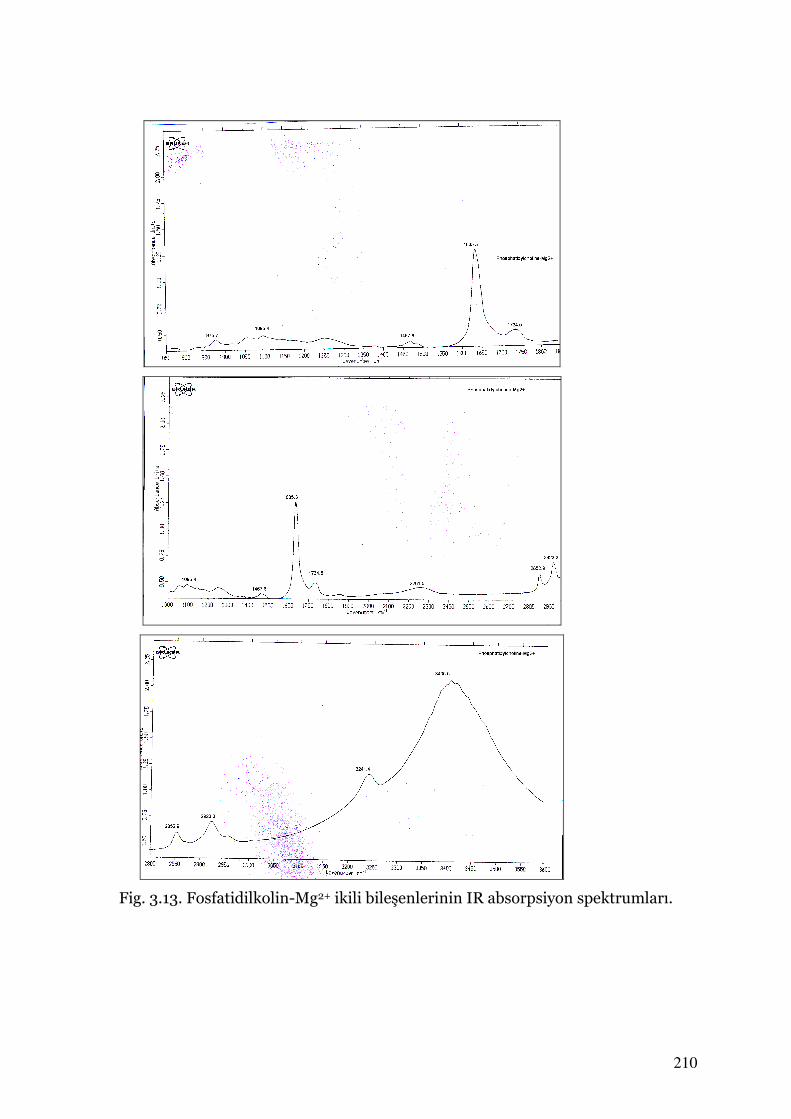
210
Fig. 3.13. Fosfatidilkolin-Mg2+ ikili bileşenlerinin IR absorpsiyon spektrumları.

211
3.2. Diferansiyel Taramalı Kalorimetre (DSC) Ölçümleri

212
Şekil 3.14. Kontrol olarak Fosfatidilkolin’in tek başına çekilen termogramı. Temodinamik parametreler termogram
üzerinde belirtilmiştir.
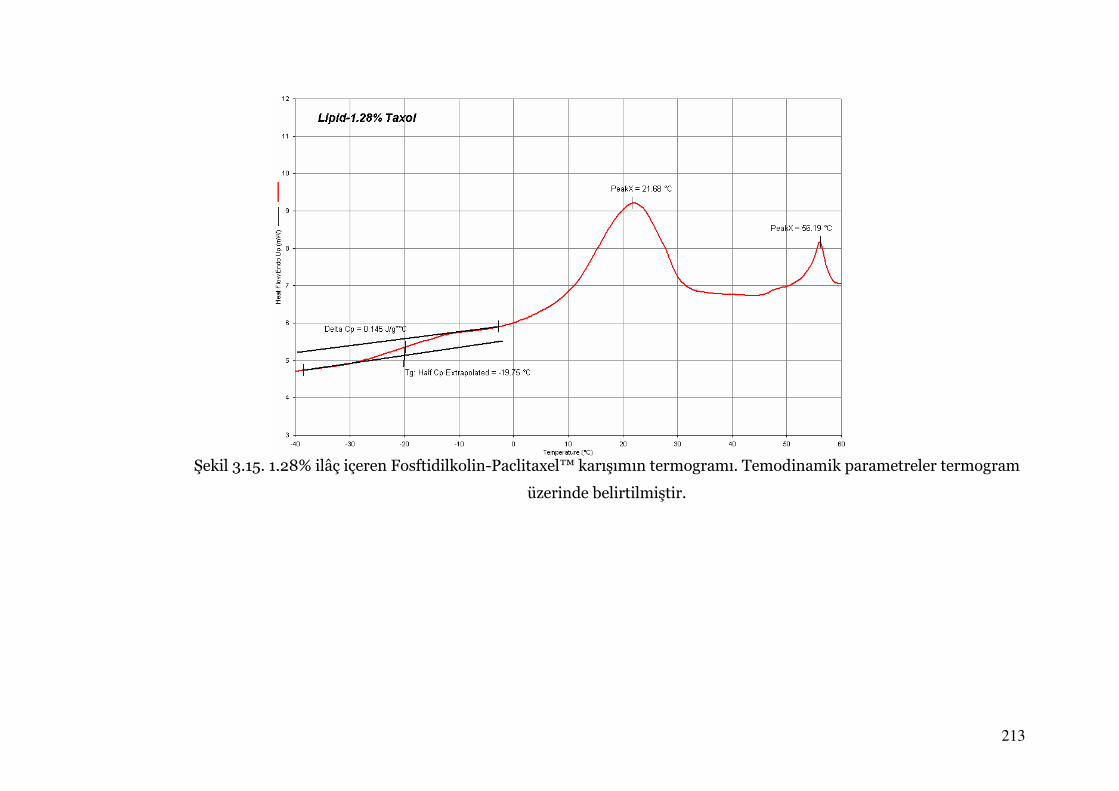
213
Şekil 3.15. 1.28% ilâç içeren Fosftidilkolin-Paclitaxel™ karışımın termogramı. Temodinamik parametreler termogram
üzerinde belirtilmiştir.

214
Şekil 3.16. 10% ilâç içeren Fosftidilkolin-Paclitaxel™ karışımın termogramı. Temodinamik parametreler termogram
üzerinde belirtilmiştir.
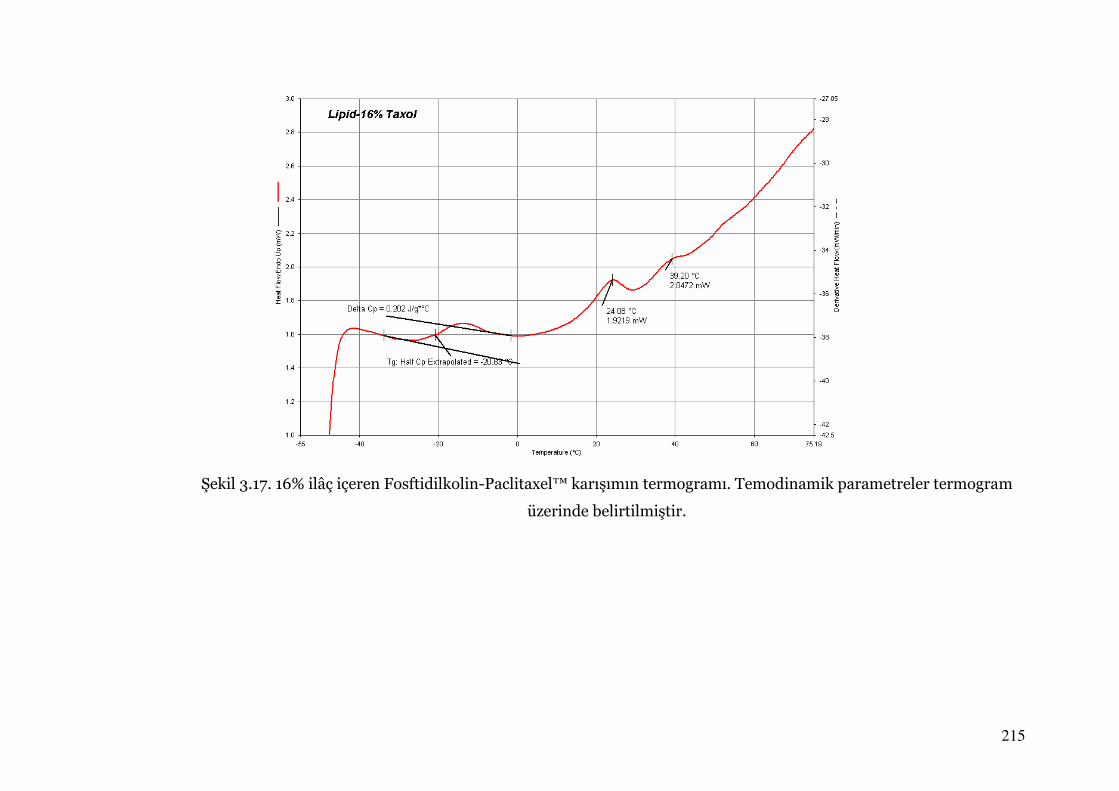
215
Şekil 3.17. 16% ilâç içeren Fosftidilkolin-Paclitaxel™ karışımın termogramı. Temodinamik parametreler termogram
üzerinde belirtilmiştir.

216
Şekil 3.18. 20% ilâç içeren Fosftidilkolin-Paclitaxel™ karışımın termogramı. Temodinamik parametreler termogram
üzerinde belirtilmiştir.

217
Şekil 3.19. Kontrol olarak Paclitaxel™’in termogramı.

163
3.3. Langmuir-Blodgett Tek Katman Ölçümleri
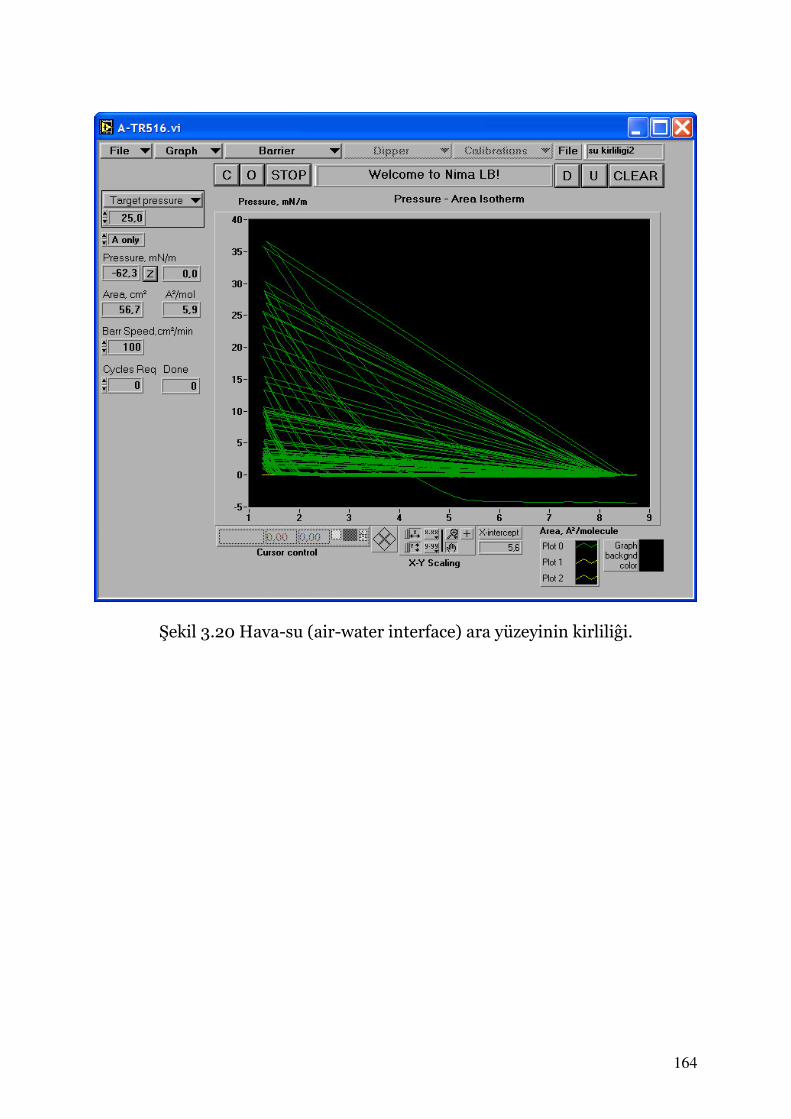
164
Şekil 3.20 Hava-su (air-water interface) ara yüzeyinin kirliliĝi.

165
Şekil 3.21 Yüzey kirliliĝinin giderilmesi.
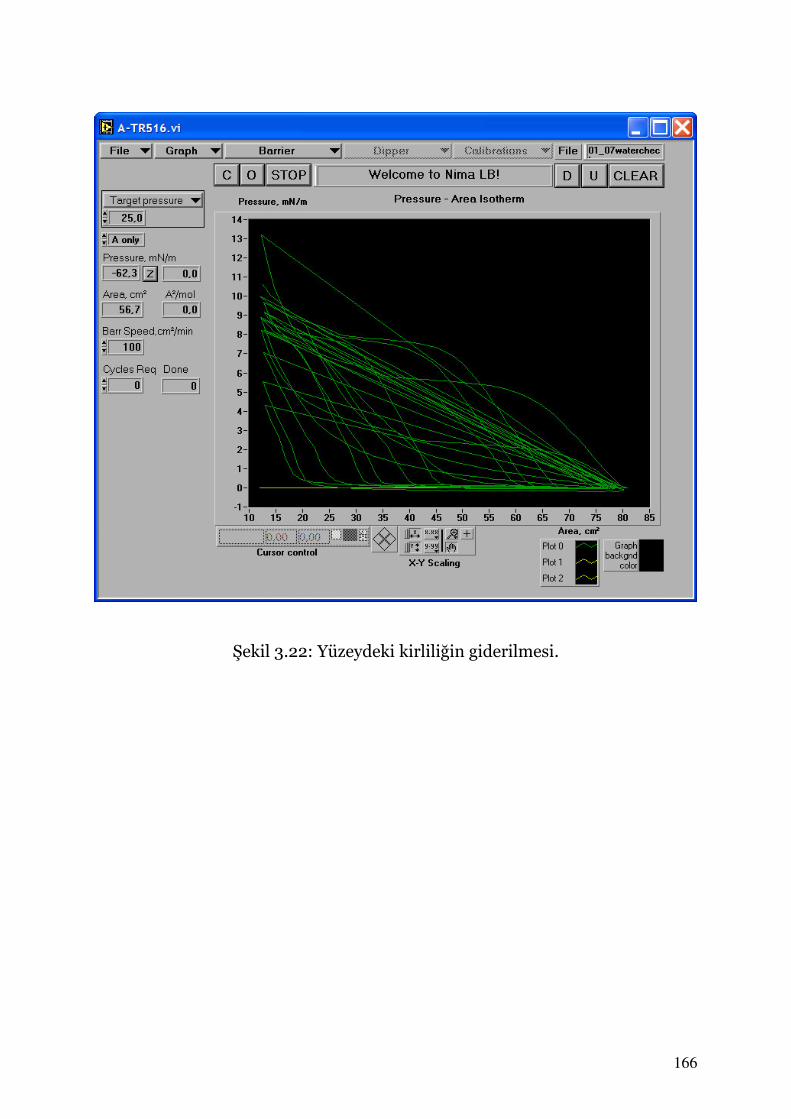
166
Şekil 3.22: Yüzeydeki kirliliğin giderilmesi.

167
Şekil 3.23. Yüzeydeki kirliliğin giderilmesi. Basıncın aşamalı olarak düşürülmesi kirliliğin giderilmesinin işaretidir.

168
Şekil 3.24. Yüzeydeki parçacıklardan kaynaklanan kirliliğin giderilmesi. Basıncın aşamalı olarak düşürülmesi kirliliğin giderilmesinin işaretidir.

169
Şekil 3.25. Yüzeydeki toz ve parçacık kirliliğinin giderilmesi.

170
Şekil 3.26. Yüzeydeki parçacık ve ortam kirliliğinin giderilmesi.
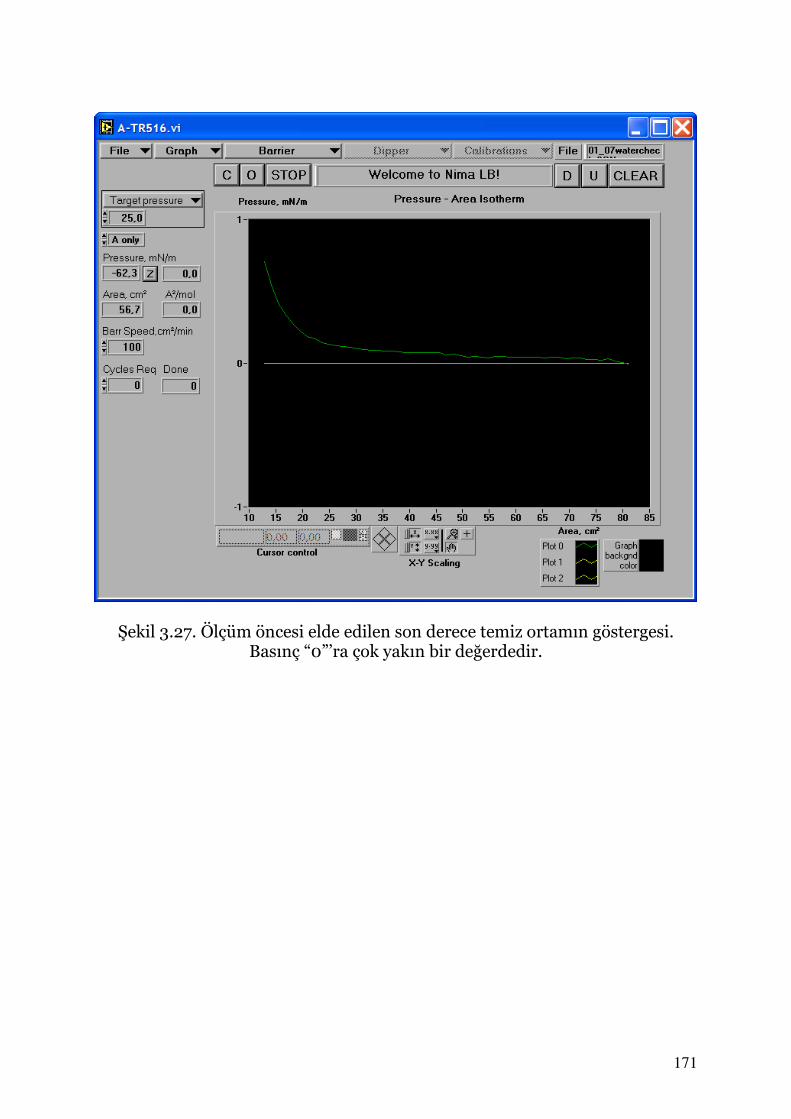
171
Şekil 3.27. Ölçüm öncesi elde edilen son derece temiz ortamın göstergesi. Basınç “0”’ra çok yakın bir değerdedir.

172
Şekil 3.28. 0.2 mM Dipalmitoilfosfatidilkolin (DPPC)’in konsantrasyon profilleri. Konsantrasyonlar yukarıya doĝru iki misli artırılmıştır. Ölçümler
için en yukarıdaki konsantrasyon esas alınmıştır.

173
Şekil 3.29. Kontrol olarak 0.2 mM Taxol®’un tek başına çekilen yüzey basınç-
Alan (π-A) izotermi.

174
Şekil 3.30. DPPC-Taxol®’un 1/5 oranındaki karışımın π-A izotermi. Karışım
yüzey üzerinde oluşturulmuştur.

175
Şekil 3.31. DPPC-Taxol®’un 1/4 oranındaki karışımın π-A izotermi. Karışım
yüzey üzerinde oluşturulmuştur.
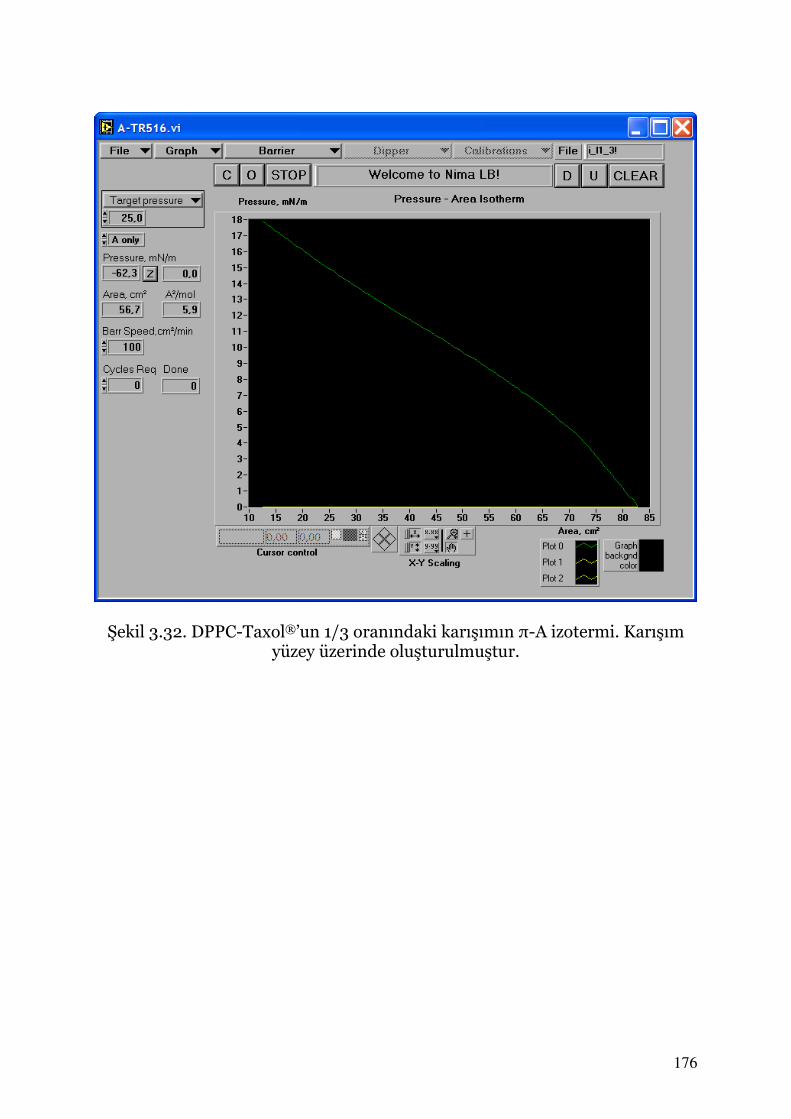
176
Şekil 3.32. DPPC-Taxol®’un 1/3 oranındaki karışımın π-A izotermi. Karışım
yüzey üzerinde oluşturulmuştur.

177
Şekil 3.33. DPPC-Taxol®’un 1/2 oranındaki karışımın π-A izotermi. Karışım
yüzey üzerinde oluşturulmuştur.

178
Şekil 3.34. DPPC- Taxol®’un 1/1 oranındaki karışımın π-A izotermi. Karışım
yüzey üzerinde oluşturulmuştur.
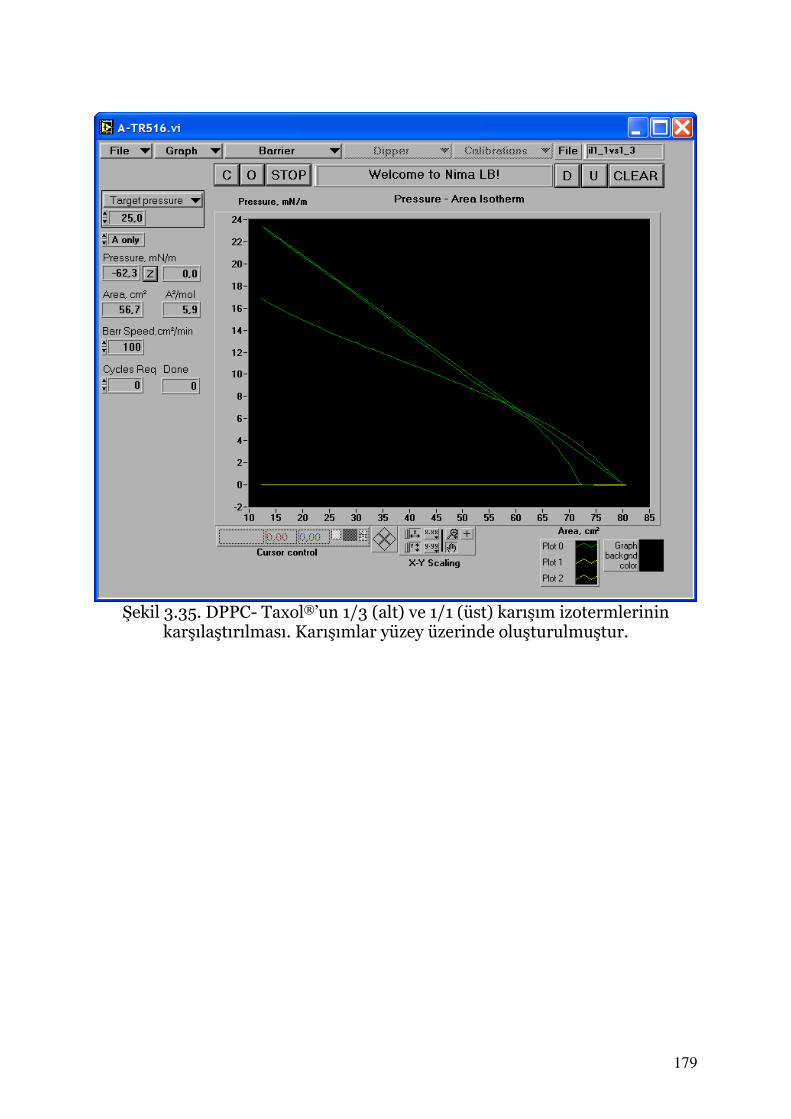
179
Şekil 3.35. DPPC- Taxol®’un 1/3 (alt) ve 1/1 (üst) karışım izotermlerinin
karşılaştırılması. Karışımlar yüzey üzerinde oluşturulmuştur.

180
Şekil 3.36. DPPC- Taxol®’un 1/3 ¼ 1/5. Karışımlar yüzey üzerinde
oluşturulmuştur.

181
Şekil 3.37. DPPC- Taxol®’un ½ karışımın alan başına molekül (Area/molecule)
olarak elde edilen izotermi. Karışımlar yüzey üzerinde oluşturulmuştur.

182
Şekil 3.38. DPPC- Taxol®’un 1/3 karışımın alan başına molekül
(Area/molecule) olarak elde edilen izotermi. Karışımlar yüzey üzerinde oluşturulmuştur.

183
Şekil 3.39. DPPC- Taxol®’un 1/5 karışımın alan başına molekül
(Area/molecule) olarak elde edilen izotermi. Karışımlar yüzey üzerinde oluşturulmuştur.

184
Şekil 3.40. DPPC- Taxol®’un 1/3 vs ¼ karışımın alan başına molekül (Area/molecule) olarak elde edilen izotermlerin karşılaştırılması. Karışımlar
yüzey üzerinde oluşturulmuştur.

185
Şekil 3.41. 1/3 vs ¼ oranında hazırlanan DPPC-Taxol® karışımları. Karışımlar
önceden Eppendorf tüplerde hazırlanarak bu şekilde uygulanmıştır.

186
Şekil 3.42. Taxol®-Dipalmitoilfosfatidilkolin (DPPC) 1/8 oranındaki π-A izotermi, kırılma noktası ve geriye doĝru izotermi.

187
Şekil 3.43. Taxol®-Dipalmitoilfosfatidilkolin (DPPC) 1/4 oranındaki izotermi,
kırılma noktası ve geriye doĝru izotermi.

188
Şekil 3.44. Taxol®-Dipalmitoilfosfatidilkolin (DPPC) 1/2 oranındaki izotermi,
kırılma noktası ve geriye doĝru izotermi.
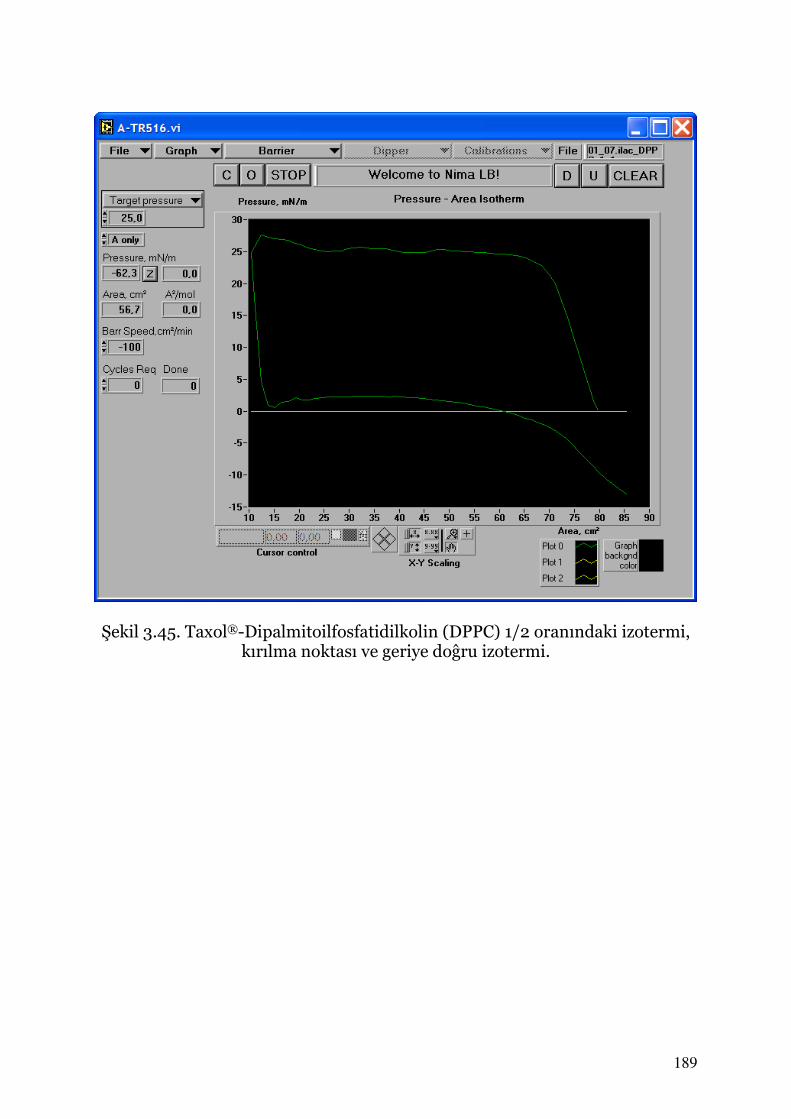
189
Şekil 3.45. Taxol®-Dipalmitoilfosfatidilkolin (DPPC) 1/2 oranındaki izotermi,
kırılma noktası ve geriye doĝru izotermi.

190
Şekil 3.46.Saf yüzey sinyalinin elde edilmesi.

191
Şekil 3.47. Yüzeyin saflık testi. Son derece temiz bir yüzey elde edilişinin kanıtı.

192
Şekil 3.48. 0.2 mM Dipalmitoilfosfatidilkolin (DPPC)’in π-A izotermi.

193
Şekil 3.49. o.2 mM Dipalmitoilfosfatidilkolin (DPPC)’in kırılma noktasi
ve geriye doĝru izotermi.

194
Şekil 3.50. 1/8 oranındaki Taxol®-DPPC kompleksin izotermi. Karışım
önceden Eppendorf tüplerde hazırlanmıştır.
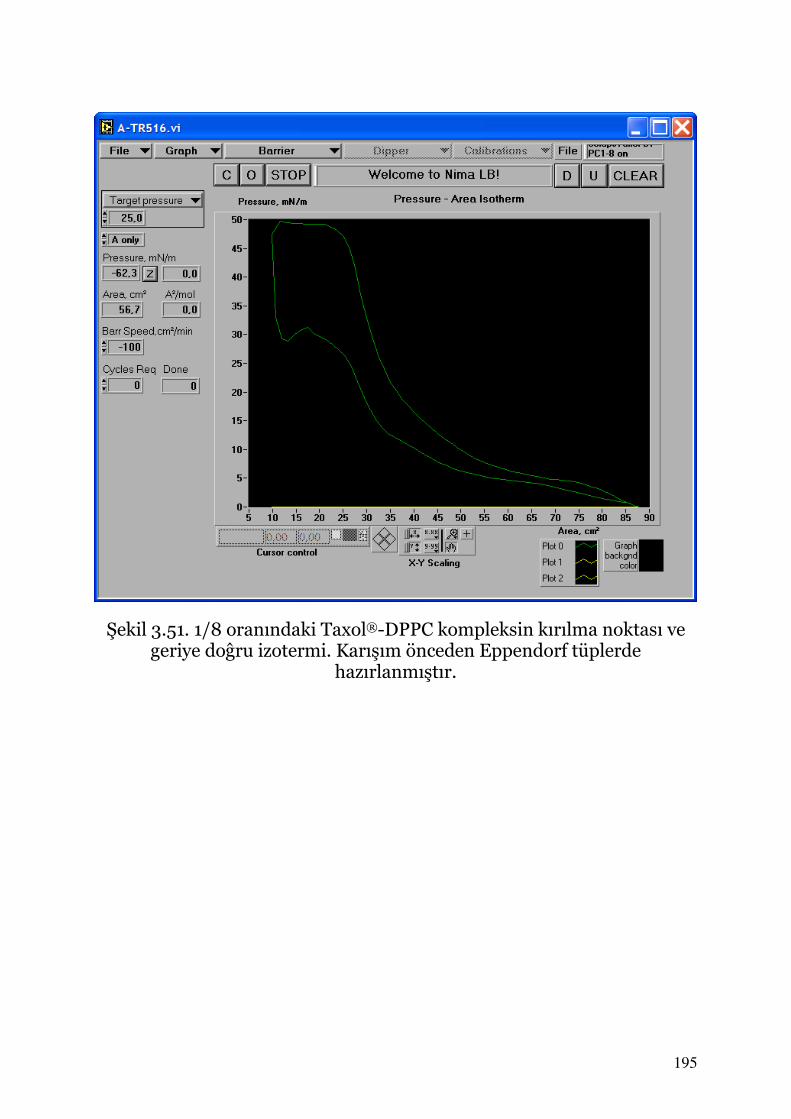
195
Şekil 3.51. 1/8 oranındaki Taxol®-DPPC kompleksin kırılma noktası ve
geriye doĝru izotermi. Karışım önceden Eppendorf tüplerde hazırlanmıştır.

196
Şekil 3.52. Taxol®-DPPC 1/8, 1/6, 1/4, 1/2 ve 1/1 (en alt) oranlardaki izotermleri. Karışım önceden Eppendorf tüplerde hazırlanmıştır.
Düz çizgiler ekstrapolasyonu göstermektedir.

197
Şekil 3.53. 1/6 oranındaki Taxol®-DPPC kompleksin kırılma noktası ve
geriye doĝru izotermi. Karışım önceden Eppendorf tüplerde hazırlanmıştır.

198
Şekil 3.54. Her deneyden önce elde edilmesi gereken yüzey saflıĝı sinyali.
Bu saflık sinyalinin gayet iyi olmasından, diĝer ölçümlere geçilebilmektedir.

199
Şekil 3.55. Saf yüzey sinyalinin elde edilmesinden sonra, esas ölçümlere geçilmeden önce, ilk önce dipalmitoylfosfatidilkolin (DPPC)’in uyumlu
izotermi bu şekilde olmalıdır.

200
Şekil 3.56. Dipalmitoylfosfatidilkolin (DPPC)’in kırılma noktası ve geriye
doĝru izotermi.

201
Şekil 3.57. Taxol®-DPPC (1/4) kompleksin bu konsantrasyondaki
yüzeysel basınç (π-A) izotermi. Karışım önceden Eppendorf tüplerde hazırlanmıştır.

202
Şekil 3.58. Taxol®-DPPC (1/4) kompleksin bu konsantrasyondaki kırılma noktası ve geriye doĝru izotermi. Karışım önceden Eppendorf tüplerde
hazırlanmıştır.

203
Şekil 3.59. Taxol®-DPPC (1/2) oranındaki kompleks oluşumunun yüzeysel basınç izotermi. Karışım önceden Eppendorf tüplerde
hazırlanmıştır.

204
Şekil 3.60. Taxol®-DPPC (1/2) kompleksin bu orandaki kırılma noktası
ve geriye doĝru izotermi. Karışım önceden Eppendorf tüplerde hazırlanmıştır.

205
Şekil 3.61. Taxol®-DPPC ekvimolar (1/1) oranındaki kompleks oluşumunun yüzeysel basınç izotermi. Karışım önceden Eppendorf
tüplerde hazırlanmıştır.

206
Şekil 3.62. Taxol®-DPPC ekvimolar (1/1) kompleksin bu orandaki kırılma noktası ve geriye doĝru izotermi. Karışım önceden Eppendorf tüplerde
hazırlanmıştır.

207
4. TARTIŞMA
Bu projemde, çok etkili bir antikanser ilâcı olan Taxol® (Paclitaxel) ile model
membran fosfolipidler arasında oluşan moleküler etkileşmeler ele alınmıştır103. Bu
konu ile ilgili elde edilecek olan bulgular, daha sonraki aşamalarda yeni ve özellikleri
iyileştirilmiş kemoterapötik formülasyonlarının geliştirilmesini mümkün kılacaktır.
Bu doğrultuda Taxol® ile tümör hücre zarı arasında meydana gelen etkileşmelerinin
incelenmesi amaçlanmıştır. Bunun için tümör hücre yüzeyi Langmuir-Blodgett tek
katman ve fosfolipid yüzeyleriyle modellnmiştir. Bu ölçümlerden elde edilen yüzey
kimyasal parametrelerine dayanarak daha sonra in vivo ortamda ilâç-hücre etkileşim
modellerinin geliştirilmesi amaçlanmıştır. Projemde deneysel tasarım olarak ilk önce
ön fizikokimyasal inceleme yapılmıştır. Bunun bilimsel dayanakları ve gerekçelri
Tablo-4.1.’de verilmiştir. Ayrıca da, bundan önce sunduğum ara raporlarda da bu
hususların önemi vurgulanmıştır. Bu doğrultuda, FTIR spektroskopik ve DSC
termodinamik ölçümleri yapılmıştır. Bu cihazların Fakültemizde bulunmaması
sebebiyle söz konusu ölçümler yeni kurulan Orta Doğu Teknik Üniversitesi Merkezi
Laboratuvarında yapılmıştır.
4.1. FTIR Ölçümleri
FTIR spektroskopisi fosfolipidlerle bağlanmasını takiben Taxol®’un
morfolojisinin değişip değişmediğini ortaya çıkarmak için uygulanmıştır. Bunun
özellikle yüksek ilâç konsantrasyonlarda ilginç olacağı düşünülmüştür. İlâcın katı
fazdaki (solid state) özelliklerinin kontrol edilmesi günümüzde farmasötik
endüstrinin başlıca amaçları arasında gösterilmektedir. Birçok ilâcın kendisine ait
farklı katı fazına bağlı olarak farklı uygulamaları söz konusudur. Bazen, bir ilâcın
sudaki veya başka çözücülerdeki çözünürlülüğü ilâcın hidrasyon seviyesi gibi katı
fazdaki özelliklerine bağlıdır. Ayrıca bir ilâcın sabitliliği (stabilitesi) bu ilâcın
morfolojik olarak birkaç farklı şekil arasında gösterdiği dönüşümüne bağlıdır. Bu
bakımdan, ilâcın katı faz kontrolleri ve karakterizasyonları önemlidir. Bu, özellikle
Taxol® için geçerlidir. Bu ilâç, karmaşık organik bileşen olup birkaç katı faz yapısı
göstermektedir (Bk.: Şekil 1.26). 4 farklı morfolojik şeklinden 3’ü-dihidrat, anhidrid
103 Projenin amacı ile ilgili daha ayrıntılı bilgi için bk.: Bölüm-2: Gereç ve Yöntem.

208
ve amorf şekilleri sabittir (Şekil 4.1). Projemde de, fosfolipidlerle tanınmasını
(recognition) takiben Taxol®’daki morfolojik değişikliklerinin meydana gelip
gelmediği konusu ele alınmıştır.
Kızılötesi (IR) spektroskopisi katı fazların incelenmesi için çok uygun bir
yaklaşımdır, çünkü ölçümler için incelenen maddenin çok az miktarını
gerektirmektedir. Bu projemde, FTIR spektroskopisi fosfolipidlerle tanınma
olayından sonra Taxol®’un meydana gelen morfolojik şekillerin incelenmesi için
uygulanmıştır.
Elde edilen bulgular Şekil 3.1-3.13’te gösterilmiştir. Şekil 3.2-3.7 gösterildiği
üzere, daha yüksek konsantrasyonlarda kullanıldığında, Taxol®’un amorf şeklinin
anhidrid şekline dönüştüğünü göstermektedir.
Şekil 3.1. 500 ile 4000 cm-1 aralığında L-α-fosfatidilkolinin tüm IR
spektumunu göstermektedir. 15’e yakın frekans bandı bu fosfolipid yapısıyla ilgili
önemli spektral ve bilgiler vermektedir (Bkz.: ayrıca Tablo-4.1). Bunların en önemli
olanları CH titreşimlerine ait olan 2750-3100 cm-1 ve CH2 wagging progression ve CH2
makaslama titreşimlerini kapsayan 1150-1400 cm-1 ve 1465-1475 cm-1 (PEVSNER ve
DIEM, 2003). Bunlardan ilki lipid açil zincirlirindeki konformasyonel düzen
hakkında, diğer ikisi ise açil zincirlerinin paketleme etkileri hakkında önemli bilgiler
vermektedir. 1258 cm-1 görülen ve PO2- titreşimlerine ait karakteristik frekans burada
1242 cm-1 ‘de görülmektedir. Aradaki 16 cm-1’lik fark kalan su etkileriyle
açıklanmaktadır104.
Taxol® 1600-1750, 1180-1300 ve 630-770 cm-1 büyük IR bandlarına sahiptir
(Bk.: Şekil 3.7, Şekil 4.1 ve Tablo 4.1). Çok küçük kaymalar dışında, tüm bu bandlar
ilâç-fosfolipid spektrumlarında da mevcuttur. Bu da, söz konusu bu komplekste
Taxol®’un bulunduğunu ve lipidle bağ oluşturduğunu göstermektedir. Tek başına
lipidin spektrumu ile ilâç-fosfolipid spektrumları karşılaştırıldığında (Şekil 3.1 ile
Şekil 3.7) Taxol®’un 1736 cm-1’daki karbonil stretching titreşimi hafifçe 1732 cm-1’ ye
104
Bu konu ile ilgili çok daha ayrıntılı bilgi Nanomedicine dergisine sunduğumuz makalemizde bulunmaktadır.

209
gitmektedir ki bu da ilâç ile fosfolipid arasında kompleks oluştuğunun bir başka
kanıtıdır. Bu, her yeni ve daha yüksek ilâç konsantrasyonlarının kullanıldığında (Şekil
3.2-3.7; Karbonil IR bandı 1736 cm-1’den→ 1707.39 cm-1’ye kaymaktadır) da
görülmektedir. Ayrıca, 3300-3500 cm-1’deki güçlü bandlar H-bağlarla bağlı olan OH
gruplarına aittir. 2880-3000 cm-1’deki karakteristi 2 band ise çok tipik bir fosfolipid
–CH ve –CH2 gruplarına aittir. İlâç ile fosfolipid arasında kompleks oluşmasını
takiben, söz konusu bandların intansitesinde önemli düşüşlerin olduğu ve ek band
kaymalarının meydana geldiği görülmektedir (Bk.: Şekil 3.1-3.7). Söz konusu
hidroksil bandları 3424 cm-1’den→ 3512 cm-1’ye kaymaktadır. Fosfolipid –CH ve –
CH2 gruplarına ait bandlar ise gittikçe yükselen Taxol® konsantrasyonlarda tamamen
yok olmaktadır (Bk.: Şekil 3.7). İlâcın tek başına ve fosfolipid kompleksindeki IR
spektrumları kıyaslandığında, 1647 cm-1’dekiC=C ve C=C’un shoulder yok olmakta ve
pik hafifçe daralmaktadır. Bu muhtemelen ilâç ile bağ yapması neticesinde, fosfolipid
tarafından alınan konformasyonlarda bir kısıtlamanın meydana geldiğinin işaretidir.
Bu da, Taxol®’un esnkeliğinde (flexibility) bir düşüşün olduğunun göstergesidir.
Ancak, bu bölgede henüz daha açıklanmayan ilâç yapılarının meydana gelebileceğini
de göstermektedir. Bu durumda, bu spektrum bölgesindeki değişikliklerden yola
çıkarak çok sağlıklı bir aypabilmek için “ağı su” (D2O) FTIR ölçüm yöntemi
uygulanmalıdır105.
Sonuç olarak, projemde kullanılan KBr tablet yöntemindeki suyun
uzaklaştırılması sonucu ilâç ile fosfolipid arasında kompleks oluşmasında genelde
fosfolipidin –CH ve –CH2 grupları ve H-bağlı yüzey OH grupları önemli ölçüde rol
oynamaktadır. İlâcın da PO2-, C=O, C=C ve CH2 grupları da bağ oluşumunda yer
almakatdır. Taxol® ile fosfolipid arasındaki kompleks oluşumundan sonra hidroksil
bandların değişikliğe uğraması, önceden varolan H-bağların koptuğunu ve bu da ilâç-
lipid kompleksinin oluştuğunun yeni bir kanıtı olarak kabul edilebilmeltedir.
Şekil 3.13. biyolojik önemi çok büyük olan lipid-metal katyon kompleks
oluşumunu göstermektedir. Burada metal katyonlar membran akışkanlığını
tetikleyen faktörler olarak incelenmiştir. Görüleceği üzere, Mg2+ ‘un sırasıyla fosfat,
105 Bu konunun önemi için Bk.: Helene Gaussier, Thierry Lefevre, and Muriel Subirade, Binding of Pediocin PA-1 with Anionic Lipid Induces Model Membrane Destabilization. Applied and Environmental Microbiology, Nov. 2003, Vol. 69, No. 11, 6777–6784.

210
karbonil ve CH2 titreşimlerine etkilerini daha iyi bir şekilde yansıtılması amacıyla üç
ayrı IR spektral bölgeye yer verilmiştir. Bu ilk iki etki, şeklin üst ve orta kısmında yer
almaktadır. Burada fosfatidilkolin üzerinde (1000-1175 cm-1) çok az bir etki
görülmektedir. Bundan farklı olarak H-bağlar ve bağlanmış ve bağlanmamış karbonil
gruplar üzerindeki etki (1635.3 ve 1734.5 cm-1 ‘de) çok daha belirgindir. Karboniller
üzerinde bu etki, karbonil grupların su moleküleriyle azalan etkileşmelere bir cevap
olarak H-bağlardaki azalmalardan kaynaklanmaktadır. Fosfat titreşim bölgesini
takiben 1467.6 cm-1 ‘de yeni bir band ortaya çıkmaktadır. Bu band, heksagonal
paketlemeye geçişi işaret eden önceden daha yüksek olan bağ frekansının
düşürülmesindeki Mg2+ iyonların etkisini göstermektedir. Şeklin alt kısmında yer
alan spektrum, 720 cm-1 ‘de görülen ve değişime uğramamış CH2 rocking
hareketlerine özgü ve lipid zincirin hareketliliğini düşürüldüğünün ve zincir düzenin
artırdığının göstergesi olan sinyalin görülmesine rağmen, lipid CH2 sinyallerinde
sadece çok hafif bir değişiminin meydana geldiğini ve lipid zincir düzeni ile ilgili
bunlardan çok az bilgi alınabileceğini göstermektedir (BINDER ve ZSCHÖRNIG,
2002). Jel fazda bulunan νas PO2- üzerindeki Mg2+’un fosfat gruplarını dehadrasyona
uğratma doğrultusundaki beklenilen etkisi burada görülmemektedir. Bu, 1115 ve 1137
cm-1 ‘de çıkması beklenilen sinyallerin yokluğundan ortaya çıkmaktadır (HAUSER,
1991; BINDER ve ZSCHÖRNIG, 2002). Fosfatidilkolinin karbonil grubunun
absorpsyon spektrumu üzerine ilginç etkilerin meydana geldiği görülmektedir. 1736.6
cm-1 ‘de görülen pik su ile etkileşmeye giren H-bağ ile bağlanmış karbonillerin
bulunduğunu göstermektedir. Mg2+ iyonları νC=O daha düşük dalga numaralarına
doğru götürmektedir. Bu etki, karbonil gruplarının iyonların hidrasyon tabakalarında
yer almalarından dolayı su molekülleri tarafından daha kolayca ulaşabilmelerinden
kaynaklanmaktadır. Buna bir alternativ olarak, bu etkinin lipidlerin polar fazlar arası
bölgede bulunduğundan, beraberinde lipid yapılarının faz değişimlerini de getirdiği,
karbonil gruplarında etkeleşme neticesinde meydana gelen konformasyonel
değişikliklerinden kaynakalanabileceği fikri de geçerli olabilir.
Mg2+ ile bağlanma, ayrıca lipidteki CH2 titreşimlerine de etki eder. Yukarıda
bahsedildiği üzere, yüzeysel su oluşumu, lipid-metal katyon ikilisinin oluşuşumunun
da önemli bir parçasıdır ve etkisi söz konusu spektrumda 3000-4000 cm-1
aralığındaki piklerden görülmektedir. Bu IR absorpsyonu, bu yüzeysel su oluşumuna

211
bağlıdır (POHLE, et. al., 2000). Lipid örneğinde kalan suyun etkisi, L-α-
fosfatidilkolinin difaransiyel taramalı kalorimetrik (DSC) termogramı ile de teyit
edilmiştir106.
Şekil 4.1. Taxol®’un çeşitli katı faz şekilleri. (Bk.:Jeong Hoon Lee, Un-Sook Gi, Jin-Hyun
Kim, Yongae Kim, Seon-Ho Kim, Hunseung Oh, and Bumchan Min. Preparation and
Characterization of Solvent Induced Dihydrated, Anhydrous, and Amorphous
Paclitaxel, Bull. Korean Chem. Soc. 2001, Vol. 22, No. 8, 925-928).
106 Termodinamik teyit ile ilgili yorum ayrıca bu projenin ara raporunda da yer almaktadır.

212
Lipid yapısındaki polar kısmına özgü sinyaller veren C-H absorpsyon
bandlarıyla polar olmayan bölgede bulunan C-H grup titreşimlerinden kaynaklanan
sinyallerin birbiriyle örtüştüğünden, yaklaşımımda söz konusu spektrum
kıyaslamaları sadece Taxol® ve L-α-fosfatidilkolinin sinyallerinin (fosfat, karbonil ve
CH titreşimleri) çakışmadığı IR bölgelerinde yapılmıştır. Bu şekilde ayrıca, ikili
kompleks iki ayrı kompleks oluşumu ile (Taxol®-fosfolipid ve fosfolipid-Mg2+)
karşılaştırılmıştır. Tüm ikili kompleks oluşum IR absorpsyon spektrumları Şekil 3.2-
3.7 ve Tablo-4.1’de verilmiştir.
FTIR ölçümlerinde katı fazda ve sıvı fazda hazırlanan örneklerle çekilen
spektrumlarının kıyaslanması gerektiğine dair gerekçelerden daha önce
bahsedilmişti. KBr yönteminin ortamdan yan sinyallere neden olan suyun
uzaklaştırması gerektiğinden, bu tür FTIR ölçümlerinde önemli br yeri vardır.
Bunların geçerliliği ile ilgili hususlara daha önce verdiğim projemin ara raporlarında
yorumlar bölümünde de değinilmiştir. Projemizde şimdiye kadar yer verdiğim
neticeler KBr tablet yöntemiyle hazırlanan örneklerle elde edilmiştir. Ancak, hücresel
ortam katı değildir. Söz konusu hücresel ortamın sıvı ve akışkan olduğunu göz
önünde bulundurarak, tasarımımda yer alan aynı ölçümleri sıvı örneklerle kontrollü
nem ortamında hazırlanan ince tabaka lipid filmi ile çekilen FTIR spektral
analizleriyle devam etmek arzusundayım. Sıvı örnekleri için: Characterization of a
Cationic Lipid-DNA Complex and Its Components, Phys. Chem. Chem. Phys., 2,
4642-4650, (2000)., FORATO, L.A., Bernardes-Filho, R. and Colnago, L.A., Protein
Struture in KBr Pellets by Infrared Spectroscopy, Anal. Biochemistry, 259, 136-141,
(1998) yayınlarda yer alan prosedürleri bir sonraki projemde esas alarak uygulamak
gerekmektedir107.
107 FTIR ölçümlerinde KBr pellet (tablet) ve sıvı örneklerin önemini vurgulayan ve birbirini tutmaları konusunda yapılan deneylerle ilgili şu kaynaklarda çok önemli protokol ayrıntıları bulunmaktadır: ANCHORDOQUY, T. J., Dean Allison, S., Molina, M. D. C., Girouard, L. G., and Carson, T. K., Physical
Stability of DNA-Based Therapeutics, Drug Disc. Today, 6, 463-470, (2001).
BANYAY, M., Sarkar, M. and Gräslund, A., A Library of IR Bands of Nucleic Acids in Solution,
Biophys. Chem., 104, 477-488, (2003).

213
BINDER, H., and Zschörnig, O., The Effect of Metal Cations on the Phase Behavior and Hydration
Charactersitics of Phospholipid Membranes, Chem. Phys. Lipids, 115, 39-61, (2002).
BLOOMFIELD, V. A., Crothers, D. M., and Tinoco, I. Jr., Nucleic Acids-Structures, Properties, and
Functions. University Science Books, Sausalito, CA 94965 (2000), Pp: 210-212.
BRUNI, P., Cingolani, F., Iacussi, M., Pierfederici, F. , and Tosi, G., The Effect of Bivalent Metal Ions
on Complexes DNA-Liposome: a FT-IR Study, 565-566, 237-245, (2001).
CLEMENT, J., Keifer, K., Kimpfler, A., Garidel, P., and Peschka-Süss, R., Large-Scale Production of
Lipoplexes With Long Shelf-Life, Eur. J. Pharmaceut. Biopharm., 59, 35-43, (2005).
FORATO, L.A., Bernardes-Filho, R. and Colnago, L.A., Protein Struture in KBr Pellets by Infrared
Spectroscopy, Anal. Biochemistry, 259, 136-141, (1998).
GARIDEL, P., Blume, A., and Hübner, W., A Fourier Transform Infrared Spectroscopic Study of the
Interaction of Alkaline Earth Cations With the Negatively Charged Phospholipid 1,2-dimyristoyl-sn-
glycero-3-phosphoglycerol,. Biochim. Biophys. Acta, 1466, 245-259, (2000).
GAUGER, D.R., Selle, C., Fritzsche, H., and Pohle, W., Chain-Length Dependence of the Hydration
Properties of Saturated Phosphatidylcholines as Revealed by FTIR Spectroscopy, J. Mol. Structure,
565-566, 25-29, (2001).
GÜNZLER, H. and Gremlich, H.-U., IR Spectroscopy. An Introduction, Wiley-VCH Verlag GmbH,
69469, Weinheim (Germany), (2002).
HACKL, E. V., Kornilova, S. V., Kapinos, L. E., Andrushchenko, V. V., Galkin, Vi L., Grigoriev, D. N.,
and Blagoi, Yu. P., Study of Ca2+, Mn2+ and Cu2+ Binding to DNA in Solution by Means of IR
Spectroscopy, J. Mol. Structure, 408/409, 229-232, (1997).
HAUSER, H., Effect of Inorganic Cations on Phase Transitions, Chem. Phys. Lipids, 57, 309-325,
(1991).
HÜBNER, W. and Blume, A., Interactions at the Lipid-Water Interface, Chem. Phys. Lipids, 96, 99-
123, (1998).
JOHNSTON, S. F., Fourier Transform Infrared – A Constantly Evolving Technology, Ellis Horwood
Limited, (1991).

214
LAI, E., van Zanten, J. H., Evidence of Lipoplex Dissociation in Liquid Formulations, J. Pharm. Sci.,
91, 1225-1232, (2002).
Le BIHAN, T., and Pézolet, M., Study of the Structure and Phase Behavior of
Dipalmitoylphosphatidylcholine by Infrared Spectroscopy: Characterization of the Pretranstion and
Subtransition, Chem. Phys. Lipids, 94, 13-33, (1998).
LEWIS, R. N. A. H., Winter, I., Kriechbaum M., Lohner, K. and McElhaney, R. N. Studies of the
Structure and Organization of Cationic Lipid Bilayer Membranes: Calorimetric, Spectroscopic, and X-
ray Diffraction Studies of Linear Saturated P-O-ethyl Phosphatidylcholines, Biophys. J., 60, 1329-
1342, (2001).
LEWIS, R.N.A.H. and McElhaney, R.N., Fourier Transform Infrared Spectroscopy in The study of
Hydrated Lipids and Lipid Bilayer Membranes. in: H.H. Mantsch and D. Chapman (Eds.), Infrared
Spectroscopy of Biomolecules. Wiley-Liss, New York, (1996), Pp: 159-202.
PEVSNER, A., and Diem, M., IR Spectroscopic Studies of Major Cellular Components. III. Hydration
of Protein, Nucleic Acid, and Phospholpid Films, Biopolymers (Biospectroscopy), 72, 282-289,
(2003).
POHLE, W., Gauger, D.R., Fritzsche, H., Rattay, B., Selle, C., Binder, H., and Böhlıg, H., FTIR-
Spectroscopic Characterization of Phosphocholine-Headgroup Model Compounds, J. Mol. Structure,
563-564, 463-467, (2001).
POHLE, W., Selle, C., Gauger, D. R., Zantl, R., Artzner, F., and Rädler, J. O., FTIR Spectroscopic
Characterization of a Cationc Lipid-DNA Complex and Its Components, Phys. Chem. Chem. Phys., 2,
4642-4650, (2000).
RUTHVEN, N. A., Lewis, H. and McElhaney, R. N., The Structure and Organization of Phospholipid
Bilayers as Revealed by Infrared Spectroscopy, Chem. Phys. Lipids, 96, 9–21, (1998).
SAENGER, W., Principles of Nucleic Acid Structure, Chapter 17: Water and Nucleic Acids. Mir
Publishers, Moscow, (1987), Pp: 63, 393-412 (The Russian translation).
SELLE, C. and Pohle, W., Fourier Transform Infrared Spectroscopy as a Probe for the Study of the
Hydration of Lipid Self-assemblies. II. Water Binding Versus Phase Transitions, Biospectroscopy, 4,
281-294, (1998).

215
SELLE, C., Pohle, W. and Fritzsche, H., FTIR Spectroscopic Features of Lyotropically Induced Phase
Transitions in Phospholipid Model Membranes, J. Mol. Structure, 480-481, 401-405, (1999).
SILVESTRO, L. and Axelsen, P. H., Infrared Spectroscopy of Supported Lipid Monolayer, Bilayer, and
Multibilayer Membranes, Chem. Phys. Lipids, 96, 69-80, (1998).
SORGI, F., and Huang, L., A Dry Powder Formulation for Gene Therapy, Pharm. Res., 12, S266,
(1995).
TAILLANDIER, I.E. and J. Liquier. (2002), in: J.M. Chalmers and Griffith (eds.), Handbook of
Vibrational Spectroscopy, John Wiley & Sons, Chichester, Vol. 5, Chapter 2, Pp. 3465-3480.
van der WEERT, M., Haris, P.I., Hennink, W.E., and Crommelin, D.J.A., Fourier Transform Infrared
Spectrometric Analysis of Protein Conformation: Effect of Sampling Method and Stress Factors, Anal.
Biochemistry, 297, 160-169, (2001).
ZUNDEL, G. Polar Interactions, Hydration, Proton Conduction and Conformation of Biological
Systems – Infrared Results, , in: W. Hoppe, W. Lohmann, H. Markl, H. Ziegler (Eds.), Biophysics,
Springer-Verlag, Berlin, Hedelberg, New York, Tokyo, (1983). Pp: 243-254.

216
Tablo 4.1 FTIR ölçümlerinde görülen önemli IR band titreşimleri.
Fosfolipid 3424.9
2922
2852.2
1736.6
700 – 1500
1467.4
1378.1
1242.6
1091.5
970.4
927.0
826.9
721.8
Su band
νs[(CH3)4]
νs[(CH3)4]
νs[C=O]
δ[C-H]
scissoring δ[(CH2)n]
δ[CH2]
νas[PO2-]
ν[PO2-]
asimetrik [+N-(CH3)3] stretching
bölgesi
ν[C-C-N]
ν [P(-O-C)n]
rocking ν[(CH2)n]
Phospholipid-Mg2+ 975.7
1095.4
1467.6
1635.3
1734.5
2261.5
2852.9
2923.3
3241.4
3405.5
ν[C-C-N]
ν[PO2-]
scissoring δ[(CH2)n]
H-bağlı olmayan CO
H-bağlı CO
CH stretching
CH2 stretching
CH2 stretching
Yüzeysel (Interstitial) su
Bağlı su
Taxol® 3064
1735
1709
1647
1072
-C-H
ester
keton
amid
Finger print

217
1244
708
Finger print
Finger print
Fosfolipid- Taxol® 709
969
1095
1245
1467
1647
1737
2853
2923
3441
Finger print
Asimetrik [+N-(CH3)3] stretching
Simetrik νs[PO2-]
Antisimetrik νas[PO2-]
Scissoring CH2δ (hexagonal)
C=O; simetrik alil C=C
C=O stretching
Simetrik νs[(CH3)4]
Simetrik νs[(CH3)4]
H-bağlı OH grupları
4.2. DSC Ölçümleri
Bu ölçümler, Taxol®’un fosfolipid yüzeyine ve faz değişimlerine etkisini ortaya
çıkarmak için yapılmıştır. Önceden hazırlanan fosfolipid türlerinin tek başına veya
farklı oranlarda Taxol® içeren komplekslerinin DSC termogramları bu raporumun sf.
113’te bulunan Şekil 3.14-3.19 verilmiştir.
Genel olarak, Taxol®’un sisteme katılması fosfatidilkolin moleküllerinde
birtakım perturbasyonlara veya daha muhtemel olarak mikrodomenlere yol
açmaktadır. Egg fosfatidilkolin kullanıldığında çok düşük oranda - 1.28% Taxol®
oranı bile glass transitiom (Tg) ısı değişim noktasının değişmesine yol açmaktadır
(Şekil 3.15). İkinci bir pikinise 56.19°C’de ortaya çıkması muhtemel olarak birden
fazla faz oluştuğunun işaretidir. Muhtemelen Taxol®’un katılmasından kaynaklana
mikrodomen oluşumunun işaretidir. Söz konusu faz değişiminin (Tg) fosfolipidlerin
polar kısmındaki yabancı moleküllerin bulunmasına son derece duyarlı olduğundan,
bu noktanın değişime uğraması sadece spesifik moleküler değişimlerle açıklanamaz.
10% Taxol® içerildiğinde, esas faz değişim ısısı (main phase transition temperature-
Tm) 26.16°C’den 28.25°C’ye gitmektedir, ancak etrafındaki termgram piki simetriktir.
Ancak, Tm piki daralmaktadır ve yüksekliğ düşmektedir. 1.28% Taxol®’un
içerildiğinde, esas faz değişimi ısı noktası (Tm) hafifçe daha düşük değerlere

218
gitmektedir (Şekil 3.15; 26.16°C→21.68°C), ancak etrafındaki termogram piki
simetrik kalmaktadır. Bu pikin genişliği daralmakatdır. Bu etki, söz konusu faz
değişimi ısı noktasının (Tm) kooperativitesinin arttığını ve fosfolipidlerin
esnekliğinidüşürmektedir. Bu durumda, çok düzenli olan all-trans hidrokarbon
fosfolipid zincir konformasyonu mevcuttur. Daha yüksekbir rotasyon serbestliğe yol
açan gauche (kinked) konformasyon ise söz konusu değildir. Taxol® içeriğinin 10%
faz değişim noktası (Tm) daha düşük değere değil, daha yüksek değerlere gitmektedir
(Şekil 3.16; 26.16°C→28.25°C). 56.19°C’deki ikinci pik artık görülmemektedir. Tm
piki ise simetrik değildir-sağ tarafında ikinci bir ufak pik belirmektedir. Bu da,
Taxol®’un daha yüksek konsantrasyonlarda fosfolipidlere karşı bir şekilde afinitesinin
arttığını göstermektedir. Tm‘e kıyasla glass transition temperature (Tg) pikinde daha
belirgin etkiler görülmektedir. Başka bir deyişle, Tg Taxol® varlığında daha büyük
değişime uğramaktadır. 1.28% Taxol® varlığında, Tg -19.75°C→-22.61°C gitmektedir
ve ayrıca da belirgin bir şekilde genişlemektedir. Aynı neticenin, 10%, 16% ve 20%
görülmesi (Şekil 3.16-3.18) görülmesi ise son derec ilginçtir. Bu neticeler, 0-10%
aralığındaki Taxol® varlığının fosfatidilkolin akışkanlğını arttırmaktadır (Tm‘i
düşürmektedir) ve esas faz değişimi noktasının (Tm) kooperativitesini arttırmaltadır
(faz geçişini daraltmaktadır). 10% Taxol® varlığında ise farklı bir durum öne
çıkmaktadır. Tm yükselmektedir ve 2nci pik yok olmaktadır. Ancak, 16% Taxol®
varlığı akışkan fazın (Tm azalmış durumdadır) tekrar ortaya çıkmasına neden
olmaktadır. Akışkan faz Şekil 4.2’de gösterilmiştir.

219
Şekil 4.2. Fosfolipid çift katmanların akışkanlığının meydana gelmesi.
20% Taxol® varlığında ise sadece Tg üzerindeki etki görülmektedir. Başka bir
deyişle, daha yüksek Taxol® konsantrasyonlarda fosfatidilkolin dispersiyonu daha
akışkan bir hale gelmektedir. Bu noktalarda glass transition noktası (Tg) esas faz
değişimi noktasından (Tm) daha önemli bilgiler vermektedir. Bu neticeler, Taxol®’un
diğer fosfolipid molekülleriyle etkileşmelerini inceleyen araştırmalardaki durumdan
çok farklıdır. Bu egg fosfatidilkolinin diğer fosfatidilkolin türlerine kıyasla farklılıklar
gösterdiğini ortaya çıkarmaktadır. Bu, Taxol®’un yapısından kaynaklanmaktadır (Bk.:
Şekil 1.26). Çok hacimli bir molekül olduğundan, Taxol® fosfolipid moleküllerinin iç
bölgelerine kadar ulaşamaz. Hidrofobik bölgelere penetrasyonu kısıtlıdır ve lipidlerin
ilk birkaç açil zinciri ile bağ yapmaktadır. Ancak, bu durum gerçekten lup olmadığı
paramanyetik özellikleri olan etiketler kullanarak Electron Spin Resonance
Spektroskopisi (ESR) ile teyit edilmelidir.
4.3. Langmuir-Blodgett Tek Katman Ölçümleri
30°C hava/su (air/water) tek başına ve Taxol® ile kompleks halindeki
fosfolipidlerin π-A bu raporun sf. 119’da bulunan 3.3. “Langmuir-Blodgett Tek
Katman Ölçümleri” başlıklı bölümde verilmiştir. Daha önce de bahsedildiği üzere
esas ölçümlere başlamadan önce, ortamın ve özellikle de söz konusu ara yüzeyin
temiz hali sağlanmalıdır. Bu yüzeyin temiz hale getirilmesi sırasıyla Şekil 3.20-

220
3.27’de gösterilmiştir. Temiz ara yüzeyin elde edilişi özellikle Şekil 3.27’de ve Şekil
3.47’de açıkça görülmektedir-bunun kriteri olarak yüzey basıncın 0 ile 1 arası olması
gerektiği hususu esas alınmaktadır (Bk.: www.nima.co.uk).
Yük taşımayan (zwitterionic, nötr) lipidlerin π-A izotermleri belirtilen
konsantrasyonda Şekil 3.28’den itibaren başlamaktadır. Bunlar gibi bir birine çok
benzeyen ve aralarında aynı polar baş grubuna sahip ve sadece hidrokarbon zincir
kısmında bir-CH2 grubu gibi çok küçük hacimli bir grupla farklılaşan fosfolipidlerde
bile farklı izotermler izlenmektedir.
Yüksek yüzey basınç bölgesinde izotermlerin şiddetli bir şekilde yükselişleri
açıkça görülmektedir (<50 Å2). Fosfolipidler arasında gözlenen bu eğri farkı, daha
uzun alifatik zincirli lipid hidrokarbom kısımların arasında meydana gelen daha
güçlü van der Waals kuvvetleri ve etkileşmeleri ile açıklanmaktadır. Daha geniş
moleküler alan, daha uzun alifatik zincire karşılık geldiği de açıkça görülmektedir.
Başka bir deyişle, bu durumda yüzey basıncın yükselmesi için daha fazla lipid
molekülleri gerekmektedir.
Şekil farklılıklarından başka, farklı fosfolipidlerde izotermlerin dikey olarak
yükseliş şekilleri arasında da (the sttepness of the isotherms) önemli farklılıklar
gözlenmektedir. Bu olgu, söz konusu lipidlerin hava/su ara yüzeyi üzerinde
oluşturdukları tilt angle ile açıklanabilir. Çünkü bu durumda geniş yüzey basınç (π)
aralığında bile bazı lipidler daha sıvı halde iken (liquid-expanded phase), başka lipid
türlerinin tek katmanları sıvı-kondanze (liquid-condensd phase) fazdadır. Bu
gerekçenin doğruluğu, bir lipidlerin açil zincir kısmı ne kadar daha uzunluğuna bağlı
olarak bu lipidin daha hdrofobik özellikleri kazanması hususu göz önünde
bulundurulduğunda açıkça ortaya çıkmaktadır.
Şekil 3.28 bir lipidin konsantrasyon ayarlanması deneylerini göstermektedir.
Şekil 3.28’den başlayarak Şekil 3.62’ye kadar farklı deneysel tasarımlar gösterilmiştir.
Örneğin, Şekil 3.30-Şekil 3.39 arasında verilen izotermler, önceden belirlenen
konsantrasyonlardaki Taxol®-fosfatidilkolin karışımlarının doğrudan Langmuir-
Blodgett tek katman üzerinde oluşturularak elde edilmiştir. Şekil 3.40 sonraki

221
şekillerde ise söz konusu karışımlar önceden Eppendorf tüplerde hazırlanıp tek
katmana uygulanmıştır. Aynı bölümde, π-A izotermleri, lipidin ulaştığı kırılma
noktası (collapse point) ve geriye doğru izotermleri gösterilmiştir. Bu şekilde molekül
başına düşen alanı (Area/molecule) hesaplamak çok daha kolaylaşmaktadır. Çünkü
bu şekilde monolayer compression-extension eğrileri kıyaslayarak tek katman
üzerindeki lipid molekülünün veya ilâç karışımının ne kadar alan kapsadığını
hesaplamak mümkündür.
Şekil 3.48 ise dipalmitoylphosphatidil choline’in (DPPC) tipik bir izotermidir.
Bu izoterm diğer lipidlere göre farklıdır, çünkü bu tipik liquid expanded→liquid-
condensed faz değişimlerini göstermektedir. Bu bakımdan, bu değişimler first-order
olarak adlandırılmaktadır.
Tek başına fosfatidilkolin’den başka bir diğer kontrol eğri de tek başına
Taxol®’un eğrisi elde edilmiştir ve Şekil 3.29’de gösterilmiştir. Görüldüğü üzere,
lipidlere kıyasla Taxol®’un izotermi son derece düzgün bir şekilde ve dalgalanma
göstermeksizin gradient olarak yükselmektedir. Bu da, Taxol®’un yüksek bir
kompresibiliteye sahip olduğunun bir göstergesidir. Başka bir deyişle, air/water ara
yüzeylerde Taxol®’un yüzeysel basıncı lipidlere göre çok daha düşüktür ve bu da
ilâcın çok düşük bir yüzey aktiviteye sahip olduğunu göstermektedir. Yüzey basıncın,
yüzey gerilimin yüzey aktivitesine sahip moleküller tarafından düşürülmesinin ifadesi
olduğundan, Taxol®’un su yüzeyinde ekspansiyonu lipidlere göre çok daha düşük
olduğu hususu ortaya çıkmaktadır. Bu da mantıklıdır, çünkü lipidlerin amfifilik
doğasından dolayı baş gruplaru su yezeyi ile tams etmek durumundadır ve zincirleri
ise hava fazına doğru yönelirler. Taxol® ise çok hidrofobiktir. Lipidler gibi air/water
ara yüzeylerde aynı eğilimi göstermemektedir. Bu yüzden, Taxol® gibi bir molekülün
yüzey basıncını çok daha düşük değerlere götürmektedir.
4.3.1. Bu Projede Yapılan Langmuir-Blodgett Tek Katman Ölçümlerinin
Daha Önce Başka Bir Cihazla Yaptığım Ölçümlerle Kıyaslanması
Bu bölümde bu projemde elde edilen neticelerin daha önce çok daha temiz
ortamlarda başka bir Langmuir-Blodgett cihazıyla ve çok deneyimli araştırmacıların
kontrolü altında yapmış olduğum deneylerle kıyaslamalara yer verilmiştir. Bu şekilde

222
bu projemde elde edilen verilerin geçerliliği ve tekrarlanabiliriliği hususu
vurgulanmaktadır. Ayrıca da, bu tür araştırmalarda farklı cihazların aynı neticeyi
verip vermediğini inceleyerek cihazlar arasında ölçüm hassasiyetinin de kontrol
edilmesi çok önemlidir.
Aşağıda gösterilen neticeler, 1998 yılında konuk araştırmacı olarak
bulunduğum Bergen Üniversitesi, Biyokimya ve Moleküler Biyoloji Bölümünde
yürütülen EU destekli “Acyl Specificity and Compartmentation in
Glycerophospholipid Metabolism During Signal Transduction in Brain” başlıklı
proje kapsamında elde edilmiştir. Bu neticeleri henüz daha yayımlamadım ve bu
raporumda bunlara söz konusu sağlıklı kıyaslamaları yapabilmek amacıyla yer
verilmiştir. Söz konusu projede nörokimyasal özellikleri olan klorpromazin ilâcı ile
model lipid tek katmanlar arasındaki moleküler etkileşmeler incelenmiştir.
Taxol® ile ilgili projemde elde ettiğim neticelerle bunların kıyaslandığında çok
iyi bir örtüşmenin söz konusu olduğu ortaya çıkmaktadır. İlk önce, Fakültemizde
yaşadığım ortam temizliği sorunu Norveç’teki çok iyi donanımlı üniversite
laboratuvarında da yaşanmıştır. Bu sorun, Şekil 4.3 ve 4.4 izotermlerine de
yansıtılmıştır. Bu çok büyük sorunu minimuma indirmek amacıyla Fakültemize
NIMA Technology’nin ürettiği Langmuir-Blodgett tek katman cihazı108 yanında
sunulan koruyucu konteyner ile alınmıştır. Bu çok tozlu olan 422 No’lu Merkez
Laboratuvarımızdaki ortamdan uzaklaşarak deneylerin temiz bir ortamda
yapılmasında çok faydalı olmuştur. Fakülteye getirdiğim cihazın bazı rakip firmaların
ürettiği (www.kibron.com ve www.ksvltd.com) üstünlükleri de vardır. Örneğin, son
derece “user-friendly” software sayesinde neticeleri doğrudan üst üstte getirip
inceleme fırsatı sunmaktadır. Ancak, yine de bu projemde elde ettiğim verileri Excel®
gibi uygun bir grafik çizim programına dönüştürüp yayın yapmak üzere tekrar
grafikleri elde edilecektir. Fakültemize getirdiğim cihaza ayrıca da çok uygun bir su
sirkülatörü monte edilmiştir. Bu şekilde sıcaklık kontrolü altında Şekil 4.10-4.11’de
gösterilen ortam sıcaklığının önemi bir kez daha vurgulanmıştır. Bu konunun önemi
daha önce çözeltilerin pH kontrolü (Bk. Şekil 4.7) ve 4.8 ve saf suyun kullanılmasının
(Bk.:Şekil 4.9) önemi ile birlikte ara raporlarımda ayrıntılı olarak ifade edilmiştir.
108 Bk.: www.nima.co.uk veya www.langmuir.com.

223
Şekil 4.3. Kibron marka (www.kibron.com) Langmuir-Blodgett cihazıyla yapılmış bir
deney. Ara yüzeyin temizlilik testi gösterilmektedir. Üstte, kirli bir yüzeyin neden
olduğu anlamsız izotemler görülmektedir. Altta verilen oklarla
dpalmitoilfosfatidilserin (DPPS) çok düzgün bir (π-A) izotermi görülmektedir.

224
Şekil 4.4. Kibron marka Langmuir-Blodgett cihazıylayüzey temizliliği testinden sonra
(üstteki eğriler) elde edilen düzgün bir lipid izoterminin elde edilişi (altta).

225
Şekil 4.5. Kibron marka Langmuir-Blodgett cihazıyla elde edilen dipalmitoylphoshpatidyl
choline-chlopromazine (DPPS-chlorpromazine) bileşimlrinin izotermleri. Farklı
orandaki karışımlar gösterilmiştir.
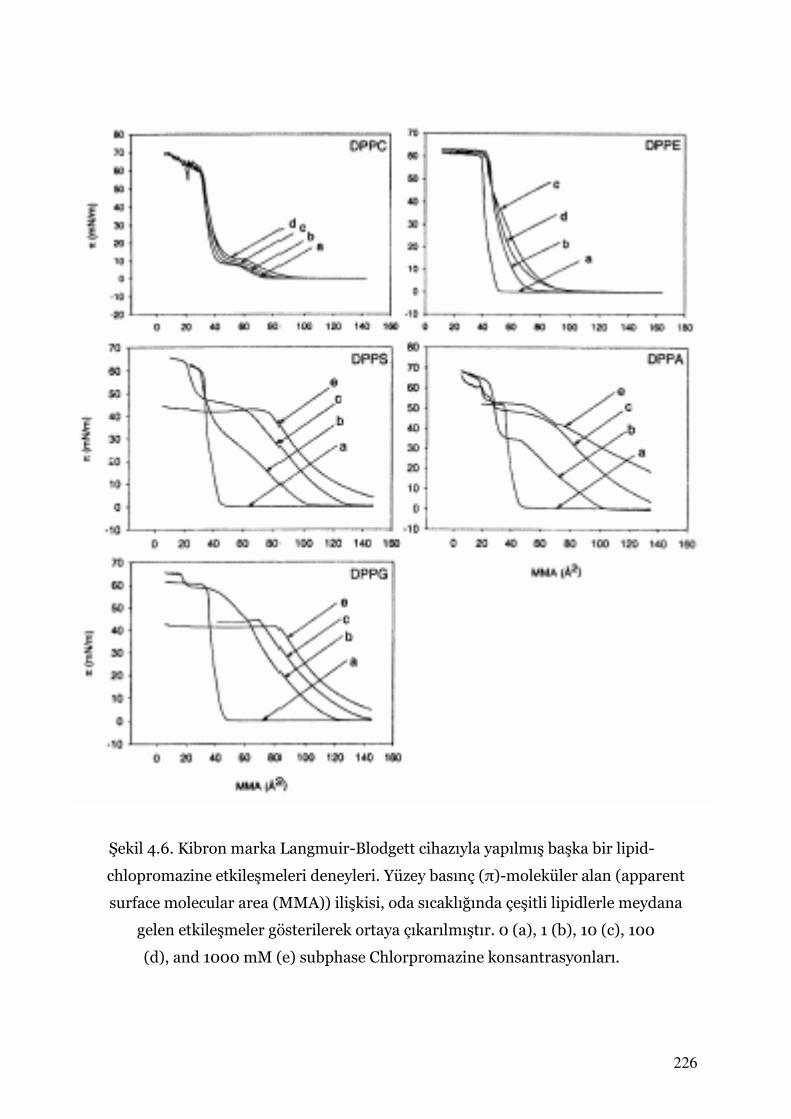
226
Şekil 4.6. Kibron marka Langmuir-Blodgett cihazıyla yapılmış başka bir lipid-
chlopromazine etkileşmeleri deneyleri. Yüzey basınç (π)-moleküler alan (apparent
surface molecular area (MMA)) ilişkisi, oda sıcaklığında çeşitli lipidlerle meydana
gelen etkileşmeler gösterilerek ortaya çıkarılmıştır. 0 (a), 1 (b), 10 (c), 100
(d), and 1000 mM (e) subphase Chlorpromazine konsantrasyonları.

227
Şekil 4.7. Yukarıdaki izotermler üzerie Na+ iyonun etkisi. Deneyler Kibron marka
Langmuir-Blodgett cihazı ile yapılmıştır. Lipid alanındaki artış açıkça görülmektedir.
Lipid olarak DPPS kullanılmıştır ve monolayer’ı oda sıcaklığında uygulanmıştır; (A) 1
mM CPZ and (B) 20 mM CPZ subphase with 0 (a),
2 (b), and 150 (c) mM NaCl.

228
Şekil 4.8. 150 mM NaCl, DPPS izotermler üzerine etkisi. Tüm izotermler plato ile biten
faz değişimi göstermektedir. Kibron marka Langmuir-Blodgett cihazı kullanılmıştır;
0 mM cations (1),
2 mM NaCl (2), 2 mM LiCl (3), 150 mM LiCl (4), and 150 mM NaCl (5).
Söz konusu iyon etkileri aşağıdaki çizimde daha ayrıntılı bir şekilde göterilmiştir109:
109 Burada sadece modelin algılanması için veridiğinden ve daha sonra buna atıfta bulunulmadığında bu şekle numara verilmemiştir.

229
Şekil 4.9. Milli-Q saf su ile yapılmış tek katman deneyleri. Kibron marka Langmuir-
Blodgett cihazı kullanılmıştır. Diğer ortam parametreleri şunlardır: Surface
pressure/area curve for a DPPS monolayer spread on
milli-Q water (1), 1 mM CPZ (2), 10 mM CPZ (3), 1 mM CPZ
(4), 10 mM CPZ (5), 1 mM CPZ (6), and 10 mM CPZ (7).
10 mM CPZ.
Şekil 4.10. Farklı sıcaklıklarda elde edilen DPPS monolayer izotermleri. Moleküler alanlar
π= 20 mN/m ortam sıcaklığı ıskalasına göre çizilmiştir. Deneyler Kibron marka
Langmuir-Blodgett cihazı ile sıcaklık kontrol sistemi kullanarak yapılmıştır.

230
Şekil 4.11. Farklı sıcaklıklarda yapılan DSPS monolayer izotermleri. Moleküler alan
hesaplamaları π= 20 mN/m’ında yapılmıştır ve ortam sıcaklığı ıskalasına göre
çizilmiştir. Deneyler Kibron marka Langmuir-Blodgett cihazı ile sıcaklık kontrol
sistemi kullanarak yapılmıştır.
4.4. Taxol®- Fosfolipid Membran Arasında Oluşan Yapı
Bu projemde kullanılan ölçümler sayesinde ortaya çıkarılan Taxol®-fosfolipid
etkileşmelerine dayanarak şu model geliştirilmiştir (Şekil 4.12).
Şekil 4.12. Moleküler düzeyde oluşan Taxol®-model membran fosfolipid yaıları.
Sağda (B) şeklinde verilen model bu projemde elde edilen neticelere bakacak
olursak çok daha muhtemeldir. Bu hususlara daha önce çok detaylı olarak yer

231
verilmiştir. Ayrıca da, bu model projeden kaynaklanacak olan makalelere de
yansıtılacaktır. Bu yüzden, bu konuya burada çok kısa olarak değenilmiştir.
4.5. Biyoteknolojik Boyut
Bu projemin 2 ayrı boyutundan biridir. Taxol®’un hidrofobik özeklerinden
kaynaklanan çözünürlülük problemleri ile ilgilidir. Şimdiye kadar geliştirilen
taşıyıcıları ile ilgili verilere dayanarak, bu formülasyonlarının lipide dayalı kontrollü
sistemlerin geliştirilmesinde önemli bir motivasyonu teşkil etmektedir. Bu bölümde,
projemde geliştirilen Taxol®-membran fosfolipid etkileşme modellerinin
biyoteknolojik tasarımlarını vurgulamaktadır. Bu boyut son derece geniş olduğundan
burada sadece önemli şekillere yer verimiştir. Buradaki modeller daha önce
hazırladığım ders notlar olarak Anabili Dalımızın isteği üzere Fakültemize bunların
bir ders olarak vermem konusunda 16.11.2007 tarihinde yazılı olarak bir başvurum
olmuştur. Ancak bu dilekçeme herhangi bir cevap (oumlu veya olumsuz, sözlü veya
yazılı) verilmemiştir. Bu modelleri istenildiği takdirde raportörlerimize seminerler
vererek açıklayabilirim. Bunları burada açıklamak bu raporumun hacmini çok
arttıracaktır. Bu modeller projeden kaynaklanacak olan makalelere de
yansıtılacağında, bunlara burada fazla yer verilmeyecektir. Burada yer alan tüm
şekillere renkli olarak www.interscience.wiley.com’dan ulaşmak mümkündür.

232
Şekil 4.13. Lipidlere dayalı antineoplastik formülasyonlarının geliştirilmesi. (Bk.: Gerben
A. Koning and Gerard C. Krijger, Targeted Multifunctional Lipid-Based Nanocarriers
for Image-Guided Drug Delivery, Anti-Cancer Agents in Medicinal Chemistry, 2007,
7, 425-440).

233
Şekil 4.14. Hücre içi (intraselüler) antikanser formülasyonlarının akıbeti. (Bk.: Gerben A.
Koning and Gerard C. Krijger, Targeted Multifunctional Lipid-Based Nanocarriers for
Image-Guided Drug Delivery, Anti-Cancer Agents in Medicinal Chemistry, 2007, 7,
425-440).
Şekil 4.15 Nanotıp konuları doğrultusunda antikanser ilâçların kullanım perspektifleri
(Bk.: Yiyao Liu, Hirokazu Miyoshi and Michihiro Nakamura, Nanomedicine for drug
delivery and imaging: A promising avenue for cancer therapy and diagnosis using
targeted functional nanoparticles, Int. J. Cancer. 120, 2527–2537 (2007).

234
Şekil 4.16. Bu projemde de ele alınan antikanser ilâç formülasyonlarının bastleştirilmiş
hazırlanma şemaları.
Şekil No. 4.17. Taxol® gibi çözünürlülük sorunları olan antikanser ilâçlarının
hedeflendirilmiş tedavi uygulamaları (Bk.: Yiyao Liu, Hirokazu Miyoshi and
Michihiro Nakamura, Nanomedicine for drug delivery and imaging: A promising
avenue for cancer therapy and diagnosis using targeted functional nanoparticles, Int.
J. Cancer. 120, 2527–2537 (2007).
.

235
Şekil 4.18 In vivo kanser kemoterapilerde kullanılmak üzere geliştirilmiş hedfelendirlimiş
antikanser tedavi profileri ve biyokonjuge Qdot’ların uygulanma presipleri (Bk.:
Yiyao Liu1, Hirokazu Miyoshi and Michihiro Nakamura, Nanomedicine for drug
delivery and imaging: A promising avenue for cancer therapy and diagnosis using
targeted functional nanoparticles, Int. J. Cancer. 120, 2527–2537 (2007)110.
110 Şekil 4.18-b)’de gösterilen in vivo görüntüleme sisteminin Fakültemize getirilmesi için proje hazırlanmıştır-bk.: Ek-17.

236
4.6. Hücresel ve Moleküler Biyolojik Boyut
Bu konu, projemin ikinci boyutunu oluşturmaktadır ve daha çok hücresel
biyolojik teorilere dayanarak Taxol®’un günümüzde gerçerli olan etki mekanizmasına
ek bilgiler katmak amacıyla geliştirilmiştir. Bu konu ile ilgili olarak aşağıda bir öncü
hipotez de sunulmuştur. Buradaki modeller, antikanser ilâçların membran
akışkanlığı üzerindeki etkilerine dayanmaktadır (Şekil 4.19-4.20 ve Tablo-4.2-4.4).
Şekil 4.19. Antikanser ilâçların hücresel hedefleri ve kanser ile bağlantı noktaları (Bk.:
Petty, 1993).

237
Tablo-4.2 Hücre zarı akışkanlığı ve kanserle bağlantısı. Tablo DPH floresan
spektroskopik ölçümlere dayanarak yapılmıştır (Bk.: Petty, 1993).
Tablo-4.3. Neoplazmik Transfromasyon ve Hücrelerdeki Morfolojik Değişiklikler (Bk.:
Petty, 1993).

238
Tablo-4.4. Neoplazmik Transformasyona Yol Açan Bazı Membran Proteinleri (Bk.: W.
Petty, 1993).

239
Şekil 4.20. Hücre membranı ve neoplazmik oluşumlarındaki rolü (Bk.: W. Petty, 1993).

240
Şekil 4.21. Taxol®’un sinyal iletim mekanizmalarındaki yeri.
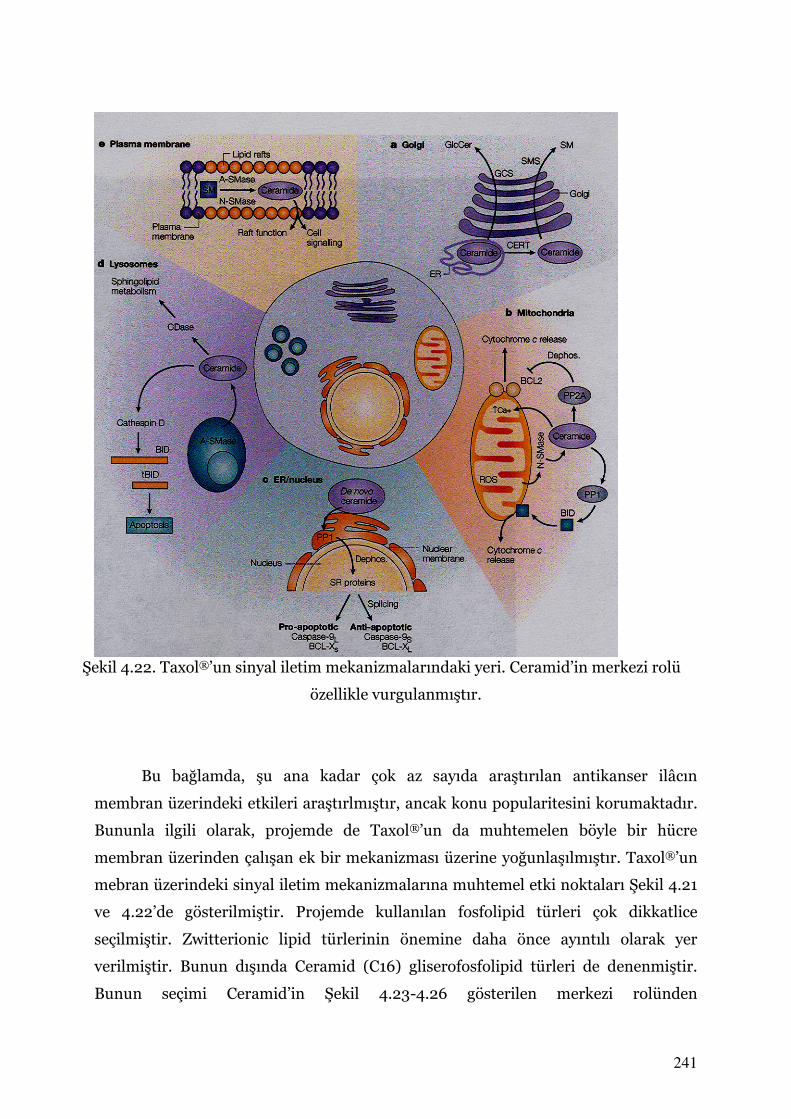
241
Şekil 4.22. Taxol®’un sinyal iletim mekanizmalarındaki yeri. Ceramid’in merkezi rolü
özellikle vurgulanmıştır.
Bu bağlamda, şu ana kadar çok az sayıda araştırılan antikanser ilâcın
membran üzerindeki etkileri araştırlmıştır, ancak konu popularitesini korumaktadır.
Bununla ilgili olarak, projemde de Taxol®’un da muhtemelen böyle bir hücre
membran üzerinden çalışan ek bir mekanizması üzerine yoğunlaşılmıştır. Taxol®’un
mebran üzerindeki sinyal iletim mekanizmalarına muhtemel etki noktaları Şekil 4.21
ve 4.22’de gösterilmiştir. Projemde kullanılan fosfolipid türleri çok dikkatlice
seçilmiştir. Zwitterionic lipid türlerinin önemine daha önce ayıntılı olarak yer
verilmiştir. Bunun dışında Ceramid (C16) gliserofosfolipid türleri de denenmiştir.
Bunun seçimi Ceramid’in Şekil 4.23-4.26 gösterilen merkezi rolünden

242
kaynaklanmaktadır. Bundan farklı olarak bir de negativ yüklü fosfatidilserin lipidleri
de incelenmiştir. Bunları seçimi de Şekil 4.27’de gösterildiği üzere söz konusu
fosfatidilserininin tümör oluşumlarında önemli olan membran lipid raftlardaki
rolünden kaynaklanmaktadır. Çünkü birçok kanser hücre membranında normal
hücrelerden farklı olarak fosfatidilserin ekspresyonu çok önemli artışlar
göstermektedir.
Şekil 4.23. Ceramid’in hücresel fosfolipid metabolizmasındaki merkezi rolü.

243
Şekil 4.24. Ceramid’in hücresel fosfolipid metabolizmasındaki merkezi rolünü gösteren başka bir şema.

244
Şekil 4.25. Ceramid’in hücresel apoptoz olaylarında ve bununla bağlantılı olarak lipid
raftların oluşumundaki rolü.
Şekil 4.26. Ceramid’in hücre mitokondri membranıyla bağlantı noktaları ve apoptozda oynadığı önemli rol.

245
Şekil 4.27. Fosfatidilserin’in antineoplazmik oluşumlarındaki rolü.
Tüm bu konular, projemden kaynaklanacak yayınlarda ele alınacağından bunlara burada daha fazla yer verilmeyektir.

246
4.7. Projemde Geliştirilen Tasarımın Diğer Analitik
Uygulamalardaki Yeri
4.7.1. İlâç Ligandlarının Ayrışımı ve Analizi için İmobilize Lipozom
Kromatografisinin Geliştirilmesi
Bu doğrultuda ilâç-fosfolipid arasındaki tanıma (drug-phospholipid
recognition) ile ilgil bu şekilde elde edilen önemli fizikokimyasal parametreler çok
yakın bir gelecekte çeşitli yapı ve işlevlere sahip farmasötik formülasyonlarının
ayrıştırılması, saflaştırılması ve analizleri için yeni ve analitik özellikleri iyileştirilmiş
kromatografik stasyoner fazların üretilmesinde ve uygulamasında kanımızca
kullanılabileceklerdir.
Projemde, bu bağlamda çeşitli katyonların da varlığında ilâç
konformasyonlarına spesifik olarak ve gerektiğinde spesifik olmayan bir şekilde
çeşitli tepkilemelerde önem taşıyan yapı-aktivite ilişkisi araştırmalarında bu
bileşenlerde moleküler ve yüzeysel etkileşmelerinin sıvı, çift-fazlı sistem olarak
kullanılmalarını vurgulamaktayız. Kromatografik kolonun jel matrikslerine böyle
kovalent olarak bağlanmış lipozomlarla çeşitli ilâç türlerinin veya plazmidler arasında
kompleks oluşumunun stabilite, bağlanma afiniteleri ve daha sonra meydana gelen
konfmasyon değişikliklerinin ortaya çıkarılmasında çok daha kolay
kullanılabilecekleri gibi lipozomlarla bağ yapan, reaksyona giren ve bileşkler
oluşturan birçok biyolojik ve farmakolojik özellikleri olan etkin makromoleküllerin de
yapılarının aydınlatılması kolaylaşacaktır. Kanımızca, böyle tasarlanmış sistemler
önemli biyospesifik tanıma olaylarında kantitativ analizleri mümkün kılacaktır ve
daha sonra diğer parametreler dışında, önemleri bu projemde de vurgulandığı gibi,
özellikle ortam sıcaklığı, pH, pI ve tampon içeriğine bağlı olarak membrana-bağlı
ilâçların nasıl değişime uğradıklarını ve bunun nasıl bir biyolojik önemi olduğunu
açıklayacaktır. Bu şekilde elde edilen verilerle biyomembran üzerinden hücre
sitozolüne geçiş yapan çeşitli ilâçları ve tedavi amaçlı sentezlenen farmasötik
formülasyon bileşenlerinin farklı boyut ve lipid türüne sahip hücre membranlarla

247
kompleks oluşumunun diğer önemli parametrelerini ve özellikle biyolojik neticeleri
olan membran akışkanlığı ve bu bağlamda patolojik durumların ortaya çıkması ile
ilgili konular aydınlatılacaktır.
Bu şekilde kolayca modifiye edilen lipozomlar, sıvı fazı olan ve stasyoner fazı
teşkil eden biyomakromoleküllerin spesifik olarak ayrıştırılması için bir model
oluşturan lipozomun iç hacmini içeren kısım ile bir çeşit sıvı çift fazlı sistem olarak
algılanmaktadır. Böylece, imobilize lipozom kromatografisi sıvı-sıvı partition
kromatografisinin (liquid-liquid partition chromatography) bir çeşidi olarak
görülmektedir. Bu tasarımımızda, lipozomun sıvı iç hacmi ile dış sıvı fazı arasında
çeşitli moleküler ağırlığa sahip maddelere karşı yüksek geçiş özelliklerine sahip olan
vezikül katmanı sayesinde bir faz farklılığı oluşmaktadır. Bu sistem, elektrostatik ve
hidrofobik bağ kuvvetlerini kullanarak konformasyonel değişiminin de meydana
geldiği veya tercihe göre getirilmediği durumlarda ilâçların imobilizasyonunda
kullanılabilmektedir. Projemde, bu bağlamda, bu çift fazlı sistem konformasyona
spesifik olarak veya tercihe göre olmayarak fosfolipidlerle nükleik asitler arasında
kompleks oluşumunun araştırılması konusu, Mg2+ veya diğer çift değerli katyonların
(Me2+) da varlığında ele alınmıştır. Lipidlerin faz değişimi ve faz geçiş sıcaklığının
üzerinde ve altındaki ısı değelerinde çalışarak, bu sistemin termostatik kontrolü de
mümkündür. Ayrıca, lipozom ve farmasötik bileşenlerinin bu yaklaşım sayesinde
değişime uğratılarak lipid membranlarının hidrofobik ve elektrostatik baş gruplarını
(membrane phospholipid head groups) yerel erişimi yükseltilir.
Lipid vezikülleri ile farklı ilâçlar arasında oluşan komplekste, bu gibi
konformasyon değişikliği parametreleri ayrıntılı olarak henüz daha araştırılmamıştır
ve hedefendirilmiş ilâç taşıyıcılarının yapı aydınlatılmasında kullanılmaları kanımca
çok ilginç olmalıdır. Bu bağlamda, söz konusu farmasötik formülasyonlarının yük
taşımayan (zwitterionic, nötr) fosfolipidlerin çeşitli katyonların varlığında membran
füzyonu meydana getirip getirmedikleri araştırılabilir. Böylece, bu şekilde kovalent
olarak kromatografik kolonun jel matriksine bağlanan lipozomların kendi
akışkanlıklarını ve yüzey tanıma özelliklierini de kontrol ettikleri fikri substrat olarak
ele alınan farklı boyut ve konformasyona sahip ilâçlar için de bu soyut varsayımlar ve
somut parametreler incelenebilecektir. Buradaki amaç, bu şekilde sterik olarak

248
stabilize edilen veziküllerin elektrostatik etkileşme kaabiliyetlerini kullanarak
lipozom yüzeyinde ilâç moleküllerinin adsorbsiyonunu gerçekleştirmektir.
Söz konusu lipid fazın içeriğini doğal hücre membranlarda bulunan bileşiklere
benzerlik gösterecek şekilde seçerek ve tampon içeriğini, pH, pI ve T°C gibi
fizikokimyasal parametreleri de bu doğal hücre ortamında ölçülen değerlerde tutarak,
çeşitli lipid’e-bağlı ilâç konformasyonları araştırılabilir. Bu şekilde kovalent olarak
imobilize edilmiş lipozomların stabiliteleri, çeşitli ilâç ve terapötik nükleik asitlerin111
membranlara bağlanma afiniteleri ve fosfolipidlere bağlı nükleik asitlerin
kompaktizasyon özellikleri incelenmektedir. Bu tasarımımız, çok katmanlı
(multilamellar vesicles, MLV) veya tek katmanlı (unilamellar vesicles, ULV)
lipozomların veya veziküllerin kromatografik kolon jel matriksine (chromatographic
gel beads) kovalent bağlama süretiyle enjektör, pompa ve UV-detektör ile donatılmış
yüksek performanslı sıvı kromatografisi (high perfomance liquid chromatography,
HPLC) ile söz konusu fizikokimyasal parametrelerinin incelenmesine dayanmaktadır.
Bu şekilde kovalent112 olarak stabilize edilmiş fosfolipid vezikülleri çok daha
akışkandır ve doğal hücre membranlarına çok daha iyi bir yaklaşım ve modeli teşkil
edip, ilâçların membran akışkanlığı üzerindeki etkilerini incelemek için çok daha
uygun bir sistemdir. Fosfolipidler, jel’e fosfatlarıyla fosfodiester bağ ve aktiv grubun
karbonat kısmıyla nükleofilik fosfat katalizi ile bağlanmaktadır. Biyomembran
ortamına yaklaştığı için imobilize edilmiş veziküllerde heterojen lipid katmanın elde
edilmesi tercih sebebidir. Bu tasarımda, tek lipid veya birkaç lipid karışımının
kullanıldığında genelde elektrostatik ve hidrofobik etkileşmeler meydana
gelmektedir. Böylece, bu tasarım sayesinde membran-ilâç arasında oluşan
kompleksin oluşma kinetiğini de incelemek mümkündür. Ayrıca, bu tasarım, serbest
iyonların ve membrana bağlı olarak işlev yapan çeşitli yüklü deterjanların nükleik asit
kondanzasyonu ve kompaktizasyonu gerçekleştiren ajanlar olarak lipid’e bağlı gen
taşıyıcı tasarımlarda da, fizikokimyasal özellikleri iyileştirilmiş moleküller olarak
111 Günümüzde birçok uluslararası bilimsel terminolojide olduğu gibi, bu projemizde de ilâç ile terapötik nükleik asit kavramları eş anlamlı olarak kullanılmaktadır. 112 Kovalent bağlanma adı altında burada sadece kromatografik jel matriksine lipozomların bağlanması kastedilmektedir. Bu yöntem ile ilgili çok sayıda protokol ve tasarım vardır. Projemizde ise yenilik olarak bu tasarıma dayanarak kovalent olarak imobilize edilmiş lipozomal stasyoner faza bir de elektrostatik veya hafif kuvvetler veya hidrofobik etkileşmeleri kullanarak çeşitli katyonlar varlığında ilgili ligandları (ilâç, nükleik asit, protein vs.) bağlayarak elüasyonlarını sağlayarak spesifik olarak bu makromolekülleri saflaştırılması yer almaktadır.

249
diziliş ve konformasyon’a bağlı nükleik asitlerin saflaştırmasındaki yeri de
incelenebilmektedir.
Bu konu özellikle gen aktarımı öncesi yüksek miktarlarda saflaştırılması
gereken plazmid türleri için geçerlidir. Farmasötik formülasyonlarda kullanımları
için büyük ölçüde, miktarda ve saflıkta plazmid DNA’nın üretilmesi insan gen
tedavisi gibi birçok biyoteknolojik uygulamalar için esastır. Özellikle plazmid DNA
saflığı söz konusu olunca, plazmidleri daha yüksek bir dinamik bağlama kapasitesine
sahip ve iyileştirilmiş ve optimize edilmiş kromatografik kolon matrikslerinin
tasarımı çok büyük bir önem kazanır. Şu ana kadar saflştırma amacıyla uygulanmakta
olan poröz matrikslere bir alternativ olarak tasarımımızda geleneksel malzemelere
nazaran iyileştirilmiş hidrodinamik özelliklere sahip membran özellikli matrikslerin
uygulamaya yönelik hususlar vurgulamaktadır. Bu bağlamda, çeşitli nükleik asit
konformasyonları fizikokimyasal ve stereokimyasal olarak ayrıştırabilen kontrollü
DNA presipitasyonlarını gerçekleştirerek tasarımımızda tek ve çift zincirli DNA’ların
saflaştırmasına da yer verilmiştir. Böylece DNA işlevsel yüzeyin yaratılmasında ve
daha sonra bu lipozomal yüzeyinde nükleik asit imobilizasyonu için kullanılabilir.
Genomik ve plazmid DNA veya RNA arasındaki ayrışım kullanılan katyona bağlı
olarak farklı kompaktizasyon globülü meydana geleceğinden mümkün olmaktadır. Bu
tasarım, yine negativ yüklü DNA ile yüksüz (nötr) veya pozitiv yüklü lipidleri çift
valanslı katyonlar (Mg2+, Ca2+, vs.) aracılıyla bir araya getiren elektrostatik
etkileşmelerinin uygulanmasına dayanmaktadır. Diğer yandan, elektrostatik
kuvvetler dışında DNA-lipid oluşumu bazlarla matriksteki ligand-taşıyan gruplar
arasında hidrofobik etkileşmeler sayesinde de meydana gelidiğinden söz konusu
konformasyonlar arası ayrışım ters-fazlı sıvı kromatografisi (Reverse Phase Liquid
Chromatography, RPLC) ile de inclenebilinmektedir. Bu durumda, bağlı olan
polinükleotidler hidrofobik bir özellik olan yükselen hidrofobisiteye göre elüe
olmaktadırlar. Böylece, daha yüksek moleküler ağırlığa sahip nükleik asitler RPLC
kolonunda daha uzun bir süre kalacaklardır. Sonuç olarak, bu tasarım sayesinde tek
sarmallı DNA’yı plazmid DNA’dan ve çift sarmallı DNA’dan saflaştırmak mümkün
olur. Süpersarmallı plazmid DNAlar genelde gen nakli için en uygun malzeme
olduklarından kromatografik yaklaşımların en önemli amacı bu konformasyonu diğer
plazmid konformasyonlardan ayrıştırmak ve saflaştırmaktır.

250
Nükleotid bazlar kromatografik kolon bağlayıcıya (colon linker) şeker, baz
veya fosfat kısmındaki çeşitli pozisyonlarda bağlanmaktadır. Gen transkripsiyonu, ve
translasyounu, virüs-hücre etkileşmeleri, nükleositoplazmik transport ve hücre
bölünmesi gibi çok önemli hücresel olaylarda rol oynayan proteinlerin büyük bir
bölümü öncelikli olarak tek zincirli veya çift zincirli nükleik asit dizilişlerine
bağlanmayı tercih ederler. Bu olaylarda önemli rol oynayan makromoleküller
membrana-bağlı olarak işlev yapmaktadır. Böylece, bu olayların moleküler
mekanizmalarını ortaya çıkarmak, bu kilit moleküllerin saflaştırmasına bağlıdır. Bu
da yeni nükleotid dizilişi ve konformasyona spesifiklik gösteren kromatografik
nükleik asit saflaştırma tasarımlarını zorunlu kılar. Mekanizmaya özgü bir afinite
kromatografi tasarımını şematik olarak geliştirebilmek son derece önemlidir.
İmobilize afinite lipozom-ilaç ve lipozom-nükleik asit kromatografisi konusunda
çalışmalarım yoğun olarak devam etmektedir ve yeni bir araştırma projesi olarak
sunulacaktır.

251
4.7.2. Projenin Devamı Olarak Yapılması Öngörülen Ortak
Çalışmalar
Bu projemle ortaya çıkarılan Taxol®-membran fosfolipid etkileşmelerinden
yola çıkarak burada sunulan modelleri in vivo ortamda denemek. Amacım, bu şekilde
karakterize edilen antikanser ilâç-lipid bileşenlerinin fizikokimyasal parametrelerine
dayanarak bu kontrollü ilâç taşıyıcı tasarımını lipidler aracılıyla gerçekleştirilen in
vivo deneylerde denemek suretiyle iyi ve güvenilir bir model terapötik Taxol®
formülasyonu elde etmek. Bunun için uluslararası düzeyde ortak çalışmalara girmeyi
ümit etmekteyim. Ancak, in vitro ve in vivo ortamda söz konusu formülasyon
çalışmalara girmenin koşulu ilk önce bu formülasyonların fizikokimyasal
karakterizasyonudur ki bunu da projemiz yüzey kimyasal, kolloid ve biyofiziksel
yaklaşımlar kullanarak başarıldığını inanmaktayım. Bu ön verilere dayanarak bu
konuyu çok daha iddialı boyutlara taşımayı arzuluyorum. İleriye yönelik yurt dışı
ortaklarımızla yapmayı planladığımız deneyler kısaca şunlardır:
• Öncelikle, fakültemizde şu ana kadar atık sorunu tehlikesi yüzünden
radyoaktiv bileşenlerle çalışmaların mümkün olmadığından, sözü edilen lipozom-
hücre etkileşmelerinin radyoaktiv lipid markerleri kullanarak incelemeyi
planlamaktayım. Örneğin, lipozomları radyoaktiv [3H]kolesteril ester ve kolesteril
[14C]-oleat işaretleriyle çifte işaretleyerek lipozomların intraselüler akıbeti hakkında
çok önmeli bilgiler elde edilebilir. Radyoaktiv işaretleme yöntemine bir alternativ
olarak piyasada uzun yıllardan beri bulunan lipidlere spesifik floresan işaretler
kullanılabilinir. Ayrıca, lipozomların iç hacminde enkapsüle edilmiş şekilde bulunan
DNA’nın kompaktizasyonu konvansyonel floresan mikroskopisi ile veya konfokal
lazer taramalı mikroskopisi (confocal laser scanning microscopy) ile incelemek çok
önemli bilgiler vermektedir. Örneğin, lipozom lipidlere spesifik olan FM 4-64
uygulandığında bu biyomoleküllerin dinamiği hakkında bilgiler vermektedir
(SÜLEYMANOĞLU, 2002).
• Projemin bir sonraki aşaması hücre kültürlerinin kullanılması ile ilgilidir.
Ancak, bu konu ile ilgili uygulanacak olan protokol hücre türüne göre spesifiklik

252
gösterdiğinden, in situ olarak elde edilecek olan neticeleri çok dikkatlice
değerlendirilmeli, çünkü her hücre türü kendine özgü ilâç internalizasyon kinetiği
göstermektedir. Diğer yandan, hücre kültürlerinin kullanılmasının büyük avantajı bu
hücre kültürlerini piyasadan edinebilme kolaylığı ve yapılmakta olan deneylerin
kontrol edilebilme fırsatıdır. Örneğin, hücre kültürü düzeneğini kullanarak kan ve
kan bileşenlerinin (serum, plazma ve çeşitli proteinler) veya farklı reseptörlerin
lipozom-hücre etkileşmelerine etkilerini ortaya çıkarmayı planlamaktayım.
• Bu projemde, nötr fosfolipidlerlerin yanı sıra negativ yüklü fosfatidilserin-
veya fosfatidilgliserol içeren lipozomların çeşitli hücre kültürleri ve türleri ile yapılan
araştırmalarca farklı hücre-lipozom etkileşmelerinin ve özellikle bağlanma
afinitelerin söz konusu olduğu ortaya çıkarılmıştır. Bu yüzden bu projede yapılan tüm
ön fizikokimyasal ölçümlerini farklı yük taşıyan lipidleri kullanarak lipozom
bileşenlerinin bağlanma afiniteleri, intraselüler salınımları ve hücre içi akıbetlerini
incelemeyi hedefliyorum.
• In vitro protokoller ortam sıcaklığına göre son derece duyarlıdır ve anlamlı
deneylerin yapılabilmesi için ortam sıcaklığının optimize edilmesi gerekmektedir.
Örneğin, bazı hücre kültürleri 4°C en etkili ilâç internalizasyon oranı gösterirken
diğer hücre türleri 37°C’de en yüksek oran gösterirler. Bu bakımdan, in vitro
deneylerini farklı ortam sıcaklığında gerçekleştirerek termodinamik olarak en etkili
transfeksyon ortamını tespit etmeyi düşünmekteyim. Bu kısmen bu projemde de
sirkülatörlü su pompasının alınmasıyla yapılmıştır.
• Ortaklarımızla müşterek olarak yapmayı planladığım projemizin ileriye
dönük boyutlardan biri de elde edilen in vitro verilerini in vivo ortamda denemektir
ve böylece in vitro – in vivo korelasyonlarını incelemektir. Bunun için sıçan ve rat
modellerini kullanarak in vivo ortamda lipozomların klirıns hızı ile ilgili
farmakokinetik veriler elde edilebilinir. Bunun için bröşürü Ek-17’e verilen in vivo
imaging cihazlarının alımı da uygun görülmüştür.
• Bir sonraki aşama olarak, projenin gidişatına göre lipozomların kinetiğini ve
doku dağılımlarını ve intra-organ dağılımlarını radyoaktiv ve/veya floresan işaretleri
kullanarak araştırmayı hedefliyorum.

253
Bu uzun vadeli çalışmalara dahil edilebilmemiz için bir önkoşul olan söz
konusu bileşenlerinin fizikokimyasal karakterizasyonu doğrultusunda, ilâç-lipid
kompleks oluşumunun daha mütevazı ancak çok orijinal bir yaklaşım olan FTIR
spektroskopisi, termodinamik ve yüzey kimyasal ölçümleri kullanarak elde etmiş
bulunmaktayım. Bu ölçümlere dayanarak, daha uzun vadeli ve uluslararası boyutu
olan projelere katılmayı ümit ediyorum. Bu konuda daha önce çalıştığım çok iyi
laboratuvar donanımlı araştırma grupları ile temas edilmiş ve çok olumlu yanıtlar
alınmıştır∇. Böylece, projemin ara raporunda da bahsedildiği gibi çalışmalarımızın
nanoteknolojik boyutuna ulaşılmış olunacak ki, bu Avrupa Birliğinin 7ci Çerçeve
Programında proje öncelik sıralamasında ilk yerlerde bulunan bir konu olarak bize
uluslararası projelere de dahil edilmemizi sağlayacaktır ∏.
4.7.3. Öncü Hipotez
Projemde geliştirilen hipotez ile ilgili burada sadece genel bilgiler
verilmektedir. Bu hipotezin dayanaklarına burada geniş olarak yer verildiği takdirde
bu raporun hacmi çok fazla artacaktır. Ancak, buradaki kısa bilgiler bile bu hipotezin
iyi anlaşılmasına yeterli olacağı kanısındayım.
Bu projemde geliştirilen Taxol®-hücre membranı etkileşme modellerine
dayanarak bu ilâcın yeni etki mekanizmalarının ortaya çıkarılması hedeflenmiştir. Bu
bağlamdai bu antikanser ilâcın membran akışkanlığını düzenleyerek başka bir etki
mekanizmasının daha mümkün olabileceği öngörülmektedir. Bunun için, bu ilâcın
sitotoksisitesini wild-type ve multidrug resistant (MDR) hayvan hücrelerinde
inceleyerek önemli kemoduyarlılık mekanizmalarının ortaya çıkarabilme imkânlarına
bağlıdır. Bu bağlamda, hipotez modelini oluşturmak için önce literatürde bulunan
mevcut bulgulara dayanarak ve ileride bu tür düzeneklerin Fakültemizde de
kurulmasıyla bu tür modellerin geçerliliğini inceleme fırsatı bulunacaktır. Örneğin,
hipoteze göre, Taxol®’un P-glikoprotein (Şekil 4.28) üzerinden değil de, farklı bir
mekanizma ile kemoduyarlılık sitotosisitenin meydana gelmesinde rol oynamaktadır.
∇ Bu konu ile ilgili çok kapsamlı uluslararası projem Ek-16’da verilmiştir. ∏ Projemizin uluslararası boyutu ve bu konuda hedeflerimiz ve beklentilerimiz ile ilgili ayrıntılı gerekçelere ayrıca ara raporumuzda yer verilmişti.

254
Bu modeli test etmek için reserpine ve verapamil gibi birçok kemoduyarlılığı
meydana getiren molekül kullanılabilir. Eğer bunların uygulanması, P-gp
bulundurmayan karsinom hücrelerindeki Taxol®’un birikmesine yol açtıkları tespit
edilebilirse bu teklif edilen mekanizmanın bir kanıtı olarak sunulabilecektir. Çünkü
bu bulgular, kemoduyarlılık yolları üzerinden etki gösteren ve P-gp’inden bağmsız
olarak ortaya çıkan membran geçirgenliğinin artmasına ilişkin başka verilele de
desteklenecektir. Ancak, bunu ortaya çıkarmak çok iyi çalışılmış Taxol®-
biyomembran etkileme incelemelerine dayanmaktadır. Bunların bir başlangıç
nktasını bu proje ile başlamış bulunmkatayız. Bu projemde vurgulanan hususlar
doğrultusunda membran akışkanlığını arttıran bazı moleküller gibi kemoduyarlılığı
arttıran moleküller (chemosensitizers) de doza ve konsantrasyona bağlı olarak söz
konusu akışkanlığının artmasına neden olmaktadır. Bu olgu, örneğin DPH
polarizasyon gibi floresan spektroskopik yöntemlerle hem lipozomlarda hem d ewild-
type ve MDR hayvan ve insan hücre membranları üzerinde test edilebilir. Böylece, bu
tür chemosntitizer moleküllerin lipid katmanlatın geçirgenliğini arttırıcı etkileri ile
anti-kanser ilâçlar karşı başlattıkları sitotoksiste arasında önemli korelasyonlar
bulunmaktadır. Sonuç olarak, bu hipotezde chemosentisizerler aracılıyla
biyomembranların yapılarında akışkanlığın değişime uğraması ve geçirgenliklerinin
artması gibi olayların, bu tür hücrelerde Taxol®’un sitotoksisitesini ortaya
çıkmasında önemli rol oynayabildiklerini teklif edilmeltedir. Bu konuda yapılacak lan
ek incelemelerin yeni tedavi profillerinin de geliştirilmesine katkı sağalayacaktır
(Şekil 4.29).

255
Şekil 4.28. P-glikoprotein (P-gp) membran üzerinden hücre dışına ilâç
moleküllerini pompalayarak kemotedavilerinin etkinliğini önemli ölçüde
düşürmetedir.

256
Şekil 4.29. Antikanser ilâç-tümör hücre etkileşmeleri.

257
4.7.4. Fakültemizde Bundan Sonraki Proje Hedeflerim
Fakültemizde göreve başladığım tarihten hemen sonra, 04.Şubat.2004 günü
bir Fakülte toplantısı düzenlenmiş, bu toplantıda o zamanki yönetim tarafından
tarafıma ve beraber çalıştığım diğer 2 uzman arkadaşıma mevcut laboratuvar
durumumuzun çok kötü olduğundan somut projeler üretmemiz konusunda talimatlar
verilmiştir. Ayrıca da, bu projelerde suiistimalleri engellemek amacıyla kesinlikle
öğretim üyelerinin yazılmaması gerektiği talimatı da tarafımıza verilmiştir. Bana
verilen bu talimatları aynen uygulayıp bu proje hazırlanmıştır. Fakültemizdeki tüm
kürsülerimize hizmet veren Merkez Laboratuvarın mevcut altyapısını güçlendirmek
amacıyla, cihaz alımı ile sonuçlanabilecek bir proje olmasına özen gösterilmiştir.
Cihaz seçiminde de son derece dikkatli davranılmıştır. Her kürsünün istifade
edebileceği bir cihaz projesi hazırlanmıştır. Bu benim Türkiye’de ilk projem
olduğundan daha kısa süreli bir proje olarak hazırlanıp, bu projede elde edilen
bulgulara dayanarak çok daha kapsamlı, uzun vadeli, yüksek bütçeli ve uluslararası
katılımlı bir proje daha hazırlanmıştır (Bk.: Ek-16). Bunun için tarafıma yurt içinden
ve yurt dışından yapılan ortak çalışma başvurularını da (Bk.: Ek-15) değerlendirip
ulusal ve uluslararası proje sponsorlarına başvurulması uygun görülmekteyim. Ancak
bu şekilde, uzun vadeli bir proje desteği ile şu andaki durum (Bk.: Ek-1 ve Ek-2)
iyileştirilebilecektir. Bu doğrultuda, bu projemle alınan Langmuir-Blodgett cihazına
eklenebilecek bazı cihazların alımı ile bu çalışmalarımın niteliği yükseltilebilecektir.
Bu bağlamda, daha 2 sene önce bu cihaza eklemek üzere yine yüzey analizlerde yoğun
olarak kullanılan Atomik Kuvvet Mikroskopu (Atomic Force Microscopy-AFM) veya
yüzeylerde faz değişimlerinin araştırılmasında kullanılan Brewster Angle Microscopy
(BAM) sistemlerinin alınması ile ilgili projeler hazırladım. Ancak, söz konusu
mikroskopi sistemleri temiz bir ortama bağlı olarak çalıştırılmaktadır. Oysa
Fakültemizde henüz daha temiz (tozsuz) bir odamız yoktur. Bu yüzden bu projelerimi
böyle bir odanın yapımından sonrası için ertelenmiştir. O zamana kadar Fakültemize
daha faydalı olabilecek ve çok orijinal alternativler üzerinde çalıştım ve bundan böyle
geliştirmek istediğim laboratuvar donanımını şöyle sıralamaktayım. Tüm ilgili
bröşürler sırasıyla E-17’de verilmiştir.
İlk önce, bu projemle alınan Langmuir-Blodgett sistemine ek olarak bir Dip
Coaters parçalarının alınması uygun olur. Söz konusu parçalar, çalışılan düzenek

258
çözeltilerine son derece hassas olarak substratların uygulanmasını sağlamaktadır.
Böyle bir ek sistemin monte edilmesi çalışmalarımızdaki hassasiyeti arttıracaktır.
Çalışmalarımıza termodinamik boyut kazandırmak amacıyla termal analiz
cihazlarının getirilmesi için gereken girişimlerde bulunulmuştur. Bu doğrultuda bir
Isothermal Titration Calorimeter cihazın alınması için başka bir proje
hazırlanacaktır. Söz konusu cihaz şu ana kadar kullandığım Differential Scanning
Calorimetry cihazlarından çok önemli üstünlüğü vardır. Örneğin, Isotehrmal
Titration Calorimetry ile ligand bağlama kinetiği ölçülmektedir ve bu çok önemli
bilgiler vermektedir.
Bir sonraki proje hedefim bir Confocal Fluorescence Microscopy
sistemidir. Söz konusu mikroskopi sistemi, bu proje ile alınan Langmuir-Blodgett tek
katman sistemine inverted versiyonu olarak eklenebilmektedir. Bu sisteme ayrıca
da Epifluorescence Microscopy sistemi de eklenebilmektedir. Bu
mikroskopilerin eklenmesi model tek katman membran çalışmalarında son derece
önemli bilgiler verecektir, çünkü bu sistemler uygun floresan işaretlerin yardımıyla
doğrudan fosfolipid akışkanlığının incelenmesini mümkün kılacaktır. Bunun bilimsel
gerekçeleri de şöyle sıralanabilir. Daha 1960lı yıllarda Almanya’da Ruska’nın
keşfettiği elektron mikroskopisi hücre biyolojisinde çok önemli rol oynamıştır.
Örneğin, virüs ve fajların bakteri yüzeyindeki yapılarını incelenmesini mümkün
kılmıştır ve birçok intraselüler organellin yapısını aydınlatmıştır. Ultrasantrifüjün
kullanıma girmesiyle bu mikroskopi yöntemi moleküler hücre biyolojisinin ve
biyokimyanın gelişmesine olağanüstü bir katkıda bulunmuştur. Ancak daha sonraları
araştırıcılar tarafından bazı uygulamaları kuşku uyandırmıştır. Örneğin, bu
mikroskopi sisteminde kullanılan sert muameleler (-94°C ortamı ve OsO4 ile
fiksaksiyon işlemleri) birçok biyolojik molekülleri olumsuz etkilemektedir. Çünkü
doğada böyle ortamlar zaten yoktur. Her nekadar elektron mikroskopisinin TEM
(Transmission Electron Microscopy) versiyonu özellikle boyut analizi için günümüzde
de kullanılıyorsa da, bu mikroskopilerle elde edilen görüntüler bu sebeplerden dolayı
artefakttır (yapaydır). Buna karşılık, floresan mikroskopisinin çok önemli üstünlüğü
bulunmaktadır. Örneğin, Gren Fluorescence Protein (GFP)-plazmidlerle klonlanan ve
hücresel ekspresyonu gerçekleştirilen bir proteinin söz konusu hücreyi hiç
öldürmeden hücre kültürü ortamında protein dinamiğini live cell imaging olarak

259
görüntülemek mümkündür. Anlaşılacağı üzere yaşayan hücrelerle çalışmak çok
önemlidir. Ayrıca da, BODIPY türü gibi lipid floresan işaretler kullanarak fosfolipid
veziküllerin (lipozomların) (Bk.: Ek-9) şeklini ve akışkanlığını incelemek mümkün
olmaktadır. Şimdiye kadar adı geçen cihazlar Langmuir-Blodgett tek katman
sistemine doğrudan eklenebilinmektedir. Bu sistem ile elde edilen veriler ve
geliştirilen modeller daha sonra in vivo ortamda uygulanabilmelidir ve geçerlilikleri
ispat edilmelidir.
Bu bakımdan, Fakültemizde bir hücre kültürü düzeneğinin kurumasından
sonraki proje amacım Raman Mikroskopi sisteminin getirilmesi. Bunu da, uzun
vadeli proje desteği ve Avrupa’daki eski hocalarımın da projelere katılımıyla bir İlâç-
Hücre Etkileşmeleri konulu uluslararası proje kapsamında getirilmesini uygun
görmekteyim, çünkü böyle bir sistem 220 000 Avro’luk bir desteği de gerektirecektir.
Ancak bu pahalı sistemin çok önemli avantajları vardır. Örneğin, her ne kadar
floresan mikroskopisi yaşayan hücredeki yapıları görüntülüyor olsa da bu yöntemde
ortaya çıkan “photobleaching” olayı tek başına hücreleri öldürebilmektedir. Bu
yüzden, bir yapı görüntülenir görüntülenmez hemen başka bir bölgeye mikroskopi
okularının yönlendirilmesi gerekmektedir. Dolayısıyla da deneyimli personele ihtiyaç
vardır. Kanımca, buna alternativ olarak böyle bir sorunu olmayan Raman
Mikroskopisi teklif edilmelidir.
Buraya kadar adı geçen cihazların her biriyle ilgili uluslararası tecrübem
bulunmaktadır ve herhangi bir eğitim için zaman kaybedilmeyecektir (Ek-18). Ancak,
tüm bu projelerin başarıyla tamamlanması Fakültemizdeki statümün özerkliğine ve
suiistimallerin engellenebilmesine bağlıdır. Herkesin projelere müdahale ettiği ve
suiistimal etmeye çalıştığı bir noktada zaten Avrupa’daki ortaklarımın bunlara
katkıda bulunmaları söz konusu olamaz.

260
5. SONUÇ
Bu projemde, kemoterapilrde yoğun olarak kullanılan Taxol®’un (Paclitaxel’in)
model olarak uygulanan fosfolipid yapılarıyla etkileşmeleri moleküler düzeyde
incelenmiştir. Yüzeyin fizikokimyasal karakterizasyonu ve farklı ilâç-lipid
bileşimlerinin yapı ve termodinamik özelliklerinin aydınlatılması için Langmuir-
Blodgett tek katman, Fourier Transfrom Infrared (FTIR) Spektroskopi ve
Diferansiyel Taramalı Kalorimetrik ölçümlerinde yararlanılmıştır. Örneğin
hazırlanmasının, lipid türünün, ilâç konsantrasyonunun gözlemlenen bu özellikler
üzerinde belirleyici etkileri vardır. Fosfatidilkolin tarafından Taxol®’un tanınması,
lipid baş gruplarınca yüzeyin tanınması, başlangıç, değişim öncesi ve ana faz değişim
dereceleri ve açil zincir gruplaşması gibi sonuçlara yol açarak model membran
yüzeylerinin fiziksel parametreleri üzerinde sıvılaştırıcı ve akışkanlığı arttırıcı etkileri
olduğu ortaya çıkarılmıştır.
Projemdeki vurgu Taxol®’un model hücre membranlar üzerindeki etkileri
hakkında yoğunlaşmış olmasına rağmen, tümör tedavisi denemelerinde hayati önem
taşıyan ilâcın etki mekanizmasındaki biyofiziksel olguların anlaşmasına katkı
sağlayabilecek sonuçlar elde edildiğini ümit etmekteyim. Bu iki yaklaşımın doğasına
uygun olarak kullanılan deneysel tasarıma uygun düşen bir öncü hipotez ve öneriler
de ayrıca tanımlanmıştır.
Projemi de önceden öngörülen zaman içerisinde gecikme yaşanmaksızın
tamamlamış bulunmaktayım. Benden kaynaklanan herhangi bir gecikme söz konusu
değildir. Tüm eksikliklere ve özellikle de temiz (tozsuz) ortamın olmamasına rağmen,
projemin başarıyla tamamlanmış olması da buaradaki neticelerin, bu raporumda yer
verdiğim ve daha önce böyle tozsuz ortamda yapmış olduğum ölçümlerle
kıyaslamalarından da görülmektedir. Sonuç olarak, yapılması son derece zor olan
ölçümler tamamlanmış olup, yeni bir düzenek Fakültemize başarıyla kurulmuştur ve
sadece Fakültemizde değil, Ankara’da bulunan diğer araştırmacılara da hizmet
vermek üzere hazır tutulmaktadır.

261
Ekler Listesi
Ek-1: Öğretim Elemanı Başına Düşen Makale Sayısına Göre Üniversite Sıralamasındaki Üniversitemizin Yeri Ek-2: Bologna Süreci Durum Değerlendirme Raporu, 2005-2007 Ek-3: Tübitak Avrupa Birliği 7. Çerçeve Programı: Nanobilimler, Nanoteknolojiler, Malzemeler ve Yeni Üretim Teknolojileri Ek-4: Gazi Üniversitesinde Nanotıp Üssünün Kurulması ile ilgili Karar Ek-5: Taxol® (Paclitaxel) ile ilgili Güncel Referanslar Ek-6: Gen Aktarımı ile ilgili Güncel Referanslar Ek-7 Taxol® (Paclitaxel’in) Fizikokimyasal Özellikleri Ek-8 Taxol®’un Sentezi Ek-9 Lipozom Hazırlama Yöntemleri Ek-10 Tek Katmanlı Lipozomların Hacmi, Çapı, Sayıları, Alanları ve Lipid Ağırlığı Arasındaki İlişkiyi Gösteren Bir Nomogram Ek-11 NIMA Technology Ltd.’in ürettiği Langmuir-Blodgett cihazları Ek-12 Langmuir-Blodgett Tek Katman Cihazın Teknik Özellikleri
Ek-13 Langmuir-Blodgett Yüzey Kimyasal ve Kolloid Düzeneklerin Teorisi
Ek-14 Kemoterapilerde Kullanılmakta Olan Diğer Güncel Anti-Kanser İlâçlar
Ek-15 Tarafıma Yurt İçinden Ve Yurtdışından Gelen Ortak Çalışma Başvuruları
Ek-16 Avrupa Birliği Destekli Proje Çerçevesine Hazırladığım Çok Katılımlı
Uluslararası Ve Uzun Vadeli Proje
Ek-17 Fakültemizde Bundan Sonraki Proje Hedeflerim
Ek-18 Projemle İlgili Tüm Evraklar ve Faturalar
Ek-19 Özgeçmiş

262
Ek-5: Taxol® (Paclitaxel) İLE İLGİLİ GÜNCEL REFERANSLAR
Abernethy DR. Pharmacokinetics and pharmacodynamics of amlodipine.
Cardiology. 1992;80 Suppl 1:31-6.

263
Adv Cancer Res. 2007;98:149-90 ve Song RX. Membrane-initiated steroid
signaling action of estrogen and breast cancer. Semin Reprod Med. 2007
May;25(3):187-97.
Advances in Molecular Oncology (Advances in Experimental Medicine and
Biology) (Advances in Experimental Medicine and Biology), by Fabrizio d'Adda di
Fagagna (Editor), Susanna Chiocca (Editor), Fraser McBlane (Editor), Ugo Cavallaro
(Editor); Springer; 1 edition (July 12, 2007).
AMA Council Report. Guidelines for Handling Parenteral Antineoplastics. J.
Amer. Med. Assoc. 1985; 253 (11): 1590-1592.
American Society of Hospital Pharmacists Technical Assistance Bulletin on
Handling Cytotoxic and Hazardous Drugs. Am J Hosp Pharm 1990; 47:1033-1049.
Aster RH. Drug-induced immune thrombocytopenia: an overview of
pathogenesis.
Barbato F, di Martino G, Grumetto L, La Rotonda MI. Prediction of drug-
membrane interactions by IAM-HPLC: effects of different phospholipid stationary
phases on the partition of bases. Eur J Pharm Sci. 2004 Jul;22(4):261-9.
Baritaki S, Apostolakis S, Kanellou P, Dimanche-Boitrel MT, Spandidos DA,
Bonavida B. Reversal of tumor resistance to apoptotic stimuli by alteration of
membrane fluidity: therapeutic implications.
Benie AJ, Blume A, Schmidt RR, Reutter W, Hinderlich S, Peters T.
Characterization of ligand binding to the bifunctional key enzyme in the sialic acid
biosynthesis by NMR: II. Investigation of the ManNAc kinase functionality. J Biol
Chem. 2004 Dec 31;279(53):55722-7.
Berquand A, Fa N, Dufrene YF, Mingeot-Leclercq MP. Interaction of the
macrolide antibiotic azithromycin with lipid bilayers: effect on membrane
organization, fluidity, and permeability. Pharm Res. 2005 Mar;22(3):465-75.

264
Berquand A, Mingeot-Leclercq MP, Dufrene YF. Real-time imaging of drug-
membrane interactions by atomic force microscopy. Biochim Biophys Acta. 2004
Aug 30;1664(2):198-205.
Bhatti M, Yahioglu G, Milgrom LR, Garcia-Maya M, Chester KA, Deonarain MP.
Targeted photodynamic therapy with multiply-loaded recombinant antibody
fragments. Int J Cancer. 2007 Oct 31.
Bianco R, Damiano V, Gelardi T, Daniele G, Ciardiello F, Tortora G. Rational
combination of targeted therapies as a strategy to overcome the mechanisms of
resistance to inhibitors of EGFR signaling. Curr Pharm Des. 2007;13(33):3358-67.
Biochem Pharmacol. 1999 Mar 1;57(5):503-10.
Biochim Biophys Acta. 2000 Nov 23;1508(1-2):210-34.
Bordi F, Cametti C, Motta A, Diociaiuti M, Molinari A. Interactions of
anthracyclines with zwitterionic phospholipid monolayers at the air-water interface.
Bioelectrochem Bioenerg. 1999 Oct;49(1):51-6.
Byrnes RW, Antholine WE, Petering DH. Interactions of 1,10-phenanthroline and
its copper complex with Ehrlich cells. Free Radic Biol Med. 1992;12(6):457-69.
Calcagnile O, Gisselsson D. Telomere dysfunction and telomerase activation in
cancer--a pathological paradox? Cytogenet Genome Res. 2007;118(2-4):270-6.
Caldwell GW, Masucci JA, Evangelisto M, White R. Evaluation of the immobilized
artificial membrane phosphatidylcholine. Drug discovery column for high-
performance liquid chromatographic screening of drug-membrane interactions.
Capobianco JO, Zakula D, Frost DJ, Goldman RC, Li L, Klein LL, Lartey PA.
Cellular accumulation, localization, and activity of a synthetic cyclopeptamine in
fungi. Antimicrob Agents Chemother. 1998 Feb; 42(2): 389-93.

265
Carrozzino JM, Khaledi MG. Interaction of basic drugs with lipid bilayers using
liposome electrokinetic chromatography. Pharm Res. 2004 Dec;21(12):2327-35.
Castaing M, Loiseau A, Dani M. Designing multidrug-resistance modulators
circumventing the reverse pH gradient in tumours. J Pharm Pharmacol. 2001
Jul;53(7):1021-8.
Castaing M, Loiseau A, Dani M. Thermal dependence of multidrug-resistant-
modulator efficiency: a study in anionic liposomes. J Pharm Pharmacol. 2000
Oct;52(10):1171-8.
Castaing M, Loiseau A, Djoudi L. Effects of cholesterol on dye leakage induced by
multidrug-resistance modulators from anionic liposomes. Eur J Pharm Sci. 2003
Jan;18(1):81-8.
Castaing M, Loiseau A, Mulliert G. Interactions between verapamil and neutral
and acidic liposomes: effects of the ionic strength. Biochim Biophys Acta. 2003 Apr
1;1611(1-2):107-14.
Castaing M, Loiseau A, Mulliert G. Multidrug resistance modulator interactions
with neutral and anionic liposomes: membrane binding affinity and membrane
perturbing activity. J Pharm Pharmacol. 2005 May;57(5):547-54.
Cauchetier E, Loiseau PM, Lehman J, Rivollet D, Fleury J, Astier A, Deniau M,
Paul M. Characterisation of atovaquone resistance in Leishmania infantum
promastigotes.
Cereijido M, Contreras RG, Flores-Benítez D, Flores-Maldonado C, Larre I, Ruiz
A, Shoshani L. New diseases derived or associated with the tight junction. Arch Med
Res. 2007 Jul;38(5):465-78.

266
Chakraborty H, Banerjee R, Sarkar M. Incorporation of NSAIDs in micelles:
implication of structural switchover in drug-membrane interaction. Biophys Chem.
2003 May 1;104(1):315-25.
Charras GT, Coughlin M, Mitchison TJ, Mahadevan L. Life and Times of a Cellular
Bleb. Biophys J. 2007 Oct 5.
Chen WM, Zhang PL, Wu B, Zheng QT. Studies on the chemical constituents of
Taxus yunnanensis. Yao Hsueh Hsueh Pao 1991;26(10):747-54.
Chen YJ, Chen CY, Shen YC, New taxoids from the seeds of taxus chinensis. J Nat
Prod 1999 Jan;62(1):149-51.
Cheung AL, Deng W. Telomere dysfunction, genome instability and cancer. Front
Biosci. 2008 Jan 1;13:2075-90.
Chidgey M, Dawson C. Desmosomes: a role in cancer? Br J Cancer. 2007 Jun
18;96(12):1783-7.
Chiquero MJ, Perez-Victoria JM, O'Valle F, Gonzalez-Ros JM, del Moral RG,
Ferragut JA, Castanys S, Gamarro F. Altered drug membrane permeability in a
multidrug-resistant Leishmania tropica line. Biochem Pharmacol. 1998 Jan
15;55(2):131-9.
Clinical Oncological Society of Australia. Guidelines and Recommendations for
Safe Handling of Antineoplastic Agents. Med J Australia 1983; 1:426-428.
Deo N, Somasundaran T, Somasundaran P. Solution properties of amitriptyline
and its partitioning into lipid bilayers. Colloids Surf B Biointerfaces. 2004 Apr
1;34(3):155-9.
Dingemans AM, Pinedo HM, Giaccone G. Clinical resistance to topoisomerase-
targeted drugs. Biochim Biophys Acta. 1998 Oct 1;1400(1-3):275-88.

267
Dittmer DP, Krown SE. Targeted therapy for Kaposi's sarcoma and Kaposi's
sarcoma-associated herpesvirus. Curr Opin Oncol. 2007 Sep;19(5):452-7.
Dubielecka PM, Trusz A, Diakowski W, Grzybek M, Chorzalska A, Jazwiec B,
Lisowski M, Jezierski A, Sikorski AF. Mitoxantrone changes spectrin-
aminophospholipid interactions. Mol Membr Biol. 2006 May-Jun;23(3):235-43.
F. Cavalli H. H. Hauser ve S. B. Kaye (Eds.): Textbook of Medical Oncology,
Taylor and Francis, New York, 2004.
Fa N, Ronkart S, Schanck A, Deleu M, Gaigneaux A, Goormaghtigh E, Mingeot-
Leclercq MP. Effect of the antibiotic azithromycin on thermotropic behavior of DOPC
or DPPC bilayers. Chem Phys Lipids. 2006 Oct;144(1):108-16.
Fan J, Tang g, Shu G, Xu X, On the conservation and regeneration of Taxus
resources. Chung Kuo Chung Yao Tsa Chih. 1996 Jul;21(7):389-91, 446.
Fardel O, Payen L, Sparfel L, Vernhet L, Lecureur V. Drug membrane transporters
in the liver: regulation of their expression and activity. Ann Pharm Fr. 2002
Nov;60(6):380-5.
Ferte J. Analysis of the tangled relationships between P-glycoprotein-mediated
multidrug resistance and the lipid phase of the cell membrane. Eur J Biochem. 2000
Jan;267(2):277-94.
Fox CB, Horton RA, Harris JM. Detection of drug-membrane interactions in
individual phospholipid vesicles by confocal Raman microscopy. Anal Chem. 2006
Jul 15;78(14):4918-24.
Fresta M, Furneri PM, Mezzasalma E, Nicolosi VM, Puglisi G. Correlation of
trimethoprim and brodimoprim physicochemical and lipid membrane interaction
properties with their accumulation in human neutrophils. Antimicrob Agents
Chemother. 1996 Dec;40(12):2865-73.

268
Fresta M, Panico AM, Bucolo C, Giannavola C, Puglisi G. Characterization and in-
vivo ocular absorption of liposome-encapsulated acyclovir. J Pharm Pharmacol. 1999
May;51(5):565-76.
Fukushima M, Takeda J, Fukamiya N, Okano M, Tagahara K, Zhang SX, Zhang
DC, Lee KH. A new taxoid, 19-acetoxytaxagifine, from Taxus chinensis. J Nat Prod
1999 Jan;62(1):140-2.
Gallois L, Fiallo M, Garnier-Suillerot A. Comparison of the interaction of
doxorubicin, daunorubicin, idarubicin and idarubicinol with large unilamellar
vesicles. Circular dichroism study. Biochim Biophys Acta. 1998 Mar 6;1370(1):31-40.
Garman KS, Nevins JR, Potti A. Genomic strategies for personalized cancer
therapy. Hum Mol Genet. 2007 Oct 15;16 Spec No. 2:R226-32.
Gasparini P, Sozzi G, Pierotti MA. The role of chromosomal alterations in human
cancer development. J Cell Biochem. 2007 Oct 1;102(2):320-31.
Goldenberg DM. "Targeted therapy with monoclonal antibodies: the new
generation of pharmaceuticals". Conf Proc IEEE Eng Med Biol Soc. 2006; 21-2.
Hamill OP, McBride DW Jr. Induced membrane hypo/hyper-mechanosensitivity:
a limitation of patch-clamp recording. Annu Rev Physiol. 1997;59:621-31.
Hendrix MJ. A morphological characterization of melanoma tumor cell
interaction with a basement membrane model in the absence and presence of an
anticancer agent. Scan Electron Microsc. 1984; (Pt 4): 1973-82.
Hideshima T, Anderson KC. Preclinical studies of novel targeted therapies.
Hematol Oncol Clin North Am. 2007 Dec;21(6):1071-91.
Howard R. Petty, Molecular Biology of Membranes-Structure and Function,
Plenum Press, New York and London, 1993, pp. 353-377.

269
Int J Parasitol. 2002 Jul;32(8):1043-51.
J Chromatogr A. 1998 Mar 27;800(2):161-9.
J Chromatogr A. 2001 Apr 13;913(1-2):123-31.
J Pharm Sci. 1998 Aug;87(8):960-6.
Johansson E, Engvall C, Arfvidsson M, Lundahl P, Edwards K. Development and
initial evaluation of PEG-stabilized bilayer disks as novel model membranes. Biophys
Chem. 2005 Feb 1;113(2):183-92.
Jones RB, et al: Safe Handling of Chemotherapeutic Agents: A Report from the
Mount Sinai Medical Center. Ca-A Cancer Journal for Clinicians 1983; (Sept/Oct)
258-263.
Korfel A, Thiel E. Targeted therapy and blood-brain barrier. Recent Results
Cancer Res. 2007;176:123-33.
Kozin SV, Gerweck LE. Related. Cytotoxicity of weak electrolytes after the
adaptation of cells to low pH: role of the transmembrane pH gradient. Br J Cancer.
1998 May; 77 (10): 1580-5.
Larsen AK, Lametsch R, Elce JS, Larsen JK, Thomsen B, Larsen MR, Lawson MA,
Greer PA, Ertbjerg P. Genetic disruption of calpain correlates with loss of membrane
blebbing and differential expression of RhoGDI-1, cofilin and tropomyosin. Biochem
J. 2007 Dec 12.
Li Y, Liu J, Pan D, Hopfinger AJ. A study of the relationship between cornea
permeability and eye irritation using membrane-interaction QSAR analysis. Toxicol
Sci. 2005 Dec;88(2):434-46.

270
Liang J, Kingston DG. Two new taxane diterpenoids from Taxus mairei. J Nat
Prod 1993 Apr;56(4):594-9.
Liang JY, Min ZD, Iinuma M, Tanaka T, Mizuno M, Two new antineoplastic
diterpenes from Taxus mairei. Chem Pharm Bull (Tokyo) 1987 Jun;35(6):2613-4.
Liang Y, Liu LR, Zhang QQ. Structure and degradation property of the PVA-
collagen complex drug membrane. Zhongguo Yi Xue Ke Xue Yuan Xue Bao. 2004
Feb;26(1):18-23.
Liu H, Carter GT, Tischler M. Immobilized artificial membrane chromatography
with mass spectrometric detection: a rapid method for screening drug-membrane
interactions. Rapid Commun Mass Spectrom. 2001;15(17):1533-8.
Liu XY, Nakamura C, Yang Q, Kamo N, Miyake J. Immobilized liposome
chromatography to study drug-membrane interactions. Correlation with drug
absorption in humans. J Chromatogr A. 2002 Jun 28;961(1):113-8.
Liu XY, Yang Q, Kamo N, Miyake J. Effect of liposome type and membrane
fluidity on drug-membrane partitioning analyzed by immobilized liposome
chromatography.
Lundquist A, Engvall C, Boija E, Kurtovic S, Chattopadhyaya J, Hagglund CL,
Lundahl P. Interactions of drugs and an oligonucleotide with charged membranes
analyzed by immobilized liposome chromatography. Biomed Chromatogr. 2006
Jan;20(1):83-7.
Luxo C, Jurado AS, Custodio JB, Madeira VM. Toxic effects of tamoxifen on the
growth and respiratory activity of Bacillus stearothermophilus. Toxicol In Vitro. 2001
Aug-Oct;15(4-5):303-5.
Ma DW. Lipid mediators in membrane rafts are important determinants of
human health and disease. Appl Physiol Nutr Metab. 2007, Jun;32(3):341-50.

271
Ma JY, Barger MW, Ma JK, Castranova V. Inhibition of respiratory burst activity
in alveolar macrophages by bisbenzylisoquinoline alkaloids: characterization of drug-
cell interaction. Exp Lung Res. 1992 Nov-Dec;18(6):829-43.
Ma W, Stahlhut RW, Adams TL, Park GL, Evans WA, Blumenthal SG, Gomez GA,
Nieder MH, Hylands PJ. Yunnanxane and its homologous esters from cell cultures of
Taxus chinensis var. mairei. J Nat Prod 1994 Sep;57(9):1320-4.
Majd S, Mayer M. Hydrogel stamping of arrays of supported lipid bilayers with
various lipid compositions for the screening of drug-membrane and protein-
membrane interactions. Angew Chem Int Ed Engl. 2005 Oct 21;44(41):6697-700.
Chakraborty H, Roy S, Sarkar M. Interaction of oxicam NSAIDs with DMPC
vesicles: differential partitioning of drugs. Chem Phys Lipids. 2005 Dec;138(1-2):20-
8.
Malheiros SV, Brito MA, Brites D, Meirelles NC. Membrane effects of
trifluoperazine, dibucaine and praziquantel on human erythrocytes. Chem Biol
Interact. 2000 May 1;126(2):79-95.
Mao JJ, Sun J, He ZG. Biopartitioning micellar chromatography: its application in
evaluating drug membrane transport and activity. Yao Xue Xue Bao. 2005
Oct;40(10):865-70.
Martínez MC, Kunzelmann C, Freyssinet JM. Plasma membrane remodelling and
cell stimulation. Med Sci (Paris). 2004 Feb;20(2):189-95.
Mavromoustakos T, Zoumpoulakis P, Kyrikou I, Zoga A, Siapi E, Zervou M,
Daliani I, Dimitriou D, Pitsas A, Kamoutsis C, Laggner P. Efforts to understand the
molecular basis of hypertension through drug:membrane interactions. Curr Top Med
Chem. 2004;4(4):445-59.

272
Meng QC, Johansson JS, Eckenhoff RG. Chromatographic approach for
determining the relative membrane permeability of drugs. J Chromatogr B Analyt
Technol Biomed Life Sci. 2002 Jul 5;774(1):89-95.
Middleton DA, Reid DG, Watts A. Combined quantitative and mechanistic study
of drug-membrane interactions using a novel 2H NMR approach. J Pharm Sci. 2004
Feb;93(2):507-14.
Min ZD, Jiang H, Liang JY. Studies on the taxane diterpenes of the heartwood
from Taxus mairei. Yao Hsueh Hsueh Pao 1989;24(9):673-7.
Molecular Biology of Cancer: Mechanisms, Targets, and Therapeutics,by Lauren
Pecorino, Oxford University Press, USA; 1 edition (May 21, 2005).
Montenez JP, Van Bambeke F, Piret J, Brasseur R, Tulkens PM, Mingeot-Leclercq
MP. Interactions of macrolide antibiotics (Erythromycin A, roxithromycin,
erythromycylamine [Dirithromycin], and azithromycin) with phospholipids:
computer-aided conformational analysis and studies on acellular and cell culture
models. Toxicol Appl Pharmacol. 1999 Apr 15;156(2):129-40.
National Study Commission on Cytotoxic Exposure - Recommendations for
Handling Cytotoxic Agents. Available from Louis P. Jeffrey, ScD, Chairman, National
Study Commission on Cytotoxic Exposure. Massachusetts College of Pharmacy and
Allied Health Sciences, 179 Longwood Avenue, Boston, Massachusetts, 02115.
Nusbaum P, Lainé C, Seveau S, Lesavre P, Halbwachs-Mecarelli L. Early
membrane events in polymorphonuclear cell (PMN) apoptosis: membrane blebbing
and vesicle release, CD43 and CD16 down-regulation and phosphatidylserine
externalization. Biochem Soc Trans. 2004 Jun;32(Pt3):477-9.
Oberdoerffer P, Sinclair DA. The role of nuclear architecture in genomic
instability and ageing. Nat Rev Mol Cell Biol. 2007 Sep;8(9):692-702.

273
Ornskov E, Gottfries J, Erickson M, Folestad S. Experimental modelling of drug
membrane permeability by capillary electrophoresis using liposomes, micelles and
microemulsions. J Pharm Pharmacol. 2005 Apr;57(4):435-42.
OSHA Work-Practice Guidelines for Personnel Dealing with Cytotoxic
(Antineoplastic) Drugs. Am J Hosp Pharm 1986; 43:1193-1204.
Pajak B, Molnar J, Engi H, Orzechowski A. Preliminary studies on phenothiazine-
mediated reversal of multidrug resistance in mouse lymphoma and COLO 320 cells.
In Vivo. 2005 Nov-Dec;19(6):1101-4.
Pajeva I, Todorov DK, Seydel J. Membrane effects of the antitumor drugs
doxorubicin and thaliblastine: comparison to multidrug resistance modulators
verapamil and trans-flupentixol. Eur J Pharm Sci. 2004 Feb;21(2-3):243-50.
Paulussen JJ, Fischer MJ, Zuidam NJ, v Miltenburg JC, de Mol NJ, Janssen LH.
Influence of the antiallergic drug oxatomide and derivatives on membrane structures:
relation with inhibition of calcium influx in rat basophilic leukemia cells.
Porcar I, Codoner A, Gomez CM, Abad C, Campos A. Interaction of quinine with
model lipid membranes of different compositions. J Pharm Sci. 2003 Jan;92(1):45-
57.
Principles of Molecular Oncology, by Miguel H. Bronchud, MaryAnn Foote,
Giuseppe Giaccone, Olufunmilayo I. Olopade, Paul Workman (Editors), Humana
Press; 3rd ed. edition (November 30, 2007).
Rabow AA, Shoemaker RH, Sausville EA, Covell DG. Mining the National
Cancer Institute's tumor-screening database: identification of compounds with
similar cellular activities. J Med Chem. 2002 Feb 14; 45(4): 818-40.
Rakotomanga M, Saint-Pierre-Chazalet M, Loiseau PM. Alteration of fatty acid
and sterol metabolism in miltefosine-resistant Leishmania donovani promastigotes

274
and consequences for drug-membrane interactions. Antimicrob Agents Chemother.
2005 Jul;49(7):2677-86.
Recommendations for the Safe Handling of Parenteral Antineoplastic Drugs.
NIH Publication No. 83-2621. For sale by the Superintendent of Documents, US
Government Printing Office, Washington, DC 20402.
Robertson IG, Atwell GJ, Baguley BC. The relationship between lipophilic-
hydrophilic balance, uptake and anti-bacteriophage lambda activity of experimental
anti-tumour bisquaternary salts. Chem Biol Interact. 1988; 65(1): 85-95.
Rodrigues C, Gameiro P, Reis S, Lima JL, de Castro B. Derivative
spectrophotometry as a tool for the determination of drug partition coefficients in
water/dimyristoyl-L-alpha-phosphatidylglycerol (DMPG) liposomes. Biophys Chem.
2001 Dec 11;94(1-2):97-106.
Rodriguez-Casado A, Molina M, Carmona P. New accessory for studies of
isotopic 1H/2H exchange and biomolecular interactions using transmission infrared
spectroscopy. Anal Bioanal Chem. 2006 May; 385(1): 134-8.
Rojo F, Dalmases A, Corominas JM, Albanell J. Pharmacodynamics: biological
activity of targeted therapies in clinical trials. Clin Transl Oncol. 2007 Oct;9(10):634-
44.
Schop H, Wiese M, Cordes HP, Seydel JK. Partial resistance of E. coli mutants
against 2, 4-diamino-5-benzylpyrimidines by interactions with bacterial membrane
lipopolysaccharides. Derivation of quantitative structure-binding relationships.
Schreier S, Malheiros SV, de Paula E. Surface active drugs: self-association and
interaction with membranes and surfactants. Physicochemical and biological aspects.
Schultz H, Hume J, Zhang de S, Gioannini TL, Weiss JP. A novel role for the
bactericidal/permeability increasing protein in interactions of gram-negative
bacterial outer membrane blebs with dendritic cells. J Immunol. 2007 Aug
15;179(4):2477-84.

275
Semin Hematol. 1999 Jan;36(1 Suppl 1):2-6.
Shanghai Kou Qiang Yi Xue. 2004 Aug;13(4):252-5.
Sharma SV, Settleman J. Oncogene addiction: setting the stage for molecularly
targeted cancer therapy. Genes Dev. 2007 Dec 15;21(24):3214-31.
Sharp SY, O'Neill CF, Rogers P, Boxall FE, Kelland LR. Retention of activity by
the new generation platinum agent AMD0473 in four human tumour cell lines
possessing acquired resistance to oxaliplatin. Eur J Cancer. 2002 Nov;38(17):2309-
15.
Sheetz MP, Sable JE, Döbereiner HG. Continuous membrane-cytoskeleton
adhesion requires continuous accommodation to lipid and cytoskeleton dynamics.
Annu Rev Biophys Biomol Struct. 2006;35:417-34.
Shen YC, Chen CY, Kuo YH. A new taxane diterpenoid from Taxus mairei. J
Nat Prod 1998 Jun 26;61(6):838-40.
Shen YC, Chen CY. Taxane diterpenes from Taxus mairei. Planta Med 1997
Dec;63(6):569-70.
Shen YC, Tai HR, Chen CY. New taxane diterpenoids from the roots of Taxus
mairei. J Nat Prod 1996 Feb;59(2):173-6.
Shi QW, Oritani T, Sugiyama T, Yamada T. Isolation and structural
determination of a novel bicyclic taxane diterpene from needles of the Chinese yew,
Taxus mairei. Biosci Biotechnol Biochem 1999 Apr;63(4):756-9.
Shi QW, Oritani T, Sugiyama T. Kiyota H. Three novel bicyclic 3,8-secotaxane
diterpenoids from the needles of the chinese yew, taxus chinensis var. mairei. J Nat
Prod 1998 Nov;61(11):1437-40.

276
Shi QW, Oritani T, Sugiyama T. Three novel bicyclic taxane diterpenoids with
verticillene skeleton from the needles of Chinese yew, Taxus chinensis var. mairei.
Planta Med 1999 May;65(4):356-9.
Shi QW, Oritani T, Sugiyama T. Two bicyclic taxane diterpenoids from the
needles of taxus mairei. Phytochemistry 1999 Feb;50(4):633-6.
Skipper H. E. Historic milestones in cancer biology: a few that are important to
cancer treatment. Semin. Oncol., 1979, 6: 506-514.
Skrede S, Holmsen H. A role for antipsychotic agents in the membrane
hypothesis of schizophrenia? Tidsskr Nor Laegeforen. 2003 Sep 25;123(18):2568-70.
Srinivasan V, Ciddi V, Bringi V. Shuler ML, Metabolic inhibitors, elicitors, and
precursors as tools for probing yield limitation in taxane production by Taxus
chinensis cell cultures. Biotechnol Prog 1996 Jul-Aug;12(4):457-65.
Sugawara M, Takekuma Y, Yamada H, Kobayashi M, Iseki K, Miyazaki K. A
general approach for the prediction of the intestinal absorption of drugs: regression
analysis using the physicochemical properties and drug-membrane electrostatic
interaction.
Sun J, Zhang T, He Z. Immobilized artificial membrane chromatography and
its application in profiling the drug membrane transport. Se Pu. 2005 Jul;23(4):378-
83.
Targeted Molecular Imaging in Oncology, by E. Edmund Kim and David J.
Yang (Editors), Springer; 1 edition (January 15, 2001).
The Biology of Cancer, by Robert A. Weinberg, Garland Science; 1 edition
(June 9, 2006).
The Molecular Biology of Cancer, by Michael Khan, Stella Pelengaris
(Editors), Wiley-Blackwell; 1 edition (February 28, 2006).

277
Thein-Han WW, Stevens WF. Transdermal delivery controlled by a chitosan
membrane. Drug Dev Ind Pharm. 2004 Apr;30(4):397-404.
Tokes ZA, Rogers KE, Rembaum A. Synthesis of adriamycin-coupled
polyglutaraldehyde microspheres and evaluation of their cytostatic activity. Proc Natl
Acad Sci U S A. 1982 Mar; 79(6): 2026-30.
Tournaviti S, Hannemann S, Terjung S, Kitzing TM, Stegmayer C, Ritzerfeld J,
Walther P, Grosse R, Nickel W, Fackler OT. SH4-domain-induced plasma membrane
dynamization promotes bleb-associated cell motility. J Cell Sci. 2007 Nov 1;120(Pt
21):3820-9.
Uchiyama Y. Apoptosis: The history and trends of its studies. Arch Histol
Cytol. 1995 Jun;58(2):127-37.
Venkitaraman AR. Chromosomal instability in cancer: causality and
interdependence. Cell Cycle. 2007 Aug;6(19):2341-3. 2007 Jul 18.
Venter BR, Venter JC, Kaplan NO. Affinity isolation of cultured tumor cells by
means of drugs and hormones covalently bound to glass and Sepharose beads. Proc
Natl Acad Sci U S A. 1976 Jun; 73(6): 2013-7.
Vereb G Jr, Panyi G, Balazs M, Matyus L, Matko J, Damjanovich S. Effect of
cyclosporin A on the membrane potential and Ca2+ level of human lymphoid cell
lines and mouse thymocytes. Biochim Biophys Acta. 1990 Aug 30;1019(2):159-65.
Verkleij AJ, Post JA. Physico-chemical properties and organization of lipids in
membranes: their possible role in myocardial injury. Basic Res Cardiol. 1987;82
Suppl 1:85-91.
Voltz E, Gronemeyer H. A new era of cancer therapy: Cancer cell targeted
therapies are coming of age. Int J Biochem Cell Biol. 2008;40(1):1-8.

278
Wang Y. Chromosome instability in yeast and its implications to the study of
human cancer. Front Biosci. 2008 Jan 1;13:2091-102.
Wang YJ, Sun J, He ZG. Liposome electrokinetic chromatography and its
application in evaluating drug-membrane interactio. Yao Xue Xue Bao. 2006
Jul;41(7):583-8.
Wiese M, Pajeva IK. Molecular modeling study of the multidrug resistance
modifiers cis- and trans-flupentixol. Pharmazie. 1997 Sep;52(9):679-85.
Yang Q, Liu XY, Ajiki S, Hara M, Lundahl P, Miyake J. Avidin-biotin
immobilization of unilamellar liposomes in gel beads for chromatographic analysis of
drug-membrane partitioning. J Chromatogr B Biomed Sci Appl. 1998 Apr 10;707(1-
2):131-41.
Yang Q, Liu XY, Yoshimoto M, Kuboi R, Miyake J. Covalent immobilization of
unilamellar liposomes in gel beads for chromatography. Anal Biochem. 1999 Mar
15;268(2):354-62.
Yoon KA, Burgess DJ. Mathematical modelling of drug transport in emulsion
systems. J Pharm Pharmacol. 1998 Jun;50(6):601-10.
Zhang S, Chen WM, Chen YH. Isolation and identification of two new taxane
diterpenes from Taxus chinensis (Pilger)Rehd. Yao Hsueh Hsueh Pao
1992;27(4):268-72.
Zhang S, Tung-Ling Lee C, Kashiwada Y, Chen K, Zhang DC, Lee KH,
Yunantaxusin A, a new 11(15-->1)-abeo-taxane from Taxus yunnanensis. J Nat Prod
1994 Nov;57(11):1580-3.
Zhao L, Feng SS. Effects of lipid chain unsaturation and headgroup type on
molecular interactions between paclitaxel and phospholipid within model
biomembrane. J Colloid Interface Sci. 2005 May 1;285(1):326-35.

279
Zhao L, Feng SS. Effects of cholesterol component on molecular interactions
between paclitaxel and phospholipid within the lipid monolayer at the air-water
interface. J Colloid Interface Sci. 2006 Aug 1;300(1): 314-26.

280
EK-6: GEN AKTARIMI İLE İLGİLİ GÜNCEL REFERANSLAR

281
ADRIAN-SCOTTO M, Vasileva D., Mallet G., Vasilescu D., About Hydration of Mg2+:
a quantum DFT study, Internet Electron. J. Mol. Des., 1, 1-13, (2003).
AHMAD R., Arakawa H., Tajmir-Riahi H. A., A Comparative Study of DNA
Complexation with Mg(II) and Ca(II) in Aqueous Solution: Major and Minor Grooves
Bindings, Biophys. J., 84, 2460-2466, (2003).
AKAO T., Fukumoto T., Ihara H., Ito A., Conformational Change in DNA Induced by
Cationic Bilayer Membranes, FEBS Lett., 391 (1-2), 215-8, (1996).
AKHTAR, S., Basu, S., Wickstrom, E., Juliano, R. L., Cationic liposome-mediated
transfection with lipofection reagent Methods of Molecular Biology, 91-98, (1991).
ANGELOVA M. I., Tsoneva I., Interactions of DNA with Giant Liposomes, Chem.
Phys. Lipids, 101, 1, 123-137, (1999).
ANCHORDOQUY, T. J., Dean Allison, S., Molina, M. D. C., Girouard, L. G., and
Carson, T. K., Physical Stability of DNA-Based Therapeutics, Drug Disc. Today, 6,
463-470, (2001).
ANWER K. B,, Rhee G., Mendiratta S.K., Recent Progress in Polymeric Gene Delivery
Systems, Crit. Rev. Ther. Drug Carrier. Syst., 20(4), 249-293, (2003).
AVRAMOVIC-ZIKIC O., Colbow K., Turbidity Changes of Lipid Vesicles Near the
Phase Transition Temperature as an Indication of Fusion, Biochim Biophys Acta, Sep
11, 512(1), 97-104, (1978).
AZZAM T., Domb A. J., Current Developments in Gene Transfection Agents, Curr
Drug Deliv., Apr;1(2), 165-93, (2004).
BALKRISHNAN K., Mehdi S. Q., McConnell H. M., Binding and Mobility of Anti-
Dinitrophenyl Monoclonal Antibodies on Fluid-like Langmuir-Blodgett Phospholipid
Monolayers Containing Dinitrophenylated Phospholipids, Biochim. Biophysic. Acta,
1064, 219-228, (1991).

282
BANYAY, M., Sarkar, M. and Gräslund, A., A Library of IR Bands of Nucleic Acids in
Solution, Biophys. Chem., 104, 477-488, (2003).
BARTOSCH B., Cosset F. L., Strategies for Retargeted Gene Delivery using Vectors
Derived from Lentiviruses, Curr Gene Ther., Dec;4(4), 427-43, (2004).
BICHENKOV E. E., Budker V. G., Korobeinicheva I. K., Savchenko E. V., Filimonov
V. V., DNA Interaction with Phosphatidylcholine Liposomes. Melting of DNA and
Phase Transition of the Lipid Membrane in the Complex, Biologicheski Membrani, 5,
8, 843-851 (1988).
BINDER, H., and Zschörnig, O., The Effect of Metal Cations on the Phase Behavior
and Hydration Charactersitics of Phospholipid Membranes, Chem. Phys. Lipids, 115,
39-61, (2002).
BLOOMFIELD V. A., Crothers D. M., Tinoco I. Jr., Nucleic Acids-Structures,
Properties, and Functions, University Science Books, Sausalito, California, (2000).
BRAUN C. S., Kueltzo L. A., Midaugh C. R., Ultraviolet Absorption and Circular
Dichroism Spectroscopy of Nonviral Gene Delivery Complexes, In: Methods in
Molecular Medicine, vol. 65: Nonviral Vectors for Gene Therapy, M. A. Findeis (Ed.),
Humana Press Inc., Totowa, NJ., (2001), pp.253-284.
BRUNI, P., Cingolani, F., Iacussi, M., Pierfederici, F. , and Tosi, G., The Effect of
Bivalent Metal Ions on Complexes DNA-Liposome: a FT-IR Study, 565-566, 237-245,
(2001).
BUELER H.,Adeno-Associated Viral Vectors for Gene Transfer and Gene Therapy,
Biol Chem., Jun;380(6), 613-22, (1999).
BURCKBUCHLER V., Wintgens V., Lecomte S., Percot A., Leborgne C., Danos O.,
Kichler A., Amiel C., DNA Compaction into New DNA Vectors Based on Cyclodextrin

283
Polymer: Surface Enhanced Raman Spectroscopy Characterization, Biopolymers, Apr
5;81(5), 360-70, (2006).
BURTON E. A., Fink D. J., Glorioso J. C., Gene Delivery Using Herpes Simplex Virus
Vectors, DNA Cell Biol., Dec;21(12), 915-36, (2002).
CAFFREY M., Lipidat, CRC Press, Inc. (1993).
CARACCIOLO G., Pozzi D., Amenitsch H., Caminiti R., Multicomponent Cationic
Lipid-DNA Complex Formation: Role of Lipid Mixing, Langmuir, Dec 6; 21 (25),
11582-7, (2005).
CASTELLI F., Raciti G. ,A Calorimetric Study of the Influence of Divalent Cations on
the Thermotropic Behaviour of some Phosphatidylcholines, Thermochim. Acta, 186,
205-215, (1991).
CHABAUD P., Camplo M., Payet D., Serin G., Moreau L., Barthelemy P., Grinstaff M.
W., Cationic Nucleoside Lipids for Gene Delivery, Bioconjug Chem., Mar-Apr;17(2),
466-72, (2006).
CHALIKIAN T. V., Völker G., Plum E., Breslauer K. J., A More Unified Picture for the
Thermodynamics of Nucleic Acid Duplex Melting: A Characterization by Calorimetric
and Volumetric Techniques, Proc. Natl. Acad. Sci. USA, 96, 7853-7858, (1999).
CHONG C. S., Colbow K., Light Scattering and Turbidity Measurements of Lipid
Vesicles, Biochim. Biophys. Acta, 436, 260-282, (1976).
CHU D., Sullivan C. C., Weitzman M. D., Du L., Wolf P. L., Jamieson S. W.,
Thistlethwaite P. A., Direct Comparison of Efficiency and Stability of Gene Transfer
into the Mammalian Heart Using Adeno-Associated Virus Versus Adenovirus
Vectors, J. Thorac Cardiovasc Surg., Sep;126(3), 671-9, (2003).

284
CLEMENT, J., Keifer, K., Kimpfler, A., Garidel, P., and Peschka-Süss, R., Large-Scale
Production of Lipoplexes With Long Shelf-Life, Eur. J. Pharmaceut. Biopharm., 59,
35-43, (2005).
CONWAY J. E., Zolotukhin S., Muzyczka N., Hayward G. S., Byrne B. J.,
Recombinant Adeno-Associated Virus Type 2 Replication and Packaging is Entirely
Supported by a Herpes Simplex Virus Type 1 Amplicon Expressing Rep and Cap., J.
Virol., Nov;71(11), 8780-9, (1997).
DANG J. M., Leong K. W., Natural Polymers for Gene Dlivery and Tissue
Engineering, Adv Drug Deliv Rev., Jun 6; (2006).
DASS R., Cytotoxicity Issues Pertinent to Lipoplex-Mediated Gene Therapy In-Vivo,
J. Pharm. Pharmacol., 54, 593-601, (2002).
de GENNES P-G., Problems of DNA Entry into a Cell, Physica A, 274, 1-7, (1999).
DRITSCHILO, A., Thierry, A.R., Rahman, A., Targeted delivery of DNA by liposomes
and polymers, J. of Contr. Release, 19, 269-274, (1992).
DUGUID J. G., Bloomfield V. A., Aggregation of Melted DNA by Divalent Metal Ion-
Mediated Cross-Linking, Biophys J., 69, 2642-2648, (1995).
EGBARIA K., Weiner N., Hydroxylated Nylon Polymers Hydroxylated Nylons Based
on Unprotected Esterified D-Glucaric Acid by Simple Condensation Reactions, J.
Amer. Chem. Soc., 116, 571-578, (1994).
EL-ANEED A., An Overview of Current Delivery Systems in Cancer Gene Therapy, J.
Control Release., Jan 8;94(1),1-14, (2004).
ESPONDA P., Goldstein M., Witkin S. S., In Vitro Transfection of the Human Vas
Deferens Using DNA-Liposome and DNA-Neutral Lipid Complexes, Fertility and
Sterility, 81;1, 171-175, (2004).

285
FALKNER F. G., Holzer G. W., Vaccinia Viral/Retroviral Chimeric Vectors, Curr
Gene Ther., Dec;4(4), 417-26, (2004).
FELGNER P. L., Liposomes for the transformation of eukaryotic cells, Biochim.
Biophysica Acta, 1097, 1-17, (1991).
FLETCHER S., Ahmad A., Perouzel E., Heron A., Miller A. D., Jorgensen M. R., In
Vivo Studies of Dialkynoyl Analogues of DOTAP Demonstrate Improved Gene
Transfer Efficiency of Cationic Liposomes in Mouse Lung, J. Med Chem., Jan 12; 49
(1), 349-57, (2006).
FLETCHER S., Ahmad A., Perouzel E., Jorgensen M. R., Miller A. D., A Dialkynoyl
Analogue of DOPE Improves Gene Transfer of Lower-Charged, Cationic Lipoplexes,
Org Biomol Chem., Jan 21; 4 (2), 196-9, (2005).
FLOTTE T. R., Laube B. L., Gene Therapy in Cystic Fibrosis, Chest., Sep;120(3
Suppl), 124S-131S, (2001).
FORATO, L.A., Bernardes-Filho, R. and Colnago, L.A., Protein Struture in KBr
Pellets by Infrared Spectroscopy, Anal. Biochemistry, 259, 136-141, (1998).
FRAÚSTO DA SILVA J. J. R., Williams R. J. P., The Biological Chemistry of the
Elements. The Inorganic Chemistry of Life, Chapter 9: The Biological Chemistry of
Magnesium:Phosphate Metabolism, 2nd. ed., Oxford University Press, Oxford, New
York, (2001), pp. 251-276.
FUJII G. ,To Fuse or Not to Fuse: The Effects of Electrostatic Interactions,
Hydrophobic Forces, and Structural Amphiphilicity on Protein-Mediated Membrane
Destabilization, Adv Drug Deliv Rev., 20; 38(3), 257-277, (1999).
GAMON B. L., Virden J. W., Berg J. C., The Aggregation Kinetics of an
Electrostatically Stabilized Dipalmitoyl Phosphatidylcholine System, J. Coll. Interf.
Science, 132, 1, 125-138, (1989).

286
GARIDEL, P., Blume, A., and Hübner, W., A Fourier Transform Infrared
Spectroscopic Study of the Interaction of Alkaline Earth Cations With the Negatively
Charged Phospholipid 1,2-dimyristoyl-sn-glycero-3-phosphoglycerol,. Biochim.
Biophys. Acta, 1466, 245-259, (2000).
GAUGER, D.R., Selle, C., Fritzsche, H., and Pohle, W., Chain-Length Dependence of
the Hydration Properties of Saturated Phosphatidylcholines as Revealed by FTIR
Spectroscopy, J. Mol. Structure, 565-566, 25-29, (2001).
GEBHART C. L., Kabanov A. V., Evaluation of Polyplexes as Gene Transfer Agents, J.
Contr. Releas., 73, 401-416, (2001).
GEILEN C. C., Wieder T., Haase A., Reutter W., Morre D. M., Moore D. J.,
Pharmacokinetic Behavior and Antineoplastic Activity of Liposomal-
Hexadecylphosphocholine, Cancer Chemotherapy & Pharmacology, 34, 393-398,
(1994).
GELBART W. M., Bruinsma R. F., Pincus P. A., Parsegian V. A., DNA-Inspired
Electrostatics, Physics Today, Sep., 38-44, (2000).
GODBEY W. T., Atala A., In Vitro Systems for Tissue Engineering, Ann N Y Acad Sci.,
Jun;961, 10-26, (2002).
GRDADOLNIK J., Hadzi D., Conformational Effects of Metal Salt Binding to the
Polar Head of Phosphatidylcholines iIvestigated by FTIR Spectroscopy, Chem. Phys.
Lipids, 65, 121-132, (1993).
GREGORIADIS G. (Ed.)., Liposome Technology, Vol 1, 2, and 3, CRC Press, Inc.,
(1993).
GUNSTONE F. D., Harwood J. L.and Padley, F. B., The Lipid Handbook, 2nd Ed.,
CRC Pres., (1994).

287
GÜNZLER, H. and Gremlich, H.-U., IR Spectroscopy. An Introduction, Wiley-VCH
Verlag GmbH, 69469, Weinheim (Germany), (2002).
HACKL, E. V., Kornilova, S. V., Kapinos, L. E., Andrushchenko, V. V., Galkin, Vi L.,
Grigoriev, D. N., and Blagoi, Yu. P., Study of Ca2+, Mn2+ and Cu2+ Binding to DNA in
Solution by Means of IR Spectroscopy, J. Mol. Structure, 408/409, 229-232, (1997).
HART S.L. Lipid Carriers for Gene Therapy, Curr Drug Deliv., Oct; 2 (4), 423-8,
(2005).
HAUSER H., Effect of Inorganic Cations on Phase Transitions, Chem. Phys. Lipids ,
57, 309-325, (1991).
HENDRIE P. C., Russell D. W., Gene Targeting with Viral Vectors, Mol Ther.,
Jul;12(1), 9-17, (2005).
HENDRIKS W. T., Ruitenberg M. J., Blits B., Boer G. J., Verhaagen J., Viral Vector-
Mediated Gene Transfer of Neurotrophins to Promote Regeneration of the Injured
Spinal Cord, Prog Brain Res., 146, 451-76, (2004).
HERMANSON, G. T., Hermanson, G. T., Bioconjugate Techniques, Academic Pres,
(1996).
HINRICHS W. L., Mancenido F. A., Sanders N. N., Braeckmans K., De Smedt S. C.,
Demeester J., Frijlink H. W., The Choice of a Suitable Oligosaccharide to Prevent
Aggregation of PEGylated Nanoparticles During Freeze Thawing and Freeze Drying,
Int J Pharm., Mar 27; 311 (1-2), 237-44, (2006).
HOLT J. R., Johns D. C., Wang S., Chen Z. Y., Dunn R. J., Marban E., Corey D. P.,
Functional Expression of Exogenous Proteins in Mammalian Sensory Hair Cells
Infected with Adenoviral Vectors, J. Neurophysiol., Apr;81(4), 1881-8, (1999).
HUD N. V. Polak M., DNA – Cation Interactions: The Major and Minor Grooves are
Flexible Ionophores, Curr Opin Struct Biol., 11, 293-301, (2001).

288
HUG P., Slight, R. G., Liposomal delivery as a new approach to transport antisense
oligonucleotides, Gene Regulation, Erickson, R. P. & Izant, G., (Eds.), (1992), Raven
Press, New York., Pp: 147-161.
HÜBNER, W. and Blume, A., Interactions at the Lipid-Water Interface, Chem. Phys.
Lipids, 96, 99-123, (1998).
IGARASHI S, Hattori Y, Maitani Y, Biosurfactant MEL-A Enhances Cellular
Association and Gene Transfection by Cationic Liposome, J. Control Release, May
30; 112(3), 362-8, (2006).
IVANOV I. I., Chapter V: Calorimetric Methods of Studying Biopolymers and
Membrane Systems, in: Modern Methods of Biophysical Investigations – a Practicum
of Biophysics, Ed. A. B. Rubin, Vishaya Shkola, Moscow, (1988), pp 203 – 216.
JANG J. H., Houchin T. L., Shea L. D., Gene Delivery from Polymer Scaffolds for
Tissue Engineering, Expert Rev Med Devices, Sep;1(1),127-38, (2004).
JANOFF, A. S. (Ed.), Liposomes: Rational Design, Marcel Dekker (1998).
JEWELL C. M., Hays M. E., Kondo Y., Abbott N. L., Lynn D. M., Ferrocene-
Containing Cationic Lipids for the Delivery of DNA: Oxidation State Determines
Transfection Activity, J. Control Release, May 1; 112(1), 129-38, (2006).
JOHNSTON, S. F., Fourier Transform Infrared – A Constantly Evolving Technology,
Ellis Horwood Limited, (1991).
JULIANO R. L., Peptide and protein drugs: I. Therapeutic applications, absorption
and parenteral administration, Inter. J. Pharm., 75, 97-115, (1991).
JULLIEN L., Cottet H., Hamelin B., Jardy A., Thermodynamic Behaviour of a
Supramolecular System Self-Assembled by Electrostatic Interaction in Aqueous

289
Solution. Results and Theoretical Analysis, J. Phys. Chem. B, 103, 10866-10875,
(1999).
KABANOV A.V., Kabanov V. A., DNA Complexes with Polycations for the Delivery of
Genetic Material into Cells, Bioconjug Chem., Jan-Feb;6(1), 7-20, (1995).
KAMIYA H., Akita H., Harashima H., Pharmacokinetic and Pharmacodynamic
Considerations in Gene Therapy, Drug Discov Today, 1; 8(21), 990-6, (2003).
KAPLITT M. G., Makimura H., Defective Viral Vectors as Agents for Gene Transfer in
the Nervous System, J. Neurosci Methods, Jan;71(1), 125-32, (1997).
KARMALI P. P., Majeti B. K., Sreedhar B., Chaudhuri A., In Vitro Gene Transfer
Efficacies and Serum Compatibility Profiles of Novel Mono-, Di-, and Tri-
Histidinylated Cationic Transfection Lipids: A Structure-Activity Investigation,
Bioconjug Chem., Jan-Feb; 17(1):, 59-71, (2006).
KAUFMANN-KOLLE P., Drevs J., Berger M. R., Kotting J., Marschner N., Unger C.,
Eibl H., Topical Administration of Hexadecyl-Phosphocholine in Patients with
Cutaneous Lymphomas: Results of a Phase I/II study, J. of the American Academy of
Dermatology, 29, 963-970, (1993).
KAWAKAMI S., Ito Y., Fumoto S., Yamashita F., Hashida M., Enhanced Gene
Expression in Lung by a Stabilized Lipoplex Using Sodium Chloride for Complex
Formation, J Gene Med., Dec; 7 (12), 1526-33, (2005).
KEILY D. E., Chen, L., Lin T. H., Alkyl Phosphocholine Structure of Glucagon-Like
Peptide (7-36) Amide in a Dodecyl-Phosphocholine Micelle as Determined by 2D
NMR, Biochemistry, 33, 3532-3539, (1994).
KHALIL I. A., Kogure K., Akita H., Harashima H., Uptake Pathways and Subsequent
Intracellular Trafficking in Nonviral Gene Delivery, Pharmacol Rev., Mar; 58(1), 32-
45, (2006).

290
KHARAKOZ D. P., Khusainova R. S., Gorelov A. V., Dawson K. A., Stoichiometry of
Dipalmytoilphosphatidylcholine-DNA Interaction in the Presence of Ca2+: a
Temperature-Scanning Ultrasonic Study, FEBS Lett., 446, 27-29, (1999).
KHAZANOV E., Simberg D., Barenholz Y., Lipoplexes Prepared from Cationic
Liposomes and Mammalian DNA Induce CpG-Independent, Direct Cytotoxic Effects
in Cell Cultures and in Mice, J. Gene Med., (2006).
KIRBY C. J., Gregoriadis G., Liposomes, in: E. Mathiowitz (Ed.), Encyclopedia of
Controlled Drug Delivery, Vol. 1, John Wiley &Sons, Inc., (1999), pp. 461-492.
KOBAYASHI, N., Nishikawa, M., and Takakura, Y., Gene Therapy and Gene Delivery,
In: Drug Delivery: Principles and Applications, Binghe Wang, Teruna Siahaan, and
Richard Soltero (Eds.), John Wiley & Sons, Inc., (2005), Pp: 305-318.
KOOTSTRA, N. A., Verma I. M., Gene Therapy with Viral Vectors, Ann. Rev.
Pharmacol. Toxicol., 43, 413-39, (2003).
KOYNOVA R., Koumanov A., Tenchov B., Metastable Rippled Gel Phase in Saturated
Phosphatidylcholines: Calorimetric and Densitometric Characterization, Biochim.
Biophys. Acta, 1285, 101-108, (1996).
KOYNOVA R., Wang L., Tarahovsky Y., MacDonald R. C., Lipid Phase Control of
DNA Delivery, Bioconjug Chem., Nov-Dec;16 (6), 1335-9, (2005).
KUVITCHKIN V. V., Sukhomudrenko A. G., Interaction of Natural and Synthetic
Polynucleotides with Liposomes in the Presence of Divalent Cations, Biophysics,
XXXII, 4, 628-633 (1987).
LAI, E., van Zanten, J. H., Evidence of Lipoplex Dissociation in Liquid Formulations,
J. Pharm. Sci., 91, 1225-1232, (2002).
LAM M. I., Cullis P. R., Calcium Enhances the Transfection Potency of Plasmid DNA-
Cationic Liposome Complexes, Biochim. Biophys. Acta, 1463, 279-290, (2000).

291
LAMBERT G., Fattal E., Couvreur P., Nanoparticulate Systems for the Delivery of
Antisence Oligonucleotides, Adv. Drug Del. Rev., 47, 99-112, (2001).
LASIC D. D. & Yechezkel B., Handbook of Nonmedical Applications of Liposomes,
Vol. I, II, III, & IV, CRC Press, Inc., (1996).
LASIC D. D. (Ed.), Liposomes in Gene Delivery, CRC Press, Inc., (1997).
LASIC D. D. and Papahadjopoulos D. (Ed.), Medical Applications Of Liposomes,
Elsevier Press.
LASIC D. D. and Templeton N. S. (Ed.), Gene Therapy: Therapeutic Mechanisms
and Strategies, Marcel Dekker (2000).
LASIC, D. D., Targeted liposomes and intracellular delivery of macromolecules,
Progress in Clinical and Biology Research, 343, 95-102, (1990).
Le BIHAN, T., and Pézolet, M., Study of the Structure and Phase Behavior of
Dipalmitoylphosphatidylcholine by Infrared Spectroscopy: Characterization of the
Pretranstion and Subtransition, Chem. Phys. Lipids, 94, 13-33, (1998).
LESERMAN, L., Machy, P., Leonetti, P., Jilhaud, P. G., Degols, G., Lebleu, B.,
Liposomal Drug Delivery System, Polymembrane Preparation ,Vol. 31, 159-160,
(1990).
LEVENTIS R., Silvius J. R., Systemic Gene Expression After Intravenous DNA
Delivery in Adult Mice, Science, 261, 209-211, (1993).
LEWIS, R. N. A. H., Winter, I., Kriechbaum M., Lohner, K. and McElhaney, R. N.
Studies of the Structure and Organization of Cationic Lipid Bilayer Membranes:
Calorimetric, Spectroscopic, and X-ray Diffraction Studies of Linear Saturated P-O-
ethyl Phosphatidylcholines, Biophys. J., 60, 1329-1342, (2001).

292
LEWIS, R.N.A.H. and McElhaney, R.N., Fourier Transform Infrared Spectroscopy in
The study of Hydrated Lipids and Lipid Bilayer Membranes. in: H.H. Mantsch and D.
Chapman (Eds.), Infrared Spectroscopy of Biomolecules. Wiley-Liss, New York,
(1996), Pp: 159-202.
LIN A. J., Slack N. L., Ahmad A., George C. X., Samuel C. E., Three-Dimensional
Imaging of Lipid Gene-Carriers: Membrane Charge Density Controls Universal
Transfection Behaviour in Lamellar Cationic Liposome-DNA Complexes, Biophys. J.,
84, 3307-3316, (2003).
LIU F., Huang L., Development of Non-Viral Vectors for Systemic Gene Delivery, J.
Contr. Release, 78, 259-266, (2002).
Liposome Preparation, Pharmaceutical Manufacturing of Liposomes, Martin F. J.
(1990), In: Liposomes as Drug Delivery Systems, Part I: Preparation,
Pharmaceutical Technology Volume 16, (1992), Pp: 96-106.
LOBO B. A., Rogers S. A., Wiethoff C. M., Choosakoonkriang S., Bogdanovich-Knipp
S., Middaugh C. R., Characterization of Cationic Vector-Based Gene Delivery Vehicles
Using Isothermal Titration and Differential Scanning Calorimetry, in: Methods in
Molecular Medicine, Vol. 65, Nonviral Vectors for Gene Therapy, Eds. M. A. Findeis,
Humana Press Inc., Totowa, New Jersey, USA, (2001), pp 319-348.
LOHMEYER M., Bittman R., Uptake, Subcellular Distribution and Metabolism of the
Phospholipid Analogue Hexadecylphosphocholine in MCDK Cells, Biochemica et
Biophysica Acta, 1211, 14-22, (1994).
LOPEZ-BARCONS L. A., Polo D., Llorens A., Reig F., Fabra A., Targeted Adriamycin
Delivery to MXT-B2 Metastatic Mammary Carcinoma Cells by Transferrin
Liposomes: Effect of Adriamycin ADR-to-Lipid Ratio, Oncol Rep., Nov; 14 (5), 1337-
43, (2005).

293
LOSER P., Huser A., Hillgenberg M., Kumin D., Both G. W., Hofmann C., Advances
in the Development of Non-Human Viral DNA-Vectors for Gene Delivery, Curr Gene
Ther., May;2(2), 161-71, (2002).
MACDONALD P. M., Leisen J., Marassi F. M., Responce of Phosphatidylcholine in
the Gel and Liquid-Crystalline States to Membrane Surface Charges, Biochemistry,
30, 3558-3566, (1991).
MACDONALD R. C., Gorbonos A., Momsen M. M., Brockman H. L., Surface
Properties of Dioleoyl-sn-Glycerol-3-Ethylphosphocholine, A Cationic
Phosphatidylcholine Transfection Agent, Alone and in Combination with Lipids or
DNA, Langmuir, Mar 14; 22(6), 2770-9, (2006).
MAH C., Byrne B. J., Flotte T. R., Virus-Based Gene Delivery Systems, Clin
Pharmacokinet., 41(12), 901-11, (2002).
MAHESHRI N., Koerber J. T., Kaspar B. K., Schaffer D. V., Directed Evolution of
Adeno-Associated Virus Yields Enhanced Gene Delivery Vectors, Nat Biotechnol.,
Feb;24(2):198-204, (2006).
MARSH D., Handbook of Lipid Bilayers, CRC Press, Inc., (1990).
MARUYAMA, K., Kennel, S. & Huang, L., Liposomes designed to avoid the
reticuloendothelial system, Progress in Clinical and Biological Research, 343, 85-93,
(1990).
MATSUZAKİ K. O., Murase K., Sugishita S., Yoneyama K., Akada M., Ueha A.,
Nakamura Kobayashi S., Optical Characterization of Liposomes by Right Angle Light
Scattering and Turbidity Measurement, Biochim. Biophys. Acta, 1467, 219-226,
(2000).
MAXWELL I. H., Spitzer A. L., Long C. J., Maxwell F., Autonomous Parvovirus
Transduction of a Gene Under Control of Tissue-Specific or Inducible Promoters,
Gene Ther., Jan;3(1), 28-36, (1996).

294
McCULLOUGH, H. N, Juliano, R. L., Liposomes: a pulmonary perspective,
In:Liposomes as Drug Carriers, Gregoriadis, G., (Ed.), (1988), Pp: 679-694.
Measured by Total Internal Reflection Fluorescence Microscopy, Biophys. J., 58,
1235-1249, (1990).
MEIDAN V. M, Glezer J., Salomon S., Sidi Y., Barenholz Y., Cohen J. S., Lilling G.,
Specific Lipoplex-Mediated Antisense Against Bcl-2 in Breast Cancer Cells: A
Comparison Between Different Formulations, J. Liposome Res., Mar;16(1), 27-43,
(2006).
MEL'NIKOV S. M., Dias R., Mel'nikova Y. S., Marques E. F., Miguel M. G., Lindman
B., DNA Conformational Dynamics in the Presence of Catanionic Mixtures, FEBS
Lett., 453, 113-118, (1999).
METZ M. Z., Best D. M., Kane S. E., Harvey Murine Sarcoma Virus/MDR1 Retroviral
Vectors: Efficient Virus Production and Foreign Gene Transduction Using MDR1 as a
Selectable Marker, Virology, Apr 20;208(2), 634-43, (1995).
MIHALKO P., Schreier, H., Abra, R. M., Liposomes for pulmonary applications, In:
Modern Medicine of Canada, Volume 45, (1990), Pp: 233-237.
MILLER A. D., Nonviral Liposomes. Methods Mol. Med., 90, 107-38, (2004).
MILLER D., The Problem with Cationic Liposome/Micelle-Based Non-Viral Vector
Systems for Gene Therapy, Curr. Med. Chem., 10, 1195-1211, (2003).
MOHR L., Geissler M., Gene Therapy: New Developments, Schweiz Rundsch Med
Prax., Dec 18;91(51-52), 2227-35, (2002).
MOLDAWER L. L, Edwards P. D., Josephs M., Minter R. M., Copeland E. M. 3rd,
MacKay S. L., Application of Gene Therapy to Acute Inflammatory Diseases, Shock,
Aug;12(2), 83-101, (1999).

295
MORET-TATAY I., Sanmartin I., Marco F. M., Diaz J., Alino S. F., Nonviral
Therapeutic Cell Vaccine Mediates Potent Antitumor Effects, Vaccine, May 1; 24(18),
3937-45, (2006).
MÖNKKÖNEN J., Urtti A., Lipid Fusion in Oligonucleotide and Gene Delivery with
Cationic Lipids, Adv. Drug Deliv. Rev., 34, 37-49, (1998).
NICOLAU C.,Cudd A., Amphipathic Antigens Availability of Dinitrophenylated Lipids
Haptens for Specific Antibody Binding Depends on the Physical Properties of Host
Bilayer Membranes, J. Biol. Chem., 257, 6434-6439, (1982).
NICOLAU, C. & Cudd, A., Interactions of antisense DNA oligonucleotide analogs with
phospholipid membranes (liposomes), Nucleic Acids Research, 19, 5551-5559, (1991).
OKADA T., Caplen N. J., Ramsey W. J., Onodera M., Shimazaki K., Nomoto T., Ajalli
R., Wildner O., Morris J., Kume A., Hamada H., Blaese R. M., Ozawa K., In situ
Generation of Pseudotyped Retroviral Progeny by Adenovirus-Mediated
Transduction of Tumor Cells Enhances the Killing Effect of HSV-tk Suicide Gene
Therapy In Vitro and In Vivo, J. Gene Med., Mar;6(3),288-99, (2004).
OPANASOPIT P., Nishikawa M., Hashida M., Factors Affecting Drug and Gene
Delivery: Effects of Interaction with Blood Components, Crit Rev Ther Drug Carrier
Syst., 19(3), 191-233, (2002).
OTTOVA-LEITMANNOVA, A. and Tien H. Ti, Membrane Biophysics: As Viewed
from Experimental Bilayer Lipid Membranes (Planar Lipid Bilayers and Spherical
Liposomes)., Elsevier, (2001).
PAASONEN L., Korhonen M., Yliperttula M., Urtti A., Epidermal Cell Culture Model
with Tight Stratum Corneum as a Tool for Dermal Gene Delivery Studies, Int J
Pharm., Jan 13; 307 (2), 188-93, (2006).
PALMER J. A., Branston R. H., Lilley C. E., Robinson M. J., Groutsi F., Smith J.,
Latchman D. S., Coffin R.S., Development and Optimization of Herpes Simplex Virus

296
Vectors for Multiple Long-Term Gene Delivery to the Peripheral Nervous System, J.
Virol., Jun;74(12), 5604-18, (2000).
PAPAHADJOPOULOS, D. & Gabizon, A., Encapsulation/Delivery of DNA
Interactions of cationic lipid vesicles with negatively charged phospholipid vesicles
and biological membranes, Biochemistry, 27, 3917-3925, (1988).
PASSINI M. A., Watson D. J., Wolfe J. H., Gene Delivery to the Mouse Brain with
Adeno-Associated Virus, Methods Mol Biol.,;246, 225-36, (2004).
PEETERS L., Sanders N. N., Braeckmans K., Boussery K., Van de Voorde J., De
Smedt S. C., Demeester J., Vitreous: A Barrier to Nonviral Ocular Gene Therapy,
Invest Ophthalmol Vis Sci., Oct; 46 (10), 3553-61, (2005).
PELISEK J., Gaedtke L., DeRouchey J., Walker G. F., Nikol S., Wagner E., Optimized
Lipopolyplex Formulations for Gene Transfer to Human Colon Carcinoma Cells
Under In Vitro Conditions, J Gene Med., Feb; 8 (2), 186-97, (2006).
PETERS A. R., Dekker, N., van den Berg, L., Boelens, R., Kaptein, R., Slotboom, A. J.,
de Haas G. H., Antitumor ether lipids and alkyl phosphocholines, Drugs of the
Future, 19, 1021-1037, (1994).
PEVSNER, A., and Diem, M., IR Spectroscopic Studies of Major Cellular
Components. III. Hydration of Protein, Nucleic Acid, and Phospholpid Films,
Biopolymers (Biospectroscopy), 72, 282-289, (2003).
PHILLIP R., Liggitt D., Phillip M., Dazin P., Debs R., Liposomes as carriers of DNA,
Crit. Rev. Ther. Drug Carr. Syst., 6, 239-271, (1989).
PISARCHICK M. L., Thompson N. L., Immune Clearance of Liposomes Inhibited by
an Anti-FC Receptor Antibody in vivo, Proc. Nat. Acad. Sci. USA,83, 2699-2703,
(1986).

297
POHLE, W., Gauger, D.R., Fritzsche, H., Rattay, B., Selle, C., Binder, H., and Böhlıg,
H., FTIR-Spectroscopic Characterization of Phosphocholine-Headgroup Model
Compounds, J. Mol. Structure, 563-564, 463-467, (2001).
POHLE, W., Selle, C., Gauger, D. R., Zantl, R., Artzner, F., and Rädler, J. O., FTIR
Spectroscopic Characterization of a Cationc Lipid-DNA Complex and Its
Components, Phys. Chem. Chem. Phys., 2, 4642-4650, (2000).
RASMUSSEN U. B., Benchaibi M., Meyer V., Schlesinger Y., Schughart K, Novel
Human Gene Transfer Vectors: Evaluation of Wild-Type and Recombinant Animal
Adenoviruses in Human-Derived Cells, Hum Gene Ther., Nov 1;10(16), 2587-99,
(1999).
REMAUT K., Lucas B., Braeckmans K., Sanders N. N., Demeester J., De Smedt S. C.
Protection of Oligonucleotides Against Nucleases by Pegylated and Non-Pegylated
Liposomes as Studied by Fluorescence Correlation Spectroscopy, J. Control Release,
Dec 10; 110 (1), 212-26, (2005).
RITTER T., Lehmann M., Volk H. D., Improvements in Gene Therapy: Averting the
Immune Response to Adenoviral Vectors, BioDrugs,;16(1),3-10, (2002).
ROBBINS P. D., Ghivizzani S. C., Viral Vectors for Gene Therapy, Pharmacol Ther.,
Oct; 80(1), 35-47, (1998).
ROMANCZUK H., Galer C. E., Zabner J., Barsomian G., Wadsworth S. C., O'Riordan
C. R., Modification of an Adenoviral Vector with Biologically Selected Peptides: A
Novel Strategy for Gene Delivery to Cells of Choice, Hum Gene Ther., Nov 1;10(16),
2615-26, (1999).
ROMOREN K., Fjeld X. T., Poleo A. B., Smistad G., Thu B. J., Evensen O.,
Transfection Efficiency and Cytotoxicity of Cationic Liposomes in Primary Cultures of
Rainbow Trout (Oncorhynchus mykiss) gill cells, Biochim Biophys Acta, Nov 10; 1717
(1), 50-7, (2005). BOULANGER C., Di Giorgio C., Vierling P., Synthesis of Acridine-
Nuclear Localization Signal (NLS) Conjugates and Evaluation of Their Impact on

298
Lipoplex and Polyplex-Based Transfection, Eur J Med Chem., Dec; 40 (12), 1295-306,
(2005).
RUTHVEN, N. A., Lewis, H. and McElhaney, R. N., The Structure and Organization
of Phospholipid Bilayers as Revealed by Infrared Spectroscopy, Chem. Phys. Lipids,
96, 9–21, (1998).
SAENGER W., Principles of Nucleic Acid Structure, Springer-Verlag, Berlin,
Heidelberg, MIR Publishers, Moscow, (1987).
SAFINYA C. R., Structures of Lipid-DNA Complexes: Supramolecular Assembly and
Gene Delivery, Curr. Opin. Struct. Biol., Aug. 11 (4), 440-8, (2001).
SALVATI A., Ciani L., Ristori S., Martini G., Masi A., Arcangeli A., Physico-Chemical
Characterization and Transfection Efficacy of Cationic Liposomes Containing the
pEGFP Plasmid, Biophys Chem., Apr 20; 121 (1), 21-9, (2006).
SANTEL A., Aleku M., Keil O., Endruschat J., Esche V., Durieux B., Loffler K.,
Fechtner M., Rohl T., Fisch G., Dames S., Arnold W., Giese K., Klippel A., Kaufmann
J., RNA Interference in the Mouse Vascular Endothelium by Systemic Administration
of siRNA-Lipoplexes for Cancer Therapy, Gene Ther., Apr 20, (2006).
SARAF A., Mikos A. G., Gene Delivery Strategies for Cartilage Tissue Engineering,
Adv Drug Deliv Rev., Jun 9, (2006).
SATO Y., Nomura S.-I., Yoshikawa K., Enhanced Uptake of Giant DNA in Cell-Sized
Liposomes, Chem, Phys. Lett., 380, 279-285, (2003).
SAVVA M., Chen P., Aljaberi A., Selvi B., Spelios M., In Vitro Lipofection with Novel
Asymmetric Series of 1,2-dialkoylamidopropane-Based Cytofectins Containing Single
Symmetric Bis-(2-dimethylaminoethane) Polar Headgroups, Bioconjug Chem., Nov-
Dec;16 (6), 1411-22, (2005).

299
SELLE, C. and Pohle, W., Fourier Transform Infrared Spectroscopy as a Probe for the
Study of the Hydration of Lipid Self-assemblies. II. Water Binding Versus Phase
Transitions, Biospectroscopy, 4, 281-294, (1998).
SELLE, C., Pohle, W. and Fritzsche, H., FTIR Spectroscopic Features of Lyotropically
Induced Phase Transitions in Phospholipid Model Membranes, J. Mol. Structure,
480-481, 401-405, (1999).
SHEK P. N., Barber R. F., Jurimaromet, M., Topical Applications of Liposomes, In:
Liposomes as a topical drug delivery system, Polym. Mater. Sci. Eng., 63, 59-63,
(1990).
SILMAN N. J., Fooks A. R., Biophysical Targeting of Adenovirus Vectors for Gene
Therapy, Curr Opin Mol Ther., Oct;2(5), 524-31, (2000).
SILVESTRO, L. and Axelsen, P. H., Infrared Spectroscopy of Supported Lipid
Monolayer, Bilayer, and Multibilayer Membranes, Chem. Phys. Lipids, 96, 69-80,
(1998).
SIMOES S., Filipe A., Faneca H., Mano M., Penacho N., Duzgunes N., de Lima M. P.,
Cationic Liposomes for Gene Delivery, Expert Opin Drug Deliv., Mar;2 (2), 237-54,
(2005).
SINGH M., Ariatti M., A Cationic Cytofectin with Long Spacer Mediates Favourable
Transfection in Transformed Human Epithelial Cells, Int J Pharm., Feb 17; 309 (1-2),
189-98, (2006).
SORGI, F., and Huang, L., A Dry Powder Formulation for Gene Therapy, Pharm.
Res., 12, S266, (1995).
SMALL D., Handbook of Lipid Research Vol. 4, The Physical Chemistry of Lipids,
Plenum Pres, (1986).

300
SMITH J. G., Walzem R. L., German G. B., Liposomes as Agents of DNA Transfer,
Biochim. Biophys. Acta, 1154, 327-340, (1993).
SOROKIN V. A., Valeyev V. A., Gladchenko G. O., Blagoi Yu. P., Features of the
Interaction of Bivalent Metal Ions with Homopolynucleotides in the Multichain
Conformation, Biophysics, 1994; 39,5, 821-832, (1996).
STEBELSKA K., Dubielecka P. M., Sikorski A. F., The Effect of PS Content on the
Ability of Natural Membranes to Fuse with Positively Charged Liposomes and
Lipoplexes, J. Membr Biol., Aug; 206(3), 203-14, (2005).
STONE D., David A., Bolognani F., Lowenstein P. R., Castro M. G., Viral Vectors for
Gene Delivery and Gene Therapy Within the Endocrine System, J. Endocrinol.,
Feb;164(2), 103-18, (2000).
STRAYER D. S., Viral Gene Delivery, Expert Opin Investig Drugs., Dec;8(12), 2159-
2172, (1999).
SÜLEYMANOĞLU E., Turbidimetric Study of Interaction Between
Polyribonucleotides, Phosphatidylcholine Vesicles and Metal Ions, NATO-Advanced
Study Institute Proceedings: Trafficking of Intracellular Membranes: From Molecular
Sorting to Membrane Fusion, June 19-30, (1994), Espinho, Portugal.
SÜLEYMANOĞLU E., M. Sc. Thesis, Middle East Technical University, Ankara,
Turkey, (1995).
SÜLEYMANOĞLU E., Preparation and Phase Behaviour of Surface-Active
Pharmaceuticals: Self-Assembly of DNA and Surfactants with Membranes.
Differential Adiabatic Scanning Microcalorimetric Study, Farmaco, 60, (8), 701-10,
(2005).
SÜLEYMANOĞLU E., Fluorescence Microscopy of Condensed DNA Conformations
of Bacterial Cells, J. Microbiol., 40, (4), 319-326, (2002).

301
SÜLEYMANOĞLU, E., A Nanoscale Polynucleotide-Neutral Liposome Self-
Assemblies Formulated for Therapeutic Gene Delivery, Electron. J. Biomed., 2, 13-35,
(2004).
SZOKA, Jr. F. & Papahadjopoulos, D. Comparative Properties and Methods of
Prepartaion of Lipid Vesicles (Liposomes), Ann. Rev. Biophys. Bioeng., 9, 467-508,
(1980).
TAILLANDIER, I.E. and J. Liquier. (2002), in: J.M. Chalmers and Griffith (eds.),
Handbook of Vibrational Spectroscopy, John Wiley & Sons, Chichester, Vol. 5,
Chapter 2, Pp. 3465-3480.
TAKAHASHI T., Harada A., Emi N., Kono K., Preparation of Efficient Gene Carriers
Using a Polyamidoamine Dendron-Bearing Lipid: Improvement of Serum Resistance,
Bioconjug Chem., Sep-Oct; 16 (5), 1160-5, (2005).
TALSMA, H & Crommelin, D. J. A., Liposomes as Drug Delivery Systems, Part III:
Stabilization, Pharmaceutical Technology, Volume 17, (1992), Pp: 48-59.
TALSMA, H. & Cormmelin, D. J. A., Liposomes as Drug Delivery Systems, Part II:
Characterization, Pharmaceutical Technology, Volume 16, (1992), Pp: 52-58.
TALSMA, H. & Crommelin, D. J. A., Drug Delivery Overview: Liposomes: Synthetic
lipid microspheres serve as multipurpose vehicles for the delivery of drugs, genetic
material and cosmetics, American Scientist, 80, 20-31, (1992).
TARAHOVSKY Y. S., Khusainova R. S., Gorelov A. V., Nikolaeva T. I., Deev A. A.,
Dawson A. K., Ivanitsky G. R., DNA Initiates Polymorphic Structural Transitions in
Lecithin, FEBS Lett., 390, 133-136, (1996).
TATULIAN S. A., Gordeliy V. I., Sokolova A. E., Syrykh A.G., A Neutron Diffraction
Study of the Ions on Phospholipid Membrane Interactions, Biochim. Biophys. Acta,
1070, 143-151, (1991).

302
TEMPLETON N S., Gene and Cell Therapy-Therapeutic Mechanisms and Strategies,
2nd Ed., Marcel Dekker, (2003).
TEMPLETON N. S., Developments in Liposomal Gene Delivery Systems, Expert
Opin. Biol. Ther., 1, 1-4, (2001).
TEMPLETON N. S., Liposomal Delivery of Nucleic Acids In Vivo. DNA Cell Biol., 12,
857-867, (2002).
THIERRY A. R., Abes S., Resina S., Travo A., Richard J. P., Prevot P., Lebleu B.,
Comparison of Basic Peptides- and Lipid-Based Strategies for the Delivery of Splice
Correcting Oligonucleotides, Biochim Biophys Acta, Mar; 1758 (3), 364-74., (2006).
THORNTON K., Gorenstein D.G., Conformational changes in phospholipase A2 upon
binding to micellar interfaces in the absence and presence of competitive inhibitors.
A 1H and 15N NMR study, Biochemistry, 31, 10021-10030, (1992).
UHRIKOVA D., Rapp G., Kunst B. H., Balgavy P., Interaction of DNA with DPPC
Bilayers in the Presence of Mg (II) Ions, 1998; EMBL DESY Outstation – Hamburg;
Project No: ML-98-6.
ULRICH A. S., Biophysical Aspects of Using Liposomes as Delivery Vehicles,
Bioscience Rep., 22, 2, 129-149, (2002).
van der WEERT, M., Haris, P.I., Hennink, W.E., and Crommelin, D.J.A., Fourier
Transform Infrared Spectrometric Analysis of Protein Conformation: Effect of
Sampling Method and Stress Factors, Anal. Biochemistry, 297, 160-169, (2001).
VARGA C. M, Wickham T. J., Lauffenburger D. A., Receptor-Mediated Targeting of
Gene Delivery Vectors: Insights from Molecular Mechanisms for Improved Vehicle
Design, Biotechnol Bioeng., Dec 20; 70(6), 593-605, (2000).
VLASSOV V. V., Balakireva L. A., Yakubov L. A., Transport of Oligonucleotides
Across Natural and Model Membranes, Biochim Biophys Acta, 1197, 95-108, (1994).

303
WALTHER W., Stein U., Viral Vectors for Gene Transfer: A Review of Their Use in
the Treatment of Human Diseases, Drugs, Aug;60(2), 249-71, (2000).
WANG X. T., Liu P. Y., Xin K. Q., Tang J. B., Tendon Healing In Vitro: bFGF Gene
Transfer to Tenocytes by Adeno-Associated Viral Vectors Promotes Expression of
Collagen Genes, J. Hand Surg [Am]. Nov;30(6), 1255-61, (2005).
WANG, Binghe, Teruna Siahaan, and Richard Soltero (Eds.), Drug Delivery:
Principles and Applications, John Wiley & Sons, Inc., (2005).
WANG, G., Liposomes as Drug Delivery Vehicles, In: Drug Delivery: Principles and
Applications, Binghe Wang, Teruna Siahaan, and Richard Soltero (Eds.), John Wiley
& Sons, Inc., (2005), Pp: 411-434.
WASUNGU L., Scarzello M., van Dam G., Molema G., Wagenaar A., Engberts J. B.,
Hoekstra D., Transfection Mediated by pH-Sensitive Sugar-Based Gemini
Surfactants; Potential for In Vivo Gene Therapy Applications, J. Mol Med., Jun 8,
(2006).
WHITTLESEY K. J., Shea L. D., Nerve Growth Factor Expression by PLG-Mediated
Lipofection, Biomaterials, Apr; 27 (11), 2477-86, (2006).
WILSON D. R., Viral-Mediated Gene Transfer for Cancer Treatment, Curr Pharm
Biotechnol., Jun;3(2), 151-64, (2002).
YOUNG L. S., Searle P. F., Onion D., Mautner V., Viral Gene Therapy Strategies:
From Basic Science to Clinical Application, J. Pathol., Jan;208(2), 299-318, (2006).
ZHANG X., Godbey W.T., Viral Vectors for Gene Delivery in Tissue Engineering, Adv
Drug Deliv Rev., Jun 6, (2006).
ZHOU X, Huang L., Improved encapsulation of DNA in pH sensitive liposomes for
transfection, Liposome Research, 2, 125-139, (1992).

304
ZHOU X. H., Po, A. L. W. Pulmonary Delivery of Drugs Organ-selective action of an
antitumor drug: pharmacologic studies of liposome-encapsulated beta-cytosine
arabinoside administered via the respiratory system of the rat, J. Natl. Cancer Inst.,
63, 727-731, (1979).
ZHOU X., Klibanov A. L., Huang L., Encapsulation/Delivery of Proteins Interactions
of proteins and drugs with liposomes, In: Liposomes, Ostro M., (Ed.), Marcel Dekker,
New York, (1983) Pp: 53-86.
ZHU N., Liggitt D., Phillip M., Dazin P., Debs R., In Vivo Gene Delivery. Efficient
Transfection of T lymphocytes in Adult Mice, J. Biol. Chem., 268, 16087-16090,
(1993).
ZUNDEL, G. Polar Interactions, Hydration, Proton Conduction and Conformation of
Biological Systems – Infrared Results, , in: W. Hoppe, W. Lohmann, H. Markl, H.
Ziegler (Eds.), Biophysics, Springer-Verlag, Berlin, Hedelberg, New York, Tokyo,
(1983). Pp: 243-254.

305
Gen Taşıyıcı Tasarımlarında Kullanılan Çeşitli DNA Kompaktizasyon
Molekülleri
Tablo yapımında kullanılan kaynaklar62:
[1] http://www.wiley.co.uk/wileychi/genmed/clinical/index.html.; [2] ZIAUDDIN J.,
Sabatini D. M., Microarrays of cells expressing defined cDNAs. Nature,411 (6833),
107–10, (2001). [3] MARTIN-HERRANZ A., Ahmad A., Evans H. M., Ewert K.,
Schulze U., Safinya C. R., Surface functionalized cationic lipid-DNA complexes for
gene delivery: PEGylated lamellar complexes exhibit distinct DNA-DNA interaction
regimes. Biophys J 86(2), 1160–8, (2004). [4] BOULANGER C., Di Giorgio C.,
Gaucheron J., Vierling P., Transfection with fluorinated lipoplexes based on new
fluorinated cationic lipids and in the presence of a bile salt surfactant. Bioconjug
Chem 15(4), 901–8, (2004). [5] SEGURA T., Shea L. D., Materials for non-viral gene
delivery, Annu Rev Mater Res, 31, 25–46, (2001). [6] Anderson D. G., Lynn D. M.,
62
Tablo takibi daha kolay olması açısından burada yer alan kaynaklar Yararlanılan Kaynak Listesine dahil edilmemiştir.

306
Langer R., Semi-automated synthesis and screening of a large library of degradable
cationic polymers for gene delivery, Angew Chem Int Ed Engl, 42(27), 3153–8,
(2003). [7] EWERT K., Ahmad A., Evans H. M., Safinya C. R., Cationic lipid–DNA
complexes for non-viral gene therapy: relating supramolecular structures to cellular
pathways. Expert Opin Biol Ther, 5(1), 33–53, (2005). [8] TRANCHANT I.,
Thompson B., Nicolazzi C., Mignet N., Scherman D., Physicochemical optimisation of
plasmid delivery by cationic lipids. J Gene Med, 6 (Suppl 1), S24–35, (2004). [9]
BOUSSIF O., Lezoualc’h F., Zanta M. A., Mergny M. D., Scherman D., Demeneix B.,
et al. A versatile vector for gene and oligonucleotide transfer into cells in culture and
in vivo: polyethylenimine. Proc. Natl. Acad Sci USA, 92(16), 7297–301, (1995). [10]
BRAUN C. S., Vetro J. A., Tomalia D. A., Koe G. S., Koe J. G., Russell Middaugh C.,
Structure/function relationships of polyamidoamine/DNA dendrimers. as gene
delivery vehicles. J Pharm Sci , 94 (2), 423–36, (2005). [11] WANG J., Zhang P. C.,
Mao H. Q., Leong K. W., Enhanced gene expression in mouse muscle by sustained
release of plasmid DNA using PPE-EA as a carrier, Gene Ther,9 (18), 1254–61,
(2002). [12] AKINC A., Lynn D. M., Anderson D. G., Langer R., Parallel synthesis and
biophysical characterization of a degradable polymer library for gene delivery, J Am
Chem Soc, 125 (18), 5316–23, (2003). [13] NEU M., Fischer D., Kissel T., Recent
advances in rational gene transfer vector design based on poly(ethylene imine) and
its derivatives, J Gene Med, 7 (8), 992–1009, (2005). [14] SCHAFFER D. V.,
Fidelman N. A., Dan N., Lauffenburger D. A.. Vector unpacking as a potential barrier
for receptor-mediated polyplex gene delivery, Biotechnol Bioeng., 67(5), 598–606,
(2000). [15] AGARWAL A., Unfer R., Mallapragada S. K., Novel cationic pentablock
copolymers as non-viral vectors for gene therapy. J Control Release,;103 (1), 245–58,
(2005). [16] REINEKE T. M., Davis M. E., Structural effects of carbohydrate-
containing polycations on gene delivery. 1. Carbohydrate size and its distance from
charge centers, Bioconjug Chem, 14 (1), 247–54, (2003). [17] POPIELARSKI S. R.,
Mishra S., Davis M. E.. Structural effects of carbohydrate-containing polycations on
gene delivery. 3. Cyclodextrin type and functionalization, Bioconjug Chem,14 (3),
672–8, (2003).
Ek-9 LİPOZOM HAZIRLAMA YÖNTEMLERİ63,64
63
Bu bölümde lipozomlarla ilgili sadece genel bilgiler yer almaktadır. Modern farmasötik biliminde çok sayıda kullanım alanlarıyla ilgili güncel makalelerin ve derlemelerin de bulunduğu Referanslar listesine başvurulmalıdır.

307
Lipid Vezikülleri (Lipozom) Hazırlama Yöntemleri
Lipozomlar veya veziküller üç boyutlu olmaları itibariyle doğal hücrelere
benzeyen fosfolipidler gibi amfifilik (Şekil E5.1) moleküllerin kendi kendine
oluşturdukları (self-assembling) yapılara sahip yuvarlak agregatlardır. Lipozomlar ilk
olarak A. D. Bangham65,66 1960lı yıllarda keşfedildiler. Yüksek biyoyararlılıkları ve
çeşitli molekül ve maddelerin vezikül hacmi içinde yüksek miktarda enkapsüle
64 Bu bölümün hazırlanmasında: Wang, G., Liposomes as Drug Delivery Vehicles, In: Drug Delivery:
Principles and Applications, Binghe Wang, Teruna Siahaan, and Richard Soltero (Eds.), John Wiley & Sons,
Inc., (2005), Pp: 411-434; Kobayashi, N., Nishikawa, M., and Takakura, Y., Gene Therapy and Gene Delivery,
In: Drug Delivery: Principles and Applications, Binghe Wang, Teruna Siahaan, and Richard Soltero (Eds.), John
Wiley & Sons, Inc., (2005), Pp: 305-318; Binghe Wang, Teruna Siahaan, and Richard Soltero (Eds.), Drug
Delivery: Principles and Applications, John Wiley & Sons, Inc., (2005)’dan yararlanılmıştır.
65
Bangham, A. D., Horne, R. W., Nature, 196, (1962), 952-953. 66
Bangham, A. D., Horne, R. W., J. Mol. Biol., 8, (1964), 660-668.

308
edilebilmeleri bakımından, lipid vezikülleri o yıllarda mükemmel bir ilâç taşıma aracı
olarak değerlendirilmişlerdir.
Lipozomların çeşitli türleri (MLV, SUV, LUV, FRV) uygulanmakta olan birçok
yöntemi kullanarak hazırlamak mümkündür (Şekil E5.2 ve E5.6). Projemizde çok
popüler bir yöntem olan “hand-shaken vesicles preparation” yöntemi uygulanmıştır.
Lipozomların herhangi bir formülasyonunu hazırlamak için lipid moleküllerini önce
sıvı ortamına aktarmak gerekmektedir. Orijinal prosedüre uygun olarak, ince tabaka
lipid filmi yuvarlak balon jojeye konulmak suretiyle bunların daha sonra sıvı
ortamında çalkalanması ve çözülmesi sağlanmaktadır (Şekil E5.3 ve E5.4).
İlâç formülasyonları, gen taşıyıcıları ve kanser tedavilerinde kullanılan çeşitli
farmasötik bileşenlerinin enkapsülasyonu gibi alanlardaki uygulanmaları dışında,
lamelar veziküller biyomembran modelleri olarak da çok önem kazanmışlardır
(WANG, et. al., 2005; KOBAYASHI, et. al, 2005 WANG, et. al., 2005). Lipozomlar
özellikle bağışıklık sisteminin fagositlerine yönelik ilâç hedeflendirilmesinde etkilidir.
Lipozomlara dayalı ilâç taşıyıcısistemleri terapötik endeksi ve biyoyararlılığı
iyileştirebildikleri gibi, kontrollü salınım sistemlerinin etkinliğini artırırlar ve
toksisiteyi düşürürler. Bu bileşenler, antikanser, DNA ve proteinlerin, diabet ve
kardiyovasküler hastalıklara karşı ilâçların taşıyıcıları olarak kullanılmışlardır.
Keşfedildiklerinden itibaren, birçok lipozomal formülasyon klinik araştırmalarda
uygulanmıştır ve birkaç başarılı lipozom’a dayalı ilâç formülasyonu patent olarak
Food and Drug Administration (FDA) tarafından kabul edilmiş ve
ruhsatlandırılmıştır. Bunlar arasında doxorubicin (Doxil ve Myocet), daunorubicin
(DaunoXome) gibi antikanser ilâçları ve mantarlara karşı kullanılan amphotericin B
(AmBisome, Amphotect, ABELCET) formülasyonları örnek olarak verilebilir. Bu
ilâçların lipozomal formülasyonları önemli ölçüde toksisiteyi düşürmüşlerdir ve etken
maddelerin etkinliğini korumuş veya yükseltmişlerdir.
Son yıllarda lipozomlar viral olmayan gen taşıyıcı tasarımlarında da yoğun
olarak yer almakta ve uygulanmaktadır67.
67 Bkz: Referanslar listesini.

309
Ancak, lipozomal ilâç ve gen taşıyıcı sistemlerinin terapötik potansiyellerine tam
olarak henüz daha ulaşılamamıştır. Bunun nedenlerinden biri, bu taşıyıcı sistemlerle
karşılaşılan birçok tasarım ve uygulama sorunlarının olmasıdır. Bu sorunlar arasında
sabitlilik, salınım zamanı ve hızı, formülasyon maaliyeti, kısa süren biyoyararlılık ve
birçok ilâçla etkisiz etkileşmelerin olduğu durumlar sayılabilir. Ayrıca, sirkülasyonda
sabitliliğini destekleyerek aynı zamanda söz konusu ilâçın hedef dokudaki
biyoyararlılığını seçici olarak artırmak çok güçtür (WANG, 2005).
1. Vezikül Oluşumunun Mekanizması.
Lipozomlar (lipid vezikülleri) ince tabaka lipid filmlerinin veya diğer lipid
yüzeylerinin hidrasyonu sonucu oluşurlar (Şekil E5.1). Bu yapılarda sıvı kristal
şeklinde bulunan lipid katmanları akışkan hale gelir. Hidrasyonu meydana gelmiş
lipid katmanların agitasyon ve çalkalama esnasında bulundukları balon jojenin veya
kabbın iç yüzeyinden ayrılırlar ve daha büyük yuvarlak çok katmanlı veziküller (Large
Multilamellar Vesicles, LMV) oluşur. Kenarlarda katmanın hidrokarbon kısmının su
ile etkileşmeleri engellendiğinden, bu yapı termodinamik açıdan da bakılacak olursa
çok sağlam ve sabittir. Parçacıkların oluşumundan sonra boyutlarının
düşürülmesisonik enerjinin (sonication) veya mekanik enerjinin (extrusion)
harcanmasını gerektirmektedir.
2. Lipozom Hazırlama Yöntemleri.
Lipid formülasyonların özellikleri lipid içeriğine (katyonik, anyonik veya nötr
lipidler) bağlıdır, ancak, aynı hazırlama prosedürü tüm lipid veziküllerin
hazırlanmasında lipid türü önem taşımaksızın uygulanabilmektedir (Şekil E5.2).
Genel olarak, prosedür hidrasyon için lipidin hazırlanması, eşzamanlı
gerçekleştirilen hidrasyon, agitasyon ve çalkalama ve veziküllerin homojen
dağlımının sağlanması için yapılan boyutlandırma aşamalarından ibarettir.

310
Şekil E5.1: Lipozomların genel yapısı. Kaynak: www.cem.msu.edu/~reusch/
VirtualText/lipids.htm
Şekil E5.2: Çeşitli lipozom formülasyonlarının hazırlama yöntemlerini özetleyen
genel klasifikasyon. SUV, small unilamellar vesicles; MLV, multilamellar vesicles;
LUV, large unilamellar vesicles; FRV, freeze-drying rehydration vesicles. Kaynak:
LASCH, J. Weissig, V. and Brandl, M. Preparation of liposomes, In: Liposomes. A

311
Practical Approach, 2nd Ed. (Torchilin Vl. P. And Weissig, V., Eds.), Oxford
University Press, (2003), Pp: 3-29.
a. Hidrasyon İşlemi.
Lipid karışımlarından oluşan lipozomları hazırlarken, lipidler ilk önce organik
çözgende çözülmeli ve homojen lipid karışımının elde edilmesi için iyice
karıştırılmalıdır. Genelde, bu işlem kloroform veya kloroform:metanol karışımı
kullanılarak gerçekleştirilir (TORCHILIN, et. al., 2003). Burada amaç, lipidlerin tam
olarak karışımını sağlamak için temiz lipid çözeltinin elde edilmesidir. Genel olarak,
lipid çözeltileri 10-20 mg lipid/ml organik çözgen konsantrasyonunda hazırlanır.
Ancak, lipid çözünürlülüğü veya lipidlerin tam olarak karışımı isteniliyorsa, daha
yüksek konsantrasyonlarla çalışmak avantajlıdır (TORCHILIN, et. al., 2003). Organik
çözgende lipidlerin çözülmesinden sonra, bu çözücüyü azot veya argon gazı ile
uzaklaştırarak ince lipid filmi elde edilir. Daha küçük organik çözgen hacimleri ile
çalışılıyorsa (<1 ml), bu işlem iyi neticeler verir (TORCHILIN, et. al., 2003). Daha
büyük organik çözgen hacimlerinin söz konusu olduğunda ise uzaklaştırma işlemi
rotary evaporator ile gerçekleştirilir. Her iki işlem sonucunda da balon jojenin
dibinde ince lipid filmi veya tabakası oluşmaktadır. Muhtemelen bu ortamda kalan
organik çözgeni uzaklaştırmak amacıyla, bu film daha sonra bir gece boyunca vaküm
pompa ile kurutulur. Kloroform kullanımı tercih edilmiyorsa, onun yerine lipidleri
tersiyer butanol veyasikloheksanda çözmek de mümkündür. Lipid çözeltisi kaplara
aktarılır ve kuru buz veya kuru buz-aseton karışımıyla veya alkol (etanol veya
metanol) banyosuna konularak dondurulur. Banyoda konulduğu takdirde, ani ısı
değişiklerinden kaynaklanabilecek olan muhtemel kap çatlamasını engellemek için
dikkat edilmeli. Tam olarak dondurulduktan sonra, lipidin içinde bulunduğu kap
vaküm pompaya bağlanır ve lipid filminin kuruyuncaya kadar liyofilize edilir. Bu
işlem elde edilmesi istenilen lipid hacmine göre değişir ve genelde 1 ile 3 gün arasında
sürer. Bunun ardından, kurutulmuş lipid filmi bulunduran balon joje vaküm
pompadan çıkarılır ve tekrar bir sonraki hidrasyon için dondurulmuş vaziyette
muhafaza edilir.
b. Lipid Filminin Hidrasyonu.
Kuru lipid filminin hidrasyonu basitçe sıvı ortamın veya tampon çözeltisinin
lipidin bulunduğu balon jojeye katarak ve çalkalayarak gerçekleştirilir. Hidrasyon

312
ortamının ısısı kullanılan lipidin genel jel-sıvı kristal faz geçiş ısısından (Tc veya Tm)
daha yüksek olmalı. Bu işlemden sonra lipid süspansiyonu, bulunduğu kendi ortam
ısısının söz konusu Tm ‘in üzerinde olacak şekilde muhafaza edilmelidir. Yüksek faz
geçişine sahip lipidler için bu işlemi balon joje içinde bulunan lipidleri rotary
evaporator sistemine bağlayarak vaküm kullanılmaksızın gerçekleştirmek
mümkündür. Daha sonra, bu balon jojeyi yine önceden lipid süspansyon ısısısnın
üzerinde bulunan bir ısı ortamında ısıtılmış su banyosuna aktarılarak ve çalkalayarak
lipidin hidrasyonunu gerçekleştirerek lipidlerin akışkan fazı elde edilmiş olur. Lipid
türüne bağlı olarak hidrasyon için gereken zaman değişmekte, ancak bu işlemin
çalkalama ve vorteksleme zamanları ile birlikte, genelde bir saati aşmamasına özen
gösterilir. Bazı durumlarda, veziküllerin boyut dağılımı ve homojeniteleri
iyileştirildiğinden, bundan sonraki aşama olan vezikül boyutlandırma işlemine
geçmeden önce, lipid süspansyonu bu şekilde bir gece bekletilir68.
Hidrasyon ortamını genelde lipid veziküllerin uygulama amacı tayin eder. Uygun
hidrasyon ortamı, distile su, tampon çözeltileri, tuzları, ve şeker çözeltileri gibi
elektrolit olmayan bileşenleri içerir. In vivo uygulamalar için fizyolojik osmolalitede
(290 mOsm/kg) çalışmak tavsiye edilir (TORCHILIN, et. al., 2003). Bu
gereksinimleri karşılayan genel olarak kullanılan bir çözelti: 0.9% tuz, 5% dekstroz ve
10% sukroz’u içerir. Hidrasyon esnasında, bazı lipidler yapılarına özgü kompleksler
oluştururlar. Yüksek düzeyde yük taşıyan lipidlerin hidrasyonları esnasında ve düşük
pI’e sahip çözeltilerde, vis köz jel oluşturdukları görülmüştür (TORCHILIN, et. al.,
2003). Bu sorun, ortama tuzların katılması veya vezikül boyutlarının düşürülmesiyle
giderilebilir. Fosfatidiletanolamin gibi hidrasyonu zor olan lipidler kendi kendine
agregatlar oluştururlar. >60% fosfatidiletanolamin içeren veziküller vezikül etrafında
oluşan düşük hidrasyon tabakasına sahiptir. Bu durumda bu fosfolipidten oluşan
veziküller arasında uzaklaştırıcı kuvvetlerin zayıflaması sonucu birbirine çok yaklaşıp
birbirleri bağlanarak daha büyük agregatlar oluştururlar (). Bu parçacıklar çözeltide
daha büyük dispers flokülasyonlar olarak meydana gelir. Hidrasyonun sonucunda,
büyük çok katmanlı veziküller (Large Multilamellar Vesicles, LMV) oluşur.
Katmanlar arasında bulunan lipid yüzeyleri su katmanı ile çevrilidir. Aralarında
oluşan mesafe lipid katmanlarındaki lipid içeriğine bağlıdır. Örneğin, polihidre 68
Bu olay “aging” olarak da adlandırılmaktadır. Ancak, lipid hidrolizinin meydana gelme olasılığının yüksek derecelerde artığından, bu işlem yüksek faz geçişine sahip lipidlere uygulanması tercih edilmez (TORCHILIN, et. al., 2003).

313
katmanlar, elektrostatik uzaklaştırma neticesinde oluşan yüklü lipid katmanlarına
göre birbirine daha yakın bir mesafede oluşurlar. Sabit, hidrasyonu gerçekleştirilmiş
çok katmanlı veziküllerin (Large Multilamellar Vesicles, LMV) elde edilmesinden
sonra, bu parçacıkların boyutları sonication ve extrusion gibi birçok yöntem
kullanılarak düşürülür (TORCHILIN, et. al., 2003).
c. Lipid Süspansyonlarının Boyutlandırılması
Sonication
LMV süspansyonlarının sonik enerji (sonication) kullanarak parçalanması
tipik olarak çapları 15-50 nm arasında değişen küçük, tek katmanlı veziküllerin (SUV)
oluşmuuna neden olur. Bunu gerçekleştirmek için kullanılan cihazlar “probe tip
sonicators” olarak bilinmektedir. Ancak bu yöntem birçok tercih edilmeyen yan
etkilere de sahiptir. Örneğin, bu yöntemi kullanarak elde edilen küçük tek katmanlı
veziküller sabit değildir ve zamanla agregasyonlar neticesinde çapları daha büyük
olan agregatlara dönüşürler. Sonuç olarak, elde edilen lipozom formülasyonları son
derece heterojendir. Ayrıca, sonikasyon lipid peroksidasyonlara da neden olmuştur.
Bu peroksidasyonları engellemek amacıyla araştırmacılar lipozom formülasyonlarına
lipid koruyucu ve antioksidan etkilerine sahip α-tokoferol (Vitamin E) katmışlardır.
Ancak bu vitaminin büyük hacmi lipozomların daha sonraki boyut incelemelerini ve
spektroskopik araştırmalarını çok sayıda yan sinyal verdiklerinden olumsuz olarak
etkilemştir. Bu yüzden günümüzde tek katmanlı lipid veziküllerinin elde edilmesinde
kullanılan diğer bir yöntem olan extrusion yöntemi tercih edilmektedir.
Extrusion
Bu yöntemde lipid süspansyonu önceden bilinen ve tercih edilen por
büyüklüğüne sahip 2 polikarbonat filtre üzerinden 10 defa geçirilerek bu por
büyüklüğüne yakın boyuttaki tek katmanlı veziküller elde edilir. Bu yöntem
sayesinde ortalama olarak söz konusu tek katmanlı veziküllerin boyut dağlımı son
derece homojendir. Şimdiye kadar bahsedilen tüm diğer lipozom boyutunun
düşürülmesinde kullanılan yöntemlerde olduğu gibi burada da bu işlem lipidin faz
geçiş derecesinin üzerinde bir değerde çalışılması gerekmektedir. Aksi takdirde, bu
derecenin altında lipidlerin rijid olduklarından ve bu yüzden de polikarbonat
filtresindeki porlara yapışıp tıkanmalarına neden olduklarından, bu işlem

314
başarısızlıkla sonuçlanacaktır. 100 nm por büyüklüğüne sahip filtreden
geçirildiklerinde MLV genelde ortalama olarak çapları 120-140 nm arasında değişen
büyük, tek katmanlı (large unilamellar vesicles (LUV))’i meydana getirir. Bu işlem
sunucunda, elde edilen lipozomların boyutu particle sizer cihazıyla teyit edilir. Bu
ortalama partikül boyutu ayrıca lipid içeriğine bağlıdır ancak son derece
tekrarlanabilir bir prosedürdür.
Şekil E5.3 Çok katmanlı veziküllerin hazırlanması enasında oluşan fosfolipid yapıları.
Kaynak: http://www.avantilipids.com

315
Şekil E5.4 MLV’lerin hazırlanmasındaki üç aşamanın şematik gösterimi. 1: Önceden
oluşturulan kurutulmuş ince tabaka lipidlere sıvı fazın katılması; 2: Balon jojeyi
çalkalama ve ısıtma neticesinde lipid filmin erimesi ve çözülmesi; 3: Ekvilibre edilmiş
MLVlerin sütsü süspansyonu. Kaynak: http://www.avantilipids.com

316
Şekil.E5.5: Makromoleküler enkapsülasyon amaçlı tasarlanan çeşitli lipozomlar.
Kaynak: www.unizh.ch/ onkwww/lipos.htm

317
Şekil E5.6: Çeşitli Lipozom boyutları. Kaynak: www.azonano.com/details.
asp?ArticleID=1243
EK-10 TEK KATMANLI LİPOZOMLARIN HACMİ, ÇAPI, SAYILARI, ALANLARI
VE LİPİD AĞIRLIĞI ARASINDAKI İLİŞKİYİ GÖSTEREN BİR NOMOGRAM69
69 Johnes, G. R. and Cossins, A. R., Lposomes-A Practical Approach, IRL Pres, (1990)’tan alınmıştır.

318

319
Ek-18 ÖZGEÇMİŞ

320
CURRICULUM VITAE
Name and Surname: Erhan Süleymanoğlu
Date of Birth: 22.03.1967 in Sofia, Bulgaria.
Citizenship: The Netherlands, Bulgaria and Turkey (Trial Citizenship)
Home Address:
Gazi Mahallesi,
Polatlı Caddesi, No:115/5,
Yenimahalle,
06560-Ankara,
Turkey
Tel.: (00-90)-5423527265
Home tel.: (00-90-312)-211-19-47
E-mail: [email protected],
http://erhan.boom.ru
Non-academic Affiliations:
November 1992-January 1993: Official translator and consultant on Russian
Federation of Sutek Trade and Construction Co. Ltd.- Ankara, Turkey.
1993-present: Authorized translator of 9th Notarial Service - Ankara, Turkey.
2001-2002: Foreign Affairs Officer of DEM İLÂÇ PHARMACEUTICALS Co. Ltd. -
official distributor of Instituto Grifols, S.A. (Barcelona, Spain) in Turkey and advisor
on authorization of medicinal products to be submitted to The Turkish Ministry of
Health for further approval.
2004-present: Member of The Editorial Board of Electronic Journal of Biomedicine
(University Hospital Yägue-Burgos, Spain): http://biomed.uninet.edu

321
2006-present: Member of The Editorial Board of International Journal of
Biomedical Sciences (www.ijbs.org).
Primary Education:
1974-1985: Russian Language High School - Sofia and in parallel UNESCO English
Language School - Sofia, Bulgaria.
Academic Achievements and Research Experience:
1989: B.Sc. Programme in Biotechnology - The Biotechnology Centre of The Sofia
University - Sofia, Bulgaria.
1992: B.Sc. in Biology - Middle East Technical University, Department of Biology -
Ankara, Turkey.
1995: M.Sc. in Biochemistry, Institute of Natural and Applied Sciences, Department
of Biochemistry - Middle East Technical University - Ankara, Turkey (M.Sc. Thesis
Topic: "Effect of Metal Ions on Interaction Between Nucleic Acid, Protein (Melittin)
and Drug (Tamoxifen) with Model Membranes").
Predoctoral Training:
October 1996-March 1997: University of Vienna, Medical Faculty, VIENNA
BIOCENTER, Molecular Virology Group - Vienna, Austria (Research Topic: "Study of
the Interaction Between Neutralizing Antibodies with Human Rhinoviruses from the
Minor Receptor Group").
2001: Ph.D. in Biochemsitry, Institute of Biochemistry, University of Vienna, Medical
Faculty, VIENNA BIOCENTER (Ph.D. Thesis Topic: "Cloning and Characterization of
Human Heterogeneous Nuclear Ribonucleoprotein (hnRNP) A3 - a Novel Member of
the hnRNP A/B Family"; Abstract available at: www.arcs.ac.at/dissdb/rn036425).

322
Post-doctoral Research:
January 2000-September 2001: Swammerdam Institute for Life Sciences, Section of
Molecular Cytology, University of Amsterdam, BioCentrum Amsterdam -
Amsterdam, The Netherlands (Research Topic: "Prokaryotic DNA Organization:
Balancing Physical and Physiological Forces").
Assistantships and Other Research Appointments:
1. September 1994-April 1996: Research assistantship at Gazi University, Medical
Faculty, Department of Biophysics - Ankara, Turkey (Duties included teaching
general physics with laboratory section to medical students and performing research
on effects of electromagnetic fields on living cells and tissues).
2. January-February 1997: Guest scientist at University of Bergen, Faculty of
Medicine, Pre-clinical Institutes, Institute of Biochemistry and Molecular Biology -
Bergen, Norway (Research Topic: "Acyl Chain Specificity in Phospholipid Metabolism
and Signal Transduction in Brain" - The European Union supported project).
3. June 1997: Guest scientist at J.A. Comenius University, Faculty of Pharmacy,
Department of Physical Chemistry, Biophysics Section - Bratislava, Slovakia, and at
The Slovak Academy of Sciences, Institute of Experimental Physics, Department of
Biophysics - Kosice, Slovakia (Research Topic: "Differential Scanning Calorimetric
Studies on the Effect of Metal Ions on Interaction Between Phospholipid Membranes
with Nucleic Acids, Immunomodulators and Surfactants").
4. April-May 1999: Guest researcher at Institut Curie, UMR-216 du CNRS - Genetique
et Chemie des Genomes Eucaryotes; bat. 110, Centre Universitair Paris - Sud, Orsay
Cedex - France (Research Topic: "Circular Dichroism and Fluorescence Spectroscopy
of RNA-Protein Folding Patterns in Ribonucleoprotein Particles and Their Role in
Autoimmune Diseases").
5. March 1999: Guest researcher at Institute of Chemical Technology, NMR
Laboratory - Prague, Czech Republic (Research Topic: "NMR Spectroscopy of RNA-
Protein Recognition in hnRNP particles - Implications for Disease Specific Epitope
Formation").

323
6. October 2000 - February 2001: Guest researcher at Department of Molecular Cell
Physiology and Mathematical Biochemistry, BioCentrum Amsterdam, Free
University of Amsterdam - Amsterdam, The Netherlands (Research Topic: "Physics
and Molecular Physiology of DNA Supercoiling").
Scholarships Awarded:
1. 1985-1989: Scholarship of The Chemicopharmaceutical Research Institute
(НИХФИ-СОФИЯ) - Sofia, Bulgaria.
2. 1993-1995: Scholarship of The Technical and Research Council of Turkey
(TÜBİTAK) for research on protein folding and denaturation mechanisms of citrate
synthase - in collaboration with University of Bath - United Kingdom.
3. 1996-1999: Ph. D. Scholarship of The Austrian Government.
Publications:
1. Severcan, F., Süleymanoğlu, E., Boyar, H. Turbidity studies of the effect of divalent
cations on Tamoxifen - model membrane interactions. Biochem. Soc. Trans., 25 (3):
493S, (1997).
2. Huber, W., Süleymanoğlu, E., et. al. The hnRNP-D/AUF proteins - a novel group of
autoantigens in rheumatology - In: Pathogenesis and Diagnostic Relevance of
Autoantibodies; Report on the 4th Dresden Symposium on Autoantibodies, hold in
Dresden - Germany on Octomber 21-24, (1998); Conrad, K., Humbel, R-L., Muerer,
M., Shoenfeld, Y., E.M.T., (Eds.) PABST Publishers.
3. Skriner, K., Huber, W., Süleymanoğlu, E., Smolen, J., Steiner, G. Are mRNA decay
complexes target structures in systemic autoimmune diseases? Arthritis and
Rheumatism, 43 (9), Suppl. S, Sept. (2000).
4. Cunha, S., Odijk, T., Süleymanoğlu, E., Woldringh, C.L. Isolation of Escherichia
coli nucleoid. Biochimie, Feb., 83 (2): 149-154, (2001).

324
5. Süleymanoğlu, E. Electrorelease of Escherichia coli nucleoids. Folia Microbiol. 47
(4): 365-370, (2002).
6. Süleymanoğlu, E. Fluorescence microscopy of condensed DNA conformations of
bacterial cells. J. Microbiol. 40 (4): 319-326, (2002).
7. Steiner, G., Cervenak, Z., Skriner, K. and Süleymanoğlu, E. Molecular cloning,
expression and immunochemical properties of new human hnRNP A/B type
autoantigen, Biomedica, 19: 8-17, Jul.-Dec. (2003).
8. Süleymanoğlu, E. Molecular phylogenetics and functional evolution of major RNA
recognition domains of recently cloned and characterized autoimmune RNA-binding
particle, Geno., Prot. & Bioinfo. 1 (4): 310-320, (2003).
9. Süleymanoğlu, E. Circular dichroism and fluorescence spectroscopic study of RNA-
protein folding patterns in human hnRNP A3 and their implications in human
autoimmune diseases. Prog. Biochem. Biophys. 31 (3): 219-224, (2004).
10. Süleymanoğlu E. Preparation and phase behaviour of surface-active
pharmaceuticals: self-assembly of DNA and surfactants with membranes. Differential
adiabatic scanning microcalorimetric study. Il Farmaco, Aug; 60 (8): 701-710,
(2005).
11. Süleymanoğlu E. Phoshpolipid-Nucleic Acid Recognition: Energetics of DNA-
Mg2+-Phosphatidylcholine Ternary Complex Formation and its Further Compaction
as a Gene Delivery Formulation, PDA J. Pharmaceut. Sci. Technol. 60 (4): 218-231,
(2006).
12. Phoshpolipid-Nucleic Acid Recognition: Developing an Immobilized Liposome
Chromatography for DNA Separation and Analysis, PDA J. Pharmaceut. Sci.
Technol. 60 (4): 232-239, (2006).
15. Manavbasi Y, Suleymanoglu E., Probing Taxol (TM) (Paclitaxel)-cell membrane
interactions with Langmuir-Blodgett monolayer, Fourier transform infrared
spectroscopy and differential scanning calorimetry, FEBS JOURNAL 273: 354-354
Suppl. 1, JUN (2006).

325
16. Manavbaşı Y. and Süleymanoğlu E., Nucleic acid-phospholipid recognition:
Fourier transform infrared spectrometric characterization of ternary phospholipid-
inorganic cation-DNA complex and its relevance to chemicopharmaceutical design of
nanometric liposome based gene delivery formulations. Arch Pharm Res., Aug; 30
(8): 1027-40, (2007).
17. Karl Skriner, Wolfgang Hueber, Erhan Süleymanoglu, Elisabeth Höfler, Veit Kren,
Josef Smolen and Günter Steiner, The Regulator of TNFα mRNA Decay AU-rich
Element Binding Factor 1 is Targeted by Autoantibodies of Patients with Systemic
Rheumatic Diseases, (accepted for publication in Rheumatism and Arthritis-2007).
Books:
Hayriye S. Yenisoy, Nergiz Aksoy and Erhan Süleymanoglu, Contemprorary
Bulgarian, published by Kurmay Yayınevi (www.kurmayyayinevi.com)-2007.
Conferences and Workshops Attended:
1. Intensive Course on Protein Structure, Function and Stability, April 18-22, 1994,
Ankara - Turkey; Organized by Middle East Technical University - Ankara, Turkey
and University of Bath - United Kingdom.
2. Turbidimetric Study of Interaction Between Polyribonucleotides,
Phosphatidylcholine Vesicles and Metal Ions, NATO Advanced Study Institute:
Trafficking of Intracellular Membranes: From Molecular Sorting to Membrane
Fusion, June 19-30, 1994, Espinho, Portugal.
3. Spectrophotometric Study of the Interaction Between Polyribonucleotides,
Phosphatidylcholine Vesicles and Metal Ions, VIth National Biophysical Congress,
September 28-30, 1994, Silivri - Istanbul, Turkey.
4. Calcium/Magnesium Specificity in Modulation of Phospholipid Phase Transition
Induced by Tamoxifen, NATO Advanced Study Institute: Molecular Dynamics of
Biomembranes, June 19 - July 1, 1995, Cargese, Corsica, France.
5. Self-Assembly of Lipid Vesicles and Polyribonucleotides - An Inorganic
Perspectives for the Origin of Life, NATO Advanced Study Institute: Solvents and
Self-Organization of Polymers, July 23, 1995, Belek, Antalya, Turkey.

326
6. Self-Assembly of Lipid Droplets and Polyribonucleotides or Primordial Soup
Revisited, ISSOL'96: International Conference on The Origin of Life, July 7 - 12,
1996, Orleans, France.
7. Self-Association of Melittin with Model Membranes, Winter School of Biophysics:
Biophysical Aspects of Protein Folding; Organized by Stockholm Graduate School in
Biophysics, as a joint programme between The Karolinska Institute, The Royal
Institute of Technology and Stockholm University, April 1 - 4, 1997.
8. International Workshop on Modern Spectroscopic Techniques in Biophysics, June
1 - 5, 1998, Neptun, Romania.
9. 1998 Joint Meeting and Symposium of Clinical Biochemistry: New Trends in
Clinical Biochemistry of Transplantation, Artis Tower Hotel - Vienna, Austria;
Organized by Austrian Society for Clinical Chemistry and Instituto di Semiotica
Medica, Universita degli Studi Padova - Italia and Society Italiana Di Biochemica
Clinica.
10. The hnRNP-D/AUF Proteins - A Novel Group of Autoantigens in Rheumatology,
National Scientific Meeting, Arthritis and Rheumatism - Abstract Supplement, Vol.
41, No: 9, Sept. S349, November 8-12, 1998, San Diego, California, U.S.A.
11. The hnRNP-D/AUF Proteins - a Novel Group of Autoantigens in SLE, MCTD and
RA, XVIII European Workshop for Rheumatology Research, March 12 - 15, 1998,
Athens, Greece.
12. ISPPP98 - 18th International Symposium on the Separation and Analysis of
Proteins, Peptides and Polynucleotides, November 1 - 4, 1998, Hilton Hotel - Vienna,
Austria.
13. The hnRNP A/B type proteins - major molecular targets in rheumatic
autoimmune disease, RNA'99: The 4th Annual Meeting of the RNA Society, 23-27
June, 1999, University of Edinburgh, Scotland.
14. Cloning of a novel hnRNP A/B type protein highly homologous to hnRNP-A3
from X. laevis, RNA'99: The 4th Annual Meeting of the RNA Society, 23-27 June,
1999, University of Edinburgh, Scotland.

327
15. Identification of a novel autoantigen highly related to hnRNP-A2/RA33, The
19th European Workshop for Rheumatology Research, 1999, Oslo, Norway.
16. Epitope Mapping of hnRNP-D/AUF-1 proteins, The 19th European Workshop for
Rheumatology Research, 1999, Oslo, Norway.
17. The hnRNP-D/AUF-1 Proteins - Novel Autoantigens in Rheumatology, 4th
European Conference on Systemic Lupus Erythematosus; Organized by Vienna
Academy of Postgraduate Medical Education and Research, January 23 - 26, 1999,
Vienna, Austria.
18. Thermodynamics of nucleic acid-membrane-self-assembly as a precellular
precursors and the emerging role of Ca2+ and Mg2+ as condensing agents, 12th
International Conference on the Origin of Life and 9th ISSOL Meeting, 11-16, 1999,
San Diego, California, U.S.A.
19. Are mRNA stability complexes target structures in systemic autoimmunity?,
14th European Immunology Meeting, EFIS, Sept. 23-27, 2000, Poznań, Poland.
20. Confocal Light Microscopy: Fundamentals and Biological Applications;
BioCentrum Course with practical training; University of Amsterdam, May 8-12,
2000, Amsterdam, The Netherlands.
21. Visualizing Cellular Processes, 8 June, 2000, Free University of Amsterdam,
Amsterdam, The Netherlands.
22. Analysis of Compaction Forces and Supercoil Content of Isolated E. coli
nucleoids, International Summer School on DNA and Chromosomes: Physical and
Biological Approaches, July 31 - August 12, 2000, Cargese, Corsica, France.
23. Analysis of DNA Compaction within the Bacterial Cell, EMBO Workshop: The
Cell Cycle and Nucleoid Organization in Bacteria; Netherlands Institute for Sea
Research (NIOZ), September 2-6, 2000, Island of Texel, The Netherlands.
24. AiK (The Association for Physics Students) Symposium: Complex Systems,
November 3, 2000, Free University of Amsterdam, Amsterdam, The Netherlands.

328
25. Fall Symposium of The Dutch Society for Biochemistry and Molecular Biology:
Into the Living Cell and Beyond, November 7, 2000, BioCentrum Amsterdam - Free
University of Amsterdam, Amsterdam, The Netherlands.
26. Autoantibodies to the mRNA destabilizing protein hnRNP-D/AUF1 in patients with systemic autoimmune diseases, 21st European Workshop for Rheumatology Research, Parkhotel Schönbrunn, Vienna, Austria. 1-4 March 2001, published in Arthritis Res. 2001, 3(Suppl A): P012.
26. Symposium on Live Cell Imaging; Organized by PerkinElmer Life Sciences and
hold at Institute of Molecular Pathology, April 10, 2001, Vienna, Austria.
27. IMMPC - 1: First International Meeting on Medical and Pharmaceutical
Chemistry, September 25-28, 2002, Hotel Ankara Dedeman - Ankara, Turkey.
28. Circular Dichroism and Fluorescence Spectroscopy of RNA - Protein Folding
Patterns and Their Role in Autoimmune Diseases, The Inaugural Austral - Asian
Biospectroscopy Conference: Organized by Monash University - Australia and hold at
the Suranaree University of Technology, Nakhon Ratchasima (Korat) - Thailand,
February 3-7, 2003.
29. Fluorescence Microscopy of Polymer - Induced Phase Separation and Divalent
Cation - Dependent Compaction and Structural Integrity of Bacterial Chromosome,
Bionanotechnology - EuroConference on Biomolecular Devices - Granada, Spain, 9-
14 July, 2003.
30. Macromolecular Crowding and Bacterial DNA Cytoarchitecture - Conference
for Young Scientists, Ph. D. Students and Students on Moleculur Biology and
Genetics - Dedicated to 50 years of the double helix and 30 anniversary of the
Institute of Molecular Biology and Genetics of National Academy of Sciences of
Ukraine, 25 - 27 September 2003, Kiev-Ukraine.
31. Adiabatic Differential Scanning Microcalorimetric Study of Divalent Cation -
Induced Neutral Lipid - DNA Nanocondensate Formation Assambled for Gene
Delivery - Conference for Young Scientists, Ph. D. Students and Students on
Moleculur Biology and Genetics - Dedicated to 50 years of the double helix and 30

329
anniversary of the Institute of Molecular Biology and Genetics of National Academy
of Sciences of Ukraine, 25 - 27 September 2003, Kiev-Ukraine.
32. Light Microscopy of Polyethylene Glycol-Induced Phase Separation and
Divalent Cation-Dependent Compaction of Prokaryotic Nucleoids - ELSO-2003:
European Life Scientist Organization - 2003, Dresden, Germany.
33. On-line Dissolution, Advanced Detection Methods and Authomated Method
Development. Workshop organized by Waters Corporation (U.S.A.), Gazi University,
Faculty of Pharmacy 11.03.2004, Ankara, Turkey.
34. Biophysical Aspects of RNA-protein Folding Features in A/B Type hnRNP
Proteins and Their Implications in Human Systemic Autoimmune Pathology, 30th
FEBS Congress and 9th IUBMB Conference - The Protein World, 2 - 7 July, 2005,
Budapest, Hungary.
35. Workshops of Middle East Technical University-Central Laboratory, 1.
Rheological Characterization of Materials; 2. Particle Size Measurement Using
Laser Diffraction Technique; 3. Particle Size Measurement Using Dynamic Light
Scattering Techniques and Zeta Potential Measurement; 4. Thermal Analysis
Techniques, 30.11.2005-21.12.2005, Ankara, Turkey.
36. Thermal Analysis and Rheological Applications in Polymer Field, Workshop
organized by TA Instruments (UK), Gazi University, Faculty of Pharmacy, 21.12.2005,
Ankara, Turkey.
37. Probing Taxol™ (Paclitaxel)-Cell Membrane Interations with Langmuir-Blodgett Monolayer, Fourier Transform Infrared Spectroscopy and Differential Scanning Calorimetry, 1st Multidisciplinary Cancer Research Symposium, Uludağ-Bursa, Turkey, 2006.
38. Preparation and Physicochemical Characterization of Biomolecular DNA-
Zwitterionic Phospholipid Self-Assemblies and Their Implications in
Chemicopharmaceutical Design of Liposome Based Gene Delivery
Nanoformulation, NanoTR-II: NanoScience and NanoTechnology 2006, 3-5 May
2006, Middle East Technical University, Ankara, Turkey.
39. Assessment of Structures of Poly(ribo)nucleotide-Phospholipid Self-Assemblies and Their Implıcations in Gene Transfection and DNA Chromatography, 8th

330
International Symposium on Pharmaceutical Sciences (ISOPS-8), 13-16 June, 2006, Ankara, Turkey.
40. Probing Taxol™ (Paclitaxel)-Cell Membrane Interations with Langmuir-
Blodgett Monolayer, Fourier Transform Infrared Spectroscopy and Differential
Scanning Calorimetry, FEBS-2006, Istanbul, Turkey.
41. METU-CENTER WORKSHOP on Modern Electron Paramagnetic Resonance
(EPR) Spectroscopy, 6-9 November 2006, Middle East Technical University (METU)
Central Laboratory.
42. Fluorescence Microscopy and Interfacial Characterization of DNA
Condensation, Compaction and Release from Stable Virus-Resembling
Phospholipid-Nucleic Acid Self-Assemblies and Their Implications as Non-Viral
Gene Delivery Nanopharmaceuticals, NANO-TRIII 2007-Bilkent University, Ankara,
Turkey.
43. An FTIR Spectrometric Study of the Physicochemical Characteristics of Nucleic
Acid-Phospholipid Nanoformulations Designed for Gene Delivery at the Molecular
Level, X. National Congress of Spectroscopy, 04-07 July 2007, Izmir Institute of
Technology, Urla, Izmir.
44. Applying multilayer films and planar lipid bilayers (BLMs) for designing
nucleic acid nanobiosensors: some examples of bioengineering at the molecular
level, NANOMAT 2007, International Workshop on nanobiotechnology & Genome
Technologies, October 31-November 03, 2007, Antalya, Turkey.
Memberships:
Member of The New York Academy of Sciences, The Biophysical Society, The British
Biophysical Society, The Society for Mathematical Biology, The European Society of
Neurochemistry, The Austrian Society of Clinical Chemistry, The International
Society for the Study of the Origin of Life, Metbio™.

331
Fluency in Foreign Languages:
Excellent fluency in English, Russian, Bulgarian, Turkish and week knowledge of
German.