effects of serotonin on the intrinsic membrane properties of layer ii medial entorhinal cortex...
TRANSCRIPT

Effects of Serotonin on the Intrinsic Membrane Propertiesof Layer II Medial Entorhinal Cortex Neurons
Li Ma,1,2 Mark H. Shalinsky,1 Angel Alonso,1{ and Clayton T. Dickson2*
ABSTRACT: Although serotonin (5-HT) is an important neuromodula-tor in the superficial layers of the medial entorhinal cortex (mEC), thereis some disagreement concerning its influences upon the membraneproperties of neurons within this region. We performed whole cellrecordings of mEC Layer II projection neurons in rat brain slices inorder to characterize the intrinsic influences of 5-HT. In current clamp,5-HT evoked a biphasic response consisting of a moderately short la-tency and large amplitude hyperpolarization followed by a slowly devel-oping, long lasting, and small amplitude depolarization. Correspond-ingly, in voltage clamp, 5-HT evoked a robust outward followed by asmaller inward shift of holding current. The outward current evoked by5-HT showed a consistent current/voltage (I/V) relationship across cellswith inward rectification, and demonstrating a reversal potential thatwas systematically dependent upon the extracellular concentration ofK+, suggesting that it was predominantly carried by potassium ions.However, the inward current showed a less consistent I/V relationshipacross different cells, suggesting multiple independent ionic mecha-nisms. The outward current was mediated through activation of 5-HT1Areceptors via a G-protein dependent mechanism while inward currentswere evoked in a 5-HT1A-independent fashion. A significant proportionof the inward current was blocked by the Ih inhibitor ZD7288 andappeared to be due to 5-HT modulation of Ih as 5-HT shifted the activa-tion curve of Ih in a depolarizing fashion. Serotonin is thus likely toinfluence, in a composite fashion, the information processing of Layer IIneurons in the mEC and thus, the passage of neocortical information viathe perforant pathway to the hippocampus. VVC 2006 Wiley-Liss, Inc.
KEY WORDS: 5-HT; stellate cells; perforant path; GIRK; Ih
INTRODUCTION
The anatomical connections between the neocortex and the hippo-campal formation, which converge in the medial temporal lobe, have
been described as the brain’s hardware for specificforms of memories in humans and animals. Damageto structures within medial temporal lobe circuitryresults in prominent memory disturbances and am-nesic syndromes, both anterograde and retrograde(Scoville and Milner, 1957; Squire and Zola-Morgan,1996; Corkin et al., 1997; Nadel and Moscovitch,1997; Stefanacci et al., 2000). The entorhinal cortex(EC) is located in an anatomically nodal position inthis circuitry. Cells in the superficial layers of the ECreceive neocortical input that is, in turn, conveyed tothe hippocampal formation. Conversely, cells in thedeep layers of the EC receive the results of hippocam-pal processing and then provide a reciprocal output tothe same cortical regions that began this cortico-hip-pocampo-cortical loop (Lavenex and Amaral, 2000).
Anatomically, the EC appears to function as abidirectional relay for neocortical and hippocampalcommunications. However, it is more than a passiveconveyer of information since physiological analyseshave demonstrated that the input/output properties ofmost EC projection neurons are decidedly nonlinear,except perhaps for those found in Layer III (Alonsoand Llinas, 1989; Alonso and Klink, 1993; Jones,1994; Dickson et al., 1997; Gloveli et al., 1997; vander Linden and Lopes da Silva, 1998; Schmitz et al.,1998a; Hamam et al., 2000, 2002). Furthermore,both superficial and deep layers of the EC are subjectto neuromodulatory influences from numerous ascend-ing neuromodulatory systems (Swanson et al., 1987),which play a role in dynamically altering the physio-logical properties of single neurons and synaptic trans-mission in addition to the patterns of rhythmical syn-chronization of populations of neurons in the localnetwork (Vanderwolf, 1988; Bland and Colom, 1993;Pralong and Jones, 1993; Dickson et al., 1994, 1995,2000a; Pralong and Magistretti, 1995; Dickson andAlonso, 1997; Klink and Alonso, 1997a,b; Stenkampet al., 1998; Schmitz et al., 1998c; Chrobak et al.,2000; Caruana et al., 2006; Rosenkranz and Johnston,2006).
One such neuromodulator is serotonin (5-HT), amonoaminergic neurotransmitter, that derives fromthe raphe and other closely opposed nuclei in thebrainstem. The EC is prominently innervated by sero-tonergic fibers from both the dorsal and median raphenuclei and all layers except for Layer III have beenshown to display high densities of 5-HT binding sites
{Deceased.Mark H. Shalinsky is currently at Massachusetts General Hospital,Harvard University, Charlestown, MA, USA.Grant sponsor: Medical Research Council/Canadian Institutes of HealthResearch; Grant sponsor: Natural Sciences and Engineering ResearchCouncil; Grant number: RGPIN 249861.
1Department of Neurology and Neurosurgery, Montreal NeurologicalInstitute, McGill University, Montreal, Quebec, Canada; 2Departmentof Psychology, Centre for Neuroscience, University of Alberta, Edmon-ton, Alberta, Canada
*Correspondence to: Clayton Dickson, Department of Psychology, Centrefor Neuroscience, University of Alberta, Edmonton, AB, Canada, T6G2E9. E-mail: [email protected] for publication 3 November 2006DOI 10.1002/hipo.20250Published online 4 December 2006 in Wiley InterScience (www.interscience.wiley.com).
HIPPOCAMPUS 17:114–129 (2007)
VVC 2006 WILEY-LISS, INC.

(Kohler and Steinbusch, 1982; Kohler, 1984). The neurophys-iological effects of 5-HT in the EC are thus poised to shapeneuronal responsiveness in this region. Indeed, 5-HT has beensuggested to contribute in a state-dependent fashion to the hip-pocampal theta rhythm via the EC (Vanderwolf, 1988; Drin-genberg and Vanderwolf, 1998). For an opposing view, seeVertes and Kocsis (1997).
Initial studies of the neurophysiological effects of 5-HTapplication upon EC principal neurons (Sizer et al., 1992;Grunschlag et al., 1997) were consistent with intrinsic mem-brane effects found in other central nervous system cells, includ-ing a prominent and consistent hyperpolarization concomitantwith increased membrane conductance that was mediated bystimulation of 5-HT1A receptors. These studies also showedthat 5-HT depressed electrically mediated excitatory synapticresponses, responsiveness to exogenously applied glutamate, butdid not alter glutamate release or paired pulse facilitation,suggesting that any influence on excitatory neurotransmissionwere mediated by the postsynaptic influence of 5-HT. How-ever, later studies in a single laboratory (using higher concentra-tions of 5-HT up to 100 lM) focused exclusively on thepresynaptic depressant effects of 5-HT on excitatory synapticmechanisms (Schmitz et al., 1995a,b, 1997, 1998d, 1999).In fact, these researchers reported that intrinsic effects uponsuperficial EC principal neurons (including those from LayerIII) were inconsistent and small in amplitude (Schmitz et al.,1998b) in stark contrast to strong intrinsic effects in the hippo-campus (Schmitz et al., 1998c).
Given the obvious discrepancy of findings concerning intrin-sic versus synaptic effects of 5-HT in the EC, we sought toreinvestigate the intrinsic actions of this neuromodulator usingwhole cell techniques in Layer II medial EC principal neurons.Our findings conclusively demonstrate a strong and compositeaction of 5-HT upon the intrinsic membrane properties ofboth stellate and pyramidal neurons.
MATERIALS AND METHODS
General
Brain slices were prepared from male Long-Evans rats (100–200 g) using standard procedures (Alonso and Klink, 1993),which were approved by the Animal Care Committee of theMontreal Neurological Institute, and also following principlesoutlined by the Canadian Council for Animal Care, the Societyfor Neuroscience, and the American Physiological Society.Animals were quickly decapitated, and the brain was rapidlyremoved, blocked, and placed in a cold (4–68C) oxygenatedmodified Ringer’s solution containing: 124 mM NaCl, 5 mMKCl, 1.3 mM NaH2PO4, 2.0 mM CaCl2, 2.0 mM MgSO4,26 mM NaHCO3, and 10 mM glucose (pH 7.4, by saturatingwith 95% O2 and 5% CO2). Horizontal slices from the retro-hippocampal region were cut at a thickness of 400 lm using a
vibratome (Pelco, Redding, CA). After at least a 1-h recoveryperiod, during which they were submerged in the same oxygen-ated solution at room temperature (23–258C), individual sliceswere transferred to a recording chamber located on the stageof an upright microscope equipped with 603 water-immersionobjective (Zeiss, Oberkochen, FRG). Slices were submergedand superfused with a continuous supply of oxygenatedRinger’s solution maintained at room temperature. In mostcases, this superfusion Ringer’s solution was identical to thatdescribed earlier. In voltage clamp experiments, 1 lM tetrodo-toxin (TTX) was used to block sodium channels. In furtherexperiments, in which divalent cations were added, a modifiedRinger’s solution without phosphate and sulfate was used. Thiswas composed of (in mM): 125 NaCl, 5 KCl, 2 MgCl2,26 NaHCO3, 2 CaCl2, and 10 glucose, saturated with 95%O2 and 5% CO2 to maintain a pH of 7.4. Using DIC opticsand a near-infrared charge-coupled device camera (SonyXC-75), Layer II neurons of the EC could be visually distin-guished based on their shape and position within the slices(Dickson et al., 2000b).
Recordings
Somatic whole-cell recordings were performed under visual-ized control in all types of EC Layer II neurons using 4–7 MOelectrodes loaded with a pipette solution containing: 130 mMK-Gluconate, 5 mM NaCl, 2 mM MgCl2, 10 mM HEPES,0.5 mM EGTA, 2 mM ATP-Mg, and 0.4 mM GTP-Na(pH 7.2–7.4 with 1 M KOH; 290–300 mOsm). Membranevoltage signals were amplified using an Axoclamp-2A (AxonInstruments, Union City, CA) in current clamp (bridge) modewhile membrane current was measured using an Axopatch-200Bamplifier (Axon Instruments) in voltage clamp mode. The low-pass filter (�3 dB) was set at 1 kHz. Series resistance (between12 and 30 MO in our study) was compensated >40% with theamplifier’s built-in compensation section. The holding potentialwas always �60 mV, unless otherwise noted. The liquid junc-tion potential between pipette and bath solutions was empiri-cally determined using the method of Neher (1992) and wasfound to be �8 mV. However, no correction of membranepotential was applied since this correction is only consideredaccurate if the cell contents are effectively and completelydialyzed by the pipette solution (Barry and Lynch, 1991). Thisis unlikely to be the case in a neuron with long and extensiveprocesses like those of medial entorhinal cortex (mEC) Layer IIneurons, especially stellate neurons.
All recordings were stored on VHS tape by PCL coding usinga Neurocorder converter (Neurodata, NY). Voltage protocolswere commanded and current signals were acquired with aPentium PC interfaced to an Axon Digidata 1320 interface,using pClamp software (v8.1, Axon Instruments). Data storedon VHS tape were digitized off-line by sampling at 2 kHz in asimilar manner.
For current clamp recordings, the effects of 5-HT on mem-brane potential were monitored at resting membrane potentials
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 115
Hippocampus DOI 10.1002/hipo

and changes in input resistance were monitored by measuringthe voltage deflections caused by the intracellular injection of500–800 ms square hyperpolarizing current pulses applied at afrequency of 0.2 Hz. For voltage clamp recordings, the effectsof 5-HT were assessed at a holding potential of �60 mV, butmembrane current/voltage (I/V) relationships were assessed byapplying slow voltage ramp commands from �120 to �50 mV(0.02 V/s) at intervals of 2 min throughout the recordings.The I/V relationship of 5-HT-induced outward or inward cur-rents was computed by subtracting ramp-evoked current traceselicited during the respective steady state conditions following5-HT application from control traces elicited prior to 5-HTapplication and plotting the results with respect to voltage(Figs. 2 and 3).
To study the modulation of the delayed hyperpolarizationactivated inward rectifying current (Ih), a slightly differentprotocol was used. Once whole-cell configuration was achieved,slices were perfused at a high rate (>1.5 ml/min) with themodified Ringer’s solution without phosphate and sulfate towhich 2 mM BaCl and 1 lM TTX were added. Voltage clampprotocols described later were begun after the slice had beenperfused with this solution for >5 min.
The activation curve of Ih was estimated using a tail currentprotocol described previously by Dickson et al. (2000b). Briefly, afamily of tail currents were recorded at �80 mV followinga series of voltage-clamp steps to potentials ranging from �30to �130 mV. The peak amplitude of each of these tail currentswas normalized to the maximal value (Imax) and plotted as afunction of the membrane potential during the hyperpolarizingprepulse. The data were well fitted with a Boltzmann equationof the form
I=Imax ¼ 1þ eðVm�V1=2Þ=k� ��1
where Vm is the membrane potential of the prepulse, V½ themembrane potential at which Ih is half activated, k a slopefactor, and I the amplitude of the tail current recorded afterthe prepulse.
Traces were plotted and measurements made with the use ofClampfit 8.0 (Axon Instruments) and Origin (Microcal, North-ampton, MA) software packages. Average values were expressedas mean 6 standard deviation (SD) and statistical significancewas evaluated by means of the two-tail Student’s t test.
Drugs and Reagents
All chemicals and reagents were purchased from Sigma(St. Louis, MO), except TTX (Alamone Labs), 8-OH-DPAT(Tocris), and GTPgs (RBI). WAY-100635 was a generous giftfrom Dr. Pierre Blier (McGill University). All drugs weremixed fresh just prior to application via the bath perfusate.In some cases, ascorbic acid was added to solutions containing5-HT in order to prevent oxidation. No differences in 5-HT-induced effects were observed in these experiments.
RESULTS
Current Clamp Recordings of 5-HT-MediatedEffects
The effects of 5-HT on medial EC Layer II principal neu-rons were first investigated in current clamp mode. Cells werecharacterized as being either stellate or pyramidal based on acombination of observations: their morphology as visualized innear infrared DIC images (stellates tended to be slightly largerand multipolar) and their electrophysiological characteristics(stellates showed prominent subthreshold membrane potentialoscillations (Alonso and Klink, 1993)). Following the establish-ment of stable resting membrane potential values after forma-tion of the whole-cell recording configuration, 100 lM 5-HTwas bath applied for a duration of 1–2 min to both visuallyand electrophysiologically identified stellate and pyramidalneurons. As shown in Figure 1, for a stellate cell, this applica-tion led to a large amplitude and long-lasting hyperpolariza-tion, accompanied by a decrease in apparent input resistance asmeasured by decreasing deflections in the voltage response tohyperpolarizing constant current pulses (shown in Fig. 1 asexpansions for positions 1 and 2). This effect was identical forpyramidal neurons (not shown). The hyperpolarization wasmaximal at a latency of less than 2 min and gradually washedout over a period of 10–18 min. As shown in Figure 1, thisrepolarization occurred concomitantly with increasing inputresistance. The mean amplitude of the hyperpolarization was7.0 6 1.5 mV and 6.1 6 1.2 mV in stellate (n ¼ 11) andpyramidal cells (n ¼ 10), respectively (not significantly different:
FIGURE 1. In current clamp mode, 5-HT induced a stronghyperpolarization followed by a smaller depolarization in bothsubtypes of medial EC (mEC) Layer II projection neurons. Bathapplication of 100 lM 5-HT (indicated by the thick horizontalbar) to a Layer II stellate neuron led to a quickly developingand long-lasting hyperpolarization. Following washout of the5-HT-induced hyperpolarization, a small depolarization could beobserved. The hyperpolarization was accompanied by a reductionin apparent input resistance as revealed by a reduction in the volt-age deflections to hyperpolarizing constant current pulses appliedthroughout the experiment (Expansions 1 and 2).
116 MA ET AL.
Hippocampus DOI 10.1002/hipo
P > 0.05). The change in input resistance, expressed by mea-suring a ratio of stable response amplitudes to injection ofcurrent pulses before and after application of 5-HT, wasreduced on average to (64.6 6 10.6)% and (62.6 6 11.9)%of control values in stellate (n ¼ 11) and pyramidal cells (n ¼ 10),respectively (not significantly different: P > 0.05). Repeatedapplications of 5-HT (2 or even 3 times) following recovery inthe same neuron consistently evoked a similar hyperpolarizingresponse (n ¼ 4, data not shown).
In addition to the hyperpolarizing responses directly follow-ing the application of 5-HT, a small depolarization (on average2.9 6 1.1 mV and 2.7 6 1.2 mV in stellates and pyramids,respectively) could also be observed (14 out of 18 cases),following washout of the 5-HT-induced hyperpolarization(Fig. 1). Although in all cells, the repolarization–depolarizationsequence following the initial hyperpolarization was correlatedwith an increase in apparent input resistance. When compared
with the control levels (pre-5-HT), the values of input resist-ance during the depolarizing response could show either anincrease, decrease, or no change. Furthermore, although thehyperpolarizing response was robust to repeated applications of5-HT, the depolarizing response was less so, becoming smallerin subsequent applications (n ¼ 4; data not shown).
Voltage Clamp Recordings of 5-HT-MediatedEffects
Consistent with current clamp data, bath application of100 lM 5-HT for a duration of 30 s in voltage clamp modeevoked a strong and long-lasting outward shift of the holdingcurrent when neurons were held at a potential approximatelyequivalent to resting membrane potential levels (�60 mV)(Fig. 2A). This response was maximal at a latency of 2–3(2.6 6 0.3) min and gradually washed out over a period of
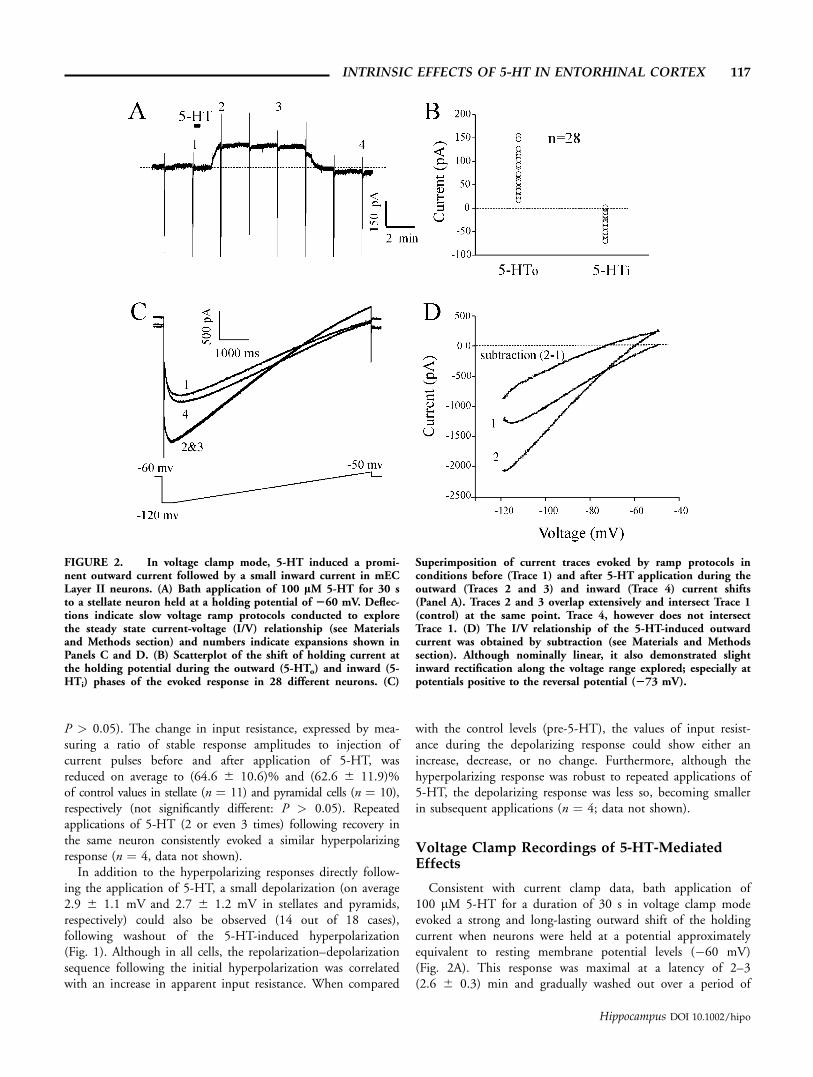
FIGURE 2. In voltage clamp mode, 5-HT induced a promi-nent outward current followed by a small inward current in mECLayer II neurons. (A) Bath application of 100 lM 5-HT for 30 sto a stellate neuron held at a holding potential of 260 mV. Deflec-tions indicate slow voltage ramp protocols conducted to explorethe steady state current-voltage (I/V) relationship (see Materialsand Methods section) and numbers indicate expansions shown inPanels C and D. (B) Scatterplot of the shift of holding current atthe holding potential during the outward (5-HTo) and inward (5-HTi) phases of the evoked response in 28 different neurons. (C)
Superimposition of current traces evoked by ramp protocols inconditions before (Trace 1) and after 5-HT application during theoutward (Traces 2 and 3) and inward (Trace 4) current shifts(Panel A). Traces 2 and 3 overlap extensively and intersect Trace 1(control) at the same point. Trace 4, however does not intersectTrace 1. (D) The I/V relationship of the 5-HT-induced outwardcurrent was obtained by subtraction (see Materials and Methodssection). Although nominally linear, it also demonstrated slightinward rectification along the voltage range explored; especially atpotentials positive to the reversal potential (273 mV).
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 117
Hippocampus DOI 10.1002/hipo

7–12 (9.2 6 1.5) min. Following washout, holding currentlevels were occasionally shifted inwards when compared withthe initial values obtained preapplication (Fig. 2A). Althoughthe initial outward shift evoked by 5-HT was both robust andconsistent (occurring in all 41 cells tested in these conditions),the subsequent inward shift present following washout of theoutward component was less so. With bath applications of100 lM 5-HT for 30 s, only 71% (20 out of 28) of neuronsrecorded from for a sufficient duration following applicationof 5-HT displayed a visible inward component (>5 pA) uponwashout of 5-HT at a holding potential of �60 mV. Thesedata are summarized in scatterplot form in Figure 2B.
In an initial attempt to elucidate the ionic basis of both theoutward and inward current shifts evoked by 5-HT (5-HToand 5-HTi, respectively), we performed comparisons of thesteady state I/V relationships of neurons prior and following5-HT application with the use of slow voltage ramps (denotedby deflections in Fig. 2A with respective expansions shownin Fig. 2C). As illustrated in Figure 2C, by superimposingramp-evoked currents at these different points, we were able tocompare the steady state I/V relationships during these 5-HT-evoked changes. In all neurons recorded, current traces obtainedduring the outward shift of holding current displayed an inter-section point with the control (pre-5-HT) trace within therange of ramp voltages used (�120 to �50 mV). The I/Vrelationship of the 5-HT-induced outward current was obtainedby subtraction and a representative plot is shown in Figure 2D.On average, this current reversed at �71.0 6 4.8 mV across allcells recorded and tended to show a slight inward rectificationalong the voltage range explored. Importantly, the I/V relation-ship calculated for the 5-HT-induced outward current did notchange appreciably during the early period of the outwardcurrent shift. This is shown in Figure 2C as an extensive over-lap of Traces 2 and 3, which were obtained immediately and�5 min following the development of the 5-HT induced out-ward shift. This finding suggests that there is little temporaloverlap between the development of the early components ofthe outward and inward components of the 5-HT-inducedresponse.
The I/V relationship of the 5-HT evoked inward shift wascalculated using a similar protocol (i.e., subtraction of currenttraces evoked by voltage ramps before and during the 5-HTevoked inward component). A ramp-evoked current trace dur-ing the 5-HT-induced inward shift is shown in Figure 2C(Trace 4). In this example, there was no intersection with thecontrol (pre 5-HT) trace. From all experiments, in which aninward shift occurred, there were three general and differentpatterns for the I/V relationship (represented in Fig. 3). In thefirst case illustrated (Fig. 3A: data derived from the cell shownin Fig. 2), the inward current shift appeared across the entirerange of voltages tested. In these cases (8 out of 20 total), therewas no intersection of the current trace evoked in 5-HT andthe control trace and therefore no reversal potential foundwithin the range of voltages tested. This suggests that the majorcomponent of the inward response was produced throughthe activation of a current with a reversal potential positive to
�50 mV. In the second case illustrated (Fig. 3B), the I/V rela-tionship for the inward component appeared as an inversion ofthe outward component, reversing at a similar potential butdemonstrating the opposite polarity. This pattern was observedin 8 of the 20 cases and suggested a similar ionic mechanismfor both the inward and outward shifts, albeit having oppositepolarities. This implies in one case, a channel opening, and inthe other, a channel closing. In the third case illustrated, whichoccurred in the final 4 of the 20 cases, a peculiar I/V relation-ship was evident in which the reversal potential was evident atmore hyperpolarized potentials (between �80 and �120 mV)suggesting a mixed inward/outward ionic mechanism. The re-versal potentials for both the outward and inward componentsof the 5-HT evoked responses across all neurons are shown inFigure 3D. Although the reversal potentials for the outwardcomponent were highly consistent, there were three clusters ofreversal potential values for the inward component. Based onthe variability of the I/V relationships and estimated reversalpotentials, there appeared to be at least two independent ionicmechanisms for the inward shift and these two mechanismscould cooccur and have overlapping influences.
Although no significant differences in the 5-HT-evokedresponses between the two subtypes of EC Layer II neurons(stellate and pyramidal) were observed in current clamp record-ings, we could evaluate this more precisely under voltage clampconditions. As shown in Table 1, no differences were observedin either the amplitude or the reversal potential of the outwardcurrent evoked by 5-HT across the two cell types. Similarly,no differences were observed in the amplitude of the inwardcurrent evoked by 5-HT. Because of this homogeneity ofresponses, we elected to pool subsequent results of the effectsof 5-HT across both subtypes of cells.
Pharmacology of 5-HT-Induced Currents
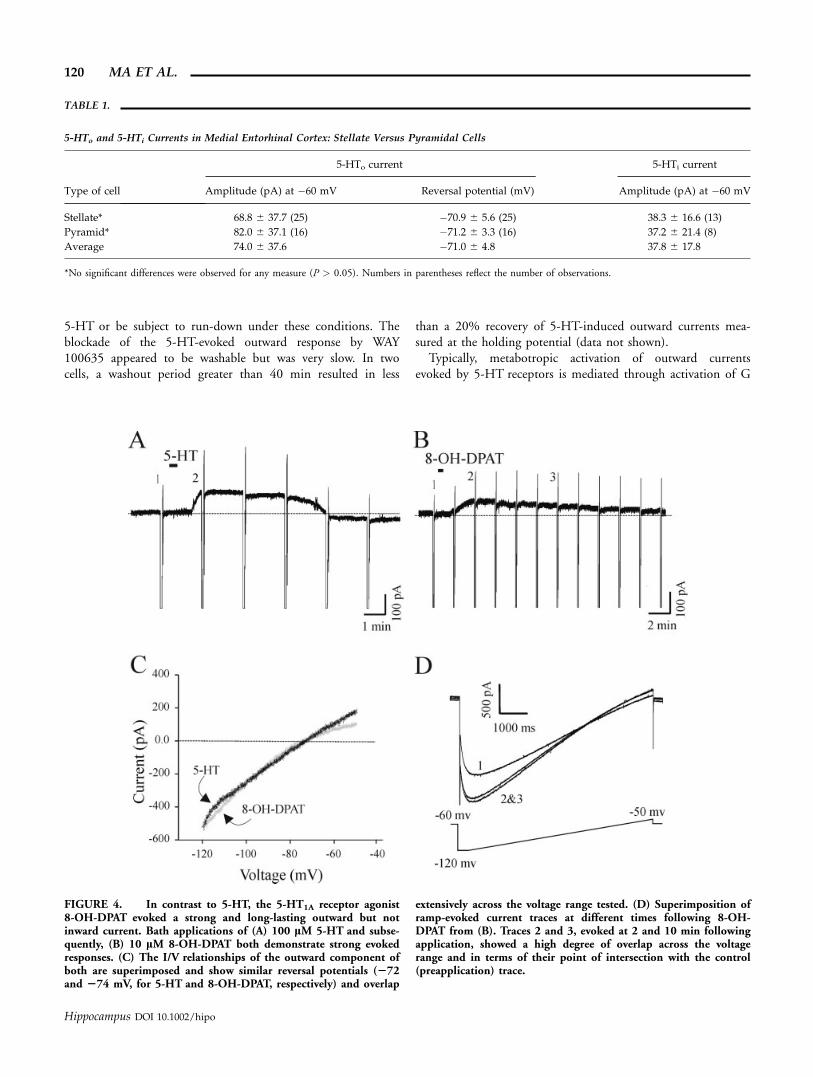
To study the possible receptor subtypes involved in themediation of the 5-HT-induced responses, we employed thetwo drugs 8-OH-DPAT (a selective 5-HT1A agonist; (Gozlanet al., 1983) and WAY 100635 (a selective 5-HT1A antagonist;(Craven et al., 1994; Corradetti et al., 1996; Fletcher et al.,1996)). Consistent with results using 5-HT, bath application of10 lM 8-OH-DPAT also evoked an outward current at a shortlatency, however, the duration of this outward shift was muchlonger, lasting for the length of the recording period (maxi-mum 25–30 min), when compared with an average of 9.5 61.4 min for 100 lM 5-HT previously applied to the same neu-ron (Fig. 4). Exploration of the I/V relationship of this current(Fig. 4C) revealed a high degree of similarity to that evoked by5-HT. The average reversal potential (�74.2 6 2.9 mV) wasnot significantly different to that observed with 5-HT (P > 0.05,n ¼ 6) although the degree of inward rectification appearedmore pronounced (Fig. 4C). The mean amplitude of the out-ward current evoked by 8-OH-DPAT at a holding potential of�60 mV was 53.7 6 22.6 pA.
Another striking difference between the effects of 5-HT and8-OH-DPAT was that inward shifts were never observed using
118 MA ET AL.
Hippocampus DOI 10.1002/hipo

the 5-HT1A receptor agonist. There are several possibilities forthis effect: (1) a prior application of 5-HT might have desensi-tized or occluded this response, or during the intervening timebetween applications, the intracellular mechanisms for its elici-tation might have rundown, (2) any inward current might havebeen masked by the strong and long-duration activation of the
outward current, or (3) that another receptor subtype could beinvolved in the activation of the inward current. We felt thatthe latter possibility was more probable since: (1) a similar lackof inward current was noted with applications of 8-OH-DPATalone (not shown: n ¼ 5) and (2) the I/V relationship and thereversal potential (intersection point) of the outward currentevoked by 8-OH-DPAT was consistent across a time span dur-ing, which the inward current was normally evoked with 5-HT(see overlapping plots in Fig. 4D).
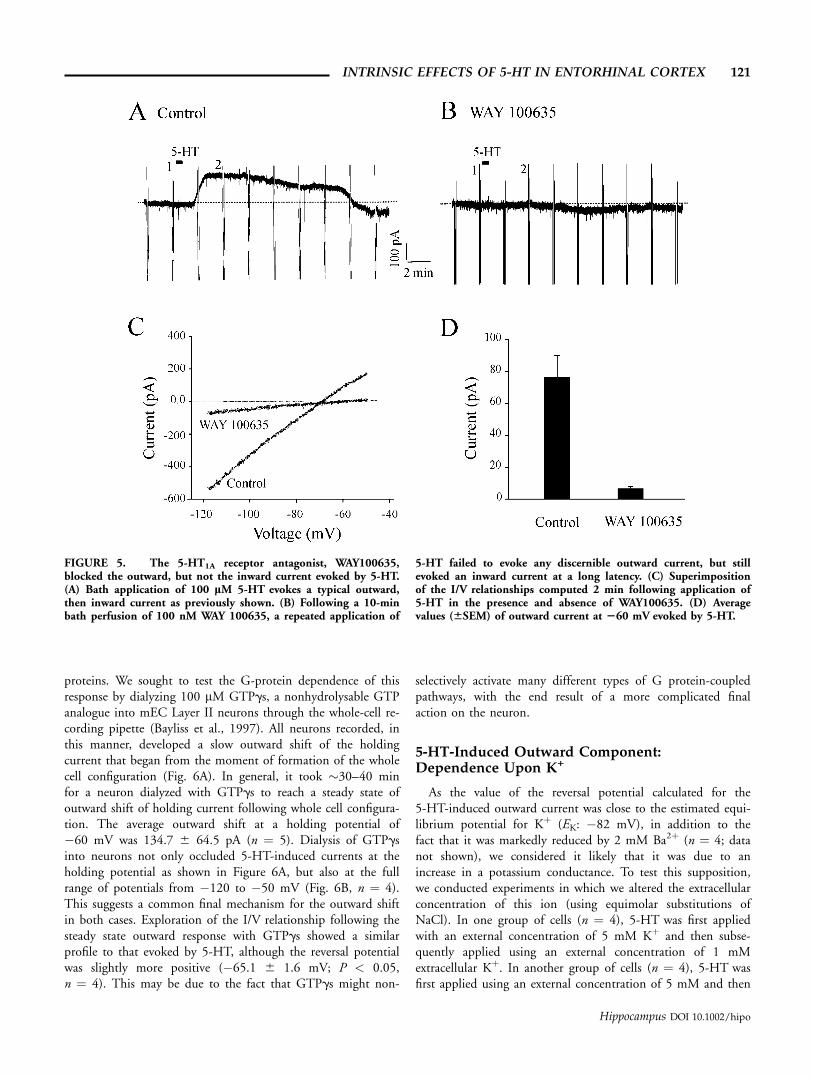
Consequently, we tested the 5-HT-induced response in thepresence of the selective 5-HT1A receptor antagonist, WAY100635. Following an initial application of 5-HT in control so-lution, it was reapplied in the presence of 100 nM WAY100635, which was perfused for at least 10 min before the sec-ond application of 5-HT. In all cases, the 5-HT-induced out-ward current was dramatically blocked in the presence of WAY1000635 when assessed at a holding potential of �60 mV(cf. Panels A and B) as well as across the voltage range exploredwith ramp protocols (Panel C) (Fig. 5). On average, the out-ward current evoked by 5-HT in the presence of 100 nM WAY100635 was 6.6 6 4.3 pA at a holding potential of �60 mV,which is significantly smaller than that observed in the absenceof the antagonist (Fig. 5D: P < 0.05, n ¼ 6). These resultssupport the idea that the outward current is selectively inducedby stimulation of 5-HT1A receptors. Interestingly, in 3 out of 6neurons tested in this fashion, there was a development of anoticeable inward current at �60 mV that was maximal at alatency of 6 to 8 (7.3 6 0.7) min following the drug applica-tion. As shown in Figure 5B, this current developed at latenciesthat were longer than those for the outward current but shorterthan those for the inward current evoked by 5-HT in controlconditions. The peak amplitude of this inward current was alsoless than that evoked during control conditions, suggesting thatit could show desensitization for subsequent applications of
FIGURE 3. I/V relationships of 5-HT-induced outward currentwere consistent, while 5-HT-induced inward current were inconsis-tent, across the neurons studied. The steady-state I/V relationshipof the 5-HT evoked outward (5-HTo) and inward (5-HTi) currentswere computed as described and shown in Figure 2. In all casesillustrated (A–C), the I/V relationships for the outward component(5-HTo) are highly comparable, showing similar shapes and rever-sal potentials (273, 272, and 269 mV, respectively). However,the I/V relationships for the inward component (5-HTi) are all dis-similar. (A) In this example (same data as from Fig. 2), the I/Vrelationship of 5-HTi is negative throughout the range of voltagestested and demonstrates no reversal potential. By linear extrapola-tion, a reversal potential could be estimated at 235 mV. (B) Inthis example, the I/V relationship of 5-HTi is inverted with respectto that of 5-HTo but interestingly, they both demonstrate a rever-sal potential at a similar voltage (271 mV). (C) In this example,the I/V relationship of 5-HTi appears to demonstrate a mix of therelationships previously shown in (A) and (B). Although the cur-rent reverses within the voltage range tested, it is shifted to a morehyperpolarized potential (2105 mV). (D) A scatterplot of the val-ues of reversal potentials observed across all cells shows the consis-tency of the outward (5-HTo) vs. the inward (5-HTi) componentof the 5-HT-evoked response.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 119
Hippocampus DOI 10.1002/hipo
5-HT or be subject to run-down under these conditions. Theblockade of the 5-HT-evoked outward response by WAY100635 appeared to be washable but was very slow. In twocells, a washout period greater than 40 min resulted in less
than a 20% recovery of 5-HT-induced outward currents mea-sured at the holding potential (data not shown).
Typically, metabotropic activation of outward currentsevoked by 5-HT receptors is mediated through activation of G
FIGURE 4. In contrast to 5-HT, the 5-HT1A receptor agonist8-OH-DPAT evoked a strong and long-lasting outward but notinward current. Bath applications of (A) 100 lM 5-HT and subse-quently, (B) 10 lM 8-OH-DPAT both demonstrate strong evokedresponses. (C) The I/V relationships of the outward component ofboth are superimposed and show similar reversal potentials (272and 274 mV, for 5-HT and 8-OH-DPAT, respectively) and overlap
extensively across the voltage range tested. (D) Superimposition oframp-evoked current traces at different times following 8-OH-DPAT from (B). Traces 2 and 3, evoked at 2 and 10 min followingapplication, showed a high degree of overlap across the voltagerange and in terms of their point of intersection with the control(preapplication) trace.
TABLE 1.
5-HTo and 5-HTi Currents in Medial Entorhinal Cortex: Stellate Versus Pyramidal Cells
Type of cell
5-HTo current 5-HTi current
Amplitude (pA) at �60 mV Reversal potential (mV) Amplitude (pA) at �60 mV
Stellate* 68.8 6 37.7 (25) �70.9 6 5.6 (25) 38.3 6 16.6 (13)
Pyramid* 82.0 6 37.1 (16) �71.2 6 3.3 (16) 37.2 6 21.4 (8)
Average 74.0 6 37.6 �71.0 6 4.8 37.8 6 17.8
*No significant differences were observed for any measure (P > 0.05). Numbers in parentheses reflect the number of observations.
120 MA ET AL.
Hippocampus DOI 10.1002/hipo
proteins. We sought to test the G-protein dependence of thisresponse by dialyzing 100 lM GTPgs, a nonhydrolysable GTPanalogue into mEC Layer II neurons through the whole-cell re-cording pipette (Bayliss et al., 1997). All neurons recorded, inthis manner, developed a slow outward shift of the holdingcurrent that began from the moment of formation of the wholecell configuration (Fig. 6A). In general, it took �30–40 minfor a neuron dialyzed with GTPgs to reach a steady state ofoutward shift of holding current following whole cell configura-tion. The average outward shift at a holding potential of�60 mV was 134.7 6 64.5 pA (n ¼ 5). Dialysis of GTPgsinto neurons not only occluded 5-HT-induced currents at theholding potential as shown in Figure 6A, but also at the fullrange of potentials from �120 to �50 mV (Fig. 6B, n ¼ 4).This suggests a common final mechanism for the outward shiftin both cases. Exploration of the I/V relationship following thesteady state outward response with GTPgs showed a similarprofile to that evoked by 5-HT, although the reversal potentialwas slightly more positive (�65.1 6 1.6 mV; P < 0.05,n ¼ 4). This may be due to the fact that GTPgs might non-
selectively activate many different types of G protein-coupledpathways, with the end result of a more complicated finalaction on the neuron.
5-HT-Induced Outward Component:Dependence Upon K+
As the value of the reversal potential calculated for the5-HT-induced outward current was close to the estimated equi-librium potential for Kþ (EK: �82 mV), in addition to thefact that it was markedly reduced by 2 mM Ba2þ (n ¼ 4; datanot shown), we considered it likely that it was due to anincrease in a potassium conductance. To test this supposition,we conducted experiments in which we altered the extracellularconcentration of this ion (using equimolar substitutions ofNaCl). In one group of cells (n ¼ 4), 5-HT was first appliedwith an external concentration of 5 mM Kþ and then subse-quently applied using an external concentration of 1 mMextracellular Kþ. In another group of cells (n ¼ 4), 5-HT wasfirst applied using an external concentration of 5 mM and then
FIGURE 5. The 5-HT1A receptor antagonist, WAY100635,blocked the outward, but not the inward current evoked by 5-HT.(A) Bath application of 100 lM 5-HT evokes a typical outward,then inward current as previously shown. (B) Following a 10-minbath perfusion of 100 nM WAY 100635, a repeated application of
5-HT failed to evoke any discernible outward current, but stillevoked an inward current at a long latency. (C) Superimpositionof the I/V relationships computed 2 min following application of5-HT in the presence and absence of WAY100635. (D) Averagevalues (66SEM) of outward current at 260 mV evoked by 5-HT.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 121
Hippocampus DOI 10.1002/hipo

subsequently, 10 mM extracellular Kþ. As shown in Figure 7A,the reversal potential of the 5-HT-induced outward current was�69.5 mV with 5 mM extracellular Kþ and was shifted to�102.5 mV when the external concentration was changed to1 mM Kþ. Furthermore, as shown in Figure 7B, the reversalpotential of this current was shifted from �70.5 mV in 5 mMextracellular Kþ to �49.2 mV in 10 mM extracellular Kþ.A plot of the average reversal potential as a function of the log-arithmic Kþ concentration in the extracellular fluid showed amarked linearity, suggesting that the 5-HT-induced outwardcurrent is predominantly Kþ dependent (Fig. 7C).
5-HT-Induced Inward Current:Partial Dependence Upon Ih
Although the inward component evoked by 5-HT appearedto be a possible mix of different current subtypes, at least one
subtype appeared to be inward across the entire potential rangeof our voltage ramp protocols (Fig. 3A). A candidate currentfor this putatively independent effect is the hyperpolarization
FIGURE 6. 5-HT-induced outward currents were likely medi-ated via G-protein activation. (A) Whole cell dialysis with 100 lMGTPgs caused a slowly developing but prominent outward shift ofholding current in the absence of 5-HT. At a stable level, applica-tion of 5-HT (horizontal bar) failed to induce any further outwardshift. The initial artifact in the recording trace indicated by theopen arrow refers to the initiation of whole cell mode (break ofmembrane patch). (B) I/V relationship of current induced byGTPgs (difference of Traces 2 and 1) and 5-HT (difference ofTraces 3 and 2). 5-HT failed to evoke any current above andbeyond that induced by GTPgs.
FIGURE 7. The I/V relationship and reversal potential of5-HT-induced outward current was shifted with changes in theexternal potassium concentration ([K+]o). (A) Substitution of1 mM [K+]o for 5 mM [K+]o shifted the I/V plot and the reversalpotential of the 5-HT-induced outward current to a more negativepotential while in another neuron, (B) substitution of 10 mM [K]ofor 5 mM [K]o shifted the I/V plot and the reversal potential to aless negative (more positive) potential. The reversal potential ofthe current at 10 mM [K]o was estimated by linear extrapolation(dotted line). (C) Plot of the averaged reversal potential (66SEM)of the 5-HT evoked outward current for various manipulations of[K+]o (log scale) and the least squares regression fit to the data(solid line).
122 MA ET AL.
Hippocampus DOI 10.1002/hipo

activated delayed inward rectifier (Ih), which is also a substan-tive component of the total membrane current of mEC LayerII cells, especially stellates (Alonso and Llinas, 1989; Alonsoand Klink, 1993; Dickson et al., 2000b). To test this hypoth-esis, we utilized the bradycardic agent ZD7288, reported tobe a selective and specific blocker of Ih (BoSmith et al.,1993; Briggs et al., 1994; Harris and Constanti, 1995; Mac-caferri and McBain, 1996; Gasparini and DiFrancsco, 1997;Dickson et al., 2000b). A measure of 10-lM ZD7288 wasfound to block Ih in EC Layer II neurons adequately,although slowly, usually requiring �20–25 min to reach max-imal blockade in our experimental conditions (Fig. 8A). Onaverage, 10 lM ZD7288 reduced Ih measured at a membrane
potential of �110 mV to 11.8% 6 3.1% of control (n ¼ 8).Because of this slow action, we typically began to apply thedrug 5–10 min before breaking of membrane patch in theneurons.
A typical 5-HT-induced response in the presence of 10 lMZD7288 is shown in Figure 8B. As previously described, appli-cation of 5-HT still led to a rapidly developing and long-lastingoutward shift of the holding current, however, the inward shiftupon washout of the 5-HT-induced outward current appearedto be reduced to a considerable extent. The steady-state I/V rela-tionship for both the outward and inward currents evoked by5-HT in ZD7288 perfusion conditions are shown in Figure 8C.The outward current evoked by 5-HT appeared similar to that
FIGURE 8. 5-HT-induced currents in the presence of the Ihblocker, ZD7288. (A) Hyperpolarization activated inward currents(Ih) were recorded in control conditions and subsequently after24 min of perfusion with 10 lM ZD7288 by applying long dura-tion (>>1 s) hyperpolarizing voltage steps from a holding potentialof 260 to 2110 mV. The amplitude of Ih was measured by sub-tracting the instantaneous current (*) from steady state current(**). (B) In the presence of 10 lM ZD7288, bath application of100 lM 5-HT led to an outward current very similar to thatshown previously (Fig. 2). However, in this example, very littleinward current was observed upon washout of 5-HT. (C) I/V rela-
tionship of 5-HT-induced outward and inward currents wereobtained in the same way as in Figures 2 and 3. The reversalpotential for both was very similar (272 and 269 mV, for 5-HToand 5-HTi, respectively) (D) Mean amplitude (66SEM) of 5-HT-induced outward (5-HTo) and inward (5HTi) currents in controland ZD7288 experiments (left panel), and a scatter plot of thereversal potentials of both the 5-HTo and 5-HTi currents (rightpanel). Although the inward component was markedly reduced, itsreversal potential was highly consistent and similar to that foundin the outward component under these conditions.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 123
Hippocampus DOI 10.1002/hipo

observed previously in control conditions (cf. Figs. 2 and 3),however, the amplitude of the current measured at �60 mV(Fig. 8D, left panel) was found to be significantly low (76% ofcontrol values; P < 0.05, n ¼ 17) (Fig. 8D, left panel). Theaverage reversal potential for the outward current in theseconditions (�70.2 6 1.9 mV) was not found to be signifi-cantly different from that found in normal perfusion medium(P > 0.05, n ¼ 17) (Fig. 8D, right panel). This suggested thatZD7288 might have a nonspecific blocking effect upon theoutward current evoked by 5-HT. Confirmation of this block-ing effect of ZD7288 on the outward current was realized inexperiments in which we perfused higher concentrations ofZD7288 (100 lM). In these cases, very little outward currentcould be evoked by 5-HT (n ¼ 6; not shown).
The inward component evoked by 5-HT was also reduced inthe presence of 10 lM ZD7288. The mean amplitude of theinward current measured at �60 mV was decreased signifi-cantly from control values (61% of control values; P < 0.05,n ¼ 8, Fig. 8D, left panel). Perhaps more interestingly, andunlike control conditions, the I/V relationship of any inwardshift under these conditions was highly consistent across allneurons. As shown in Figure 8C, this current showed a similarI/V relationship to that presented previously in Figure 3B,which reversed at a similar potential to that of the outwardcomponent evoked by 5-HT. This suggests that while modula-tion of Ih may contribute significantly to the inward compo-nent evoked by 5-HT, it is certainly not the only componentof this response.
To directly examine a potential modulatory role of 5-HTupon Ih in mEC Layer II neurons, we performed a series ofexperiments in conditions designed to isolate Ih. All recordingswere undertaken in the presence of 2 mM Ba2þ to block boththe instantaneous inward rectifier and to reduce the outwardcurrent evoked by 5-HT. Furthermore, they were also con-ducted in the presence of the 5-HT1A receptor blocker WAY100635 to isolate the inward current evoked by 5-HT. In placeof voltage ramps, we performed a tail current analysis at apotential of �80 mV following long duration (>1 s) voltage-clamp steps to potentials ranging from �30 to �130 mV inorder to study the activation of Ih (see Materials and Methodssection and Fig. 9). This protocol was conducted, before, dur-ing, and following washout of the inward current evoked by5-HT. We also used a faster and longer perfusion protocol for5-HT in order to evoke a larger amplitude and longer lastinginward shift of the holding current.
Prior to the application of 5-HT, the activation range of Ihwas observed between �35 and �120 mV with a mean valuefor V1/2 of �79.3 6 0.9 mV and a slope factor (k) of 11.6 60.2 as illustrated in Figure 9D. These values agreed with thosepreviously observed for cells of this region (Dickson et al.,2000b). Following the application of 5-HT, a prominent andlong-lasting inward shift of the holding current recorded at�60 mV was developed. This current had an average peakamplitude of 32.2 6 6.6 pA (n ¼ 5), which maximized at alatency of 4–5 min and gradually washed out over a period of12–15 min. During the peak of inward current, the activation
curve of Ih was shifted in the depolarizing direction by 3.1 60.5 mV (n ¼ 5; P < 0.05).
DISCUSSION
In other brain regions (including the hippocampus), theintrinsic electrophysiological actions of 5-HT on neurons aremultiple and complex, which is likely due to the existence of aplethora of different 5-HT receptor subtypes (Larkman andKelly, 1995; Andrade, 1998). Our present results in the medialEC are consistent with these composite intrinsic actions.Although initial studies in the EC (Sizer et al., 1992;Grunschlag et al., 1997) showed prominent 5-HT1A-mediatedhyperpolarizing responses, the bulk of recent studies in the EChave primarily focused on the modulatory action of 5-HT onexcitatory synaptic transmission since intrinsic responsivenesswas both inconsistent and small (Schmitz et al., 1998c). Ourresults suggest, in contrast to these latter researchers, that theintrinsic hyperpolarizing action of 5-HT upon medial EC LayerII principal neurons is both consistent and prominent. Theintrinsic effects of 5-HT in the EC are likely to be relevant forneural processing in the medial temporal lobe due to the nodalanatomical position occupied by these cells and the fact thatendogenous serotonin release is thought to modulate the activ-ity of this region in a state-dependent fashion (Vanderwolf,1988; Dringenberg and Vanderwolf, 1998).
5-HT-Induces Biphasic and Opposing Responsesin mEC Projection Neurons
In our current clamp experiments, 5-HT produced an initialrobust hyperpolarization followed by a weaker but long-lastingdepolarization. Consistent with these results, our voltage clampexperiments revealed that at a holding potential level equivalentto rest, 5-HT evoked an initial robust outward shift of holdingcurrent levels followed by a weaker and less consistent inwardshift. Although the outward and inward current shifts could beexpressed in the same neurons, the characteristics of the tworesponses appeared very different. While the outward currentshifts were robust, reliable, and repeatable, the same was notnecessarily true of the inward current shifts. It is of interest tonote that despite this being the first report of 5-HT-induceddepolarization in mEC neurons, inspection of traces in Figure 1Aof Grunschlag et al. (1997) and Figure 2 of Sizer et al. (1992),show clear, albeit small, depolarizations evoked by 5-HTfollowing the typical (and large) hyperpolarizing response. Byusing low-access resistance whole-cell recording techniques,we may have been better able to quantify this depolarizingresponse when it occurred.
The implication from these initial data (and supported byour later experiments) was the two opposing responses thatcould involve differences both in their ionic substrates and thereceptor/intracellular activation pathways responsible for each.Dual opposing actions of 5-HT on individual cells is consistentwith previous studies in other neurons, such as hippocampal
124 MA ET AL.
Hippocampus DOI 10.1002/hipo

CA1 pyramidal cells (Andrade and Nicoll, 1987; Colino andHaliwell, 1987), neurons of dorsal raphe nucleus (Cravenet al., 2001), and cortical neurons (Davies et al., 1987).
5-HT-Induced Outward Current
Ionic mechanism
The I/V relationship of the 5-HT evoked outward shift asexplored in voltage clamp experiments was highly consistentacross all neurons tested. It reversed at �71 mV, and showedmoderate rectification across the voltage range between �120and �50 mV. It was markedly reduced by the addition ofBa2þ to the extracellular fluid and was affected by alterationsof extracellular Kþ. Based upon these data, it appears highly
likely that this outward current evoked by 5-HT represents theopening of a potassium channel.
Although the value of the reversal potential of this currentwas not equivalent to the value of the equilibrium potential forKþ in our experiments as calculated via the Nernst equation(�82 mV for 5 mM external Kþ), when accounting for thevalue of the junction potential between the pipette and extrac-ellular solution in our experiments (8 mV), it was highlysimilar (�79 mV). Furthermore, and perhaps more convinc-ingly, alterations of the extracellular concentration of Kþ sys-tematically shifted the values of the reversal potential of thiscurrent in a linear relationship to the log of the Kþ concentra-tion. This linear shift paralleled the predicted shift of the Kþ
equilibrium potential as well.
FIGURE 9. 5-HT modulates Ih in medial EC Layer II projec-tion neurons via non-5-HT1A receptor stimulation. (A) In thepresence of 150 nM WAY100635 and 2 mM Ba2+, application of100 lM 5-HT under fast perfusion conditions for 3 min elicited aprominent inward shift of the holding current at 260 mV. Deflec-tions show a family of applied voltage pulses (expanded and super-imposed in Panel B) to examine the I/V relationship and the acti-vation profile of Ih before and after 5-HT application. (B) Currenttraces evoked by voltage step protocols (i) before, (ii) during, and
(iii) after washout of 5-HT-induced inward current shift. Net steadystate currents were larger during the inward shift. (C) Activationcurves for Ih were established by plotting normalized maximal tailcurrents (see Materials and Methods section). 5-HT shifted theactivation curve to the right in this example (left panel) andon average in 5 neurons (right panel). The half-activation voltage(V1/2) for the neuron shown in Figure 9 was 282.2 and 278.4 mVin control and postapplication of 5-HT, respectively, which yieldeda 3.8 mV positive shift.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 125
Hippocampus DOI 10.1002/hipo

Pharmacology
In other central nervous system neurons, the mechanism of5-HT-evoked outward currents has been consistently shown tobe due to the opening of Kþ channels via activation of a5-HT1A receptor-coupled G protein, (Andrade and Nicoll,1987; Penington et al., 1993; Hwang and Dun, 1998). Inagreement with these results, our pharmacological studies alsoconfirmed that the 5-HT-evoked outward current in mECLayer II neurons required stimulation of the 5-HT1A receptorand involvement of G-protein activation. First, the selective5-HT1A-receptor agonist, 8-OH-DPAT, mimicked the outwardshift evoked by 5-HT. In fact, the 8-OH-DPAT-evoked effectlasted remarkably longer than 5-HT in the neurons, perhapsdue to a higher binding affinity of 8-OH-DPAT for 5-HT1A
receptors than 5-HT (Gozlan et al., 1983). Long-lasting actionsof 8-OH-DPAT have been previously reported in other centralnervous system neurons (Andrade and Nicoll, 1987; Leishmanet al., 1994; Hwang and Dun, 1998). Second, the selective5-HT1A-receptor antagonist WAY 100635 abolished the 5-HT-evoked outward current (Fig. 5). Third, intracellular dialysis ofGTPgs, a nonhydrolysable GTP analogue, induced an outwardshift of the holding current with similar properties to the out-ward current evoked by 5-HT and, perhaps more importantly,occluded any further responsiveness of the neuron to 5-HT.
The pronounced hyperpolarization and outward currentevoked in mEC neurons with application of 5-HTmay be asso-ciated with the high density of 5-HT1A receptors expressed inthis part of the brain. A variety of approaches, including auto-radiography, in situ hybridization, and immunohistochemistryhave demonstrated that the EC, like both the hippocampalformation and the raphe nuclei, expresses very high densities of5-HT1A receptors (Pazos and Palacios, 1985; Chalmers andWatson, 1991; Wright et al., 1995; Zhou et al., 1999). Stimu-lation of 5-HT1A receptors in EC has also been shown to resultin high G-protein activity (Sim et al., 1997). Furthermore, theintracellular signaling mechanisms of 5-HT-induced hyperpo-larization in central nervous system neurons has been elucidated(Oh et al., 1995) [for a review, see Yamada et al. (1998)]. Inbrief, activation of a 5-HT1A-coupled G-protein results in thedissociation of two G subunits; subunit a (Ga) and subunitbg (Gbg). This, in turn, results in the binding of Gbg toG-protein-regulated Kþ channels (GIRKs), which leads tochannel opening. Interestingly, GIRK channels are also highlyexpressed in EC (Karschin et al., 1996). In summary, all of theabove-mentioned evidence support that 5-HT action in ECneurons should exert strong hyperpolarization via activation ofoutward Kþ currents. Although, other authors have describedsuch consistent intrinsic robust hyperpolarizing responses inEC neurons (Sizer et al., 1992; Grunschlag et al., 1997), resultsfrom another laboratory report inconsistent and rather smallintrinsic hyperpolarizing responses in the EC (Schmitz et al.,1998b). It is unlikely that the concentration of 5-HT could bea likely explanation since our study used concentrations compa-rable to those used by Schmitz and colleagues in which theyreported a lack of intrinsic effects (Schmitz et al., 1995b,
1998b,d). Indeed, the studies of Sizer et al. (1992) and Grun-schlag et al. (1997) obtained prominent intrinsic effects atlower concentrations of 5-HT. One possible limitation of ourstudy was that we did not attempt to differentiate the influenceof a variety of concentrations of 5-HT.
ZD7288 nonspecific block
Another interesting and original feature concerning theoutward shift evoked by 5-HT was that it was partially blockedby low concentrations and almost completely blocked by highconcentrations of ZD7288, a supposedly specific blocker of Ih.Our results suggest that the actions of ZD7288 may not be asspecific as previously touted. Typically, most studies concerningthe specificity of ZD7288 have focused on inwardly rectifyingcurrents in the voltage range hyperpolarized to the restingmembrane potential. In these experiments, ZD7288 clearlyblocks Ih while having little to no effect on the instantaneousinward rectifier (IKir) (BoSmith et al., 1993; Briggs et al., 1994;Harris and Constanti, 1995; Maccaferri and McBain, 1996;Gasparini and DiFrancsco, 1997). However, a recent study hasshown that ZD7288 blocks both native and reconstitutedlow-threshold (T type) Ca2þ currents in sperm cells (Felixet al., 2003). Our data suggest that ZD7288 may also have aneffect upon GIRK-type channels in excitable membranes as well.
5-HT-Induced Inward Currents
The variability in the inward currents evoked by 5-HTsuggested at least two independent and overlapping mecha-nisms. In other central nervous systems cells, 5-HT-evokeddepolarizations have been shown to arise through two separateand independent ionic mechanisms: one involving a decreasedpotassium conductance (Andrade and Nicoll, 1987; Colino andHaliwell, 1987; Larkman and Kelly, 1998) and the otherinvolving an enhancement of Ih (Bobker and Williams, 1989;Pape and McCormick, 1989; Larkman and Kelly, 1992;Gasparini and DiFrancesco, 1999). Our findings are consistentwith both these mechanisms, and indeed, a combination ofthese effects in single mEC Layer II neurons.
Although we did not directly investigate the possibility of apotassium conductance decrease as a component of the inwardcurrent shift, two pieces of evidence support this idea. First, the5-HT-induced inward current in some neurons demonstratedan I/V relationship that shared the same reversal potential asthe 5-HT outward current but appeared as a near inversion.We presumed these to be carried out by the same ion species,(namely Kþ) but to reflect channel closure in place of opening.Second, inward currents could still be observed during coperfu-sions with low concentrations of ZD7288 that blocked Ih andin these cases, similar observations were made regarding theI/V relationship and the reversal potential of the inward shift.Thus, in mEC Layer II cells, it would appear that a proportionof the inward shift evoked by 5-HT could be due to the
126 MA ET AL.
Hippocampus DOI 10.1002/hipo

closure of potassium channels. The type and characteristics ofthese channels remain to be discovered.
We did directly confirm a role for 5-HT modulation of Ih inLayer II mEC neurons. We first considered this as a possibilitywhen, in some cells, the inward current evoked by 5-HT ap-peared to reverse at membrane potentials positive to �50 mV.Consistent with a role of Ih in these inward currents, blockadeof Ih using low concentrations of ZD7288 also reduced theamplitude of the inward shifts evoked by 5-HT, suggesting thatit was at least one component of the inward current. Further-more, by examining the activation curve of Ih prior and duringthe inward current evoked by 5-HT, we were able to directlydemonstrate a rightward shift. Essentially, this shift wouldbe expected to allow a greater proportion of Ih to be activatedwithin a more depolarized range of potentials.
The effects of Ih enhancement would be expected to havemultiple actions upon the responsiveness of Layer II mECneurons. We have previously shown that even slight modula-tion of this current can have profound effects in EC Layer IIstellates due to the high density of this current expressedin these cells (Dickson et al., 2000b). The primary effectwould be an increase in excitability due to its depolarizingeffect on the resting membrane potential. However, given theimportant role that Ih has been shown to play in the oscillatoryproperties of mEC Layer II stellates (Dickson et al., 2000b), inaddition to the resonant properties of other central nervoussystem cells (Hutcheon et al., 1996; Hu et al., 2002), another,and perhaps more interesting effect would be an alteration ofthese dynamic properties. This, in turn, could influence therhythmic patterned behavior of the cell, its response to inputsand oscillatory population dynamics in the superficial Layer IImEC network.
The pharmacology of the 5-HT receptor subtype that medi-ated the inward current in the EC remains unknown. However,it is clear from our data that it was independent of stimulationof the 5-HT1A receptor due to the fact that it was inducedin the presence of WAY 100635 and not by 8-OH DPAT. Thisis consistent with studies in other neurons (Andrade, 1998;Cardenas et al., 1999; Chapin and Andrade, 2001).
CONCLUSION
The coexistence of opposing hyperpolarizing and depolariz-ing actions of 5-HT upon the intrinsic membrane properties ofsingle mEC Layer II neurons could confer a multitude of mod-ulatory effects that, in vivo, could yield complex and time-dependent actions not only on the input–output properties ofthese nodally-located cells but could also bestow differentdynamic interactive states of activity patterns within the localnetwork. In concert with the reported actions of 5-HT uponsynaptic transmission within this region, modulation of theintrinsic properties of Layer II EC cells could play an impor-tant role in shaping \information flow" from the neocortex tothe hippocampal formation.
Acknowledgments
A.A. was an MNI Killam Scholar and C.T.D. is an AlbertaHeritage Medical Scholar.
REFERENCES
Alonso A, Klink R. 1993. Differential electroresponsiveness of stellateand pyramidal-like cells of medial entorhinal cortex layer II. J Neu-rophysiol 70:128–143.
Alonso A, Llinas RR. 1989. Subthreshold Naþ-dependent theta-likerhythmicity in stellate cells of entorhinal cortex layer II. Nature342:175–177.
Andrade R. 1998. Regulation of membrane excitability in the centralnervous system by serotonin receptor subtypes. Ann N Y Acad Sci861:190–203.
Andrade R, Nicoll RA. 1987. Pharmacologically distinct actions ofserotonin on single pyramidal neurones of the rat hippocampusrecorded in vitro. J Physiol 394:99–124.
Barry PH, Lynch JW. 1991. Liquid junction potentials and small celleffects in patch-clamp analysis. J Membr Biol 121:101–117.
Bayliss DA, Li YW, Talley EM. 1997. Effects of serotonin on caudalraphe neurons: Activation of an inwardly rectifying potassium con-ductance. J Neurophysiol 77:1349–1361.
Bland BH, Colom LV. 1993. Extrinsic and intrinsic properties under-lying oscillation and synchrony in limbic cortex. Prog Neurobiol41:157–208.
Bobker DH, Williams JT. 1989. Serotonin augments the cationic cur-rent Ih in central neurons. Neuron 2:1535–1540.
BoSmith RE, Briggs I, Sturgess NC. 1993. Inhibitory actions ofZENECA ZD7288 on whole-cell hyperpolarization activatedinward current (If ) in guinea-pig dissociated sinoatrial node cells.Br J Pharmacol 110:343–349.
Briggs I, BoSmith RE, Heapy CG. 1994. Effects of Zeneca ZD7288in comparison with alinidine and UL-FS 49 on guinea pig sinoa-trial node and ventricular action potentials. J Cardiovasc Pharmacol24:380–387.
Cardenas CG, Mar LP, Vysokanov AV, Arnold PB, Cardenas LM,Surmeier DJ, Scroggs RS. 1999. Serotonergic modulation of hyper-polarization-activated current in acutely isolated rat dorsal rootganglion neurons. J Physiol 518 (Pt 2):507–523.
Caruana DA, Sorge RE, Stewart J, Chapman CA. 2006. Dopamine hasbidirectional effects on synaptic responses to cortical inputs in layerII of the lateral entorhinal cortex. J Neurophysiol 96:3006–3015.
Chalmers DT, Watson SJ. 1991. Comparative anatomical distributionof 5-HT1A receptor mRNA and 5-HT1A binding in rat brain—Acombined in situ hybridisation/in vitro receptor autoradiographicstudy. Brain Res 561:51–60.
Chapin EM, Andrade R. 2001. A 5-HT(7) receptor-mediated de-polarization in the anterodorsal thalamus. II. Involvement of thehyperpolarization-activated current I(h). J Pharmacol Exp Ther 297:403–409.
Chrobak JJ, Lorincz A, Buzsaki G. 2000. Physiological patterns in thehippocampo-entorhinal cortex system. Hippocampus 10:457–465.
Colino A, Halliwell JV. 1987. Differential modulation of 3 separateK-conductances in hippocampal CA1 neurons by serotonin. Nature328:73–77.
Corkin S, Amaral DG, Gonzalez RG, Johnson KA, Hyman BT. 1997.H.M.’s medial temporal lobe lesion: Findings from magnetic reso-nance imaging. J Neurosci 17:3964–3979.
Corradetti R, Le-Poul E, Laaris N, Hamon M, Lanfumey L.1996. Electrophysiological effects of N-(2-(4-(2-methoxyphenyl)-1-piperazinyl)ethyl)-N-(2-pyridinyl) cyclohexane carboxamide (WAY100635) on dorsal raphe serotonergic neurons and CA1 hippocam-pal pyramidal cells in vitro. J Pharmacol Exp Ther 278:679–688.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 127
Hippocampus DOI 10.1002/hipo

Craven R, Grahame-Smith D, Newberry N. 1994. WAY-100635 andGR127935: Effects on 5-hydroxytryptamine-containing neurones.Eur J Pharmacol 271:R1–R3.
Craven RM, Grahame-Smith DG, Newberry NR. 2001. 5-HT1A and5-HT2 receptors differentially regulate the excitability of 5-HT-containing neurones of the guinea pig dorsal raphe nucleus in vitro.Brain Res 899:159–168.
Davies MF, Deisz RA, Prince DA, Peroutka SJ. 1987. Two distincteffects of 5-hydroxytryptamine on single cortical neurons. BrainRes 423:347–352.
Dickson CT, Alonso A. 1997. Muscarinic induction of synchronouspopulation activity in the entorhinal cortex. J Neurosci 17:6729–6744.
Dickson CT, Trepel C, Bland BH. 1994. Extrinsic modulation oftheta field activity in the entorhinal cortex of the anesthetized rat.Hippocampus 4:37–52.
Dickson CT, Kirk IJ, Oddie SD, Bland BH. 1995. Classification oftheta-related cells in the entorhinal cortex: Cell discharges arecontrolled by the ascending brainstem synchronizing pathway inparallel with hippocampal theta-related cells. Hippocampus 5:306–319.
Dickson CT, Mena AR, Alonso A. 1997. Electroresponsiveness ofmedial entorhinal cortex layer III neurons in vitro. Neuroscience81:937–950.
Dickson CT, Magistretti J, Shalinsky M, Hamam B, Alonso A. 2000a.Oscillatory activity in entorhinal neurons and circuits: Mechanismsand function. Ann N Y Acad Sci 911:127–150.
Dickson CT, Magistretti J, Shalinsky MH, Fransen E, Hasselmo M,Alonso A. 2000b. Properties and role of Ih in the pacing ofsubthreshold oscillations in entorhinal cortex layer II neurons.J Neurophysiol 83:2562–2579.
Dringenberg HC, Vanderwolf CH. 1998. Involvement of direct andindirect pathways in electrocorticographic activation. NeurosciBiobehav Rev 22:243–257.
Felix R, Sandoval A, Sanchez D, Gomora JC, De-la-Vega-Beltran JL,Trevino CL, Darszon A. 2003. ZD7288 inhibits low-thresholdCa(2þ) channel activity and regulates sperm function. BiochemBiophys Res Commun 311:187–192.
Fletcher A, Forster EA, Bill DJ, Brown G, Cliffe IA, Hartley JE, JonesDE, McLenachan A, Stanhope KJ, Critchley DJ, Childs KJ, Mid-dlefell VC, Lanfumey L, Corradetti R, Laporte AM, Gozlan H,Hamon M, Dourish CT. 1996. Electrophysiological, biochemical,neurohormonal and behavioural studies with WAY-100635, apotent, selective and silent 5-HT1A receptor antagonist. BehavBrain Res 73:337–353.
Gasparini S, DiFrancesco D. 1997. Action of the hyperpolarization-activated current (Ih) blocker ZD 7288 in hippocampal CA1neurons. Pflugers Arch 435:99–106.
Gasparini S, DiFrancesco D. 1999. Action of serotonin on the hyper-polarization-activated cation current (Ih) in rat CA1 hippocampalneurons. Eur J Neurosci 11:3093–3100.
Gloveli T, Schmitz D, Empson RM, Dugladze T, Heinemann U.1997. Morphological and electrophysiological characterization oflayer III cells of the medial entorhinal cortex of the rat. Neuro-science 77:629–648.
Gozlan H, El-Mestikawy S, Pichat L, Glowinski J, Hamon M. 1983.Identification of presynaptic serotonin autoreceptors using a newligand: 3H-PAT. Nature 305:140–142.
Grunschlag CR, Haas HL, Stevens DR. 1997. 5-HT inhibits lateralentorhinal cortical neurons of the rat in vitro by activation ofpotassium channel-coupled 5-HT1A receptors. Brain Res 770:10–17.
Hamam BN, Kennedy TE, Alonso A, Amaral DG. 2000. Morphologi-cal and electrophysiological characteristics of layer V neurons ofthe rat medial entorhinal cortex. J Comp Neurol 418:457–472.
Hamam BN, Amaral DG, Alonso AA. 2002. Morphological andelectrophysiological characteristics of layer V neurons of the ratlateral entorhinal cortex. J Comp Neurol 451:45–61.
Harris NC, Constanti A. 1995. Mechanism of block by ZD 7288 ofthe hyperpolarization-activated inward rectifying current in guineapig substantia nigra neurons in vitro. J Neurophysiol 74:2366–2378.
Hu H, Vervaeke K, Storm JF. 2002. Two forms of electrical resonanceat theta frequencies, generated by M-current, h-current and persis-tent Naþ current in rat hippocampal pyramidal cells. J Physiol 545(Pt 3):783–805.
Hutcheon B, Miura RM, Puil E. 1996. Subthrehold membraneresonance in neocortical neurons. J Neurophysiol 76:683–697.
Hwang LL, Dun NJ. 1998. 5-Hydroxytryptamine responses in imma-ture rat rostral ventrolateral medulla neurons in vitro. J Neurophy-siol 80:1033–1041.
Jones RSG. 1994. Synaptic and intrinsic properties of neurons oforigin of the perforant path in layer II of the rat entorhinal cortexin vitro. Hippocampus 4:335–353.
Karschin C, Dissmann E, Stuhmer W, Karschin A. 1996. IRK(1–3)and GIRK(1–4) inwardly rectifying Kþ channel mRNAs are dif-ferentially expressed in the adult rat brain. J Neurosci 16:3559–3570.
Klink R, Alonso A. 1997a. Ionic mechanisms of muscarinic depolari-zation in entorhinal cortex layer II neurons. J Neurophysiol 77:1829–1843.
Klink R, Alonso A. 1997b. Muscarinic modulation of the oscillatoryand repetitive firing properties of entorhinal cortex layer II neurons.J Neurophysiol 77:1813–1828.
Kohler C. 1984. The distribution of serotonin binding sites in thehippocampal region of the rat brain. An autoradiographic study.Neuroscience 13:667–680.
Kohler C, Steinbusch H. 1982. Identification of serotonin and non-serotonin containing neurons of the mid-brain raphe projecting tothe entorhinal area and the hippocampal formation. A combinedimmunohistochemical and fluorescent retrograde tracing study inthe rat brain. Neuroscience 7:951–975.
Larkman PM, Kelly JS. 1992. Ionic mechanisms mediating 5-hydroxy-tryptamine- and noradrenaline-evoked depolarization of adult ratfacial motoneurones. J Physiol 456:473–490.
Larkman PM, Kelly JS. 1995. The use of brain slices and dissociatedneurones to explore the multiplicity of actions of 5-HT in thecentral nervous system. J Neurosci Methods 59:31–39.
Larkman PM, Kelly JS. 1998. Characterization of 5-HT-sensitivepotassium conductances in neonatal rat facial motoneurones invitro. J Physiol 508 (Pt 1):67–81.
Lavenex P, Amaral DG. 2000. Hippocampal-neocortical interaction:A hierarchy of associativity. Hippocampus 10:420–430.
Leishman DJ, Boeijinga PH, Galvan M. 1994. Differential effects ofcentrally-active antihypertensives on 5-HT1A receptors in rat dorso-lateral septum, rat hippocampus and guinea-pig hippocampus. Br JPharmacol 111:318–324.
Maccaferri G, McBain CJ. 1996. The hyperpolarization-activatedcurrent (Ih) and its contribution to pacemaker activity in rat CA1hippocampal stratum oriens-alveus interneurones. J Physiol 497:119–130.
Nadel L, Moscovitch M. 1997. Memory consolidation, retrogradeamnesia and the hippocampal complex. Curr Opin Neurobiol 7:217–227.
Neher E. 1992. Correction for liquid junction potentials in patchclamp experiments. Methods Enzymol 207:123–131.
Oh U, Ho YK, Kim D. 1995. Modulation of the serotonin-activatedKþ channel by G protein subunits and nucleotides in rat hippo-campal neurons. J Membr Biol 147:241–253.
Pape H-C, McCormick DA. 1989. Noradrenaline and serotoninselectively modulate thalamic burst firing by enhancing a hyperpo-larization-activated cation current. Nature 340:715–718.
Pazos A, Palacios JM. 1985. Quantitative autoradiographic mappingof serotonin receptors in the rat brain. I. Serotonin-1 receptors.Brain Res 346:205–230.
128 MA ET AL.
Hippocampus DOI 10.1002/hipo

Penington NJ, Kelly JS, Fox AP. 1993. Whole-cell recordings ofinwardly rectifying Kþ currents activated by 5-HT1A receptors ondorsal raphe neurones of the adult rat. J Physiol 469:387–405.
Pralong E, Jones RS. 1993. Interactions of dopamine with glutamate-and GABA-mediated synaptic transmission in the rat entorhinalcortex in vitro. Eur J Neurosci 5:760–767.
Pralong E, Magistretti PJ. 1995. Noradrenaline increases K-conduct-ance and reduces glutamatergic transmission in the mouse entorhi-nal cortex by activation of a2-adrenoreceptors. Eur J Neurosci 7:2370–2378.
Rosenkranz JA, Johnston D. 2006. Dopaminergic regulation of neuro-nal excitability through modulation of Ih in layer V entorhinalcortex. J Neurosci 26:3229–3244.
Schmitz D, Empson RM, Gloveli T, Heinemann U. 1995a. Serotoninreduces synaptic excitation of principal cells in the superficial layersof rat hippocampal-entorhinal cortex combined slices. NeurosciLett 190:37–40.
Schmitz D, Empson RM, Heinemann U. 1995b. Serotonin and8-OH-DPAT reduce excitatory transmission in rat hippocampalarea CA1 via reduction in presumed presynaptic Ca2þ entry. BrainRes 701:249–254.
Schmitz D, Empson RM, Gloveli T, Heinemann U. 1997. Serotoninblocks different patterns of low Mg2þ-induced epileptiform activityin rat entorhinal cortex, but not hippocampus. Neuroscience 76:449–458.
Schmitz D, Gloveli T, Behr J, Dugladze T, Heinemann U. 1998a.Subthreshold membrane potential oscillations in neurons of deeplayers of the entorhinal cortex. Neuroscience 85:999–1004.
Schmitz D, Gloveli T, Empson RM, Draguhn A, Heinemann U.1998b. Serotonin reduces synaptic excitation in the superficialmedial entorhinal cortex of the rat via a presynaptic mechanism.J Physiol 508 (Pt 1):119–129.
Schmitz D, Gloveli T, Empson RM, Heinemann U. 1998c. Compari-son of the effects of serotonin in the hippocampus and the entorhi-nal cortex. Mol Neurobiol 17:59–72.
Schmitz D, Gloveli T, Empson RM, Heinemann U. 1998d. Serotoninreduces polysynaptic inhibition via 5-HT1A receptors in the super-ficial entorhinal cortex. J Neurophysiol 80:1116–1121.
Schmitz D, Gloveli T, Empson RM, Heinemann U. 1999. Potentdepression of stimulus evoked field potential responses in the medialentorhinal cortex by serotonin. Br J Pharmacol 128:248–254.
Scoville WB, Milner B. 1957. Loss of recent memory after bilateralhippocampal lesions. J Neurol Neurosurg Psychiatry 20:11–21.
Sim LJ, Xiao R, Childers SR. 1997. In vitro autoradiographic localiza-tion of 5-HT1A receptor-activated G-proteins in the rat brain.Brain Res Bull 44:39–45.
Sizer AR, Kilpatrick GJ, Roberts MH. 1992. A post-synaptic depres-sant modulatory action of 5-hydroxytryptamine on excitatoryamino acid responses in rat entorhinal cortex in vitro. Neurophar-macol 31:531–539.
Squire LR, Zola-Morgan S. 1996. Structure and function of declara-tive and nondeclarative memory systems. Proc Natl Acad Sci USA93:13515–13522.
Stefanacci L, Buffalo EA, Schmolck H, Squire LR. 2000. Profoundamnesia after damage to the medial temporal lobe: A neuroana-tomical and neuropsychological profile of patient E. P. J Neurosci20:7024–7036.
Stenkamp K, Heinemann U, Schmitz D. 1998. Dopamine suppressesstimulus-induced field potentials in layer III of rat medial entorhi-nal cortex. Neurosci Lett 255:119–121.
Swanson LW, Kohler C, Bjorklund A. 1987. The limbic region. I. Theseptohippocampal system. In: Bjorklund A, Hokfelt T, Swanson LW,editors. Handbook of Chemical Neuroanatomy, Vol 5. Amsterdam:Elsevier.
van der Linden S, Lopes da Silva FH. 1998. Comparison of the elec-trophysiology and morphology of layers III, II neurons of the ratmedial entorhinal cortex in vitro. Eur J Neurosci 10:1479–1489.
Vanderwolf CH. 1988. Cerebral activity and behavior: Control bycentral cholinergic and serotoninergic systems. Int Rev Neurobiol30:225–340.
Vertes RP, Kocsis B. 1997. Brainstem-diencephalo-septohippocampalsystems controlling the theta rhythm of the hippocampus. Neuro-science 81:893–926.
Wright DE, Seroogy KB, Lundgren KH, Davis BM, Jennes L. 1995.Comparative localization of serotonin1A, 1C, and 2 receptor sub-type mRNAs in rat brain. J Comp Neurol 351:357–373.
Yamada M, Inanobe A, Kurachi Y. 1998. G protein regulation ofpotassium ion channels. Pharmacol Rev 50:723–760.
Zhou FC, Patel TD, Swartz D, Xu Y, Kelley MR. 1999. Productionand characterization of an anti-serotonin 1A receptor antibodywhich detects functional 5-HT1A binding sites. Brain Res MolBrain Res 69:186–201.
INTRINSIC EFFECTS OF 5-HT IN ENTORHINAL CORTEX 129
Hippocampus DOI 10.1002/hipo