electrotonic coupling in a network of compartmental external urethral sphincter motoneurons of...
TRANSCRIPT

*Corresponding author.E-mail address: [email protected] (M. Heldoorn).
Neurocomputing 38}40 (2001) 647}658
Electrotonic coupling in a network of compartmentalexternal urethral sphincter motoneurons of
Onuf 's nucleus
Marcel Heldoorn��*, Johan L. Van Leeuwen�, Jan Vanderschoot�,Enrico Marani���
�Department of Neurosurgery, Leiden University Medical Center (LUMC), P.O. Box 9604,Wassenaarseweg 62, NL-2300 RC Leiden, Netherlands
�Experimental Zoology Group, Wageningen Institute of Animal Sciences (WIAS),Wageningen University, Marijkeweg 40, P.O. Box 338, NL-6700 AH Wageningen, Netherlands
�Department of Medical Informatics, Leiden University Medical Center (LUMC),Wassenaarseweg 62, P.O. Box 2086, NL-2301 CB Leiden, Netherlands
�Department of Signals and Systems, Faculty of Electroengineering and BMTI,University Twente, Enschede, Netherlands
Abstract
A compartmental model of motoneurons of Onuf 's nucleus, the external urethral and analsphincter motor nucleus, was constructed from morphological and electrophysiological datapreviously reported in the literature. The model neurons have active somata and initial axonsegments. The dendritic trees are represented by one equivalent dendritic cylinder. These modelneurons were used to perform network simulations. The cells in the network were mutuallyconnected by dendrodendritic gap junctions, which were modeled as ohmic resistances betweentwo neuronal compartments. Circularly propagating waves and extinction of meeting waves areto be expected. Increasing random input injection will cause fastening of the network activation,which could be related to Onuf 's function. This could mean that a small continuous inputto this nucleus from a higher center is only needed to cause a fast massive activation ofthe network, resulting in fast contraction of the sphincter. � 2001 Elsevier Science B.V. Allrights reserved.
Keywords: Multi-compartment model; External urethral sphincter; Onuf 's nucleus; Moto-neurons; Gap junction
0925-2312/01/$ - see front matter � 2001 Elsevier Science B.V. All rights reserved.PII: S 0 9 2 5 - 2 3 1 2 ( 0 1 ) 0 0 4 2 3 - 4

1. Introduction
Onuf 's nucleus in man and animals is restricted to the ventral horn of the "rst,second and third sacral segment in the spinal cord. This group of neurons isresponsible for steering the urethral and anal sphincters. Although serious debates arefound in literature concerning the sympathic, parasympathic, dual or somatic charac-ter of these neurons, it is well established that they constitute a specialized spinal cordnucleus. The dorsomedial subnucleus innervates the rectal striated sphincter, whilethe ventrolateral subgroup connects to the urethral striated sphincter [2]. Histochem-istry showed these neurons to contain peptides related to the autonomic system, whilethe synaptic distribution resembles somatic or parasympathic input. However, en-kephalin immunoreactive varicosities are abundantly present in this nucleus [20], inthis respect it distinguishes itself from somatic motoneurons, as does the genderdi!erence in this nucleus. Ultrastructural studies showed longitudinally runningbundles of closely packed dendrites in Onuf 's nucleus [2,23,26]. Casual appositionand specialized electrotonical contacts are present between the dendrites [26]. Mor-phology also showed recurrent axon collaterals. The dendrites extended outside thenucleus to other spinal cord gray matter areas.The importance of this nucleus was rediscovered in relation to the preservation of
urethral and anal functions in Amyotrophic Lateral Sclerosis [17]. Onuf 's nucleuswas also spared in several other diseases (e.g. spinal muscular atrophy and healedpoliomyelitis), while other somatic sacral motoneurons degenerated [25]. The reverseis found in multiple system atrophy and Shy-Drager Syndrome [6,17].Since there is no model of the urethral or anal sphincter re#ex loop in literature,
a project was initiated to model this re#ex loop consisting of a muscular partand a neuronal part. This paper concerns the modeling of a neuronal network.The network is based on simple model neurons, in which Onuf 's morphologicaldendritic characteristics together with �-motoneuron membrane properties are imple-mented, since only a few physiological characteristics of neurons in Onuf 's nucleus areknown.
2. Material and methods
2.1. Physiology
A variety of passive electrical membrane properties [10,21] and detailed descrip-tions of the morphology [2,22] of Onuf 's external urethral and anal sphincter (EUSand EAS, respectively) motoneurons have been published. Features of these mo-toneurons that are of particular interest include their low rheobase values [10,21] andtheir lack of recurrent inhibition [14,16] in spite of the presence of recurrent axoncollaterals [22]. In Table 1 these characteristic values of EUS motoneurons are given[10]. The only signi"cant di!erence between EUS and EAS neurons is the membranetime constant which is twice as long in EAS neurons. Relating these observations tothe electrical properties of hindlimb motoneurons, the similarity between EUS and
648 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658

Table 1Electrical properties of EUS motoneurons. Mean values and standard deviation are presented with samplesizes in parenthesis. Values according to Hochman et al. [10]
Electrical property Value for EUS neuron
Action potential amplitude (71$10) mV [14]Conduction velocity (47$14) m/s [17]Membrane time constant (2.1$0.2) ms [3]Input resistance (2.0$0.7) M� [13]Electrotonic length (1.37$0.05) [3]Rheobase (3.1$1.7) nA [10]Threshold voltage (7.1$3.0) mV [9]Rheobase � resistance (5.2$1.4) mV [9]After hyperpolarization duration (97$22) ms [11]AHP peak voltage (1.5$0.7) mV [12]
&tonic' or &slow' hindlimb motoneurons is striking: the values for input resistance,rheobase, and threshold voltage are comparable [5,7,9,11].The whole neuron surface has not been measured, but the total cell capacitance, an
estimate of cell surface can be calculated from the membrane time constant, electro-tonic length and input resistance values [19]. A calculated cell capacitance of 3.4 nFfor EUS neurons is half the value reported for slow hindlimb motoneurons [11],suggesting that the EUS neurons are smaller than slow hindlimb motoneurons. Thissuggestion is consistent with the lower conduction velocity and smaller reported somasize [2].
2.2. Morphology
Sasaki [22] found long dendrites directed rostrally or caudally within Onuf 'snucleus, more prominently in EUS motoneurons. EAS motoneurons had longer endbranches compared with EUS neurons, which had a relatively smaller number of endbranches, summed length and surface area of a dendrite. The dendritic surface areaand the dendritic volume were tightly correlated with the diameter of the "rst-orderdendrite. The latter relation was almost identical between EUS, EAS, and hindlimb �-and �-motoneurons. The large values for the dendritic-to-soma surface area ratio (31in EUS, 50 in EAS) con"rm the well-developed dendritic arborization which iscomparable to hindlimb motoneurons.
2.2.1. Dendritic trees equivalent to a cylinderRall [18] developed the concept of &trees equivalent to a cylinder'. Rall argued that
when a cylinder with diameter D bifurcates into two daughter branches with dia-meters d
�and d
�, the branch point behaves as a continuous cable for current that
#ows from the parent to the daughters, if D���"�d����
. The mean ratio of �d����
toD��� was 0.99 in EUS and 1.08 in EAS motoneurons. �d���
�mostly equalled
M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658 649

D��� irrespective of the thickness of the parent dendrites and of the distance fromthe cell body (see Fig. 11 in [22], indicating that the 3/2 power constraint atbranching points is nearly satis"ed, and suggesting that Rall's equivalent-cylindermodel is applicable to sphincter motoneurons. In addition, the EUS motoneuronshave equivalent cylinder electrotonic lengths similar to those of hindlimb mo-toneurons [9,11].
2.3. Compartmental model
The previous sections indicate that EUS motoneurons are both physiologically andmorphologically comparable to hindlimb motoneurons. This enables us to use hind-limb motoneurons data to substitute unknown parameter values. In order to consti-tute a neuronal network a single cell model of Onuf 's neurons is a necessaryprerequisite. Therefore, an equivalent cylinder model of the single cell was construc-ted. The neuronal unit was modeled using compartmental methods (see for a review:Segev et al. [24]. The model neurons consist of a soma segment, an initial segment andan equivalent dendritic cylinder. The dimensions of the compartments were based onreconstructions of EUS neurons reported in literature [22]. The model soma diameteris 29.9 �m, which is the mean soma diameter of EUS neurons according to Sasaki[22]. Initial segment length and diameter were not measured, but Jones and Bawa[15] used 100 �m and 5 �m, respectively.The passive parameters determining the cable properties of the model neuron were
set to the values reported by Hochman et al. [10] for EUS cells and Fleshman et al.[8] for �-motoneurons. Speci"c membrane capacitance C
�was 1.0 �F cm��, speci"c
cytoplasmatic resistivity R�was 70 � cm and speci"c membrane resistivity R
�was
6.9 k� cm� for the soma and dendrites, respectively.In the model neuron, all dendritic trees are represented by one equivalent cylinder.
The electrotonic length ¸ of the equivalent cylinder was calculated by the formula[13] ¸"�/�(�
/�
�)!1. The electrophysiological properties used to match cell
morphology consist of the time constants of voltage decay after short current pulses,�"6.9 ms, �
�"1.2 ms [22]. The electrotonic length of the equivalent cylinder ¸ is
1.44. The diameter of the equivalent cylinder d��
was estimated from the equationd��
"(�D���
)���. The 3/2 power value of a stem dendrite's diameter Dis a mor-
phological parameter related to the input conductance of the dendrite, so the summedvalue of D���
of all stem dendrites (�D���
) is related to the input conductance of the
whole of dendritic branches of a cell [18,19]. The mean value for �D���
in EUSneurons is 70.0 �m��� [22], so the diameter of the equivalent cylinder d
��is 17.0 �m.
The electrotonic length of a cable of physical length l and diameter d is ¸"l/�, wherethe space constant �"�dR
�/4R
�. The space constant � of the equivalent cylinder is
3485 �m. The physical length of the cylinder l is 5017 �m (¸�"1.44�3485 �m). Thedendrite equivalent cylinder terminated in a closed end, as the initial segment does.The equivalent dendrite was subdivided in 10 compartments of 502 �m, so that nocompartment had an electrotonic length '0.2� [24]. To refer to the di!erentdendritic compartments, the compartments were named d0,2, d9, with the compart-ment most proximal to the soma being d0 (Fig. 1).
650 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658

Fig. 1. Single cell model with initial segment (IS), soma compartment and ten dendritic compartments(d0,2, d9).
Active properties were similar to the S type motoneuron model of Jones and Bawa[15]. Five voltage-gated ionic currents, simulated by standard Hodgkin}Huxleyequations [12] were associated with the initial segment and soma compartments only(see Appendix A). The soma compartment contained a fast sodium conductance (seeEq. (15)), a fast potassium conductance (see Eq. (12)) and a slow potassium conduc-tance (see Eq. (20)), which gave rise to the afterhyperpolarization. The initial segmentcontained a fast delayed recti"er type potassium conductance (see Eq. (4)) and a fastsodium conductance (see Eq. (7)).
2.4. The network model connected by gap junctions
The single cell model was used to construct a neuronal network. Cells are laid downon a rectangular grid. The network model consists of a matrix of 25�25 neuronalunits. The neurons were mutually connected by dendrodendritic gap junctionsbetween the three most proximal dendritic compartments d0, d1, and d2. The gapjunctions were modeled as ohmic resistances of 1 M�.
2.5. Simulations
All simulations were performed with the GENESIS software [4] on a IntelPentium-III 450 MHz.
3. Results
3.1. Single cell and two coupled cells
In Fig. 2 a voltage trace of a single model neuron is shown. The trace was evoked bya triangular shaped current injection of 5 nA at the soma. The phase of the signal was2703 and the frequency was 1 Hz. Increase of the injected current is indeed followed bythe neuron model with a frequency increase of the action potentials.In Fig. 3 the voltage traces of two model neurons are shown. The neurons were
mutually connected by dendrodendritic gap junctions between the three most prox-imal dendritic segments (d0}d2). A triangular shaped current of 0.5 nA was injected atthe soma of neuron EUS[0] (see Fig. 3). The bidirectional electrotonic coupling of thetwo neurons allows the #ow of current from one to the other cell. Notice that the "re
M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658 651
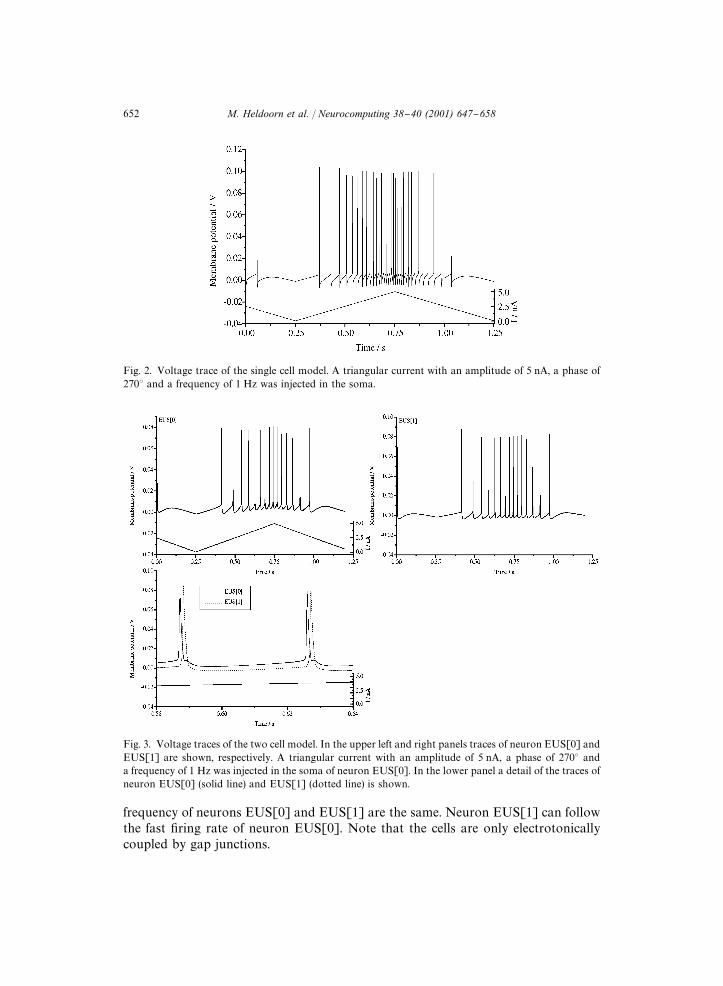
Fig. 2. Voltage trace of the single cell model. A triangular current with an amplitude of 5 nA, a phase of2703 and a frequency of 1 Hz was injected in the soma.
Fig. 3. Voltage traces of the two cell model. In the upper left and right panels traces of neuron EUS[0] andEUS[1] are shown, respectively. A triangular current with an amplitude of 5 nA, a phase of 2703 anda frequency of 1 Hz was injected in the soma of neuron EUS[0]. In the lower panel a detail of the traces ofneuron EUS[0] (solid line) and EUS[1] (dotted line) is shown.
frequency of neurons EUS[0] and EUS[1] are the same. Neuron EUS[1] can followthe fast "ring rate of neuron EUS[0]. Note that the cells are only electrotonicallycoupled by gap junctions.
652 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658

Fig. 4. Simulation experiments with the network model. The two upper rows show a 12.5 ms simulation: inthe "rst column the network state at t"2.5 ms is shown; the time gap between two frames is 2.5 ms. Thegray values in the graphs refer to the scale bar which shows the values of the membrane potential in thesomata in volts (V). Upper row: simulation of the network with one neuron getting a current input (1 Hztriangular shaped current input with an amplitude of 20 nA and a phase of 903). Middle row: simulation ofthe network with two neurons getting a current input (see description above). Lower row: a 7.5 mssimulation: in the "rst column the network state at t"1.50 ms is shown; the time gap between two frames is1.5 ms; network simulation with "ve randomly chosen cells receiving the current input.
3.2. Network simulation
Fig. 4 shows network simulations. The "rst simulation shows the network stateafter applying a current input in one cell in the center of the network. A propagatingwave of activity in the network is initiated. The waves form a closed connected front.The second simulation in Fig. 4 shows a similar experiment in which the current isapplied into two cells. The waves originating from the activated cells collide, but donot cross one another. The two waves accrete, forming one wave front. In the thirdsimulation 10 randomly chosen cells receive a current input. This leads to a fastmassive spread of activity over the whole network.
4. Discussion
Onuf 's nucleus is, besides its important role in urine and faecal neurogenic inconti-nence, a speci"ed group of motoneurons, characterized by tight electrotonic coupling.This is only inherent to a selected group of nuclei in the central nervous system.Moreover, Onuf 's nucleus is responsible for very fast and long lasting continuous
M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658 653

contraction of the urethral and anal sphincter. Due to transdi!erentiation fromsmooth to striated muscles of these sphincters during development its particularitystands out even more.Any modeling study of Onuf 's nucleus will have to emphasize these peculiarities,
which have been organized in this paper by implementing several specialized ionchannels together with electrotonic coupling.The "re capacity of the modeled neurons has been demonstrated in Fig. 2, showing
that the manner of injection is followed by the modeled neuron. Coupling of two ofthese neurons shows an acceptable time-lag in the non-injected neuron.When coupling neurons in a grid of 25�25 the propagating wave is circular-like
over the network, if injection occurs in one neuron in the center of the network.Putting in two injections on opposite sides of the network the propagating waves willextinct each other, while at random injections in the network show propagatingactivity waves, fastening the complete activation of the network.Circularly propagating waves and extinction of meeting waves are to be expected.
Current injection at random will cause fastening of the network activation, whichcould be related to Onuf 's function. This could mean that a small continuous input tothis nucleus from a higher center is only needed to cause a fast massive activation ofthe network, resulting in fast contraction of the sphincter, like in anal #atulation orupholding continence during coughing. This hypothesis is supported by the "nding ofdescending projections to Onuf 's nucleus from the BoK tzinger complex neurons, whichare responsible for generating the cough motor pattern [3] indicating a possible rolein preventing incontinence during coughing. In addition, the lack of recurrent inhibi-tion in sphincter motoneurons [16] and the presence of excitatory recurrent axoncollaterals would also accentuate the excitability of the neurons in Onuf 's nucleus.The in#uence of excitatory recurrent axon collaterals on the network behavior will
be the topic of further investigation.
Appendix A. Formulas
The constitution of the ionic currents in the soma and initial axon segment wasbased on motoneuron literature [1,15,27]. Ionic currents in the soma can be describedby
I��
"g�(<
!E
�)#(g
�#g
���� )(<
!E
�)
"g��
m�h(<
!E�)#(g�
�n�#g�
� �� p�)(<
!E
�), (A.1)
where g���
is the maximum conductance for the sodium (Na), potassium (K) and slowpotassium (K
��� ) ion channels; m, h, n and p are time- and voltage-dependent para-
meters, which determine the magnitude and time course of the activation andinactivation of the respective currents. The time and voltage dependence is given by
d�dt
"�� (1!�)!��"
��
!���
, (A.2)
654 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658

where
��"1
��#�, �
�"
����#�
. (A.3)
The parameters m, h, n and p are represented in the above general equations as �. Themagnitudes and time courses of the activation/inactivation parameters � and aregiven below.
A.1. Initial segment currents
A.1.1. K� current [1]
I��� �
"g���� �
n�(V
!E�), (A.4)
g���� �
"100 mS cm��, E�
"!10 mV,
��"
0.2#(!0.02<)
e������������!1, (A.5)
�"
0.15
e�����������������!0.01. (A.6)
A.1.2. Na� current
I��� �
"g���� �
m�h(<
!E�), (A.7)
g���� �
"500 mS cm��, E�
"115 mV,
��
"
4#(!0.4<)
e������������!1, (A.8)
�
"
(0.4<)!14
e����������!1, (A.9)
��"
0.16
e�����������������, (A.10)
�"
4
e������������#1. (A.11)
A.2. Soma currents
A.2.1. K� current [1]
I�
"g��
n�(<
!E�), (A.12)
g��
"35 mS cm��, E�
"!10 mV,
��"
0.4#(!0.02<)
e������������!1, (A.13)
M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658 655

�"
0.16
e�����������������!0.032. (A.14)
A.2.2. Na� current
I�
"g��
m�h(<
!E�), (A.15)
g��
"140 mS cm��, E�
"115 mV,
��
"
7#(!0.4<)
e���������������!1, (A.16)
�
"
(0.4<)!18
e����������!1, (A.17)
��"
0.15
e�����������������, (A.18)
�"
4
e������������#1. (A.19)
A.2.3. Slow K� current [1]
I����
"g�����
p�(<
!E�), (A.20)
g�����
"12 mS cm��, E�
"!10 mV,
��"
3.5
e�������������#1, (A.21)
�"
0.015
e�������������#1. (A.22)
References
[1] J.N. Barrett, W.E. Crill, Voltage clamp of cat motoneurone soma: properties of the fast inwardcurrent, J. Physiol. London 304 (1980) 231}249.
[2] M.S. Beattie, Q. Li, M.G. Leedy, J.C. Bresnahan, Motoneurons innervating the external anal andurethral sphincters of the female cat have di!erent patterns of dendritic arborization, Neurosci. Lett.111 (1}2) (1990) 69}74.
[3] F. Bongianni, D. Mutolo, G.A. Fontana, T. Pantaleo, Discharge patterns of BoK tzinger complexneurons during cough in the cat, Am. J. Physiol. 274 (4 Pt 2) (1998) R1015}R1024.
[4] J.M. Bower, D. Beeman, The book of GENESIS: Exploring Realistic Neural Models with theGEneral NEural SImulation System, 1st Edition, Springer, New York, 1994.
[5] R.E. Burke, G. Ten Bruggencate, Electrotonic characteristics of � motoneurones of varying size,J. Physiol. (London) 212 (1) (1971) 120.
[6] D. Chalmers, M. Swash, Selective vulnerability of urinary Onuf motoneurons in Shy}Dragersyndrome, J. Neurol. 234 (4) (1987) 259}260.
656 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658

[7] J.W. Fleshman, J.B. Munson, G.W. Sypert, W.A. Friedman, Rheobase, input resistance, and motor-unit type in medial gastrocnemius motoneurons in the cat, J. Neurophysiol. 46 (6) (1981) 1326}1338.
[8] J.W. Fleshman, I. Segev, R.B. Burke, Electrotonic architecture of type-identi"ed �-motoneurons in thecat spinal cord, J. Neurophysiol. 60 (1) (1988) 60}85.
[9] B. Gustafsson, M.J. Pinter, Relations among passive electrical properties of lumbar �-motoneuronesof the cat, J. Physiol (London) 356 (1984) 401}431.
[10] S. Hochman, B. Fedirchuk, S.J. Shefchyk, Membrane electrical properties of external urethral andexternal anal sphincter somatic motoneurons in the decerebrate cat, Neurosci. Lett. 127 (1) (1991)87}90.
[11] S. Hochman, D.A. McCrea, E!ects of chronic spinalization on ankle extensor motoneurons. III.Composite Ia EPSPs in motoneurons separated into motor unit types, J. Neurophysiol. 71 (4) (1994)1480}1490.
[12] A.L. Hodgkin, A.F. Huxley, A quantitative description of membrane current and its application toconduction and excitation in nerve, J. Physiol. London 117 (1952) 500}544.
[13] W.R. Holmes, W. Rall, Estimating the electrotonic structure of neurons with compartmental models,J. Neurophysiol. 68 (4) (1992) 1438}1452.
[14] E. Jankowska, Y. Padel, P. Zarzecki, Crossed disynaptic inhibition of sacral motoneurones, J. Physiol.(London) 285 (1978) 425}444.
[15] K.E. Jones, P. Bawa, Computer simulation of the responses of human motoneurons to composite 1AEPSPS: e!ects of background "ring rate, J. Neurophysiol. 77 (1) (1997) 405}420.
[16] R. Mackel, Segmental and descending control of the external urethral and anal sphincters in the cat,J. Physiol. (London) 294 (1979) 105}122.
[17] T. Mannen, M. Iwata, Y. Toyokura, K. Nagashima, The Onuf 's nucleus and the external analsphincter muscles in amyotrophic lateral sclerosis and Shy}Drager syndrome, Acta. Neuropathol.(Berlin) 58 (4) (1982) 255}260.
[18] W. Rall, Branching dendritic trees and motoneuron membrane resistivity, Exp. Neurol. 1 (1959)491}527.
[19] W. Rall, Core conductor theory and cable properties of neurons, in: Kandel, E (Ed.), Handbook ofPhysiology, American Physiological Society, 1977, pp. 39}97.
[20] V. RammH rez-LeoH n, T. Hokfelt, A.C. Cuello, T.J. Visser, B. Ulfhake, Enkephalin-, thyrotropin-releasing hormone- and substance P-immunoreactive axonal innervation of the ventrolateral den-dritic bundle in the cat sacral spinal cord: an ultrastructural study, J. Chem. Neuroanat. 7 (4) (1994)203}215.
[21] M. Sasaki, Membrane properties of external urethral and external anal sphincter motoneurones in thecat, J. Physiol. London 440 (1991) 345}366.
[22] M. Sasaki, Morphological analysis of external urethral and external anal sphincter motoneurones ofcat, J. Comput. Neurol. 349 (2) (1994) 269}287.
[23] H.D. Schr~der, Onuf 's nucleus X: a morphological study of a human spinal nucleus, Anat. Embryol.(Berlin) 162 (4) (1981) 443}453.
[24] I. Segev, J.W. Fleshman, J.P. Miller, B. Bunow, Modeling the electrical behavior of anatomicallycomplex neurons using a network analysis program: passive membrane, Biol. Cybern. 53 (1) (1985)27}40.
[25] J.H. Sung, A.R. Mastri, Spinal autonomic neurons in Werdnig-Ho!mann disease, mannosidosis, andHurler's syndrome: distribution of autonomic neurons in the sacral spinal cord, J. Neuropathol. Exp.Neurol. 39 (4) (1980) 441}451.
[26] K. Takahashi, T. Yamamoto, Ultrastructure of the cell group X of Onuf in the cat sacral spinal cord,Z Mikrosk. Anat. Forsch. 93 (2) (1979) 244}256.
[27] R.D. Traub, Motorneurons of di!erent geometry and the size principle, Biol. Cybern. 25 (3) (1977)163}176.
Marcel Heldoorn was born in 1973 in Zwartsluis, The Netherlands. He received the M.Sc. degree inBiomedical Sciences in 1997. He is currently working as a Ph.D. student in the Neuroregulation group inthe Department of Neurosurgery of the Leiden University Medical Center. His current research topics arethe cross striated (external urethral and anal) sphincters and Onuf 's nucleus.
M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658 657

Enrico Marani studied biology. He was sta!member of the department of Neuroanatomy and produced hisPh.D. thesis on enzyme histochemistry of the mammalian cerebellum. In 1986 he became head of theNeuroregulation group at the Department of Physiology. In 1996 he was appointed professor in Neuro-physiology at the Technical University Twente.
658 M. Heldoorn et al. / Neurocomputing 38}40 (2001) 647}658