effects of selected phytohormones on the growth …/67531/metadc163919/m2/1/high... · effects of...
TRANSCRIPT

EFFECTS OF SELECTED PHYTOHORMONES ON THE GROWTH
AND MORPHOLOGY OF ESCHERICHIA COLI
APPROVED:
/ A\ /UJ-IUAA • ^ CUA*J^ Major Professor
L L't ' /\ Mindir Professor
\ ' / ;. i-
rVr"~ V ) I Director of the Department of Biology
Dean of the Graduate School

EFFECTS OF SELECTED PHYTOHORMONES ON THE GROWTH
AND MORPHOLOGY OF ESCHERICHIA COLI
THESIS
Presented to the Graduate Council of the
North Texas State University in Partial
Fulfillment of the Requirements
For the Degree of
MASTER OF SCIENCE
By
Lynn Mallory Little, B. S.
Denton, Texas
January, 1968

TABLE OF CONTENTS
Page
LIST OF TABLES .iv
LIST OF ILLUSTRATIONS v
Chapter
I. INTRODUCTION 1
II. MATERIALS AND METHODS 8
III. RESULTS 14
IV. DISCUSSION . . . . . . .22
BIBLIOGRAPHY • . »32
ill

LIST OF TABLES
Table Page
I. Concentrations of Phytohormones added to trypti-case soy broth cultures of Escherichia coli . 11
IV

LIST OF ILLUSTRATIONS
Figure Page
1. Growth curves for cultures treated with IAA. . . . 15 *
2. Growth curves for cultures treated with GA3. . . . 16
3. Growth curves showing that a lower concentration of IAA has negligible effect on growth, but that higher concentrations of GA3 and es-pecially IAA are inhibitory 17
4. Growth curves for cultures containing a stimula-tory concentration of GA3 and ineffective concentrations of kinetin, or a stimulatory concentration of GA3 and an inhibitory con-centration of IAA .18
5. Growth curves for inhibitory concentrations of GA3 and IAA tested separately and together. . 19
6. Representative photographs of untreated cells and cells treated with 1.0 mg/ml IAA, 5.0 mg/ml IAA, or 1.0 mg/ml IAA + 1.0 mg/ml GA3 . . . . . . . . . . . . . . . . . . 2 0

CHAPTER I
INTRODUCTION
Growth in plants is regulated by groups of substances
known as phytohormones. On the basis of their structural
similarities and modes of action, three major groups of
phytohormones have been identified and characterized,
auxins, gibberellins, and kinins.
Kogl, _et a l. (13) first described indoleacetic acid
as an auxin, and this chemical was purified from plant
materials by Kogl and Kostermans (14) in 1934 and by
Thimann (24) a year later. It is now known that in the
individual plant cell, auxins affect the plasticity and
elasticity of the cell wall, cytoplasmic viscosity,
protoplasmic streaming, respiration rates, metabolic
pathways, changes in oxidative states, content of nucleic
acids, and the activities of many enzymes (17).
In 1926, Kurosawa (16) in Japan was studying a disease
of rice caused by the fungus Gibberella fujikoroi which
produced a characteristic excessive growth of the rice
plant.. The separation of the responsible growth stimulant
closely preceded the first separation of auxin from plants
in 1928, and in 1935 Yabuta (25) crystalized the compound

and named it gibberellin. In general, it appears that the
gibberelliris are involved in the regulatory systems which
bring about the developmental response of the plant to envi-
ronmental cues (17).
The generic name "kinin" was proposed for chemical
substances which stimulate cytokinesis. The purines and
other plant kinins began to receive attention in physio-
logical literature in the 1940s, and in 1955 Miller and
his team (21) separated an active stimulant of cell division
from yeast DNA. This material, which they named kinetin,
was identified as 6-furfuryl aminopurine or 6-furfuryl-
adenine (19, 20). In addition to its effects on cell
division and bud differentiation, kinetin has been found
to have a variety of other growth effects, although the
overall plant responses to kinins have been much less
dramatic than those to auxins or gibberellins (17).
The effect of the presence of an auxin on the growth
of bacterial populations was first studied by Ball (1), who
reported that indoleacetic acid in concentrations of 0.0001
mg/ml to 0.001 mg/ml increased by more than two-fold the
final stationary phase titer of Escherichia coli suspensions.
Later, Handler and Kamin (7).could not detect a stimulation
of Escherichia coli growth using the same concentrations

of the auxin. According to Beckwith and Geary (2), indole-
acetic acid at concentrations of 0.0003 mg/ntl to 0.2 mg/ml
will stimulate the growth of Escherichia coli cells taken
from the logarithmic death phase, while a concentration of
10.0 mg/ml is completely inhibitory. Fletcher (5) reported
that low concentrations of auxins can stimulate the growth
of certain bacteria, but, as he pointed out, other workers
have found that auxins have little or no effect on micro-
organisms or that at higher concentrations they are definitely
toxic. An interesting finding by Dubos (4) is that peptone
and especially tryptophan can partially or completely
reverse the bacteriostatic effect of auxins.
Saono (23) tested the effect of gibberellic acid on the
growth of thirty-eight species of bacteria and two species
each of the algae Chlorella and Scenedesmus. Except for
two strains of Azotobacter chroococcum and Actinomycetes
coelicolor, the bacteria did not react to the acid at con-
centrations of 0.0005 mg/ml to 0.05 mg/ml. However, lower
concentrations of the acid stimulated multiplication of all
four algal species.
Kennell (12) reported that Escherichia coli cultures
grown' in a salts-glucose medium showed accelerated growth
when kinetin was present at a concentration of 0.001 mg/ml.

Maruzzella and Garner (18) found that concentrations of
kinetin from 0.002152 mg/ml to 0.00000002152 mg/ml exerted
a marked stimulatory effect on the growth of Bacillus mega-
terium and Agrobacterium tumefaciens and to a lesser degree
on Escherichia coli, Staphylococcus aureus, and Erwinia
carotovara. The growth of Corynebacterium michiganense was
partially inhibited by kinetin at most concentrations used.
Interestingly, no change in cell morphology or Gram stain
reaction was observed in any of the tests.
While numerous investigators have studied the effects
of auxins, gibberellins, and kinins on the growth of various
microorganisms, there have been no reported attempts to
determine whether treatment with more than one phytohormone
could cause a synergistic response in either enhancing or
inhibiting the growth of a bacterial population. Such
synergism with these groups of phytohormones has been often
reported on the growth of higher plants (3, 6, 8, 9, 10, 11,
15, 22, 26). The present investigation was undertaken as a
preliminary study to learn the response of Escherichia coli
cells grown under identical experimental conditions to
various concentrations of indoleacetic acid, gibberellic acid,
and kinetin alone, and in combination with one another.

CHAPTER BIBLIOGRAPHY
1. Ball, Earnest, "Heteroauxin and the Growth of Escherichia coli," Journal of Bacteriology, XXXVI (1938), 559-565.
2. Beckwith, T. D. and E. M. Geary, "Effect of Indole 3 Acetic Acid on Multiplication of Esch. coli and J3. typhosa," Journal of Infectious Diseases, LXVI (1940), 78-79.
3. Brian, P. W., "Role of Gibberellin-like Hormones in Regulation of Plant Growth and Flowering," Nature CLXXXI (1958), 1122-1123.
4. Dubos, Rene J., "Inhibition of Bacterial Growth by Auxins," Proceedings of the Society for Experimental Biology, New York, LXIII (1946), 317-319.
5. Fletcher, Wm. W., "The Effect of Herbicides on Soil Micro-organisms," The Physiology and Biochemistry of Herbicides, edited by L. J. Audus, London, Academic Press, 1964, pp. 20-62.
6. Galston, A. W. and H. Warburg, "An Analysis of Auxin-Gibberellin Interaction in Pea Stem Tissue," Plant Physiology, XXXIV (1959), 16-22.
7. Handler, P. and H. Kamin, "Indole Acetic Acid and Growth of Bacteria with Varying Requirements for Nicotinic Acid and Tryptophan," Proceedings of the Society for Experimental Biology, New York, LXVI (1948), 251-254.
8. Hillman, W. S. and W. K. Purves, "Does Gibberellin Act Through an Auxin-mediated Mechanism?," Plant Growth Regulation, edited by R. M. Klein, Ames, Iowa, Iowa State University Press, 1961, pp. 589-600.
9. Kato, J., "Physiological Action of Gibberellin with Special Reference to Auxin," Plant Growth Regulation, edited by R. M. Klein, Ames, Iowa, Iowa State Univer-sity Press, 1961, pp. 601-609.

10. , "Studies on the Physiological Effect of Gibberellin II. On the Interaction of Gibberellin with Auxins and Growth Inhibitors," Physiologia Plantarum, XI (1958), 10-15.
11. Kefford, N. P., "Auxin-Gibberellin Interactions in Rice Coleoptile Elongation," Plant Physiology, XXXVII (1962), 380-386.
12. Kennell, D. E., "The Effects of Indoleacetic Acid and Kinetin on the Growth of Some Microorganisms," Experimenta1 Cell Research, XXI (1960), 19-33.
13. Kogl, F., A. J. Haagen-Smit and H. Erxleben, "Uber ein Neues Auxin ('Heteroauxin') aus Harn. XI.," Zeitschrift Physiologisch Chemie, CCXXVIII (1934), 90-103.
14. Kogl, F. and D. G. F. R. Kostermans, "Hetero-auxin als Stoffwechselprodukt Niederer Pflanzlicher Organismen. XIII.," Zeitschrift Physiologisch Chemie, CCXXVIII (1934), 113-121.
15. Kuraishi, S. and R. M. Muir, "Increase in Diffusible Auxin After Treatment with Gibberellin," Science, CXXXVII (1962), 760-761.
16. Kurosawa, E., "Experimental Studies on the Secretion of Fusarium heterosporum on Rice Plants," Trans-actions of the Natural History Society of Formosa, XVI (1926), 213-227.
17. Leopold, A. Carl, Plant Growth and Development, New York, McGraw-Hill Book Company, 1964.
18. Maruzella, Jasper C. and James G. Garner, "Effect of Kinetin on Bacteria," Nature, CC (1963), 385.
19. Miller, C. 0., F. Skoog, F. S. Okumura, M. H. von Saltza and F. M. Strong, "Isolation, Structure, and Synthesis of Kinetin, a Substance Promoting Cell Division," Journal of the American Chemical Society, LXVIII (1956), 1375-1380.

» 5 IJ L : '' "
20. "Structure and Synthesis of Kinetin,"
Journal of the American Chemical Society/ LXXVII (1955), 2662-2663.
21. Miller, C. 0., P. Skoog, M. H. von Saltza and F. M. Strong, "Kinetin: a Cell Division Factor from Deoxyribonucleic Acid, "Journal of the American Chemical Society, LXVIII (1955), 1392.
22. Phillips, I. D. J., A. J. Vlitos and H. Cutler, "The Influence of Gibberellic Acid Upon the Endogenous Growth Substances of the Alaska Pea," Contributions to the Boyce Thompson Institute, XX (1959), 111^-120.
23. Saono, Susono, "Effect of Gibberellic Acid on the Growth and Multiplication of Some Soil Microorganisms and Unicellular Green Algae," Nature, CCIV (1964), 1328-1329.
24. Thimann, K. V., "On the Plant Growth Hormone Produced by Rhizopus suinus," Journal of Biological Chemistry, CIX (1935), 279-291.
25. Yabuta, T., "Biochemistry of the 'Bakanae' Fungus of Rice," Agriculture and Horticulture (Tokyo), X (1935), 17-22.
26. Yoda, S. and J. Ashida, "Effects of Gibberellin and Auxin on the Extensibility of the Pea Stem," Plant Cell Physiology (Tokyo), I (1960), 99-105.

CHAPTER II
MATERIALS AND METHODS
Escherichia coli ATCC 11303 cells were grown in the
dark at 37° ^1° C in 10-rtil volumes of trypticase soy broth
(TSB, Baltimore Biological Laboratory) contained in 13 x 150
mm colorimetrically-matched screw-cap Pyrex test tubes. The
tubes were inoculated by transferring 1„0 ml from a 12-hr
culture through three 9-ml sterile distilled water dilution
tubes and pipeting 0.1 ml from the third dilution tube into
each culture tube. A modification of the method of Scott
and Chu (1) of obtaining synchronized cultures was employed.
This consisted of refrigerating the dilution tubes and hold-
ing the cells at 5° ^1° C for one hour before inoculating
into the 37° culture tubes. Synchrony of division, however,
was not achieved, as the generation time was found to be
approximately forty minutes. Some cultures were treated by
including in the growth medium various concentrations of
indole-3-acetic acid (IAA), gibberellic acid (GA^), and/or
kinetin, all of which were obtained from Nutritional Bio-
chemical Corporation, Cleveland, Ohio.
8

The number of cells present in a given culture was
determined in two ways. First, replicate plate counts
were made on cultures which did not contain hormone after
0, 1, 2, 3, 4, 5, 6, 7, 8, 12, 24, 48, and 96 hours of
incubation. At the same time intervals, the tubes were
read colorimetrically on a Bausch and Lomb Spectronic 20
Colorimeter at a wavelength of 650 mp.. A regression line
was plotted from the data. Second, on other cultures not
treated with hormone, replicate direct counts were made at
these same time intervals using a Petroff-Hausser Counting
Chamber. Colorimetric readings were made as before, and a
regression line was plotted from the data. Other untreated
cultures were both plated out and read directly to jfcompare
the two methods of counting. It was found that the number
of cells as determined by direct counts equalled approxi-
2
mately 1.6 x 10 times the number of cells as determined by
plate counts. In all subsequent experiments the number of
bacteria present was determined colorimetrically on the
basis pf the turbidity-versus-direct counts regression line, r 'I A
which had a standard error of -1.3953 x 10" cells/ml at a
confidence level of P = 0.05.
In experiments employing IAA and/or GA^, the desired
quantity of phytohormone was added directly to the culture

10
broth. These tubes were then heated and shaken to facilitate
dissolution of the hormones before the culture tubes were
autoclaved at 121° C for fifteen minutes. Kinetin, insoluble
in water, was dissolved in 10% NaOH before being added to the
broth. Ten per cent HCl was then added to the tubes to
bring the medium back to its original pH of 7.3 -0.2 (2).
All .experiments involving phytohormones were performed
in triplicate, and all concentrations were tested at least
twice. Controls consisted of culture tubes containing 10 ml
TSB but no hormone. Also,, in experiments employing kinetin,
additional controls consisted of tubes containing TSB plus
the same quantities of 10% NaOH and 10% HCl as were added to
the kinetin-containing culture tubes. These tubes received
no hormone.
Culture tubes were inoculated as previously described
from inoculum cultures which were checked for Escherichia
coli growth by Gram staining and streaking on eosin wethylene
blue agar plates. The culture tubes were read coloripietric-
ally upon inoculation and after 3, 4, 5, 6, 7, 8, 10, 12,
48, and 72 hours of incubation. Before each reading, each
tube was shaken on a Super-Mixer (Lab-Line Instruments, Inc.)
for twenty seconds to bring about a homogeneous dispersion
of cells. Tubes were held in a 37° water bath while out of

11
TABLE I
CONCENTRATIONS OF PHYTOHORMONES ADDED TO TRYPTICASE SOY BROTH CULTURES OF ESCHERICHIA COLI
Phytohormones Employed Singly*
IAA GAi Kinetin
0.0001 0.0001 0.0001 0.001 0.001 0.001 0.01 0.01 0.01 0.1 0.1 0.1 1.0 1.0 5.0 5.0
Phytohormones in Combination*
1.0 GA3 +0.1 IAA 1.0 GA3 + 1.0 IAA 5.0 GA3 + 1.0 IAA 0.1 GA3 +0.01 IAA
0.1 GA3 + 0.1 Kinetin 1.0 GA3 + 0.01 Kinetin 1.0 GA3 + 0.001 Kinetin 0.01 IAA +0.1 Kinetin
0.1 IAA +0.1 Kinetin +0.1 GA3
*All concentrations are given in milligrams phyto-hormone per milliliter trypticase soy broth (mg/ml) .
the incubator for reading. Gram stains were made on all cul-
tures after 8, 24, 48, and 72 hours of incubation; these
stains were examined by phase contrast microscopy under oil
immersion at 1000 x magnification using a Nikon Microflex

12
Model EFM microscope so that the morphology of hormone-
treated cells could be compared with that of cells from
control cultures. Photographs of representative cells were
taken on Kodak 35 mm TRI-X Pan film with a Nikon M-35 dark
box and Type B camera adapter.
The actual concentrations of the phytohormones which
were tested singly and in combination are listed in Table 1.
The method employed in dissolving kinetin did not make it
possible to prepare concentrations of this compound as high
as were used of IAA and GA3.

CHAPTER BIBLIOGRAPHY
1. Scott, D. B. M. and E. Chu, "Synchronized Division of Growing Cultures of 12. coli," Experimental Cell Research, XIV (1958), 166-174.
2. Skinner, C. G., personal correspondence. North Texas State University, Denton, Texas, July 31, 1967.
13

CHAPTER III
RESULTS
Growth responses greater than plus one or less than
minus one standard deviation from the control mean were
considered to be attributable to the phytohormone(s) present,
The results of experiments in which stimulation (growth
response of more than plus one standard deviation from the
control mean or inhibition (growth response of less than
minus one standard deviation from the control mean) occurred
are presented in Figures 1-5. In each case, the vertical
lines represent -1 standard deviation from the mean for
each growth curve at 24, 48, and 72 hours of incubation.
It should be noted that the numbers of cells reported
are based on the size of cells occurring in control cultures,
In cultures having larger cells, the numbers reported are
erroneously high, although accurate in terms of total cell
density per culture.
It can be seen in Figure 1 that growth was inhibited
in cultures containing 1.0 mg/ml IAfl». Less growth occurred
in cultures treated with 0.1 mg/ml IAA than in the control
cultures, but this difference was not appreciable at 48 or
72 hours.

15
u CD -P •H trH *H r-4 i—1 *H a
a. to
a; o M-l O U a)
3 !3
2 x 10 10
1 x 10" 10
1—TSB
12 24
2—0.1 mg/ml IAA
3—1.0 mg/ml IAA
36 48 60
Hours
72
Fig. 1—Growth curves for cultures treated with IAA. Vertical lines represent +1 standard deviation from the mean.
Figure 2 shows that more growth occurred in cultures
containing either 0.1 mg/ml GA3 or 1.0 mg/ml GA3 than in
TSB cultures. Only the latter GA3 concentration elicited a
level of growth greater than one standard deviation above
the control mean.
In Figure 3 it can be seen that 0.01 mg/ml IAA had no
appreciable effect on Escherichia coli growth, but that both
5.0 mg/ml GA3 and 5.0 mg/ml IAA caused inhibition. The

16
u Q) -P "H rH
•H a U <D Ok
w i~H rH <l) o
o
<D
2 x 10 10
1 x 10 10
12
1—1.0 mg/ml GA^
2—0.1 mg/ml GA3
3—TSB
24 36 48
Hours
60 72
Fig. 2—Growth curves for cultures treated with GA3. Vertical lines represent ^1 standard deviation from the mean,
latter IAA concentration proved to be so inhibitory as to
prevent the rapid growth characteristic of bacterial cultures
during the early hours of incubation.
The concentrations of kinetin used had no discernable
effect on growth when tested alone. Figure 4 shows that
combinations of 1.0 mg/ml GA-j + 0.01 mg/ml kinetin and
1.0 mg/ml GA3 -f 0.001 mg/ml kinetin gave growth responses
greater than one standard deviation above the control mean
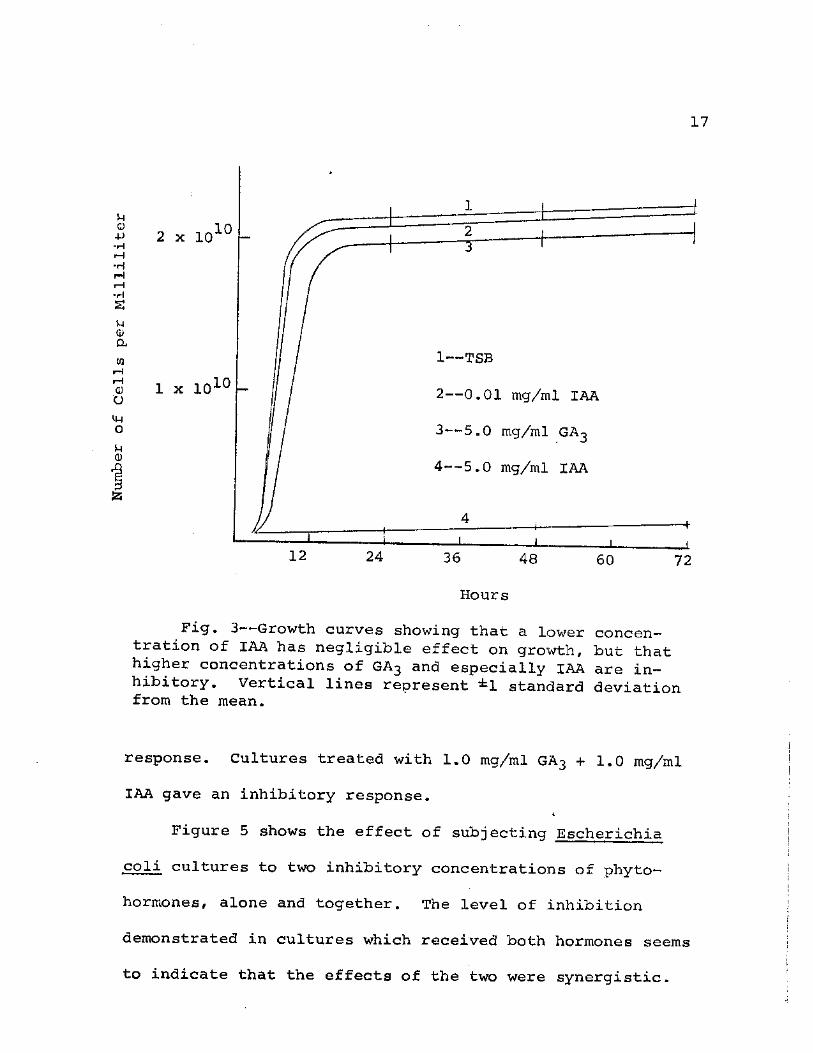
17
u Q)
•H rH -HI H iH -H a M CP O. CTJ
<D U
o n <D
3 S3
2 x 10
1 x 10 -
1—TSB
2—0,01 mg/ml IAA
3—5*0 mg/ml GA3
4—5.0 mg/ml IAA
Hours
Fig. 3—-Growth curves showing that a lower concen-tration of IAA has negligible effect on growth, but that higher concentrations of GA3 and especially IAA are in— hibitory. Vertical lines represent ±1 standard deviation from the mean.
response. Cultures treated with 1.0 mg/ml GA3 + 1.0 mg/ml
IAA gave an inhibitory response. 4
Figure 5 shows the effect of subjecting Escherichia
coli cultures to two inhibitory concentrations of phyto-
hormones, alone and together. The level of inhibition
demonstrated in cultures which received both hormones seems
to indicate that the effects of the two were synergistic.

18
2.4 x 10 10
u <y -p -H
•H a n <u a 03
r - i iH a; o m o u ai
2.2 x 10 10
2.0 x 10 10
1.8 x 10 10
12 24
1—1.0 GA3 + 0.001 Kinetin 2—1.0 GA3 + 0.01 Kinetin 3—1.0 GA3 +0.1 IAA 4—TSB 5—0.1 NaOH + 0.21 HCl 6—1.0 GAo + 1.0 IAA
36 48 60 72
Hours
Fig. 4—Growth curves for cultures containing a stimu-latory concentration of GA3 and ineffective concentrations of kinetin (1 and 2) or IAA (3), and for cultures containing a stimulatory concentration of GA3 and an inhibitory concen-tration of IAA (6). Values given in the figure are in mg/ml except for 5, the latter representing ml 10% acid or base. Vertical lines represent *1 standard deviation from the mean
The only cells that were morphologically distinguishable
from control cells were those treated with 1,0 mg/ml IAA,
5.0 mg/ml IAA, or 1.0 mg/ml IAA + 1.0 mg/ml GAg. All cultures

19
u Q) -P • H
-H g
U Q) a. CQ r—i .H <P o in o u QJ
3 53
2 x 10 10
1 x 10 10
1—TSB
2 — 5 .0 mg/ml GA3
3—1.0 mg/ml IAA
4—5.0 mg/ml GA3
+ 1.0 mg/ml IAA
12 24 36 48
Hours
60 a 72
Pig. 5—Growth curves for inhibitory concentrations of GA3 and IAA tested separately and together. Inhibition is seen to be synergistic in the latter case. The dashed line represents the growth curve that would be expected if the inhibition were only additive. Vertical lines represent ±1 standard deviation from the mean.
which received these concentrations of phytohormones contained
many cells that were larger than those found in any other
cultures. The greatest over-all increase in cell size was
seen in cultures treated with 1.0 mg/ml IAA. Many of these
cells were approximately one-and-one-half to two-and-one-half
times longer and larger in diameter than control cells, and
tended to be more nearly coccoid in shape. Cultures containing

20
mm r
i U
• o
Fig. 6—Representative photographs of untreated Escherichia coli cells (A) and of cells treated with 1.0 mg/ml IAA (B), 5.0 mg/ml IAA (C), and 1.0 mg/ml IAA + 1 . 0 mg/ml GA3. All photographs were taken using phase contrast microscopy after seventy—two hours incubation.
5.0 mg/ml IAA produced cells that were generally larger in
diameter than control cells (though not as large as cells
treated with 1.0 mg/ml IAA), and longer by one and one-half
to several times the length of control cells. Those cells
treated with 1.0 mg/ml IAA + 1 . 0 mg/ml GA3 were generally
about the same size as control cells, to slightly larger,
and tended to be more nearly coccoid.

21
No important differences in Gram stain reaction were
noted. Representative photographs of cells occurring in
cultures treated with the three concentrations of hormones
mentioned and in control cultures are included in Figure 6,

CHAPTER IV
DISCUSSION
While some of the phytohormone concentrations used may
appear unusually high in comparison with those which have
been reported to affect plant growth, they were chosen on
the basis of results of experiments performed preparatory to
this investigation in which Escherichia coli cultures were
grown in the presence of graded concentrations of the
hormones for twenty-four hours and were then plated out on
trypticase soy agar plates so that colony counts could be
made. The particular concentrations of the three phyto-
hormones chosen to be tested in combination with one another
were selected on the basis of their effects on the growth
of Escherichia coli when used singly.
The results indicate that when indole-3-acetic acid was
included in the culture medium of Escherichia coli cells,
growth was slightly inhibited by a concentration of 0.1 mg/ml,
increasingly inhibited by a concentration of 1.0 mg/ml, and
almost completely inhibited by 5.0 mg/ml. When gibberellic
acid was included in the culture medium, 0.1 mg/ml showed no
22

23
effect on growth, 1.0 mg/ml gave a substantial increase in
growth, and 5.0 mg/ml was definitely inhibitory. Kinetin
had no apparent effect on either the growth or the morphology
°f Escherichia coli in any concentration used? the same is
true for the quantities of 10% NaOH and 10% HCl which were
used.
Since 1.0 mg/ml GA^ proved stimulatory when used alone,
an experiment was designed to determine whether this concen-
tration of GA^ could still increase growth when tested in
combination with concentrations of IAA and kinetin which had
not evidently affected growth. It is interesting that both
1.0 mg/ml GA3 + 0.001 mg/ml kinetin and 1.0 mg/ml GA^ + 0.01
mg/ml kinetin stimulated growth (Figure 4), but that the
combination of 1.0 mg/ml GA^ + 0.1 mg/ml IAA gave a rate and
extent of growth which was not different from the control.
It appears that the potentially stimulatory effect of GA3
was in some way masked by the presence of IAA.
Another experiment was set up to determine whether two
hormone concentrations which were inhibitory alone might
produce a synergistic inhibitory response when tested
together. Such synergism would be apparent if the level of
inhibition (i,.e., the difference in the number of cells
between control cultures and experimental cultures) could

24
be shown to be greater in cultures containing both hormones
than the sums of the levels of inhibition caused by each
hormone alone. Synergistic inhibition did occur in cultures
containing 1.0 mg/ml IAA + 5.0 mg/ml GA3 as is shown in
Figure 5. The dashed line in this figure is the growth
curve that would be expected for a culture containing these
concentrations of hormones if the inhibitory effects were
additive, not synergistic.
The purpose of a final experiment was to determine
whether various concentrations of the three phytohormones
which had had no apparent effect on growth when tested alone
could produce an appreciable effect on growth when tested
together. The highest concentration of each hormone which
had shown no effect on growth (0.01 mg/ml IAA, 0.1 mg/ml GA3,
and 0.1 mg/ml kinetin) was selected, and these were tested
separately, in pairs, and all three together. Through
seventy-two hours of incubation, none of these cultures gave
a growth response different from the control mean by as
much ate 1 standard deviation.
Again it may be pointed out that in cultures containing
larger than normal cells, the total number of cells present
must actually have been less than the number reported since
cell-number was determined by optical density measurements

25
based on the size of cells occurring in control cultures.
Thus, the larger the individual cells in a culture, the fewer
cells needed to account for any particular optical density,
assuming that these larger cells were as dense throughout as
were control cells. This is, however, not necessarily an
entirely valid assumption since the larger cell-size may
have been a result of increased cell wall elasticity
rather than increased protoplasmic content (increased
density) per cell.
Until recently, there had been no reported explanation
of the mechanism of IAA-induced inhibition in bacterial
cells. In 1964, Fukuyama and Moyed (4) announced that ribo-
flavin-catalyzed photooxidation products are the inhibitory
agents, rather than the acid itself. One of these products
was tentatively characterized as 3-hydroxymethyloxindole,
and the other was identified as 3-methyleneoxindole. The
former compound was thought to be non-enzymatically converted
to the latter, which was in turn converted to 3-methyl-
oxindole, a non-toxic substance.
Later in the same year, Still, Fukuyama, and Moyed (15)
presented a possible mechanism of inhibition. The effect
of 3-methyleneoxindole was attributed to its high reactivity
with sulfhydryl-containing enzymes. Since inhibition of any

26
one of the large number of essential sulfhydryl enzymes
could result in inhibition of growth, these workers con-
sidered it unlikely that 3-methyleneoxindole has a single
major site of action. Instead, it is probable that the
functions of at least several critical systems are suspended.
Thus, these authors concluded, the enzymatic reduction of
3-methyleneoxindole to 3-methyloxindole may account only in
part for the transient nature of the bacteriostasis. Dubos
(3) pointed out in 1946 that peptone could partially or com-
pletely reverse the bacteriostatic effect of auxins. The
peptone content of the trypticase soy broth employed as the
growth medium in these experiments may have played a part
in reversing lAA-induced inhibition, but to what extent and
by what mechanism is not known.,
Conn and Stumpf (2) have commented that hormones have
not been recognized to function as either enzymes or co-
enzymes, but that they probably function in controlling
either;the synthesis or activation of enzymes. While it
has not been reported, it might be feasible that GA^, or
one of its catabolic products, could function by inducing
the production of an enzyme which could catalyze this
product's conversion to another substance. If this latter
substance were a substrate common to some metabolic pathway

27
of the cell, then the cell might conceivably be able to
carry on a higher rate of metabolism using this increased
supply of substrate. The fact that GA3 is stimulatory in
one concentration and inhibitory in another might suggest
that the inducing substance could cause the formation of a
product which is an inhibitor of the over-all reaction.
Thus, if too much substrate were present, the inhibitor
produced might slow the reaction by competing with the sub-
strate for some essential enzyme's active site.
In the case of kinetin, since none of the concentrations
used had any detectable effect, it cannot be.concluded that
this phytohormone altered the biosynthesis of Escherichia
coli cells.
It is of note that the only cultures which produced
cells morphologically distinguishable from untreated cells
were those containing either 1.0 mg/ml or 5.0 mg/ml IAA.
In these cultures, cells appeared which were both longer
and larger in diameter than control cells. It has been re-
ported (1, 8, 16) that the action of auxins, including IAA,
softens the walls of plant cells by increasing their plas-
ticity. With a softening of the wall, there ensues a
swelling of the cell by simple osmotic water uptake until
the restraining forces of the wall balance the osmotic

28
values of the cytoplasm. It seems reasonable that the
same phenomenon could occur in bacterial cells, thus
accounting for the enlarged appearance of cells grown in
the presence of IAA.
Gibberellin was discovered because of its ability to
increase growth in plants, and in this quality its effects
are more impressive than those of auxins (9). Whether the
gibberellin effect is due to a stimulation of cell division
or of cell enlargement has been extensively debated; "various
workers have presented data which seem to support one con-
clusion or the other (6, 7, 13, 14). In the case of bacterial
cells, the present investigation would tend to support the
argument for increased cell division, at least at some GA3
concentrations, but not for increased cell enlargement since
the only cultures treated with GA3 which produced cells larger
than control cells were also treated with IAA.
Since Miller's team separated kinetin from yeast DNA
and found it to be an active stimulant of cell division,
numerous reports have appeared on its ability to increase
cell division in plants. Kinetin also causes marked altera-
tions ;in the protein and nucleic acid components of plant
tissue (10,11), a circumstance which could be a basic part
of its effect on cell division (5). Partheir and Wollgiehn
(12) have reported that protein content, RNA, and DNA in

29
Nicotiana plants are all increased markedly by kinetin
treatment. An argument for similar activity in Escherichia
coli is not given credence by the present work, as kinetin
was not seen to have any effect on either growth rate or I
cell size.
The conclusions reached in this investigation may be
summarized by pointing out that IAA showed no effect on
growth rate at lower concentrations and was inhibitory at
higher ones, that GA3 showed no effect at lower concentrations,
stimulated growth at a higher concentration, and was inhibi-
tory at the highest concentration tested, and that kinetin
did not appear to affect Escherichia coli growth or morpho-
logy at all. GA^ alone did not seem to affect cell morphology,
while those concentrations of IAA which inhibited total
growth also caused the formation of larger than normal cells,
this latter phenomenon being a possible consequence of
increased cell wall elasticity. Disparate results have
been reported by earlier workers who have investigated the
general question of phytohormone effects on microbial cells.
While the present preliminary study provides more information
to add to that already accumulated, the underlying principles
involved in hormone-cell relationships are yet to be learned.

CHAPTER BIBLIOGRAPHY
1. Cleland, R., "A Separation of Auxin-induced Cell Wall Loosening Into Its Plastic and Elastic Components," Physiologia Plantarum, XI (1958), 599-609.
2. Conn, Eric E. and P. K. Stumpf, Outlines of Biochemistry, 2nd ed., New York, John Wiley and Sons, Inc., 1966.
3. Bubos, Rene J., "Inhibition of Bacterial Growth by-Auxins, " Proceedings of the Society for Experimental Biology, New York, LXIII (1946), 317-319.
4. Fukuyama, T. T. and H. S. Moyed, '"Inhibition of Cell Growth by Photooxidation Products of Indole-3-acetic Acid," Journal of Biological Chemistry, CCXXXIX (1964), 2392-2397.
5. Guttman, R., "Effects of Kinetin on Cell Division with Special Reference to Initiation and Duration of Mitosis," Chromosoma, VIII (1956), 341-350.
6. Haber, A. H. and H. J. Luippold, "Effects of Gibberellin on Gamma-irradiated wheat," American Journal of Botany, XLVII (1960), 140-144.
7. Hayashi, T., Y. Takijima and Y. Murakami, "The Bio-chemistry of Bakanae Fungus. 28. The Physiological Action of Gibberellin. IV, " Journal of the Agricul-tural Chemistry Society of Japan, XXVII (1927), 672-675.
8. Heyn, A. N. J., "Der Mechanismus der Zellstreckung," Rec. Tray. Bot. Neerl., XXVIII (1931), 113-244.
9. Leopold, A. Carl, Plant Growth and Development, New York, McGraw-Hill Book Cortipany, 1964.
10. Mothes, K., "Uber das Altern der Blatter und die Moglichkeit ihrer WiederverjungungV" Naturwissen-schaften, XLVII (1960), 337-350.
30

31
11. _ , L. Engelbrecht and 0. Kulajewa, ,rUber die Wirkung des Kinetins auf Stickstoffverteilung und Eiweisssynthese in isolierte Blattern. Flora (Jena), CXLVII (1959), 445-464.
12. Partheir, B. and R. Wollgiehn, "Uber den Einfluss des Kinetins auf den Eiweiss-und Nukleinsaure Stoffwecksel isolierten Tabakblattern," Ber. Deut. Botan. Ges., LXXIV (1961), 47-51.
13. Sachs, P. M., C. F. Bretz and A. Lang, "Short Histo-genesis: The Early Effects of Gibberellin Upon Stem Elongation in Two Rosette Plants," American Journal of Botany, XLVI (1959), 376-384.
14. Sawada, K. and E. Kurosawa, "On the Prevention of the Bakanae Disease of Rice," Experimental Station Bulletin of Formosa, XXI (1924), 1-19.
15. Still, Cecil C., T. T. Fukuyama and H. S. Moyed, "Inhibitory Oxidation .products of Indole-3-acetic Acid," Journal of Biological Chemistry, CCXL (1964), 2612-2618.
16. Tagawa, T. and J. Bonner, "Mechanical Properties of the Avena Coleoptile as Related to Auxin and to Ionic Interactions," Plant Physiology, XXXII (1957), 207-212.

BIBLIOGRAPHY
Books
Conn, Eric E. and P. K. Sturapf, Outlines of Biochemistry, 2nd ed., New York, John Wiley and Sons, Inc., 1966.
Fletcher, Win. W., "The Effect of Herbicides on Soil Micro-organisms, " The Physiology and Biochemistry of Herbicides, edited by L. J. Audus, London, Academic Press, 1964, pp . 2 0 - 6 2 .
Hillman, W. S. and W. K. Purves, "Does Gibberellin Act Through an Auxin-mediated Mechanism?," Plant Growth Regulation, edited by R. M. Klein, Ames, Iowa, Iowa State University Press, 1961, pp. 589-600.
Kato, J., "Physiological Action of Gibberellin with Special Reference to Auxin," Plant Growth Regulation, edited by R. M. Klein, Ames, Iowa, Iowa State University Press, 1961 , p p . 6 0 1 - 6 0 9 .
Leopold, A. Carl, Plant Growth and Development, New York, McGraw-Hill Book Company, 1964.
Articles
Ball, Earnest, "Heteroauxin and the Growth of Escherichia coli," Journal of Bacteriology, XXXVI (1938), 559-565.
Beckwith, T. D. and E. M. Geary, "Effect of Indole 3 Acetic Acid on Multiplication of Esch. coli and E. typhosa,,r
Brian, P. W., "Role of Gibberellin-like Hormones in Regulation of Plant Growth and Flowering," Nature CLXXXI (1958), 1122-1123.
Cleland, R., "A Separation of Auxin-induced Cell Wall Loosening Into Is Plastic and Elastic•Components," Physiologia Plantarum, XI (1958), 599-609.
32

33
Dubos, Rene J., "Inhibition of Bacterial Growth by Auxins," Proceedings of the Society for Experimental Biology, New York, LXIII (1946), 317-319.
Fukuyama, T. T. and H. S. Moyed, "Inhibition of Cell Growth by Photooxidation Products of Indole-3-acetic Acid," Journal of Biological Chemistry, CCXXXIX (1964), 2392-2397,
Galston, A. W. and H. Warburg, "An Analysis of Auxin-Gibberellin Interaction in Pea Stem Tissue," plant Physiology, XXXIV (1959), 16-22.
Guttman, R., "Effects of Kinetin on Cell Division with Special Reference to Initiation and Duration of Mitosis," Chromosoma, VIII (1956), 341-350.
Haber, A. H. and H. J. Luippold, "Effects of Gibberellin on Gamma-irradiated Wheat," American Journal of Botany, XLVII (1960), 140-144.
Handler, P. and H. Kamin, "Indole Acetic Acid and Growth of Bacteria with Varying Requirements for Nicotinic Acid and Tryptophan," Proceedings of the Society for Experimental Biology, New York, LXVI (1948), 251-254.
Hayashi, T., Y. Takijima and Y. Murakami, "The Bio-chemistry of Bakanae Fungus. 28. The Physiological Action of Gibberellin. IV," Journal of the Agricultural Chemistry Society of Japan, XXVII (1927), 672-675.
Heyn, A. N. J., "Der Mechanismus der Zellstreckung," Rec. Trav. Bot. Neerl., XXVIII (1931), 113-244.
Kato, J., "Studies on the Physiological Effect of Gibberellin II. On the Interaction of Gibberellin with Auxins and Growth Inhibitors," Physiologia Plantarum, XI (1958), 10-15.
Kefford, N. P., "Auxin-Gibberellin Interactions in Rice Coleoptile Elongation," Plant Physiology, XXXVII (1962), 380-386.
Kennell, D. E., "The Effects .of Indoleacetic Acid and Kinetin on the Growth of Some Microorganisms," Experi-mental Cell Research, XXI (1960), 19-33.

34
Kogl, F., A. J. Haagen-Smit and H. Erxleben, "Uber ein Neues Auxin ('Heteroauxin') aus Harn. XI.," Zeit-schrift Physiologisch Chemie, CCXXVIII (1934) 90-103.
Kogl, F. and D. G. F. R. Kostermans, "Hetero-auxin als Stoffwechselprodukt Niederer Prlanzlicher Organisraen. XIIi:.," Zeitschrift Physiologisch Chemie, CCXXVIII (1934), 113-121.
Kuraishi, S. and R. M. Muir, "Increase in Diffusible Auxin After Treatment with Gibberellin," Science, CXXXVII (1962), 760-761.
Kurosawa, E., "Experimental Studies on the Secretion of Fusarium heterosporum on Rice Plants," Transactions of the Natural History Society of Formosa, XVI (1926), 213-227.
Ma'ruzella, Jasper C. and James G. Garner, "Effect of Kinetin on Bacteria," Nature, CC (1963), 385.
Miller, C. O., F. Skoog, F. S. Okumura, M. H. von Saltza and F. M. Strong, "Isolation, Structure, and Synthesis of Kinetin, a Substance Promoting Cell Division," Journal of the American:Chemical Society, LXVIII (1956), 1375-1380.
"Structure and Synthesis of Kinetin," Journal of the American Chemical Society, LXXVII (1955), 2662-2663.
Miller, C. O., F. Skoog, M. H. von Saltza and F. M. Strong, "Kinetin: a Cell Division Factor from Deoxyribonucleic Acid," Journal of the American Chemical Society, LXVIII (1955), 1392.
Mothes, K., "Uber das Altern der Blatter und die Moglichkeit ihrer Wiederverjungung," Naturwissenschaften, XLVII (1960), 337-350.
_ , L. Engelbrecht and O. Kulajewa, "Uber die Wirkung des Kinetins auf Stickstoffverteilung und Eiweisssynthese in isolierte Blattern. Flora (Jena), CXLVII (1959), 445-464.

35
partheir, B. and R. Wollgiehn, "IJber den Einfluss des Kinetins auf den Eiweiss-und Nukleinsaure Stoffwecksel in isolierten Tabakblattern," Ber. Deut. Botan. Ges., LXXIV (1961), 47-51.
Phillips, I. D. J., A. J. Vlitos and H. Cutler, "The Influ-ence of Gibberellic Acid Upon the Endogenous Growth Substances of the Alaska Pea," Contributions to the Boyce Thompson Institute, XX (1959), 111-120.
Saono, Susono, "Effect of Gibberellic Acid on the Growth and Multiplication of Some Soil Microorganisms and Unicellular Green Algae," Nature, CCIV (1964), 1328-1329.
Sachs, F. M., C. F. Bretz and A. Lang, "Short Histogenesis: The Early Effects of Gibberellin Upon Stem Elongation in Two Rosette Plants," American Journal of Botany, XLVI (1959), 376-384.
Sawada, K. and E. Kurosawa, "On the Prevention of the Bakanae Disease of Rice," Experimental Station Bulletin of Formosa, XXI (1924), 1-19.
Scott, D. B. M. and E. Chu, "Synchronized Division of Growing Cultures of 12. coli," Experimental Cell Reaearch, XIV (1958), 166-174.
Still, Cecil C., T. T. Fukuyama and H. S. Moyed, "Inhibitory Oxidation Products of Indole-3-acetic Acid," Journal of Biological Chemistry, CCXL (1964), 2612-2618.
Tagawa, T. and J. Bonner, "Mechanical Properties of the Avena Coleoptile as Related to Auxin and to Ionic Interactions," Plant Physiology, XXXII (1957), 207-212.
Thimann, K. V., "On the Plant Growth Hormone Produced by Rhizopus suinus," Journal of Biological Chemistry, CIX (1935), 279-291.
Yabuta, T., "Biochemistry of the 'Bakanae' Fungus of Rice," Agriculture and Horticulture (Tokyo), X (1935), 17-22.
Yoda, S. and J. Ashida, "Effects of Gibberellin and Auxin on the Extensibility of the Pea Stem," Plant Cell Physiology (Tokyo), I (1960), 99-105.

36
Unpublished Materials
Skinner, C. G., personal correspondence, North Texas State University, Denton, Texas, July 31, 1967.