einfluss eines kontrollierten zinkmangels auf die...
TRANSCRIPT

Einfluss eines kontrollierten Zinkmangels
auf die Proteinexpression im Heilungsprozess
intestinaler Anastomosen der Ratte
Marion Franken

Einfluss eines kontrollierten Zinkmangels
auf die Proteinexpression im Heilungsprozess
intestinaler Anastomosen der Ratte
Von der Medizinischen Fakultät
der Rheinisch-Westfälischen Technischen Hochschule Aachen zur Erlangung des akademischen Grades
einer Doktorin der Medizin genehmigte Dissertation
vorgelegt von
Marion Franken
aus
Erkelenz
Berichter: Herr Universitätsprofessor Dr.med. Dr.E.h.(GUS) Volker Schumpelick
Herr Professor Dr.med. Andreas Prescher
Tag der mündlichen Prüfung: 17. Mai 2010
Diese Dissertation ist auf den Internetseiten der Hochschulbibliothek online verfügbar.

Für Onkel Hans

Inhaltsverzeichnis
I
Inhaltsverzeichnis
Abbildungsverzeichnis....................................................................................III
Tabellenverzeichnis.........................................................................................IV
Abkürzungsverzeichnis............................................. ......................................V
1 Einleitung .................................................................................... 1
1.1 Wundheilung ............................................................................................3
1.1.1 Physiologie der allgemeinen Wundheilung ........................................ 3 1.1.2 Bedeutung der extrazellulären Matrix in der Wundheilung ............... 4 1.1.3 Wundheilung im Gastrointestinaltrakt ................................................ 5
1.6 Notch-3................................................................................................... 16
1.7 Zink ......................................................................................................... 19
1.7.1 Allgemeine Bedeutung ..................................................................... 19 1.7.2 Zink in der Wundheilung................................................................... 21
2 Zielsetzung.............................................................................. 23
3 Material und Methoden ............................................................ 24
3.1 Tiermodell .............................................................................................. 24
3.2 Histologie............................................................................................... 25
3.4 Färbungen.............................................................................................. 27
3.4.1 TIMP-1.............................................................................................. 27 3.4.2 ß-Catenin.......................................................................................... 28 3.4.3 c-myc ................................................................................................ 29 3.4.4 Notch-3 ............................................................................................. 30
3.5 Auswertung ........................................................................................... 31
3.5.1 Remmele-Score ............................................................................... 32 3.5.2 Statistik ............................................................................................. 33

Inhaltsverzeichnis
II
4 Ergebnisse ............................................................................... 34
4.1 TIMP-1..................................................................................................... 35
4.2 ß-Catenin............................................................................................ 38
4.3 c-myc .................................................................................................. 44
4.4 Notch-3 ............................................................................................... 50
5 Diskussion .............................................................................. 55
5.1 TIMP-1..................................................................................................... 56
5.2 ß-Catenin................................................................................................ 58
5.3 c-myc...................................................................................................... 59
5.4 Notch-3................................................................................................... 61
6 Zusammenfassung................................................................. 63
7 Literaturverzeichnis ............................................................... 65
8 Danksagung............................................................................ 74
9 Datenaufbewahrung............................................................... 75
10 Lebenslauf ........................................................................... 76

Abbildungsverzeichnis
III
Abbildungsverzeichnis
Abbildung 1: Schematische Darstellung der extrazellulären Matrix ................... 4
Abbildung 2: Schematische Darstellung der Struktur von TIMP-1 ..................... 9
Abbildung 3: ß-Catenin in seiner Funktion als Strukturprotein ......................... 11
Abbildung 4: Funktion von ß-Catenin im Wnt-Signalweg ................................. 12
Abbildung 5: Abbildung des Zellzyklus und der physiologischen Zellhomöostase
........................................................................................................................... 14
Abbildung 6: Notch-Kaskade a) inaktiv; b) aktiv ............................................... 16
Abbildung 7: Notch-3 in der intestinalen Zellerneuerung ................................. 17
Abbildung 8: Beispielpräparat TIMP-1; Kontrolle, Kolotomie, schnittnah ......... 27
Abbildung 9: Beispielpräparat ß-Catenin; Kontrolle, Anastomose, schnittfern 28
Abbildung 10: Beispielpräparat c-myc; Kontrolle, Anastomose, schnittnah ..... 29
Abbildung 11: Notch-3 Präparat; Zinkmangel, Anastomose, schnittfern ......... 30
Abbildung 12: Schaubild der Untersuchungsebenen ....................................... 31
Abbildung 13: TIMP-1-Expression im Vergleich der Nahttechniken für
schnittnahe Mukosa (Kontrollgruppe)................................................................ 36
Abbildung 14: TIMP-1-Expression im Vergleich der Nahttechnik für schnittnahe
Mukosa (Zinkmangelgruppe)............................................................................. 37
Abbildung 15: ß-Catenin-Expression im Vergleich der Gruppen für schnittferne
Submukosa (Kolotomie) .................................................................................... 39
Abbildung 16: ß-Catenin-Expression im Vergleich der Gruppen für schnittferne
Submukosa (Kolotomie) .................................................................................... 39
Abbildung 17: ß-Catenin-Expression im Vergleich der Nahttechnik für
schnittnahe Mukosa (Kontrollgruppe)................................................................ 40
Abbildung 18: ß-Catenin-Expression im Vergleich der Nahttechnik für
schnittnahe Mukosa (Zinkmangelgruppe) ......................................................... 41
Abbildung 19: ß-Catenin-Expression im Vergleich der Nahttechnik für
schnittferne Submukosa (Zinkmangelgruppe)................................................... 42
Abbildung 20: ß-Catenin-Präparat der Kontrollgruppe; anastomosennah ....... 43
Abbildung 21: ß-Catenin-Präparat der Kontrollgruppe; anastomosenfern ....... 43

Abbildungsverzeichnis
IV
Abbildung 22: c-myc-Expression im Gruppenvergleich für schnittnahe Mukosa
(Anastomose)..................................................................................................... 45
Abbildung 23: c-myc-Expression im Gruppenvergleich für schnittferne Mukosa
(Anastomose)..................................................................................................... 45
Abbildung 24: c-myc-Expression im Gruppenvergleich für schnittferne
Submukosa (Anastomose) ................................................................................ 46
Abbildung 25: c-myc-Expression im Vergleich der Nahttechniken für schnittferne
Mukosa (Kontrolle)............................................................................................. 47
Abbildung 26: c-myc-Expression im Vergleich der Nahttechniken für schnittferne
Submukosa (Kontrolle) ...................................................................................... 48
Abbildung 27: c-myc-Expression im Vergleich der Nahttechniken für schnittnahe
Mukosa (Zinkmangel) ........................................................................................ 48
Abbildung 28: c-myc-Expression im Vergleich der Nahttechniken für schnittferne
Mukosa (Zinkmangel) ........................................................................................ 49
Abbildung 29: c-myc-Expression im Vergleich der Nahttechniken für schnittferne
Submukosa (Zinkmangel).................................................................................. 49
Abbildung 30: Notch-3-Expression im Gruppenvergleich für schnittferne Mukosa
(Anastomose)..................................................................................................... 51
Abbildung 31: Notch-3-Expression im Gruppenvergleich für schnittferne
Submukosa (Anastomose) ................................................................................ 51
Abbildung 32: Notch-3-Expression im Gruppenvergleich für schnittferne Mukosa
(Kolotomie)......................................................................................................... 52
Abbildung 33: Notch-3-Expression im Vergleich der Nahttechniken für
schnittferne Mukosa (Zinkmangel) .................................................................... 53
Abbildung 34: Notch-3-Expression im Vergleich der Nahttechniken für
schnittferne Submukosa (Zinkmangel) .............................................................. 54

Tabellenverzeichnis
V
Tabellenverzeichnis
Tabelle 1: p-Werte der TIMP-1-Scores im Vergleich der Gruppen .................. 35
Tabelle 2: p-Werte der TIMP-1-Scores im Vergleich der Nahttechniken ......... 36
Tabelle 3: p-Werte der ß-Catenin-Scores im Vergleich der Gruppen ............... 38
Tabelle 4: p-Werte der ß-Catenin-Scores im Vergleich der Nahttechniken...... 40
Tabelle 5: p-Werte der c-myc-Scores im Vergleich der Gruppen ..................... 44
Tabelle 6: p-Werte der c-myc-Scores im Vergleich der Nahttechniken ........... 47
Tabelle 7: p-Werte der Notch-3-Scores im Vergleich der Gruppen ................. 50
Tabelle 8: p-Werte der Notch-3-Scores im Vergleich der Nahttechniken ........ 53

Abkürzungsverzeichnis
VI
Abkürzungsverzeichnis
ADAM Adamalysin-ähnliche Proteasen
CADASIL Autosomal dominant erbliche zerebrale
Arteriopathie, die zu subkortikalen Infarkten und
Leukoenzephalopathien führt
CSL Genregulatorprotein im Notch-Pathway
DNA Desoxyribonukleinsäure
DAB 3,3 Diaminobenzidine tablets
ECM Extrazellulärmatrix
EDTA Ethylendiamintetraessigsäure
EGF Epidermal growth factor
FGF Fibroblast growth factor
HES-1 Zielgen des Notch-Pathway
IL-1 beta Interleukin-1 beta
JK Janus Kinase
Lef Lymphoid-enhancer-binding-factor
MMP Matrixmetalloproteinase
Notch-IC Notch-intrazelluläre Domäne
PBS Phosphate Buffered Saline
PDGF Platelet-derived growth factor
ppPDGF partially purified Platelet-derived growth factor
RBP Regulatorprotein im Zellzyklus (RBP/JK)
RNA Ribonukleinsäure
rTGF-ß Rekombinator tissue growth factor
TCF T cell factor
TIMP Tissue Inhibitor of Metalloproteinases
TNF-alpha Tumor-Nekrose-Faktor-alpha
Wnt Signalproteine, welche als lokale Mediatoren
agieren

1 Einleitung
1
1 Einleitung
“Anastomotic leakage remains the most important cause of morbidity and
mortality in digestive surgery.” (Kostic, 1994)
Operationen am Kolon gehören vor allem wegen der Bedeutung des
Kolonkarzinoms als zweithäufigstes Malignom bei Männern und Frauen zu den
häufigsten Eingriffen in der Viszeralchirurgie (Willis und Schumpelick, 2005).
Obwohl Operationen am eröffneten Darm heutzutage Routineoperationen
darstellen, bleibt die Nahtinsuffizienz auf der Basis einer Wundheilungsstörung
noch heute die gefährlichste Komplikation in der kolorektalen Chirurgie
(Hendriks und Mastboom, 1990). Daher bleiben Untersuchungen ihres
Heilungsprozesses von großem Interesse.
Die schnelle und komplikationsarme Wundheilung hat bei jedem invasiven
Eingriff einen großen Stellenwert. Neben Operationstechnik, Erfahrung sowie
handwerklichem Geschick des Operateurs nehmen viele, unterschiedliche
Faktoren Einfluss auf die molekularen Prozesse der Wundheilung (Willis und
Schumpelick, 2005; Stumpf et al., 2004; Burch et al., 2000). Man unterscheidet
hierbei zwischen patienteneigenen und externen Risikofaktoren sowie zwischen
systemischen und lokalen Einflüssen. Zu den letzteren beiden sind unter
anderem Veränderungen der extrazellulären Matrix (ECM) und als deren
Effektoren auch Veränderungen der Zellmigration, Zellproliferation sowie
Zelldifferenzierung zu zählen. Diese werden durch dynamische Prozesse auf
molekularer Ebene gesteuert. Die Prävention von Wundheilungsstörungen setzt
daher ein grundlegendes Verständnis der einzelnen, an der Wundheilung
beteiligten Faktoren voraus.
Das Forschungsinteresse an diesen Faktoren besitzt eine lange Tradition.
Bereits 1922 publizierte Ishido eine Studie, deren Versuchsaufbau mit dem
dieser Arbeit vergleichbar ist. Unter der Annahme, dass sich Mangelzustände
negativ auf den Zellstoffwechsel und in dessen Folge auch auf die Wundheilung
auswirken, untersuchte Ishido in unterschiedlich ernährten Versuchstiergruppen
mikroskopisch die postoperative Beschaffenheit von künstlich induzierten
Wunden im Bereich der Rückenhaut und Rückenmuskulatur von Ratten (Ishido,
1922). Unter einem allgemeinen Ernährungsmangels bzw. einer Avitaminose

1 Einleitung
2
konnte er eine verzögerte Wundheilung sowie eine Beeinträchtigung des
Immunsystems mit erhöhtem Bakteriennachweis in den Schnitten (Ishido,
1922).
Die Anastomosenheilung am Darm entspricht den drei Phasen der
Wundheilung. Dem initialen Trauma der Anastomose bzw. Kolotomie folgt
zunächst die Substratphase, welche neben Exsudation und Resorption eine
Entzündungsreaktion beinhaltet. Die unterschiedlichen Schichten der
Darmwand (Mukosa, Basalmembran und Tela submucosa) fördern über eine
Modulation der Extrazellulärmatrix (ECM) den Heilungsprozess. Zur ECM
zählen Kollagen Typ I und Typ III, Fibronektin und verschiedene Proteoglycane,
wie zum Beispiel die Tissue inhibitors of Matrixproteinasen (TIMPs). Die
primäre Naht ist in der Tunica serosa verankert. Hierdurch wird die
Vorraussetzung zur Akkumulation von Kollagenfasern in der Tela submucosa
geschaffen (Dietz und Debus 2002). Der Kollagen I/III Ratio ist ein wichtiges
mechanisches Qualitätsmerkmal für die Stabilität der Narbe (Klinge et al. 2000).
Ist dieses Verhältnis durch externe oder interne Einflüsse gestört, kommt es zu
einer Dysregulation des Kollagenstoffwechsels wodurch die Wundheilung
empfindlich gestört wird. Wichtig ist bei der Regulierung vor allem das
Gleichgewicht zwischen Synthese und Degradierung von Proteinen in der ECM.
Dieses wird durch eine fein gesteuerte Interaktion zwischen Matrix-
Metalloproteinasen (MMPs) und deren Inhibitoren TIMPs (Tissue Inhibitors of
Metalloproteinases) gewährleistet (Gomez et al., 1997; McKaig et al., 2003,
Visse und Nagase 2003). Die MMPs besitzen ein zentrales Zink-Atom, welches
als Bindungsstelle für TIMPs dient.
Das transmembranäre Protein Notch-3 sowie die Onkogene ß-Catenin und c-
myc besitzen wegen ihrer physiologischen Funktionen ebenfalls eine besondere
Bedeutung. Notch-3 besitzt als membrangebundener Transkriptionsfaktor
sowohl Verbindungen zur ECM als auch zum Intrazellulärraum (Alberts et al.,
2004). ß-Catenin und c-myc sind Protagonisten des Zellzyklus und ihr
Vorhandensein ist für die Zellerneuerung eine notwendige Vorraussetzung
(Lodish et al., 2008; Alberts et al., 2004; Knippers, 2001). Da das Prinzip der
Wundheilung neben einer Umstrukturierung der ECM auch auf Zellerneuerung
und Proliferation beruht, ist eine Beteiligung der genannten Faktoren

1 Einleitung
3
naheliegend und soll in dieser Arbeit untersucht werden. Dass eine Störung des
fragilen Gleichgewichtes der Reparaturvorgänge in der ECM bei negativer
Zinkbilanz über eine eingeschränkte Funktion der MMPs die Wundheilung
supprimieren kann, ist bekannt (Binnebösel et al., 2009). Der Einfluss von Zink
bzw. eines Zinkmangels auf die physiologische Funktion der anderen oben
angegebenen Regulatoren ist dagegen noch unbekannt.
1.1 Wundheilung
1.1.1 Physiologie der allgemeinen Wundheilung
Die Wundheilung stellt eine Untergruppe allgemeiner regeneratorischer
Prozesse dar und wird in Substratphase (Exsudationsphase und
Resorptionsphase), Proliferationsphase und Differenzierungsphase unterteilt
(Schumpelick et al., 2004). In jeder Phase sind typische Signalpeptide aktiv und
induzieren spezifische Veränderungen und Reaktionen der extrazellulären
Matrix. Eine entzündliche, exsudative und katabole Sofortreaktion führt zur
Bildung einer provisorischen Wundmatrix welche die Proliferation, die
Differenzierung und die Synthese neuer Matrixmoleküle stimuliert (Schultz et al,
1995). Durch die Sezernierung chemotaktisch wirksamer Zytokine (PDGF, r-
TGF-ß, TNF-alpha, etc.) werden Fibroblasten, Granulozyten und Monozyten zur
Einwanderung in den Wundbereich angeregt (Hofstädter, 1995; Lawrence,
1998). Neben dem zentralen Defekt ist oft auch das umgebende Gewebe
betroffen. Dort entsteht durch Hypoxie, anaerobe Glykolyse und Azidose ein für
katabole Enzyme optimales, niedrigeres pH-Niveau. Nach primärem
Substanzverlust kommt es durch Proliferation zu einer Zunahme des DNA- und
Gesamtzellgehaltes im Wundsegment (Lindner, 1995). Parallel sprossen
Kapillaren in das Wundgebiet ein. Dadurch wird der pH-Wert in den neutralen
Bereich verschoben und der Sauerstoffmangel aufgehoben (Haase und Rink,
2006). In der Differenzierungsphase sinkt die Anzahl der Bindegewebszellen im
Wundbereich und es wird vermehrt Interzellularsubstanz gebildet. Durch
Kontakt mit ECM-Proteinen und verschiedenen Wachstumsfaktoren
transformieren Fibroblasten zu Myofibroblasten, welche für die Wundkontraktion

1 Einleitung
4
verantwortlich sind (Clark, 1993; Arora et al., 1999). Je geringer der primäre
Substanzabbau in der Wundheilung war, desto besser und stabiler ist die
resultierende Narbe bezüglich ihrer Struktur und Funktion (Lindner, 1995).
1.1.2 Bedeutung der extrazellulären Matrix in der Wundheilung
Die extrazelluläre Matrix besteht aus einem dreidimensionalen Netz
verschiedener Kollagentypen sowie Elastin, Fibronektin, Glykosaminoglykanen
und Proteoglykanen. Sie umfasst alle unlöslichen Faktoren des interstitiellen
Raumes und Bestandteile der Basalmembran. Sie dient als Strukturelement
und beeinflusst außerdem die Zellentwicklung und -physiologie.
Abbildung 1: Schematische Darstellung der extrazellulären Matrix (UB, 2005)
Zu ihren Aufgaben zählen die Regulation zellulärer Funktionen, wie die der
Zelladhäsion und die der Organisation eines Transportsystems für Nähr- sowie
Abfallstoffe. Defekte der extrazellulären Matrix werden mit pathologischen
Prozessen, wie Entzündungen, chronisch-degenerativen Erkrankungen und
Tumorinvasion, in Verbindung gebracht (Gomez et al, 1997). Außerdem nimmt
die ECM an entscheidenden Prozessen der Wundheilung teil (Rhagow, 1994).
Stumpf et al wiesen in einer prospektiven Studie an Kolonkarzinom-Patienten
nach, dass sich Milieuveränderungen in der ECM störend auf die
Anastomosenheilung auswirken (Stumpf et al., 2004; Charrier, L, 2004).

1 Einleitung
5
Die Bestandteile der extrazellulären Matrix verändern sich parallel zu den
Phasen der Wundheilung. Unmittelbar nach einer Verletzung bildet sich eine
provisorische Wundmatrix, die hauptsächlich aus Fibrin und Fibronektin
besteht. Nach Ablauf einiger Tage sind von Fibroblasten synthetisierte
Kollagene und Proteoglykane die Hauptelemente der Matrix (Gailit und Clark,
1994). Pathologisch erhöhte Proteasenkonzentrationen können das Wundmilieu
in Richtung destruktiver Prozesse verschieben. Dabei werden Bestandteile der
ECM inklusive der Wachstumsfaktoren in erhöhtem Maße abgebaut. Die
Abbauvorgänge werden überwiegend durch die Matrix-Metalloproteinasen
(MMPs) vorgenommen. Die MMPs sind proteolytische Enzyme, die maßgeblich
an der Umstrukturierung der Extrazellulärmatrix im Rahmen physiologischer
und auch pathologischer Prozesse beteiligt sind. Sie sind auch in der
Umbauphase der Wundheilung aktiv. Dieser Prozess wird von den TIMPs, den
speziellen Inhibitoren der MMPs, kontrolliert. Sie verhindern, dass Teile der
funktionellen Matrix abgebaut werden. In chronischen Wunden kommt es durch
eine entzündliche Reaktion zu einer erhöhten Proteasenkonzentration und
damit zu einem Ungleichgewicht zwischen MMPs und TIMPs. Dies kann einen
pathologisch gesteigerten Matrix-Abbau zur Folge haben. Außerdem agiert die
ECM eng mit intrazellulären Prozessen zusammen. Viele Abläufe werden über
transmembranäre Rezeptoren und über die Interaktion mit Bestandteilen der
ECM vermittelt und ausgelöst (Lodish et al., 2008).
1.1.3 Wundheilung im Gastrointestinaltrakt
Grundsätzlich ist der Prozess der Wundheilung im Intestinum vergleichbar zur
Wundheilung in anderen Geweben. Aus morphologischen, funktionellen und
technischen Merkmalen ergeben sich jedoch auch Besonderheiten. Sowohl der
mehrschichtige Aufbau der Darmwand, als auch seine teilweise intraperitoneale
Lage müssen eine besondere Berücksichtigung in der Darmchirurgie finden.
Die intestinale Mukosa unterhält einen spezifischen physiologischen
Zellumsatz, der durch eine ständige Proliferation von Stammzellen in den
basalen Zellschichten der Krypten aufrecht erhalten wird und eine stetige
Selbsterneuerung innerhalb von 2-7 Tagen bewirkt (Alberts et al., 2004).

1 Einleitung
6
Auch Art, Größe und Lokalisation der Wunde nehmen Einfluss auf den
Heilungsverlauf. Durch die Keimbesiedlung im Dickdarm treten hier im Fall von
Insuffizienzen infektiöse Komplikationen häufiger auf, als im Dünndarm (Betzler
und Schmidt, 2005; Willis und Stumpf, 2004). Ein kontrovers diskutierter
Einflussfaktor einer Darm-Operation ist die postoperative Durchblutung der
Wundränder. Einige Studien dokumentieren Nahtinsuffizienzen als Folge einer
lokalen Minderperfusion (Stumpf et al, 2004; Betzler und Schmidt, 2005;
Hansen und Stelzner, 1975), andere Autoren postulieren, dass eine evtl.
entstandene ischämische Läsion klinisch zu vernachlässigen sei (Harder und
Kuhl, 1987). Die darmdurchblutenden Gefäße und ihre Verzweigungen
verlaufen größtenteils zirkulär und parallel zu den Fasern des Bindegewebes.
Manchen Autoren zufolge sollte die Schnittführung, um eine bessere
postoperative Durchblutung zu gewährleisten, senkrecht zur Darmlängsachse
erfolgen (Hansen und Stelzner, 1975). Dies ist bei der Kolotomie der Fall.

1 Einleitung
7
1.2 Nahttechniken
Anastomoseninsuffizienzen im unteren Gastrointestinaltrakt treten in ein bis
zwölf Prozent der Operationen auf und können für einen letalen
Krankheitsverlauf verantwortlich sein (Willis und Stumpf, 2004). Dies ist der
wichtigste Grund dafür, dass die intestinale Anastomose im Hinblick auf
Verbesserungen ihrer Durchführung in verschiedenen Studien immer wieder
von Interesse ist und bleibt. Die Kolonanastomose ist eine Nahtvereinigung von
zwei Organlumina nach querer Durchtrennung. Man unterscheidet zwischen
End-zu-End, End-zu-Seit, Seit-zu-Seit und Seit-zu-End Anastomosen
(Schumpelick et al., 2004).
Eine weitere Option für eine darmeröffnende Operationstechnik stellt die
Kolotomie dar. Die Kolotomie ist ein fast ausschließlich bei benignen
Grunderkrankungen verwendetes Operationsverfahren, welches beispielsweise
zur Polypentfernung eingesetzt wird (Willis und Schumpelick, 2005). Es handelt
sich hierbei, im Gegensatz zur Anastomose, um einen Längsschnitt durch alle
drei Schichten der Darmwand.
Operationsmethoden und Nahttechniken am Darm unterliegen einer stetigen
Entwicklung und spiegeln dabei den jeweiligen Kenntnisstand in Anatomie,
Physiologie, Antiseptik und Aseptik wieder (Schumacher und Lorenz, 1991;
Takeyama et al., 2007). Große Bedeutung besitzt auch die Auswahl von
Operationstechnik, Nahttechnik und Nahtmaterial. Aufgrund kontroverser
Studien bezüglich der Effizienz der unterschiedlichen Techniken gilt jedoch
auch, dass der Operateur diejenigen Methoden anwendet, welche er am besten
beherrscht (Stumpf et al., 2004).
Heutzutage werden bevorzugt einreihige Nahttechniken verwendet
(Schumacher und Lorenz, 1991). Die fortlaufende einreihige Nahttechnik mit
monophilem Nahtmaterial gilt zunehmend als Standardmethode bei intestinalen
Anastomosen. Diese wird unter Aussparung der Mukosa durchstochen und
ermöglicht eine lockere, primär dichte Adaption der Resektionsränder (Stumpf
et al., 2004). Dabei wird die Spannung über der Anastomose gleichmäßig
verteilt. Ein weiterer Vorteil dieser Technik ist, dass sie weniger zeitaufwendig
und kostengünstiger ist als die allschichtige, einreihige Einzelknopfnaht.

1 Einleitung
8
Zusätzlich wird die Menge des eingebrachten Nahtmaterials minimiert (Harder
und Kull, 1987). Die allschichtige, einreihige Einzelknopfnaht wird aber noch
alternativ verwendet. Die Submukosa, die zum großen Teil aus Kollagenfibrillen
besteht, spielt in ihrer Funktion als „Darmskelett“ die tragende Rolle in der
Darmchirurgie (Hendriks und Mastboom, 1990). Dies ist auch der Grund dafür,
dass es noch heute üblich ist, die Darmnähte in der Submukosa zu plazieren.
Das vorhandene Kollagen in der Submukosa ist somit für die Stabilität in der
frühen Phase der Wundheilung verantwortlich, während später das neu
aufgebaute Kollagen diese Funktion übernimmt (Hendriks und Mastboom,
1990; Stumpf et al., 2005; Binnebösel et al., 2009)
Dem Operateur stehen sowohl resorbierbare als auch nichtresorbierbare
Nahtmaterialien zur Verfügung. Das optimale Nahtmaterial sollte stabil sein und
durch minimales Gewebetrauma und geringe Umgebungs- und
Entzündungsreaktion eine gute Wundheilung ermöglichen. Nahtmaterialien gibt
es in der monophilen oder multiphilen Variante. Bei monophilem Material wird
ein atraumatischer Gewebedurchzug erreicht und eine Bakterienverschleppung
vermindert (Betzler und Schmidt, 2005). Die synthetische, resorbierbare,
monophile Naht ist nach Thornton und Barbul diejenige, die nach heutigen
Vorstellungen, der idealen Naht im intestinalen Wundmanagement am nächsten
kommt (Thornton und Barbul, 1997; Betzler und Schmidt, 2005; Harder und
Kuhl, 1987).
Maschinelle Klammernähte stellen eine weitere Möglichkeit zur
Anastomosierung im Intestinaltrakt dar. Häufige Verwendung findet diese
Technik sowohl bei laparoskopischen Operationen als auch bei Operationen am
rektosigmoidalen Übergang (Betzler und Schmidt, 2005; Takeyama et al.,
2007). In der Literatur ist die Rate der Insuffizienzen zwischen Klammernähten
und Handnähten vergleichbar. Für die Klammernaht wird jedoch ein größeres
Risiko von frühpostoperativen Blutungen und späten Anastomosenstenosen
beschrieben Takeyama et al., 2007; Thornton und Barbul, 1997).

1 Einleitung
9
1.3 Tissue Inhibitor of Metalloproteinases-1 (TIMP-1)
Bis heute sind vier Tissue Inhibitors of Metalloproteinases (TIMP-1 bis TIMP-4)
für Wirbeltiere beschrieben und charakterisiert worden. Sie sind als
Komponenten der extrazellulären Matrix ubiquitär als freie Moleküle in
menschlichen Geweben aber auch in Körperflüssigkeiten vorhanden (Alberts et
al., 2004; McKaig et al., 2003; Brew et al., 2000). TIMPs sind aus sechs
Schleifen aufgebaute Proteine mit sechs Disulfid-Bindungen, die aus zwei
Untereinheiten bestehen. Mit ihrem N-terminalen Ende, welches aus drei
Schleifen besteht, hemmen sie spezifisch die katalytische Domäne von Matrix-
Metalloproteinasen (MMPs) durch eine nicht-kovalente Bindung im Verhältnis
1:1 und inhibieren auf diese Weise deren Funktion (Gomez et al., 1997; Lee
und Murphy, 2004; Visse und Nagase, 2003).
Abbildung 2: Schematische Darstellung der Struktur von TIMP-1 (Cao und Zucker, 2004)
Alle vier TIMPs haben zwar fundamentale Ähnlichkeiten, weisen aber jeweils
auch markante strukturelle Eigenschaften, biochemische Wirksamkeiten und
spezifische Expressionsmuster auf (Baker et al., 2002; Brew et al., 2000).
TIMP-1 und TIMP-2 sind in der Lage, alle bisher bekannten Matrix-
Metalloproteinasen (MMPs) zu inhibieren (Gomez et al., 1997; McKaig et al.,
2003). MMPs sind eine multigene Familie von Enzymen, die zahlreiche
perizelluläre Substrate wie Kollagen, Wachstumsfaktoren, Zell-Oberflächen-
Rezeptoren, Adhäsionsmoleküle, andere Proteasen und viele mehr, bearbeiten
beziehungsweise abbauen (Thorgeirsson et al., 1996). Die MMPs enthalten

1 Einleitung
10
jeweils ein Zn²+-Molekül in ihrem aktiven Zentrum welches bei einer Inhibition
durch TIMPs als Bindungsstelle fungiert (van Waart und Birkedal-Hansen,
1990). Es besteht ein in der Regel gut organisiertes Wechselspiel zwischen
MMPs und TIMPs, welches sich wie oben beschrieben direkt auf den
Kollagenstoffwechsel auswirkt und dessen Endprodukt ein funktionierendes
Gerüst an strukturbildenden Proteinen bildet. Die negative Rückkopplung ist
eine wichtige Vorraussetzung für eine funktionierende Wundheilung. Ein
Zusammenhang zwischen der Aktivität von MMPs und dem Zink-Angebot im
Gewebe konnte von Krones bereits nachgewiesen werden (Krones, 2006).
Über ihre Kontrollfunktion auf die MMPs haben die TIMPs weitere wichtige
Funktionen in der Steuerung der Zell-Zell- bzw. Zell-Matrix-Interaktionen inne.
Sie greifen ebenso in den Ablauf intrazellulärer Prozesse ein (Alberts et al.,
2004). Auf diese Weise beeinflussen sie den Abbau und die Freisetzung von
Zellzyklus-steuernden Mediatoren, und damit die Zelldifferenzierung, die
Zellmigration, die Proliferation und das Zellüberleben. Außerdem schützen sie
Zelloberflächenrezeptoren, die für die Zelladhäsion notwendig sind, und
verhindern damit eine Zellmigration (Alberts et al., 2004). TIMPs werden vor
allem in Grenzbereichen des aktiven Proteinabbaus exprimiert, um die nicht
betroffene Matrix zu schützen (Alberts et al., 2004). Dies führt dazu, dass in
einem „nicht-physiologischen“ Gleichgewicht zwischen MMPs und TIMPs
fibrosierende Prozesse begünstigt werden; eine überhöhte TIMP-Expression ist
nachgewiesenermaßen mit gesteigerten Fibrosierungsraten vergesellschaftet
(McKaig et al., 2003; Roeb et al, 2000). Ungeklärt sind noch die Mechanismen,
durch die TIMPs einerseits das Zellwachstum fördern und andererseits Zelltod
bewirken und Proliferationssignale unterdrücken können (Sternlicht und Werb,
2001; Brew et al., 2000). Die Expression von TIMPs kann sowohl konstitutionell
als auch induzierbar sein (McKaig et al., 2003). Eine Induktion von TIMP-1 kann
zum Beispiel durch die Entzündungsmediatoren TNF-alpha und IL-1-beta
herbeigeführt werden (Borden und Heller, 1997; Voung et al, 2005; Kralisch et
al, 2006). Knittel et al. zeigten, dass das TIMP-Level direkt nach einer
Verletzung konstant bleibt, in der Entzündungsphase jedoch stark ansteigt und
während der Heilungsphase hoch bleibt (Knittel et al., 2000).
1 Einleitung
11
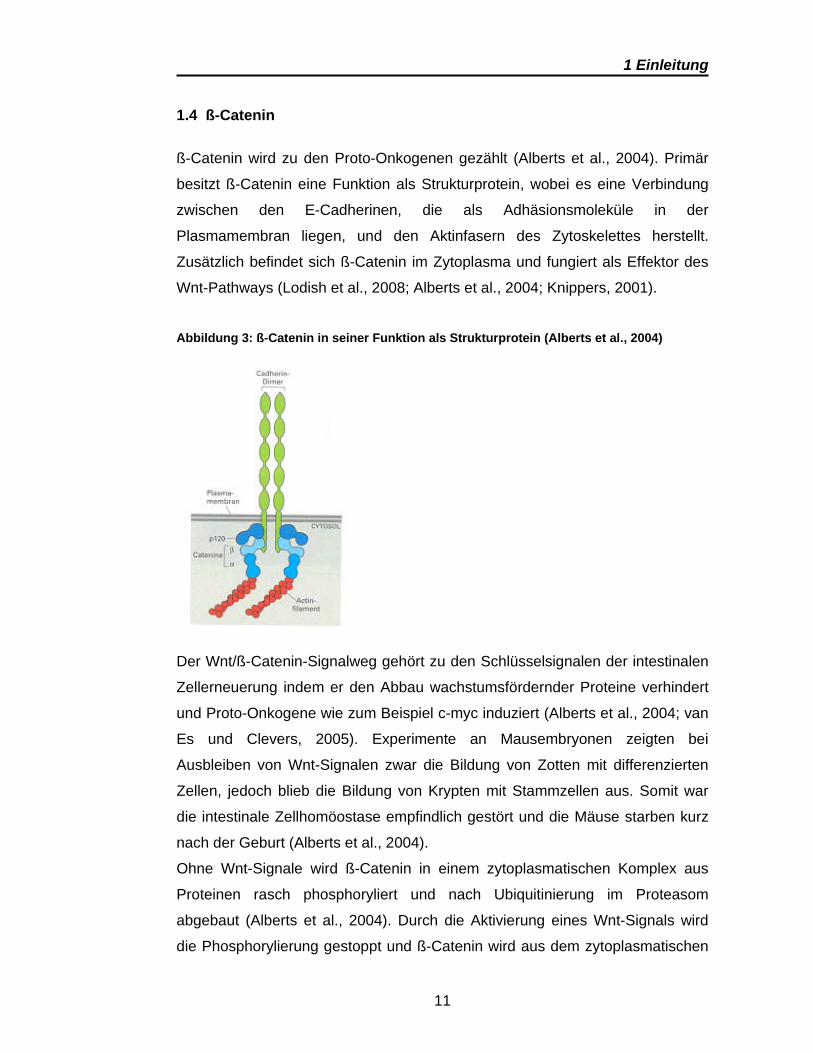
1.4 ß-Catenin
ß-Catenin wird zu den Proto-Onkogenen gezählt (Alberts et al., 2004). Primär
besitzt ß-Catenin eine Funktion als Strukturprotein, wobei es eine Verbindung
zwischen den E-Cadherinen, die als Adhäsionsmoleküle in der
Plasmamembran liegen, und den Aktinfasern des Zytoskelettes herstellt.
Zusätzlich befindet sich ß-Catenin im Zytoplasma und fungiert als Effektor des
Wnt-Pathways (Lodish et al., 2008; Alberts et al., 2004; Knippers, 2001).
Abbildung 3: ß-Catenin in seiner Funktion als Strukturprotein (Alberts et al., 2004)
Der Wnt/ß-Catenin-Signalweg gehört zu den Schlüsselsignalen der intestinalen
Zellerneuerung indem er den Abbau wachstumsfördernder Proteine verhindert
und Proto-Onkogene wie zum Beispiel c-myc induziert (Alberts et al., 2004; van
Es und Clevers, 2005). Experimente an Mausembryonen zeigten bei
Ausbleiben von Wnt-Signalen zwar die Bildung von Zotten mit differenzierten
Zellen, jedoch blieb die Bildung von Krypten mit Stammzellen aus. Somit war
die intestinale Zellhomöostase empfindlich gestört und die Mäuse starben kurz
nach der Geburt (Alberts et al., 2004).
Ohne Wnt-Signale wird ß-Catenin in einem zytoplasmatischen Komplex aus
Proteinen rasch phosphoryliert und nach Ubiquitinierung im Proteasom
abgebaut (Alberts et al., 2004). Durch die Aktivierung eines Wnt-Signals wird
die Phosphorylierung gestoppt und ß-Catenin wird aus dem zytoplasmatischen
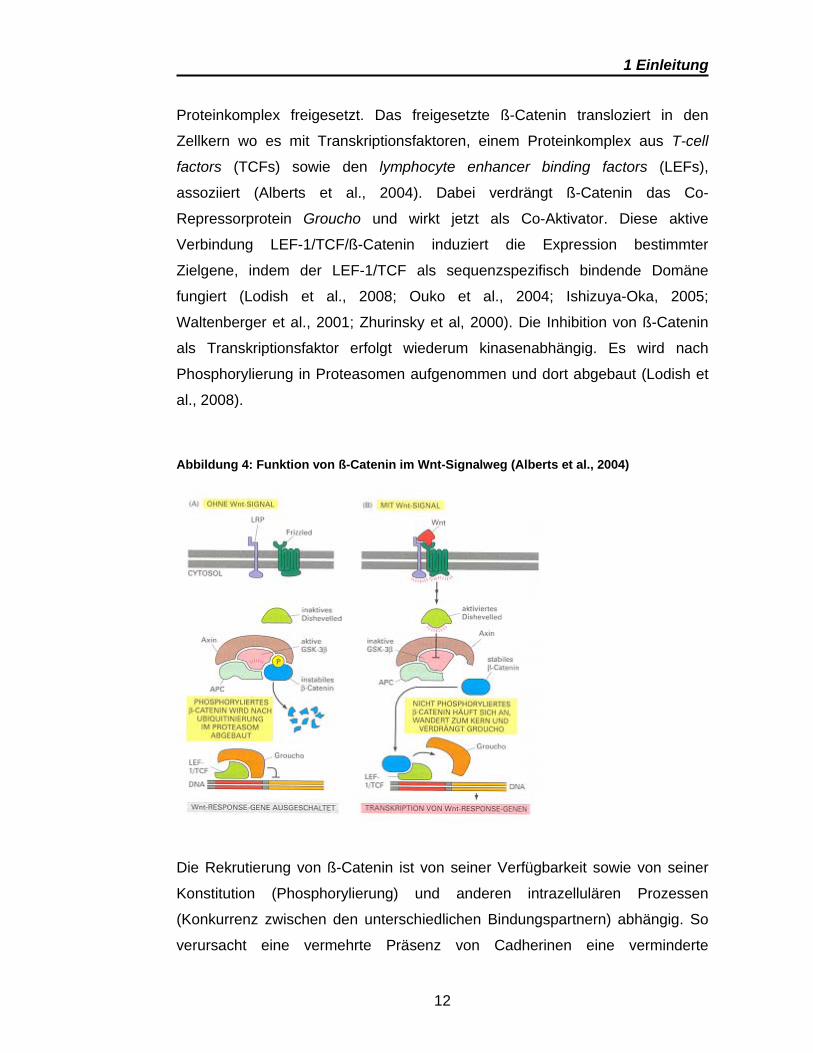
1 Einleitung
12
Proteinkomplex freigesetzt. Das freigesetzte ß-Catenin transloziert in den
Zellkern wo es mit Transkriptionsfaktoren, einem Proteinkomplex aus T-cell
factors (TCFs) sowie den lymphocyte enhancer binding factors (LEFs),
assoziiert (Alberts et al., 2004). Dabei verdrängt ß-Catenin das Co-
Repressorprotein Groucho und wirkt jetzt als Co-Aktivator. Diese aktive
Verbindung LEF-1/TCF/ß-Catenin induziert die Expression bestimmter
Zielgene, indem der LEF-1/TCF als sequenzspezifisch bindende Domäne
fungiert (Lodish et al., 2008; Ouko et al., 2004; Ishizuya-Oka, 2005;
Waltenberger et al., 2001; Zhurinsky et al, 2000). Die Inhibition von ß-Catenin
als Transkriptionsfaktor erfolgt wiederum kinasenabhängig. Es wird nach
Phosphorylierung in Proteasomen aufgenommen und dort abgebaut (Lodish et
al., 2008).
Abbildung 4: Funktion von ß-Catenin im Wnt-Signalweg (Alberts et al., 2004)
Die Rekrutierung von ß-Catenin ist von seiner Verfügbarkeit sowie von seiner
Konstitution (Phosphorylierung) und anderen intrazellulären Prozessen
(Konkurrenz zwischen den unterschiedlichen Bindungspartnern) abhängig. So
verursacht eine vermehrte Präsenz von Cadherinen eine verminderte

1 Einleitung
13
Verfügbarkeit von ß-Catenin in seiner Funktion als Transkriptionsfaktor
(Zhurinsky et al., 2000).
ß-Catenin kann aber auch eine Eskalation oben genannter Kaskaden im Sinne
einer unkontrollierten Transkriptionsaktivität auslösen und sich damit auch
negativ auf die physiologische Zellerneuerung, z.B. in chronischen Wunden,
auswirken. Die physiologische Funktion wechselt je nach Konzentration, Milieu
und Lokalisation. Punktmutationen an den beim Abbau phosphorylierten Enden
führen zu einer Stabilisierung von ß-Catenin. Auf diese Weise kann ß-Catenin,
neben seiner essentiellen Bedeutung für die Zellhomöostase des
Gastrointestinaltraktes und Zellerneuerungsvorgängen, im Falle eines
Kontrolldefizites zu diversen pathologischen Veränderungen wie z.B. Störung
der Zelldifferenzierung und -integrität. Auch in einigen Tumorarten hat man die
Akkumulation von ß-Catenin in den Zellkernen gefunden (Zhurinsky et al.,
2000). Bei einer übermäßigen Aktivierung des ß-Catenin/c-myc-Pathways
kommt es zu einer Aktivierung des Zellzyklus und zur Erschöpfung der Zellen
im Wundgebiet. In einem solchen Fall sind z.B. die Keratinozyten in einem
Zyklus von Aktivierung und Differenzierung blockiert: sie proliferieren, können
aber aufgrund eines inkompletten Aktivierungs-Pathways nicht migrieren
(Stojadinovic et al., 2005). Dies kann zur Konversion einer akuten Wunde in
eine chronische durch übermäßige ß-Catenin-Aktivierung führen (Stojadinovic
et al., 2005).
Versuche von Cheon et al. zeigen, dass Wunden mit einer hohen ß-Catenin-
Konzentration eine gesteigerte mesenchymale Proliferation mit einer größeren
Narbe aufweisen (Cheon et al., 2006). Ferner wurden persistierende
Entzündung, verminderte Aktivität von Wachstumsfaktoren und eine
beeinträchtigte Angiogenese als Folge einer übermäßigen ß-Catenin/c-myc-
Aktivierung beschrieben (Stojadinovic et al., 2005).

1 Einleitung
14
1.5 c-myc
Das Proto-Onkogen c-myc wird u.a. von ß-Catenin als Effektor des Wnt-
Signalweges aktiviert (Alberts et al., 2004; Bernhard und Eilers, 2006). Neben
dem Wnt-Signalweg können aber auch andere Onkogene und Zytokine seine
Expression beeinflussen (Chiariello et al., 2001; Xie et al., 2002). C-myc kodiert
die Translation von Myc-Proteinen, welche wiederum eine erhebliche Anzahl an
Genen regulieren (Alberts et al., 2004). Nach Transkription von c-myc und
Translation von C-Myc-Protein wandert dieses aus dem Zytoplasma in den
Zellkern. Nach Dimerisierung mit einem Partnerprotein bindet es zunächst
unspezifisch an die DNA. Diese Verbindung übernimmt eine Promotorfunktion
und vermittelt die Suche nach einer bestimmten Sequenz. Da die
Partnerproteine teilweise eigenständig als Transkriptionsfaktoren fungieren,
wird angenommen, dass C-Myc-Proteine als Kontrollinstanz dienen (Kato und
Dang, 1992). Niedrige C-Myc-Level werden durch die Partnerproteine
kompetitiv an ihrer Wirkung gehindert. In einigen Tumorarten konnte man
dagegen eine gesteigerte, deregulierte Protein-Aktivität von c-myc feststellen,
die von äußeren Signalen entkoppelt ist (Bernhard und Eilers, 2006).
C-myc hat Einfluss auf Transformation, Mitogenese, Apoptose und Förderung
der Differenzierung. C-myc wird im Zellzyklus für den Übergang von der G1 in
die S-Phase benötigt, es verstärkt die Proliferation von Übergangszellen
(„transitional cells“) (Wang et al., 2002).
Abbildung 5: Abbildung des Zellzyklus und der physiologischen Zellhomöostase (Alberts et al., 2004)

1 Einleitung
15
In gesundem Gewebe und bei unbeeinträchtigter Wundheilung ist c-myc
während der frühen Phase nicht detektierbar (Stojadinovic et al., 2005; Wang et
al., 2002), in aktiv proliferierenden Zellen ist c-myc jedoch in höherem Maße zu
finden als in ausdifferenzierten Geweben ohne Proliferationskapazität (Wang et
al., 2002). C-myc kann analog zu ß-Catenin den Heilungsprozess stören
(Stojadinovic et al., 2005). So ist in chronischen epidermalen Wunden eine
erhöhte Konzentration der beiden Modulatoren vorzufinden. Über eine
Aktivierung des Zellzyklus kommt es hier, wie schon für ß-Catenin beschrieben,
zu einer Erschöpfung der Zellen im Wundgebiet.
Repressionen der c-myc-Expression resultierten in profunden Änderungen der
Zelladhäsion und der Zellmotilität in Verbindung mit einer geringeren
Expression von ECM- und Adhäsionsmolekülen (Frye et al., 2003). Dies wirkt
sich sowohl auf das Apoptoseverhalten von Zellen als auch auf die
Wundheilung aus. Auch für c-myc sind in der Literatur kontroverse Effekte
dokumentiert, so daß Expression und Auswirkung jeweils situativ zu bewerten
sind.
1 Einleitung
16
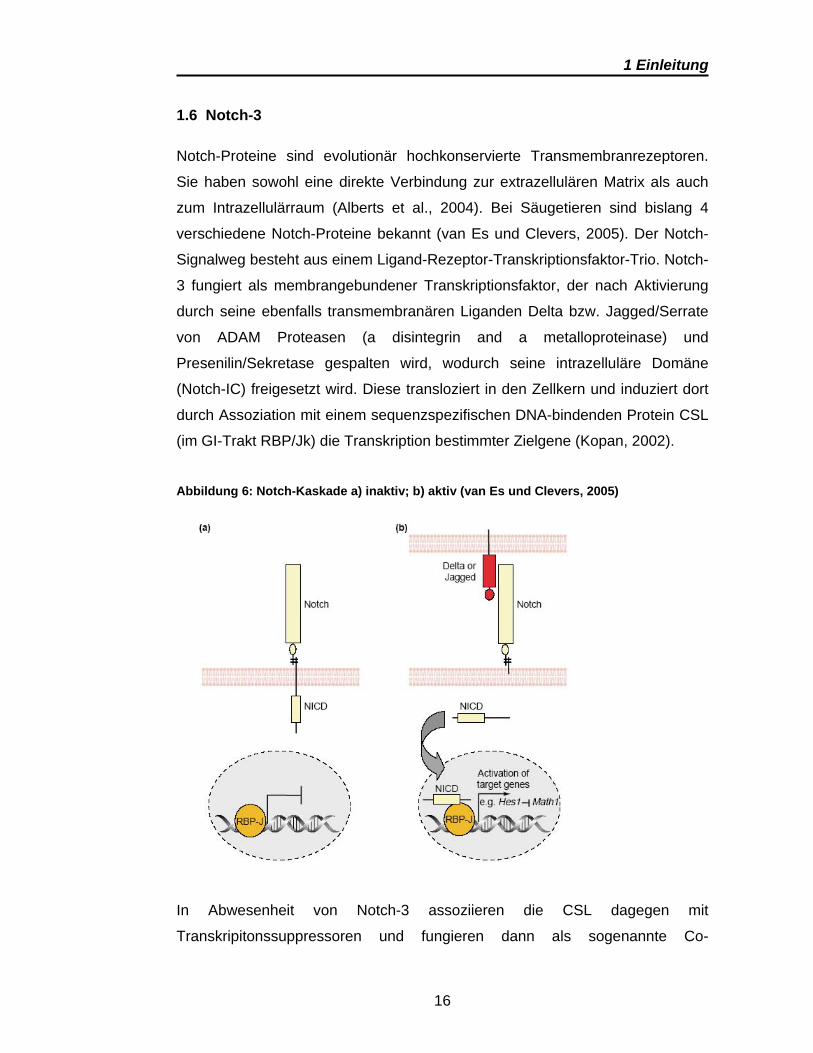
1.6 Notch-3
Notch-Proteine sind evolutionär hochkonservierte Transmembranrezeptoren.
Sie haben sowohl eine direkte Verbindung zur extrazellulären Matrix als auch
zum Intrazellulärraum (Alberts et al., 2004). Bei Säugetieren sind bislang 4
verschiedene Notch-Proteine bekannt (van Es und Clevers, 2005). Der Notch-
Signalweg besteht aus einem Ligand-Rezeptor-Transkriptionsfaktor-Trio. Notch-
3 fungiert als membrangebundener Transkriptionsfaktor, der nach Aktivierung
durch seine ebenfalls transmembranären Liganden Delta bzw. Jagged/Serrate
von ADAM Proteasen (a disintegrin and a metalloproteinase) und
Presenilin/Sekretase gespalten wird, wodurch seine intrazelluläre Domäne
(Notch-IC) freigesetzt wird. Diese transloziert in den Zellkern und induziert dort
durch Assoziation mit einem sequenzspezifischen DNA-bindenden Protein CSL
(im GI-Trakt RBP/Jk) die Transkription bestimmter Zielgene (Kopan, 2002).
Abbildung 6: Notch-Kaskade a) inaktiv; b) aktiv (van Es und Clevers, 2005)
In Abwesenheit von Notch-3 assoziieren die CSL dagegen mit
Transkripitonssuppressoren und fungieren dann als sogenannte Co-

1 Einleitung
17
Suppressoren (Lai, 2004). Der Notch-Signalweg gehört zu den
Schlüsselsignalen der intestinalen Zellerneuerung (Ishizuya-Oka, 2005). Die
muköse Homöostase beinhaltet eine ständige zelluläre Proliferation,
Differenzierung und Migration von der Basalmembran zur Oberfläche. In
proliferierenden Zellen findet man das nukleäre Notch stark hochreguliert
(Noggle et al., 2006).
Im Intestinum erfüllt der Notch-Signalweg über seinen Liganden Delta
außerdem die Aufgabe der lateralen Inhibition (Alberts et al., 2004). Der
Mechanismus der lateralen Hemmung ist für die Bildung von Grenzen zwischen
zwei Geweben oder Zelltypen notwendig (Lodish et al., 2008). Ein Signal von
Delta wirkt hemmend auf eine Empfängerzelle. Diese kann sich nicht
weiterentwickeln und ist nicht mehr dazu in der Lage, selbst das hemmende
Delta-Signal zu erzeugen. Diese kompetitive Situation führt schließlich zu einer
„Mutterzelle“, die selber keine inhibierenden Signale mehr erhält (Alberts et al.,
2004). So beeinflussen Notch-3 und Delta mit ihrer Interaktion die Entwicklung
endokriner Darmzellen. Tierversuche an Mäusen haben ergeben, dass
Mutationen, die eine Notch-Signalisierung hemmen, die Bildung endokriner
Darmzellen steigern (Alberts et al., 2004).
Abbildung 7: Notch-3 in der intestinalen Zellerneuerung (Alberts et al., 2004)

1 Einleitung
18
Notch-3 ist hauptsächlich an Gefäßen lokalisiert. Es ist in differenziertem
Gewebe nicht aktiv und kann erst durch kationische Chelatoren (z.B. EDTA)
aktiviert werden (Noggle et al, 2006; Mitsiadis et al, 2001). Bekannt ist die
Bedeutung von Notch-3 vor allem aus seiner Rolle in der Pathogenese der
Gefäßerkrankung CADASIL (autosomal dominant erbliche zerebrale
Arteriopathie, die zu subkortikalen Infarkten und Leukoenzephalopathien führt).
Mutationen von Rezeptoren und Liganden lösen bei Säugetieren bereits
während der Embryogenese durch Hämorrhagien den Tod aus.
Für verschiedene Karzinome konnte gezeigt werden, dass sie vermehrt Notch
sowie seine Liganden exprimieren und dass Notch hier über eine
Differenzierungsblockade sowie über die Inhibition von Apoptose das
Tumorwachstum fördert (Gray et al., 1999). Notch-3 führt ohne seinen Liganden
Delta zu exzessiven Mitosen der Zellen (Lodish et al., 2008).
Experimentell konnte nachgewiesen werden, dass Notch-3 mit dem Wnt-
Signalweg interagiert. Unter bestimmten Bedingungen kann Notch-3 diesen
sowohl supprimieren als auch aktivieren (Wang et al., 2007). Einerseits
stimuliert Notch-3-IC (intrazelluläre Domäne) in Verbindung mit einer
gesteigerten Phosphorylierung von ß-Catenin einen Wnt-unabhängigen TCF
(Wang et al., 2007). Hohe Notch-3-IC-Level können außerdem zur vermehrten
Expression von N-Cadherin führen und stabilisieren damit die Zellmembran
(Wang et al., 2007). Auf der anderen Seite ist Notch-3-IC dazu in der Lage,
TCFs zu supprimieren und damit den Wnt-Signalweg zu indirekt zu fördern.
ß-Catenin und Notch-3 dienen durch sich überlagernde und ergänzende Effekte
der Zellhomöostase.

1 Einleitung
19
1.7 Zink
1.7.1 Allgemeine Bedeutung
Zink ist als divalentes Kation ein essentielles Spurenelement und erfüllt im
Stoffwechsel spezifische Funktionen als Bestandteil oder Aktivator von mehr als
300 Enzymen des Protein-, Kohlenhydrat-, Fett- und
Nukleinsäurestoffwechsels, sowie als Bestandteil von Hormonen und
Rezeptoren, in der Insulinspeicherung und im Immunsystem (Rink und Gabriel,
2000; Haase und Rink, 2006). Nach Eisen ist Zink das häufigste Metallion des
Körpers (Coleman, 1992). 1932 konnte erstmals nachgewiesen werden, dass
die Carboanhydrase gebundenes Zink enthält, welches mit der katalytischen
Aktivität zusammenhängt. Damit war die Carboanhydrase das erste entdeckte
zinkhaltige Enzym. Auch bei den Matrix-Metalloproteinasen ist ein Zinkion im
aktiven Zentrum nachgewiesen und somit für den Um- und Abbau von
Geweben essentiell (Haase und Rink, 2006; Berg et al., 2003). Zink greift durch
die Beeinflussung dieser Enzyme steuernd in Differenzierung, Proliferation und
Zelltod ein.
Zink kann protektiv wirken, indem es die Zelle als Chelatbildner vor
chemischen, physikalischen oder immunologischen Einflüssen schützt.
Zinkmangel kann durch eine Aktivierung von Endonukleasen Apoptose
induzieren (Sunderman, 1995). Schon 1974 konnten Prasad und Oberleas
belegen, dass Zinkmangel die Aktivität der Thymidinkinase und der DNA-
Synthese in schnell regenerierendem Bindegewebe nachteilig beeinflusst
(Prasad und Oberleas, 1974). Später konnte von Prasad et al. eine signifikante
Beeinträchtigung des Zellzyklus beobachtet werden, welche sich im
Besonderen durch eine verminderte Progression in die S-Phase des Zellzyklus
ausdrückte (Prasad, 1996). Vor allem in stärker proliferierenden Geweben, und
insbesondere dem Immunsystem, ist Zink ein äußerst wichtiger Faktor (Krones
et al., 2005; Ibs und Rink, 2003; Rink und Gabriel, 2000). Die intestinale
Schleimhaut gehört zu den wachstumsaktiven Geweben, so dass Vermutungen
über eine besondere Bedeutung von Zink nahe liegen. Die Hauptspeicher des
Körpers für Zink sind das Skelett, sowie Muskel- und Fettgewebe. Diese
können einen Mangel für einige Wochen ohne Plasmaspiegelschwankungen

1 Einleitung
20
kompensieren (Windisch, 2003). Die Resorption findet hauptsächlich im
Dünndarm statt, wobei das Ileum die weitaus größte Kompetenz aufweist
(Antonson et al., 1979). Aber auch das Kolon spielt bei der Resorption von Zink
eine nicht unerhebliche Rolle. Es ist als zinksensitives Organ anzusehen,
welches dazu in der Lage ist, auf Zinkmangel mit der Expression von Zink-
Transporterproteinen zu reagieren (Pfaffl und Windisch, 2003). Auch bei der
Ausscheidung von Zink übernimmt der Darm die tragende Rolle. So enthalten
Pankreas- und Gallensaft sowie gastroduodenale Sekrete einen
vergleichsweise hohen Zinkgehalt, welcher bei einer Mangelsituation vollständig
rückresorbiert werden kann. Um diese physiologischen Abläufe der Resorption
experimentell zu verhindern, und um die körpereigenen Zinkvorräte zu leeren,
kann einer zinklosen Diät der Absorptionshemmer Phytat zugesetzt werden.
Phytat hemmt die Resorbierbarkeit von Zink durch die Bildung eines
schwerlöslichen Komplexes (Windisch und Kirchgessner, 1999). Weitere
Faktoren, welche die Zinkverwertung beeinflussen können, sind unter anderem
die Zusammensetzung der Nahrung (insbesondere Wechselwirkungen mit
anderen Nahrungsbestandteilen wie z.B. Calcium und Phytat),
Stresssituationen, chirurgische Eingriffe, parasitäre Erkrankungen und
Infektionen.
Ein akuter Zinkmangel ist selten. Begünstigende Situationen findet man
beispielsweise bei Malabsorptionssyndromen, parenteraler Ernährung,
Behandlung mit Chelatbildnern und großflächigen Verbrennungen vor. Ein
latenter, chronischer Zinkmangel ist allerdings auch in den westlichen Ländern,
gerade bei älteren Menschen, häufig anzutreffen (Prasad, 1996; Sandstead und
Smith, 1996).
Bei schwerem Zinkmangel treten Verminderungen der
Geschmacksempfindung, Appetitlosigkeit, Dermatitis, Haarausfall, Durchfall und
neuropsychische Störungen auf. Darüber hinaus sind Wachstumsstörungen,
Störungen der männlichen Sexualentwicklung und Reproduktionsfunktion, eine
erhöhte Infektanfälligkeit und Störungen der Wundheilung beschrieben. Ein
bekanntes Beispiel für Zinkmangel ist die autosomal rezessiv erbliche Krankheit
Acrodermatitis enteropathica. Hier handelt es sich um einen Defekt der
enteralen Zinkaufnahme, der sich in einer Thymusatrophie und einem für

1 Einleitung
21
diverse Infektionen sehr anfälligen Phänotyp manifestiert. Die Symptomatik und
die ohne Behandlung sehr kurze Lebenserwartung von wenigen Jahren kann
durch die pharmakologische Supplementierung von Zink komplett revidiert
werden (Haase und Rink, 2006).
1.7.2 Zink in der Wundheilung
Die Eigenschaften von Zink können sich auf die komplexe Regulation der
Wundheilung auswirken. Eine Dysregulation des Zinkhaushaltes kann
theoretisch positive oder negative Effekte auf die Regeneration haben. Zink
akkumuliert in experimentellen Wunden. Diese Anreicherung tritt direkt nach
einer Verletzung auf und erreicht zwischen dem 5. und 7. Tag ein Maximum.
Der Zinkgehalt normalisiert sich bis zum 15. postoperativen Tag, zu diesem
Zeitpunkt hat auch die Zugfestigkeit des heilenden Gewebes ihren Höhepunkt
erreicht. Aus diesen Ergebnissen lässt sich ableiten, dass der Heilungsprozess
in den ersten sieben Tagen ein erhöhtes Zinkniveau benötigt (Kaplan et al.,
2004; Oberleas et al, 1971). Dass in stark proliferierenden Geweben eine
höhere Zinkkonzentration vorzufinden ist, impliziert, dass Zink auch in der
Wundheilung intestinaler Gewebe eine besondere Stellung einnimmt (Haase
und Rink, 2006; Ibs und Rink, 2003). Diese Annahme wird durch Ergebnisse
von Vaxman et al. unterstützt, die den Zinkgehalt im Blut sowie im Kolon von
Kaninchen nach Anastomosierung untersuchten. Es ließ sich ein
Konzentrationsabfall im Blut und ein –anstieg in der Darmwand darstellen
(Vaxman et al., 1992). Andere Studien sprechen Zink eine besondere Stellung
während der Reifungsphase, also in späteren Phasen der Wundheilung, zu.
Hier wird Zink eine Funktion als Effektor der Proteinsynthese und somit eine
notwendige Heilungsvoraussetzung zugesprochen (Senapati und Thompson,
1985).
Die Effizienz einer Behandlung zinkhaltigen Zusätzen (in der lokalen
Applikation) konnte mit einer geringeren Entzündungsreaktion sowie einer
gesteigerten Mitoserate und einem verminderten Bakterienwachstum auch bei
nicht zinkdefizitären Patienten belegt werden (Agren, 1990). In der topischen

1 Einleitung
22
Anwendung bei oberflächlichen Wunden und Ulzera sind zinkhaltige Salben
bereits etabliert.
Zink ist Bestandteil jeder Matrixmetalloproteinase und damit auch notwendiger
Bestandteil der ECM, deren Stoffwechsel wichtige Einflussgröße auf die
Qualität der Wundheilung ist.

2 Zielsetzung
23
2 Zielsetzung
Trotz weitreichender Fortschritte in Technik und Durchführung der
Darmchirurgie ist die intestinale Wundheilung weiterhin ein zentrales Thema
chirurgischer Forschung. Anastomoseninsuffizienzen stellen immer noch den
größten Teil der lebensbedrohlichen Komplikationen bei diesen Eingriffen dar.
Verschiedene Arbeiten diskutieren bereits prädisponierende Faktoren. Zu
diesen zählt auch der Einfluss von Zink bzw. einem Zinkmangel. Insgesamt gibt
es derzeit jedoch erst einzelne Untersuchungen zu dem komplexen
Themengebiet der intestinalen Wundheilung, welche die Problematik noch nicht
in ihrer Gesamtheit klären.
Im Focus dieser Arbeit steht deshalb das Verhalten der ECM bzw. ihrer
Wechselspieler während der frühen postoperativen Phase. esonders
betrachtet werden dabei die vier Komponenten TIMP-1, ß-Catenin, c-myc und
Notch-3 und ihr Verhalten unter einem kontrollierten Zinkmangel. Unter diesen
Bedingungen werden auch die Einflüsse von Nahttechnik und Lokalisation
geprüft.
Das Ziel der Arbeit ist die Klärung folgender Fragen:
1) Verändert sich das Expressionsmuster der untersuchten Variablen
während der Wundheilung im Zinkmangel?
2) Wie greift Zink in die Funktionalität der untersuchten Faktoren ein?
3) Beeinflusst die Schnittführung – Kolotomie versus zirkuläre
Kolonanastomose – dabei die Wundheilung?

3 Material und Methoden
24
3 Material und Methoden
3.1 Tiermodell
Die Tierversuche zu dieser Studie wurden in Übereinstimmung mit den
nationalen Vorgaben des Tierschutzgesetzes und den internationalen
Richtlinien der Versuchstierhaltung durchgeführt.
Als Versuchstiere wurden 40 männliche Wistar Ratten mit einem
Ausgangsgewicht von 220 – 240 g eingesetzt. Von den 40 eingesetzten
Versuchstieren starben zwei bereits präoperativ im Zinkmangel an einer
Pneumonie. Bei drei weiteren Tieren kam es perioperativ zum Exitus. Nach
Randomisierung wurden die Tiere 4 Gruppen zugeteilt. Die Gruppen 1 (n = 7)
und 2 (n = 12) bildeten die Kontrollgruppen, während bei den Tieren aus
Gruppe 3 (n = 8) und Gruppe 4 (n = 8) ein kontrollierter Zinkmangel induziert
wurde. Alle Tiere wurden hierzu über 21 Tage in Einzelhaltung kontrolliert
ernährt. Das Futter wurde für alle Gruppen vom gleichen Hersteller bezogen
und unterschied sich in der Zusammensetzung nur im Zinkgehalt. Jedes Tier
erhielt in den ersten 14 Tagen 8 g Futter pro Tag. Diese Ration wurde in den
folgenden 7 Tagen auf 12 g pro Tag erhöht.
Vorarbeiten von Windisch zur Induktion von Zinkmangel in der adulten Ratte
entsprechend (Windisch, 2003), lag der Zinkgehalt der zinkarmen Diät bei 5,9
mg/KG (C1040 zinkarm, Altromin, Lage, Deutschland) und in der
Kontrollgruppe bei 58,4 mg/KG (C1040 mod, Altromin, Lage, Deutschland).
Nach 21 Tagen kontrollierter Ernährung wurde der in den Gruppen 3 (n = 8)
und 4 (n = 8) induzierte Zinkmangel durch Haaranalysen mittels
Atomabsorptionsspektralphotometrie (AAS) geprüft (Spectrophotometer
Quantum-2, Cortec, Moskau, Russland). Der Zinkgehalt in den Proben der
Gruppen 3 und 4 lag dabei signifikant (p < 0,05) unterhalb der Werte aus den
Kontroll-Gruppen 1 (n = 7) und 2 (n = 12).
Nach der Bestimmung des Zinkgehaltes wurden die Versuchstiere
unterschiedlichen Operationen am Kolon unterzogen. Bei den Gruppen 1
(Kontrolle) und 3 (Zinkmangel) wurden zirkuläre Kolonanastomosen und bei
den Gruppen 2 (Kontrolle) und 4 (Zinkmangel) Kolotomien mit anschließendem

3 Material und Methoden
25
Nahtverschluss durchgeführt. Alle Maßnahmen erfolgten unter sterilen
Bedingungen. Die Einleitung der Narkose erfolgte mit Isofluran und wurde über
eine subkutane Injektion von 0,3 ml/kg Medetomidin und 100 mg/kg
Ketaminhydrochlorid aufrechterhalten. Nach medianer Laparotomie wurde das
Kolon zirkulär oder antimesenterial längs eröffnet und in direktem Anschluss in
allschichtiger, einreihiger Einzelknopfnaht mit 5-0 multifilen Polyclagtin 910-
Fäden (Vicryl, Ethicon Products, Norderstedt, Deutschland) wieder
verschlossen. Postoperativ wurde die gewohnte Fütterung beibehalten. Am 7.
postoperativen Tag wurden die Tiere getötet, gewogen und die operierten
Darmschlingen explantiert.
Nach Kolonanastomose kam es sowohl in der Kontroll- und der
Zinkmangelgruppe zu je einer Insuffizienz. In der Kontrollgruppe kam es auch
nach Kolotomie zu einer Nahtundichtigkeit. Alle Insuffizienzen führten über
Peritonitis und/oder Ileus zum Tod. Bei numerischer Übereinstimmung wurde
auf eine Signifikanzprüfung verzichtet.
3.2 Histologie
Die histologische Aufarbeitung aller explantierten Anastomosen und Darmnähte
erfolgte primär über eine 24-stündige Fixation in 10 % Formalin. Darauf folgte
die Dehydrierung und Paraffineinbettung. Anschließend wurden die Proben in
10-15, 3-5 m dicke Stufenschnitte unterteilt. Zur histologischen Auswertung
wurden die Stufenschnitte mit Hämatoxylin & Eosin (H&E) gefärbt. Prinzipiell
wurden für alle weiteren Untersuchungen pro Tier Blöcke von je zwei
Lokalisationen angefertigt:
1. Anastomose, Kolotomie
2. anastomosen-/Kolotomiefern

3 Material und Methoden
26
3.3 Immunhistochemische Aufarbeitung
Bei den durchgeführten immunhistologischen Färbungen kamen käuflich
erwerbbare Antikörper unterschiedlicher Firmen zum Einsatz. Das in zehn
prozentig gepuffertem Formalin fixierte Gewebe wurde zunächst
zurechtgeschnitten, im Gewebeentwässerungsautomat (V.I.P E 150, Firma
Vogel, Gießen, Deutschland) entwässert und nachfolgend in Paraffin
eingebettet (Histoembedder, Firma Jung, Heidelberg, Deutschland). Mit Hilfe
eines Schlittenmikrotoms (Firma Jung, Heidelberg, Deutschland) erfolgte die
Herstellung von etwa 3 µm dicken Schnitten. Diese wurden anschließend bei
37°C im Brutschrank über Nacht entparaffiniert. Danach erfolgten die
unterschiedlichen immunhistologischen Färbungen.
Die Aufbereitung der Proben erfolgte nach der Avidin-Biotin Methode. Zunächst
wurde ein Primärantikörper verwendet, der jeweils gegen das zu detektierende
Antigen gerichtet war. Anschließend diente ein biotinylierter Sekundärantikörper
dazu, die gebundenen Primärantikörper zu markieren. Danach erfolgte die
Zugabe des Avidin-Biotin-Enzymkomplexes, welcher mit dem biotinylierten
Sekundärantikörper reagiert. Als Substrat zur Farbentwicklung diente DAB (3,3
Diaminobenzidine tablets, Firma Sigma, Steinheim, Deutschland). Das
Verfahren ist im Folgenden genauer beschrieben.
Nach einer Inkubation von 60 min mit einem in zwei prozentigem Milchpulver
verdünnten Normalserum wurde der jeweilige Primärantikörper hinzu pipettiert
und über Nacht bei 4°C im Kühlschrank inkubiert. Am nächsten Tag wurden die
Proben auf Raumtemperatur gebracht, mit PBS (Phosphate Buffered Saline)
gespült und dann mit dem Sekundärantikörper versehen. Nach 30-minütiger
Inkubation bei Raumtemperatur wurde nach erneuter Spülung der ABC-
Komplex hinzugefügt und eine halbe Stunde bei Raumtemperatur inkubiert.
Nach einer weiteren Spülung mit PBS erfolgte die Zugabe von aktiviertem DAB.
Eine mehrfache Waschung mit Wasser entfernte die Reste der Lösung. Zum
Abschluss wurden die Schnitte nun noch für 18 sec in Hämalaunlösung (Firma
Merck, Darmstadt, Deutschland) eingestellt, erneut gespült und durch eine
aufsteigende Alkoholreihe gezogen.
3 Material und Methoden
27
3.4 Färbungen
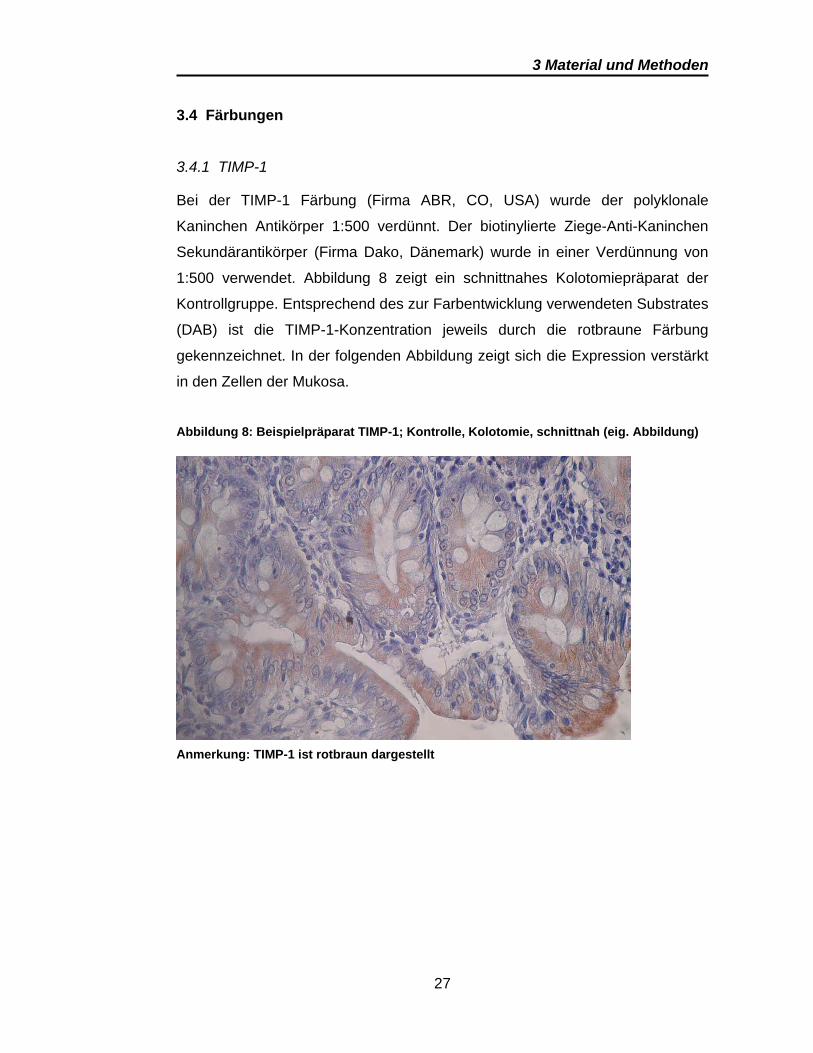
3.4.1 TIMP-1
Bei der TIMP-1 Färbung (Firma ABR, CO, USA) wurde der polyklonale
Kaninchen Antikörper 1:500 verdünnt. Der biotinylierte Ziege-Anti-Kaninchen
Sekundärantikörper (Firma Dako, Dänemark) wurde in einer Verdünnung von
1:500 verwendet. Abbildung 8 zeigt ein schnittnahes Kolotomiepräparat der
Kontrollgruppe. Entsprechend des zur Farbentwicklung verwendeten Substrates
(DAB) ist die TIMP-1-Konzentration jeweils durch die rotbraune Färbung
gekennzeichnet. In der folgenden Abbildung zeigt sich die Expression verstärkt
in den Zellen der Mukosa.
Abbildung 8: Beispielpräparat TIMP-1; Kontrolle, Kolotomie, schnittnah (eig. Abbildung)
Anmerkung: TIMP-1 ist rotbraun dargestellt

3 Material und Methoden
28
3.4.2 ß-Catenin
Bei der ß-Catenin Färbung (Firma Abcam, Cambridge, UK) wurde der
monoklonale Maus Antikörper 1:500 verdünnt. Die Vorbehandlung erfolgte in
der Mikrowelle bei 600 Watt mit Citratpuffer (pH 6) für zwei mal zehn Minuten.
Der biotinylierte Kaninchen-Anti-Maus Sekundärantikörper (Firma Dako,
Dänemark) wurde in einer Verdünnung von 1:300 verwendet. Abbildung 9 zeigt
ein schnittfernes Anastomosenpräparat der Kontrollgruppe. Entsprechend des
zur Farbentwicklung verwendeten Substrates (DAB) ist die ß-Catenin-
Konzentration jeweils durch die rotbraune Färbung gekennzeichnet. Diese zeigt
sich in dem hier dargestellten Präparat am deutlichsten in den junktionalen
Abschnitten der Mukosa.
Abbildung 9: Beispielpräparat ß-Catenin; Kontrolle, Anastomose, schnittfern (eig. Abbildung)
Anmerkung: ß-Catenin ist rotbraun dargestellt

3 Material und Methoden
29
3.4.3 c-myc
Bei der c-myc Färbung (Firma Santa Cruz, CA, USA) wurde der polyklonale
Kaninchen Antikörper 1:100 verdünnt. Der biotinylierte Ziege-Anti-Kaninchen
Sekundärantikörper (Firma Dako, Dänemark) wurde in einer Verdünnung von
1:500 verwendet. Abbildung 10 zeigt ein schnittnahes Anastomosenpräparat
der Kontrollgruppe. Entsprechend des zur Farbentwicklung verwendeten
Substrates (DAB) ist die c-myc-Konzentration jeweils durch die rotbraune
Färbung gekennzeichnet. In dem hier abgebildeten Präparat zeigt sich diese
verstärkt im Zytoplasma der Mukosazellen.
Abbildung 10: Beispielpräparat c-myc; Kontrolle, Anastomose, schnittnah (eig. Abbildung)
Anmerkung: c-myc ist rotbraun dargestellt

3 Material und Methoden
30
3.4.4 Notch-3
Der Notch-3-Antikörper (Firma Santa Cruz, CA, USA) ist ein polyklonaler
Kaninchen-Antikörper. Der Primärantikörper wurde in einer Verdünnung 1: 100
verwendet. Die Vorbehandlung erfolgte in der Mikrowelle bei 600 Watt mit
Citratpuffer (pH 6). Als Sekundärantikörper diente ein biotinylierter Ziege- Anti-
Kaninchen Antikörper (Firma Dako, Dänemark), der in einer Verdünnung von
1:500 verwendet wurde. Abbildung 11 zeigt ein schnittfernes
Anastomosenpräparat der Zinkmangelgruppe. Entsprechend des zur
Farbentwicklung verwendeten Substrates (DAB) ist die Notch-3-Konzentration
jeweils durch die rotbraune Färbung gekennzeichnet. Das abgebildete Präparat
zeigt eine relativ homogene Expression in Mukosa und Submukosa mit
Betonung der Zellkerne.
Abbildung 11: Notch-3 Präparat; Zinkmangel, Anastomose, schnittfern (eig. Abbildung)
Anmerkung: Notch-3 ist rotbraun dargestellt

3 Material und Methoden
31
3.5 Auswertung
Alle Schnittpräparate wurden über eine auf dem Mikroskop installierte
Digitalkamera (Olympus C-3030, Olympus, Hamburg, Deutschland) erfasst.
Jedes Bild wurde in 400facher Vergrößerung angefertigt und mittels der
Software Olympus DP-Soft 3.1 (Olympus, Hamburg, Deutschland) archiviert.
Es wurden für alle untersuchten Variablen (TIMP-1, c-myc, ß-Catenin und
Notch-3) zwölf Bilder der Mukosa (je sechs von Zotten und Krypten) und sechs
Bilder der Submukosa anastomosenfern bzw. schnittfern sowie zwölf Bilder der
Mukosa (je sechs von Zotten und Krypten) und sechs Bilder der Submukosa
anastomosen-/schnittnah mit Hilfe des Remmele-Scores bewertet. So konnten
insgesamt Unterschiede der Faktorexpression in Abhängigkeit vom
Zinkhaushalt und der Operationstechnik beurteilt werden. Die folgende Grafik
zeigt die Beziehungen zwischen den einzelnen gestesteten Regionen dar.
Abbildung 12: Schaubild der Untersuchungsebenen (eigene Darstellung)

3 Material und Methoden
32
3.5.1 Remmele-Score
Beurteilt wurden die Präparate nach dem Vorbild des semi-quantitativen
Remmele-Scores, der folgendermaßen modifiziert wurde (Remmele und
Stegner, 1987).
Der Bewertung der Färbeintensität liegt ein vierstufiger Score zugrunde:
1= keine Färbereaktion
2= schwache Färbereaktion
3= mäßige Färbereaktion
4= starke Färbereaktion
Die Auswertung der Anzahl der positiven Zellen erfolgte mittels eines
fünfstufigen Scores:
1= keine positive Zelle
2= <10% positive Zellen
3= 10-50% positive Zellen
4= 51-80% positive Zellen
5= >80% positive Zellen
Die Multiplikation beider Scores ergab den Gesamtscore auf einer Skala von 1-
20. Eine zehnprozentige Rate von Zellen mit erkennbarer
immunhistochemischer Reaktion wurde dabei als Schwellenwert einer
relevanten Reaktion definiert.

3 Material und Methoden
33
3.5.2 Statistik
Die statistische Analyse wurde mit dem Statistical Package for Social Sciences
Version 10.0 (SPSS®) durchgeführt. Die Insuffizienzraten der verschiedenen
Gruppen (Kolotomie vs. Kolonanastomose, Zinkmangel-Gruppe vs.
Kontrollgruppe) wurden aufgrund der numerischen Übereinstimmung nicht mit
dem Chi-Quadrat-Test auf ihre Unabhängigkeit geprüft.
Die Auswertung der Expression der einzelnen Faktoren – TIMP-1, ß-Catenin, c-
myc und Notch-3 – erfolgte mittels der zugeordneten Scores Die Expression
wurde bezüglich ihrer Beeinflussbarkeit durch die Untersuchungsgruppe
(Kontrolle vs. Zinkmangel) analysiert. Zur Überprüfung der
Gruppenunterschiede auf ihre Signifikanz kam der Mann-Whitney-Wilcoxon-
Test (U-Test) zum Einsatz. Dieser Test wurde gewählt, da ordinalskalierte
Variablen (Scores) zur Beurteilung der Merkmale vergeben wurden. Aufgrund
der mehrfachen Aufteilung des Versuchsaufbaus wurden paarweise Vergleiche
für nicht parametrische Daten durchgeführt. Im Sinne einer explorativen
Analyse wurde ein p-Wert kleiner oder gleich 0,05 als Indikator einer lokalen
statistischen Signifikanz betrachtet. Die Boxplots stellen die Ergebnisse
graphisch dar. Der Boxplot dient der Darstellung der Merkmalsverteilung
beruhend auf Quartilen. Der Kasten stellt dabei den Interquartilsabstand dar,
die davon ausgehenden Linien führen zum Minimum und zum Maximum
(maximal das 1,5-fache des Interquartilsabstandes), die waagerechte Linie gibt
den Median wieder, Ausreißer sind in Punktform dargestellt.

4 Ergebnisse
34
4 Ergebnisse
Zwei Tiere starben präoperativ im Rahmen des Zinkmangels an einer
Pneumonie. Postoperativ traten insgesamt drei Nahtinsuffizienzen auf. In der
Kontrollgruppe trat bei Anastomosen und Kolotomien jeweils eine Insuffizienz
auf, in der Zinkmangelgruppe kam es zu einer Anastomoseninsuffizienz. Alle
Insuffizienzen führten zum Tod. Aufgrund der geringen Zahl wurde auf eine
Signifikanzprüfung verzichtet. Abbildung 12 stellt den allgemeinen
Versuchsaufbau dar und zeigt die untersuchten Einflussfaktoren im
Zusammenhang. Wie abgebildet, erfolgt innerhalb der Gruppen eine
Aufspaltung unter Berücksichtigung der untersuchten Variablen. Variabel ist
jeweils die Gruppe (Kontrolle – Zinkmangel), die Nahttechnik (Anastomose –
Kolotomie), die Lokalisation (Schnittnah – Schnittfern) und die Region (Mukosa
– Submukosa). Die vier untersuchten Faktoren werden nacheinander in
separaten Kapiteln abgehandelt.
Signifikante Ergebnisse sind in den nachfolgenden Tabellen mit Sternchen
gekennzeichnet. Es werden ausschließlich die signifikanten Ergebnisse in
Boxplots dargestellt.
4 Ergebnisse
35
4.1 TIMP-1
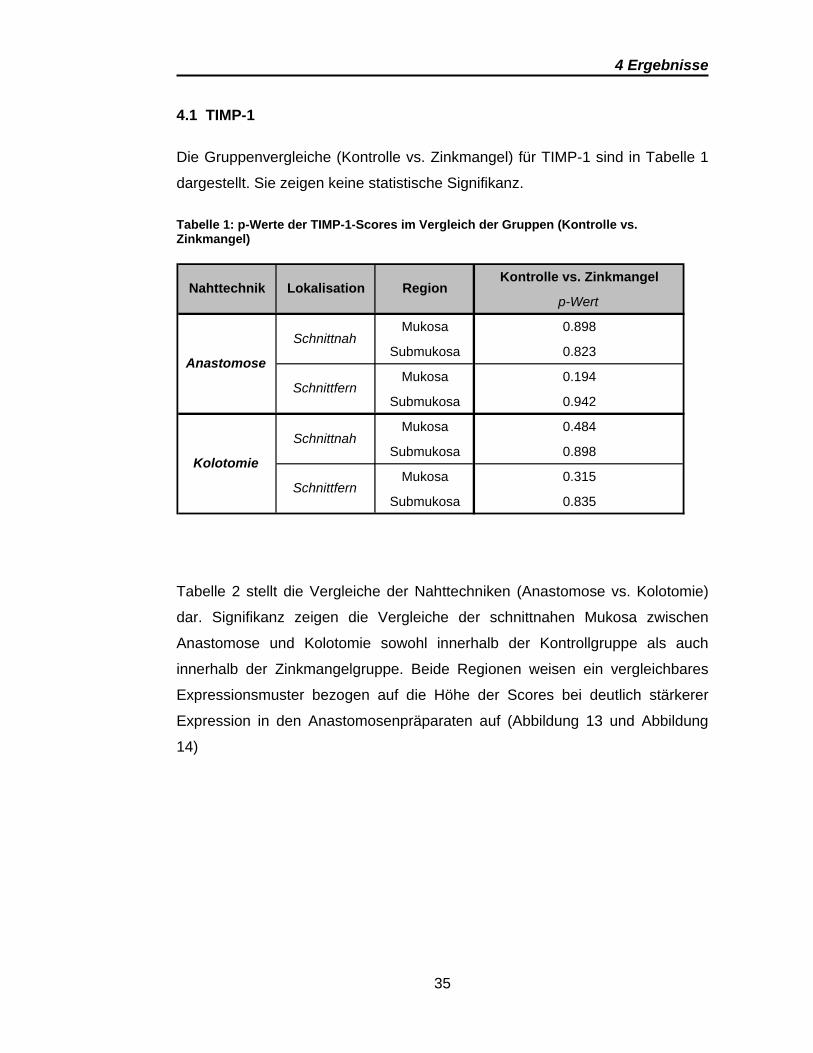
Die Gruppenvergleiche (Kontrolle vs. Zinkmangel) für TIMP-1 sind in Tabelle 1
dargestellt. Sie zeigen keine statistische Signifikanz.
Tabelle 1: p-Werte der TIMP-1-Scores im Vergleich der Gruppen (Kontrolle vs. Zinkmangel)
Kontrolle vs. Zinkmangel
p-Wert
Mukosa 0.898
Submukosa 0.823
Mukosa 0.194
Submukosa 0.942
Mukosa 0.484
Submukosa 0.898
Mukosa 0.315
Submukosa 0.835
Region
Anastomose
Kolotomie
Schnittnah
Schnittfern
Nahttechnik Lokalisation
Schnittnah
Schnittfern
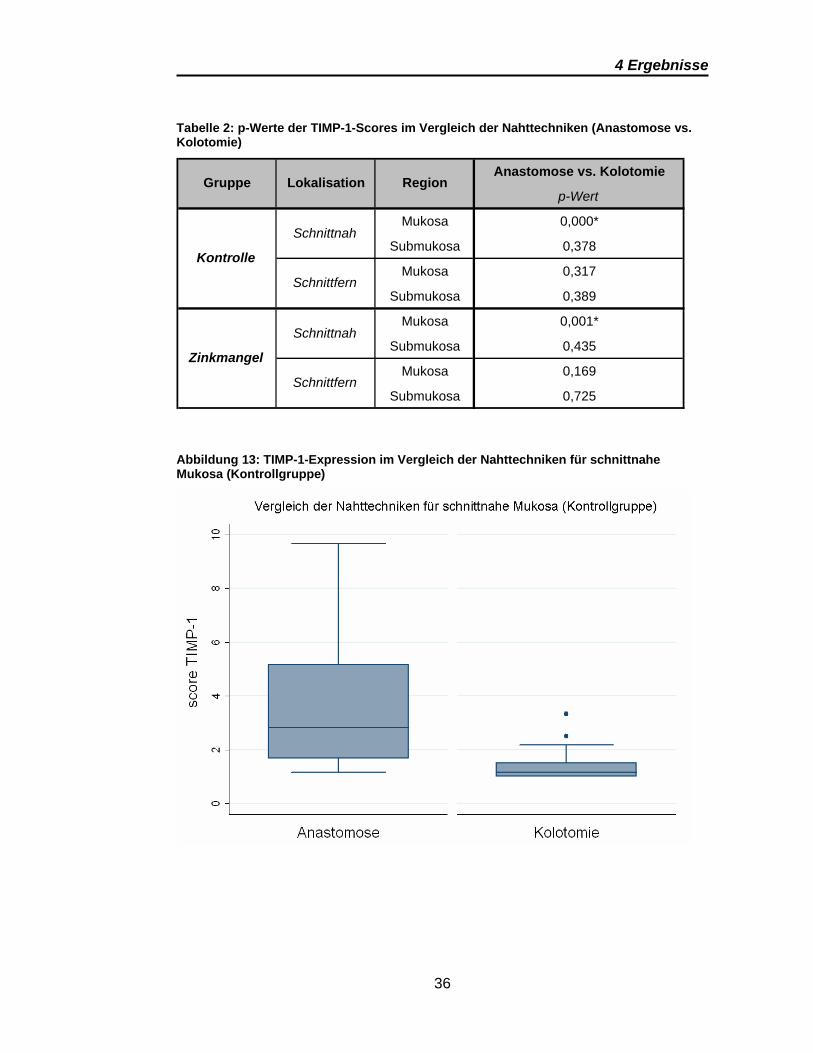
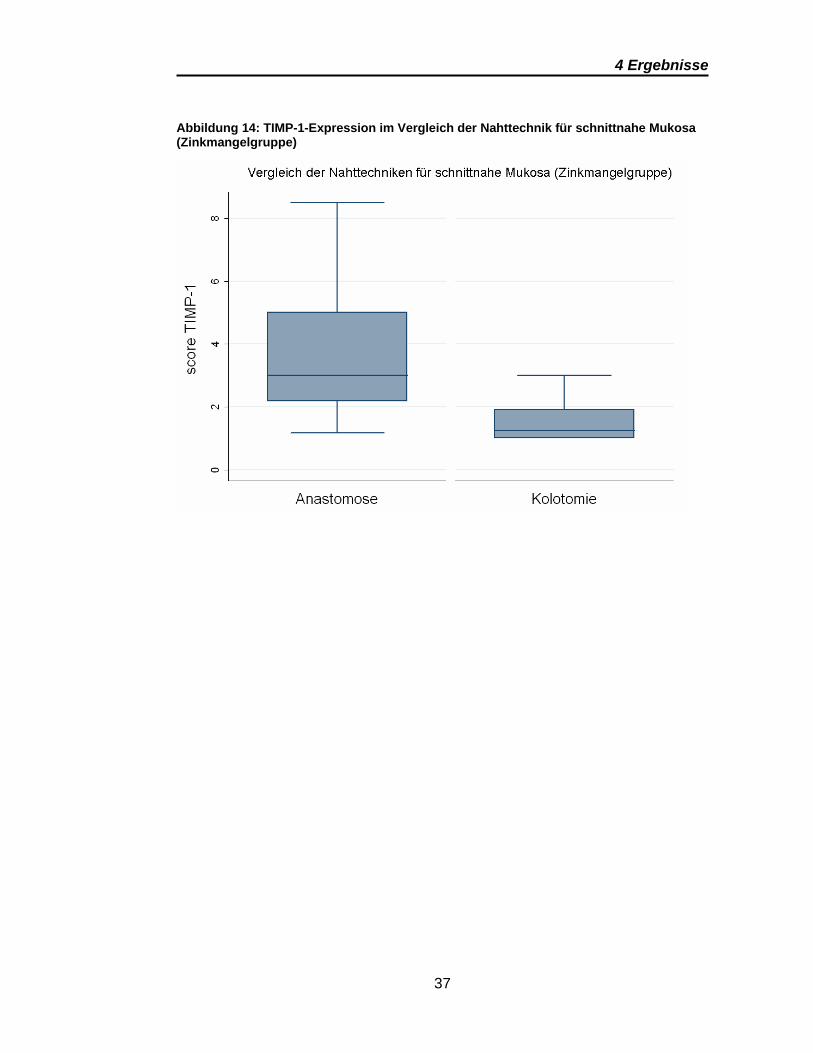
Tabelle 2 stellt die Vergleiche der Nahttechniken (Anastomose vs. Kolotomie)
dar. Signifikanz zeigen die Vergleiche der schnittnahen Mukosa zwischen
Anastomose und Kolotomie sowohl innerhalb der Kontrollgruppe als auch
innerhalb der Zinkmangelgruppe. Beide Regionen weisen ein vergleichbares
Expressionsmuster bezogen auf die Höhe der Scores bei deutlich stärkerer
Expression in den Anastomosenpräparaten auf (Abbildung 13 und Abbildung
14)
4 Ergebnisse
36
Tabelle 2: p-Werte der TIMP-1-Scores im Vergleich der Nahttechniken (Anastomose vs. Kolotomie)
Anastomose vs. Kolotomie
p-Wert
Mukosa 0,000*
Submukosa 0,378
Mukosa 0,317
Submukosa 0,389
Mukosa 0,001*
Submukosa 0,435
Mukosa 0,169
Submukosa 0,725
Region
Kontrolle
Zinkmangel
Schnittnah
Schnittfern
Gruppe Lokalisation
Schnittnah
Schnittfern
Abbildung 13: TIMP-1-Expression im Vergleich der Nahttechniken für schnittnahe Mukosa (Kontrollgruppe)
4 Ergebnisse
37
Abbildung 14: TIMP-1-Expression im Vergleich der Nahttechnik für schnittnahe Mukosa (Zinkmangelgruppe)
4 Ergebnisse
38
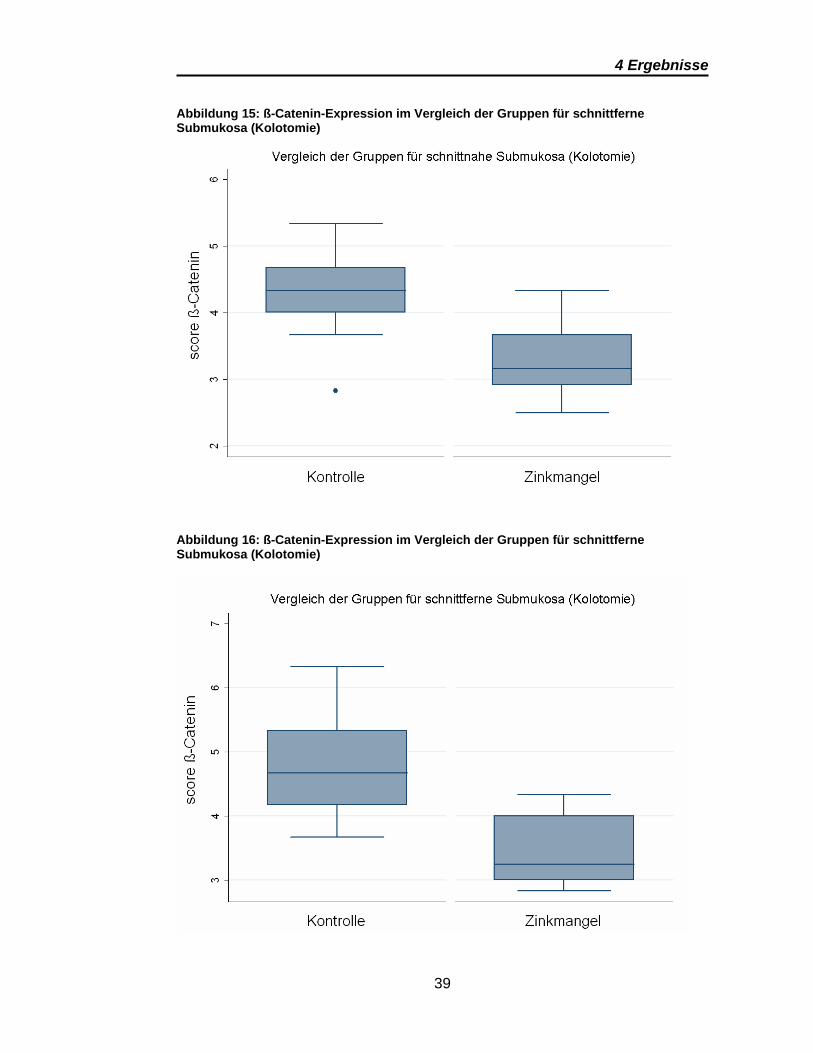
4.2 ß-Catenin
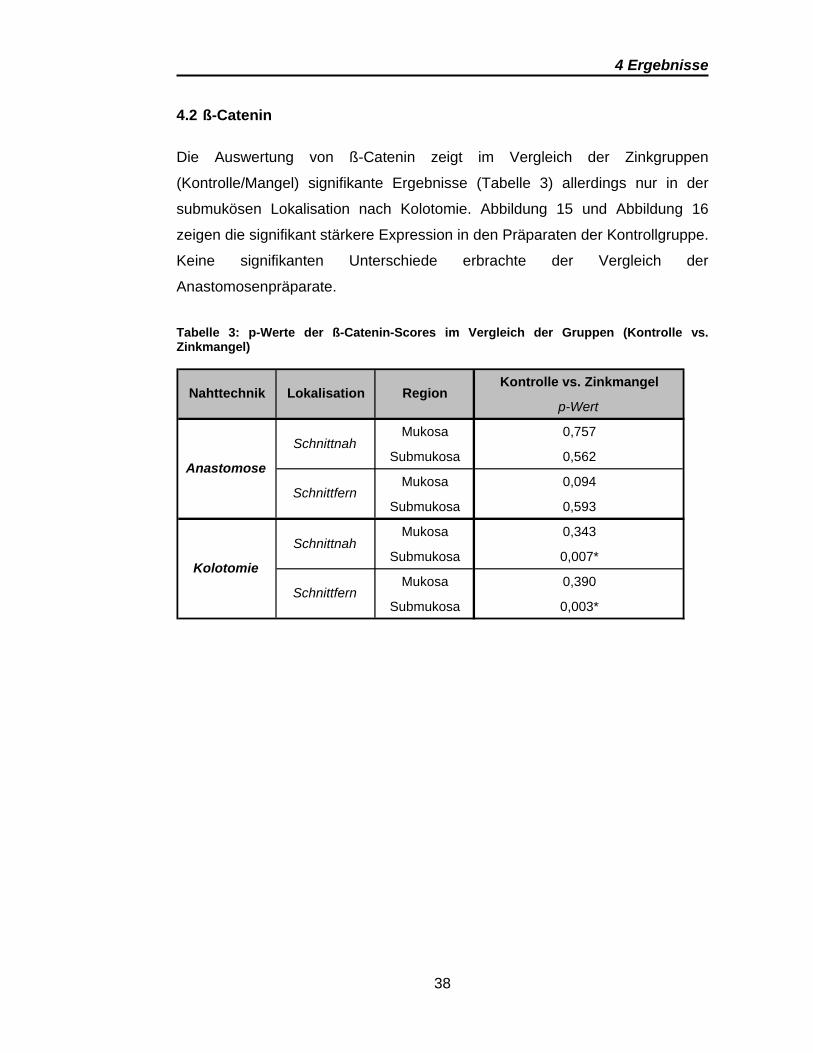
Die Auswertung von ß-Catenin zeigt im Vergleich der Zinkgruppen
(Kontrolle/Mangel) signifikante Ergebnisse (Tabelle 3) allerdings nur in der
submukösen Lokalisation nach Kolotomie. Abbildung 15 und Abbildung 16
zeigen die signifikant stärkere Expression in den Präparaten der Kontrollgruppe.
Keine signifikanten Unterschiede erbrachte der Vergleich der
Anastomosenpräparate.
Tabelle 3: p-Werte der ß-Catenin-Scores im Vergleich der Gruppen (Kontrolle vs. Zinkmangel)
Kontrolle vs. Zinkmangel
p-Wert
Mukosa 0,757
Submukosa 0,562
Mukosa 0,094
Submukosa 0,593
Mukosa 0,343
Submukosa 0,007*
Mukosa 0,390
Submukosa 0,003*
Region
Anastomose
Kolotomie
Schnittnah
Schnittfern
Nahttechnik Lokalisation
Schnittnah
Schnittfern
4 Ergebnisse
39
Abbildung 15: ß-Catenin-Expression im Vergleich der Gruppen für schnittferne Submukosa (Kolotomie)
Abbildung 16: ß-Catenin-Expression im Vergleich der Gruppen für schnittferne Submukosa (Kolotomie)
4 Ergebnisse
40
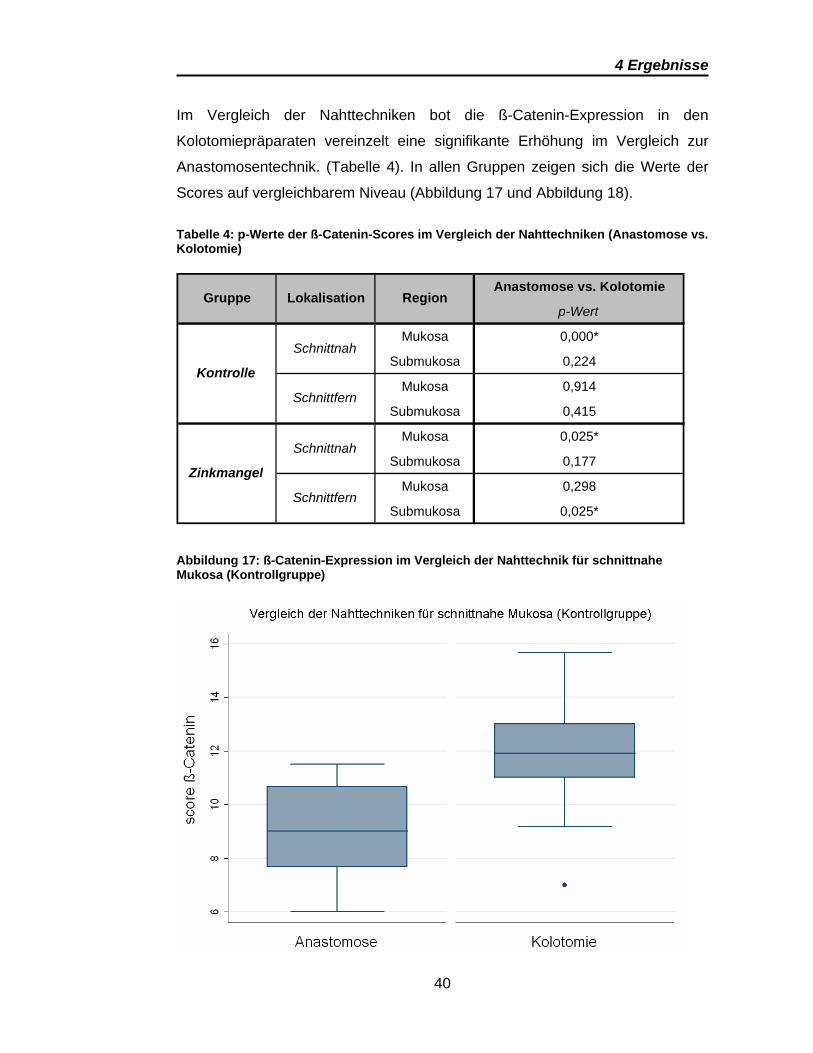
Im Vergleich der Nahttechniken bot die ß-Catenin-Expression in den
Kolotomiepräparaten vereinzelt eine signifikante Erhöhung im Vergleich zur
Anastomosentechnik. (Tabelle 4). In allen Gruppen zeigen sich die Werte der
Scores auf vergleichbarem Niveau (Abbildung 17 und Abbildung 18).
Tabelle 4: p-Werte der ß-Catenin-Scores im Vergleich der Nahttechniken (Anastomose vs. Kolotomie)
Anastomose vs. Kolotomie
p-Wert
Mukosa 0,000*
Submukosa 0,224
Mukosa 0,914
Submukosa 0,415
Mukosa 0,025*
Submukosa 0,177
Mukosa 0,298
Submukosa 0,025*
Region
Kontrolle
Zinkmangel
Schnittnah
Schnittfern
Gruppe Lokalisation
Schnittnah
Schnittfern
Abbildung 17: ß-Catenin-Expression im Vergleich der Nahttechnik für schnittnahe Mukosa (Kontrollgruppe)
4 Ergebnisse
41
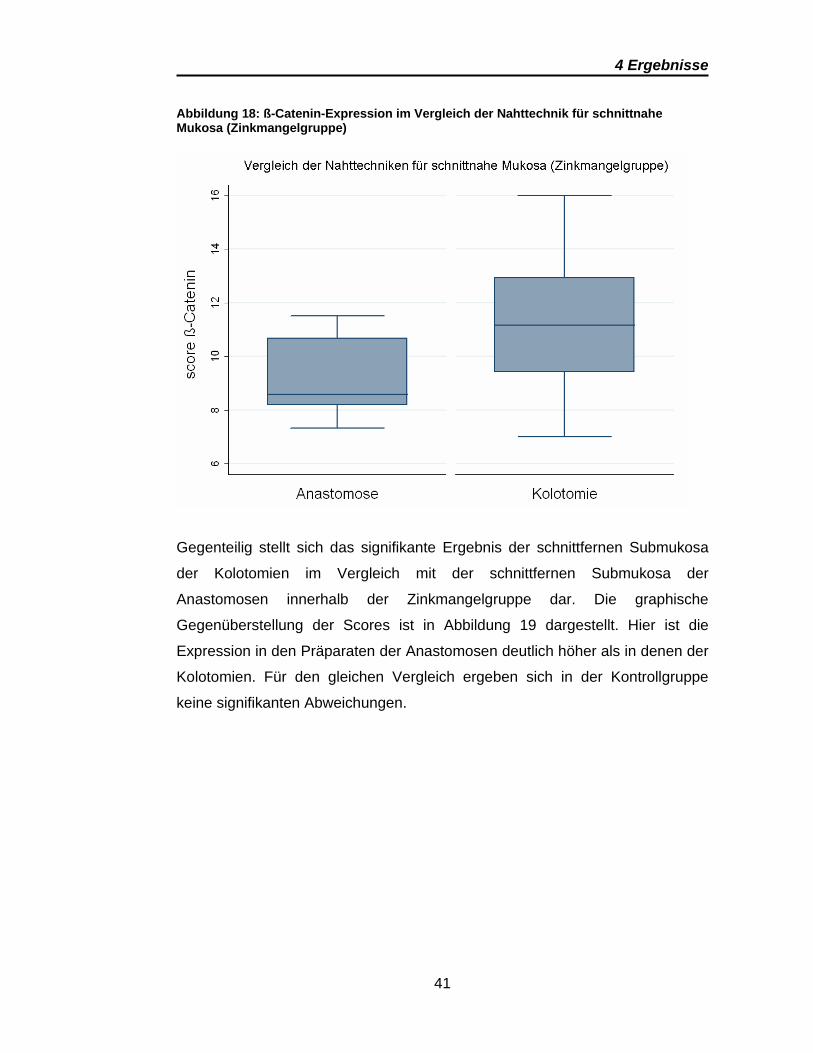
Abbildung 18: ß-Catenin-Expression im Vergleich der Nahttechnik für schnittnahe Mukosa (Zinkmangelgruppe)
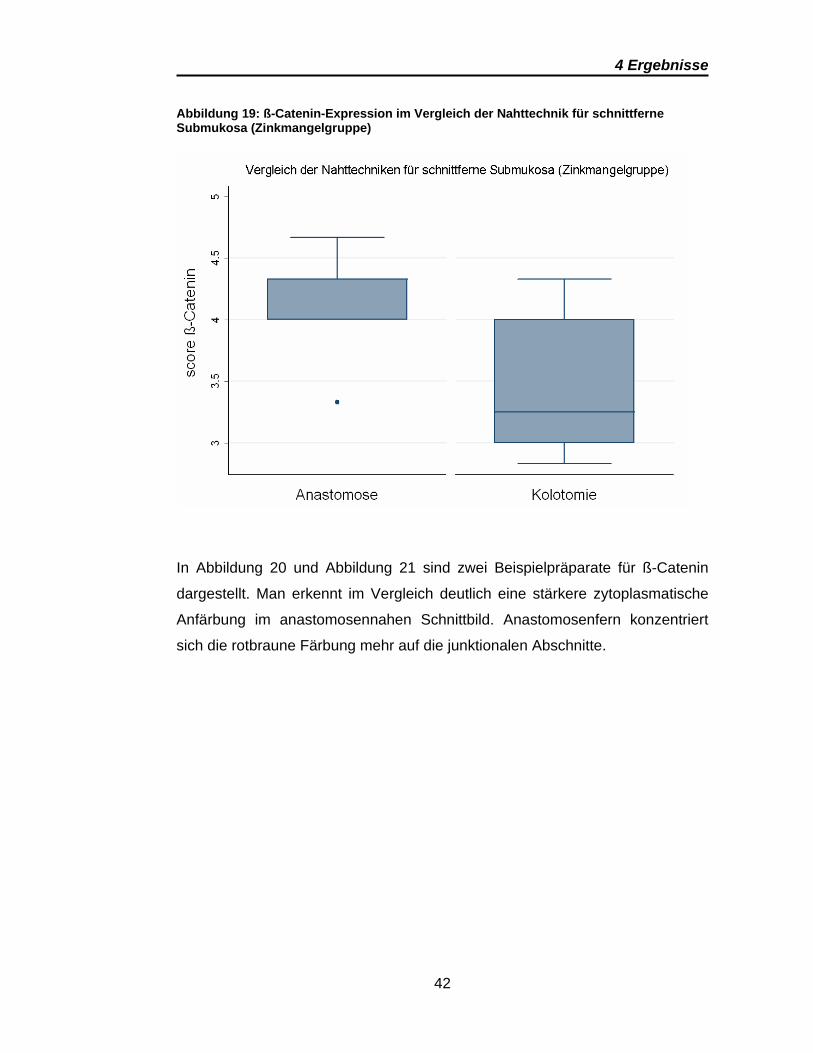
Gegenteilig stellt sich das signifikante Ergebnis der schnittfernen Submukosa
der Kolotomien im Vergleich mit der schnittfernen Submukosa der
Anastomosen innerhalb der Zinkmangelgruppe dar. Die graphische
Gegenüberstellung der Scores ist in Abbildung 19 dargestellt. Hier ist die
Expression in den Präparaten der Anastomosen deutlich höher als in denen der
Kolotomien. Für den gleichen Vergleich ergeben sich in der Kontrollgruppe
keine signifikanten Abweichungen.
4 Ergebnisse
42
Abbildung 19: ß-Catenin-Expression im Vergleich der Nahttechnik für schnittferne Submukosa (Zinkmangelgruppe)
In Abbildung 20 und Abbildung 21 sind zwei Beispielpräparate für ß-Catenin
dargestellt. Man erkennt im Vergleich deutlich eine stärkere zytoplasmatische
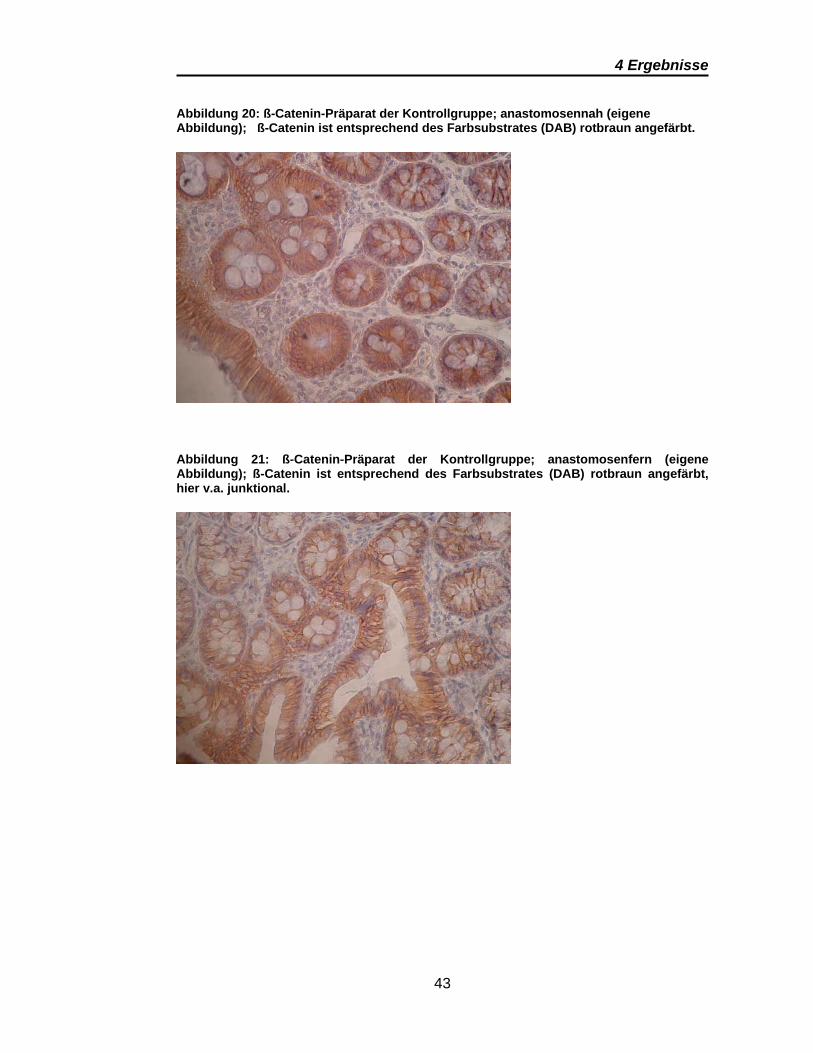
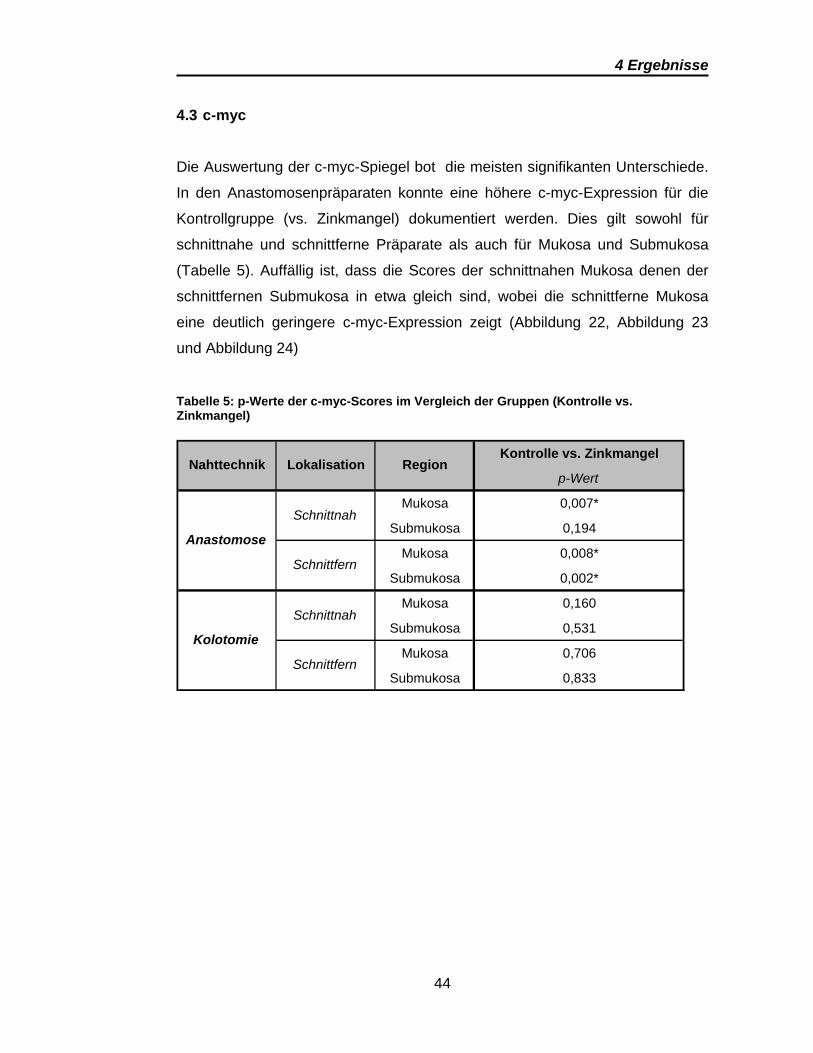
Anfärbung im anastomosennahen Schnittbild. Anastomosenfern konzentriert
sich die rotbraune Färbung mehr auf die junktionalen Abschnitte.
4 Ergebnisse
43
Abbildung 20: ß-Catenin-Präparat der Kontrollgruppe; anastomosennah (eigene Abbildung); ß-Catenin ist entsprechend des Farbsubstrates (DAB) rotbraun angefärbt.
Abbildung 21: ß-Catenin-Präparat der Kontrollgruppe; anastomosenfern (eigene Abbildung); ß-Catenin ist entsprechend des Farbsubstrates (DAB) rotbraun angefärbt, hier v.a. junktional.
4 Ergebnisse
44
4.3 c-myc
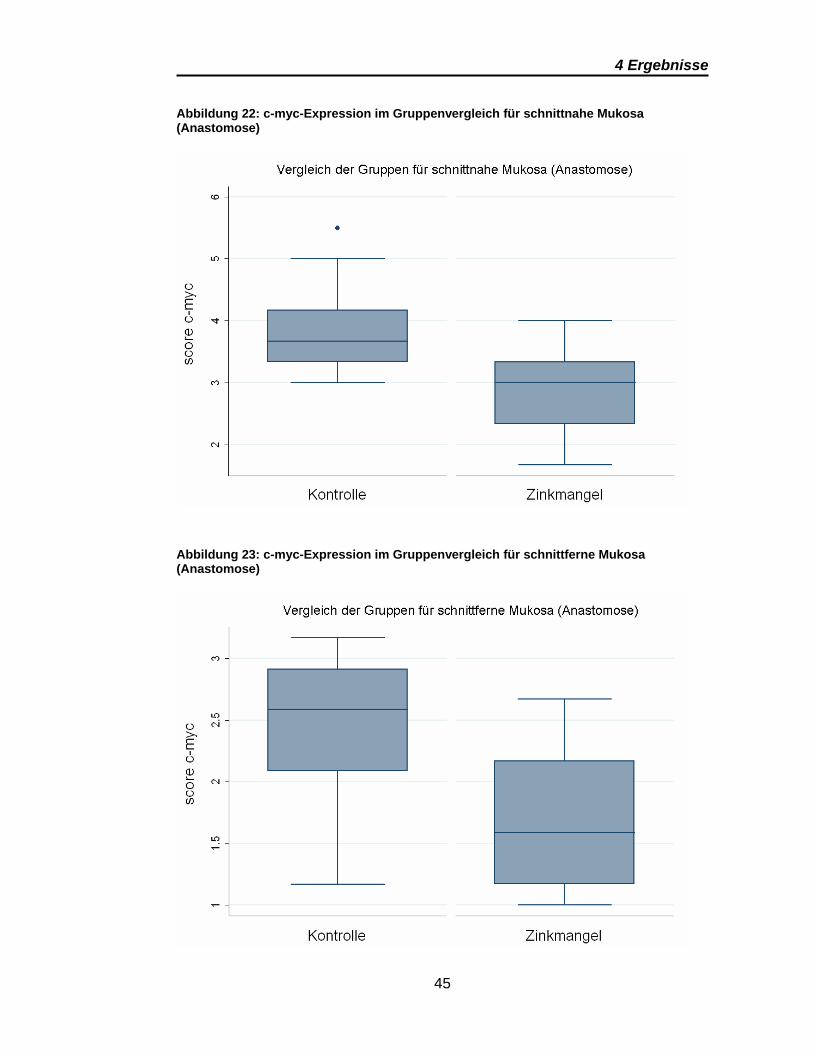
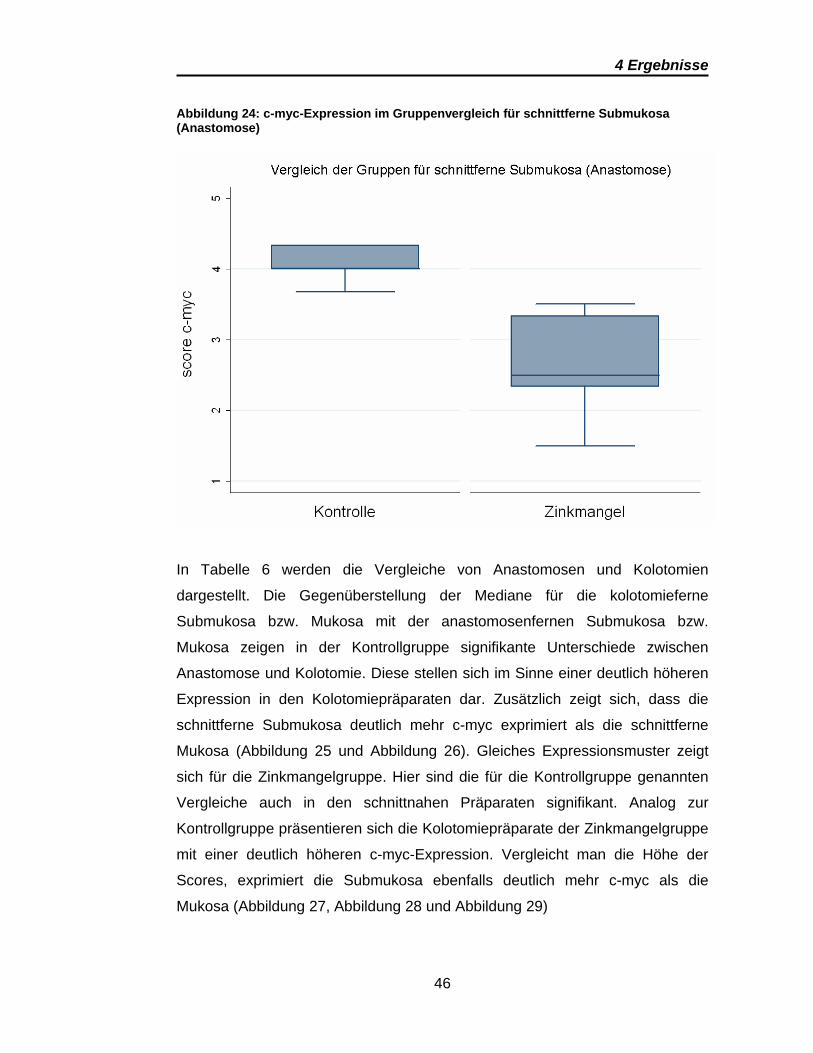
Die Auswertung der c-myc-Spiegel bot die meisten signifikanten Unterschiede.
In den Anastomosenpräparaten konnte eine höhere c-myc-Expression für die
Kontrollgruppe (vs. Zinkmangel) dokumentiert werden. Dies gilt sowohl für
schnittnahe und schnittferne Präparate als auch für Mukosa und Submukosa
(Tabelle 5). Auffällig ist, dass die Scores der schnittnahen Mukosa denen der
schnittfernen Submukosa in etwa gleich sind, wobei die schnittferne Mukosa
eine deutlich geringere c-myc-Expression zeigt (Abbildung 22, Abbildung 23
und Abbildung 24)
Tabelle 5: p-Werte der c-myc-Scores im Vergleich der Gruppen (Kontrolle vs. Zinkmangel)
Kontrolle vs. Zinkmangel
p-Wert
Mukosa 0,007*
Submukosa 0,194
Mukosa 0,008*
Submukosa 0,002*
Mukosa 0,160
Submukosa 0,531
Mukosa 0,706
Submukosa 0,833
Region
Anastomose
Kolotomie
Schnittnah
Schnittfern
Nahttechnik Lokalisation
Schnittnah
Schnittfern
4 Ergebnisse
45
Abbildung 22: c-myc-Expression im Gruppenvergleich für schnittnahe Mukosa (Anastomose)
Abbildung 23: c-myc-Expression im Gruppenvergleich für schnittferne Mukosa (Anastomose)
4 Ergebnisse
46
Abbildung 24: c-myc-Expression im Gruppenvergleich für schnittferne Submukosa (Anastomose)
In Tabelle 6 werden die Vergleiche von Anastomosen und Kolotomien
dargestellt. Die Gegenüberstellung der Mediane für die kolotomieferne
Submukosa bzw. Mukosa mit der anastomosenfernen Submukosa bzw.
Mukosa zeigen in der Kontrollgruppe signifikante Unterschiede zwischen
Anastomose und Kolotomie. Diese stellen sich im Sinne einer deutlich höheren
Expression in den Kolotomiepräparaten dar. Zusätzlich zeigt sich, dass die
schnittferne Submukosa deutlich mehr c-myc exprimiert als die schnittferne
Mukosa (Abbildung 25 und Abbildung 26). Gleiches Expressionsmuster zeigt
sich für die Zinkmangelgruppe. Hier sind die für die Kontrollgruppe genannten
Vergleiche auch in den schnittnahen Präparaten signifikant. Analog zur
Kontrollgruppe präsentieren sich die Kolotomiepräparate der Zinkmangelgruppe
mit einer deutlich höheren c-myc-Expression. Vergleicht man die Höhe der
Scores, exprimiert die Submukosa ebenfalls deutlich mehr c-myc als die
Mukosa (Abbildung 27, Abbildung 28 und Abbildung 29)
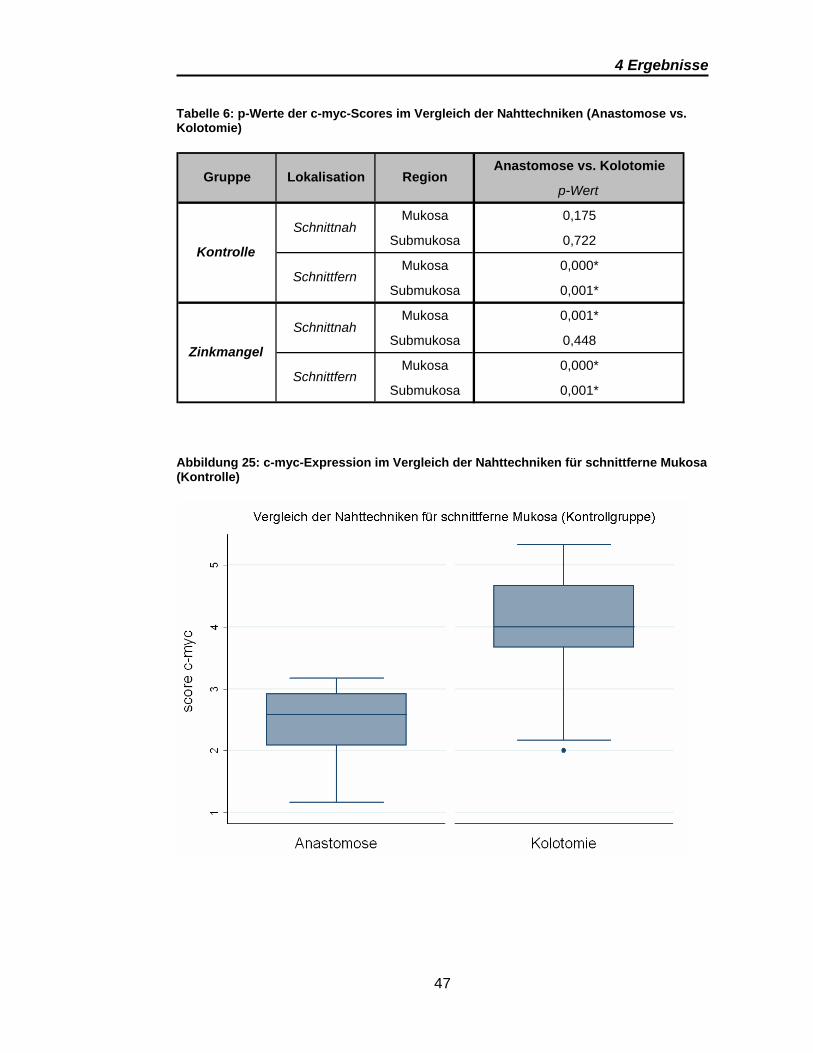
4 Ergebnisse
47
Tabelle 6: p-Werte der c-myc-Scores im Vergleich der Nahttechniken (Anastomose vs. Kolotomie)
Anastomose vs. Kolotomie
p-Wert
Mukosa 0,175
Submukosa 0,722
Mukosa 0,000*
Submukosa 0,001*
Mukosa 0,001*
Submukosa 0,448
Mukosa 0,000*
Submukosa 0,001*
Region
Kontrolle
Zinkmangel
Schnittnah
Schnittfern
Gruppe Lokalisation
Schnittnah
Schnittfern
Abbildung 25: c-myc-Expression im Vergleich der Nahttechniken für schnittferne Mukosa (Kontrolle)
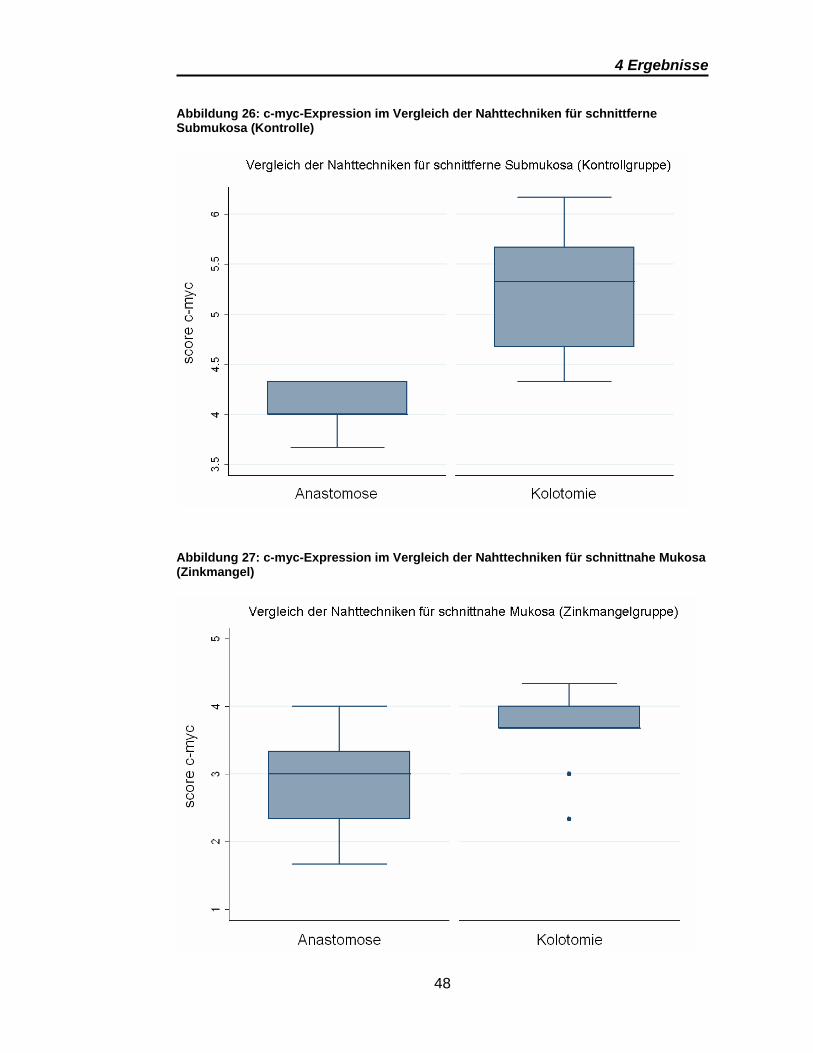
4 Ergebnisse
48
Abbildung 26: c-myc-Expression im Vergleich der Nahttechniken für schnittferne Submukosa (Kontrolle)
Abbildung 27: c-myc-Expression im Vergleich der Nahttechniken für schnittnahe Mukosa (Zinkmangel)
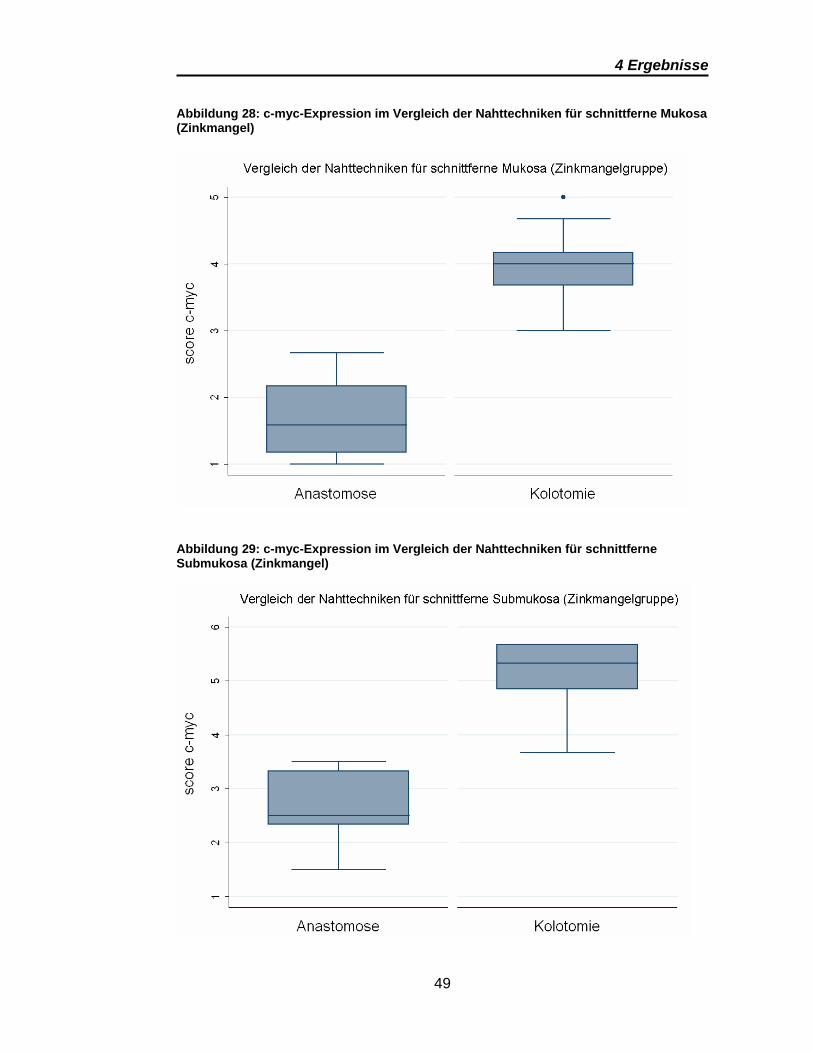
4 Ergebnisse
49
Abbildung 28: c-myc-Expression im Vergleich der Nahttechniken für schnittferne Mukosa (Zinkmangel)
Abbildung 29: c-myc-Expression im Vergleich der Nahttechniken für schnittferne Submukosa (Zinkmangel)
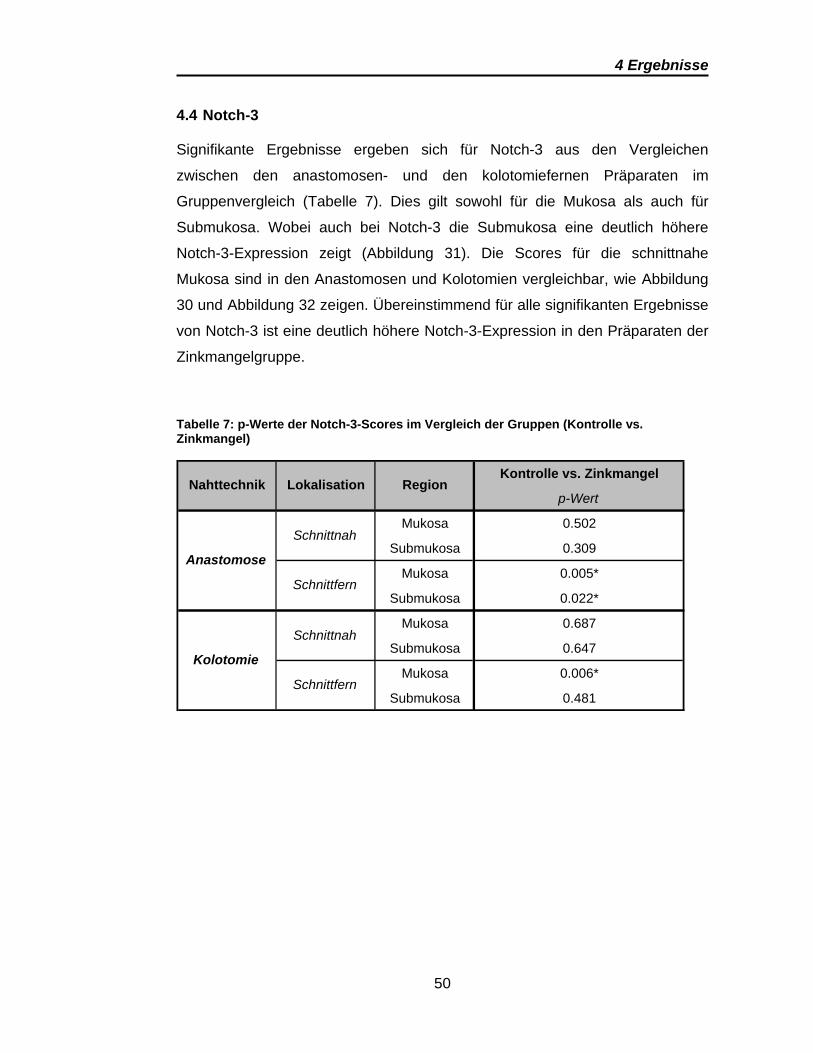
4 Ergebnisse
50
4.4 Notch-3
Signifikante Ergebnisse ergeben sich für Notch-3 aus den Vergleichen
zwischen den anastomosen- und den kolotomiefernen Präparaten im
Gruppenvergleich (Tabelle 7). Dies gilt sowohl für die Mukosa als auch für
Submukosa. Wobei auch bei Notch-3 die Submukosa eine deutlich höhere
Notch-3-Expression zeigt (Abbildung 31). Die Scores für die schnittnahe
Mukosa sind in den Anastomosen und Kolotomien vergleichbar, wie Abbildung
30 und Abbildung 32 zeigen. Übereinstimmend für alle signifikanten Ergebnisse
von Notch-3 ist eine deutlich höhere Notch-3-Expression in den Präparaten der
Zinkmangelgruppe.
Tabelle 7: p-Werte der Notch-3-Scores im Vergleich der Gruppen (Kontrolle vs. Zinkmangel)
Kontrolle vs. Zinkmangel
p-Wert
Mukosa 0.502
Submukosa 0.309
Mukosa 0.005*
Submukosa 0.022*
Mukosa 0.687
Submukosa 0.647
Mukosa 0.006*
Submukosa 0.481
Region
Anastomose
Kolotomie
Schnittnah
Schnittfern
Nahttechnik Lokalisation
Schnittnah
Schnittfern
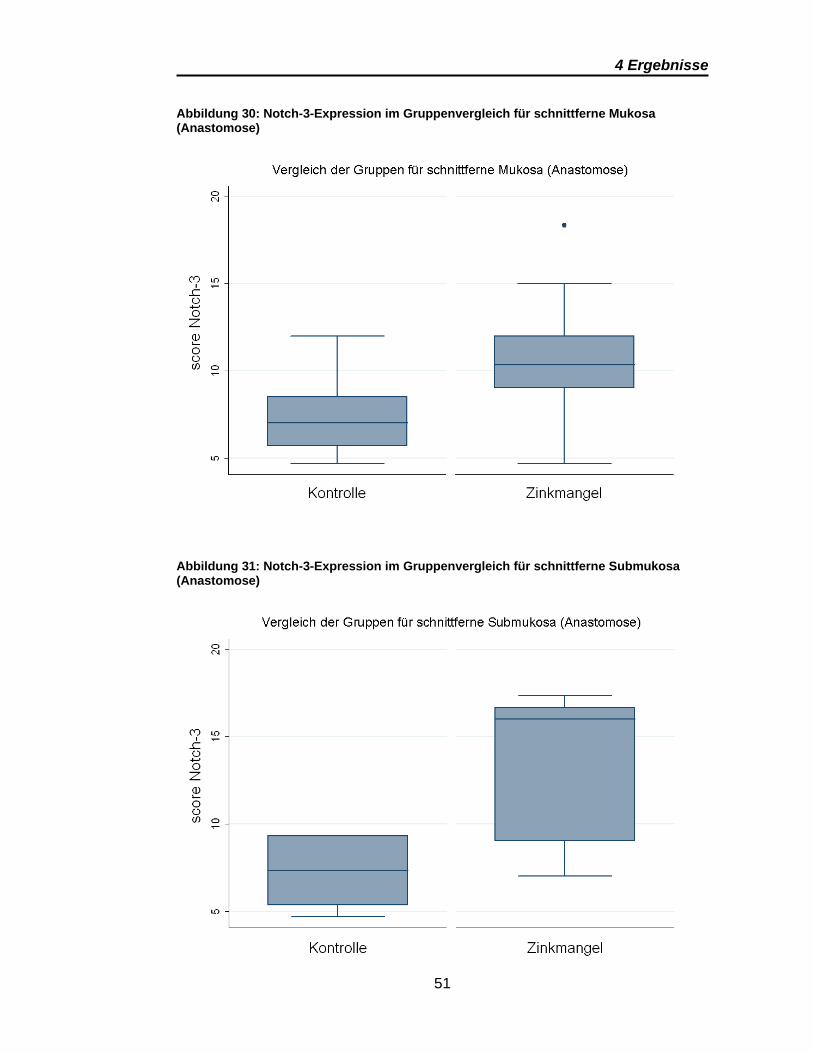
4 Ergebnisse
51
Abbildung 30: Notch-3-Expression im Gruppenvergleich für schnittferne Mukosa (Anastomose)
Abbildung 31: Notch-3-Expression im Gruppenvergleich für schnittferne Submukosa (Anastomose)
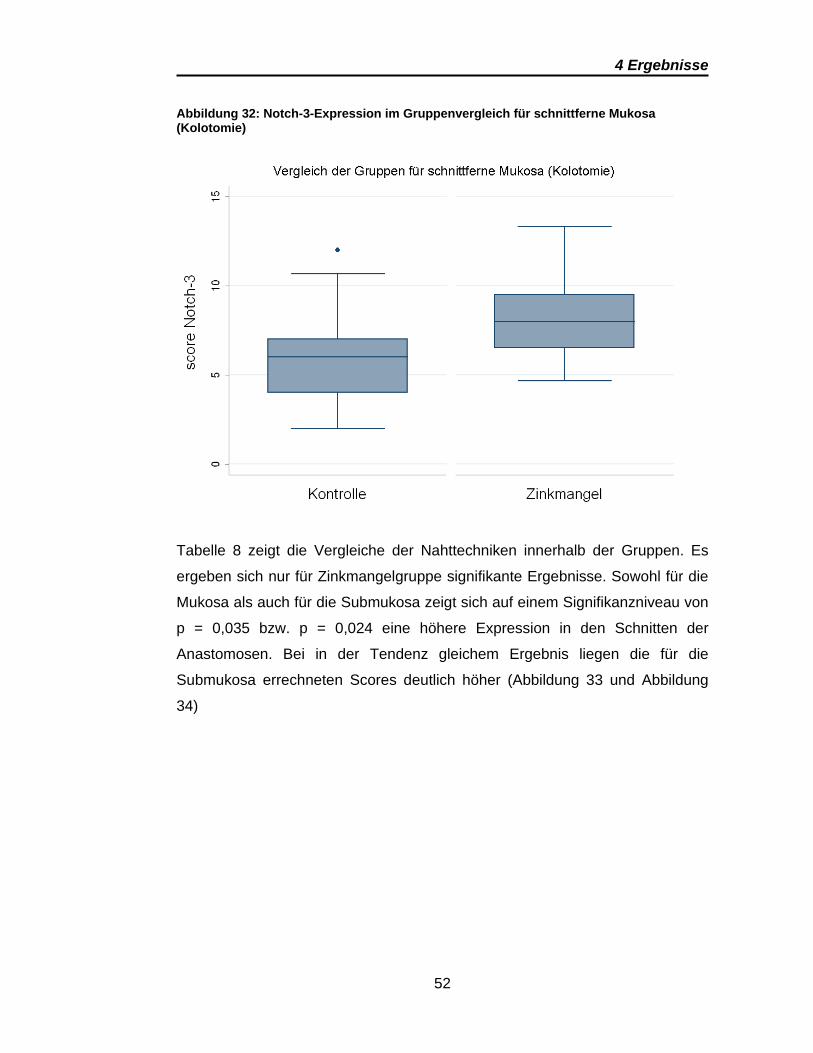
4 Ergebnisse
52
Abbildung 32: Notch-3-Expression im Gruppenvergleich für schnittferne Mukosa (Kolotomie)
Tabelle 8 zeigt die Vergleiche der Nahttechniken innerhalb der Gruppen. Es
ergeben sich nur für Zinkmangelgruppe signifikante Ergebnisse. Sowohl für die
Mukosa als auch für die Submukosa zeigt sich auf einem Signifikanzniveau von
p = 0,035 bzw. p = 0,024 eine höhere Expression in den Schnitten der
Anastomosen. Bei in der Tendenz gleichem Ergebnis liegen die für die
Submukosa errechneten Scores deutlich höher (Abbildung 33 und Abbildung
34)
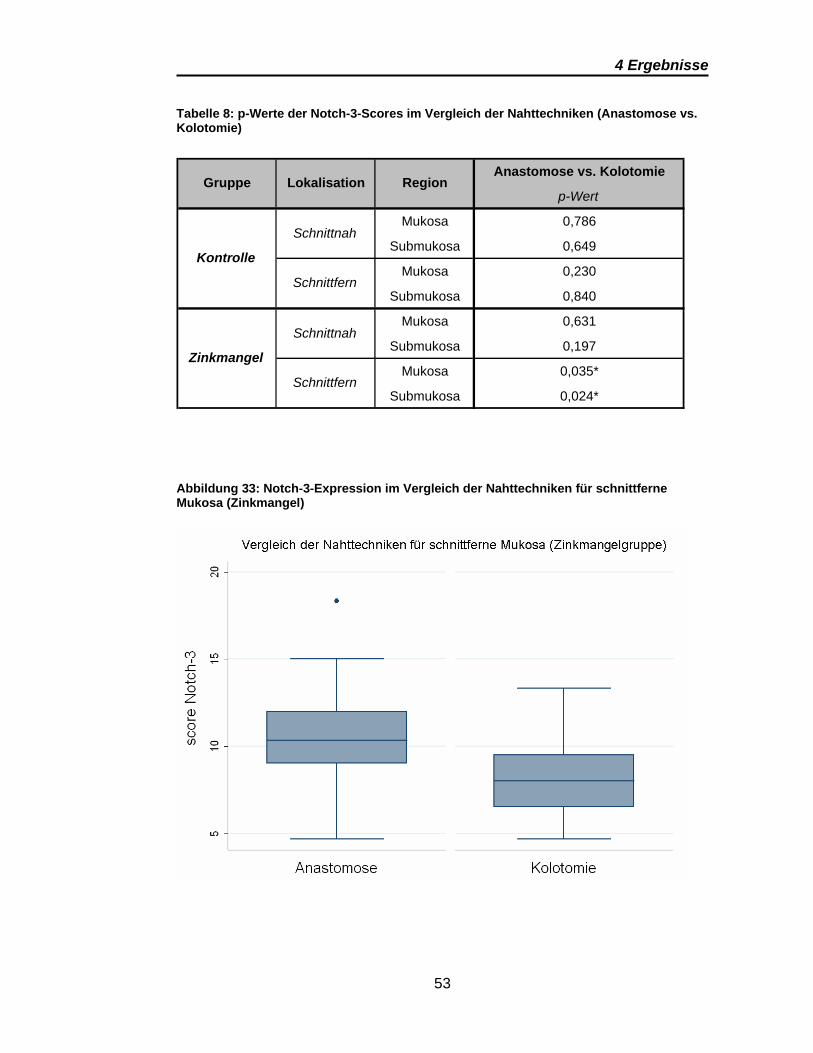
4 Ergebnisse
53
Tabelle 8: p-Werte der Notch-3-Scores im Vergleich der Nahttechniken (Anastomose vs. Kolotomie)
Anastomose vs. Kolotomie
p-Wert
Mukosa 0,786
Submukosa 0,649
Mukosa 0,230
Submukosa 0,840
Mukosa 0,631
Submukosa 0,197
Mukosa 0,035*
Submukosa 0,024*
Region
Kontrolle
Zinkmangel
Schnittnah
Schnittfern
Gruppe Lokalisation
Schnittnah
Schnittfern
Abbildung 33: Notch-3-Expression im Vergleich der Nahttechniken für schnittferne Mukosa (Zinkmangel)
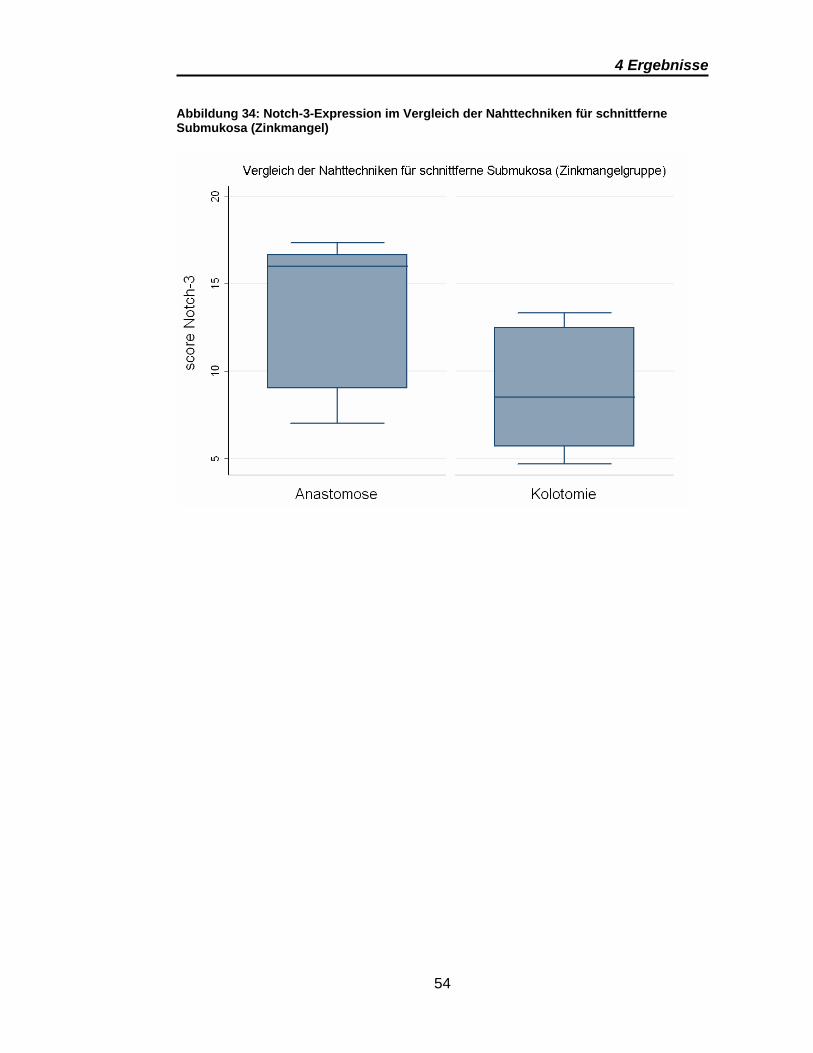
4 Ergebnisse
54
Abbildung 34: Notch-3-Expression im Vergleich der Nahttechniken für schnittferne Submukosa (Zinkmangel)

5 Diskussion
55
5 Diskussion
Wundheilungsstörungen können erhebliche Auswirkungen auf den Erfolg
operativer Eingriffe haben. Aufgrund der lebensbedrohlichen Komplikationen gilt
dies im Besonderen für die Abdominalchirurgie. Dass der Zinkhaushalt im
Rahmen der Wundheilung verschiedene zelluläre und extrazelluläre
Reparations- und Regenerationsabläufe beeinflusst, wurde in einigen
Publikationen bereits dokumentiert. Die Arbeitsgruppen von Fraser und
Lansdown wiesen nach, dass humane Wunden in der Heilungsphase Zink
anreichern (Fraser et al., 1989; Lansdown 1996). Pfaffl und Windisch
beschrieben insbesondere das Kolon als zinksensibles Organsystem (Pfaffl und
Windisch, 2003). Vaxman et al. konnten nachweisen, dass auch heilende
Kolonanastomosen im Kaninchenmodell vermehrt Zink anreichern, was
zeitgleich zu einem Absinken des Zinkspiegels im Serum führt (Vaxman et al.,
1992). Bei akut erniedrigtem Zinkangebot werden zunächst die Zinkspeicher
z.B. im Skelettsystem entleert, um den Mangel zu kompensieren (Windisch,
2003).
Neben der Wundheilung benötigt auch das Immunsystem eine stabile
Zinkhömostase (Agren et al., 1990, Krones et al. 2004-2005). In
Übereinstimmung kam es in unserer Untersuchung in der Zinkmangelgruppe
präoperativ zu zwei Todesfällen durch Pneumonie.
Einzelne Bestandteile der ECM und verschiedene Faktoren des Zellzyklus
wurden bereits auf ihre Funktion in der Wundheilung untersucht. Besonders
sensibel auf ein Zinkdefizit reagieren dabei die MMPs und daraus resultierend,
der Kollagenstoffwechsel (Krones, 2006, Binneboesel et al., 2009). Die in
dieser Studie untersuchten Parameter – TIMP-1, ß-Catenin, c-myc, Notch-3 –
wurden bereits als relevante Einflussgrößen der Zellregeneration definiert
(Alberts et al., 2004; Ishizuya-Oka, 2005; Stojadinovic et al., 2005; Wang et al.,
2002). Wie sich eine gestörte Hömöostase des Zinkstoffwechsels auf die
untersuchten Komponenten auswirkt, ist dagegen bisher nicht bekannt. Zink ist
zwar nicht direkter Bestandteil der hier untersuchten Faktoren. Es kann aber
über seine Funktion als Antioxidant die Modulation verschiedenster

5 Diskussion
56
Signalkaskaden beeinflussen. Zudem greift Zink über den MMP- und
Kollagenstoffwechsel zumindest indirekt in den TIMP-Haushalt ein.
Alle in dieser Arbeit untersuchten Faktoren – TIMP-1, ß-Catenin, c-myc, Notch-
3 – wurden zusammenfassend nur sehr eingeschränkt durch die experimentell
gestörte Zinkhomöostase beeinflusst. Überraschend erwies sich dabei selbst
TIMP-1, als einer der physiologischen Regulatoren des MMP-Stoffwechsels als
zinkunabhängig. Auch die Technikvergleiche zeigen keine einheitlichen
Unterschiede, die auf einen systemischen Einfluss rückschließen ließen. Die
verglichenen Nahttechniken spielen offensichtlich in den Heilungsvorgängen auf
molekularer Ebene keine relevante Rolle. Als grundsätzlicher Nachteil des hier
verwendeten Studienmodells muss dabei die einzeitige Beurteilung der
Expressionsmuster angesehen werden. Möglicherweise wäre retrospektiv eine
mehrzeitige Testung in Analogie zu den verschiedenen Phasen der
Wundheilung aufschlussreicher gewesen. Diese würde eine Veränderung des
Expressionsmusters einzelner Faktoren im Verlauf darstellen, womit sich die
Relevanz der Einflussnahme eines Zinkmangels besser einschätzen ließe.
.
5.1 TIMP-1
In unserem experimentellen Ansatz ließ sich keine signifikante Korrelation
zwischen der TIMP-1-Expression und dem kontrolliert induzierten Zinkmangel
nachweisen. Die gemessenen Scores erwiesen sich in beiden
Untersuchungsgruppen als weitgehend unabhängig vom Zinkangebot.
TIMPs besitzen als Proteaseninhibitoren eine Schutzfunktion für das Gewebe.
Sie hemmen vor allem die proteolytische Aktivität der MMP´s. Ergebnisse von
Knittel et al. zeigen, dass TIMPs im Wundgebiet eine höhere Konzentration
aufweisen als in gesundem Gewebe (Knittel et al., 2000). Auch Lodish et al.
konnten zeigen, dass TIMP-1 im Grenzbereich zwischen gesundem und
krankem bzw. verletztem Gewebe vermehrt exprimiert wird (Lodish et al.,
2008).
Voruntersuchungen unserer Arbeitsgruppe ergaben unter einem Zinkmangel
eine höhere MMP-Expression, was zu einer verstärkten Kollagendegradation

5 Diskussion
57
führte (Krones 2006; Binnebösel et al., 2009). Nach den Ergebnissen der
vorliegenden Arbeit resultiert der verstärkte Kollagenabbau aus einer
Verschiebung des lokalen Verhältnisses von MMP´s zu TIMP-1. Hier besteht
ein erhöhtes Substratangebot (MMPs) ohne eine konkordante up-Regulation
des entsprechenden Inhibitors (TIMPs). Diese Annahme korreliert mit einem
ungehemmten Kollagenabbau durch die MMPs bei Zinkmangel, wie es Bullen
et al. als Pendant für eine schlechtere Wundheilung für chronische Wunden
belegen konnten (Bullen et al., 1995). Ein fehlender Einfluss von Zink auf TIMP-
1 ergibt einen erhöhten Kollagenabbau bei verstärkter MMP-Expression durch
ein pathologisches Verhältnis bzw. Ungleichgewicht der beiden Gegenspieler.
In der Gegenüberstellung der beiden untersuchten Nahttechniken fanden sich
dagegen zumindest vereinzelt signifikante Unterschiede. In zwei Fällen
exprimierte die schnittnahe Mukosa der Anastomosenpräparate in beiden
Gruppen (Kontrolle und Zinkmangel) mehr TIMP-1 als die schnittnahe Mukosa
der Kolotomiepräparate. Dass die beiden konkordanten Vergleiche beider
Gruppen signifikante Ergebnisse mit gleichartigem Expressionsmuster zeigen,
bestätigt zunächst nochmals die Unabhängigkeit der TIMPs vom Zinkhaushalt
in unserem Arbeitsansatz. Die höhere Expression in der schnittnahen Mukosa
von Anastomosen im Vergleich mit den Kolotomien lässt beim größeren
Gewebetrauma der Anastomosen auf einen konsekutiv verminderten
Kollagenabbau schließen.
Zusammenfassend präsentiert sich TIMP-1 in der vorliegenden Untersuchung
als stabiler Parameter, und als von den hier untersuchten Einflussgrößen im
Wesentlichen unabhängig. Eine mehrfach vordokumentierte, generelle
Induzierbarkeit von TIMP-1 ist demnach nicht von den hier untersuchten
Variablen abhängig (Lodish et al., 2008, Senapati et al., 1985).

5 Diskussion
58
5.2 ß-Catenin
Auch das Proto-Onkogen ß-Catenin erwies sich in unserem Ansatz als
weitgehend zinkunabhängig. ß-Catenin kann auf zellulärer Ebene zwei
unterschiedliche Funktionen ausüben. Es fungiert sowohl als Strukturprotein im
Zytoskelett als auch als wichtiger Effektor des Wnt-Pathways und damit als
Schlüsselsignal in der intestinalen Zellerneuerung (Lodish et al., 2008; Alberts
et al., 2004; Knippers, 2001). Neben den beschriebenen Funktionen kann eine
unkontrollierte ß-Catenin-Exprimierung zu zellulären Fehlfunktionen im
Zusammenhang mit der Kanzerogenese führen.
Die jeweilige Funktion von ß-Catenin kann durch die in diesem Versuch
durchgeführten Färbungen nicht eindeutig unterschieden werden, das
Färbemuster lässt aber Schlussfolgerungen auf die Wirkungsweise zu. Viele
Studien haben sich bisher mit der Funktion von ß-Catenin in der
Kanzerogenese beschäftigt. Dabei konnte gezeigt werden, dass eine
verminderte junktionale ß-Catenin-Expression zu einer vermehrten
Tumorinvasion führt. Des Weiteren wurde festgestellt, dass eine vermehrte ß-
Catenin Aktivität im Wnt-Signalweg ebenfalls zu einem schnelleren
Tumorprogress führte (Alberts et al., 2004, Ouko et al., 2004; Zhurinsky et al.,
2000). Übertragen auf die zelluläre Regulation in der Wundheilung wird deshalb
eine Hochregulierung der Aktivität des Proteins erwartet. Zink moduliert
verschiedenste Signalkaskaden auch innerhalb der Wundheilung. Zudem
verändert die anti-oxidative Potenz des Atoms das lokale Milieu (Zalewski et al.,
1994; Truong-Tran et al., 2001). Obwohl Zink nicht Bestandteil von ß-Catenin
ist, ist es so grundsätzlich denkbar, dass es in seiner Funktion als Antioxidant
auch zinkunabhängige Faktoren des Zellzyklus beeinflusst.
Der Gruppenvergleich der Kolotomien zeigt ß-Catenin in der Zinkmangelgruppe
sporadisch erniedrigt. Dieses Ergebnis impliziert, dass der Wnt-Pathway in der
frühen postoperativen Phase der Kolotomie-Heilung negativ beeinflusst wird,
was auf eine verminderte Zellerneuerung schließen lässt. Ebenso kann eine
niedrigere ß-Catenin-Exprimierung aber auch eine verminderte Stabilität der
einzelnen Zellen sowie des Zellverbandes anzeigen. Betrachtet man unter
diesen Gesichtspunkten die Färbemuster der beiden Gruppen im Vergleich, so

5 Diskussion
59
sieht man in den zugehörigen Präparaten der Kontrollgruppe eine intensivere
zytoplasmatische Färbung.
Auffällig ist in allen Präparaten, dass sich die zytoplasmatische ß-Catenin-
Expression schnittfern vor allem auf die Zellen der Mukosa bezieht. In
Übereinstimmung mit Ergebnissen von Pinto et al. lässt dies auf eine höhere
proliferative Aktivität dieser Gewebe schließen. Pinto et al. beschreiben eine ß-
Catenin-Anreicherung vor allem in den regenerativ aktiven Krypten des
Dünndarmes von Mäusen (Pinto et al., 2003). Ein abweichendes
Expressionsmuster zeigen im Vergleich hierzu die Zellen der Submukosa, die
ß-Catenin deutlich stärker zellwandnah exprimieren. Nach der Färbung sind die
junktionalen Abschnitte hier mehr betont als das Zytoplasma. Eine mögliche
Erklärung wäre, dass die Submukosa als „Darmskelett“ in erster Linie der
Stabilität dient, der Zellumsatz jedoch geringer ist.
Vergleiche der schnittnahen Mukosa zeigen eine höhere ß-Catenin-Expression
in den Kolotomiepräparaten. Hierzu widersprüchlich bleiben jedoch die
Untersuchungen der schnittfernen Submukosa. Dort zeigen die Anastomosen
eine höhere Expression von ß-Catenin. Dieses Ergebnis unterstreicht nochmals
den fehlenden Einfluss der Nahttechniken auf die ß-Catenin-Expression.
5.3 c-myc
C-myc ließ sich in unseren Untersuchungen ebenfalls durch den Zinkmangel
nicht beeinflussen. C-myc ist als Protoonkogen Effektor des Wnt-Signalweges
und dabei an die Induktion durch ß-Catenin und andere Wachstumsfaktoren
gebunden (Albertz et al., 2004; Cheng et al., 1999; Chiariello et al., 2001). Eine
verminderte c-myc-Expression stört die Zelladhäsion und Zellmotilität sowie die
Zusammensetzung von ECM- und Adhäsionsmolekülen (Frye et al., 2003).
Andererseits kann c-myc analog zu ß-Catenin bei Überaktivierung zu einer
funktionellen Erschöpfung der Zellen im Wundgebiet führen (Stojadinovic et al.,
2005).
Die bekannte Interdependenz zwischen ß-Catenin und c-myc (Ouko et al.,
2004; Waltenberger et al., 2001; Zhurinsky et al., 2000) bestätigte sich in

5 Diskussion
60
unseren Experimenten erwartungsgemäß mit analogen Expressionsmustern in
den Gruppenvergleichen. Für c-myc wurde wie für ß-Catenin unabhängig von
der Lokalisation eine sporadisch niedrigere Expression unter Zinkmangel
beobachtet. Dies deckt sich zumindest tendentiell mit Ergebnissen von Prasad,
der im Zinkmangel eine verminderte DNA-Synthese nachwies (Prasad, 1996).
Übereinstimmend zeigten die Präparate der Kontrollgruppe tendentiell höhere
Proliferationsraten. Diese Beobachtung korreliert mit einer verminderten
Proliferationsrate bei kontrolliertem Zinkmangels in der Arbeit von Binnebösel et
al. (Binnebösel et al., 2009).
Im Vergleich der Nahttechniken weichen die Messdaten von c-myc
überraschend von den Ergebnissen der ß-Catenin-Spiegel ab. Die
Kolotomiepräparate zeigten vereinzelt eine signifikant höhere c-myc-
Konzentration als die Anastomosen. Dies könnte auf günstigere Bedingungen
der Zellerneuerung unter dem geringeren Trauma der Kolotomie schließen
lassen. Ursächlich käme hier die postoperativ bessere Durchblutung der
Kolotomien im Vergleich zu den Anastomosen in Frage.
Alternativ könnten die Abweichungen aber auch durch die Doppelfunktion von
ß-Catenin verursacht sein. Schließlich kann auch eine Wnt-unabhängige (und
damit auch ß-Catenin-unabhängige) c-myc Induktion über Wachstumsfaktoren
wie PDGF zu unterschiedlichen Ergebnissen führen (Xie et al. 2002; Chariello
et al., 2001). Lynch et al. konnten bereits 1987 in vitro eine gesteigerte
Zellproliferation sowohl von Bindegewebe als auch der Epidermis nach Zugabe
von ppPDGF nachweisen. Ebenso stiegen der zelluläre Proteingehalt und die
ECM-Produktion. Geht man also von einer postoperativ besseren Durchblutung
der Kolotomien aus, kann mehr PDGF über die Thrombozyten in die Wunde
gelangen. Dann erklärt sich ein gesteigerter Zellzyklus mit höherem c-myc-
Nachweis durch eine gesteigerte proliferative Kapazität mit hohem Zell-
Turnover.
Wie genau sich die Expression in der späteren postoperativen Phase darstellt
und wie sich erhöhte c-myc-Level auf chronische Wundheilungsstörungen im
Sinne einer Zellerschöpfung (Stojadinovic et al., 2005) auswirken, muss wegen
der uneindeutigen Ergebnisse jedoch Gegenstand weiterer Untersuchungen
sein.

5 Diskussion
61
5.4 Notch-3
In der vorliegenden Arbeit ergeben sich für die Notch-3-Expression in den
Präparaten der Zinkmangelgruppe signifikant höhere Scores als in der
Kontrollgruppe, was eine Zinksensibilität von Notch-3 impliziert. Diese Tendenz
gilt einheitlich für beide Nahttechniken in den schnittfernen Präparaten. Damit
ist dieses Ergebnis reziprok zu den beiden vorher getesteten Faktoren ß-
Catenin und c-myc. Alle drei gehören zu wichtigen Bestandteilen des Zellzyklus
mit sowohl kon- als auch diskordanten Einflüssen. Aus vorherigen Studien ist
eine hohe Notch-3-Expression in intestinalen Stammzellen bekannt (Ishizuya-
Oka, 2005). Aufgrund der Tatsache, dass Notch-3 kontextspezifisch nicht nur
dazu in der Lage ist, Proliferation zu stimulieren, sondern auch die
Zelldifferenzierung und sogar die Apoptose zu induzieren, müssen die
Ergebnisse jedoch situativ interpretiert werden. Im vorliegenden Fall muss
angenommen werden, dass die Funktion von Notch-3 in der Apoptoseinduktion
besteht. Dies wird unterstützt durch die Tatsache, dass Zinkmangel oxidativen
Stress im Gewebe induziert, wodurch die Apoptose-Rate ebenfalls ansteigt
(Zalewski et al., 1994; Truong-Tran et al., 2001). Es wird angenommen, dass
Notch-3 den Wnt-Signalweg sowohl analog als auch kontrovers beeinflussen
kann (Lodish et al., 2008; Alberts et al., 2004). Deregowski et al. zeigen, dass
Notch-3-Proteine mit dem Wnt-Signalweg umgekehrt proportional interagieren
(Deregowski et al., 2006). In diesem Fall scheint es wahrscheinlich, dass die
geringere Expression von Notch-3 in der Kontrollgruppe über den Wnt-
Signalweg die Proliferation stimuliert und in der Zinkmangelgruppe diese
blockiert bzw. Apoptose induziert.
Die negative Korrelation von Notch-3 mit c-myc zeigt sich sowohl im Vergleich
der beiden Zink-Gruppen als auch der Nahttechniken. Hier ergeben sich für
Notch-3 im Gegensatz zu c-myc höhere Werte in den Anastomosen. Auch hier
spricht die reziproke Darstellung im Vergleich mit c-myc für eine umgekehrt
proportionale Interaktion mit dem Wnt-Signalweg. Als ursächlich kommt im
Vergleich der Nahttechniken auch ein größeres Gewebetrauma mit
kompromittierter Durchblutung und konsekutiv verändertem Stoffwechsel in
Frage. Da Zink ebenfalls kein molekularer Bestandteil von Notch-3 ist, scheint

5 Diskussion
62
Notch-3 am ehesten sensibel auf die durch den Zinkmangel veränderten
Umgebungsreaktionen (z.B. oxidativen Stress) zu reagieren.

6 Zusammenfassung
63
6 Zusammenfassung
Obwohl Darmoperationen Routineeingriffe sind, bleibt die Nahtinsuffizienz auch
heute noch die gefährlichste Komplikation. Daher behalten Untersuchungen zur
Wundheilung am Darm zentrale Bedeutung. Neben Operationstechnik und
Erfahrung nehmen auch molekulare Faktoren Einfluss auf die Wundheilung.
Dazu gehören auch Veränderungen der Zusammensetzung der
Extrazellulärmatrix (ECM). Die Prävention von Wundheilungsstörungen setzt
daher ein grundlegendes Verständnis der einzelnen, an der Wundheilung
beteiligten Faktoren voraus.
Voraussetzung der Wundheilung ist auch am Darm ein intakter
Kollagenstoffwechsel. Dieser bildet im Wechselspiel zwischen Synthese und
Degradation abschließend eine stabile Narbe. Die Regulation erfolgt dabei
primär über zinkabhängige Proteasen (MMP), die jedoch von weiteren Faktoren
der Wundheilungskaskade unterstützt oder reguliert werden. Eine negative
Zinkbilanz kann über eine Funktionsstörung der MMPs die Wundheilung
unterdrücken. Der Einfluss von Zink bzw. eines Zinkmangels auf die
physiologische Funktion der anderen oben angegebenen Regulatoren ist
dagegen noch unbekannt. Diese Arbeit untersucht deshalb unter einem
kontrollierten Zinkmangel die Expressionsmuster der primär nicht zink-
abhängigen ECM-Faktoren TIMP-1, ß-Catenin, c-myc und Notch-3 in der
Wundheilung des Ratten-Kolons. In einem zweiten Ansatz wurden zudem der
Einfluss unterschiedlicher Nahttechniken auf die Proteinexpression beurteilt.
Zusammenfassend ließ sich die Expression der meisten Zielparameter durch
den Zinkmangel und die Nahttechniken nur gering beeinflussen. TIMP-1, ß-
Catenin und c-myc erwiesen sich als weitgehend zinkunabhängig. Lediglich die
Notch-3-Expression bot in der Zinkmangelgruppe zumindest vereinzelt
signifikant erhöhte Werte auf. Die Nahttechnik (Anastomose versus Kolotomie)
hatte dagegen bei allen Zielgrößen keinen einheitlichen Einfluss.
Die Versuchsreihe schließt damit einen relevanten Einfluss von Zinkmangel und
OP-Technik auf die untersuchten Zielparameter aus. Die einzeitige Beurteilung
der Expressionsmuster muss dabei aber als Einschränkung angesehen werden,
da sie die Phasen der Wundheilung nicht abbilden kann.

6 Zusammenfassung
64
Summary
Although intestinal operations are routine surgery, insufficiency is still the most
life-threatening complication. That is why research on intestinal wound healing
is still of particular importance. Besides operation technique and experience of
the surgeon influences of molecular factors are considered to be important for a
successful wound healing. The change of the extracellular matrix is one of
these factors. Prevention of disturbances of wound healing requires a
fundamental knowledge of molecular processes.
Precondition for intestinal wound healing is an intact collagen metabolism. This
kind of interplay between synthesis and degradation constitutes a functional
scar. Its regulation is ruled by zinc-dependent proteases (MMPs), which
themselves are regulated by other factors such as TIMPs. A negative zinc
balance can disturb the function of MMPs and suppress wound healing. The
influence of zinc on physiologic function of the factors, tested in this work, is not
known so far. In this study, the expression of TIMP-1, ß-Catenin, c-myc and
Notch-3 are checked concerning changes when zinc deficiency is induced in
adult rats. Another examination evaluates the influence of different operation
method (anastomosis and colotomy) on protein expression.
In summary, the expression of the reviewed factors is influenced little by zinc or
the operation method. TIMP-1 is totally zinc-independent, ß-Catenin and c-myc
are to a large extent zinc-independent. Notch-3 is influenced to some extent
due to significant higher scores in zinc deficiency. The operation method is most
important for c-myc, which has a higher expression in colotomies.
This series of experiments excludes a relevant influence of zinc deficiency and
operation method on the here examined factors. The one-phase evaluation of
the expression cluster should be considered as a limitation, because it cannot
display the different phases of wound healing.

7 Literaturverzeichnis
65
7 Literaturverzeichnis
Agren MS. Studies on zinc in wound healing. Acta Dermato-Venerologica.
Supple-mentum. 154:1-36. 1990
Alberts B, Johnson A, Lewis J, Raff M, Roberts K, Walter P. Molekularbiologie
der Zelle. Vierte Auflage. Wiley-VCH Verlag GmbH & Co. KgaA. 2004
Arora PD, Narani N, McCulloch CA. The compliance of collagen gels regulates
transforming growth factor-beta induction of alpha-smooth muscle actin in
fibroblasts. American Journal of Pathology 154(3):871-882. 1999
Antonson DL, Barak AJ, Vanderhoof JA. Determination of the site of zinc
absorption in rat small intestine. Journal of Nutrition 1: 142-47. 1979
Baker AH, Edwards DL, Murphy G. Metalloproteinase inhibitors: biological
actions and therapeutic opportunities. Journal of Cell Science 115: 3719-
3727. 2002
Berg JM, Tymoczko JL, Stryer L. Biochemie. Spektrum Akademischer Verlag
GmbH Heidelberg. P.261,263. 2003
Bernhard S, Eilers M. Control of cell proliferation and growth by Myc proteins.
Book series: Results and problems in cell differentiation 42: 329-42. 2006
Betzler M, Schmidt C. Standard und Risiko von Kolonanastomosen. Viszeral-
chirurgie 40:8-11. 2005
Binnebösel M, Grommes J, Koenen B, Junge K, Klink CD, Stumpf M, Ottinger
AP, Schumpelick V, Klinge U, Krones CJ. Zinc deficiency impairs wound
healing of colon anastomosis in rats. International Journal of Colorectal
Disease: Published online: 27 October 2009. 2009
Borden P, Heller RA. Transcriptional control of matrix metalloproteinases and
the tissue inhibitors of matrix metalloproteinases. Critical Reviews in
Eukaryotic Gene Expression. 7(1-2):159-78. 1997
Brew K, Dinakarpandian D, Nagase H. Tissue Inhibitors of Metalloproteinases:
Evolution, Structure and Function. Biochimica et Biophysica Acta 1477:
267-283. 2000
Bullen EC, Longaker MT, Updike DL, Benton R, Ladin D, Hou Z, Howard EW.
TIMP-1 is decreased and activated Gelatinases are increased in chronic
wounds. Journal of Investigative Dermatology 104: 236-240. 1995

7 Literaturverzeichnis
66
Burch JM, Franciose RJ, Moore EE, Biffl WJ, Offner PJ. Single-Layer
Continuous versus Two-Layer Interrupted Intestinal Anastomosis. Annals
Of Surgery 231 (6): 832-837. 2000
Cao J, Zucker S. Biology and chemistry of matrix metalloproteinases. URL:
http://www.abcam.com/index.html?pageconfig=resource&rid=11034#2.
2004
Charrier L, Yan Y, Driss A, Laboisse CL, Sitaraman SL, Merlin D. ADAM-15
inhibits wound healing in human intestinal epithelial cell monolayers.
American Journal of Physiology - Gastrointestinal and Liver Physiology
288: 346-353. 2004
Cheon SS, Wei Q, Gurung A, Youn A, Bright T, Poon R, Whetstone H, Guha A,
Alman BA. Beta-catenin regulates wound size and mediates the effect of
TGF-beta in cutaneous healing. The Journal of the Federation of American
Societies for Experimental Biology 20: 692-701. 2006
Chiariello M, Marinissen MJ, Gutkind JS. Regulation of c-myc expression by
PDGF through Rho GTPases. Nature Cell Biology 3: 580–586. 2001
Clark RA. Biology of dermal wound repair. Clinical Dermatology 11(4):647-666.
1993
Coleman JE. Zinc proteins: enzymes, storage proteins, transcription factors,
and replication proteins. Annual Review of Biochemistry 61 : 897-946.
1992
Deregowski V, Gazzerro E, Priest L, Rydziel S, Canalis E. Notch 1
Overexpression Inhibits Osteoblastogenesis by Suppressing Wnt/ß-
Catenin but Not Bone Morphogenetic Protein Signaling. The Journal of
Biological Chemistry 281(10): 6203-6210. 2006
Dietz UA, Debus ES. Intestinal anastomosis prior to 1982; a legacy of ingenuity,
persistence and research from a foundation for modern gastrointestinal
surgery. World Journal of Surgery. 29: 396-401. 2002
Fraser, WD, Taggart, DP, Fell, GS, Lyon, TD, Wheatley, D, Garden, OJ,
Shenkin, A. Changes in iron, zinc, and copper concentrations in serum and
in their binding to transport proteins after cholecystectomy and cardiac
surgery. Clinical Chemistry 35: 2243-2247. 1989

7 Literaturverzeichnis
67
Fre S, Huyghe M, Mourikis P, Robine S, Louvard D, Artavanis-Tsakonas S.
Notch signals control the fate of immature progenitor cells in the intestine.
Nature 435 (7044): 964-968. 2005
Frye M, Gardner C, Li ER, Arnold I, Watt FM. Evidence that myc activation
depletes the epidermal stem cell compartment by modulating adhesive
interactions with the local microenvironment. Development 130: 2793-
2808. 2003
Gailit J, Clark RAF. Wound repair in the context of extracellular matrix. Current
Opinion in Cell Biology 6: 717-725. 1994
Gomez DE, Alonso DF, Yoshiji H, Thorgeirsson UP. Tissue Inhibitors of
Metalloproteinases: Structure, Regulation and biological function.
European Journal of Cell Biology 74(2):111-22. 1997
Gray GE, Mann RS, Mitsiadis E, Henrique D, Carcangiu ML, Banks A, Leiman
J, Ward D, Ish-Horowitz D, Artavanis-Tsakonas S. Human Ligands of the
Notch Receptor. American Journal of Pathology 154: 785-794. 1999
Haase H, Rink L. Influence of zinc on immune function. Molecular aspects of
immunology 18, 2. 2006
Hansen HH, Stelzner F. Zur chirurgischen Anatomie der Arterienversorgung der
Dickdarmwand. Langenbeck`s Archives of Surgery 340: 63-74. 1975
Harder F, Kull C. Fortlaufende einreihige Darmanastomose. Der Chirurg 58:
269-273. 1987
Hendriks T, Mastboom WJB. Healing of Experimental Intestinal Anastomosis:
Parameters for Repair. Diseases of the Colon & Rectum 33: 891-901.
1990
Hofstädter F. Pathologie der Wundheilung. Der Chirurg 66: 174-181. 1995
Ibs K-H, Rink L. Zinc-Altered Immune Function. Journal of Nutrition 133: 1452-
1456. 2003
Ishido B. Über Beziehungen der Avitaminose zur Wundheilung. Aus der
experimentell-biologischen Abteilung des Pathologischen Instituts der
Universität Berlin. 1922
Ishizuya-Oka A: Epithelial-Connective Tissue Cross-Talk is Essential for
Regenera-tion of Intestinal Epithelium. Journal of Nippon Medical School
72: 13-18. 2005

7 Literaturverzeichnis
68
Kaplan B, Gönül B, Dincer S, Dincer Kaya FN, Babül, A. Relationships Between
Tensile Strength, Ascorbic Acid, Hydroxyproline, and Zinc Levels of Rabbit
Full-Thickness Incision Wound Healing. Surgery Today 34: 747-751. 2004
Kato GJ, Dang CV. Function of the c-Myc oncoprotein. The Journal of the
Federation of American Societies for Experimental Biology 6: 3065-3072.
1992
Klinge U, Si ZY, Zheng H, Schumpelick V, Bhardwaj RS, Klosterhafen B.
Abnormal collagen I to III distribution in the skin of patients with incisional
hernia. European Surgical Research 32: 43-48. 2000
Knippers R. Molekulare Genetik, Stuttgart: Thieme, 2001
Knittel T, Mehde M, Grundmann A, Saile B, Scharf JG, Ramadori G. Expres-
sion of matrix metalloproteinases and their inhibitors during hepatic tissue
repair in the rat. Histochemistry and Cell Biology 113 (6): 443-453. 2000
Kopan R. Notch: a membran-bound transcription factor. Journal of Cell Science
115: 1095-1097. 2002
Kostic LL. Sutures in digestive surgery. Acta Chirurgica Iugoslavica 41: 211-20.
1994
Kralisch S, Lossner U, Bluher M, Paschke R, Stumvoll M, Fasshauer M. Tissue
inhibitor of metalloproteinase 1 expression and secretion are induced by ß-
adrenergic stimulation in 3T3-L1 adipocytes. Journal of Endocrinology 189,
665-670. 2006
Krones CJ: Der Einfluss vaskulärer und molekularbiologischer Faktoren auf die
Heilung intestinaler Anastomosen. Habilitationsschrift. 2006
Krones CJ, Klosterhalfen B, Anurov M, Stumpf M, Klinge U, Oettinger AP,
Schumpelick V. Missing effects of zinc in a porcine model of recurrent
endotoxemia. BMC Surgery 5: 22. 2005
Krones CJ, Klosterhalfen B, Butz N, Hoelzl F, Junge K, Stumpf M, Peiper C,
Klinge U, Schumpelick V. Effect of zinc pretreatment on pulmonary
endothelial cells in vitro and pulmonary function in a porcine model of
endotoxemia. Journal of Surgical Research 123(2): 251-6. 2005
Krones CJ, Klosterhalfen B, Fackeldey V, Junge K, Rosch R, Schwab R,
Stumpf M, Schumpelick V. Deleterious effect of zinc in acute endotoxemia.
Journal of Investigative Surgery 17: 249-56. 2004

7 Literaturverzeichnis
69
Lai EC. Notch signalling: control of cell communication and cell fate.
Development 131: 965-973. 2004
Lansdown AB. Zinc in the healing wound. The Lancet 347: 706-707, 1996.
Lawrence WT. Physiology of the acute wound. Clinics in Plastic Surgery: 25(3):
321-40. 1998
Lee MH, Murphy G. Matrix metalloproteinases at a glance. Journal of Cell
Science 117: 4015-4016. 2004
Lindner J. Morphologie und Biochemie der Wundheilung. Langenbeck`s
Archives of Surgery 358: 153-160 (Kongressbericht 1982). 1995
Lodish H, Berk A, Kaiser CA, Krieger M, Scott MP, Bretscher A, Ploegh H,
Matsudeira P. Molecular Cell Biology. Sixth Edition. W.H. Freeman and
Company, New York. 2008
Lynch SE, Nixon JC, Colvin RB, Antoniades HN. Role of platelet-derived growth
factor in wound healing: Synergistic effects with other growth factors.
Proceedings of the National Academy of Sciences of the United States of
America 84: 7696-7700. 1987
McKaig BC, McWilliams D, Watson SA, Mahida YR. Expression and Regulation
of Tissue Inhibitor of Metalloproteinase-1and Matrix Metalloproteinases by
Intestinal Myofibroblasts in Inflammatory Bowel Desease. American
Journal of Pathology 162: 1355-1360. 2003
Mitsiadis TA, Fried K, Goridis C. Reactivation of Delta–Notch Signaling after
Injury: Complementary Expression Patterns of Ligand and Receptor in
Dental Pulp. Biochemical Pharmacology. 61: 1439-1448. 2001
Noggle SA, Weiler D, Condie BG. Notch Signaling Is Inactive but Inducible in
Human Embryonic Stem Cells. Stem Cells 24: 1646-1653. 2006
Oberleas D, Seymour JK, Lenaghan R, Hovanesian J, Wilson RF, Prasad AS.
Effect of Zinc Deficiency on Wound-Healing in Rats. The American Journal
of Surgery 121: 566-68. 1971
Olender S, Maldonado H, Randriamananjo S, Chalkiadakis G, Lambert A,
Wittmann T, Volkmar P, Grenier JF. Variations of magnesium, iron, copper
and zinc during the colonic wound healing process: experimental study on
rabbits. 1: European Surgical Research 24(5):283-90. 1992

7 Literaturverzeichnis
70
Ouko L, Ziegler TR, Gu LH, Eisenberg LM, Yang VW. Wnt11 Signaling
Promotes Proliferation, Transformation and Migration of IEC6 Intestinal
Epithelial Cells. The Journal of Biological Chemistry 279 (25): 26707-
26715. 2004
Pfaffl MW, Windisch W. Influence of zinc deficiency on the mRNA expression of
zinc transporters in adult rats. Journal of Trace Elements in Medicine &
Biology. 17 (2): 97-106. 2003
Pinto D, Gregorieff A, Begthel H, Clewers H. Canonical Wnt signals are
essential for homöostasis of intestinal epithelium. Genes & Development
17: 1709-13. 2003
Prasad AS. Zinc: The Biology and Therapeutics of an Ion. Annals of Internal
Medicine. 125 (2): 142-43. 1996
Prasad AS, Beck FWJ, Endre L, Handschu W, Kukuruga M, Kumar G. Zinc
deficiency effects cell cycle and deoxythymidin kinase gene expression in
HUT-78 cells. Journal of Laboratory and Clinical Medicine 128: 51-60.
1996
Prasad AS, Oberleas D. Thymidine kinase activity and incorporation of
thymidine into DNA in zinc-deficient tissue. Journal of Laboratory and
Clinical Medicine 83: 634-9. 1974
Rhagow R. The role oft extracellular matrix in postinflammatory wound healing
and fibrosis. The Journal of the Federation of American Societies for
Experimental Biology 8: 823-831. 1994
Remmele W, Stegner, HE. Recommendation for uniform definition of an
immunoreactive score (IRS) for immunohistochemical estrogen receptor
detection (ER-ICA) in breast cancer tissue. Der Pathologe 8: 138-140.
1987
Rink L und Gabriel P. Zinc and the immune system. Proceedings of the
Nutrition Society 59: 541–552. 2000
Rink L und Haase H. Zinc homeostasis and immunity. Trends in Immunology
28: 1-4. 2007
Roche Lexikon Medizin (5. Auflage). Urban & Fischer. 2003, URL:
http://www.tk-online.de/rochelexikon/

7 Literaturverzeichnis
71
Roeb E, Behrmann I, Grötzinger J, Breuer B. An MMP-9 mutant without
gelatinolytic activity as a novel TIMP-1-antagonist. The Journal of the
Federation of American Societies for Experimental Biology 14(12): 1671-3.
2000
Sandstead HH, Smith JC Jr. Deliberations and evaluations of approaches,
endpoints and paradigms for determining zinc dietary recommendations.
Journal of Nutrition 126: 2410S-18S. 1996
Schumacher I, Lorenz D. Historische Entwicklung der Darmnaht. Der Chirurg
62: 71-74. 1991
Schultz GS, Ladwig G, Wysocki A. Extracellular matrix: Review of its roles in
acute and chronic wounds. URL:
http://www.worldwidewounds.com/2005/august/ Schultz/Extrace-Matric-
Acute-Chronic-Wounds.html. 2005
Schumpelick V, Bleese N, Mommsen, U. Kurzlehrbuch Chirurgie, Auflage 6.
2004
Senapati A, Thompson RP. Zinc deficiency and the prolonged accumulation of
zinc in wounds. British Journal of Surgery 72(7): 583-4. 1985
Sternlicht MD, Werb T. How matrix metalloproteinases regulate cell behaviour.
Annual Review of Cell and Developmental Biology 17: 463-516. 2001
Stojadinovic O, Brem H, Vouthounis C, Lee B, Fallon J, Stallcup M, Merchant A,
Galiano RD, Tomic-Canic M. Molecular Pathogenesis of Chronic Wounds:
The Role of ß-Catenin and c-myc in the Inhibition of Epithelialization and
Wound Healing. American Journal of Pathology 167: 59-69. 2005
Stumpf M, Klinge U, Mertens PR. Anastomosenheilung - Prognostische
Faktoren. Der Chirurg 75: 1056-1062. 2004
Stumpf M, Klinge U, Wilms A, Zabrocki R, Rosch R, Junge K, Krones C,
Schumpelick V. Changes of the extracellular matrix as a risk factor for
anastomotic leakage after large bowel surgery. Surgery. 137(2): 229-34.
2005
Sunderman FW Jr: The influence of zinc on apoptosis. Annals of clinical and
laboratory science 25 (2): 134-42. 1995
Takeyama H, Sawai H, Sato M, Akamo Y, Yamamoto M, Funahashi H, Manabe
T. Surgical endoscopy 21: 1891-1894. 2007

7 Literaturverzeichnis
72
Thorgeirsson UP, Yoshiji H, Sinha CC, Gomez DE. Breast cancer; tumor
neovasculature and the effect of tissue inhibitor of metalloproteinases-1
(TIMP-1) on angiogenesis. In Vivo 10(2): 137-44. 1996
Thornton FJ, Barbul A. Healing in the gastrointestinal tract. Surgical Clinics of
North America 77: 549-73. 1997
Truong-Tran AQ, Carter J, Ruffin RE, Zalewski PD. The role of zinc in caspase
activation and apoptotic cell death. Biometals 14: 315-30. 2001
Universität Bielefeld, Sonderforschungsbereich 549, URL: http://www.uni-
bielefeld.de/SFB549/. 2005
van Es JH, Clevers H. Notch and Wnt inhibitors as potential new drugs for
intestinal neoplastic disease. Trends in molecular medicine 11(11) 496-
502. 2005
Van Waart H, Birkedal-Hansen H. The cysteine switch: A priciple of regulation
of metalloproteinase activity with potential applicability to the entire matrix
metalloproteinase gene family. Proceedings of the National Acadamy of
Science 87(14): 5578–5582. 1990
Visse R, Nagase H. Matrix Metalloproteinases and Tissue Inhibitors of Metallo-
proteinase Inhibitors. Structure, Function and Biochemistry. Circulation
Research 92: 827-839. 2003
Waltenberger J, Jehle PM, Engele J, Kühl M, Wedlich D, Reinshagen M.
Wachstumsfaktormodulation als therapeutisches Prinzip; Deutsches
Ärzteblatt, Heft 51-52. 2001
Wang W, Campos AH, Prince CZ, Mou Y, Pollman MJ. Coordinate Notch-3-
Hairy-related Transkription Factor Pathway Regulation in Response to
Arterial Injury. The Journal of Biological Chemistry 277(26): 23165 -
23171. 2002
Wang T, Holt CM, Xu C, Ridley C, Jones RPO, Baron M, Trump D. Notch3
activation modulates cell growth behaviour and cross-talk to Wnt/TCF
signalling pathway. Cellular Signaling. 19 (12): 2458-2467. 2007
Willis S, Schumpelick V. Open colon surgery: Offene Kolonchirurgie. Der
Chirurg 76 (11): 1073-1081. 2005
Willis S, Stumpf M. Insuffizienzen nach Eingriffen am unteren
Gastrointestinaltrakt. Der Chirurg 75: 1071-1078. 2004

7 Literaturverzeichnis
73
Windisch W. Development of zinc deficiency in zinc labelled, fully grown rats as
a model for adult individuals. Journal of Trace Elements in Medicine and
Biology 17 (2): 91-96. 2003
Windisch W, Kirchgessner M. Zinc absoption and excretion in adult rats at zinc
deficiency induced by dietary phytate additions: 1. Quantitative zinc
metabolism of Zink- labelled adult rats at zinc deficiency. Journal of Animal
Physiology and Animal Nutrition 82: 106-15. 1999
Xie S, Lin H, Sun T, Arlinghaus RB. Jak2 is involved in c-myc induction by Bcr-
Abl. Oncogene 21, 7137-7146. 2002
Young DA, Billingham O, Sampieri CL, Edwards DR, Clark IM. Differential
effects of histone deacetylase inhibitors on phorbol ester- and TGF-ß1
induced murine tissue inhibitor of metalloproteinase-1 gene expression.
The Journal of the Federation of American Societies for Experimental
Biology 272: 1912-1926. 2005
Zalewski PD, Forbes IJ, Seamark RF, Borlinghaus R, Betts WH, Lincoln SF,
Ward AD. Flux of intracellular labile zinc during apoptosis (gene-directed
cell death) revealed by a specific chemical probe, Zinquin. Chemical
Biology 1: 153-161, 1994.
Zhurinsky J, Shtutman M, Ben-Ze´ev A. Plakoglobin and ß-catenin: protein
inter-actions, regulation and biological roles. Journal of Cell Science 113:
3127-3139. 2000

8 Danksagung
74
8 Danksagung
Mein erster und besonderer Dank gilt Herrn Professor Dr. med. Dr. h.c. Volker
Schumpelick, Direktor der chirurgischen Klinik der RWTH Aachen, für die
freundliche Überlassung des Dissertationsthemas.
Ganz herzlich bedanke ich mich bei Dr. med. Marcel Binnebösel und PD Dr.
med. Carsten J. Krones für die Betreuung dieser Arbeit, für Ihre Anregungen
und Korrekturen. Sie standen mir stets mit Rat und Tat zur Seite.
Mein weiterer Dank gilt den Mitarbeitern des Joint Institute For Surgical
Research Aachen-Moskau unter der Leitung von Professor Alexander Öttinger
und Dr. med. Jochen Grommes, die maßgeblich am Gelingen dieses Projekts
beteiligt waren.
Großer Dank gilt Frau Ellen Krott für die fachkundige Anleitung in der
Laborarbeit und ihre Hilfe bei der Anfertigung der histologischen Präparate und
Färbungen.
Schließlich möchte ich mich bei meinem Freund und meiner Familie für ihre
Unterstützung und ihr Verständnis bedanken. Mein Dank gilt außerdem Frau
Dr. rer. nat. Maritta Jacobs, die durch ihre Anregungen gute Impulse für das
Gelingen dieser Arbeit gegeben hat.

9 Datenaufbewahrung
75
9 Datenaufbewahrung
Erklärung § 5 Abs. 1 zur Datenaufbewahrung
Hiermit erkläre ich, dass die dieser Dissertation zu Grunde liegenden
Originaldaten
bei mir,
Marion Franken, Thingstraße 12, 41812 Erkelenz
hinterlegt sind.

10 Lebenslauf
76
10 Lebenslauf
PERSÖNLICHES
Name Marion Franken
Geburtsdatum 07.07.1981
Geburtsort Erkelenz
Eltern Karin Franken, geb. Wirth, Lehrerin
Hans-Ludwig Franken, Diplom-Agraringenieur
Familienstand ledig
SCHULISCHE AUSBILDUNG
1988 -1992 Katholische Grundschule Kückhoven
1992 -2001 Cornelius-Burgh-Gymnasium, Erkelenz
07/2000 Praktikum bei Caterpillar in Bury St. Edmunds/ UK
2001 Abitur
BERUFLICHE AUSBILDUNG
10/2001 Beginn des Studiums der Humanmedizin an der
RWTH Aachen
09/2003 Physikum
03/2004 - 04/2004 Famulatur Innere Medizin, St.Theresien-
Krankenhaus Nürnberg
08/2004 - 09/2004 Famulatur Psychiatrie, Uniklinikum der RWTH
Aachen
02/2005 - 02/2005 Famulatur Gynäkologie, Weyertaler Krankenhaus in
Köln
03/2005 - 04/2005 Famulatur Radiologie, Gemeinschaftspraxis Drs.
Bruners, Esser, Kick, Mauer in Erkelenz

10 Lebenslauf
77
08/2005 - 08/2005 Famulatur Augenheilkunde, Gemeinschaftspraxis
Drs. Neuhann, Remky, Frei in München
07/2007 - 08/2007 Famulatur im Rhein-Valley-Hospital, Nakuru, Kenia
12/2006 PJ-Tertial Innere Medizin, Krankenhaus Bethesda,
Mönchengladbach
04/2007 PJ-Tertial Chirurgie, Krankenhaus Bethesda,
Mönchengladbach
07/2007 PJ-Wahlfach Anästhesie, Universiteit St.Radboud,
Nijmegen, Niederlande
01/2008 Amerikanisches Examen, USMLE Step 1,
Amsterdam, Niederlande
04/2008 Schriftliches Hammerexamen, Aachen
05/2008 Amerikanisches Examen, USMLE Step 2, München
06/2008 Mündliche Prüfung im Krankenhaus Bethesda,
Mönchengladbach, Erlangung der Approbation
07/2008 - heute Assistenzärztin im Hermann-Josef-Krankenhaus
Erkelenz, Innere Medizin