endocitosis - iib.unsam.edu.ar · fagocitosis la fagocitosis es un tipo de endocitosis en donde se...
TRANSCRIPT

ENDOCITOSIS

Las células eucarióticas poseen un mecanismo mediante el cualpueden incorporar moléculas del exterior llamado ENDOCITOSIS,y otro mecanismo para secretar moléculas al exterior llamadoEXOCITOSIS.Ambos mecanismos están compuestos por un sistema complejo de trafico de vesículasque involucra la membrana celular y las membranas del ER (retículo endoplásmico) y elaparato de Golgi, entre muchas otras.
Verde: vía biosinteticaRojo: vía secretoraAzul: vía de recuperación


Gaitan, Cell 2003
El proceso de endocitosis involucra la internalización de parches de membrana plasmática por distintos mecanismos
El destino del material endocitado depende, en parte, del mecanismo de endocitosis involucrado.

Las células internalizan fragmentos de su membrana plasmática en forma depequeñas vesículas y en un proceso continuo.
Existen dos formas principales de endocitosis:
Pinocitosis: internalización de pequeñasmacromoléculas y fluidos extracelulares. (<150 nm de
diámetro)
Fagocitosis: ingestión de partículas grandescomo microorganismos o debris celular. (> 250nm de
diámetro)

Todas las células eucarióticas están continuamente
ingiriendo fluidos y moléculas por pinocitosis, solocélulas especializadas (fagocíticas: macrófagos,neutrófilos) ingieren partículas grandes.
La tasa de internalización de membrana plasmáticadifiere entre células; los macrófagos y fibroblastosinternalizan el equivalente al 100% de su membranaplasmática cada 30 y 120 min, respectivamente.

El área de la membrana plasmática y el volumen de lascélulas permanece constante durante el proceso activo de
endocitosis, lo cual implica que es balanceado por un procesoinverso denominado exocitosis (es un proceso mediante elcual las vesículas internas de la célula se fusionan con la
membrana plasmática para secretar fluidos o material)
membranaENDOCITOSIS EXOCITOSIS membrana

PINOCITOSIS
La mayoría de las células eucarióticas incorporancontinuamente fluidos y solutos por medio de pinocitosis.
Gran cantidad de la membrana plasmática de la célula esincorporada dentro de ella por endocitosis y retorna a lasuperficie celular por exocitosis.
El tipo de pinocitosis más estudiada es la denominadaendocitosis mediada por receptor, en donde se internalizanmoléculas específicas selectivamente. Estas moléculas seencuentran en el espacio extracelular en concentracionesbajas, como pueden ser hormonas, factores de crecimiento,anticuerpos, hierro, enzimas, vitaminas y colesterol.

Las moléculas específicas que son internalizadas se pegan aproteínas denominadas receptores, que están localizadas enel exterior de la superficie celular y poseen un sitio específicode pegado a la molécula.
Una vez que la molécula se pega a su receptor, estos semueven en la membrana plasmática y se concentran enpequeñas depresiones llamadas clathrin-coated pits.
Estas depresiones son invaginaciones de la membrana celulary se encuentran cubiertos por una proteína denominadaclatrina. Estas invaginaciones son utilizadas para la víaendocítica, en donde macromoléculas específicas sonincorporadas dentro de la célula.

LAS INVAGINACIONES DE CLATRINAOCUPAN EL 2% DE LA MEMBRANA
PLASMATICA.
LAS VESICULAS (RECUBIERTAS DECLATRINA) SE FORMAN EN 1 MINUTO Y EN
SEGUNDOS SE FUSIONAN CON LOSENDOSOMAS TEMPRANOS.
EN FIBROBLASTOS SE FORMAN 2500VESICULAS POR MINUTO

Cuando se desprenden las vesículas de la membranaplasmática y se liberan al espacio intracelular, se remueve lacubierta de clatrina.
Las vesículas que aun transportan los receptores y lasmoléculas unidas específicamente se fusionan con lamembrana de otra organela denominada endosoma.
La función de los endosomas es empaquetar a las moléculasen nuevas vesículas y “enviarlas” a diferentes lugares dentrode la célula.
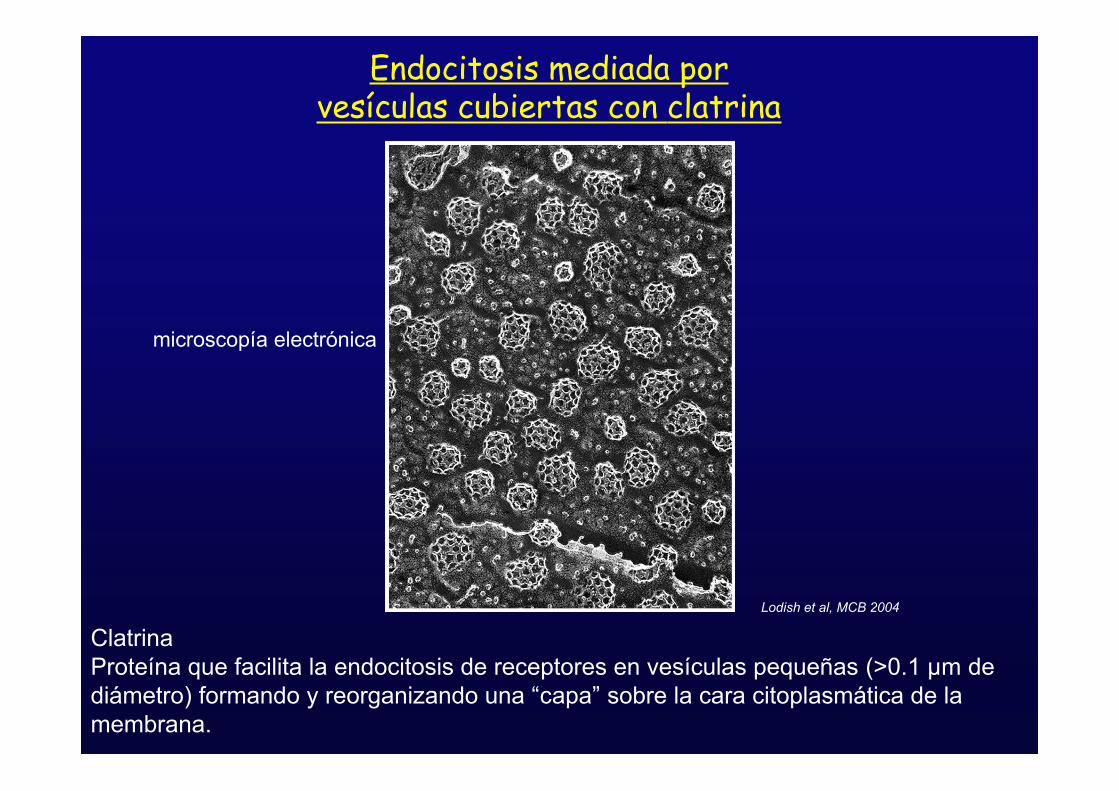
Endocitosis mediada porvesículas cubiertas con clatrina
Lodish et al, MCB 2004
microscopía electrónica
ClatrinaProteína que facilita la endocitosis de receptores en vesículas pequeñas (>0.1 µm dediámetro) formando y reorganizando una “capa” sobre la cara citoplasmática de lamembrana.

Formación de vesículas cubiertas con clatrina en la membrana celular

Secuencia de eventos moleculares durante la endocitosis mediada por vesículas de clatrina
Proteínas adaptadoras se asocian a la membrana, reclutan proteínas cargo y clatrina.Dinamina media la fisión de la vesícula con hidrólisis de GTP. Proteínas accesoriasdesensamblan la cubierta en un proceso que depende de la hidrólisis de ATP.
ATP hidrólisis
Gundelfinger, NRMCB 2003
GTP
hydrolysis

Proteínas adaptadoras reconocen señales de endocitosis en el tallo citosólico de proteínas de transmembrana
AP2 ! NPXY ej. integrinas, receptor de LDLAP2 ! YXXØ ej: receptores de transferrina, M6PAP2 ! LL ej: caderinas, receptor de FcEps ! ubiquitina ej: EGFR, E-caderinas
AP2X representa cualquier aminoácidoØ representa un residuo hidrofóbico
señales reconocidas por proteínas adaptadoras citosólicas
N: Asn Y: TyrP: Pro L: Leu

La distinción de endosomas tempranos y
tardíos se basa en el tiempo post
internalización, localización subcelular,
pH, y en la presencia de proteínas
marcadoras.
Las moléculas internalizadas pueden reciclar tantoa partir de endosomas tempranos como tardíos.Los cuerpos multivesiculares se forman por lainvaginación de la membrana de los endosomastardíos.
Distintos compartimientoso "estaciones" del flujovesicular endocítico
pH6
pH5
pH4.5
MT
El transporte endocítico depende demicrotúbulos (MT)
Sorkin & Zastrow, NRMCB 2002

Destino de las proteínasDestino de las proteínasendocitadasendocitadas
" reciclado de receptores" reciclado de receptores y sus ligandos" desensibilización y reciclado de receptores" degradación en lisosomas
La diferencia en los receptores incorporados en la célula medianteel proceso de endocitosis mediada por receptor es que una vezque liberan las moléculas que transportaron y que abandonan elendosoma pueden dirigirse a diferentes destinos. No como ocurredurante la fagocitosis en donde todo el material que entra a lacélula termina en los lisosomas para su degradación

!Reciclado de receptores
Un ejemplo de endocitosis mediada por receptor es laincorporación de colesterol en células de mamíferos.
Este descubrimiento fue realizado por Michael Brown yJoseph Goldstein, quienes estudiaban la internalizacióndel colesterol en células del torrente sanguíneo. Dichoestudio fue galardonado con el Premio Nobel enMedicina en el año 1985.

Cuando las células necesitan colesterol para la síntesis demembrana, sintetizan el receptor de LDL que es una proteína detransmembrana. Estos receptores se localizan en lasinvaginaciones (“pits”) de clatrina y al unir LDL comienza laendocitosis.
El colesterol sanguíneo es transportado encomplejos lipoproteicos = partículas de LDL(low density lipoprotein)
Una fracción de la membrana endocitada retorna a la superficie (reciclado). Elreceptor de LDL es reciclado mas de 100 veces antes de ser degradado. Cada ciclodura ~10-20 min.

Reciclado de receptores
Las partículas de colesterol LDL se unen areceptores de LDL en la superficie de lascélulas y son endocitadas en vesículas declatrina. El receptor LDL posee la señal deendocitosis NPXY la cual es reconocida porpartículas AP2 y clatrina.Los receptores endocitados se disocian delas partículas LDL en el medio ácido de losendosomas tardíos y son reciclados a lasuperficie.
N: Asn Y: TyrP: Pro

Las partículas de LDL son transportadas al lisosoma endonde enzimas digestivas las degradan en aminoácidos,colesterol y ácidos grasos. Luego el colesterol es liberado alcitoplasma donde es utilizado para la síntesis de membrananueva.
Cuando se acumula colesterol en la célula se apaga lasíntesis del receptor LDL dentro de la célula.
Aderem, Alan, and David M. Underhill. "Mechanisms of Phagocytosis in Macrophages." Annual Review ofImmunology 17 (1999): 593–623.
Alberts, Bruce, et al. Molecular Biology of the Cell, 4th ed. New York: Garland Publishing, 2000.
Mukherjee, Sushmita, Richik N. Ghosh, and Frederick R. Maxfield. "Endocytosis." Physiological Reviews77 (1997): 759–803.
Schmid, Sandra L. "Clathrin-Coated Vesicle Formation and Protein Sorting: An Integrated Process."Annual Review of Biochemistry 66 (1997): 511–548.
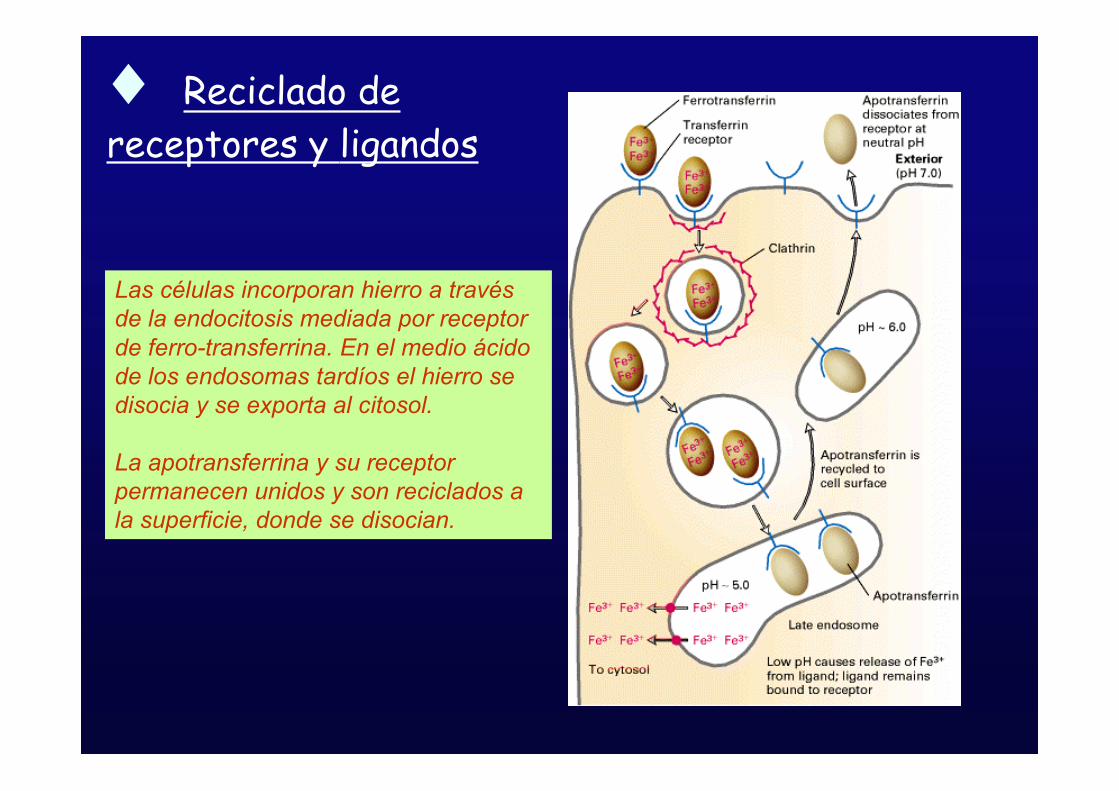
! Reciclado de
receptores y ligandos
Las células incorporan hierro a travésde la endocitosis mediada por receptorde ferro-transferrina. En el medio ácidode los endosomas tardíos el hierro sedisocia y se exporta al citosol.
La apotransferrina y su receptorpermanecen unidos y son reciclados ala superficie, donde se disocian.

!Desensibilización y reciclado de receptores
Los receptores unidos al EGF seautofosforilan y activan vías de señalizaciónintracelulares.Los receptores se endocitan en vesículascon clatrina.En los endosomas tempranos una fracciónde los receptores son defosforilados yreciclados a la superficie.
Sorkin & Zastrow, NRMCB 2002
EGF
dephosphorylation
auto
phosphorylation
EGF: epidermal growth factor

!Transporte de receptores y ligandos a lisosomas
Endosomas tempranos que transportan proteínas endocitadas (azul) y vesículas que transportanproteínas lisosomales desde el trans-Golgi (rojo y verde) se fusionan con endosomas tardíos. Lasproteínas a ser degradadas se internalizan por invaginación formando cuerpos multivesiculares.Eventualmente, los cuerpos multivesiculares se fusionan con lisosomas.
Sorkin & Zastrow, NRMCB 2002
la internalización de receptorestirosina kinasas (EGFR) en los
cuerpos multivesiculares impidesu reciclado y termina la
señalización.
citosol

La mono-ubiquitinilación actúa como una señal de direccionamiento al lisosoma
Mosesson / Yarden, IMAJ 2006
Receptores tirosina kinasa activados (ej. EGFR), E-caderinas, etc. pueden ser monoubiquitinados por E3-ubiquitina ligasas (ej. Cbl). Esto constituye una señal que es reconocida por proteínas adaptadoras (ej. Eps15,Hrs, STAM) que contribuyen a concentrar los receptores en cuerpos multivesiculares (MVB) y a su degradaciónen lisosomas.

FAGOCITOSIS
La fagocitosis es un tipo de endocitosis en donde seingieren partículas grandes o microorganismos enteros. Esun proceso mediado por actina. Extensiones de la membranaplasmática envuelven el material ingerido formando vesículasgrandes llamadas FAGOSOMAS.
Eucariontes unicelulares (Protozoa) se alimentan a través deeste medio, mientras que los animales superiores poseencélulas especializadas para tal fin como los macrófagos oneutrófilos.Estas células ingieren microorganismos y también estáninvolucradas en la limpieza de células apoptóticas osenescentes.

Neutrófilos: se encuentran en gran cantidad ensangre, pueden entrar rápidamente a los tejidos yfagocitan patógenos durante una inflamación aguda.
Macrófagos: están fuertemente relacionados con losmonocitos en sangre, estas células predominan eninflamaciones crónicas.
Células dendríticas y linfocitos B: la fagocitosis enestas células es importante durante la elaboración deuna respuesta inmune específica y no durante ladestrucción directa del patógeno

La fagocitosis consiste en la ingesta de partículasgrandes a partir de la formación de pseudópodos
Microscopía electrónica de barrido de un macrófago fagocitando dos eritrocitos. Las flechas señalan los bordes de las extensiones citoplasmáticas ricas enactina denominadas pseudópodos, que engolfan los eritrocitos.
Alberts et al., MBC 2002

Microscopía electrónica de un neutrófilo fagocitando unabacteria, que está en proceso de división.


Existe fagocitosis de:
MicroorganismosPartículas insolublesCélulas dañadas o muertasDebris celular
Diferentes etapas durante la fagocitosis
QuimiotaxisAdherenciaFormación de pseudópodosFormación del fagosomaFormación del fagolisosoma

La fagocitosis se inicia por el pegado de partículas(material inerte, microorganismos o células muertas) areceptores en la superficie celular, posiblemente aextensiones cortas, “pseudopodos”, presentes en lasuperficie de células como pueden ser los macrófagos.

Posteriormente los eventos rápidos de señalización promueven lareorganización del citoesqueleto (mayoritariamente actina) y comoconsecuencia la extensión de los pseudopodos. A pesar que la mayoría delos modelos de fagocitosis proponen a la membrana plasmática como la
principal fuente de membrana para la formación de los fagosomas(proceso de invaginación) (Aa) se ha demostrado que otrasendomembranas también contribuyen a este proceso.

Los endosomas reciclados, que son reclutados a la superficiecelular mediante el proceso denominado exocitosis (Ab), hansido identificados como potencial fuente de membrana para la
formación de los fagosomas.

También se ha demostrado que otra importante fuente de membrana queestá involucrada en la fagocitosis es el retículo endoplásmico (ER), en unproceso denominado fagocitosis mediada por ER (Ac). Existen muchostrabajos en donde se muestra que las proteínas que usualmente están en lamembrana plasmática o en los endosomas reciclados (Rab4 y Rab11) y enER (calreticulina y calnexina) también están presentes en los fagosomasindicando la potencial participación de estas membranas en la fagocitosis.

En algunos casos se demostró que los lisosomas también se fusionan con lamembrana plasmática durante la fagocitosis (Ad). Se pudo observar que enalgunos fagosomas se encuentran asociadas proteínas lisosomales (LAMP)y proteínas reticulares (calnexina) lo que indica que ambas organelasparticiparon en la contribución de membrana para la biogénesis de dichocompartimiento.

Luego de su formación, los fagosomas maduran en fagolisosomas mediante una seriede eventos de fusión con endosomas tempranos (EE), endosomas tardíos (LE) ylisosomas (Ly). Este proceso de maduración permite a los fagosomas adquirirpropiedades microbicidas y la habilidad de procesar antígenos.

Factores importantes que ayudan a destruir
microorganismos en los fagolisosomas
Proteínas NADPH oxidasa: están en las membranas de losfagolisosomas y generan radicales oxígeno que reaccionan con proteínas,lípidos y otras moléculas biológicas.
Oxido nítrico: reacciona con radicales oxígeno y generan moléculas quedañan otras moléculas biológicas.
Proteínas antimicrobianas: proteasas, elastasas. Son esenciales paraeliminar varias bacterias. Otra proteína antimicrobiana es la lisosima que actúasobre la pared celular de algunas bacterias gram positivas.
Péptidos antimicrobianos: defensinas y otros péptidos. Atacan lamembrana celular de la bacteria.
Transportadores de hidrógeno: acidifican el fagolisosoma provocando laeliminación de varios microorganismos. También activan proteasas
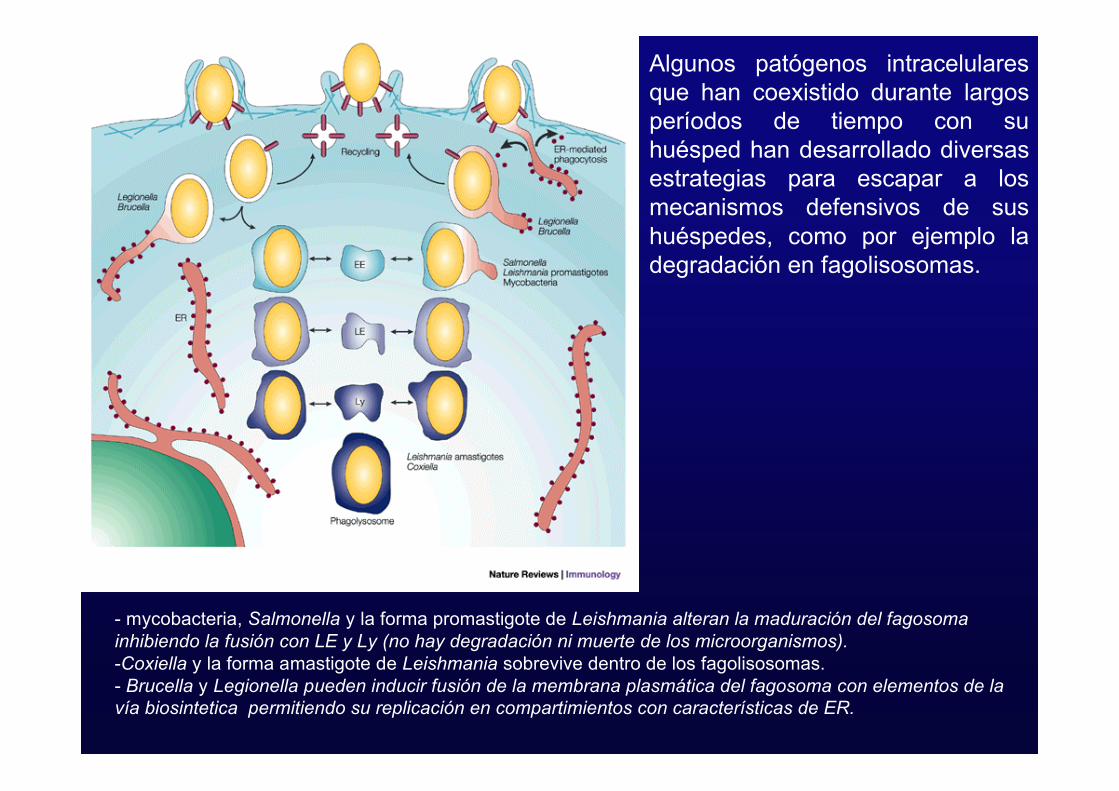
- mycobacteria, Salmonella y la forma promastigote de Leishmania alteran la maduración del fagosomainhibiendo la fusión con LE y Ly (no hay degradación ni muerte de los microorganismos).-Coxiella y la forma amastigote de Leishmania sobrevive dentro de los fagolisosomas.- Brucella y Legionella pueden inducir fusión de la membrana plasmática del fagosoma con elementos de lavía biosintetica permitiendo su replicación en compartimientos con características de ER.
Algunos patógenos intracelularesque han coexistido durante largosperíodos de tiempo con suhuésped han desarrollado diversasestrategias para escapar a losmecanismos defensivos de sushuéspedes, como por ejemplo ladegradación en fagolisosomas.

Endocitosis independiente de clatrina
Las caveolas están presentes en la mayoría de las células. Se forman en dominios especializados de lamembrana plasmática ricos en colesterol, glicoesfingolípidos y proteínas unidas por GPI (glico fosfatidil inositol)denominados "lipid rafts".
Caveolina: es la principal proteína estructural de las caveolas. La selección del cargo esta determinada por lacomposición lipídica de la membrana de la caveola.
vesícula conclatrina
Microscopía electrónica de caveolas
caveolas
membranaplasmática
100 nm
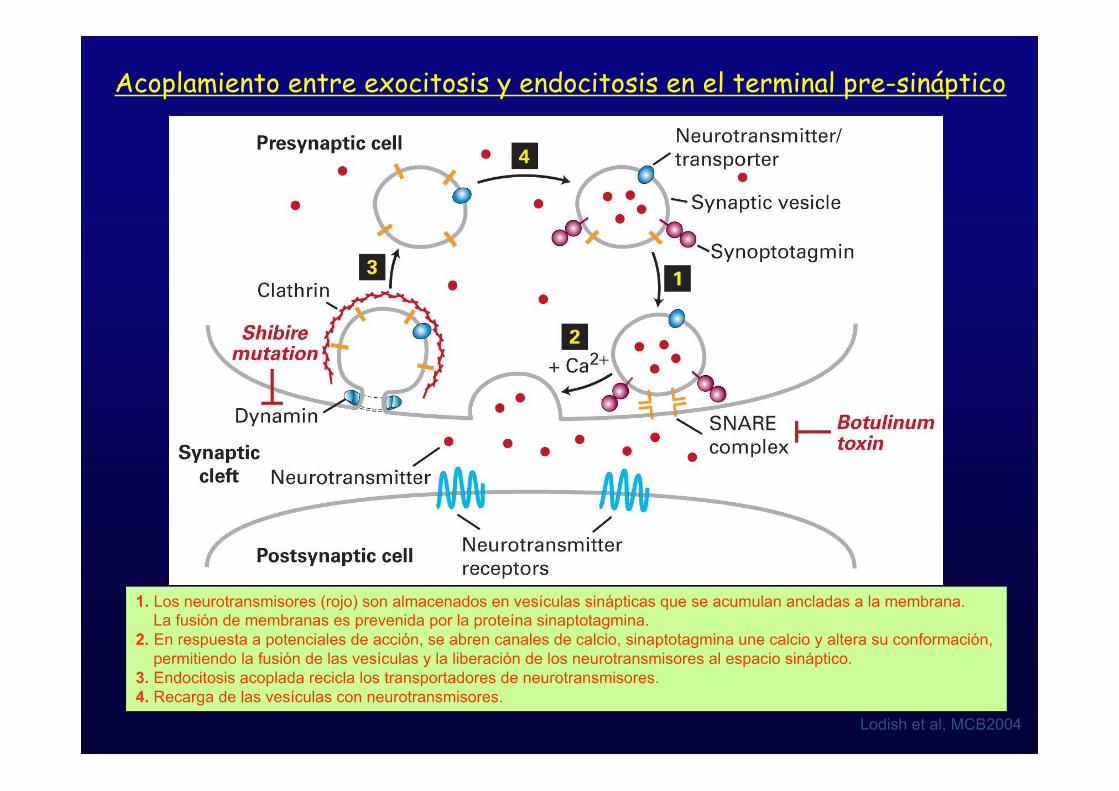
Acoplamiento entre exocitosis y endocitosis en el terminal pre-sináptico
Lodish et al, MCB2004
1. Los neurotransmisores (rojo) son almacenados en vesículas sinápticas que se acumulan ancladas a la membrana. La fusión de membranas es prevenida por la proteína sinaptotagmina.2. En respuesta a potenciales de acción, se abren canales de calcio, sinaptotagmina une calcio y altera su conformación, permitiendo la fusión de las vesículas y la liberación de los neurotransmisores al espacio sináptico.3. Endocitosis acoplada recicla los transportadores de neurotransmisores.4. Recarga de las vesículas con neurotransmisores.
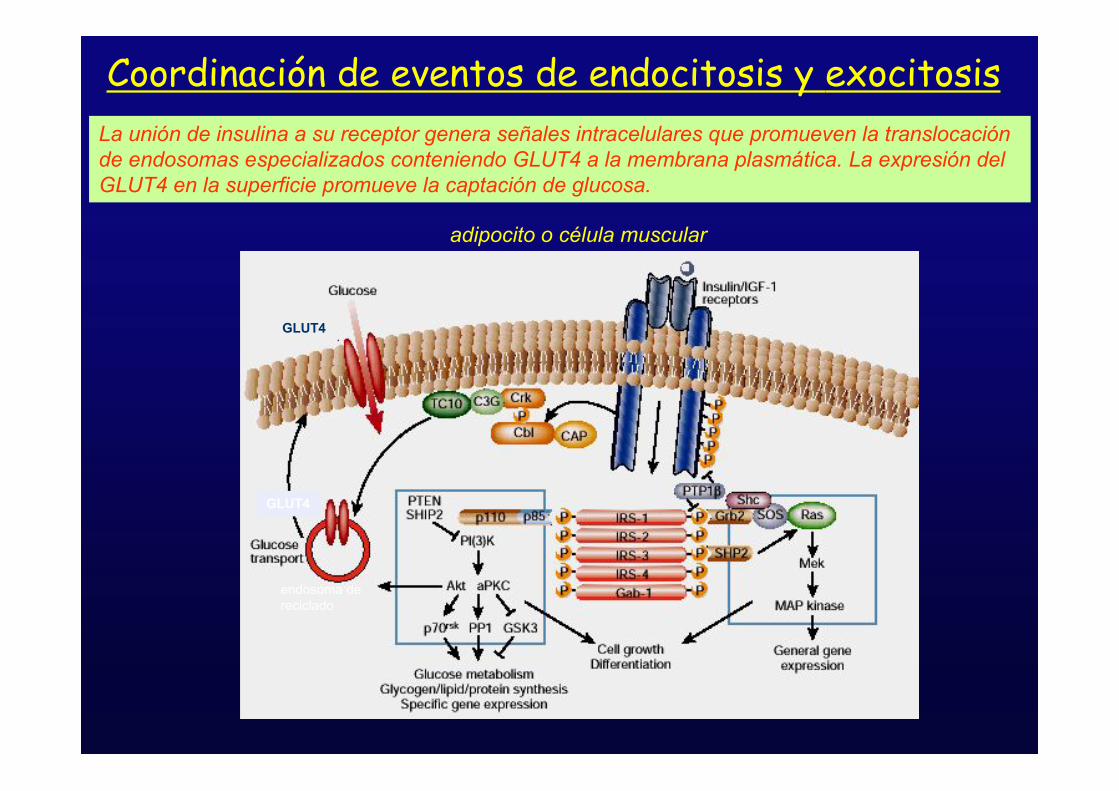
GLUT4
Coordinación de eventos de endocitosis y exocitosis
La unión de insulina a su receptor genera señales intracelulares que promueven la translocaciónde endosomas especializados conteniendo GLUT4 a la membrana plasmática. La expresión delGLUT4 en la superficie promueve la captación de glucosa.
GLUT4
adipocito o célula muscular
endosoma dereciclado

Autofagocitosis

Autofagocitosis o Autofagia esel término que se utiliza para
la degradación decomponentes citoplasmáticos
en los lisosomas.

Secuestro de material(transporte a los lisososmas)
Degradación
Generación de aminoácidos / péptidos(utilización de los productos obtenidos)
Pasos secuenciales de la autofagia
Cada uno de estos pasos cumple diferentes funciones de acuerdo en el contextocelular en el que se encuentre.

Existen tres tipos principales de autofagia
- Microautofagia
- Macroautofagia
-Autofagia mediada por chaperonas
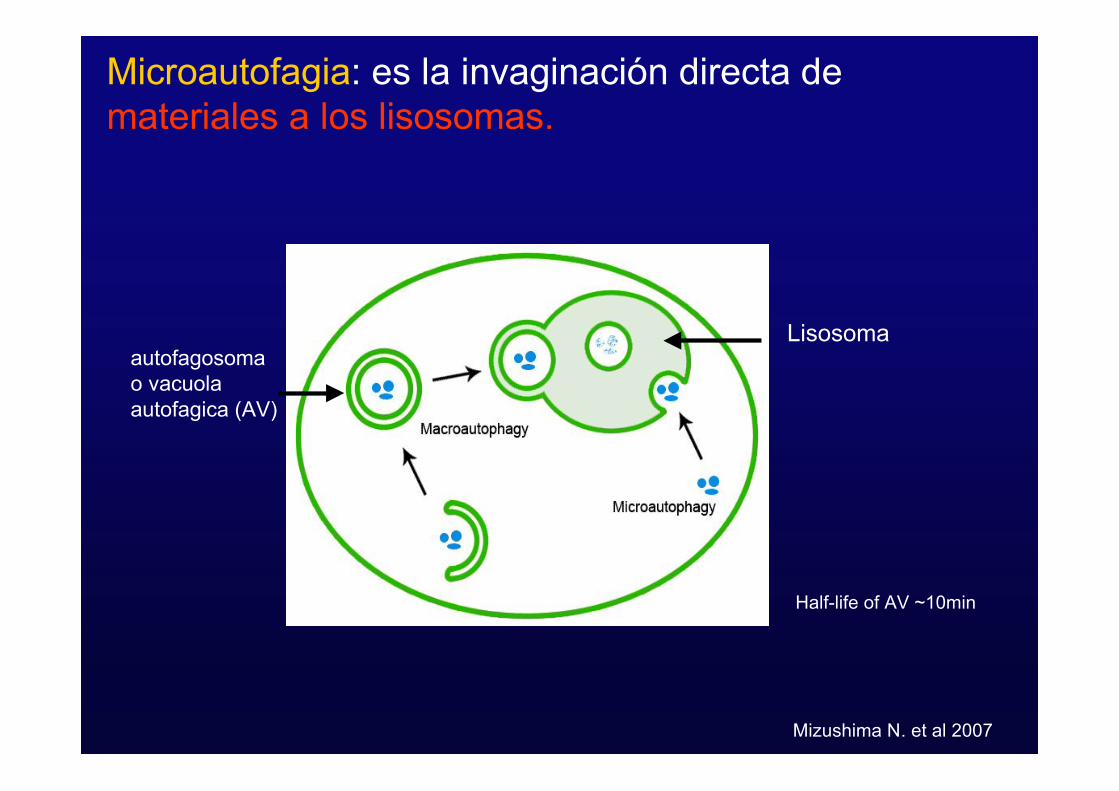
Microautofagia: es la invaginación directa demateriales a los lisosomas.
Half-life of AV ~10min
Lisosomaautofagosomao vacuolaautofagica (AV)
Mizushima N. et al 2007
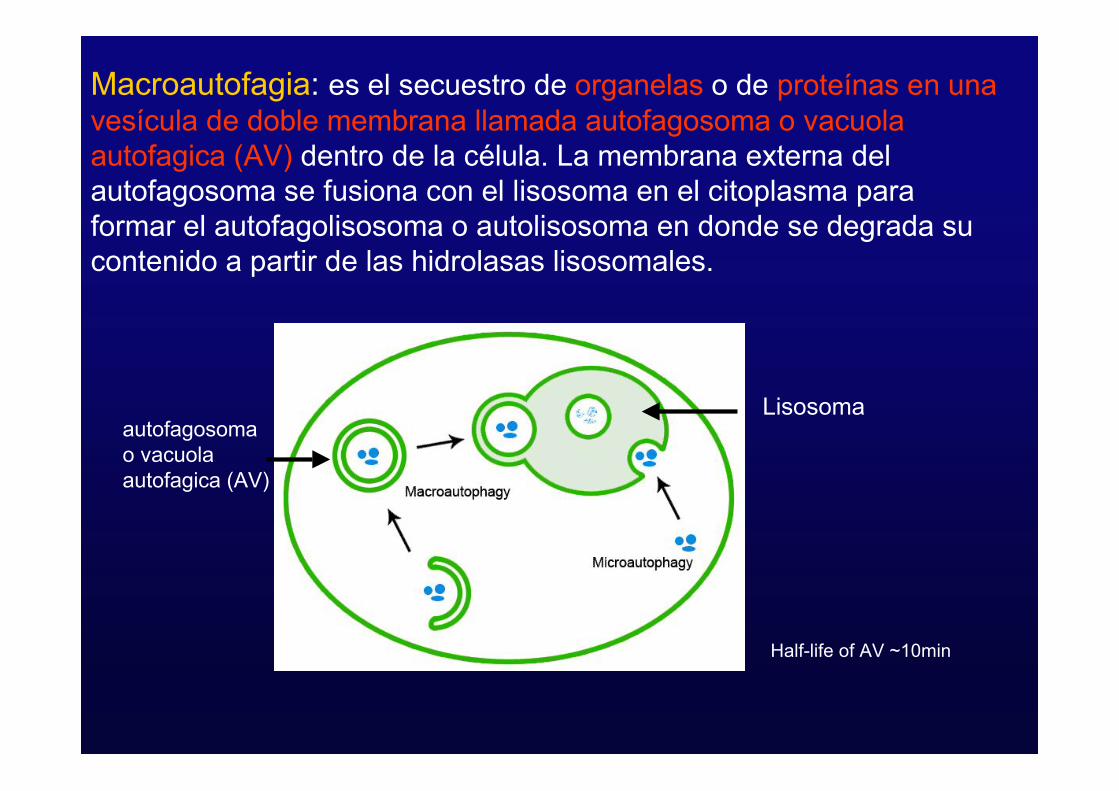
Macroautofagia: es el secuestro de organelas o de proteínas en unavesícula de doble membrana llamada autofagosoma o vacuolaautofagica (AV) dentro de la célula. La membrana externa delautofagosoma se fusiona con el lisosoma en el citoplasma paraformar el autofagolisosoma o autolisosoma en donde se degrada sucontenido a partir de las hidrolasas lisosomales.
Half-life of AV ~10min
Lisosomaautofagosomao vacuolaautofagica (AV)

Autofagia mediada por chaperonas (CMA): es eltérmino que se utiliza para describir la degradación de
proteínas citosólicas específicas que contienen unasecuencia peptídica determinada. Las chaperonas se
unen a estas proteínas y las transportan a los
lisosomas por medio de un receptor. (este es un
proceso selectivo y sólo involucra proteínas, no
transportan organelas, contrariamente a la autofagia
en donde el sistema de degradación es en general no
selectivo).

Proteína 31AtgHalf-life of AV ~10min
Lisosomaautofagosomao vacuolaautofagica (AV)

Mizushima N. et al 2007

Uno de los principales factores que es considerado comodisparador del proceso de autofagia es la reducción denutrientes.
Levaduras: falta de nitrógeno es el principal estimulo parainducir autofagia.
Plantas: falta de nitrógeno y carbono.
Mamíferos: sistema altamente complicado. Depleción total deaminoácidos. Efecto individual de los aminoácidos. Dependeel tipo de célula.Aun no se conoce como censan la falta de alguno de los aminoácidos.(candidato: GCN2, proteína kinasa de pegado a tRNA)
Mizushima N. et al 2007

Otra postura es la que considera que la autofagia deberíaestar regulada por un sistema altamente organizado.
Se cree que el sistema endocrino, en particular la insulina,regula la autofagia in vivo. (Autofagia en hígado es suprimidapor insulina y aumentada por glucagon).También contribuyen a la regulación de la autofagia otrashormonas y factores de crecimiento.
Mizushima N. et al 2007

Una vez que el material es degradado en los lisosomas /vacuolas, las unidades monoméricas (ej. Aminoácidos) sonexportados al citosol para ser reusados.
Atg22 (levaduras)Avt3, Avt4 (levaduras) -SLC36A1 / LYAAT-1 transportador lisosomal deaminoácidos (mamíferos).
Aun no se conoce contribución de la autofagia para lareutilización de lipidos y carbohidratos.
DEGRADACIÓN y REUTILIZACIÓN

Autofagia es una vía de degradación lisosomal que esescencial para la sobrevida, la diferenciación, el desarrolloy el mantenimiento de la célula.
Autofagia cumple principalmente un rol adaptativo deproteger a los organismos de diversas patologías, comopueden ser infecciones, cancer, enfermedadesneurodegenerativas.
Sin embargo, esta función puede serdestructiva o nociva para el organismo.
Excesiva degradación por autofagia causamuerte celular.

Control de calidad
Eliminacion de exceso de organelas no necesarias (peroxisomas, mitocondrias)
Streptococcus, Shigella, Mycobacterium. Algunos microorganismos (Brucella,Legionella) utilizan la via autofagica para sobrevivir. (vacuolas protectivas)


