electron microscopy of replicating dna reveals replicating bubbles. how does one prove bidirectional...
TRANSCRIPT

Electron Microscopy of replicating DNA revealsreplicating bubbles. How does one prove
bidirectional fork movement?

Pulse with radiolabeled nucleotide; chase with coldnucleotide. Then do autoradiography

DNA Replication
DNA replication is semi-conservative, one strand serves as the template for the second strand. Furthermore, DNA replication only occurs at a specific step in the cell cycle. The following table describes the cell cycle for a hypothetical cell with a 24 hr cycle.
Stage Activity Duration G1 Growth and increase in cell size 10 hr S DNA synthesis 8 hr G2 Post-DNA synthesis 5 hr M Mitosis 1 hr
DNA replication has two requirements that must be met:
1. DNA template 2. Free 3' -OH group


DNA Replication
DNA replication is semi-conservative, one strand serves as the template for the second strand. Furthermore, DNA replication only occurs at a specific step in the cell cycle. The following table describes the cell cycle for a hypothetical cell with a 24 hr cycle.
Stage Activity Duration G1 Growth and increase in cell size 10 hr S DNA synthesis 8 hr G2 Post-DNA synthesis 5 hr M Mitosis 1 hr
DNA replication has two requirements that must be met:
1. DNA template 2. Free 3' -OH group


Proteins of DNA ReplicationDNA exists in the nucleus as a condensed, compact structure. To prepare DNA for replication,a series of proteins aid in the unwinding and separation of the double-stranded DNA molecule. These proteins are required because DNA must be single-stranded before replication can proceed.
1. DNA Helicases - These proteins bind to the double stranded DNA and stimulate theseparation of the two strands.
2. DNA single-stranded binding proteins - These proteins bind to the DNA as a tetramer and stabilize the single-stranded structure that is generated by the action of the helicases. Replication is 100 times faster when these proteins are attached to the single- stranded DNA.
3. DNA Topoisomerase - This enzyme catalyzes the formation of negative supercoils that is thought to aid with the unwinding process.
In addition to these proteins, several other enzymes are involved in bacterial DNA replication.

4. DNA Polymerase - DNA Polymerase I (Pol I) was the first enzyme discovered withpolymerase activity, and it is the best characterized enzyme. Although this was the first enzyme to be discovered that had the required polymerase activities, it is not the primary enzyme involved with bacterial DNA replication. That enzyme is DNA Polymerase III (Pol III). Three activities are associated with DNA polymerase I;
* 5' to 3' elongation (polymerase activity) * 3' to 5' exonuclease (proof-reading activity) * 5' to 3' exonuclease (repair activity)
The second two activities of DNA Pol I are important for replication, but DNA Polymerase III(Pol III) is the enzyme that performs the 5'-3' polymerase function.
5. Primase - The requirement for a free 3' hydroxyl group is fulfilled by the RNA primers that are synthesized at the initiation sites by these enzymes.
6. DNA Ligase - Nicks occur in the developing molecule because the RNA primer is removedand synthesis proceeds in a discontinuous manner on the lagging strand. The final replicationproduct does not have any nicks because DNA ligase forms a covalent phosphodiester linkagebetween 3'-hydroxyl and 5'-phosphate groups.

A General Model for DNA Replication
1. The DNA molecule is unwound and prepared for synthesis by the action of DNA gyrase, DNAhelicase and the single-stranded DNA binding proteins.
2. A free 3'OH group is required for replication, but when the two chains separate no group of thatnature exists. RNA primers are synthesized, and the free 3'OH of the primer is used to begin replication.
3. The replication fork moves in one direction, but DNA replication only goes in the 5' to 3' direction.This paradox is resolved by the use of Okazaki fragments. These are short, discontinuous replicationproducts that are produced off the lagging strand. This is in comparison to the continuous strand that ismade off the leading strand.
4. The final product does not have RNA stretches in it. These are removed by the 5' to 3' exonucleaseaction of Polymerase I.
5. The final product does not have any gaps in the DNA that result from the removal of the RNA primer. These are filled in by the 5’ to 3’ polymerase action of DNA Polymerase I.
6. DNA polymerase does not have the ability to form the final bond. This is done by the enzyme DNA ligase.

RNA primed DNA replication

A General Model for DNA Replication
1. The DNA molecule is unwound and prepared for synthesis by the action of DNA gyrase, DNAhelicase and the single-stranded DNA binding proteins.
2. A free 3'OH group is required for replication, but when the two chains separate no group of thatnature exists. RNA primers are synthesized, and the free 3'OH of the primer is used to begin replication.
3. The replication fork moves in one direction, but DNA replication only goes in the 5' to 3' direction.This paradox is resolved by the use of Okazaki fragments. These are short, discontinuous replicationproducts that are produced off the lagging strand. This is in comparison to the continuous strand that ismade off the leading strand.
4. The final product does not have RNA stretches in it. These are removed by the 5' to 3' exonucleaseaction of Polymerase I.
5. The final product does not have any gaps in the DNA that result from the removal of the RNA primer. These are filled in by the 5’ to 3’ polymerase action of DNA Polymerase I.
6. DNA polymerase does not have the ability to form the final bond. This is done by the enzyme DNA ligase.


A General Model for DNA Replication
1. The DNA molecule is unwound and prepared for synthesis by the action of DNA gyrase, DNAhelicase and the single-stranded DNA binding proteins.
2. A free 3'OH group is required for replication, but when the two chains separate no group of thatnature exists. RNA primers are synthesized, and the free 3'OH of the primer is used to begin replication.
3. The replication fork moves in one direction, but DNA replication only goes in the 5' to 3' direction.This paradox is resolved by the use of Okazaki fragments. These are short, discontinuous replicationproducts that are produced off the lagging strand. This is in comparison to the continuous strand that ismade off the leading strand.
4. The final product does not have RNA stretches in it. These are removed by the 5' to 3' exonucleaseaction of Polymerase I.
5. The final product does not have any gaps in the DNA that result from the removal of the RNA primer. These are filled in by the 5’ to 3’ polymerase action of DNA Polymerase I.
6. DNA polymerase does not have the ability to form the final bond. This is done by the enzyme DNA ligase.

Removal of RNA primers and filling of gaps

A General Model for DNA Replication
1. The DNA molecule is unwound and prepared for synthesis by the action of DNA gyrase, DNAhelicase and the single-stranded DNA binding proteins.
2. A free 3'OH group is required for replication, but when the two chains separate no group of thatnature exists. RNA primers are synthesized, and the free 3'OH of the primer is used to begin replication.
3. The replication fork moves in one direction, but DNA replication only goes in the 5' to 3' direction.This paradox is resolved by the use of Okazaki fragments. These are short, discontinuous replicationproducts that are produced off the lagging strand. This is in comparison to the continuous strand that ismade off the leading strand.
4. The final product does not have RNA stretches in it. These are removed by the 5' to 3' exonucleaseaction of Polymerase I.
5. The final product does not have any gaps in the DNA that result from the removal of the RNA primer. These are filled in by the 5’ to 3’ polymerase action of DNA Polymerase I.
6. DNA polymerase does not have the ability to form the final bond. This is done by the enzyme DNA ligase.


ATP is an integral part of the ligation reaction
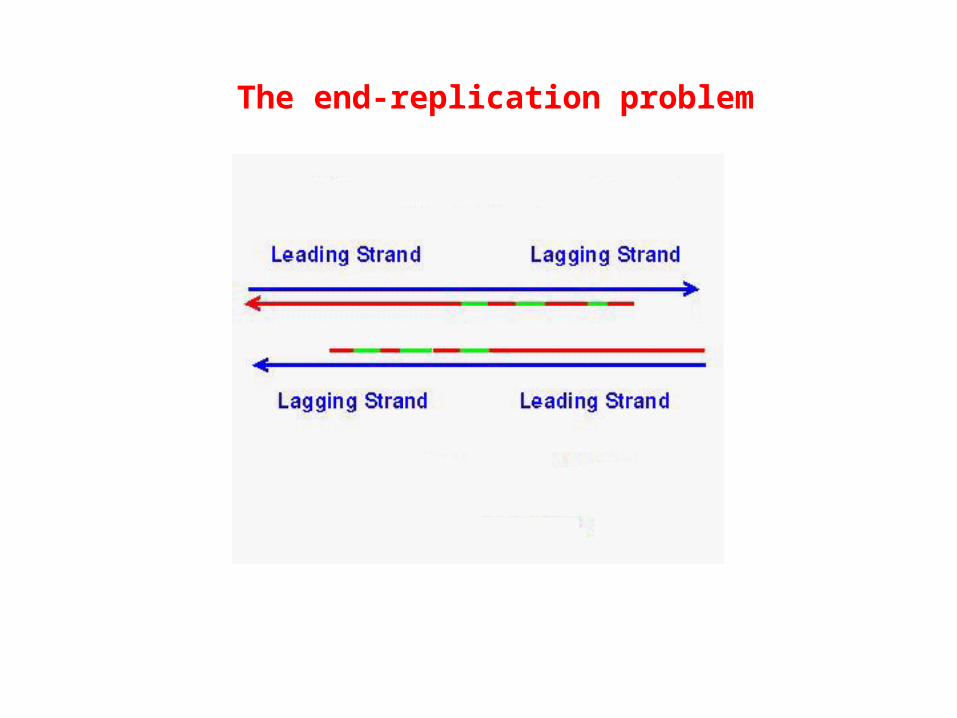
The end-replication problem


Group Organism Telomeric repeat (5' to 3' toward the end)
Vertebrates Human, mouse, Xenopus TTAGGG
Filamentousfungi Neurospora TTAGGG
Slime molds Physarum, Didymium TTAGGG Dictyostelium AG(1-8)
Ciliatedprotozoa Tetrahymena, Glaucoma TTGGGG
Paramecium TTGGG(T/G)Oxytricha, Stylonychia, TTTTGGGGEuplotes
Fission yeasts Schizosaccharomyces pombe TTAC(A)(C)G(1-8)
Budding yeasts Saccharomyces cerevisiae TGTGGGTGTGGTG
Telomeres contain arrays of DNA repeats

Telomerase isa reversetranscriptasetogether witha template RNA
It is active ingerm cells, notin somatic cells,and is activatedin cancers

Finishing school for telomeres

* The position on the DNA at which replication start points are found. (4 methods discussed)
* A DNA sequence that when added to a non-replicating DNA causes it to replicate.
* A DNA sequence whose mutation abolishes replication.
* A DNA sequence that in vitro is the binding target for enzyme complexes known to function in initiation of DNA replication.
Characteristics used to define Origins of Replication

1) In synchronously replicating cells, density transfer experiments can reveal which sequences replicate earliest. What is the direction of fork
movement here?
Time
% o
f p
rob
e in
HL
DN
A0
100
A B C
C B A
Genes on achromosome

EcoRIBamHI
BamHI EcoRI
2) Electron microscopy of bubbles after restrictionenzyme digestion


3) Hybridizing lagging strand nascent DNA to strand-specific fragmentsshows site of switch from leading to lagging strand synthesis
A B C DUpper strand fragments
Lower strand fragments
E F G H
Nascent DNA used to probe DNA fragments:
A B C D
E F G H

4) 2D Gel electrophoresis of replication intermediates followed byhybridization with a DNA fragment reveals whether a replication
bubble originates in the fragment
Dimension 1: separates by size; dimension 2: separates by shape

Sequence that when added to a non-replicating DNA causes it toreplicate; this approach was used to identify
Autonomously Replicating Sequences (ARS) in yeast.

Sequence that when mutated causes a replicating DNA tofail to replicate; this approach was used to identify
essential elements in an (ARS) in yeast
Yeast containing Leu2 on a plasmid
Plate yeast on Leucine minus plates to estimate rate of plasmid loss
Grow under non-selective conditions

Linker scanning mutations in yeast ARS1
% U
RA
+ c
olo
nie
sExamples of mutants
AB1B2B3WT-ARS1


OriC in E. coli chromosomal DNA

Initiation of DNA replication in E. coli

Two primosomes for E. coli chromosomal replication
Site of primosome assembly oriC, A-site n'-pas, D-loop, R-loop
Recognition DnaA protein PriA protein
Auxilliary proteins for (HU for oriC) PriBloading helicase PriC
DnaT
Helicase DnaB/DnaC
Priming Primase
DnaA and oriC independent replication initiation in E. colifrom hairpins, D-loops and R loops

SV40: a polyomavirus that is very useful forstudying eukaryotic replication enzymes

Early genes : proteins essential for viral DNA replication

T antigen binding sites cluster around the ori

* 98% nuclear: NLS * origin-specific DNA binding * DNA independent ATPase * ATP dependent DNA helicase * Binds to DNA polymerases, AP2, p53, Rb * Undergoes phosphorylation etc.
Properties of SV40 T antigen

COS cells (infected with SV40)(expresses T Ag)
+ plasmid + dNTP + rNTP+ 32PdATP
37o C, 60 min
Analyze products by electrophoresis and autoradiography
SV40 ori
SV40 - + ori - +
Li & KellySV40 DNA replicationIn vitro.PNAS (1984) 81:6973
What else could they doto validate their system?

Fractionation to homogeneity
COS cells (infected with SV40)(expresses T Ag)
T Antigen frombaculovirus expression system
Human HeLa or 293 cells
cell lysate
PC
0.2 M KCl 0.66 M KCl
QS
0.2 M KCl 0.4 M KCl (PCNA)
ssDNA
0.6 M KCl 1 M KClRPA
+
(RF-C, pol + pol )

Prokaryotic Function Eukaryotic
SSB ss DNA coating RPA
PRIMASE RNA primer synth. Primase subunitof DNA POL ,
5'-3' polymerase DNA POL ,DNA POL
POL III CORE 3'-5' exonuclease DNA POL 5'-3' exonuclease Fen1
COMPLEX ATP dependentclamploader
RF-C
CLAMP processivity factor PCNA
LIGASE seal nicks Lig1
DnaB Helicase ?
ELONGATION FACTORS ARE CONSERVED


Looping the lagging strand to make both polymerases move in the same direction

The discovery of DNA polymerase.Arthur Kornberg and Bob Lehman pursued an enzyme in bacterial extracts that would elongate a chain of deoxyribonucleic acid just like glycogen synthase elongates a chain of glycogen.
The enzymatic activity was unusual:
1) Needed a template which dictates what nucleotide was added: substrate was directing enzymatic activity2) Needed a primer annealed to the template.




Wait a minute!
John Cairns mutated the gene for DNA polymerase, polA, and the bacteria grew just fine!
Either the polymerase hypothesis was all wrong,…… or there were other DNA polymerases in E. colithat carried out DNA synthesis in the polA strains.
0.2M
0.4M
100
200
20 30 40
Fractions
polA- (Cairns)
I
III
II
100
200
600
3H T
hym
idin
e in
corp
ora
tio
n (
pm
ol)
0.2M
0.4M
Ph
osp
hat
e (M
)
polA+ (wild type)
+NEM
IIIII
I

Subunit
kDa Gene Subassembly
130 dnaE | 5'-3' polymerase 27.5 dnaQ
(mutD)| POL III CORE 5'-3'
exonuclease 10 | 3'-5'
exonuclease
71 dnaX
47.5 dnaX | 35 | ATP dependent' 33 | COMPLEX clamploader 15 | 12 |
40.6 dnaN CLAMP processivityfactor
DNA POLYMERASE III

Subunit
Gene Bacterial Function Eukaryotic
dnaE | 5'-3'polymerase
DNA POL
dnaQ(mutD)
| POL IIICORE
3'-5'exonuclease
DNA POL
| 5'-3'exonuclease
Fen1
dnaX
dnaX | | ATP' |
COMPLEXdependentclamploader
RF-C
| |
dnaN CLAMP processivityfactor
PCNA
CONSERVATION FROM PROKARYOTES TOEUKARYOTES

P POL dNTP
Challenge with vast excess of cold primer-template
Gel electrophoresis of products
Challenge - + - +
POL-X POL-Y
Which polymerase is processive?
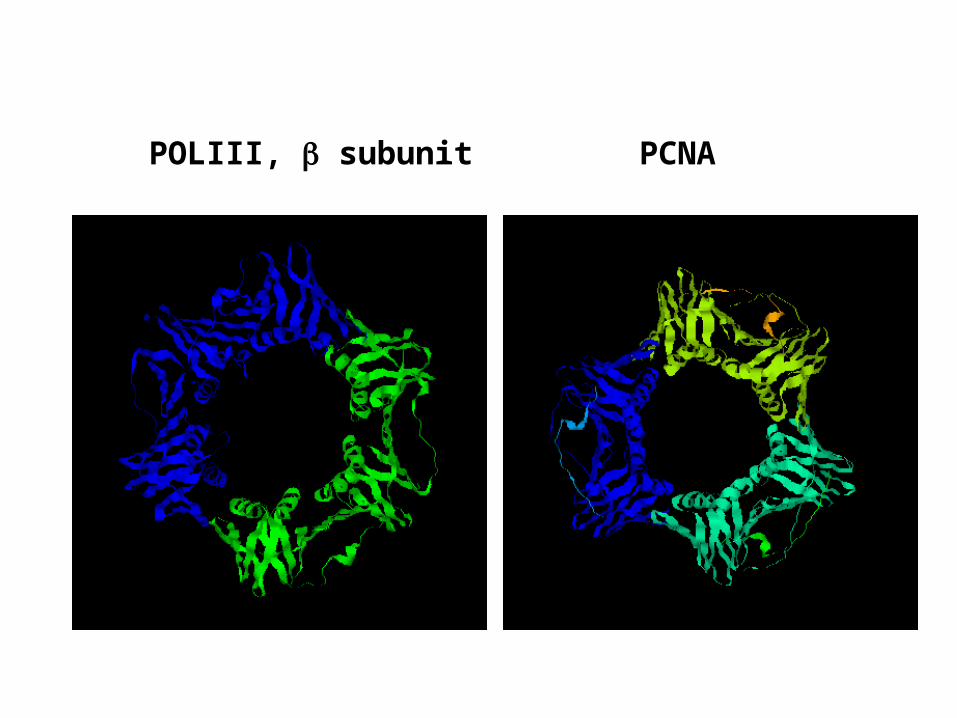
POLIII, subunit PCNA

ATP
ATP
3‘OH
ATP
ADP + PPi
Clamp
Clamp-loader
Clamp loaders hydrolyze ATP to load clamp
How does one provethat the clamp ringis opened duringloading?



Structure of a DNA polymerase (gp43 from phage RB69)
Side view:Polymerase active site
Top view withtemplate-primer:Polymerase siteAndproofreading site

* Topoisomerases II change the linking number in steps of 2 by passing both strands of double-stranded DNA through a break. * Eukaryotic topoisomerases isolated to date only relax supercoiled DNA, while prokaryotic topoisomerases (gyrases) can, given ATP, add supercoils.* TopoII releases catenated daughter molecules at the end of replication. Inhibitors like etoposide are used in chemotherapy.
Topoisomerases relax DNA by changing the DNA linking number

* Topoisomerases I change the linking number in steps of 1. They pass a single DNA strand through a nick.Topoisomerase I is a protein of the metaphase chromosome scaffold. * In interphase, topoisomerase is bound to the nuclear matrix. * The DNA replication machinery also appears bound to the matrix. * Inhibitor (camptothecin) also used in chemotherapy.

Topoisomerase action can be divided into three steps: nicking (1), strand passage (2); resealing (3).
5‘ end of DNA in gate segment is covalently linked to the OH oftyrosine in the active site of topo.

1
3
2
4
Cycle of topoisomerase activity inferred from structure
How would you test that the subunits have to open at the lower end to release the T segment?





RF-C is a five-subunit complexAll subunits are related in sequence and have ATP binding motifsATP hydrolysis by RF-C is associated with the loading of PCNARF-C is the functional homolog of the clamp-loader complex


Polymerase switching occurs even on lagging strands; pol does most of DNA synthesis


How do you think the PCNA is removed after ligation?


PCNA interacts with RF-C, pol , Fen1, DNA ligase,CAF1 and MCMT
Several of these have a common motifused in the interaction: Q-X-X-L/I/M-X-X-F-F/Y
p21/CIP1/WAF1, a protein induced by the tumor suppressor p53 uses the same motifto interact with PCNA
What effect is p21 expected to have on DNA replication/repair?

FEN1 interacts with PCNA
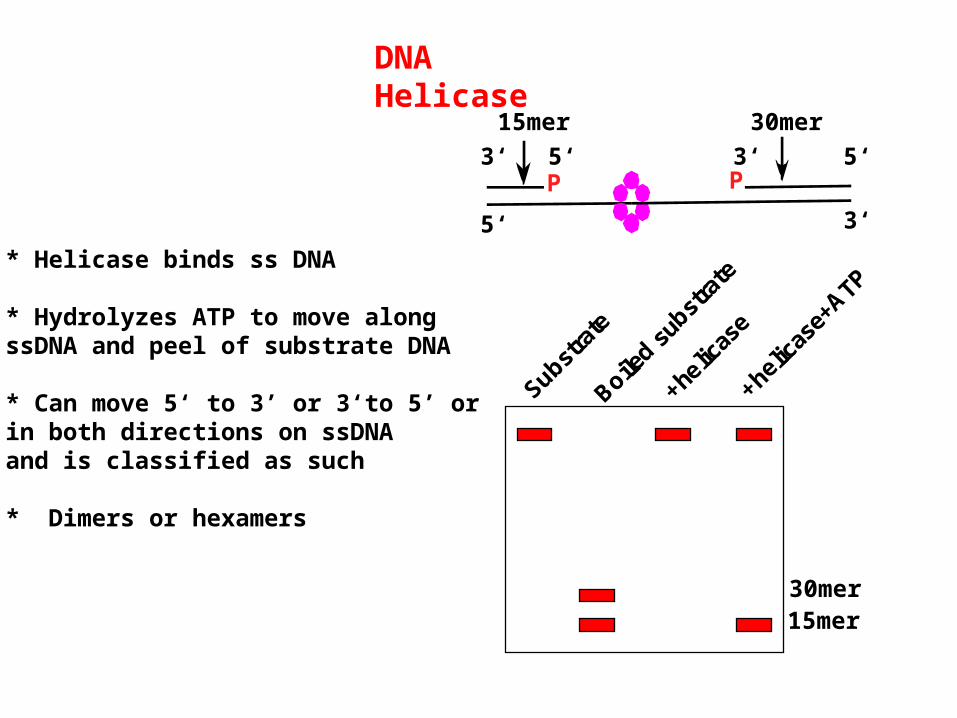
DNA Helicase
* Helicase binds ss DNA
* Hydrolyzes ATP to move alongssDNA and peel of substrate DNA
* Can move 5‘ to 3’ or 3‘to 5’ or in both directions on ssDNAand is classified as such
* Dimers or hexamers
P P
5‘ 3‘
3‘ 5‘ 5‘3‘
Substra
te
Boiled su
bstra
te
+he
licas
e
+helic
ase+
ATP
15mer 30mer
15mer30mer

Conserved helicase motifs:
Includes the following for ATPbinding and hydrolysisI = Walker A motif GxGxGKTII = Walker B motif DEad
Binds ATP
Tight conformation
Hydrolyzes ATP
Relaxed conformation
Helicases are ATP driven molecular motors
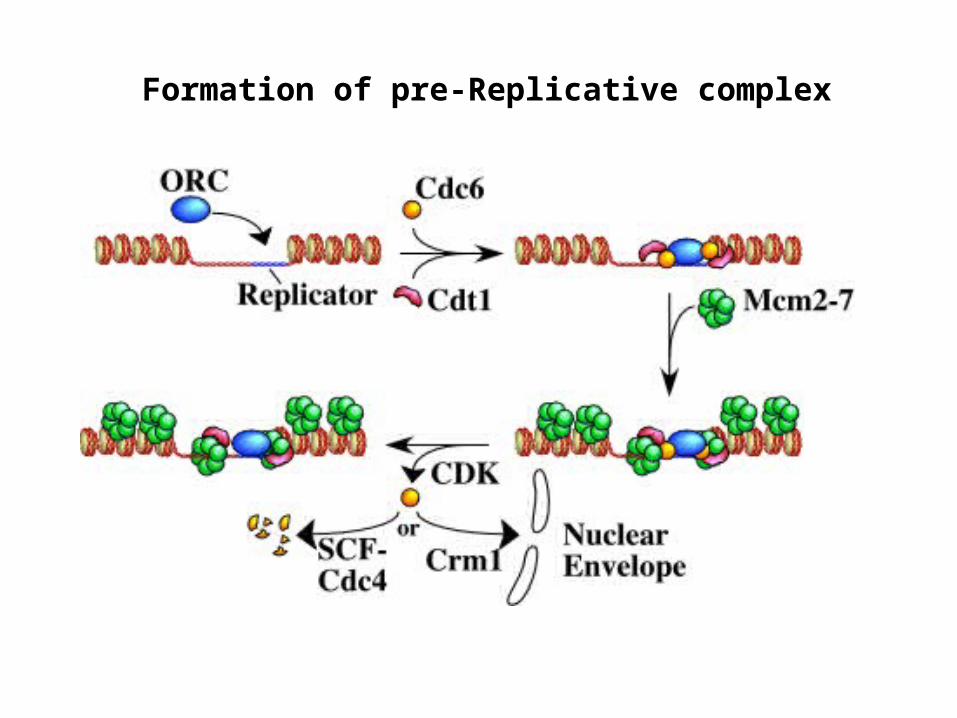
Formation of pre-Replicative complex
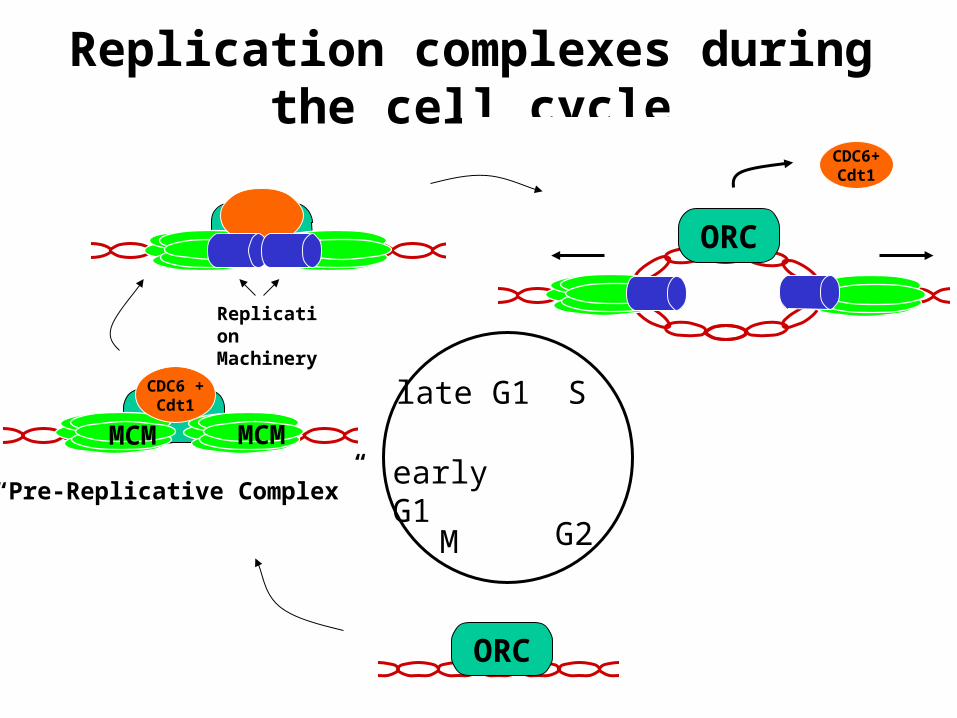
G2M
ORC
MCM MCM
CDC6 +Cdt1
Replication Machinery
“Pre-Replicative Complex”
Replication complexes during the cell cycle
ORC
CDC6+Cdt1
Slate G1
early G1

Eukaryotic initiation complex
ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein.Subunits are named according to their size, with ORC1 being the largest and ORC6 being the smallest subunit.Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
CDC6/Cdc18 : An essential factor for the assembly of the pre-replicative complexes that co-operates with Cdt1 to load MCM2-7Proteolyzed in yeasts or exported out of the nucleus in mammalian cells at the G1- S transition.Overexpression of Cdc6 in yeast causes multiple rounds of DNA replication without intervening mitosis, making it a critical regulator of DNA replication.
MCM2-7 : A family of six related proteins (MCM2-MCM7) which seem to function together in a large multi-subunit protein complex.The role of MCMís in DNA replication is still speculative, ranging from mediation of protein-DNA interactions to DNA helicase activity.

A/T DUEOREAuxiliaryelements
Auxiliaryelements
Origin recognitionproteins
TranscriptionFactors
TranscriptionFactors
Leading str
Leading str
Lagging str
Lagging str
Core origin
Origins are multi-partite

The Structure of ARS1(S. cerevisiae)
B1 ACSB2B3
ABF1
ORC
“Post-RC” protection
“Pre-RC” protection
13
456
4 1
22


ORC uses different strategies for binding DNAin different species

Pre-RC formation
ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein.Subunits are named according to their size, with ORC1 being the largest and ORC6 being the smallest subunit.Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
CDC6/Cdc18 : An essential factor for the assembly of the pre-replicative complexes that co-operates with Cdt1 to load MCM2-7Proteolyzed in yeasts or exported out of the nucleus in mammalian cells at the G1- S transition.Overexpression of Cdc6 in yeast causes multiple rounds of DNA replication without intervening mitosis, making it a critical regulator of DNA replication.
MCM2-7 : A family of six related proteins (MCM2-MCM7) which seem to function together in a large multi-subunit protein complex.The role of MCMís in DNA replication is still speculative, ranging from mediation of protein-DNA interactions to DNA helicase activity.

The Xenopus Cdc6 Protein Is Essential for the Initiation of a Single Round of DNA Replication in Cell-Free Extracts Made
from Xenopus egg extracts
Rxn Time:0-30 min30-60 min60-90 min
+ + +
+ + +
+ + +
+ + +
32P labeledDNAreplicationproduct
Replication initiation required ondouble-stranded DNA substrate
Only elongation required onsingle-stranded DNA substrate

Orc is required to load CDC6;CDC6 is required to load MCM

Immunostaining for CDC6 can pick outproliferating cells in Pap smears

Pre-RC formation
ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein.Subunits are named according to their size, with ORC1 being the largest and ORC6 being the smallest subunit.Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
CDC6/Cdc18 : An essential factor for the assembly of the pre-replicative complexes that co-operates with Cdt1 to load MCM2-7Proteolyzed in yeasts or exported out of the nucleus in mammalian cells at the G1- S transition.Overexpression of Cdc6 in yeast causes multiple rounds of DNA replication without intervening mitosis, making it a critical regulator of DNA replication.
MCM2-7 : A family of six related proteins (MCM2-MCM7) which seem to function together in a large multi-subunit protein complex.The role of MCMís in DNA replication is still speculative, ranging from mediation of protein-DNA interactions to DNA helicase activity.

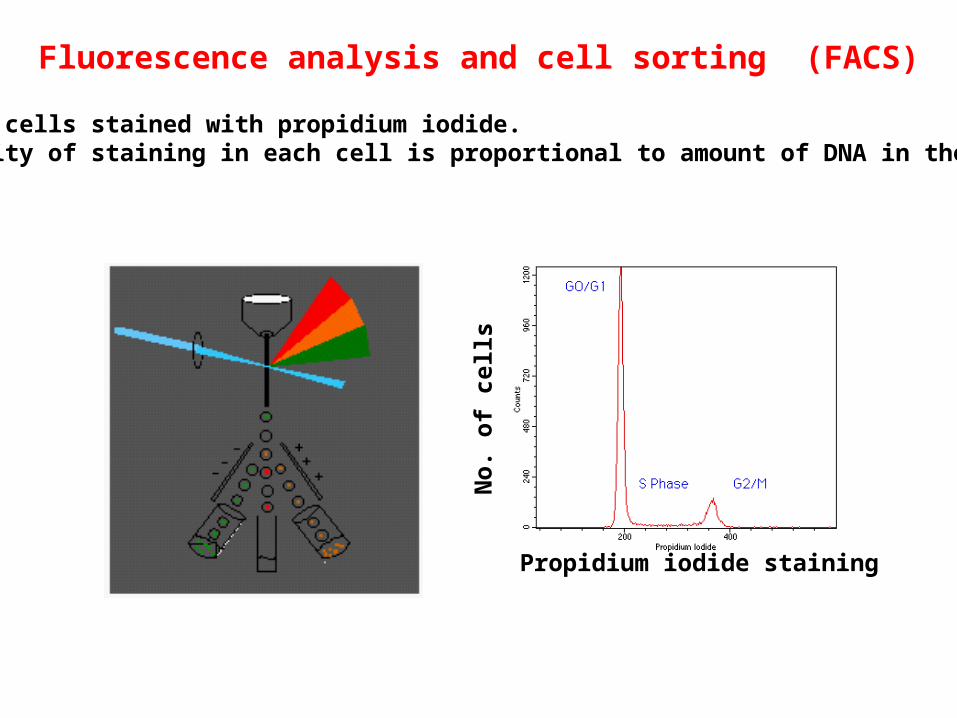
Propidium iodide staining
No
. o
f ce
lls
Fluorescence analysis and cell sorting (FACS)
DNA in cells stained with propidium iodide.Intensity of staining in each cell is proportional to amount of DNA in the cell.

Cross-link protein-DNA I.P. with anti MCM antibody
Reverse cross-link Purify DNA in I.P.
PCR to detect if your sequence was I.P.ed
MCM
anti-MCM Ab
Chromatinimmunoprecipitation
(ChIP) tests whether a given DNA sequence is bound by a protein
in vivo


MCM homolog fromM. thermoautotrophicum(an archaebacterium)has helicase activity.
What direction?
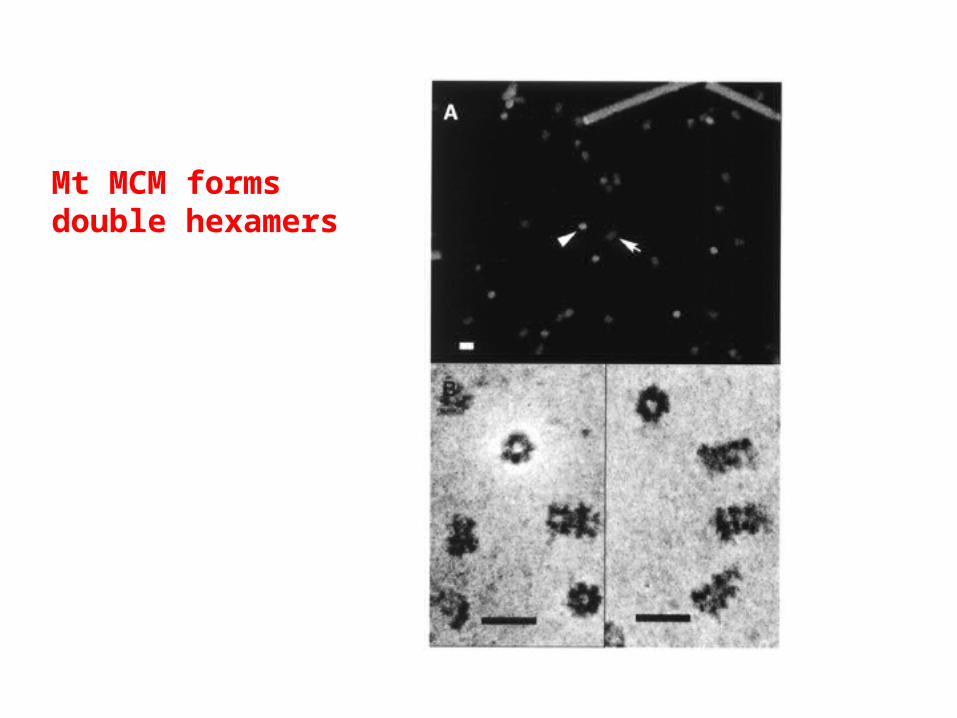
Mt MCM formsdouble hexamers

Pre-RC formation
ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein.Subunits are named according to their size, with ORC1 being the largest and ORC6 being the smallest subunit.Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.
CDC6/Cdc18 : An essential factor for the assembly of the pre-replicative complexes that co-operates with Cdt1 to load MCM2-7Proteolyzed in yeasts or exported out of the nucleus in mammalian cells at the G1- S transition.Overexpression of Cdc6 in yeast causes multiple rounds of DNA replication without intervening mitosis, making it a critical regulator of DNA replication.
MCM2-7 : A family of six related proteins (MCM2-MCM7) which seem to function together in a large multi-subunit protein complex.The role of MCMís in DNA replication is still speculative, ranging from mediation of protein-DNA interactions to DNA helicase activity.

Mammalian cells can survive with 0.1x the normal ORC2 level : WT vs cells

oriP
EB
NA
1
EBNA1 protein
Replication from an origin in Epstein-Barr virus:a plasmid expressing EBNA1 and containing oriP will
replicate and be maintained as an episome inmammalian cells
Hygromycin-Resistance
gene

Establishment of drug-resistant coloniesafter transfection of EBV-based plasmid

Transition to replication: from pre-RC to unwinding

MCM10 : essential function in the initiation of DNA replication in yeast. Required in Xenopus after the loading of Mcm2-7, before the loading of CDC45
CDC7/DBF4 : A DBF4 dependent kinase (DDK) which, together with Dbf4, is required for the initiation of DNA replication. Possible substrates of this kinase are the MCMs. How do we know this?
Cdk/Cyclin: A protein kinase essential for the initiation of DNA replication. Substrates that are activated by Cdk for replication initiation are unclear. MCM, CDC6 and some ORC subunits are phosphorylated, but this phosphorylation inactivates the proteins and prevents re-replication.
CDC45 : it seems to act together with ORC and the MCMs to initiate DNA replication.Has a role in the loading of DNA primase/polymerase complex.It and the MCM proteins appear to move along with the replication fork. How do we know this?
Transition to Replication
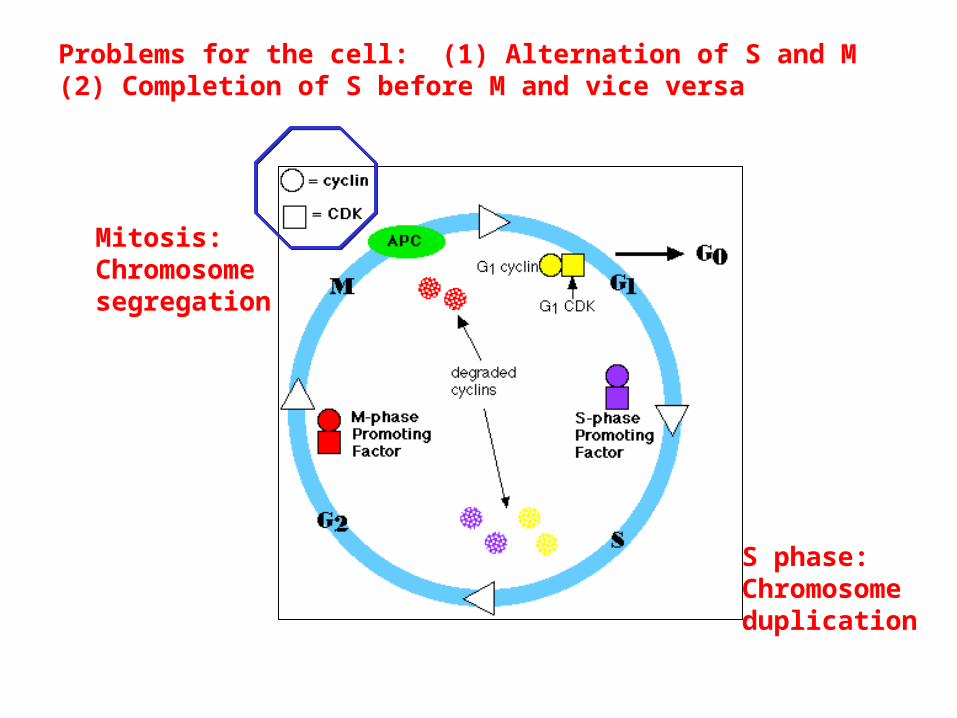
S phase:Chromosomeduplication
Mitosis:Chromosomesegregation
Problems for the cell: (1) Alternation of S and M(2) Completion of S before M and vice versa

With their multiple origins, how does the eukaryotic cell know which origins have been already replicated and which still await replication?
Two observations:When a cell in G1of the cell cycle is fused with a cell in S phase, the DNA of the G1 nucleus begins replicating early. Thus S phase cells have a positive factor for DNA replication that G1 cells lack.
When a cell in G2 of the cell cycle is fused with a cell in S phase, the DNA of the G2 nucleus does not begin replicating again even though replication is proceeding normally in the S-phase nucleus. Not until mitosis is completed, can freshly-synthesized DNA be replicated again. Thus cells in G2 have a negative factor that disappears in mitosis and/or lack a positive replication licensing factor until they pass through mitosis.
Control of Replication through the cell cycle

Cyclin/cdk protein kinase (i) yeast with temperature sensitive mutation in CDC28 (a cdk) arrest in G1(ii) addition of a cdk inhibitor (e.g. p21) to Xenopus egg extracts in G1 inhibits replication
DBF4/CDC7 protein kinase(i) ts mutation of CDC7 in yeast blocks cell cycle in G1 (by-passed by a mutation in MCM5)(ii) antibody mediated depletion in Xenopus egg extracts
Positive factor present in S phase cells

CDC6(i) degraded in yeasts after G1/S(ii) displaced out of the nucleus after G1/S in mammalian cells
MCM2-7(i) displaced out of the nucleus after G1/S in yeast(ii) loses tight association with chromatin after G1/S in mammalian cells
CDC45(i) loses tight association with chromatin after G1/S in mammalian cells
Cdt1(i) degraded in mammalian cells during S phase.(ii) inactivated by interaction with a protein called geminin.
ORC: may be inactivated by phosphorylationMcm10: ?
Positive factor (replication licensing factor) absent in G2 cells, but present in G1 cells

HsCDC6 G1 S
H2N COOHCy
S S
Wild-type
H2N COOHCy
S S
Cy
H2N COOHCy
D D
H2N COOHCy
A A
Ser Ala
Ser Asp
Proper Subcellular Localizati on of HsCDC6 is Dependent onBoth Intact Phosphorylation Sites and a Functional Cy Motif
CDC6 in mammalian cells is exported out of the nucleusIn S phase due to phosphorylation by cyclin/cdk
G1
S
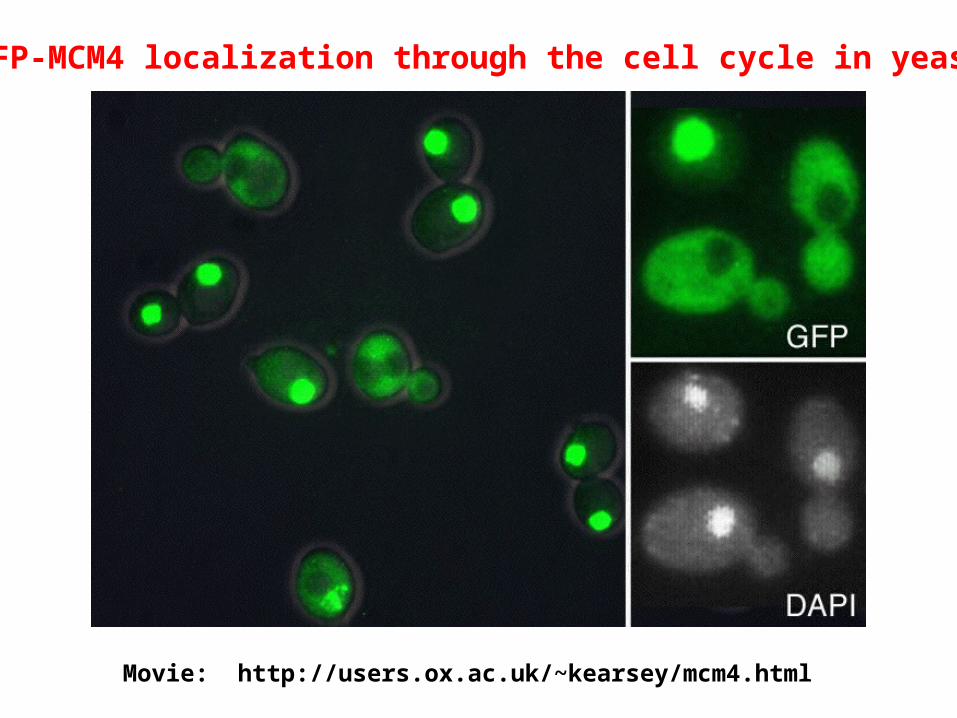
GFP-MCM4 localization through the cell cycle in yeast
Movie: http://users.ox.ac.uk/~kearsey/mcm4.html

Highly active cyclin/cdk protein kinase(i) inactivated in mitosis due to degradation of cyclins. (ii) artificial inactivation in G2 yeast with ts mutations in the cdk or by induction of a cdk inhibitor results in re-replication of DNA without mitosis.(iii) the nuclear export (in mammalian cells) or proteolysis (in yeast) of CDC6 is due to phosphorylation by cyclin/cdk. Therefore the negative factor present in G2 cells may also be responsible for the absence of a positive replication licensing factor in G2 nuclei!
Geminin(i) elevated in G2 cells and degraded in mitosis(ii) addition of a stable form of the protein to Xenopus egg extract inhibits the loading of MCM2-7 on chromatin and prevents DNA replication(iii) interacts with Cdt1.
Negative factor present in G2 cells, but absent in G1 cells

Low cyclin/cdk activity is required at G1/S to initiate DNA replicationLow cyclin/cdk activity in G1/S is inadequate to initiate mitosis
High cyclin/cdk activity in G2 inhibits DNA replicationHigh cyclin/cdk activity in G2 is required for mitosis
Solution to the alternation problem
Solution to the completion problem
For mitosis to be completed, cyclin-cdk has to be inactivated. Thus S phase cannotoccur before mitosis is completed.
Why doesn’t mitosis occur before S phase is completed? Checkpoint pathways thatare still being worked out.
