effects of the fibroblast activation protein on the invasion and migration of gastric cancer
TRANSCRIPT

Experimental and Molecular Pathology 95 (2013) 350–356
Contents lists available at ScienceDirect
Experimental and Molecular Pathology
j ourna l homepage: www.e lsev ie r .com/ locate /yexmp
Effects of the fibroblast activation protein on the invasion and migrationof gastric cancer
Rui-FenWang a, Li-Hong Zhang b, Li-Hui Shan c,Wen-Guang Sun d, Cui-Cui Chai c, Hong-MeiWu c, Juan C. Ibla e,Li-Feng Wang a,⁎, Jia-Ren Liu e
a Department of Pathology, Xin Hua Hospital affiliated to Shanghai JiaoTong University School of Medicine, Shanghai 200092, People's Republic of Chinab Department of Radiology, The First Clinical College of Harbin Medical University, 23 YouZheng Street, NanGang District, Harbin 150001, People's Republic of Chinac Department of Pathology, The First Clinical College of Harbin Medical University, 23 YouZheng Street, NanGang District, Harbin 150001, People's Republic of Chinad Department of Clinic Nutrition, Affiliated Sixth People's Hospital of Shanghai JiaoTong University, 600 Yi-shan Road, Shanghai 200233, People's Republic of Chinae Boston Children's Hospital and Harvard Medical School, 300 Longwood Ave, Boston, MA 02115, United States
⁎ Corresponding author.E-mail addresses: [email protected] (L.-F. Wang), Jia-
(J.-R. Liu).
0014-4800/$ – see front matter © 2013 Elsevier Inc. All rihttp://dx.doi.org/10.1016/j.yexmp.2013.10.008
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 25 September 2013and in revised form 17 October 2013Available online 25 October 2013Keywords:Gastric cancerFibroblast activation protein (FAP)Cancer-associated fibroblasts (CAFs)MGC-803 cells
Objective: Cancer-associated fibroblasts (CAFs) are one of the most important components of tumor microenvi-ronment. CAFs are believed to play an important role in tumor invasion andmetastasis. Recently, fibroblast acti-vation protein (FAP), a type II integral membrane glycoprotein belonging to the serine protease family, hasemerged as a specific marker of CAFs. FAP was overexpressed in stromal fibroblasts of solid malignancies,however, the role of FAP on the process of invasion and metastasis of gastric carcinomas is still unknown.Methods: Expression of FAP level was detected by immunohistochemistry in 60 gastric cancer surgical specimens(28 with omentum metastasis and 32 without), 20 normal human gastric tissues and omentum of 10 non-neoplastic gastric diseases. Fibroblasts were isolated from patient's tissues in the distal normal zones andtumor zones respectively, which were correspondingly designated as normal zone fibroblasts (NFs) and
cancer-associated fibroblasts (CAFs). To explore the effects of FAP on NFs or CAFs, fibroblasts were co-culturedwith human gastric cancer cell line MGC-803 cells. The ability of invasion and migration of MGC-803 cells wasevaluated after transfecting FAP siRNA into CAFs of gastric carcinomas.Results: We investigated the level of expression of FAP in surgical specimens, and found overexpressed in CAFsand non-expressed in NFs. Expression of FAP level in CAFs is significantly associated with Lauren classification,the degree of differentiation, depth of tumor invasion and TNM stage, but it is not correlated to age and genderin gastric carcinoma patients. There was positive correlation between the FAP level withmetastasis to the omen-tum (p b 0.05, R2=0.2736, p b 0.05, R2=0.1479). In addition, the invasion and migration abilities of MGC-803cells were significantly increasedwhen cellswere co-culturedwith CAFs. On the other hand, invasion andmigra-tion abilities were significantly decreased by 46.9 and 50.3%, respectively, after knocking down FAP in CAFs.Further, NFs did not have appreciable effect on the invasion and migration of MGC-803 cells.Conclusions:Ourfindings showed that FAPwas overexpressed inCAFs of gastric carcinomas, and siRNA-mediatedknock down of FAP significantly suppressed invasion and migration of MGC-803 cells. FAP may be an importantregulator in the invasion and migration of gastric cancer and may provide a novel therapeutic target in gastriccarcinomas.© 2013 Elsevier Inc. All rights reserved.
Introduction
Gastric cancer is the second leading cause of death due to cancerworldwide. The 5-year survival rate is 30–40%, with a poor prognosisfor advanced tumor (Cappellani et al., 2010; Kim et al., 2009). Thehigh mortality is closely related to tumor invasion and metastasis. Gen-erally, the complex process of metastasis formation can be divided intoseveral stages: a) migration from the primary tumor, invasion of the
ghts reserved.
surrounding tissue and its extracellular matrix (ECM); b) intravasationinto the circulation or the lymphatic system via transmigration throughthe endothelial lining and the basement membrane; and c) extravasa-tion and metastasis formation at target sites (Hugo et al., 2007).
Previous studies have indicated that tumor progression is not con-trolled by tumor cells independently, and it is also closely related tothe tumor stroma. As the most abundant cells in tumor stroma,cancer-associated fibroblasts (CAFs) have distinctly different morpho-logical and biological characteristics compared with normal fibroblasts(NFs). CAFs may promote tumorigenesis and progression through mul-tiple mechanisms, including increased angiogenesis, proliferation, inva-sion, and inhibition of tumor cell death. These effects are mediated

351R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
through the expression and secretion of numerous growth factors, cyto-kines, proteases, and extracellularmatrix proteins (Brennen et al., 2012;Ishii et al., 2010; Orimo et al., 2005). A key characteristic of CAFs is theexpression of fibroblast activation protein (FAP). FAP is a type II integralmembrane serine protease of the prolyl oligopeptidase family andclassified into the dipeptidyl peptidase (DPP) subfamily. The crystalstructure analysis of FAP has documented that the enzyme exists as ahomodimer and that dimerization is necessary for enzymatic function(Aertgeerts et al., 2005). It has been reported that FAP is overexpressedin cancer-associated fibroblasts in many types of carcinomas, includingcolorectal, ovarian, breast, bladder, and lung cancer (Cheng et al., 2002;Tahtis et al., 2003). FAP overexpression has been associated with anoverall poorer prognosis in many cancers, including pancreatic (Cohenet al., 2008), hepatocellular (Ju et al., 2009), colon (Saigusa et al.,2011), ovarian (Zhang et al., 2007), and gastrointestinal carcinomas(Saadi et al., 2010). However, the effect of FAP on gastric cancer progres-sion still remains unclear. Our previous study reported a high level ofFAP in the invasion front of gastric cancer (Shan et al., 2012). Thus, theobjective of this study was to further explore the effects of FAP on theinvasion and migration of gastric cancer by the genetic knock-downFAP in CAFs.
Materials and methods
Tissue specimens
All samples used in this studywere surgical specimens frompatientstreated at the First Clinical College ofHarbinMedical University (Harbin,China) from 2008 to 2010 after informed consent. Patients who had re-ceived preoperative radiotherapy or chemotherapywere excluded fromthis study. Sixty cases of gastric cancer were selected as the experimen-tal group (28 with omentum metastasis and 32 without omentummetastasis), 20 normal gastric tissues and 10 non-neoplastic gastric dis-eases were selected as control groups. The patients' mean age was 51.5(range, 21–72) years. Of the 60 gastric cancers, the histologic classifica-tion was characterized according to Lauren classification, and the TNMstage was done according to AJCC/UICC (American Joint CancerCommittee/Union for International Cancer Control, 2009 edition).
Immunohistochemistry (Liu et al., 2008)
Tissue sections (4μm) were prepared from formalin-fixed paraffin-embedded blocks. Fibroblast activation protein (FAP) was determinedby immunostaining. Briefly, the deparaffin sections were quenchedwith 3% H2O2 for 10min. After washing with phosphate-buffered saline(PBS), the sections were incubated in 5% bovine serum albumin for20min, followed by incubation with FAP primary monoclonal antibody(2 μg/mL) (R&D Systems, Minneapolis, MN) overnight at 4 °C and sec-ondary antibodies for 40 min at room temperature. The location ofFAPwas visualized by 3,3-diaminobenzidine (Slack et al., 2007) solutionfor 3–5 min. Finally, the slides were washed with water before beingcounterstainedwith hematoxylin. PBSwas substituted for primary anti-body as a negative control. FAP expressionwas quantified as the relativepercentage of the stroma area with positive staining area in slides. Thedegree of FAP staining in gastric cancer stroma was classified intothree groups (Sugiura et al., 2009): +++, strong staining in N50% ofstroma fibroblasts; ++, moderate staining in N50% of stroma fibro-blasts; and +, faint or weak staining in N50% of stroma fibroblasts.
Isolation of primary CAFs and NFs
Tissue samples corresponding to the tumor zone (tissue within thetumor boundary) and the normal zone (more than 5 cm far from thetumor zone) were minced into 1–2 mm3 fragments, washed twice inantibiotic-containing phosphate-buffered saline (PBS) and disaggre-gated overnight in serum-free DMEM with 0.1% collagenase at 37 °C
on a rotator. After 24h, the epithelial cells were separated from the stro-mal cells by differential centrifugation, as described by Speirs et al.(1996). Stromal cells were washed twice in antibiotic-containing PBSand plated in 35 mm dishes at 37 °C in an incubator containing 5%CO2. The CAFs were isolated from tumor zone and NFs from normalzones. All experiments were performed using fibroblast cultured for3–10 passages.
Characterization of primary fibroblasts (Zhang et al., 2012)
Cultured cells were plated in 24-well glass chambers, rinsed twicewith cold PBS, fixed in 4%paraformaldehyde for 20min at room temper-ature and permeabilized with 0.1% Triton X-100 for 30min on ice. Pri-mary FAP antibody solutions (R&D Systems, Inc.) were added to eachchamber and the slides were incubated overnight at 4°C. After washingwith PBS, the cells were incubated with a FITC-conjugated second anti-body (1:100) and a TRITC-conjugated second antibody (1:100) for 1h atroom temperature. The cells were then covered with a coverslip. Theslides were observed under an immunofluorescent microscope.
Small interfering RNA transfection
CAFs and NFs were recovered and cultured in DMEM containing of20% FBS without penicillin–streptomycin in 24-well plates (0.5 × 105
cells/well). The cells were incubated overnight. The following day,10 nmol/L siRNA of FAP-2252 was transiently transfected into cellsusing siLentFect™ Lipid Reagent (BIO-RAD) in Opti-MEM. Non-targeting siRNA labeled by FAM was regarded as the negative control.No siRNAwas transfected into cells in blank control group. The efficien-cy of transfectionwas evaluated by counting the number of cells labeledby FAM in a fluorescence microscope after 24h.
Migration and invasion assays (Li et al., 2011)
The gastric cancer cell line MGC-803 was purchased from the ChinaCenter for Type Culture Collection, Harbin Medical University (Harbin,China). For cell migration assay, MGC-803 cells (2× l04 cells/well) wereseeded on transwell inserts with 8-μm pores (Corning Incorporated,Corning, NewYork, USA) and cultured in serum-free DMEM. NFs orCAFs (5 × l04 cells/well) were transiently transfected with siRNA ofFAP-2252 or without siRNA for 24h andwere placed in the lower cham-ber as chemoattractant. After 24h of incubation, the cells which did notmigrate through the pores were removed by scraping the membranewith a cotton swab. Cells which migrated through the membraneswere fixed in 95% ethanol and stained with crystal violet. For cell inva-sion assay, cellsmustmigrate through an extracellularmatrix (ECM)bar-rier by enzymatic degradation. Thus, the transwell inserts were coatedwith a uniform layer of 50μL of a 1:3 dilution ofMatrigel basementmem-brane matrix (BD Biosciences, Bedford, Massachusetts, USA) per well.MGC-803 cells were seeded on transwell inserts (2× l04 cells/well) andcultured for 24 h. Invasive cells that penetrated through the pores andmigrated to the underside of the membrane were stained with crystalviolet. The number of cells was counted under a microscope (×200) infive random fields by two independent observers with double-blindedsamples.
Western blotting (Dong et al., 2013)
NFs or CAFs were cultured to 100% confluence, transfected, washed,and then serum-starved in RPMI with 0.1% bovine serum albumin for24 h. The cells were lysed in ice-cold radioimmunoprecipitation assaybuffer. Cell lysates were clarified by centrifugation at 12,000 rpm for10 min and protein concentrations were measured using the BCA™Protein Assay Kit. A total of 30 μg of protein was separated by an 8%SDS-Page gel and transferred to a polyvinylidene difluoride (PVDF)membrane. The membrane was blocked for 90 min with 5% milk in
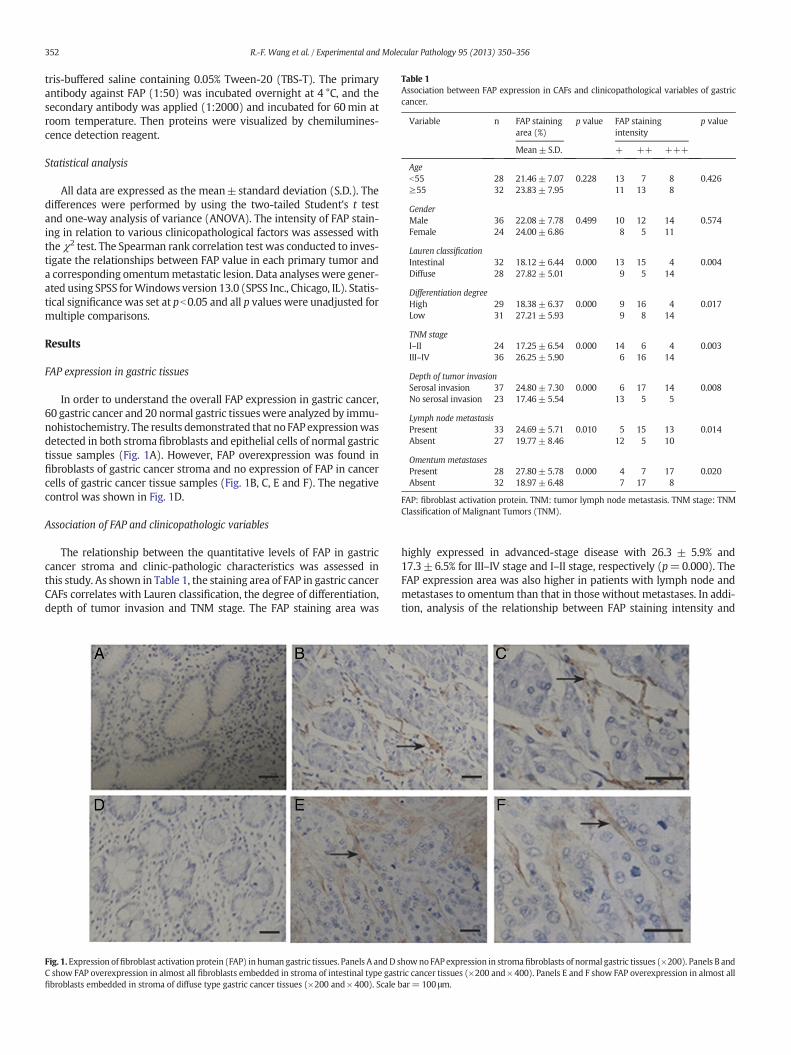
Table 1Association between FAP expression in CAFs and clinicopathological variables of gastriccancer.
Variable n FAP stainingarea (%)
p value FAP stainingintensity
p value
+ ++ +++Mean±S.D.
Ageb55 28 21.46± 7.07 0.228 13 7 8 0.426≥55 32 23.83± 7.95 11 13 8
GenderMale 36 22.08± 7.78 0.499 10 12 14 0.574Female 24 24.00± 6.86 8 5 11
Lauren classificationIntestinal 32 18.12± 6.44 0.000 13 15 4 0.004Diffuse 28 27.82± 5.01 9 5 14
Differentiation degreeHigh 29 18.38± 6.37 0.000 9 16 4 0.017Low 31 27.21± 5.93 9 8 14
TNM stageI–II 24 17.25± 6.54 0.000 14 6 4 0.003III–IV 36 26.25± 5.90 6 16 14
Depth of tumor invasionSerosal invasion 37 24.80± 7.30 0.000 6 17 14 0.008No serosal invasion 23 17.46± 5.54 13 5 5
Lymph node metastasisPresent 33 24.69± 5.71 0.010 5 15 13 0.014Absent 27 19.77± 8.46 12 5 10
Omentum metastasesPresent 28 27.80± 5.78 0.000 4 7 17 0.020Absent 32 18.97± 6.48 7 17 8
FAP: fibroblast activation protein. TNM: tumor lymph node metastasis. TNM stage: TNMClassification of Malignant Tumors (TNM).
352 R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
tris-buffered saline containing 0.05% Tween-20 (TBS-T). The primaryantibody against FAP (1:50) was incubated overnight at 4 °C, and thesecondary antibody was applied (1:2000) and incubated for 60min atroom temperature. Then proteins were visualized by chemilumines-cence detection reagent.
Statistical analysis
All data are expressed as the mean±standard deviation (S.D.). Thedifferences were performed by using the two-tailed Student's t testand one-way analysis of variance (ANOVA). The intensity of FAP stain-ing in relation to various clinicopathological factors was assessed withtheχ2 test. The Spearman rank correlation test was conducted to inves-tigate the relationships between FAP value in each primary tumor anda corresponding omentummetastatic lesion. Data analyses were gener-ated using SPSS forWindows version 13.0 (SPSS Inc., Chicago, IL). Statis-tical significance was set at pb0.05 and all p values were unadjusted formultiple comparisons.
Results
FAP expression in gastric tissues
In order to understand the overall FAP expression in gastric cancer,60 gastric cancer and 20 normal gastric tissueswere analyzed by immu-nohistochemistry. The results demonstrated that no FAP expressionwasdetected in both stroma fibroblasts and epithelial cells of normal gastrictissue samples (Fig. 1A). However, FAP overexpression was found infibroblasts of gastric cancer stroma and no expression of FAP in cancercells of gastric cancer tissue samples (Fig. 1B, C, E and F). The negativecontrol was shown in Fig. 1D.
Association of FAP and clinicopathologic variables
The relationship between the quantitative levels of FAP in gastriccancer stroma and clinic-pathologic characteristics was assessed inthis study. As shown in Table 1, the staining area of FAP in gastric cancerCAFs correlates with Lauren classification, the degree of differentiation,depth of tumor invasion and TNM stage. The FAP staining area was
Fig. 1.Expression offibroblast activation protein (FAP) inhuman gastric tissues. Panels A andD sC show FAP overexpression in almost all fibroblasts embedded in stroma of intestinal type gastfibroblasts embedded in stroma of diffuse type gastric cancer tissues (×200 and× 400). Scale b
highly expressed in advanced-stage disease with 26.3 ± 5.9% and17.3±6.5% for III–IV stage and I–II stage, respectively (p=0.000). TheFAP expression area was also higher in patients with lymph node andmetastases to omentum than that in those without metastases. In addi-tion, analysis of the relationship between FAP staining intensity and
howno FAP expression in stromafibroblasts of normal gastric tissues (×200). Panels B andric cancer tissues (×200 and×400). Panels E and F show FAP overexpression in almost allar= 100 μm.

353R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
clinic-pathological factors also showed a significant association withLauren classification, degree of differentiation, depth of tumor invasion,and TNMstage. No correlationwas found between the staining intensityof FAP and age or gender (Table 1).
Identification of primary NFs and CAFs
The fibroblasts from the normal and tumor zone tissues were desig-nated to NFs and CAFs. In our study, NFs and CAF populations were suc-cessfully isolated from gastric tissues of the same patient. Fibroblastshad the typical characteristics: a long spindle-like morphology, strongexpression of the fibroblastic marker (vimentin) but negative expres-sion for epithelial marker (cytokeratin) (Fig. 2A). To identify the NFsand CAFs, FAP expression was determined by immunofluorescence.The results showed that FAP protein was overexpressed in CAFs when
Fig. 2. Characteristics of primary fibroblasts from the different zones of human gastrictumor tissue. (A) NFs and CAFs were examined expression of vimentin and cytokeratinby immunocytochemistry. The fibroblasts fromNFs andCAFs had a long, spindle-likemor-phology and the immunostaining of vimentin was positive. These cells were negativecytokeratin expression. (B) NFs and CAFs were universally expressed FAP by immunoflu-orescence. FAP was significantly overexpressed in CAFs compared with the levels in NFs.(C) Flow cytometry results showed that FAP was variably expressed in the isolated NFsand CAFs, with the higher expression in CAFs. Bar=100 μm.
compared to NFs (Fig. 2B). Additional results from flow cytometry alsoshowed an overexpression of FAP in CAFs (Fig. 2C).
FAP siRNA inhibited the invasion and migration of gastric cancer cellsin vitro
MGC-803 cells co-cultured with CAFs and NFs showed increased in-vasion and migration in comparison with MGC-803 cells alone. CAFssignificantly increased the invasion and migration of MGC-803 cellswhen compared to NFs (Fig. 3A–D) (pb 0.05). In addition, when siRNAagainst FAP was transfected into CAFs, MGC-803 cells were decreasedby 46.9% when co-cultured with the CAFs in comparison with the con-trol group. NFs transfected with FAP siRNA did not increase MGC-803cell invasion when compared to the control group (Fig. 3A and B). Thesame pattern was also observed in the migration of MGC-803 cells. Mi-gration rate of MGC-803 cells co-cultured with CAFs was decreased by50.3% when compared to the control group (Fig. 3C and D) (p b 0.05).Western blot analysis using mAb anti-FAP was performed on samplesthat were suspended in SDS sample buffer but not boiled to preservethe 170kDa FAP dimer. Dimer (170kDa) of FAP is essential to its enzy-matic activity. Our result showed the high expression of FAP in CAFs.The expression of FAP (170 kDa and 97 KDa dimer) was not detectedin CAFs or NFs when transfected with FAP-2252 siRNA (Fig. 3E and F)(pb0.05).
FAP expression in omentum
To explore the relationship between FAP overexpression andmetas-tases to omentum, FAP expression intensity in omentumwasmeasuredin 50 patients, including 10 non-neoplastic gastric diseases, 20 gastriccancers with omentum metastasis, and 20 gastric cancers withoutomentummetastasis. No FAP expression was detected in stromal fibro-blasts of omentum tissue samples in the non-neoplastic gastric diseases(Fig. 4A). In omentum tissue sampleswith gastric cancer cell metastasis,the tumor cells formed solid invading nests, surrounded by omentumadipose and CAFs whichwas expressed as FAP protein. With the omen-tummetastasis enlarged, the stroma cells were found to have with highexpression of FAP protein (Fig. 4B). One striking finding was that someomentum tissue samples free of gastric metastasis also demonstratedthe overexpression of FAP protein (Fig. 4C). The correlation of FAP over-expression in primary tumor CAFs and corresponding omentummetastasis lesions was also analyzed in this study. Our results showedsignificant positive correlation between FAP expression in primarytumor CAFs and corresponding omentum with/without gastric cancermetastasis (Fig. 4D–E).
Discussion
CAFs play important roles in tumor invasion and metastasis (Cirriand Chiarugi, 2011; Kunz-Schughart and Knuechel, 2002a; Kunz-Schughart and Knuechel, 2002b). A key characteristic of CAFs is the ex-pression of FAP (O'Brien and O'Connor, 2008). Although FAP was foundto be overexpressed in stroma of multiple cancers, there is little infor-mation about FAP expression in gastric cancer. In the present study,FAP expressionwas detected in gastric cancer stroma and its correlationexploredwith clinic-pathological characteristics. Our results showed noFAP expression in stroma of normal gastric tissue samples, howeverthere was overexpression in CAFs of gastric cancer tissue samples. FAPimmunostaining intensity was associatedwith Lauren classification, de-gree of differentiation, depth of tumor invasion and TNMstage. Our pre-vious findings showed that stroma fibroblasts from gastric cancerinvasion front (the interface zone fibroblasts, INFs) had strong positiveFAP expression. The degree of FAP expression was higher in INFs thanthat in NFs and CAFs. In addition, the degree of FAP expression in INFswas significantly related to Lauren classification, degree of differentia-tion, depth of tumor invasion and TNM stage (Shan et al., 2012). It

Fig. 3. Effects of FAP siRNA in CAFs or NFs on invasion and migration of MGC-803 cells. Invading and migrating cells on the reverse side of the membranes were stained by crystal violet.CAFs transfectedwith FAP-2252 siRNA showed a suppressed invasion andmigration of MGC-803 cell when compared to the control group (pb0.05). However, NFs transfectedwith FAP-2252 siRNAdid not increase invasion andmigration ofMGC-803 cell (Fig. 3Aand C,×200). These results showed that the FAP siRNA inCAFs remarkably suppressed invasion andmigrationofMGC-803 cells comparedwith control (Fig. 3B andD). Both CAFs andNFs increased invasion andmigration ofMGC-803 cells; the CAFshad a far greater potential to increase invasion andmigration ofMGC-803 cells, comparedwith NFs (Fig. 3A–D) (pb0.05). The results also revealed that expression of FAP level in CAFs is stronger than those inNFs. FAP (170kDa and 97KDadimer) were not detected in CAFs or NFs transfected with FAP-2252 siRNA when compared to the control group (Fig. 3E, 3F) (p b 0.05).
354 R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
suggested that CAFs from the gastric cancer invasion front may supplytheir own optimal microenvironment for tumor cells invasion andmigration.
Fig. 4. FAP expression in omentum tissues of surgical specimens by immunostaining. (A) No FAPwith gastric cancer cells showed a large number of CAFs which was overexpressed as FAP (macells in patients with gastric carcinoma also have CAFs with FAP overexpression (×200 and ×expression in primary tumor and corresponding omentum with/without metastasis (p b 0.05)
CAFs and NFs were isolated from the tumor zone and normal zonerespectively of the same patients with gastric cancer. FAP was foundto be overexpressed in CAFs in comparison with NFs. CAFs significantly
expression in thefibroblasts of normal omentum tissues. (B) Omentum tissues implantedgnification × 200, inset: magnification ×400); (C) Omentum tissue free detectable cancer400); scale bar= 100 μm. (D–E) There was a significant positive correlation between FAP.

355R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
promoted the invasion and migration of MGC-803 cells when com-pared to NFs. Our previous study also found that INFs isolated fromthe gastric cancer invasion front had a high expression of FAP pro-tein when compared to CAFs and NFs. This indicated that INFshave a promoted effect on invasion and migration of MGC-803cells. To explore whether FAP was a regulator of gastric cancer cellinvasion and migration, FAP siRNA was transfected into CAFs by ge-netic knock-down FAP. It was found that the invasion and migrationof MGC-803 cells were significantly suppressed. We demonstratedthat CAFs could promote the invasion and migration of gastric can-cer cells by up-regulating FAP which may be a crucial factor intumor invasion and migration of gastric cancer. Other studies alsohave shown that FAP increased the invasion, proliferation and mi-gration of HO-8910PM ovarian cancer cells (Chen et al., 2009).FAP-expressing cells were an immune-suppressive component ofthe tumor microenvironment (Kraman et al., 2010). In anotherstudy, knock-down of FAP expression by siRNA lentiviral vectortransfection resulted in a low or no expression of FAP in CAFs to in-hibit cell growth. These results indicate that FAP was an importantregulator of the microenvironment in tumor formation (Lai et al.,2012). In the present study, a seemly paradox phenomenon regard-ing FAP overexpression appeared in the gastric cancer tissue of sur-gical specimens and NF from primary cell culture in vitro. We foundthat FAP expression was negative in fibroblasts of normal gastrictissues of surgical specimens, weakly positive expression in NFsfrom primary cell culture, and high expression of FAP in CAF. Theability of invasion and migration did not change when MGC-803cells were co-cultured with NF knock-down FAP expression.
In the present study, we describe an interesting phenomenon inwhich CAFs with FAP overexpression not only existed in omentumwith gastric cancer cells metastasis, but also existed in omentumof some patients without metastasis. This suggested that CAFs ar-rive at target organs before the implantation of cancer cells. TheCAFs in omentum may promote cancer cell adherence, allowingfor further growth, invasion, and metastases. We also found a sig-nificant positive correlation between FAP expression in primarytumor CAFs and corresponding omentum lesions with/without gas-tric cancer metastasis. A high concordance of FAP expression inpairs of primary tumor and their corresponding omentum metasta-sis may suggest that individual cancer in metastases sites will pro-duce their own optimal microenvironment to support migration. Ithas also been demonstrated that CAFs existed not only in omentumwith epithelial ovarian cancer (EOC) metastasis but also in omen-tum of patients without metastasis (Zhang et al., 2011). Anotherstudy showed that TGF-β1 (transforming growth factor-β1), HGF(hepatocyte growth factor) and MMP-2 (matrix metalloproteinase-2)may be involved in the adhesion and invasion of EOC cells, indicatingthat activated fibroblasts in omentum form an optimal environmentto promote EOC cells implantation (Cai et al., 2012). In the presentstudy, FAP expression in stroma of gastric cancers with omentummetastases and lymph node metastases was much higher thanthat of without metastases. This may be related to the collagenolyticactivity of FAP resulting in increased invasion and metastases by re-gional blood vessels and lymph nodes (Forssell et al., 2007; Maliket al., 2011).
In summary, CAFs can promote gastric cancer cell invasion and mi-gration by up-regulating FAP. Therefore, inhibition of FAP expressioncould be used as a newpotential therapeutic strategy against gastric can-cer. The FAP inhibitor PT630 has been found to have potent anticancereffects in several mouse models (Santos et al., 2009). Another anti-FAPantibody FAP5-DM1 can induce long-lasting inhibition of tumor growth(Ostermann et al., 2008). FAP-based pre-clinic drug strategy has beenshown as promising in achieving targeted delivery of anticanceragents (Liu et al., 2012). However, the effects of FAP on tumorgrowth and invasion, and the exact molecular mechanisms remainto need further study.
Conflict of interest statement
They declared no conflicts of interests.
Acknowledgments
This workwas supported by Natural Science Foundation of HeiLong-Jiang Province (D201103) and National Natural Science Foundationof China (No. 81072296). Li-Feng Wang and Jia-Ren Liu are co-corresponding authors. Rui-Fen Wang and Li-Hong Zhang are co-firstauthors.
References
Aertgeerts, K., et al., 2005. Structural and kinetic analysis of the substratespecificity of human fibroblast activation protein alpha. J. Biol. Chem. 280,19441–19444.
Brennen, W.N., et al., 2012. Rationale behind targeting fibroblast activation protein-expressing carcinoma-associated fibroblasts as a novel chemotherapeutic strategy.Mol. Cancer Ther. 11, 257–266.
Cai, J., et al., 2012. Fibroblasts in omentum activated by tumor cells promote ovariancancer growth, adhesion and invasiveness. Carcinogenesis 33, 20–29.
Cappellani, A., et al., 2010. Clinical and biological markers in gastric cancer: update andperspectives. Front. Biosci. (Schol. Ed.) 2, 403–412.
Chen, H., et al., 2009. TGF-beta induces fibroblast activation protein expression; fibroblastactivation protein expression increases the proliferation, adhesion, and migration ofHO-8910PM [corrected]. Exp. Mol. Pathol. 87, 189–194.
Cheng, J.D., et al., 2002. Promotion of tumor growth by murine fibroblast activationprotein, a serine protease, in an animal model. Cancer Res. 62, 4767–4772.
Cirri, P., Chiarugi, P., 2011. Cancer associated fibroblasts: the dark side of the coin. Am.J. Cancer Res. 1, 482–497.
Cohen, S.J., et al., 2008. Fibroblast activation protein and its relationship to clinical out-come in pancreatic adenocarcinoma. Pancreas 37, 154–158.
Dong, H.W., et al., 2013. beta-Ionone arrests cell cycle of gastric carcinoma cancer cells bya MAPK pathway. Arch. Toxicol. 87, 1797–1808.
Forssell, J., et al., 2007. Highmacrophage infiltration along the tumor front correlates withimproved survival in colon cancer. Clin. Cancer Res. 13, 1472–1479.
Hugo, H., et al., 2007. Epithelial–mesenchymal and mesenchymal–epithelial transitions incarcinoma progression. J. Cell. Physiol. 213, 374–383.
Ishii, G., et al., 2010. Fibroblasts associated with cancer cells keep enhanced migrationactivity after separation from cancer cells: a novel character of tumor educated fibro-blasts. Int. J. Oncol. 37, 317–325.
Ju, M.J., et al., 2009. Peritumoral activated hepatic stellate cells predict poor clinical out-come in hepatocellular carcinoma after curative resection. Am. J. Clin. Pathol. 131,498–510.
Kim, J.W., et al., 2009. Clinicopathological characteristics and predictive markers of earlygastric cancer with recurrence. J. Korean Med. Sci. 24, 1158–1164.
Kraman, M., et al., 2010. Suppression of antitumor immunity by stromal cells expressingfibroblast activation protein-alpha. Science 330, 827–830.
Kunz-Schughart, L.A., Knuechel, R., 2002a. Tumor-associated fibroblasts (part I): activestromal participants in tumor development and progression? Histol. Histopathol.17, 599–621.
Kunz-Schughart, L.A., Knuechel, R., 2002b. Tumor-associated fibroblasts (part II): func-tional impact on tumor tissue. Histol. Histopathol. 17, 623–637.
Lai, D., et al., 2012. Fibroblast activation protein regulates tumor-associated fibroblastsand epithelial ovarian cancer cells. Int. J. Oncol. 41, 541–550.
Li, Y., et al., 2011. gamma-Tocotrienol inhibits angiogenesis of human umbilical veinendothelial cell induced by cancer cell. J. Nutr. Biochem. 22, 1127–1136.
Liu, J.R., et al., 2008. beta-Ionone suppresses mammary carcinogenesis, proliferative activ-ity and induces apoptosis in the mammary gland of the Sprague–Dawley rat. Int.J. Cancer 122, 2689–2698.
Liu, R., et al., 2012. Fibroblast activation protein: a potential therapeutic target in cancer.Cancer Biol. Ther. 13, 123–129.
Malik, A.N., et al., 2011. Mitochondrial DNA as a non-invasive biomarker: accurate quan-tification using real time quantitative PCR without co-amplification of pseudogenesand dilution bias. Biochem. Biophys. Res. Commun. 412, 1–7.
O'Brien, P., O'Connor, B.F., 2008. Seprase: an overview of an important matrix serine pro-tease. Biochim. Biophys. Acta 1784, 1130–1145.
Orimo, A., et al., 2005. Stromal fibroblasts present in invasive human breast carcinomaspromote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion.Cell 121, 335–348.
Ostermann, E., et al., 2008. Effective immunoconjugate therapy in cancermodels targeting a serine protease of tumor fibroblasts. Clin. Cancer Res. 14,4584–4592.
Saadi, A., et al., 2010. Stromal genes discriminate preinvasive from invasive disease, pre-dict outcome, and highlight inflammatory pathways in digestive cancers. Proc. Natl.Acad. Sci. U. S. A. 107, 2177–2182.
Saigusa, S., et al., 2011. Cancer-associated fibroblasts correlate with poor prognosis in rec-tal cancer after chemoradiotherapy. Int. J. Oncol. 38, 655–663.
Santos, A.M., et al., 2009. Targeting fibroblast activation protein inhibits tumorstromagenesis and growth in mice. J. Clin. Invest. 119, 3613–3625.

356 R.-F. Wang et al. / Experimental and Molecular Pathology 95 (2013) 350–356
Shan, L.H., et al., 2012. Roles of fibroblasts from the interface zone in invasion, migration,proliferation and apoptosis of gastric adenocarcinoma. J. Clin. Pathol. 65, 888–895.
Slack, G.W., et al., 2007. Flow cytometric detection of ZAP-70 in chronic lymphocyticleukemia: correlation with immunocytochemistry and Western blot analysis. Arch.Pathol. Lab. Med. 131, 50–56.
Speirs, V., et al., 1996. Collagenase III: a superior enzyme for complete disaggregation andimproved viability of normal and malignant human breast tissue. In Vitro Cell. Dev.Biol. Anim. 32, 72–74.
Sugiura, T., et al., 2009. VEGF-C and VEGF-D expression is correlated with lymphatic ves-sel density and lymph nodemetastasis in oral squamous cell carcinoma: implicationsfor use as a prognostic marker. Int. J. Oncol. 34, 673–680.
Tahtis, K., et al., 2003. Expression and targeting of human fibroblast activation protein in ahuman skin/severe combined immunodeficient mouse breast cancer xenograftmodel. Mol. Cancer Ther. 2, 729–737.
Zhang, M.Z., et al., 2007. Expression of seprase in effusions from patients with epithelialovarian carcinoma. Chin. Med. J. (Engl.) 120, 663–668.
Zhang, Y., et al., 2011. Ovarian cancer-associated fibroblasts contribute to epithelial ovar-ian carcinoma metastasis by promoting angiogenesis, lymphangiogenesis and tumorcell invasion. Cancer Lett. 303, 47–55.
Zhang, H.W., et al., 2012. Toxicological insight from AP-1 silencing study on proliferation,migration, and dedifferentiation of rat vascular smooth muscle cell. Cardiovasc.Toxicol. 12, 25–38.