electron microscopy and restriction enzyme mapping reveal
TRANSCRIPT

Cell, Vol. 14, 629-639, July 1976, Copyright 0 1976 by MIT
Electron Microscopy and Restriction Enzyme Mapping Reveal Additional Intervening Sequences in the Chicken Ovalbumin Split Gene
A. C. Garapin, B. Cami, W. Roskam and P. Kourilsky Groupe de Biologie Moleculaire du Gene ERA, CNRS 201 lnstitut Pasteur Paris, France J. P. Le Pennec, F. Perrin, P. Gerlinger, M. Cachet and P. Chambon Laboratoire de Genetique Moleculaire des Eucaryotes du CNRS et U 44 de I’INSERM Faculte de Medecine Strasbourg, France
Summary
The Eco RI fragment “b” of chicken DNA (Breath- nach, Mandel and Chambon, 1977), which con- tains the sequences coding for the 5’ quarter of ovalbumin mRNA (ov mRNA), has been isolated by molecular cloning using a “shotgun” ap- proach. Electron microscopy and restriction en- zyme analysis have revealed that the sequences coding for the 5’ quarter (-500 nucleotides) of ov mRNA are split into four regions separated by three intervening sequences. The cloning proce- dure seems to be reliable, since the restriction enzyme pattern of the cloned Eco RI fragment “b” is similar to that of the corresponding chromo- somal DNA fragment. There is no evidence sup- potting the existence of a 150-200 nucleotide long sequence at the 5’ end of the ov mRNA similar to the “leader” sequences found at the 5’ end of some adenovirus and SV40 mRNAs.
Introduction
The discovery of mosaic adenovirus and SV40 mRNAs (Chambon, 1977; Chow et al., 1977; Dunn and Hassell, 1977; Klessig, 1977; Lewis, Anderson and Atkins, 1977; Sambrook, 1977) has acted as an illuminating stimulus for the groups engaged in the study of eucaryotic gene structure. Several reports have appeared during recent months that a given mRNA is encoded by separate blocks of DNA se- quences. Jeffreys and Flavell (1977) have found a 600 base pair (bp) DNA segment inserted some- where within the coding sequence for amino acid residues 101-120 of the 146 residue rabbit p-globin chain. A similar intervening sequence (for termi- nology, see Mandel et al., 1978) interrupts the mouse P-globin gene immediately following the codon corresponding to amino acid 104 (Leder et al., 1977; Tilghman et al., 1978). An intervening sequence of 1250 bp separating the variable and constant gene sequences was also found within the coding sequence of a cloned mouse plasmocy-
toma h light chain (Brack and Tonegawa, 1977). Rabbitts and Forster (1978) have also obtained evidence that the rearrangement of mouse immu- noglobulin light chain genes which occurs in mye- loma cells may not produce contiguous variable and constant genes in the DNA. A 93 bp insertion was detected and sequenced in the 5’ region of the protein coding sequence of a cloned immuno- globulin variable gene of mouse embryo (S. Tone- gawa and W. Gilbert, personal communication). We have shown that the ovalbumin gene is inter- rupted at least twice in the sequence which codes for chicken ovalbumin mRNA (Breathnach et al., 1977); similar results have been obtained by Doel et al. (1977) and by Weinstock et al. (1978). Inter- vening sequences are also found in genes which do not code for mRNA. The presence of insertions has been reported in Drosophila ribosomal DNA (Glover and Hogness, 1977; Pelligrini, Manning and Davidson, 1977; Wellauer and Dawid, 1977; White and Hogness, 1977), and short intervening sequences have been found in yeast tRNAs (Good- man, Olson and Hall, 1977; Valenzuela et al., 1978). It appears, therefore, that split genes characterized by intervening sequences interrupting structural sequences may have some generality in the eucar- yotic kingdom, and in this respect, such genes seem to differ fundamentally from those of the procaryotes.
To understand the structural and functional or- ganization of the ovalbumin gene at the molecular level, we have started to isolate restriction enzyme fragments of the ovalbumin gene by molecular cloning. In a previous report (Garapin et al., 1978), we described the cloning and structural analysis of the Eco RI fragment “a” of the split ovalbumin gene, which contains the sequence coding for the 3’ half of the ovalbumin mRNA (Breathnach et al., 1977). We report here the cloning of the 2.35 kb (kilobase pairs) Eco RI fragment “b” of the chicken chromosomal DNA, which contains the sequences coding for the 5’ quarter of the ov mRNA (Breath- nach et al., 1977). Restriction enzyme mapping and electron microscopy studies have revealed that the coding sequences present in this cloned fragment are interrupted by three intervening sequences which were not detected in previous studies of the chicken ovalbumin gene (Breathnach et al., 1977; Weinstock et al., 1978).
Results
Cloning of the Ovalbumin Gene Fragment “b” from Chicken DNA We have previously reported the cloning in a A vector of the Eco RI fragment “a” after a 20-50 fold enrichment by preparative gel electrophoresis of
Cell 630
digested chicken DNA (Garapin et al., 1978). In that experiment, 30,000 plaques were screened by in situ hybridization, and three independent recom- binants carrying fragment “a” were obtained with approximately the expected frequency, in agree- ment with our estimate that chicken DNA is di- gested into -300,000 different fragments by Eco RI. This suggested that fragment “a” could have been isolated from a total unfractionated Eco RI digest of chicken DNA, provided that a sufficient number of independent recombinants (3-10 x 105) were constructed and screened. In addition, such a “shotgun” experiment should lead to the isola- tion of other fragments such as the 2.35 kb Eco RI fragment “b”.
Chicken DNA was digested to completion by Eco RI (Breathnach et al., 1977) and recombined in vitro with the purified arms of hgtWES.hc (Enquist et al., 1976) in a 1 :l mass ratio. Recombinant DNA was packaged in vitro as described previously (Hohn and Murray, 1977; Garapin et al., 1978). From 95 pg of vector and 95 kg of cellular DNA, a total of -3 x lo6 independent recombinant phages were produced in four different experiments. Plate stocks were made to yield a total of -10’ phages. Approximately lo6 phages were screened on 80 plates (-12,000 plaques per plate) by in situ hybrid- ization (Garapin et al., 1978). Several positive plaques were detected and reisolated twice. Four phages capable of hybridization to the 32P-labeled double-stranded cDNA probe were obtained. Stocks were made, and phage DNA was extracted from the purified particles. A discussion of this “shotgun” experiment and the characterization of the phages obtained in this way, and not carrying the 2.35 kb Eco RI fragment, will be reported elsewhere (manuscript in preparation).
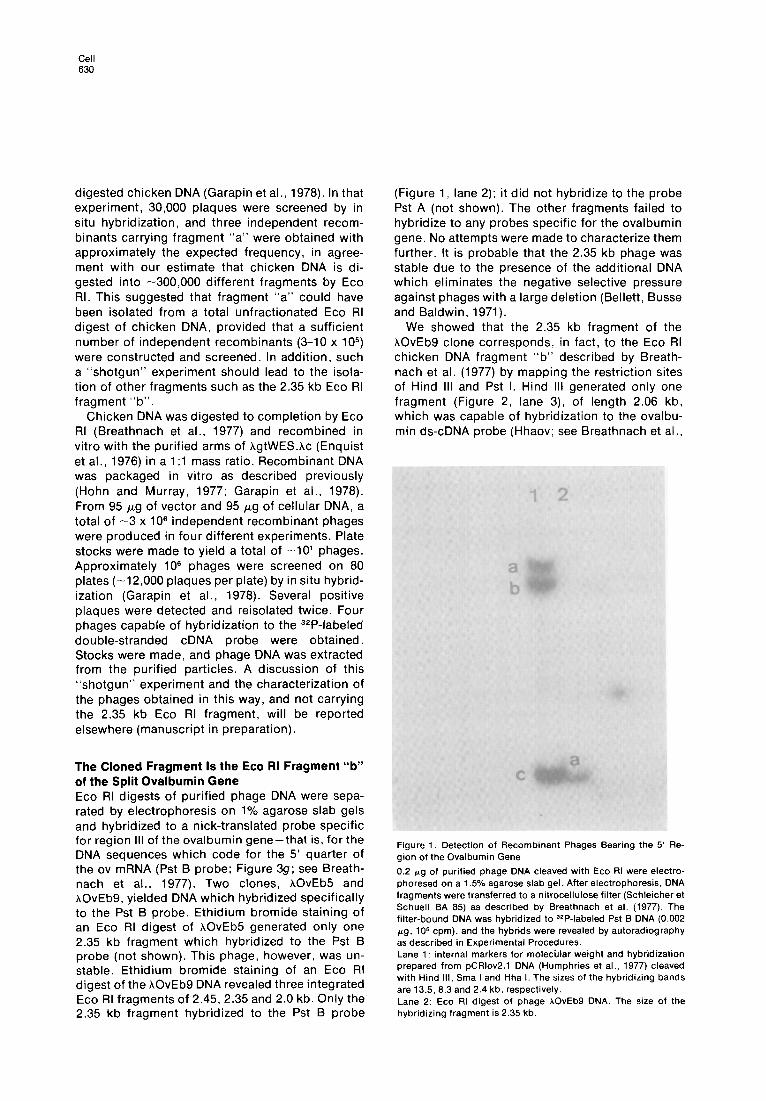
The Cloned Fragment Is the Eco RI Fragment “b” of the Split Ovalbumin Gene Eco RI digests of purified phage DNA were sepa- rated by electrophoresis on 1% agarose slab gels and hybridized to a nick-translated probe specific for region III of the ovalbumin gene-that is, for the DNA sequences which code for the 5’ quarter of the ov mRNA (Pst B probe; Figure 3g; see Breath- nach et al., 1977). Two clones, hOvEb5 and hOvEb9, yielded DNA which hybridized specifically to the Pst B probe. Ethidium bromide staining of an Eco RI digest of hOvEb5 generated only one 2.35 kb fragment which hybridized to the Pst B probe (not shown). This phage, however, was un- stable. Ethidium bromide staining of an Eco RI digest of the AOvEb9 DNA revealed three integrated Eco RI fragments of 2.45, 2.35 and 2.0 kb. Only the 2.35 kb fragment hybridized to the Pst B probe
(Figure 1, lane 2); it did not hybridize to the probe Pst A (not shown). The other fragments failed to hybridize to any probes specific for the ovalbumin gene. No attempts were made to characterize them further. It is probable that the 2.35 kb phage was stable due to the presence of the additional DNA which eliminates the negative selective pressure against phages with a large deletion (Bellett, Busse and Baldwin, 1971).
We showed that the 2.35 kb fragment of the AOvEb9 clone corresponds, in fact, to the Eco RI chicken DNA fragment “b” described by Breath- nach et al. (1977) by mapping the restriction sites of Hind III and Pst I. Hind III generated only one fragment (Figure 2, lane 3), of length 2.06 kb, which was capable of hybridization to the ovalbu- min ds-cDNA probe (Hhaov; see Breathnach et al.,
Figure 1. Detection of Recombinant Phages Bearing the 5’ Re- gion of the Ovalbumin Gene
0.2 pg of purified phage DNA cleaved with Eco RI were electro- phoresed on a 1.5% agarose slab gel. After electrophoresis. DNA fragments were transferred to a nitrocellulose filter (Schleicher et Schuell BA 65) as described by Breathnach et al. (1977). The filter-bound DNA was hybridized to J2P-labeled Pst B DNA (0.002 pg. lo5 cpm), and the hybrids were revealed by autoradiography as described in Experimental Procedures. Lane 1: internal markers for molecular weight and hybridization prepared from pCRlov2.1 DNA (Humphries et al., 1977) cleaved with Hind III, Sma I and Hha I. The sizes of the hybridizing bands are 13.5, 6.3 and 2.4 kb, respectively. Lane 2: Eco RI digest of phage hOvEb9 DNA. The size of the hybridizing fragment is 2.35 kb.

Intervening Sequences in the Ovalbumin Gene 631
1977). Pst I cuts the 2.35 kb fragment into two hybridizing fragments “a” and “b,” which are 1940 and 400 bp long, respectively (Figure 2, lane 7). Since the Pst I fragment “a” shows the strongest hybridization with the Hhaov probe, it must contain the bulk of the sequences coding for the 5’ region of the ov mRNA (Figure 3). Fragment “a” is not cut by Hind Ill, while fragment “b” is cut and generates a 130 bp hybridizing fragment “b,” which is too faint to be seen in Figure 2, lane 2, but is visible on the original autoradiograph. The Pst I and Hind III sites may thus be mapped within the Eco RI 2.35 kb fragment, as shown in Figure 3f. It is clear from
12, 3 4 56 7 8 81011
b
b
Figure 2. Mapping of Hha I, Pst I and Hind III Sites within the Cloned 2.35 kb Fragment
0.2 pg of an Eco RI digest of cloned AOvEb9 DNA were digested with Hha I, Pst I, Hind III or a combination of these enzymes, and the hybridizing fragments were revealed after electrophoresis by hybridization with a 32P-labeled Hhaov (0.01 fig, 5 X lo5 cpm) probe as described in the legend to Figure 1. The lower part of the figure is a 5 fold longer exposure than the upper part. Lanes 1, 5. 10 and 11: internal markers prepared from pCRlov2.1 DNA by cleavage with Hae Ill (lanes 1 and lo), Hinf I (lane 5) or Hha I (lane 11) restriction endonucleases. The sizes of the fragments hybridizing to the Hhaov probe are (in kb): lanes 1 and 10-(a) 1.41, (b) 0.72; lane 5-(a) 0.88, (b) 0.75, (c) 0.3 plus 0.27; lane 11 -2.4. Lanes 2, 3, 4, 6, 7 and 8: Hind IIVPst I, Hind III, Hind III/Hha I, Hha I/Pst I, Pst I and Hha I digests of the hOvEb9 DNA, digested with Eco RI (lane 9), respectively. The sizes of the hybridizing frag- ments are (in kb): lane 2-(a) 1.94, (b) 0.13; lane 3-2.06; lane 4- (a) 1.73; lane 8-(a) 1.8, (b) 0.4; lane 7-(a) 1.94. (b) 0.40; lane 8- 2.01 ; lane 9-2.35.
the position of these sites that they correspond to the Hind III and Pst I sites previously mapped in the cellular Eco RI fragment “b” (Breathnach et al., 1977; see Figure 6, c). The unique Pst I site in the 2.35 kb fragment generates two hybridizing frag- ments; this Pst I site should therefore correspond to the Pst I site present in the ovalbumin ds-cDNA (Figure 3g; Breathnach et al., 1977). Additional evidence supporting this conclusion is provided by the observation that a Pvu II site is very close to this Pst I site in both the cloned 2.35 kb fragment and the Hhaov ds-cDNA (not shown).
Electron Microscopic Evidence That the DNA Se- quence Coding for the 5’ Region of the Ovalbumin mRNA Is Split into Four Segments Heat-denatured hOvEb9 DNA was hybridized to ov mRNA at 54.5% in the presence of formamide under conditions where the formation of RNA-DNA hybrids is favored. The unexpected structure of the resulting hybrid molecules is shown in Figure 4a. A double-stranded region of -500 bp (defined by arrows 1 and 2) is joined to a nonhybridized mRNA tail (defined by arrows 2 and 3); three single- stranded loops (A, B and C) emerge from the hybrid region and must correspond to single-stranded DNA, since they are not removed by RNAase A treatment of the hybrid molecule (not shown).
Measurements carried out on 46 hybrid mole- cules gave lengths of 549 +- 40 bp and 1215 5 50 nucleotides for the hybridized and nonhybridized portions of the ov mRNA, respectively, the sum accounting within the limits of experimental error for the total length of ov mRNA (Rosen et al., 1976; Garapin et al., 1978). The fact that it is the 5’ region of ov mRNA which is contained within the hybrid regions was established as follows. The 2.35 kb Eco RI fragment of clone AOvEb9 which codes for the 5’ quarter of ov mRNA (see above), and the Eco RI-Hind III 2.6 kb fragment of clone hOvEa which codes for the 3’ half of ov mRNA (Garapin et al., 1978), were purified and hybridized to ov mRNA. Two RNA-DNA hybrid regions are seen per RNA molecule in Figure 4 c at each end of the ov mRNA molecule. One such region, 1080 ? 50 bp long, corresponds to a hybrid between the 2.6 kb frag- ment and the 3’ half of ov mRNA (Garapin et al., 1978). The other hybrid region presents the three characteristic loops (A, B and C) clearly obtained with cloned hOvEb9 DNA, and therefore corre- sponds to a hybrid between the 2.35 kb fragment and the 5’ region of ov mRNA (Figure 4~). The two hybrid regions are separated by -400 nucleotides of ov mRNA which are coded for by region II of the ovalbumin gene (Breathnach et al., 1977), which is not contained in either of these two cloned frag-

Cell 632
(a) 590 f 90 142 + 26 350 2 50 124 2 30 360 f; 60 bp
2350 bp a b
(b) Pst I I I I
(c) Hae III I b c a
I I I
(d) Hinf I 1 b a
------------_- I
c b (e) Mbo II I ,--_____ I _________ Id -----____ I
a I I
(f)
(9)
Xbal Eco RI
I3
;i’l, Hae 1 Sst 2 , Hiyf I Mbo 2 MboS- I
Mbo5 Mbo4
0
5’ 5’ 3’ 1 I . Sst I I 6St;:P”” II
TRANSCRIPTION TRAk’CP,2llr Mbo II
2350 bp
160 bp
I I Pst B PROBE
Figure 3. Localization of the Restriction Enzyme Sites on the Cloned 2.35 kb DNA Fragment and Correlation with a Schematic Representation of the Electron Microscopy Results and the Ovalbumin ds-cDNA Restriction Map
(a) Schematic representation of the electron microscopy results. The heavy lines a, b, c and d represent RNA-DNA hybrid regions. A, B and C correspond to the intervening sequences. (b) Pst I fragments of the 2.35 kb DNA: (a) 1.94, (b) 0.4 kb. (c) Hae Ill fragments of the 2.35 kb DNA: (a) 1.45. (b) 0.65, (c) 0.24 kb. (d) Hinf I fragments of the 2.35 kb DNA: (a) 1.32, (b) 0.75 kb. (e) Mbo II fragments of the 2.35 kb DNA: (a) 0.7, (b) 0.6, (c) 0.4, (d) 0.1 kb. (f) Localization of restriction endonuclease sites on the 2.35 kb fragment: a, b, c and d represent the hybridizing regions which can be correlated with corresponding regions of the ovalbumin ds-cDNA [see @)I. @) Restriction endonuclease map of ovalbumin ds-cDNA. Only the relevant sites are given. The locations of the restriction sites are given from the 3’ end of the ovalbumin mRNA (Breathnach et al., 1977). The thin lines link homologous sites in the cloned 2.35 kb fragment (f) and the ov ds-cDNA (g). The dashed lines in (d) and (e) represent fragments which are not revealed by hybridization to the 32P-labeled Hhaov probe.
Figure 4. Electron Micrography of the Hybrid Molecules between Cloned DNA and ov mRNA
(a) Hybrid between hOvEb9 DNA and ovalbumin mRNA. The hybrid region (549 f 40 bp) is defined by arrows 1 and 2. Arrows 2 and 3 define the ends of a single-stranded mRNA tail of 1215 + 50 nucleotides. The single-stranded DNA loops A, Band C define four double-stranded regions a, b. c and d (see text). (b) Hybrid between the cloned purified 2.35 kb Eco RI fragment DNA and ovalbumin mRNA. The 2.35 kb fragment extends between arrows I and II. The extent of the RNA-DNA hybrid regions is indicated by arrows 1 and 2. and the single-stranded RNA tail is defined by arrows 2 and 3. The lengths of the three loops of single-stranded DNA, A, B and C, are 194 f 45, 596 r 90 and 350 + 50 nucleotides, respectively. The lengths of the four RNA-DNA hybrid regions a, b. c and d are 221 ? 50, 70 2 25, 142 r 26 and 124 2 30 bp, respectively. Single- stranded regions lying between arrows I and 1, and 2 and II, are 297 2 50 and 360 & 60 nucleotides, respectively. Hybrids were prepared as described in Experimental Procedures. (c) Hybrid between ovalbumin mRNA, the cloned 2.35 kb Eco RI fragment “b,” and the 2.6 kb Eco RI-Hind III fragment of Eco RI fragment “a.” which codes for the 3’ half of ov mRNA (Garapin et al., 1976). The extents of the 2.35 kb Eco RI fragment and the Eco RI-Hind Ill fragment are given by arrows I and II, and IV and V, respectively. The 5’ and 3’ ends of the ov mRNA molecule are defined by arrows 1 and 5. respectively. The region of hybridization between the mRNA and the 2.35 kb Eco RI fragment extends between arrows 1 and 2 (549 ? 40 bp). while the hybridization region between the mRNA and the Eco RI-Hind III fragment lies between arrows 4 and 5 (1060 ? 50 bp). The single-stranded RNA fragment of -400 nucleotides which lies between arrows 2 and 4 is coded for by region II of the ovalbumin gene, a region not contained in either of the two cloned fragments. A, B and C. and a, b. c and d refer to features defined in the legend to (b). (d) DNA heteroduplex between the cloned 2.35 kb Eco RI fragment and Hhaov. Arrows I and II define the extent of the 2.35 kb fragment. The end of the Hhaov DNA region which is not hybridized is shown by arrow Ill. The lengths of the three single-stranded loops A, B and C are 236 f 45,620 ? 90 and 423 * 100 nucleotides, respectively. The lengths of the duplex regions a, b. c and d are 140 + 50,70 2 22,120 ? 36 and 116 ? 36 bp, respectively. Hybrid molecules were prepared as described in Experimental Procedures. Bars = 0.1 p.

Intervening Sequences in the Ovalbumin Gene 633

Cell 634
ments. The relative positions on the mRNA of the hybrid regions with the 2.6 and 2.35 kb fragments show that the loop structures must lie in the 5’ quarter of ov mRNA.
From the above results, we conclude that three intervening sequences are present in the cloned ovalbumin gene Eco RI fragment “b,” which con- tains the sequence coding for the 5’ region of ov mRNA. The three loops A, B and C define four segments of the 2.35 kb fragment (a, b, c and d, from the 5’ end of ov mRNA) which are comple- mentary to ov mRNA (Figure 4b). From measure- ments carried out on 50 molecules, the lengths of segments a, b, c and d were 221 r 50,70 lr 25,142 & 26 and 124 2 30 bp, respectively, while the lengths of loops A, B and C were 194 ? 45, 590 & 90 and 350 t 50 nucleotides, respectively. The DNA region coding for the 5’ end of the ov mRNA is located 297 ? 50 nucleotides from the closer Eco RI end of the 2.35 kb fragment, whereas the 3’ end of the ov mRNA coding segment “d” is located 360 ? 60 nucleotides from the other end (see Figure 3a).
When heteroduplexes were formed between the cloned 2.35 kb fragment and the Hhaov ds-cDNA, the same three loop structure was obtained (Figure 4d). Length measurements gave 238 5 45, 620 -c 90 and 423 + 100 nucleotides for the three loops A, B and C, and 140 ? 50,70 2 22,120 2 36 and 118 f 36 bp for the DNA-DNA hybrid regions a, b, c and d, respectively, in good agreement with the above values. The DNA-DNA hybrid region “a” (140 ? 50 bp) was shorter by 60-100 bp than the corre- sponding RNA-DNA hybrid region “a,” but had no tail at the 5 hybridizing end of the Hhaov. The latter result indicates that the DNA sequence corre- sponding to the extreme 5’ end (-80 nucleotides) of ov mRNA is not present in the Hhaov ds-cDNA, and also that the corresponding Hha I site belongs to ovalbumin ds-cDNA and not to the vector plas- mid pCR1, as was previously thought (Humphries et al., 1977).
Digestion of the 2.35 kb fragment with Hha I yields a unique fragment 2010 bp which hybridizes with the Hhaov probe (Figure 2, lane 8). This fragment can be further cut by Pst I to give two hybridizing fragments “a” and “b” of 1600 and 400 bp, respectively (Figure 2, lane 6), or by Hind III to give one hybridizing fragment of 1730 bp (Figure 2, lane 4, a). These observations allow unambiguous mapping of the Hha I site.
The positions of two Hinf I sites, Hinf 1 and 2, were deduced from the following results. Hinf I digestion yields two hybridizing fragments “a” and “b” of 1320 and 750 bp (Figure 5, lane 3). The 1320 bp fragment can be further cut with Pst I to yield the hybridizing fragments “c” and “d” (Figure 5, lane 4), which are 920 and 400 bp long, respec- tively. In addition, the Hinf I “b” fragment of 750 bp
The Presence of Three Intervening Sequences in the Cloned 2.35 kb DNA Fragment Is Confirmed by Restriction Enzyme Mapping The existence of the three intervening sequences visualized by electron microscopy was supported by comparing the restriction enzyme map of the 2.35 kb fragment with that of the corresponding regions of the ovalbumin Hhaov ds-cDNA (Figure 39). The restriction enzyme map of the 2.35 kb fragment, shown in Figure 3f, was established as follows, using the blotting technique and 32P- Hhaov ds-cDNA as a probe for ov mRNA coding sequences.
Figure 5. Mapping of Hae III and Hinf I Sites within the Cloned 2.35 kb Eco RI Fragment
0.2 rg of an Eco RI digest of cloned hOvEb9 DNA were digested with Hae Ill and Hinf I alone, or in combination with Hha I or Pst I, and processed as described in the legend to Figure 1. The ovalbumin-containing fragments were visualized by autoradiog- raphy as described in the legend to Figure 2. Lanes 1, 5 and 9: Hae Ill (lanes 1 and 9) and Hinf I (lane 5) internal markers prepared as described in the legend to Figure 2. Lanes 2, 3, 4, 6, 7 and 6: Hinf I/Hha I, Hinf I, Hinf IlPst I, Hae Ill/ Hha I, Hae III and Hae IlllPst I digests, respectively, of the 2.35 kb Eco RI hOvEb9 DNA fragment. The sizes of the hybridizing fragments are (in kb): lane 2-(a) 1.32, (e) 0.42; lane 3-(a) 1.32. (b) 0.75; lane 4-(c) 0.92, (b) 0.75, (d) 0.4; lane 6-(a) 1.46, (f) 0.26, (c) 0.24; lane 7-(a) 1.46, (b) 0.65, (c) 0.24; lane 6-(d) 1.05. (b) 0.65, (e) 0.4, (c) 0.24.
Intervening Sequences in the Ovalbumin Gene 635
is converted by Hha I to a 420 bp hybridizing fragment “e” (Figure 5, lane 2). The results localize one Hinf I site (Hinf 2) at 420 bp to the right of the Hha I site, and the other (Hinf 1) at 920 bp to the left of the Pst I site (Figure 3f). No other Hinf I sites are present in the 2.35 kb fragment, as determined from partial digestion studies (not shown).
The following results unambiguously localize two Hae III sites, Hae 1 and 2. Hae III digestion yields three hybridizing fragments “a,” “b” and “c” (Figure 5, lane 7) of 1460, 650 and 240 bp, respectively. Two fragments “d” and “e” of 1050 and 400 bp are obtained from the 1460 bp Hae III fragment “a” by further digestion with Pst I (Figure 5, lane 8). Hha I cuts the 650 bp Hae III fragment “b” (Figure 5, lane 7) to yield the 280 bp hybridizing fragment “f” (Figure 5, lane 6).
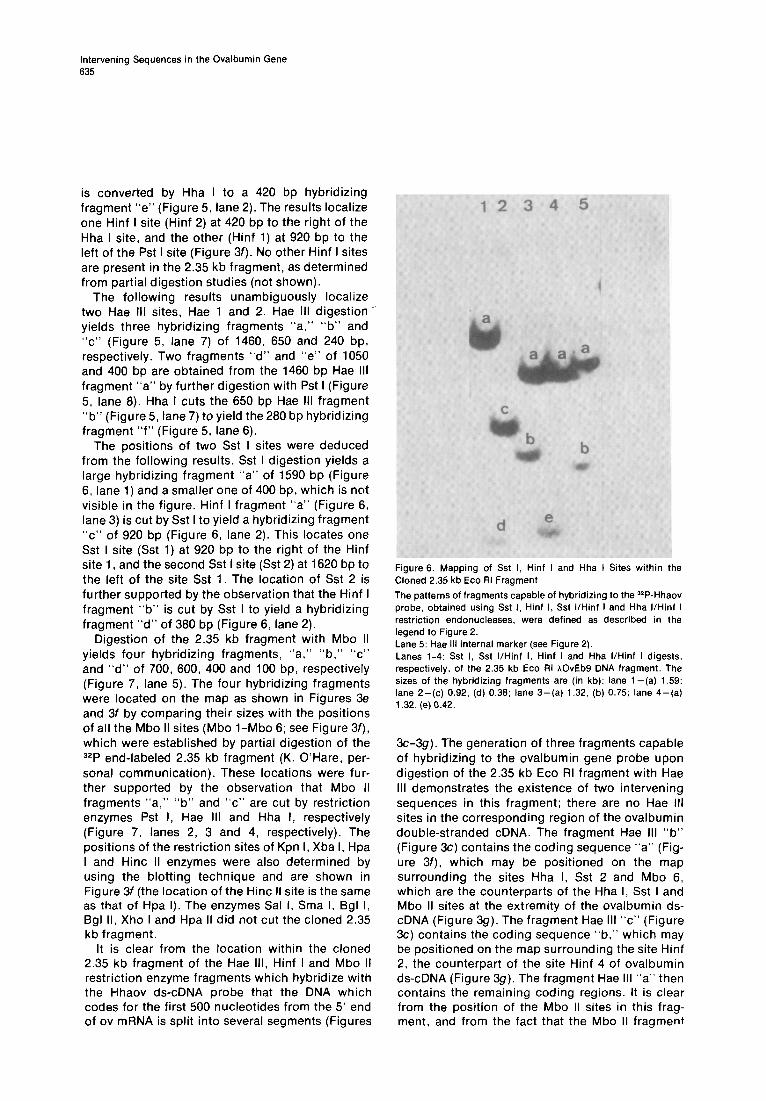
The positions of two Sst I sites were deduced from the following results. Sst I digestion yields a large hybridizing fragment “a” of 1590 bp (Figure 6, lane 1) and a smaller one of 400 bp, which is not visible in the figure. Hinf I fragment “a” (Figure 6, lane 3) is cut by Sst I to yield a hybridizing fragment “c” of 920 bp (Figure 6, lane 2). This locates one Sst I site (Sst 1) at 920 bp to the right of the Hinf site 1, and the second Sst I site (Sst 2) at 1620 bp to the left of the site Sst 1. The location of Sst 2 is further supported by the observation that the Hinf I fragment “b” is cut by Sst I to yield a hybridizing fragment “d” of 380 bp (Figure 6, lane 2).
Digestion of the 2.35 kb fragment with Mbo II yields four hybridizing fragments, “a,” “b,” “c” and “d” of 700, 600, 400 and 100 bp, respectively (Figure 7, lane 5). The four hybridizing fragments were located on the map as shown in Figures 3e and 3f by comparing their sizes with the positions of all the Mbo II sites (Mbo l-Mb0 6; see Figure 3f), which were established by partial digestion of the 32P end-labeled 2.35 kb fragment (K. O’Hare, per- sonal communication). These locations were fur- ther supported by the observation that Mbo II fragments “a, ” “b” and “c” are cut by restriction enzymes Pst I, Hae III and Hha I, respectively (Figure 7, lanes 2, 3 and 4, respectively). The positions of the restriction sites of Kpn I, Xba I, Hpa I and Hint II enzymes were also determined by using the blotting technique and are shown in Figure 3f (the location of the Hint II site is the same as that of Hpa I). The enzymes Sal I, Sma I, Bgl I, Bgl II, Xho I and Hpa II did not cut the cloned 2.35 kb fragment.
It is clear from the location within the cloned 2.35 kb fragment of the Hae III, Hinf I and Mbo II restriction enzyme fragments which hybridize with the Hhaov ds-cDNA probe that the DNA which codes for the first 500 nucleotides from the 5’ end of ov mRNA is split into several segments (Figures
Figure 6. Mapping of Sst I, Hinf I and Hha I Sites within the Cloned 2.35 kb Eco RI Fragment
The patterns of fragments capable of hybridizing to the 32P-Hhaov probe, obtained using Sst I. Hinf I, Sst I/Hinf I and Hha I/Hinf I restriction endonucleases, were defined as described in the legend to Figure 2. Lane 5: Hae Ill internal marker (see Figure 2). Lanes l-4: Sst I, Sst IlHinf I, Hinf I and Hha I/Hinf I digests, respectively, of the 2.35 kb Eco RI hOvEb9 DNA fragment. The sizes of the hybridizing fragments are (in kb): lane 1 -(a) 1.59; lane 2-(c) 0.92, (d) 0.36; lane 3-(a) 1.32. (b) 0.75; lane 4-(a) 1.32, (e) 0.42.
3c-3g). The generation of three fragments capable of hybridizing to the ovalbumin gene probe upon digestion of the 2.35 kb Eco RI fragment with Hae III demonstrates the existence of two intervening sequences in this fragment; there are no Hae III sites in the corresponding region of the ovalbumin double-stranded cDNA. The fragment Hae III “b” (Figure 3c) contains the coding sequence “a” (Fig- ure 3f), which may be positioned on the map surrounding the sites Hha I, Sst 2 and Mbo 6, which are the counterparts of the Hha I, Sst I and Mbo II sites at the extremity of the ovalbumin ds- cDNA (Figure 3s). The fragment Hae III “c” (Figure 3c) contains the coding sequence “b,” which may be positioned on the map surrounding the site Hinf 2, the counterpart of the site Hinf 4 of ovalbumin ds-cDNA (Figure 3s). The fragment Hae III “a” then contains the remaining coding regions. It is clear from the position of the Mbo II sites in this frag- ment, and from the fact that the Mbo II fragment

Cell 636
sults obtained by restriction mapping and electron microscopy (compare Figures 3a and 3f).
Figure 7. Analysis of Fragments Capable of Hybridizing to the 32P-Hhaov Probe, Produced by Digestion with Mbo II of the Cloned DNA
An EC0 RI digest of cloned hOvEb9 DNA (0.2 fig) was further digested with Mbo II, either alone or in combination with Hha I, Pst I or Hae III. The samples were electrophoresed on 2% agarose gels, and the hybridizing fragments were detected as described in the legend to Figure 2. The lower part of the figure (band “d”) corresponds to a 40 fold longer exposure than the upper part. Lanes 1 and 6: Hinf I and Hae Ill internal markers, respectively (see Figure 2). Lanes 2, 3, 4 and 5: Mbo II/Pst I, Mbo II/Hae Ill, Mbo WHha I and Mbo II digests, respectively, of the cloned 2.35 kb Eco RI frag- ment. The sizes of the hybridizing fragments are (in bp): lane 2- (b) 600, (c) 400, (e) 290, (d) 100; lane 3-(a) 700, (c) 400, (f) 245, (d) 100; lane 4-(a) 700, (b) 600, (d) 100; lane 5-(a) 700, (b) 600, (c) 400, (d) 100.
“d” (Figure 3e) hybridizes to ovalbumin ds-cDNA (see above), that the coding regions which it con- tains must be in two parts, a region “c” (Figure 3f), containing the sites Mbo 2 and Mbo 3 [counterparts of the two Mbo II sites located at positions 1460 and 1560 in the ds-cDNA (Figure Z&I)], separated from a region “d” (positioned around the Pst I, Pvu II and Sst 1 sites, also present in the ds-cDNA) by an intervening sequence containing the Mbo II site 1 (Figures 3f and 3s).
From the above results, we conclude that the DNA region III which codes for the 5’ region of ov mRNA (Breathnach et al., 1977) contains three intervening sequences which define four messen- ger-coding segments, located as indicated in Fig- ure 3f. There is excellent agreement between re-
Evidence That the Cloned Ovalbumin 2.35 kb Fragment “b” Is a Faithful Copy of the Corre- sponding Chromosomal DNA Fragment Since recombinations, deletions or translocations could have occurred during the DNA cloning proc- ess, we compared the restriction enzyme pattern obtained by digestion of the 2.35 kb cloned frag- ment “b” with that of the Eco RI fragment “b” purified by reverse-phase chromatography (RPC5) from cellular DNA (Mandel et al., 1978).
The Hinf I digest shows two hybridizing frag- ments “a” and “b” (1.32 and 0.75 kb, respectively) both with the cellular and the cloned 2.35 kb DNA fragments (Figure 8, lanes 9 and 3). The digestion of the cellular DNA was incomplete, and fragment b’ (1.03 kb) is a partial digest. Mbo II digestion generates the three hybridizing fragments “a,” “b” and “c” of 0.7, 0.6 and 0.4 kb on both cloned and chromosomal DNAs (Figure 8, lanes 5 and 11, for the cloned and cellular DNAs, respectively). The 0.1 kb Mbo II fragment “d,” which was seen in the digest of the cloned DNA in Figure 7, lane 5, could not be detected with the amount of DNA used in these experiments. Hae III digestion of the cloned 2.35 kb fragment gives two hybridizing fragments “a” and “b” of 1.46 and 0.65 kb, respectively (Figure 8, lane 4), in agreement with the results presented in Figure 5, lane 7. The small hybridizing fragment Hae “c” of 0.24 kb (Figure 5, lane 7) is not visible for the reason given above for the Mbo II fragment “d.” The cellular DNA, when cut by Hae III, shows two hybridizing bands “a” and “b” of 1.7 and 0.65 kb, respectively (Figure 8, lane 10). The size of fragment “a,” 1.7 kb, is the same as the sum of the sizes of Hae III fragments “a” and “c” derived from the cloned 2.35 kb fragment. Using increased amounts of Hae III restriction enzyme did not result in any further cleavage of the cellular DNA. This observation suggests that the Hae 1 site (Figures 3c and 3f) is either blocked or missing.
Sst I generates only one hybridizing fragment of 1.59 kb on both DNAs (Figure 8, lanes 6 and 12 for cloned and chromosomal DNA, respectively). Since the sizes of the hybridizing fragments generated by Sst I, Hinf I and Mbo II on the cellular DNA and on the cloned DNA are identical, we conclude that no important rearrangements occurred during the cloning of the DNA. The difference observed with Hae Ill enzyme is discussed below.
Discussion
Our results show that it is possible to clone a specific DNA fragment in A phage from an unfrac-

Intervening Sequences in the Ovalbumin Gene 637
Figure 6. Comparison of Restriction Fragments Produced from the Cloned and the Chromosomal 2.35 kb Eco RI Fragment “b” upon Digestion with Restriction Enzymes Hinf I, Hae Ill, Mbo II and Sst I
An Eco RI digest of hOvEb9 DNA (0.01 fig) or Eco RI fragment “b” DNA (1 pg), partially purified on RPC5 (see the text; also Mandel et al., 1976), were digested with Hinf I, Hae Ill, Mbo II or Sst I. The DNA fragments obtained were electrophoresed and revealed by hybridization as described in the legend to Figure 7. Lane 1: Hhaov DNA marker (2.4 kb). Lanes 2,8 and 13: Hae Ill internal marker (see Figure 2). Lane 7: Hinf I marker (see Figure 2). Lanes 3-6: digests of the cloned 2.35 kb DNA fragment with Hinf I (lane 3), Hae Ill (lane 4), Mbo II (lane 5) and Sst I (lane 6). Lanes 9-12: digests of RPCS partially purified chromosomal Eco RI fragment “b” with Hinf I (lane 9), Hae Ill (lane lo), Mbo II (lane 11) and Sst I (lane 12). The sizes of the hybridizing fragments are (in kb): lane 3-(a) 1.32. (b) 0.75; lane 4-(a) 1.46, (b) 0.65; lane 5-(a) 0.7, (b) 0.6, (c) 0.4; lane 6-(a) 1.59; lane 9-(a) 1.32, (b’) 1.03. (b) 0.75; lane 10-(a) 1.7. (b) 0.65; lane 11 -(a) 0.7, (b) 0.6, (c) 0.4; lane 12-(a) 1.59.
tionated Eco RI digest of chicken DNA by a “shot- gun” approach. Two independent recombinants (hOvEb5 and AOvEb9) containing the 2.35 kb oval- bumin gene Eco RI fragment “b” have been ob- tained. In clone AOvEb9, the presence of additional fragments which did not hybridize to the ovalbumin gene probes was not unexpected, since the recom- bination was carried out ata 1 :l weight ratio for h- DNA and Eco RI-digested cellular DNA, a high molar ratio for the low molecular weight Eco RI fragments. Most of’ our studies have been per- formed on clone hOvEb9, because clone AOvEb5 was unstable, probably for the reason mentioned above in Results.
Our electron microscopy and restriction enzyme mapping studies establish unequivocally that the sequence coding for the 5’ quarter of the ovalbu- min mRNA (-500 nucleotides) is split into four regions separated by three intervening sequences. The coding regions range in size from -70-220 bp, whereas the three intervening sequences range
from -200-600 bp. Taken together with the results of Breathnach et al. (1977) and Mandel et al. (1978), our present results demonstrate that the ovalbumin gene is a “highly” split gene. The possible func- tional relevance of the clustering of the intervening sequences in the regions coding for the 5’ moiety of the ovalbumin mRNA is discussed by Mandel et al. (1978).
Studies on the cloned AOvEb5 DNA give results identical to those obtained with clone AOvEb9 (J. P. LePennec, unpublished data), suggesting that the cloning procedure is reliable. This is verified by the fact that the restriction enzyme pattern of the chromosomal DNA fragment “b” is, apart from the Hae III pattern, identical to that of the correspond- ing cloned fragment. Two possibilities might ac- count for the failure of the enzyme Hae III to cut the chromosomal DNA fragment at a site Hae 1 (Figure 8). First, the Hae 1 site could be altered in the cellular DNA by a modification acting like methyla- tion, which is known to prevent digestion by some restriction endonucleases (Bird and Southern, 1978). Second, since the chromosomal DNA which served for cloning and that which was used in the digestion studies shown in Figure 8 did not origi; nate from the same hen, the possibility exists that the absence of the Hae 1 site is related to an allelic variation similar to that described by Mandel et al. (1978). Should this be the case, it is interesting to note that the allelic variation would be located in an intervening sequence.
Studies on adenovirus type 2 and SV40 have revealed the presence of “leader” sequences at the 5’ end of some of their messenger RNAs (see Introduction for references). For instance, many late adenovirus mRNAs consist of four separate blocks of, sequences. In every case, three small RNA fragments coded for by three different seg- ments of the genome, remote from the DNA coding for the main body of the mRNAs, are joined to form a common “leader” sequence of 150-200 nucleo- tides at the 5’ end of these late mRNAs. In SV40, the “leader” sequence which is present at the 5’ end of the late mRNAs is coded for by a single region of the genome. It is clear from our present results that there is not such a long “leader” sequence at the 5’ end of ovalbumin mRNA (we cannot exclude from the electron microscope data a “leader” sequence shorter than 50 nucleotides). Studies on other mRNAs, now in progress in our laboratory, will indicate whether the absence of a long “leader” sequence is general for cellular mes- sengers. As pointed out by Crick (1978), common “leader” sequences could very well be restricted to viruses with a small genome, where repetition of DNA sequences might lead to unacceptable rates of recombination.

Cell 636
Experimental Procedures
Cloning Procedure XgtWES.Ac DNA was digested by Eco RI; the arms of the vector were isolated by sucrose gradient sedimentation as described by Kourilsky et al. (1976) and concentrated by ethanol precipitation. In vitro recombination, in vitro packaging, plating on E. coli strain 603r-lrm-k SupE SupF (Murray, grammar and Murray, 1976) and in situ hybridization were carried out as described by Garapin et al. (1976). The clones carrying the 2.35 kb Eco RI fragment “b” were termed XOvEb where ov, E and b denote ovalbumin, Eco RI and fragment “b,” respectively.
Electron Microscopy Hybrids between the ovalbumin mRNA and DNA of either clone XOvEb9 or the 2.35 kb Eco RI fragment (purified by sucrose gradient centrifugation) were prepared as follows. Ovalbumin mRNA (3 pg/ml) and the appropriate heat-denatured DNA (3 fig/ ml) were incubated at 545°C in 10 mM Tris-HCI (pH 8.5), 0.3 M NaCl. 1 mM EDTA, 70% formamide for 5 hr (Casey and Davidson, 1977). Hybrids of ov mRNA with both the 2.35 kb Eco RI fragment “b” and the 2.6 kb Eco RI-Hind Ill fragment (purified by prepara- tive agarose gel electrophoresis from a Hind Ill digest of the 9.2 kb Eco RI fragment “a”) were obtained as follows. 2.35 kb fragment “b”-ov mRNA hybrids were prepared as described above, and then hybridized to heat-denatured 2.6 kb Eco RI-Hind Ill fragments (5 *g/ml). Heteroduplexes between Hhaov DNA (Humphries et al., 1977) and the 2.35 kb Eco RI fragment “b” were prepared as follows. Samples of each DNA (3 pglml) were heat-denatured in 10 mM Tris-HCI (pH 6.5). 0.3 M NaCl, 1 mM EDTA, 70% formamide at 75°C for 5 min. and then incubated at 25°C for 30 min. In all cases, the mixtures were diluted 10 fold before spreading, as described in Garapin et al. (1976). GX174 (RF) and single-stranded DNAs were used as length standards for double- and single-stranded DNA molecules, respectively.
Other Procedures and Materials Chromosomal Eco RI fragment “b” was partially purified by reverse-phase chromatography (RPC5) as described by Mandel et al. (1976). 32P-labeled ovalbumin double-stranded cDNA probes (Hhaov, Pst A and Pst B) were prepared as described by Breath- nach et al. (1977) and Mandel et al. (1976). Restriction enzyme digestions were carried out as recommended in the Biolabs booklet. DNA restriction enzyme fragments were electropho- resed, transferred to nitrocellulose filters, hybridized to 3ZP-la- beled ovalbumin probes and revealed by autoradiography as described by Breathnach et al. (1977) and Mandel et al. (1976).
All other methods and materials were as described by Breath- nach et al. (1977). Garapin et al. (1978) and Mandel et al. (1976).
Biohazards associated with the experiments described in this publication were examined previously by the French National Control Committee. The experiments were carried out accord- ingly under L3B2 conditions in the nomenclature adopted by the French Committee (Le Progres Scientifique N” 191, November- December 1977, in press: L3B2 is considered equivalent to P3 EK2 in the NIH nomenclature).
Acknowledgments
We thank Mrs. E. Sittler, Miss E. Regnier, Mr. E. Taubert and Mr. J. M. Garnier for excellent technical assistance in Strasbourg. We are grateful to A. Krust and M. Le Meur (Strasbourg) for gifts of materials. Electron microscopy facilities were made available to F. P. in Strasbourg by the CNRS and the INSERM. This work was supported in Paris by grants to P. K. from the CNRS, the DGRST and the INSERM, and in Strasbourg by grants to P. C. from the INSERM, the CNRS and the Fondation pour la Recherche Medi- tale Francaise. W. R. and J. P. Le P. were supported by DGRST fellowships.
The costs of publication of this article were defrayed in part by
the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 16 U.S.C. Section 1734 solely to indicate this fact.
Received March 31, 1976
References
Bellett. A. J. D., Busse, H. G. and Baldwin, R. L. (1971). Tandem genetic duplications in a derivative of phage lambda. In The Bacteriophage Lambda, A. D. Hershey, ed. (New York: Cold Spring Harbor Monograph Series), pp. 501-513.
Bird, A. P. and Southern, E. M. (1976). Use of restriction enzymes to study eukaryotic DNA methylations. J. Mol. Biol. 118, 27-47.
Brack, C. and Tonegawa, S. (1977). Variable and constant parts of immunoglobulin light chain gene of a mouse myeloma cell are 1250 nontranslated bases apart. Proc. Nat. Acad. Sci. USA 74, 5652-5656.
Breathnach. R., Mandel, J. L. and Chambon, P. (1977). Ovalbu- min gene is split in chicken DNA. Nature 270, 314-319.
Casey, J. and Davidson, N. (1977). Rates of formation and thermal stabilities of RNA-DNA and DNA-DNA duplexes at high concentra- tions of formamide. Nucl. Acids Res. 4, 1539-1552.
Chambon, P. (1977). Molecular biology of eukaryotic genome is coming of age. Cold Spring Harbor Symp. Quant. Biol. 42, 1211- 1235.
Chow, L. T., Gelinas, R. E., Broker, T. R. and Roberts, R. J. (1977). An amazing sequence arrangement at the 5’ ends of adenovirus 2 messenger RNA. Cell 72, 1-8.
Crick, F. M. C. (1978). Chromosome structure and function. Future prospects. Eur. J. Biochem. 83, l-3.
Doel, M. T., Houghton, M.. Cook, E. A. and Carey, N. H. (1977). The presence of ovalbumin mRNA coding sequences in multiple restriction fragments of chicken DNA. Nucl. Acids Res. 4, 3701- 3713.
Dunn, A. R. and Hassell, J. A. (1977). A novel method to map transcripts: evidence for homology between an adenovirus mRNA and discrete multiple regions of the viral genome. Cell 12, 23-36.
Enquist, L.. Tiemeier. D., Leder. P., Weisbert, R. and Sternberg, N. (1976). Safer derivates of bacteriophage Agt.AC for use in cloning of recombinant DNA molecules. Nature 259, 596-598.
Garapin, A. C., Le Pennec, J. P., Roskam, W., Perrin, F., Cami, 8.. Krust, A., Breathnach, R., Chambon, P. and Kourilsky, P. (1978). Isolation by molecular cloning of a fragment of the split ovalbumin gene. Nature 273, 349-354.
Glover, D. M. and Hogness, D. S. (1977). A novel arrangement of the 18s and 28s sequences in a repeating unit of Drosophila melanogaster rDNA. Cell 10, 167-176.
Goodman, H. M.. Olson, M. V. and Hall, B. D. (1977). Nucleotide sequence of mutant eukaryotic gene: the yeast tyrosine-inserting ochre supressor SVP4-0. Proc. Nat. Acad. Sci. USA 74, 5453- 5457.
Hohn, 8. and Murray, K. (1977). Packaging recombinant DNA molecules into bacteriophage particles in vitro. Proc. Nat. Acad. Sci. USA 74, 3259-3263.
Humphries, P., Cachet. M., Krust. A., Gerlinger, P., Kourilsky, P. and Chambon, P. (1977). Molecular cloning of extensive se- quences of the in vitro synthesized chicken ovalbumin structural gene. Nucl. Acids Res. 4, 2389-2406.
Jeffreys, A. J. and Flavell, R. A. (1977). The rabbit p-globin gene contains a large inset-f in the coding sequence. Cell 12, 1097- 1108.
Klessig. D. F. (1977). Two adenovirus mRNAs have a common 5’ terminal leader sequence encoded at least 10 kb upstream from their main coding regions. Cell 72, 9-21.
Kourilsky, P., Perricaudet, M.. Gros, D.. Garapin, A., Gottesman,

Intervening Sequences in the Ovalbumin Gene 639
M., Fritsch, A. and Tiollais, P. (1978). Description and properties of bacteriophage A vectors useful for the cloning of Eco RI DNA fragments. Biochimie 60, 183-187.
Leder, P., Tilghman. S. M., Tiemeier. D. C., Polsky, F. I., Seid- man, J. G.. Edgell, M. H., Enquist, L. W., Leder. A. and Norman, 6. (1977). The cloning of mouse globin and surrounding gene sequences in bacteriophage A. Cold Spring Harbor Symp. Quant. Biol. 4.2, in press.
Lewis, J. B., Anderson, C. W. and Atkins, J. F. (1977). Further mapping of late adenovirus genes by cell-free translation of RNA selected by hybridization to specific DNA fragments. Cell 12, 37- 44.
Mandel, J. L., Breathnach, Ft., Gerlinger, P., Le Meur, M., Gan- non, F. and Chambon, P. (1978). Organization of coding and intervening sequences in the chicken ovalbumin split gene. Cell 14, 641-653.
Murray, N. E.. Brammar. W. J. and Murray, K. (1976). Lambdoid phages that simplify recovery of in vitro recombinants. Mol. Gen. Genet. 150, 53-61.
Pellegrini, M., Manning, J. and Davidson, N. (1977). Sequence arrangement of the rDNA of Drosophila melanogaster. Cell 70, 213-224.
Rabbitts. T. H. and Forster, A. (1978). Evidence for noncontiguous variable and constant region genes in both germ line and mye- loma DNA. Cell 73, 319-327.
Rosen, J. M., Woo, S. L. C., Means, A. R. and O’Malley. B. W. (1976). Purification and characterization of ovalbumin mRNA in the chick oviduct. In Methods in Molecular Biology, 8, Eukaryotes at the Subcellular Level, J. Last, ed. (New York and Basel: Marcel Dekker), pp. 369-434.
Sambrook. J. (1977). Adenovirus amazes at Cold Spring Harbor. Nature 268, 101-104.
Tilghman, S. M., Tiemeier. D. C., Seidman, J. G.. Peterlin, B. M., Sullivan, M., Maizel, J. V. and Leder. P. (1978). An intervening sequence of DNA identified in the structural portion of a mouse p-globin gene. Proc. Nat. Acad. Sci. USA 75, 725-729.
Valenzuela. P., Venegas, A., Weinberg, F., Bishop, R. and Rutter, W. J. (1978). Structure of yeast phenylalanine-tRNA genes. An intervening DNA segment within the region coding for the tRNA. Proc. Nat. Acad. Sci. USA 75, 190-194.
Weinstock, R., Sweet, R., Weiss, M., Cedar, H. and Axel. R. (1978). lntragenic DNA spacers interrupt the ovalbumin gene. Proc. Nat. Acad. Sci. USA 75, 1299-1303.
Wellauer, P. K. and Dawid. I. 8. (1977). The structural organiza- tion of ribosomal DNA in Drosophila melanogaster. Cell 70, 193- 212.
White, R. L. and Hogness, D. S. (1977). R loop mapping of the 18s and 28s sequences in the long and short repeating units of Drosophila melanogaster rDNA. Cell 10, 177-192.