endogenous retinoid x receptor ligands in mouse ... · retinoid x receptors (rxrs) are members of...
TRANSCRIPT

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
B IOCHEM ISTRY
1Departmentof InternalMedicine,WashingtonUniversitySchoolofMedicine, St. Louis,MO63110, USA. 2Diabetic Cardiovascular Disease Center, Washington University School ofMedicine, St. Louis,MO63110,USA. 3Departmentof Biochemistry andMolecular Biophysics,Washington University School of Medicine, St. Louis, MO 63110, USA. 4Myocardial Patho-physiology Area, Centro Nacional de Investigaciones Cardiovasculares Carlos III, Madrid28029, Spain.*Corresponding author. Email: [email protected]
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
Copyright © 2017
The Authors, some
rights reserved;
exclusive licensee
American Association
for the Advancement
of Science. No claim
to original U.S.
Government Works
Dow
n
Endogenous retinoid X receptor ligands in mousehematopoietic cellsHaixia Niu,1 Hideji Fujiwara,2 Orsola di Martino,1 Gayla Hadwiger,1 Thomas E. Frederick,3
María P. Menéndez-Gutiérrez,4 Mercedes Ricote,4 Gregory R. Bowman,3 John S. Welch1*
The retinoid X receptora (RXRA) has been implicated in diverse hematological processes. To identify natural ligands ofRXRA that are present in hematopoietic cells, we adapted an upstream activation sequence–green fluorescent protein(UAS-GFP) reporter mouse to detect natural RXRA ligands in vivo. We observed reporter activity in diverse types ofhematopoietic cells in vivo. Reporter activity increased during granulocyte colony-stimulating factor (G-CSF)–inducedgranulopoiesis and after phenylhydrazine (PHZ)–induced anemia, suggesting the presence of dynamically regulatednatural RXRA ligands in hematopoietic cells. Mouse plasma activated Gal4-UAS reporter cells in vitro, and plasma frommice treatedwithG-CSF or PHZ recapitulated the patterns of reporter activation thatweobserved in vivo. Plasma frommicewithdietary vitaminAdeficiencyonlymildly reducedRXRA reporter activity,whereasplasma frommiceona fattyacid restriction diet reduced reporter activity, implicating fatty acids as plasma RXRA ligands. Through differentialextraction coupledwithmass spectrometry, we identified the long-chain fatty acid C24:5 as a natural RXRA ligand thatwas greatly increased in abundance in response to hematopoietic stress. Together, these data suggest that naturalRXRA ligands are present and dynamically increased in abundance in mouse hematopoietic cells in vivo.
load
on June 14, 2019http://stke.sciencemag.org/
ed from
INTRODUCTIONRetinoid X receptors (RXRs) are members of the nuclear receptor su-perfamily (1). Like other nuclear receptors, the transcriptional activity ofRXRs is ligand-dependent. Ligand binding results in a conformationalshift of the terminala helix [activation function 2 (AF2) domain], whichdisplaces bound corepressors and facilitates the binding of coactivators.
Diverse molecules have been implicated as natural RXR ligands, in-cluding both retinoic acids and fatty acids (2). It is unknown whetherany of these are present in hematopoietic cells in physiologically rele-vant quantities or whether they are dynamically regulated during hema-topoietic stress. The vitamin A derivative 9-cis retinoic acid (9-cis-RA)has been described as a natural RXR ligand (3, 4), yet several groupshave reported 9-cis-RA to be absent or below detectable limits in testis,liver, heart, lung, and serum (5, 6). Another vitaminAmetabolite, 9-cis-13,14-dihydroretinoic acid (9-cis-13,14-DHRA), was reported to be anendogenous RXR ligand inmouse serum, brain, and liver, although it isunclear whether it is present in hematopoietic cells (7). Using a lucifer-ase reporter assay, Lengqvist et al. (8) showed that unsaturated fattyacids such as docosahexaenoic acid (DHA; C22:6), docosapentaenoicacid (DPA; C22:5), and arachidonic acid (AA; C20:4) activate RXRa(RXRA) in vitro, whereas saturated fatty acids such as arachidic acid(C20:0) and stearic acid (C18:0) do not.
Three separate genes encode three RXR isoforms—RXRA, RXRb(RXRB), and RXRg (RXRG)—at least one of which is present in everymammalian cell (9). These subtypes are highly conserved, with nearlyidentical ligand-binding pocket conformations (1). RXRs bind to DNAas either a homodimer (10) or a heterodimer (1); the heterodimericpartners of RXR include retinoic acid receptors (RARs), thyroid hormonereceptors (TRs), the vitaminD receptor (VDR), peroxisome proliferator–activated receptors (PPARs), liver X receptors, the pregnane X receptor,
and the constitutive androstane receptor (11–15). By heterodimerizingwiththese partners, RXRs participates in diverse essential biological processes,such as development, metabolism, cell differentiation, and cell death.
RXRs play important roles in hematopoiesis. RXRA abundance in-creases upon monocytic differentiation (16). Ectopic overexpression ofRXRA in hematopoietic stem and progenitor cells inhibits granulopoiesisby impairing proliferation and differentiation, whereas expression of adominant negative form of RXRApromotes the generation of late-stagegranulocytes in vitro (17). Rxra binds to the promoter of Epo, the geneencoding the cytokine erythropoietin (EPO), and promotes its tran-scription during the embryonic day 9.5 (E9.5) to E11.5 phase of fetalliver erythropoiesis before subsequently being supplanted by hepatocytenuclear factor 4–dependent transcription of Epo from E11.5 onward(18). Rxrs also participate in osteoclast differentiation and postnatalbone remodeling (19). Conditional deletion of both Rxra and Rxrb inmouse hematopoietic cells (Rxrg is not expressed in hematopoietic cells)generates giant, nonresorbing osteoclasts, increases bone mass in malemice, and protects female mice from osteoporotic bone loss after ovari-ectomy. No information is available about the presence or distributionof natural ligands of RXRA in hematopoietic cells.
Here, we demonstrate that natural RXRA ligands are present inmouse hematopoietic cells and plasma in vivo and are predominantlyactive inmyeloid cells.We find that concentrations of RXRA ligands aredynamically increased in response tomyeloid stress, such as exposure togranulocyte colony-stimulating factor (G-CSF) and phenylhydrazine(PHZ). Moreover, we identify the long-chain fatty acid C24:5 as an en-dogenousRXRA ligand that undergoes dynamic increases in concentra-tion during mouse hematopoietic stress.
RESULTSAdaptation of Gal4–upstream activation sequence reporterto detect RXRA ligands in vivoTo determine whether natural RXRA ligands are present in hemato-poietic cells in vivo, we used transgenic upstream activation sequence–green fluorescent protein (UAS-GFP) reporter mice (20). UAS promotersequences are recognized by the yeast transcription factor Gal4 and are
1 of 12
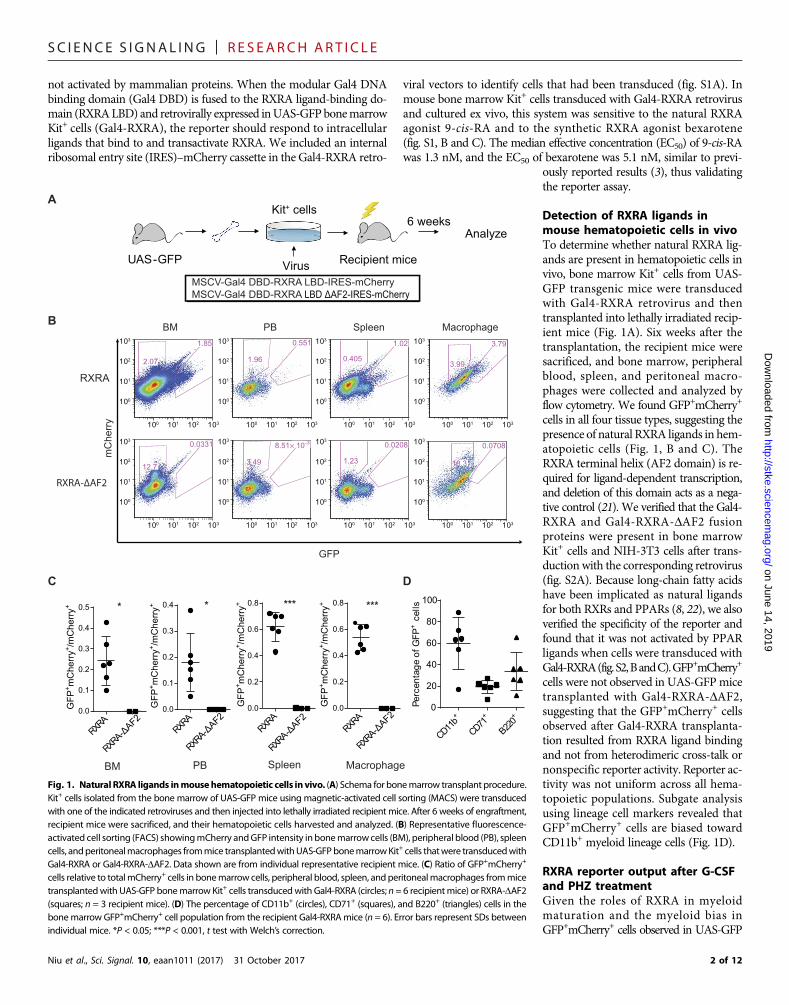
SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
not activated by mammalian proteins. When the modular Gal4 DNAbinding domain (Gal4 DBD) is fused to the RXRA ligand-binding do-main (RXRALBD) and retrovirally expressed inUAS-GFPbonemarrowKit+ cells (Gal4-RXRA), the reporter should respond to intracellularligands that bind to and transactivate RXRA. We included an internalribosomal entry site (IRES)–mCherry cassette in the Gal4-RXRA retro-
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
viral vectors to identify cells that had been transduced (fig. S1A). Inmouse bone marrow Kit+ cells transduced with Gal4-RXRA retrovirusand cultured ex vivo, this system was sensitive to the natural RXRAagonist 9-cis-RA and to the synthetic RXRA agonist bexarotene(fig. S1, B and C). The median effective concentration (EC50) of 9-cis-RAwas 1.3 nM, and the EC50 of bexarotene was 5.1 nM, similar to previ-
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
ously reported results (3), thus validatingthe reporter assay.
Detection of RXRA ligands inmouse hematopoietic cells in vivoTo determine whether natural RXRA lig-ands are present in hematopoietic cells invivo, bone marrow Kit+ cells from UAS-GFP transgenic mice were transducedwith Gal4-RXRA retrovirus and thentransplanted into lethally irradiated recip-ient mice (Fig. 1A). Six weeks after thetransplantation, the recipient mice weresacrificed, and bone marrow, peripheralblood, spleen, and peritoneal macro-phages were collected and analyzed byflow cytometry. We found GFP+mCherry+
cells in all four tissue types, suggesting thepresence of natural RXRA ligands in hem-atopoietic cells (Fig. 1, B and C). TheRXRA terminal helix (AF2 domain) is re-quired for ligand-dependent transcription,and deletion of this domain acts as a nega-tive control (21). We verified that the Gal4-RXRA and Gal4-RXRA-DAF2 fusionproteins were present in bone marrowKit+ cells and NIH-3T3 cells after trans-duction with the corresponding retrovirus(fig. S2A). Because long-chain fatty acidshave been implicated as natural ligandsfor both RXRs and PPARs (8, 22), we alsoverified the specificity of the reporter andfound that it was not activated by PPARligands when cells were transduced withGal4-RXRA(fig.S2,BandC).GFP+mCherry+
cells were not observed in UAS-GFPmicetransplanted with Gal4-RXRA-DAF2,suggesting that the GFP+mCherry+ cellsobserved after Gal4-RXRA transplanta-tion resulted from RXRA ligand bindingand not from heterodimeric cross-talk ornonspecific reporter activity. Reporter ac-tivity was not uniform across all hema-topoietic populations. Subgate analysisusing lineage cell markers revealed thatGFP+mCherry+ cells are biased towardCD11b+ myeloid lineage cells (Fig. 1D).
RXRA reporter output after G-CSFand PHZ treatmentGiven the roles of RXRA in myeloidmaturation and the myeloid bias inGFP+mCherry+ cells observed in UAS-GFP
1.85
2.07
BM0.551
1.96
PB1.02
0.405
Spleen3.79
3.99
Macrophage
8.51× 10–3
3.49
0.0331
12.7
0.0208
1.23
0.0708
18.3
RXRA
RXRA-ΔAF2
GFP
mC
herr
y
A
UAS-GFP Recipient mice
Analyze6 weeks
Virus
Kit+ cells
B
RXRA
RXRA-Δ
AF20.0
0.1
0.2
0.3
0.4
0.5
GFP
+ mC
herr
y+ /mC
herr
y+
RXRA
0.0
0.1
0.2
0.3
0.4
RXRA
0.0
0.2
0.4
0.6
0.8
RXRA
0.0
0.2
0.4
0.6
0.8
BM PB Spleen Macrophage
C
RXRA-Δ
AF2
RXRA-Δ
AF2
RRX
A-ΔAF2
* * *** ***
CD11b+
CD71+
B220+
0
20
40
60
80
100
Perc
enta
geof
GFP
+ce
lls
D
MSCV-Gal4 DBD-RXRA LBD-IRES-mCherryMSCV-Gal4 DBD-RXRA LBD ΔAF2-IRES-mCherry
GFP
+ mC
herr
y+ /mC
herr
y+
GFP
+ mC
herr
y+ /mC
herr
y+
GFP
+ mC
herr
y+ /mC
herr
y+
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
Fig. 1. Natural RXRA ligands inmousehematopoietic cells invivo. (A) Schema for bonemarrow transplant procedure.Kit+ cells isolated from the bone marrow of UAS-GFP mice using magnetic-activated cell sorting (MACS) were transducedwith one of the indicated retroviruses and then injected into lethally irradiated recipient mice. After 6 weeks of engraftment,recipient mice were sacrificed, and their hematopoietic cells harvested and analyzed. (B) Representative fluorescence-activated cell sorting (FACS) showingmCherry andGFP intensity inbonemarrowcells (BM), peripheral blood (PB), spleencells, andperitonealmacrophages frommice transplantedwithUAS-GFPbonemarrowKit+ cells thatwere transducedwithGal4-RXRA or Gal4-RXRA-DAF2. Data shown are from individual representative recipient mice. (C) Ratio of GFP+mCherry+
cells relative to totalmCherry+ cells in bonemarrow cells, peripheral blood, spleen, and peritonealmacrophages frommicetransplantedwith UAS-GFP bonemarrow Kit+ cells transducedwith Gal4-RXRA (circles; n = 6 recipientmice) or RXRA-DAF2(squares; n = 3 recipient mice). (D) The percentage of CD11b+ (circles), CD71+ (squares), and B220+ (triangles) cells in thebonemarrow GFP+mCherry+ cell population from the recipient Gal4-RXRAmice (n = 6). Error bars represent SDs betweenindividual mice. *P < 0.05; ***P < 0.001, t test with Welch’s correction.
2 of 12
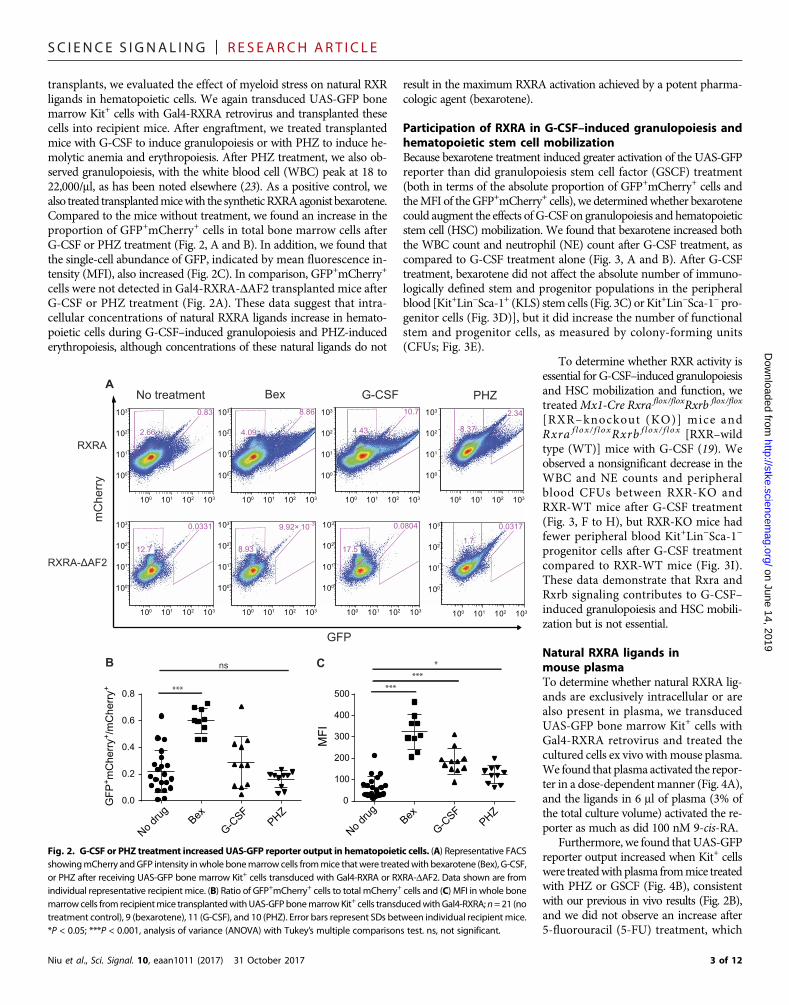
SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
D
transplants, we evaluated the effect of myeloid stress on natural RXRligands in hematopoietic cells. We again transduced UAS-GFP bonemarrow Kit+ cells with Gal4-RXRA retrovirus and transplanted thesecells into recipient mice. After engraftment, we treated transplantedmice with G-CSF to induce granulopoiesis or with PHZ to induce he-molytic anemia and erythropoiesis. After PHZ treatment, we also ob-served granulopoiesis, with the white blood cell (WBC) peak at 18 to22,000/ml, as has been noted elsewhere (23). As a positive control, wealso treated transplantedmicewith the syntheticRXRAagonist bexarotene.Compared to the mice without treatment, we found an increase in theproportion of GFP+mCherry+ cells in total bone marrow cells afterG-CSF or PHZ treatment (Fig. 2, A and B). In addition, we found thatthe single-cell abundance of GFP, indicated by mean fluorescence in-tensity (MFI), also increased (Fig. 2C). In comparison, GFP+mCherry+
cells were not detected in Gal4-RXRA-DAF2 transplanted mice afterG-CSF or PHZ treatment (Fig. 2A). These data suggest that intra-cellular concentrations of natural RXRA ligands increase in hemato-poietic cells during G-CSF–induced granulopoiesis and PHZ-inducederythropoiesis, although concentrations of these natural ligands do not
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
result in the maximum RXRA activation achieved by a potent pharma-cologic agent (bexarotene).
Participation of RXRA in G-CSF–induced granulopoiesis andhematopoietic stem cell mobilizationBecause bexarotene treatment induced greater activation of the UAS-GFPreporter than did granulopoiesis stem cell factor (GSCF) treatment(both in terms of the absolute proportion of GFP+mCherry+ cells andtheMFI of theGFP+mCherry+ cells), we determinedwhether bexarotenecould augment the effects of G-CSF on granulopoiesis and hematopoieticstem cell (HSC) mobilization. We found that bexarotene increased boththe WBC count and neutrophil (NE) count after G-CSF treatment, ascompared to G-CSF treatment alone (Fig. 3, A and B). After G-CSFtreatment, bexarotene did not affect the absolute number of immuno-logically defined stem and progenitor populations in the peripheralblood [Kit+Lin−Sca-1+ (KLS) stem cells (Fig. 3C) orKit+Lin−Sca-1−pro-genitor cells (Fig. 3D)], but it did increase the number of functionalstem and progenitor cells, as measured by colony-forming units(CFUs; Fig. 3E).
on June 14, 2019http://stke.sciencem
ag.org/ow
nloaded from
To determine whether RXR activity isessential for G-CSF–induced granulopoiesisand HSC mobilization and function, wetreatedMx1-Cre Rxra flox/floxRxrb flox/flox
[RXR–knockout (KO)] mice andRxra f lox/ f loxRxrb f lox / f lox [RXR–wildtype (WT)] mice with G-CSF (19). Weobserved a nonsignificant decrease in theWBC and NE counts and peripheralblood CFUs between RXR-KO andRXR-WT mice after G-CSF treatment(Fig. 3, F to H), but RXR-KO mice hadfewer peripheral blood Kit+Lin−Sca-1−
progenitor cells after G-CSF treatmentcompared to RXR-WT mice (Fig. 3I).These data demonstrate that Rxra andRxrb signaling contributes to G-CSF–induced granulopoiesis and HSC mobili-zation but is not essential.
Natural RXRA ligands inmouse plasmaTo determine whether natural RXRA lig-ands are exclusively intracellular or arealso present in plasma, we transducedUAS-GFP bone marrow Kit+ cells withGal4-RXRA retrovirus and treated thecultured cells ex vivo withmouse plasma.We found that plasma activated the repor-ter in a dose-dependentmanner (Fig. 4A),and the ligands in 6 ml of plasma (3% ofthe total culture volume) activated the re-porter as much as did 100 nM 9-cis-RA.
Furthermore, we found thatUAS-GFPreporter output increased when Kit+ cellswere treatedwithplasma frommice treatedwith PHZ or GSCF (Fig. 4B), consistentwith our previous in vivo results (Fig. 2B),and we did not observe an increase after5-fluorouracil (5-FU) treatment, which
No treatment
100 101 102 103
100
101
102
103 0.83
2.66
8.86
4.09
Bex10.7
4.43
G-CSF
8.37
PHZ
0.0331
12.7
9.92× 10–3
8.93
0.0804
17.5
0.0317
1.7
RXRA
RXRA-ΔAF2
GFP
mC
herr
y
A
B
No drug Bex
G-CSF
PHZ0.0
0.2
0.4
0.6
0.8
GFP
+ mC
herr
y+ /m
Che
rry+
C
0
100
200
300
400
500
MFI
***
ns
******
*
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
100 101 102 103
100
101
102
103
2.34
No drug Bex
G-CSF
PHZ
Fig. 2. G-CSF or PHZ treatment increased UAS-GFP reporter output in hematopoietic cells. (A) Representative FACSshowingmCherry andGFP intensity inwhole bonemarrow cells frommice thatwere treatedwith bexarotene (Bex), G-CSF,or PHZ after receiving UAS-GFP bone marrow Kit+ cells transduced with Gal4-RXRA or RXRA-DAF2. Data shown are fromindividual representative recipient mice. (B) Ratio of GFP+mCherry+ cells to total mCherry+ cells and (C) MFI in whole bonemarrowcells from recipientmice transplantedwithUAS-GFPbonemarrowKit+ cells transducedwithGal4-RXRA; n=21 (notreatment control), 9 (bexarotene), 11 (G-CSF), and 10 (PHZ). Error bars represent SDs between individual recipient mice.*P < 0.05; ***P < 0.001, analysis of variance (ANOVA) with Tukey’s multiple comparisons test. ns, not significant.
3 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
causes global hematopoietic aplasia followed by stem cell expansion andhematopoietic recovery (fig. S3A). As expected, reporter activity was ab-sent in cells transduced with Gal4-RXRA-DAF2 (fig. S3B) and was notobserved in cells transduced with Gal4-RARA (fig. S3C), again exclud-ing the possibility of reporter activation through heterodimeric cross-talk or nonspecific activation of the reporter. Although both G-CSFand PHZ induced granulopoiesis (23) and increasedUAS-GFP reporteractivity in our assays, the response after PHZ treatment was modestlygreater than that after G-CSF treatment (Fig. 4B). Therefore, we focused
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
our subsequent ligand identification efforts primarily on plasma frommice treated with PHZ.
Dietary vitamin A deficiency and fatty acid restrictionTo determine whether the reporter activity induced by plasmamight bedue to vitaminA derivatives (for example, 9-cis-RA or 9-cis-13,14-DHRA)(3, 4, 7), we analyzed plasma from vitamin A–deficient (VAD) mice (24).Before PHZ treatment, we confirmed vitaminAdeficiency in thesemiceusing mass spectrometry (MS) detection of plasma retinol (Fig. 5A).
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
Unexpectedly, activation of theGFP re-porter by plasma from VAD mice wasnot statistically different from activa-tion by plasma fromVAD controlmice(VADC), suggesting that prolonged vi-tamin A deficiency did not cause a re-duction in plasma ligands that activateRXRs in hematopoietic cells. Further-more, plasma from VAD mice treatedwith PHZ and plasma from VADCmice treated with PHZ activated theGFP reporter equally, suggesting thatnatural RXR ligands can be generatedinmouse plasma in the absence of vita-min A (Fig. 5B).
To determine whether the RXRAligands that increase after PHZ treat-ment are fatty acids, such as DHA orAA (8), we fed mice a no-fat (NF) dietor NF control (NFC) diet for 4 weeks, aperiod not associated with sequelae be-cause of essential fatty acid deficiency(25). Unlike vitamin A, nonessentialfatty acids can be synthesized in vivo.After 4 weeks on the NF diet, the plas-ma concentration of palmitoleic acidC16:1 (a common glyceride constitu-ent) was reduced about twofold (Fig. 5C).Unlike plasma frommice fed the VADdiet, plasma from mice fed the NF dietstimulated less GFP activation than didplasma from mice on the control diet(Fig. 5D). This presumably reflects re-duced concentrations of RXR ligandsin the plasma both at baseline and afterPHZ treatment. These data suggest thatnatural RXR ligands in mouse plasmaeither are essential fatty acids presentin the diet or require intake of essentialfatty acids for their synthesis.
Identification of plasmaRXRA ligandsTo identify RXRA ligands by MS, wegenerated a stable 293T cell line thatcontains the UAS-GFP reporter andthe fusion protein Flag–Gal4 DBD–RXRA LBD (293T-FXP). These cellsproduce GFP when treated with eitherplasma or serum and are amenable to
BWBCA NE
***
***
***
***
******
KLS Progenitors
***
*****
CFUC ED
**
**ns ***
*** ns
RXR-WT: Rxraflox/floxXRxrbflox/flox
RXR-KO: Rxraflox/floxXRxrbflox/flox X Mx1-Cre
F G H IWBC NE CFU
**ns ns
ns
Control G-CSF G-CSF + Bex0
10
20
30
40
50
K/μ
l
Control G-CSF G-CSF + Bex0
10
20
30
K/μ
l
Control G-CSF G-CSF + Bex0
5
10
15
20
K/1
0 μl
Control G-CSF G-CSF + Bex0
10
20
30
40
50
K/1
0 μl
Control G-CSF G-CSF + Bex0
20
40
60
80
100
Col
ony
num
ber/1
0 μl
RXR-KO
RXR-WT
0
5
10
15
20
25
K/μ
l
0
5
10
15
K/μ
l
0
20
40
60
80
100
Col
ony
num
ber/1
0 µl
RXR-KO
RXR-WT
RXR-KO
RXR-WT
ns
Kit+ Sca-1+ Kit+ Sca-1–0
20
40
60
K/1
0 μl
Fig. 3. RXRA inG-CSF–induced granulopoiesis andHSCmobilization. (A) WBC and (B) NE counts of C57BL6mice treatedwith or without G-CSF and bexarotene as indicated. n = 16 (no treatment control), 20 (G-CSF), and 19 (G-CSF + bexarotene).(C) Absolute number of KLS cells and (D) Kit+Lin−Sca-1− (progenitor) cells and (E) CFU in 10 ml of peripheral blood fromC57BL6 mice treated with or without G-CSF and bexarotene as indicated. n = 11 (no treatment control), 15 (G-CSF), and 15(G-CSF + bexarotene). (F) WBC and (G) NE count of RXR-KO and RXR-WT mice treated with G-CSF. (H) CFU and (I) absolutenumber of KLS cells and Kit+Lin−Sca-1− (progenitors) cells in 10 ml of peripheral blood of RXR-KO and RXR-WT mice. n = 3RXR-KO and 4 RXR-WT mice. Error bars represent SDs between individual mice. *P < 0.05; **P < 0.01; ***P < 0.001, ANOVAwith Tukey’s multiple comparisons test.
4 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
A***
Retinol
VADC VAD0
100
200
300
400
Rat
io to
inte
rnal
sta
ndar
d
B
*
C16:1
NFC NF0.0
0.5
1.0
1.5
C D
* : NFC vs NFC-PHZ * : NF vs NF-PHZ * : NFC vs NF * : NFC-PHZ vs NF-PHZ
Rat
io to
inte
rnal
sta
ndar
d
log(μl serum)
GFP
+ mC
herr
y+ /m
Che
rry+
0.4 0.6 0.8 1.00.00
0.05
0.10
0.15
0.20
0.25
NFC
NFC-PHZ
NF
NF-PHZ
*** *** *** ***** *** ** *** ***
* ***** ** ** *
0.0 0.2 0.4 0.6 0.8 1.00.0
0.1
0.2
0.3
0.4
0.5
log(μl serum)
GFP
+ mC
herr
y+ /m
Che
rry+ VADC-PHZ
VAD-PHZVADCVAD
* : VADC vs VADC-PHZ * : VAD vs VAD-PHZ
*** *** *** **** ** *** ** *
Fig. 5. Effect of vitaminAdeficiency and fatty acid deficiency onplasmaRXRA ligands. (A) Plasma concentration of retinol in VADandVADCmice as determined byMS. n=5 VADC and 5 VADmice. (B) Ratio of GFP+mCherry+ cells to totalmCherry+ cells in UAS-GFP bonemarrowKit+ cells transducedwith Gal4-RXRA retrovirus and treatedwith plasmafromVADor VADCmicewith andwithout PHZ treatment.n=4VAD, 4 VAD+PHZ, 5 VADC, and 5VADC+PHZ. (C) Plasma concentration of the fatty acid C16:1 inmice fed anNForNFCdiet as determinedbyMS.n=5NFand4NFC. (D) Ratio of GFP+mCherry+ cells to totalmCherry+ cells inUAS-GFPbonemarrowKit+ cells transducedwithGal4-RXRA retrovirusand treatedwith plasma fromNF or NFCmicewith andwithout PHZ treatment. n=3NFC, 2 NFC +PHZ, 3 NF, and 4NF+ PHZ. Error bars represent SDs betweenmeasurements ofplasma obtained from separate mice. *P < 0.05; **P < 0.01; ***P < 0.001, ANOVA with Tukey’s multiple comparisons compared results obtained at each plasma concentration.
No drug
Bex (1
00 nM)
9-cis
-RA (1
00nM)
HX531(1
μM)2 μ
l4 μl
6 μl8 μl
1 0μl0.00
0.05
0.10
0.15
0.20G
FP+ m
Che
rry+ /
mC
herr
y+BA
Plasma volumeadded to 200 μl of medium
* : PHZ vs no treatment* : G-CSF vs no treatment
Plasma volumeadded to 200 μl of medium
*****
****** *** *** *** *** *** **
*** *** *** *
log(μl serum)
No treatmentG-CSFPHZ
GFP
+ mC
herr
y+ /m
Che
rry+
0.0 0.2 0.4 0.6 0.8 1.00.00
0.05
0.10
0.15
0.20
0.25
Fig. 4. RXRA reporter activated by mouse plasma. (A) Ratio of GFP+mCherry+ cells to total mCherry+ cells in UAS-GFP bone marrow Kit+ cells transduced with Gal4-RXRAretrovirus and treated in culture with the RXRA agonists bexarotene or 9-cis-RA, the RXRA antagonist HX531, or mouse plasma as indicated. Two-way t test compared againstuntreated control. (B) Ratio of GFP+mCherry+ cells to total mCherry+ cells in UAS-GFP bone marrow Kit+ cells transduced with Gal4-RXRA retrovirus and treated in culture withplasma frommice that had been treatedwith PHZ, G-CSF, or vehicle control. Error bars represent SDs betweenmeasurements of plasmaobtained from individualmice (n=3miceper treatment group). ANOVA with Tukey’s multiple comparisons compared results obtained at each plasma concentration. *P < 0.05; **P < 0.01; ***P < 0.001.
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017 5 of 12
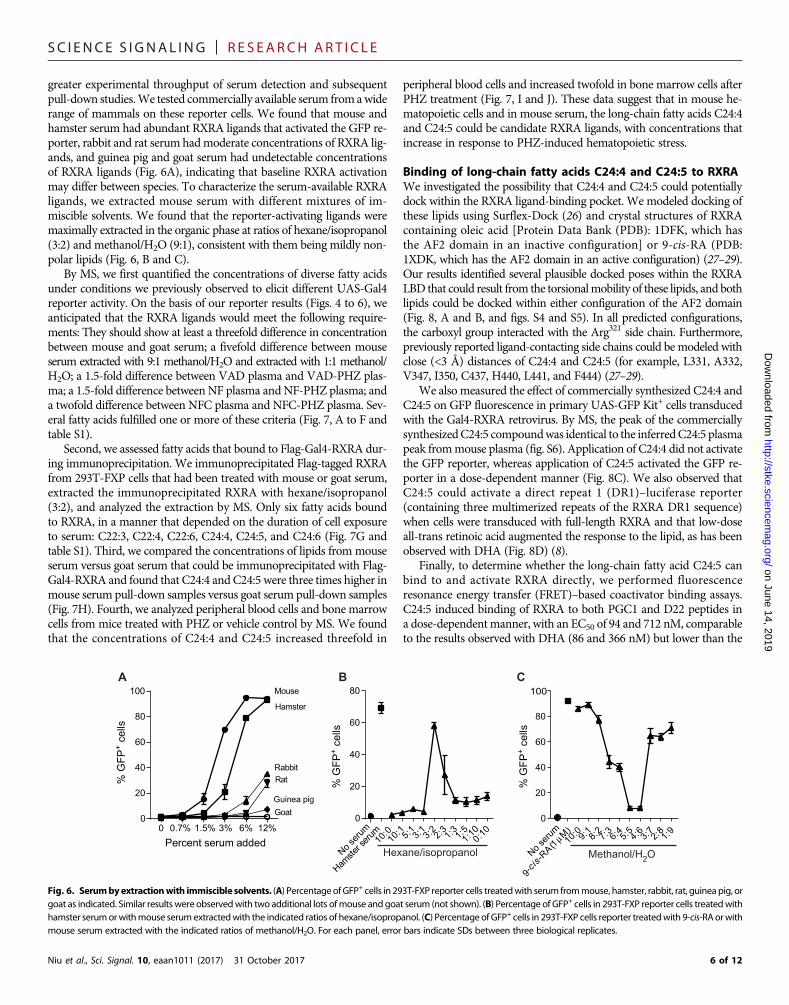
SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
greater experimental throughput of serum detection and subsequentpull-down studies.We tested commercially available serum from awiderange of mammals on these reporter cells. We found that mouse andhamster serum had abundant RXRA ligands that activated the GFP re-porter, rabbit and rat serum hadmoderate concentrations of RXRA lig-ands, and guinea pig and goat serum had undetectable concentrationsof RXRA ligands (Fig. 6A), indicating that baseline RXRA activationmay differ between species. To characterize the serum-available RXRAligands, we extracted mouse serum with different mixtures of im-miscible solvents. We found that the reporter-activating ligands weremaximally extracted in the organic phase at ratios of hexane/isopropanol(3:2) and methanol/H2O (9:1), consistent with them being mildly non-polar lipids (Fig. 6, B and C).
By MS, we first quantified the concentrations of diverse fatty acidsunder conditions we previously observed to elicit different UAS-Gal4reporter activity. On the basis of our reporter results (Figs. 4 to 6), weanticipated that the RXRA ligands would meet the following require-ments: They should show at least a threefold difference in concentrationbetween mouse and goat serum; a fivefold difference between mouseserum extracted with 9:1 methanol/H2O and extracted with 1:1 methanol/H2O; a 1.5-fold difference between VAD plasma and VAD-PHZ plas-ma; a 1.5-fold difference between NF plasma and NF-PHZ plasma; anda twofold difference between NFC plasma and NFC-PHZ plasma. Sev-eral fatty acids fulfilled one or more of these criteria (Fig. 7, A to F andtable S1).
Second, we assessed fatty acids that bound to Flag-Gal4-RXRA dur-ing immunoprecipitation. We immunoprecipitated Flag-tagged RXRAfrom 293T-FXP cells that had been treated with mouse or goat serum,extracted the immunoprecipitated RXRA with hexane/isopropanol(3:2), and analyzed the extraction by MS. Only six fatty acids boundto RXRA, in a manner that depended on the duration of cell exposureto serum: C22:3, C22:4, C22:6, C24:4, C24:5, and C24:6 (Fig. 7G andtable S1). Third, we compared the concentrations of lipids frommouseserum versus goat serum that could be immunoprecipitated with Flag-Gal4-RXRA and found that C24:4 and C24:5 were three times higher inmouse serum pull-down samples versus goat serum pull-down samples(Fig. 7H). Fourth, we analyzed peripheral blood cells and bone marrowcells from mice treated with PHZ or vehicle control by MS. We foundthat the concentrations of C24:4 and C24:5 increased threefold in
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
peripheral blood cells and increased twofold in bone marrow cells afterPHZ treatment (Fig. 7, I and J). These data suggest that in mouse he-matopoietic cells and in mouse serum, the long-chain fatty acids C24:4and C24:5 could be candidate RXRA ligands, with concentrations thatincrease in response to PHZ-induced hematopoietic stress.
Binding of long-chain fatty acids C24:4 and C24:5 to RXRAWe investigated the possibility that C24:4 and C24:5 could potentiallydock within the RXRA ligand-binding pocket. We modeled docking ofthese lipids using Surflex-Dock (26) and crystal structures of RXRAcontaining oleic acid [Protein Data Bank (PDB): 1DFK, which hasthe AF2 domain in an inactive configuration] or 9-cis-RA (PDB:1XDK, which has the AF2 domain in an active configuration) (27–29).Our results identified several plausible docked poses within the RXRALBD that could result from the torsionalmobility of these lipids, and bothlipids could be docked within either configuration of the AF2 domain(Fig. 8, A and B, and figs. S4 and S5). In all predicted configurations,the carboxyl group interacted with the Arg321 side chain. Furthermore,previously reported ligand-contacting side chains could be modeled withclose (<3 Å) distances of C24:4 and C24:5 (for example, L331, A332,V347, I350, C437, H440, L441, and F444) (27–29).
We also measured the effect of commercially synthesized C24:4 andC24:5 on GFP fluorescence in primary UAS-GFP Kit+ cells transducedwith the Gal4-RXRA retrovirus. By MS, the peak of the commerciallysynthesizedC24:5 compoundwas identical to the inferredC24:5 plasmapeak frommouse plasma (fig. S6). Application of C24:4 did not activatethe GFP reporter, whereas application of C24:5 activated the GFP re-porter in a dose-dependent manner (Fig. 8C). We also observed thatC24:5 could activate a direct repeat 1 (DR1)–luciferase reporter(containing three multimerized repeats of the RXRA DR1 sequence)when cells were transduced with full-length RXRA and that low-doseall-trans retinoic acid augmented the response to the lipid, as has beenobserved with DHA (Fig. 8D) (8).
Finally, to determine whether the long-chain fatty acid C24:5 canbind to and activate RXRA directly, we performed fluorescenceresonance energy transfer (FRET)–based coactivator binding assays.C24:5 induced binding of RXRA to both PGC1 and D22 peptides ina dose-dependentmanner, with an EC50 of 94 and 712 nM, comparableto the results observed with DHA (86 and 366 nM) but lower than the
A B
Percent serum added
% G
FP+
cells
0 0.7% 1.5% 3% 6% 12%0
20
40
60
80
100 Mouse
Hamster
RabbitRat
Guinea pigGoat
% G
FP+
cells
10:15:13:13:22:31:31:51:1
00
20
40
60
80
Hamste
r seru
m
C
Methanol/H2ONo seru
m
9-cis
-RA(1μM)
10:09:18:27:36:45:54:63:72:81:9
0
20
40
60
80
100
Hexane/isopropanol10
:00:1
0
No seru
m
% G
FP+
cells
Fig. 6. Serumbyextractionwith immiscible solvents. (A) PercentageofGFP+ cells in 293T-FXP reporter cells treatedwith serum frommouse, hamster, rabbit, rat, guinea pig, orgoat as indicated. Similar results were observedwith two additional lots ofmouse andgoat serum (not shown). (B) Percentage of GFP+ cells in 293T-FXP reporter cells treatedwithhamster serumorwithmouse serumextractedwith the indicated ratios of hexane/isopropanol. (C) Percentage of GFP+ cells in 293T-FXP cells reporter treatedwith 9-cis-RA orwithmouse serum extracted with the indicated ratios of methanol/H2O. For each panel, error bars indicate SDs between three biological replicates.
6 of 12
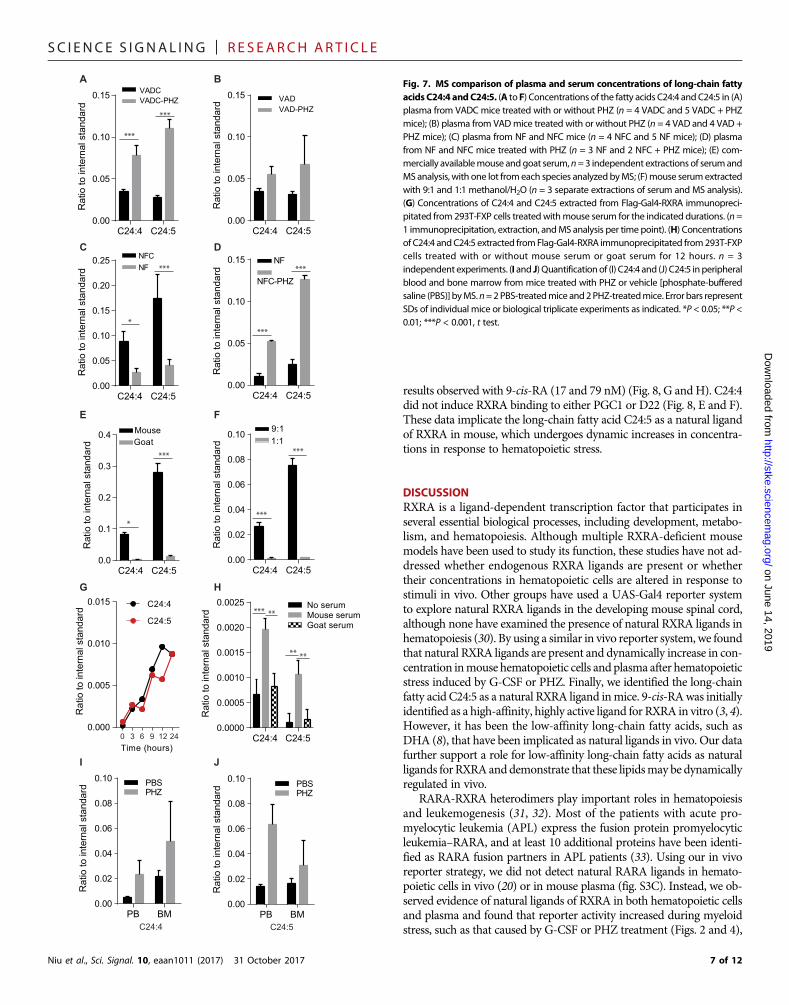
SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
http:D
ownloaded from
results observed with 9-cis-RA (17 and 79 nM) (Fig. 8, G andH). C24:4did not induce RXRA binding to either PGC1 or D22 (Fig. 8, E and F).These data implicate the long-chain fatty acid C24:5 as a natural ligandof RXRA in mouse, which undergoes dynamic increases in concentra-tions in response to hematopoietic stress.
on June 14, 2019//stke.sciencem
ag.org/
DISCUSSIONRXRA is a ligand-dependent transcription factor that participates inseveral essential biological processes, including development, metabo-lism, and hematopoiesis. Although multiple RXRA-deficient mousemodels have been used to study its function, these studies have not ad-dressed whether endogenous RXRA ligands are present or whethertheir concentrations in hematopoietic cells are altered in response tostimuli in vivo. Other groups have used a UAS-Gal4 reporter systemto explore natural RXRA ligands in the developing mouse spinal cord,although none have examined the presence of natural RXRA ligands inhematopoiesis (30). By using a similar in vivo reporter system, we foundthat natural RXRA ligands are present and dynamically increase in con-centration inmouse hematopoietic cells and plasma after hematopoieticstress induced by G-CSF or PHZ. Finally, we identified the long-chainfatty acid C24:5 as a natural RXRA ligand inmice. 9-cis-RAwas initiallyidentified as a high-affinity, highly active ligand for RXRA in vitro (3, 4).However, it has been the low-affinity long-chain fatty acids, such asDHA (8), that have been implicated as natural ligands in vivo. Our datafurther support a role for low-affinity long-chain fatty acids as naturalligands forRXRAanddemonstrate that these lipidsmay be dynamicallyregulated in vivo.
RARA-RXRA heterodimers play important roles in hematopoiesisand leukemogenesis (31, 32). Most of the patients with acute pro-myelocytic leukemia (APL) express the fusion protein promyelocyticleukemia–RARA, and at least 10 additional proteins have been identi-fied as RARA fusion partners in APL patients (33). Using our in vivoreporter strategy, we did not detect natural RARA ligands in hemato-poietic cells in vivo (20) or in mouse plasma (fig. S3C). Instead, we ob-served evidence of natural ligands of RXRA in both hematopoietic cellsand plasma and found that reporter activity increased during myeloidstress, such as that caused by G-CSF or PHZ treatment (Figs. 2 and 4),
B
C24:4 C24:50.00
0.05
0.10
0.15 VADVAD-PHZ
***
***
A
C24:4 C24:50.00
0.05
0.10
0.15R
atio
to in
tern
al s
tand
ard
VADCVADC-PHZ
***
*
C
C24:4 C24:50.00
0.05
0.10
0.15
0.20
0.25 NFCNF ***
***
D
C24:4 C24:50.00
0.05
0.10
0.15 NF
NFC-PHZ
***
*
E
C24:4 C24:50.0
0.1
0.2
0.3
0.4 MouseGoat
C24:5
J
PB BM0.00
0.02
0.04
0.06
0.08
0.10 PBSPHZ
C24:4
I
PB BM0.00
0.02
0.04
0.06
0.08
0.10 PBSPHZ
***
***
F
C24:4 C24:50.00
0.02
0.04
0.06
0.08
0.10 9:11:1
*** **
** **
H
C24:4 C24:50.0000
0.0005
0.0010
0.0015
0.0020
0.0025 No serumMouse serumGoat serum
G
0.000
0.005
0.010
0.015
Time (hours)
C24:4
C24:5
0 3 6 9 12 24
Rat
io to
inte
rnal
sta
ndar
dR
atio
to in
tern
al s
tand
ard
Rat
io to
inte
rnal
sta
ndar
dR
atio
to in
tern
al s
tand
ard
Rat
io to
inte
rnal
sta
ndar
dR
atio
to in
tern
al s
tand
ard
Rat
io to
inte
rnal
sta
ndar
dR
atio
to in
tern
al s
tand
ard
Rat
io to
inte
rnal
sta
ndar
d
Fig. 7. MS comparison of plasma and serum concentrations of long-chain fattyacids C24:4 andC24:5. (A toF) Concentrations of the fatty acids C24:4 andC24:5 in (A)plasma from VADC mice treated with or without PHZ (n = 4 VADC and 5 VADC + PHZmice); (B) plasma from VADmice treated with or without PHZ (n = 4 VAD and 4 VAD +PHZ mice); (C) plasma from NF and NFC mice (n = 4 NFC and 5 NF mice); (D) plasmafrom NF and NFC mice treated with PHZ (n = 3 NF and 2 NFC + PHZ mice); (E) com-mercially availablemouse andgoat serum,n=3 independent extractions of serumandMS analysis, with one lot fromeach species analyzed byMS; (F)mouse serumextractedwith 9:1 and 1:1 methanol/H2O (n = 3 separate extractions of serum and MS analysis).(G) Concentrations of C24:4 and C24:5 extracted from Flag-Gal4-RXRA immunopreci-pitated from 293T-FXP cells treatedwithmouse serum for the indicated durations. (n=1 immunoprecipitation, extraction, andMS analysis per time point). (H) Concentrationsof C24:4 andC24:5 extracted fromFlag-Gal4-RXRA immunoprecipitated from293T-FXPcells treated with or without mouse serum or goat serum for 12 hours. n = 3independent experiments. (I and J) Quantificationof (I) C24:4 and (J) C24:5 in peripheralblood and bone marrow from mice treated with PHZ or vehicle [phosphate-bufferedsaline (PBS)] byMS.n=2PBS-treatedmice and2PHZ-treatedmice. Error bars representSDs of individual mice or biological triplicate experiments as indicated. *P < 0.05; **P <0.01; ***P < 0.001, t test.
7 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
but not after stem cell stress, such as that induced by 5-FU (fig. S3A).We observed no reporter activity when bone marrow cells were trans-duced with a form of RXRA lacking the AF2 domain, which is requiredfor ligand-dependent activation of the reporter, suggesting that theincreased reporter activity after induction of myeloid stress was dueto increased concentrations of natural RXRA ligands. Because this de-letion only prevents ligand-dependent activation of the reporter, it doesnot control for alternative, ligand-independent effects on RXRA thatcould result from G-CSF or PHZ treatment, and this limitation mustbe acknowledged, although increased concentrations of RXRA ligandsDHA and C24:5 were observed in parallel by MS.
RXRs are central members of the nuclear receptor superfamily (1)andcan functionashomodimers (10) orheterodimers (11–15). PPAR-RXRsare permissive heterodimers that can be activated by either PPAR li-gands or RXR ligands (34). In contrast, TR-RXR and VDR-RXR arenonpermissive heterodimers, which only can be activated in the pres-ence of TR ligands or VDR ligands, respectively (35). PPARs participatein self-renewal of hematopoietic stem and progenitor cells (36, 37), TRsinfluence proliferation and differentiation of erythroid progenitors (38),
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
and VDR deficiency promotes survival of hematopoietic stem and pro-genitor cells in the spleen (39). Both RXRs and PPARs can be activatedby long-chain fatty acids. However, we did not observe activation of thereporter when cells were treated with PPAR ligands (8, 22), suggestingthat the UAS-Gal4 systemwas responding specifically to RXRA ligands(fig. S2, B and C).
We found that RXRA reporter activity increased during GSCF-induced granulopoiesis. Through studies with RXR-KO mice, wefound that RXR activation contributed to G-CSF–inducedmobilizationof HSCs (although it was not absolutely required) and that G-CSF–induced mobilization of HSCs was augmented by the addition of phar-macologic concentrations of RXR ligands (Fig. 3, A to E).
The vitamin A derivatives 9-cis-RA and 9-cis-13,14-DHRA havebeen proposed as natural RXRA ligands in mouse serum, brain, andliver (3, 4, 7). However, when we examined plasma in a VAD mousemodel, in which plasma retinol concentrations were undetectable(Fig. 5A), we nevertheless detected activation of the RXRA reporter,consistent with the presence of RXRA ligands (Fig. 5B). These data sug-gest that the predominant plasma RXRA ligands both under baseline
FD22 peptide
log (nM compound)
C24:5
9-cis-RADHA
100 101 102 103 1040.0
0.5
1.0
1.5
2.0
2.5
Emis
sion
ratio
(520
:495
)
C24:4
EPGC1α peptide
log (nM compound)
Emis
sion
ratio
(520
:495
)
100 101 102 103 1040.0
0.2
0.4
0.6
0.8
1.0 9-cis-RA
C24:5DHA
C24:4
C24:5 (n-3) configuration 13 - 1DKF C24:5 (n-3) configuration 3 - 1XDKA B
C
log (nM compound)
GFP
+ mC
herr
y+ /m
Che
rry+
100 101 102 103 104 1050.0
0.1
0.2
0.3
0.4
C24:4C24:5
UAS-GFP × Gal4-RXRAD
DR1x3-TK-Luc × RXRA
μM C24:510 nM ATRA
Ligh
t uni
ts
0–
20–
50–
75–
0+
20+
50+75+
0
5,000
10,000
15,000
20,000
25,000
*
*******
R321A318
L446
L441
F444
H440
L331
F351
V347
R321A318
L446
L441
F444H440
L331
V347
V354
Fig. 8. Transactivation of RXRA by C24:5. (A and B) In silico models of C24:5 docked in RXRA in an active configuration [(A) PDB: 1DKF] or an inactive configuration [(B) PDB:1XDK]. Blue, nitrogen; red, oxygen; green, carbon in the lipid; gray, hydrogen and carbon in the protein. (C) Transactivation of UAS-GFP bone marrow Kit+ cells transduced withGal4-RXRA and treated in culture with commercially synthesized C24:5 or C24:4. Error bars indicate SDs of independent triplicates. (D) Transactivation of multimerized DR1promoter by full-length RXRA and indicated ligands. Error bars indicate SDs of independent triplicates compared by t test. ATRA, all-trans retinoic acid; Luc, luciferase. (E andF) TR-FRET showing RXRA binding to PGC1a (E) and D22 (F) coactivator peptides in the presence of the indicated concentrations of 9-cis-RA, DHA, C24:5, or C24:4. Error barsrepresent SDs of independent triplicate experiments. EC50 determined by fitting the data to a log(agonist) variable slope with four parameters. *P < 0.05; **P < 0.01; ***P < 0.001.
8 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
conditions and after PHZ treatment are unlikely to be vitamin Aderivatives.
In contrast, multiple groups have suggested that fatty acids mightserve as activating RXR ligands. The crystallographic analysis of aconstitutively active mutant form of mouse RXRA (RXRAF318A) re-vealed a continuous U-shaped electron density in the ligand-bindingcavity, suggesting the presence of a bound ligand and consistent witha fatty acid of 16 or 18 carbon atoms (27). C18:1 can activate the RXRAreporter ex vivo butwith low efficiency (8).We detected fatty acidC18:1inmouse serum and found that it wasmore abundant inmouse than ingoat serum, but its concentration was not affected by PHZ treatment(table S1). Moreover, we found that C18:1 could be extracted bymethanol/H2O (1:1), yet the products extracted by methanol/H2O(1:1) did not activate the RXRA reporter (Fig. 6C). The unsaturatedfatty acids C20:4, C22:5, and C22:6 have been identified as Rxr ligandsin mouse brain (8). We detected C22:5 in both mouse and goat serum,although goat serum did not activate the RXRA reporter (Fig. 6A andtable S1). We also found that C22:5 did not coimmunoprecipitate withRXRA when the reporter cells were incubated with serum, althoughC22:6 did (table S1). However, PHZ treatment did not consistentlyaffect C22:6 concentrations in VAD mice and NF mice (table S1),suggesting that C22:6 may contribute to basal RXR activation but doesnot appear to increase in concentration in response to PHZ. In contrast,C24:5 was consistently detected by MS under conditions that lead toactivation of the RXRA reporter and was dynamically increased inplasma concentration after PHZ treatment. Commercially synthesizedC24:5 activated the RXRA reporter, activated a DR1-luciferase reporterwhen coexpressed with full-length RXRA, and induced dose-dependentcoactivator binding to purified RXRA (Figs. 7 and 8), suggesting thatthis lipid is a natural RXRA ligand that increases in plasma concentrationin response to hematopoietic signals.
In summary, we observed evidence of activating natural ligands forRXRA in mouse hematopoietic cells, as well as in plasma and serum.Concentrations of these ligands were increased after G-CSF– orPHZ-inducedmyeloid stress, and we identified C24:5 as a likely naturalRXRA ligand that undergoes dynamic increases in concentrationduring myeloid stress. These data do not address further importantquestions: Which pathways lead to alterations in serum concentrationsof natural RXRA ligands? Do differences in diet augment or diminishRXRA signaling and the subsequent consequences on inflammationand bone metabolism? Does intervention with RXRA agonists improvestem cell mobilization or augment G-CSF–induced NE count recoveryafter chemotherapy? Additional studies will be required to furtherdefine the function of RXRs and the outcome of their in vivo regulation.
MATERIALS AND METHODSReagentsBexarotenewas fromLCLaboratories. PHZ, 5-FU, corn oil, hexane, andisopropanol were from Sigma-Aldrich. G-CSF was from Amgen.GW6471, GW7647, pioglitazone, and tesaglitazar were from Tocris.Anti-mouse CD11b (M1/70)–BV421, anti-mouse c-Kit (2B8)–BV421,anti-mouse B220 (RA3-6B2)–phycoerythrin (PE)–Cy7 were from BDBiosciences. Anti-mouseCD8 (53 to 6.7)–eFluor 450, anti-mouseCD71(R17217)–eFluor 450, anti-mouse Sca-1 (D7)–APC (allophycocyanin),anti-mouseGr-1 (RB6-8C5)–APC, anti-mouseCD4 (GK1.5)–APC, anti-mouse Ter119 (TER119)–APC, anti-mouse c-Kit (2B8)–PE-Cy7, anti-mouse CD19 (eBio1D3)–PE-Cy7, anti-mouse Ter119 (TER119)–PE-Cy7,anti-mouseCD127 (ATR34)–PE-Cy7, anti-mouseCD8 (53-6.7)–PE-Cy7,
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
anti-mouse CD4 (RM4-5)–PE-Cy7, and anti-mouse CD3e (145-2C11)–PE-Cy7 were from eBioscience. Sera from mouse, hamster, rabbit, rat,guinea pig, and goat were obtained fromEquitech-Bio Inc., and sera wereobtained while animals were maintained on a standard diet. C24:4[(9Z,12Z,15Z,18Z,21Z)-tetracosa-9,12,15,18,21-tetraenoic acid] andC24:5 [(9Z,12Z,15Z,18Z,21Z)-tetracosa-9,12,15,18,21-pentaenoic acid]were synthesized fromAvanti Polar Lipids Inc. TheDR1-luciferase repor-ter and pBABE-RXRA plasmids were gifts from V. Arora, WashingtonUniversity.
Hematopoietic cell cultureMouse bone marrow Kit+ cells were isolated using an autoMACS Pro(Miltenyi Biotec) per themanufacturer’s protocol. Kit+ cells were platedin progenitor expansion medium [RPMI 1640, 15% fetal bovine serum(FBS), SCF (50 ng/ml), interleukin-3 (IL-3; 10 ng/ml), Fms-relatedtyrosine kinase 3 (Flt3; 25 ng/ml), thrombopoietin (Tpo; 10 ng/ml),L-glutamine (2 mM), sodium pyruvate (1 mM), Hepes buffer (10 mM),penicillin/streptomycin (100 U/ml), and b-mercaptoethanol (50 mM)]overnight and transduced by spinfection with polybrene (10 mg/ml)and 10 mM Hepes at 2400 rpm, 30°C for 90 min in an Eppendorf5810R centrifuge. Fluorescence was detected on a FACS scan or Galliosinstrument (Beckman Coulter).
MiceUAS-GFP and RXR-KO mice were bred as described (19, 20). RXRdeletion was induced by injecting RXR-KO mice intraperitoneallywith polyinosinic:polycytidylic acid (pI:pC) (300 mg per mouse); fourdoses were given every other day. RXR deletion was confirmed by poly-merase chain reaction 4 weeks after mice were treated with pI:pC. Bex-arotene was administrated by oral gavage, suspended in sterile corn oil,1 mg per mouse per day for 3 days, and mice were sacrificed at day 4.G-CSF was administrated by subcutaneous injection, 125 mg/kg every12 hours for nine injections, and mice were sacrificed 2 hours after thelast injection. All mice were cared for in the experimental animal center ofWashington University School of Medicine. The Washington UniversityAnimal Studies Committee approved all animal experiments.
VAD mouse modelVADmice were generated as described (24). Briefly, friend virus B(FVB) female mice were fed with VAD diet (TD.86143; Teklad) orvitamin A control diet (TD.91280; Teklad) during pregnancy. Theoffspring received the same diet for at least 12 weeks. Plasma con-centrations of retinol were undetectable by MS before experimentalintervention.
Diet-restricted fatty acid mouse modelA diet-restricted fatty acid mouse model was generated as described(25). Briefly, FVB mice were fed with NF diet (TD.03314; Teklad)and NFC diet (TD.130321; Teklad) for 4 weeks before experimental in-tervention. Plasma concentrations of palmitoleic acid (C16:1)were eval-uated by MS before experimental intervention.
Retrovirus productionRetrovirus productionwasperformedas described (40).Calciumchloridetransfection and low-passage 293T cells were used for virus packaging.293T cells (5 × 106) were seeded in 10-cm dishes in Dulbecco’s modifiedEagle’s medium (high glucose) + 10% FBS 18 to 24 hours before trans-fection andwere grown to 80% confluence. Twelve-microgramMSCV–Gal4 DBD–RXRA LBD–IRES–mCherry (Gal4-RXRA), MSCV–Gal4
9 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
DBD–RXRA LBD DAF2–IRES–mCherry (Gal4-RXRA DAF2), orMSCV–Gal4 DBD–RARA LBD–IRES–mCherry (Gal4-RARA), 8 mgof Ecopak, and 155 ml of 2 M CaCl2 were mixed, and the volume wasadjusted to 1.25 ml by adding H2O. Hepes buffer (2×; 1.25 ml) wasdrop-wise added to the mixture. The mixture was incubated for 20 minat room temperature and then drop-wise added onto 293T cells. Freshmediumwas changed after 12 hours transfection. Virus was collected at48 and 72 hours and stored at −80°C.
Bone marrow transplantationFemurs, tibias, and pelvises were isolated from 6- to 8-week-old UAS-GFPmice. Bonemarrow cells were collected by centrifuging bones at 6000 rpmfor 2 min. Red blood cells were lysed in ACK Buffer (NH4Cl, 150 mM;KHCO3, 10 mM; Na2EDTA, 0.1 mM) on ice for 5 min. Kit+ cells wereisolated by MACS using autoMACS Pro (Miltenyi Biotec) per the manu-facturer’s protocol. Kit+ cells were cultured in transplant medium [RPMI1640 + 15% FBS + mSCF (20 ng/ml) + mFlt3L (25 ng/ml) + mIL3(10 ng/ml) + Tpo (10 ng/ml) + 50 mM b-mercaptoethanol] overnightand transduced with Gal4-RXRA or Gal4-RXRA-DAF2 retrovirus byspinfection with polybrene (10 mg/ml) and 10 mM Hepes at 2400 rpm,30°C for 90min in an Eppendorf 5810R centrifuge. Kit+ cells were spin-fected with the same virus for a second time after 24 hours. Transducedcells were injected to lethally irradiated recipient mice 2 hours after thesecond spinfection. After 6 weeks of engraftment, mice were sacrificedand analyzed.
Flow cytometryBone marrow, peripheral blood, spleen, and peritoneal macrophageswere collected from the bone marrow of bone marrow transplant reci-pient mice after engraftment (typically ~6 weeks). For hematopoieticstem and progenitor cell analysis, cells were stained with Lineage-PE-Cy7, c-Kit–BV421, and Sca-1–APC. For myeloid lineage analysis,cells were stainedwith c-Kit–PE-Cy7, CD11b-BV421, andGr-1–APC.For erythroid lineage analysis, cells were stained with c-Kit–PE-Cy7,CD71–eFluor 450, and Ter119-APC. For lymphoid lineage analysis,cells were stained with B220-PE-Cy7, CD4-APC, and CD8–eFluor450. Fluorescence was detected using a Gallios flow cytometer (BeckmanCoulter).
Plasma or serum extractionFifty microliters of mouse plasma or serum was mixed with 500 ml ofhexane/isopropanol ormethanol/H2O and vortexed vigorously for 30 s.After centrifugation at 12,000 rpm for 5 min, the upper phase (or-ganic phase) was collected into a new tube and stored at −80°C forlater analysis.
Pull-down assaysWe generated a line of 293T cells stably expressing UAS-GFP and 3×Flag–Gal4 DBD–RXRA LBD–IRES–Puro (293T-FXP). 293T-FXP (1 ×106 cells per well) were seeded in six-well plates and treated with mouseserum or goat serum. Cells were collected and lysed with radioimmu-noprecipitation assay buffer (50 mM tris-HCl, 150 mM NaCl, 1% NP-40, 1%deoxycholate, 0.1% SDS, 1mMEDTA, and proteinase inhibitor)at 0, 3, 6, 9, 12, and 24 hours. 293T-FXP cell lysate was incubated withprewashed anti-Flag magnetic beads and rotated overnight at 4°C.Beads were washed with tris-buffered saline and Tween 20 solution(TBST) three times. Washed beads were resuspended in 50 ml of TBSTand then extracted with hexane/isopropanol (3:2). The organic phasewas collected and stored in −80°C for later analysis.
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
Mass spectrometryFor retinol analysis, 100 ml of plasma extraction samplewas used. BeforeMS analysis, 50 ng of retinol-d6 as the internal standard was added toeach sample. The solvents in the sample were dried under a stream ofnitrogen. The dried samplewas redissolved in 1ml of ethanol/H2O (1:1)for MS analyses.
For fatty acids analysis, 100 ml of plasma extraction sample or 400 mlof pull-down sample was used for DMAPA (dimethylaminopropyla-mine) derivatization for improving MS sensitivity of fatty acids. Beforethe derivatization, 50 ng (10 ng for the pull-down sample) of AA-d8 asthe internal standard was added to each sample. The solvents in thesample were dried under a stream of nitrogen. To derivatize the sample,50 ml of 100mMEDC [N-(3-dimethylaminopropyl)-N-ethylcarbodiimidehydrochloride] and 50 ml of 50 mM DMAPA and 50 mM DMAP(4-dimethylaminopyridine) were added to the dried fatty acid samplesand heated at 50°C for 30 min. The samples were dried under nitrogenand then dissolved in 1 ml of ethanol/H2O (1:1) for MS analysis.
In silico ligand docking analysisDocking of C24:5 and C24:4 against crystal structures of RXRA (PDB:1DKF and 1XDK) was performed with Surflex-Dock (26). MOL2 filesfor each compound were generated from SMILES (simplifiedmolecular-input line-entry system) strings using Open Babel (41). Surflex-Dockreceptor protomols were generated with a threshold of 0.25 and a bloatof 2.0 and subsequently docked using the default “-pgeom” docking ac-curacy parameter set. Residues lining the binding pocket were definedas those within 4 Å of oleic acid (PDB: 1DKF) or 9-cis-RA (PDB:1XDK). Images were generated in Pymol (version 1.8.6.0; Schrodinger).
Luciferase detection293T cells were transfected with pBABE-RXRA in combination withDR1x3-TK-luciferase using Lipofectamine 2000 (Invitrogen). Six hoursafter transfection, the cells were collected and plated into a 48-well platein 1% bovine serum albumin (BSA) medium in triplicate. DHA, fattyacid 24:5, bexarotene, or 9-cis-RA were added to the cells as a BSAcomplex (BSA, Fraction V, fatty acid–free, Calbiochem), with equalamounts of BSA added to all samples (42). After a 40-hour incubation,the cells were harvested and assayed for luciferase (luciferase assay sys-tem with reporter lysis buffer; Promega) in a Beckman Coulter LD400plate reader.
Time resolution–FRET assayThe assay was performed using LanthaScreen TR-FRET Retinoic XReceptor alpha Coactivator assay (Invitrogen). Reaction mixturecontained 10 nM RXRA LBD–GST (glutathione S-transferase), 5 nMterbium-labeled anti-GST antibody, 500 nM fluorescein-PGC1a orD22 peptide, 5 mM dithiothreitol, 1% dimethyl sulfoxide, and testingcompounds. Twenty microliters of the reaction mixture was analyzedafter 1-hour incubation at room temperature and analyzed using a Syn-ergy2 plate reader (BioTek Instruments), using a 340-nm filter with a30-nm bandwidth, and an emission filter was centered at 520 nmwith a25-nm bandwidth and at 495 nm with a 10-nm bandwidth. The TR-FRET ratio was calculated by dividing the emission signal at 520 bythe emission signal at 495.
Data analysisFlow cytometry data were analyzed with FlowJo software version 10.Statistical analysis was performed using Prism (GraphPad). ANOVAand t tests were performed, as appropriate. FRET studies were analyzed
10 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
using a sigmoidal dose response with variable slope. All studies wereperformed in triplicate, unless otherwise indicated. Error bars representSDs. Data points without error bars have SDs belowGraphPad’s limit todisplay.
SUPPLEMENTARY MATERIALSwww.sciencesignaling.org/cgi/content/full/10/503/eaan1011/DC1Fig. S1. Ex vivo validation of the UAS-GFP reporter transgene.Fig. S2. Reporter analysis.Fig. S3. Analysis of RXRA-DAF2 and RARA to natural ligands in mouse sera.Fig. S4. Sixteen different docking configurations of C24:5 in RXRA (1XDK).Fig. S5. Sixteen different docking configurations of C24:4 in RXRA (1XDK).Fig. S6. Comparison of liquid chromatography–MS/MS peaks of C24:5.Table S1. Analysis of long-chain fatty acids in serum samples by MS.
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from
REFERENCES AND NOTES1. R. M. Evans, D. J. Mangelsdorf, Nuclear receptors, RXR, and the Big Bang. Cell 157,
255–266 (2014).2. M. I. Dawson, Z. Xia, The retinoid X receptors and their ligands. Biochim. Biophys. Acta
1821, 21–56 (2012).3. G. Allenby, M.-T. Bocquel, M. Saunders, S. Kazmer, J. Speck, M. Rosenberger, A. Lovey,
P. Kastner, J. F. Grippo, P. Chambon, A. A. Levin, Retinoic acid receptors andretinoid X receptors: Interactions with endogenous retinoic acids. Proc. Natl. Acad. Sci.U.S.A. 90, 30–34 (1993).
4. R. A. Heyman, D. J. Mangelsdorf, J. A. Dyck, R. B. Stein, G. Eichele, R. M. Evans, C. Thaller,9-cis Retinoic acid is a high affinity ligand for the retinoid X receptor. Cell 68,397–406 (1992).
5. S. L. Arnold, J. K. Amory, T. J. Walsh, N. Isoherranen, A sensitive and specific method formeasurement of multiple retinoids in human serum with UHPLC-MS/MS. J. Lipid Res.53, 587–598 (2012).
6. J. W. Jones, K. Pierzchalski, J. Yu, M. A. Kane, Use of fast HPLC multiple reactionmonitoring cubed for endogenous retinoic acid quantification in complex matrices.Anal. Chem. 87, 3222–3230 (2015).
7. R. Rühl, A. Krzyżosiak, A. Niewiadomska-Cimicka, N. Rochel, L. Szeles, B. Vaz,M. Wietrzych-Schindler, S. Álvarez, M. Szklenar, L. Nagy, A. R. de Lera, W. Krężel,9-cis-13,14-Dihydroretinoic acid is an endogenous retinoid acting as RXR ligand in mice.PLOS Genet. 11, e1005213 (2015).
8. J. Lengqvist, A. Mata de Urquiza, A.-C. Bergman, T. M. Willson, J. Sjövall, T. Perlmann,W. J. Griffiths, Polyunsaturated fatty acids including docosahexaenoic and arachidonicacid bind to the retinoid X receptor a ligand-binding domain. Mol. Cell. Proteomics3, 692–703 (2004).
9. A. Nohara, J. Kobayashi, H. Mabuchi, Retinoid X receptor heterodimer variants andcardiovascular risk factors. J. Atheroscler. Thromb. 16, 303–318 (2009).
10. X.-K. Zhang, J. Lehmann, B. Hoffmann, M. I. Dawson, J. Cameron, G. Graupner, T. Hermann,P. Tran, M. Pfahl, Homodimer formation of retinoid X receptor induced by 9-cis retinoicacid. Nature 358, 587–591 (1992).
11. T. H. Bugge, J. Pohl, O. Lonnoy, H. G. Stunnenberg, RXR a, a promiscuous partner ofretinoic acid and thyroid hormone receptors. EMBO J. 11, 1409–1418 (1992).
12. S. A. Kliewer, K. Umesono, D. J. Mangelsdorf, R. M. Evans, Retinoid X receptor interactswith nuclear receptors in retinoic acid, thyroid hormone and vitamin D3 signalling.Nature 355, 446–449 (1992).
13. S. A. Kliewer, K. Umesono, D. J. Noonan, R. A. Heyman, R. M. Evans, Convergence of 9-cisretinoic acid and peroxisome proliferator signalling pathways through heterodimerformation of their receptors. Nature 358, 771–774 (1992).
14. A. C. Calkin, P. Tontonoz, Transcriptional integration of metabolism by the nuclear sterol-activated receptors LXR and FXR. Nat. Rev. Mol. Cell Biol. 13, 213–224 (2012).
15. T. M. Willson, S. A. Kliewer, PXR, CAR and drug metabolism. Nat. Rev. Drug Discov. 1,259–266 (2002).
16. H. Defacque, T. Commes, E. Legouffe, C. Sevilla, J. F. Rossi, C. Rochette-Egly, J. Marti,Expression of retinoid X receptor a is increased upon monocytic cell differentiation.Biochem. Biophys. Res. Commun. 220, 315–322 (1996).
17. S. Taschner, C. Koesters, B. Platzer, A. Jörgl, W. Ellmeier, T. Benesch, H. Strobl, Down-regulation of RXRa expression is essential for neutrophil development from granulocyte/monocyte progenitors. Blood 109, 971–979 (2007).
18. T. Makita, G. Hernandez-Hoyos, T. H.-P. Chen, H. Wu, E. V. Rothenberg, H. M. Sucov, Adevelopmental transition in definitive erythropoiesis: Erythropoietin expression issequentially regulated by retinoic acid receptors and HNF4. Genes Dev. 15, 889–901(2001).
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
19. M. P. Menéndez-Gutiérrez, T. Rőszer, L. Fuentes, V. Núñez, A. Escolano, J. M. Redondo,N. De Clerck, D. Metzger, A. F. Valledor, M. Ricote, Retinoid X receptors orchestrateosteoclast differentiation and postnatal bone remodeling. J. Clin. Invest. 125, 809–823(2015).
20. H. Niu, J. Chacko, G. Hadwiger, J. S. Welch, Absence of natural intracellular retinoidsin mouse bone marrow cells and implications for PML-RARA transformation.Blood Cancer J. 5, e284 (2015).
21. B. Mascrez, M. Mark, A. Dierich, N. B. Ghyselinck, P. Kastner, P. Chambon, The RXRa ligand-dependent activation function 2 (AF-2) is important for mouse development.Development 125, 4691–4707 (1998).
22. J. T. Huang, J. S. Welch, M. Ricote, C. J. Binder, T. M. Willson, C. Kelly, J. L. Witztum,C. D. Funk, D. Conrad, C. K. Glass, Interleukin-4-dependent production of PPAR-g ligandsin macrophages by 12/15-lipoxygenase. Nature 400, 378–382 (1999).
23. B. S. Dornfest, D. M. Lapin, B. A. Naughton, S. Adu, L. Korn, A. S. Gordon, Phenylhydrazine-induced leukocytosis in the rat. J. Leukoc. Biol. 39, 37–48 (1986).
24. K. Ikami, M. Tokue, R. Sugimoto, C. Noda, S. Kobayashi, K. Hara, S. Yoshida, Hierarchicaldifferentiation competence in response to retinoic acid ensures stem cell maintenanceduring mouse spermatogenesis. Development 142, 1582–1592 (2015).
25. M. V. Chakravarthy, Z. Pan, Y. Zhu, K. Tordjman, J. G. Schneider, T. Coleman, J. Turk,C. F. Semenkovich, “New” hepatic fat activates PPARa to maintain glucose, lipid, andcholesterol homeostasis. Cell Metab. 1, 309–322 (2005).
26. A. N. Jain, Surflex-Dock 2.1: Robust performance from ligand energetic modeling,ring flexibility, and knowledge-based search. J. Comput. Aided Mol. Des. 21, 281–306(2007).
27. W. Bourguet, V. Vivat, J.-M. Wurtz, P. Chambon, H. Gronemeyer, D. Moras, Crystal structureof a heterodimeric complex of RAR and RXR ligand-binding domains. Mol. Cell 5,289–298 (2000).
28. V. Pogenberg, J.-F. Guichou, V. Vivat-Hannah, S. Kammerer, E. Pérez, P. Germain,A. R. de Lera, H. Gronemeyer, C. A. Royer, W. Bourguet, Characterization of the interactionbetween retinoic acid receptor/retinoid X receptor (RAR/RXR) heterodimers andtranscriptional coactivators through structural and fluorescence anisotropy studies.J. Biol. Chem. 280, 1625–1633 (2005).
29. P. F. Egea, A. Mitschler, D. Moras, Molecular recognition of agonist ligands by RXRs.Mol. Endocrinol. 16, 987–997 (2002).
30. L. Solomin, C. B. Johansson, R. H. Zetterström, R. P. Bissonnette, R. A. Heyman, L. Olson,U. Lendahl, J. Frisén, T. Perlmann, Retinoid-X receptor signalling in the developingspinal cord. Nature 395, 398–402 (1998).
31. B. S. Johnson, R. A. Chandraratna, R. A. Heyman, E. A. Allegretto, L. Mueller, S. J. Collins,Retinoid X receptor (RXR) agonist-induced activation of dominant-negative RXR-retinoicacid receptor a403 heterodimers is developmentally regulated during myeloiddifferentiation. Mol. Cell. Biol. 19, 3372–3382 (1999).
32. Y. A. Wang, K. Shen, Y. Ishida, Y. Wang, A. Kakizuka, S. C. Brooks, Induction of murineleukemia and lymphoma by dominant negative retinoic acid receptor a. Mol. Carcinog.44, 252–261 (2005).
33. E. De Braekeleer, N. Douet-Guilbert, M. De Braekeleer, RARA fusion genes in acutepromyelocytic leukemia: A review. Expert Rev. Hematol. 7, 347–357 (2014).
34. B. M. Forman, K. Umesono, J. Chen, R. M. Evans, Unique response pathways areestablished by allosteric interactions among nuclear hormone receptors. Cell 81, 541–550(1995).
35. A. I. Shulman, D. J. Mangelsdorf, Retinoid x receptor heterodimers in the metabolicsyndrome. N. Engl. J. Med. 353, 604–615 (2005).
36. K. Ito, R. Turcotte, J. Cui, S. E. Zimmerman, S. Pinho, T. Mizoguchi, F. Arai, J. M. Runnels,C. Alt, J. Teruya-Feldstein, J. C. Mar, R. Singh, T. Suda, C. P. Lin, P. S. Frenette, K. Ito,Self-renewal of a purified Tie2+ hematopoietic stem cell population relies onmitochondrial clearance. Science 354, 1156–1160 (2016).
37. H.-Y. Lee, X. Gao, M. I. Barrasa, H. Li, R. R. Elmes, L. L. Peters, H. F. Lodish, PPAR-a andglucocorticoid receptor synergize to promote erythroid progenitor self-renewal. Nature522, 474–477 (2015).
38. A. Bauer, W. Mikulits, G. Lagger, G. Stengl, G. Brosch, H. Beug, The thyroidhormone receptor functions as a ligand-operated developmental switch betweenproliferation and differentiation of erythroid progenitors. EMBO J. 17, 4291–4303(1998).
39. N. T. Jeanson, D. T. Scadden, Vitamin D receptor deletion leads to increasedhematopoietic stem and progenitor cells residing in the spleen. Blood 116, 4126–4129(2010).
40. H. Niu, G. Hadwiger, H. Fujiwara, J. S. Welch, Pathways of retinoid synthesis in mousemacrophages and bone marrow cells. J. Leukoc. Biol. 99, 797–810 (2016).
41. N. M. O’Boyle, M. Banck, C. A. James, C. Morley, T. Vandermeersch, G. R. Hutchison, OpenBabel: An open chemical toolbox. J. Cheminform. 3, 33 (2011).
42. P. K.-Y. Chang, A. Khatchadourian, R. A. McKinney, D. Maysinger, Docosahexaenoic acid(DHA): A modulator of microglia activity and dendritic spine morphology.J. Neuroinflammation 12, 34 (2015).
11 of 12

SC I ENCE S I GNAL ING | R E S EARCH ART I C L E
Acknowledgments: We thank the Alvin J. Siteman Cancer Center at Washington UniversitySchool of Medicine and Barnes-Jewish Hospital in St. Louis, MO for the use of the FlowCytometry Core. The Siteman Cancer Center is supported in part by a National Cancer InstituteCancer Center Support Grant P30 CA91842. We thank the High-Throughput ScreeningCenter at Washington University School of Medicine in St. Louis, MO, and the MS facility atWashington University. We thank D. Laflamme for technical assistance and F. Gao for statisticalassistance. Funding: This work was supported by NIH R01 HL128447 (J.S.W.), NIH P50 CA171963(project 1; J.S.W.), and grants from the Spanish Ministry of Economy and Competitiveness(SAF2015-64287R and SAF2015-71878-REDT) (M.R.). The MS facility at Washington University issupported by NIH P30 DK020579 (D. Ory). Author contributions: J.S.W., H.N., and M.R.designed the experiments, performed the experiments, and wrote the manuscript. H.F., O.d.M.,G.H., M.P.M.-G., T.E.F., and G.R.B. designed and performed the experiments. Competing
Niu et al., Sci. Signal. 10, eaan1011 (2017) 31 October 2017
interests: The authors declare that they have no competing interests. Data and materialsavailability: MS data are available at the University of California, San Diego MetabolomicsWorkbench (www.metabolomicsworkbench.org; accession: hhfuji11_20170922_072445.
Submitted 6 March 2017Accepted 20 September 2017Published 31 October 201710.1126/scisignal.aan1011
Citation: H. Niu, H. Fujiwara, O. di Martino, G. Hadwiger, T. E. Frederick, M. P. Menéndez-Gutiérrez, M. Ricote, G. R. Bowman, J. S. Welch, Endogenous retinoid X receptor ligands inmouse hematopoietic cells. Sci. Signal. 10, eaan1011 (2017).
12 of 12
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from

Endogenous retinoid X receptor ligands in mouse hematopoietic cells
Ricote, Gregory R. Bowman and John S. WelchHaixia Niu, Hideji Fujiwara, Orsola di Martino, Gayla Hadwiger, Thomas E. Frederick, María P. Menéndez-Gutiérrez, Mercedes
DOI: 10.1126/scisignal.aan1011 (503), eaan1011.10Sci. Signal.
ligands for RXRA in hematopoietic cells.likely endogenous ligand for RXRA in this context. These findings establish fatty acids as dynamically controlled naturalin fatty acids. Mass spectrometry and other biochemical methods identified the long-chain fatty acid C24:5 as the most these mice also stimulated RXRA activation, even if the mice were deficient in vitamin A but not if the mice were deficientincreased in hematopoietic cells when mice were subjected to treatments that stimulated myeloid cells. Plasma from
. found that RXRA activityet alligands for RXRA. Using a reporter system for detecting RXRA activation in vivo, Niu derived retinoic acids and fatty acids have been implicated as endogenous−ligand-dependent manner. Both vitamin A
(RXRA) stimulates the transcription of target genes in a αLike other nuclear receptors, retinoid X receptor Endogenous RXRA ligands in hematopoietic cells
ARTICLE TOOLS http://stke.sciencemag.org/content/10/503/eaan1011
MATERIALSSUPPLEMENTARY http://stke.sciencemag.org/content/suppl/2017/10/27/10.503.eaan1011.DC1
CONTENTRELATED
http://immunology.sciencemag.org/content/immunology/1/3/eaaf8864.fullhttp://stke.sciencemag.org/content/sigtrans/10/508/eaar6003.fullhttp://stke.sciencemag.org/content/sigtrans/9/430/ra55.fullhttp://stke.sciencemag.org/content/sigtrans/10/476/eaah6275.fullhttp://stke.sciencemag.org/content/sigtrans/10/496/eaal0180.full
REFERENCES
http://stke.sciencemag.org/content/10/503/eaan1011#BIBLThis article cites 42 articles, 12 of which you can access for free
PERMISSIONS http://www.sciencemag.org/help/reprints-and-permissions
Terms of ServiceUse of this article is subject to the
registered trademark of AAAS.is aScience Signaling Association for the Advancement of Science. No claim to original U.S. Government Works. The title
York Avenue NW, Washington, DC 20005. 2017 © The Authors, some rights reserved; exclusive licensee American (ISSN 1937-9145) is published by the American Association for the Advancement of Science, 1200 NewScience Signaling
on June 14, 2019http://stke.sciencem
ag.org/D
ownloaded from