effects of cross-mating on susceptibility of synonymous ... · brugia malayi cross-mating...
TRANSCRIPT

Contents lists available at ScienceDirect
Acta Tropica
journal homepage: www.elsevier.com/locate/actatropica
Effects of cross-mating on susceptibility of synonymous mosquitoes,Anopheles paraliae and Anopheles lesteri to infection with nocturnallysubperiodic Brugia malayi
Watcharatip Dedkhada, Lyric C. Bartholomayb, Bruce M. Christensenb, Deepak Joshic,Kritsana Taaid, Chayanit Hempolchoma, Atiporn Saeunge,⁎
aGraduate PhD’s Degree Program in Parasitology, Faculty of Medicine, Chiang Mai University, Chiang Mai, 50200, ThailandbDepartment of Pathobiological Sciences, University of Wisconsin, 1656 Linden Drive, Madison, WI, 53706, USAc Department of Entomology, University of Kentucky, Lexington, KY, 40546-0091, USAd Faculty of Veterinary Medicine, Western University, Kanchanaburi, 71170, Thailande Department of Parasitology, Faculty of Medicine, Chiang Mai University, Chiang Mai, 50200, Thailand
A R T I C L E I N F O
Keywords:Anopheles lesteriAnopheles paraliaeBrugia malayiCross-mating experimentintrogressionmelanization
A B S T R A C T
In Southeast Asia, Anopheles lesteri (recently synonymized with An. paraliae) is a competent vector forPlasmodium parasites, but its ability to transmit parasites that cause lymphatic filariasis has yet to be determined.In this study, the susceptibility of An. lesteri and An. paraliae to Brugia malayi parasites was determined bycomparing with the control mosquito, Aedes togoi. We found that the infection prevalence per infected mosquitoin An. paraliae was significantly lower than that in Ae. togoi in all experiments (p < 0.05). Reciprocal crosses(female An. paraliae x male An. lesteri) produced highly susceptible F1-hybrid progeny, with increased infectionprevalence when compared to parental stocks (p < 0.05). Subsequently, the possibilities of introgression be-tween high and low/moderate parasite susceptibility genes were investigated by cross-mating experiments(parental, reciprocal crosses, back crosses and repeated backcrosses). The results showed the possibility of in-trogression of B. malayi-susceptible genes between An. paraliae (low/moderate susceptibility) and An. lesteri(high susceptibility) based on increasing or decreasing susceptibility and normal larval development in thethoracic muscles of F3-hybrids. Additionally, melanization, an innate immune response with proven involvementin the susceptibility or refractoriness of mosquitoes to B. malayi parasites, was examined. Parasite degenerationand cell aggregation, and melanization were observed for first-stage larvae in the thoracic muscle fibers ofhybrid mosquitoes.
1. Introduction
Lymphatic filariasis (LF) is a Neglected Tropical Disease (NTD).Currently, 856 million people in 52 countries are threatened by LF andrequire preventive chemotherapy. An estimated 120 million people intropical and subtropical areas of the world are infected with this disease(WHO, 2017). Chronic filarial worm infections lead to a severe damageof the lymphatic system, and patients suffer from a permanent disabilityand psychosocial impacts of associated stigmatization (WHO, 2017).The Global Programme to Eliminate Lymphatic Filariasis (GPELF) wasestablished to improve the quality of life by stopping the spread ofinfection through large-scale annual treatment of the population in anarea or region where infection is present, and to alleviate suffering
caused by LF through increased morbidity management and disabilityprevention activities (WHO, 2017). In addition to Mass Drug Admin-istration (MDA), vector control should be implemented to reduce thetransmission in both areas where the disease is prevalent and eradi-cated. As we move towards the endpoint goals set by GPELF, under-standing vector-parasite interactions is increasingly becoming im-portant (Gleave et al., 2016). In Thailand, LF is caused by both W.bancrofti and B. malayi (Saeung et al., 2013). After the infection pre-valence (microfilaria prevalence) in the country decreased from 0.77%in 2003 to 0.09% in 2010 (WHO, 2013), the target set by WHO was met(less than 0.1%) and MDA was stopped. However, no post-MDA pre-valence studies on the local population have been reported in thecountry (Dickson et al., 2017).
https://doi.org/10.1016/j.actatropica.2018.07.020Received 7 June 2018; Received in revised form 19 July 2018; Accepted 24 July 2018
⁎ Corresponding author.E-mail addresses: [email protected] (W. Dedkhad), [email protected] (L.C. Bartholomay), [email protected] (B.M. Christensen),
[email protected] (D. Joshi), [email protected] (K. Taai), [email protected] (C. Hempolchom), [email protected] (A. Saeung).
Acta Tropica 187 (2018) 65–71
Available online 25 July 20180001-706X/ © 2018 Elsevier B.V. All rights reserved.
T

Within the mosquito vector, some organs and tissues serve as bar-riers to further development of filarial parasites, i.e., cibarial andpharyngeal armatures (foregut) by physical killing of parasites in theforegut, peritrophic matrix formation and blood clotting in the midgut,and hemolymph factors that kill the parasite in the thoracic muscu-lature, and lead to melanotic encapsulation (Bartholomay, 2014). Thesebarriers affect compatibility of the vector-pathogen association (vectorcompetence) (Beerntsen et al., 2000; Bartholomay and Christensen,2002). Genetic factors, primarily controlled by a sex-linked recessivegene, also influence vector competence in various mosquito strains andspecies (Macdonald, 1962; Zielke, 1973; Coluzzi and Cancrini, 1974;McGreevy et al., 1974; Sulaiman and Townson, 1980; Severson et al.,1994).
Anopheles (Anopheles) in the Hyrcanus Group includes at least 27species (Harbach, 2018). Studies based on mosquitoes collected fromthe wild as well as laboratory experiments have shown that somemembers of this group can hybridize (Choochote et al., 2014) and passon parasite susceptibility genes/traits (Saeung et al., 2014). Anopheleslesteri is a highly competent malaria vector (Joshi et al., 2009). Thereare some degree of confusion in the morphological classifications of An.lesteri (Korea strain) and Anopheles paraliae (Thailand strain). Recently,Taai et al. (2013) were able to confirm that these two are synonymousspecies by performing cross-mating experiments between An. paraliae(Thailand strain) and An. lesteri (Korea strain) and comparing sequencefor the Internal Transcribed Spacer (ITS2) and cytochrome c oxidasesubunits I and II. After the recent resurgence of malaria in the Asiancontinent, studies of medically important mosquito vector species havegained renewed attention. Efforts have been paid to characterize thevector competence of the indigenous species to malaria; however, veryfew or no reports exist regarding their ability to transmit LF parasites.Therefore, the current study aims to compare susceptibility rate of An.lesteri and An. paraliae to infection with B. malayi and to determine thepossible occurrence of the introgression phenomenon when repeatedbackcrosses between these two synonymous species were carried out ina laboratory setting.
2. Materials and methods
2.1. Mosquito species
Anopheles lesteri (AL): wild-caught, fully engorged females wereoriginally collected from So-Rae District, Incheon City in South Korea.Exact species identification was based on a combination of morpholo-gical characters and a DNA-based assay described earlier (Joshi et al.,2010).
Anopheles paraliae (AP): wild-caught, fully engorged females An.paraliae were collected by using cow-baited traps from three differentlocalities in Thailand, including Ratchaburi, Nakhon Si Thammarat andSongkhla Provinces (Taai et al., 2013). Species identification was basedon a combination of morphological characters, metaphase chromosomeanalysis and DNA based assays (Rattanarithikul et al., 2006; Taai et al.,2013; Saeung et al., 2014).
Aedes togoi (AT) were used as a positive control for parasite devel-opment. This colony was established using adult female mosquitoescollected from Chanthaburi Province. This autogenous strain of Ae.togoi is a proven efficient laboratory vector for a wide range of generaand species of filarial nematodes, including the nocturnally subperiodicB. malayi (Saeung and Choochote 2013; Saeung et al., 2013, 2014).
The mosquito colonies were established successfully for many con-secutive generations in the insectary of the Department of Parasitology,Faculty of Medicine, Chiang Mai University, Thailand, at 27 ± 2 °C,70-80% relative humidity, using a 12:12 day night ratio adjusted withfluorescent lighting and natural light coming from the windows(Choochote and Saeung, 2013).
2.2. Cross-mating experiments
Cross-mating experiments between An. lesteri and An. paraliae wereperformed as shown in Table 1. Virgin females and males were usedfollowing the techniques previously described by Saeung et al. (2014).
2.3. Blood containing nocturnally subperiodic (NSP) B. malayimicrofilariae
The B. malayi used in these studies originated from a patient whowas a resident of Narathiwat Province, southern Thailand. Maintenanceof the parasites was transferred to Mongolian jirds (Meriones un-guiculatus) and ever since has been maintained at the Faculty ofMedicine, Chiang Mai University, Chiang Mai, Thailand (Choochoteet al., 1986). Preparations of blood containing B. malayi microfilariaeare described elsewhere (Saeung et al., 2013). Briefly, the jirds wereinoculated intraperitoneally with infective larvae of B. malayi andmaintained for at least 3 to 5 months before microfilariae were col-lected by injecting 3ml of Hank’s Balanced Salt Solution (HBSS, pH 7.2-7.4) into the peritoneal cavity and then withdrawing by peritonealwashing. Then, 0.05ml of enriched microfilariae was mixed with 5mlof human heparinized blood, which had been taken from principle in-vestigator (10 units of heparin/ml of blood) and microfilarial densitywas adjusted to approximately 200-350 microfilariae (mf)/20 μl priorto feeding mosquitoes. The reason for using a microfilarial densityranging from about 200 to 300 mf per 20 μl was based on previouslyproven experiments that yielded satisfactorily susceptible Ae. togoi toNSP B. malayi (susceptibility rate= 80-95%) (Saeung and Choochote,2013; Jariyapan et al., 2013; Saeung et al., 2013).
2.4. Infection of mosquitoes with B. malayi microfilariae
Five-day-old adult female Ae. togoi, An. paraliae, An. lesteri, hybridprogeny derived from parental, reciprocal, back crosses and repeatedbackcrosses, were starved for 24 hours then provided simultaneouslywith blood-containing B. malayi microfilariae (mf density= 210, 275,300 mf/20 μl in experiment 1, 2 and 3, respectively) through an arti-ficial membrane (Choochote and Saeung, 2013). Subsequently, theblood-fed mosquitoes were separated from the unfed mosquitoes afterexposure. Fourteen days after feeding, all infected mosquitoes weredissected in normal saline solution. The number of mosquitoes with atleast one third-stage larva (L3) in any part of the body (head, thorax orabdomen) (20 female mosquitoes/species) were recorded for de-termining the prevalence of infection and mean intensity (total numberof recovered parasites divided by total number of infected mosquitoes).
2.5. Determination of the possible factor(s) influencing the degree ofsusceptibility
Ten midguts of blood-fed An. paraliae and An. lesteri, and their F1-F3-hybrids were extracted immediately after full engorgement. Theingested blood meal was pulled out with the gut and thick smears wereprepared and stained with Giemsa (pH 7.2). Microscopic examinationwas done to quantify the number of mf ingested using a compound
Table 1Cross-mating experiments between An. paraliae (AP) and An. lesteri (AL) andtheir progenies.
Experiments Female Male Progenies
Reciprocal crosses AP AL F1-hybridsAL AP F1-hybrids
Back crosses (AP x AL)F1 AL F2-hybrids(AL x AP)F1 AP F2-hybrids
Repeated backcrosses (AP x AL)F2 x AL AL F3-hybrids(AL x AP)F2 x AP AP F3-hybrids
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
66

microscope (BX53, Olympus, Japan). Four days after feeding, 10thoraces of each cohort were torn with dissecting tools in a drop ofnormal saline solution. First stage larvae (L1) were counted and scoredas “normal” if alive with intact morphology, as “melanized” if there wasevidence of melanin associated with the worm, as “degenerated” if theydemonstrated vacuolated internal organs without any evidence ofmelanization, and as “cell aggregated” if they had one or more cellsattached to the surface.
2.6. Data analysis
Chi-square test was used to compare the infection prevalence.Student’s t-test was performed for comparisons of mean intensity. Thelevel of significance was set at 5% (p-value< 0.05). All data wereanalyzed using IBM SPSS statistics, version 24 for Windows (Chicago,SPSS Inc.).
2.7. Ethical clearance
The protocols used in these studies were approved by the AnimalEthics Committee (PAR-2558-03170, No. 05/2015), Faculty ofMedicine, Chiang Mai University, Chiang Mai, Thailand.
3. Results
3.1. Susceptibility level of hybrid progenies to infection with NSP B. malayi
Overall, comparative statistical analyses of the infection prevalencewere carried out between Ae. togoi and An. paraliae and An. lesteri. Theresults revealed that the infection rates of Ae. togoi and An. paraliaewere significantly different (Chi-square test, p<0.05), whereas theydid not differ significantly between Ae. togoi and An. lesteri (Chi-squaretest, p>0.05) in all three experiments (Table 2). In addition, meanintensity between Ae. togoi and An. paraliae or An. lesteri as well as
parental (An. paraliae and An. lesteri) and their F1 to F3-hybrids progenywere not different (Student´s t-test, p>0.05) in all three experiments(Table 2).
In experiment I, the results of reciprocal crosses [(An. paraliae x An.lesteri)F1 and (An. lesteri x An. paraliae)F1] demonstrated that the in-fection prevalence and mean intensity of F1-hybrids progeny was 65%and 4.69 and 80% and 10.44, respectively. Comparisons of infectionprevalence between parental and offspring progeny were significantlydifferent (Chi-square test, p=0.026) only between An. paraliae and(An. paraliae x An. lesteri)F1 (Table 2).
In experiment II, the results of back crosses [(An. paraliae x An.lesteri)F1 x An. lesteri and (An. lesteri x An. paraliae)F1 x An. paraliae]showed that the infection prevalence and mean intensity of F2-hybridsprogeny was 60% and 7.33 and 75% and 5.87, respectively.Comparisons of infection prevalence between parental and offspringprogenies were not statistically different (Chi-square test, p>0.05) forboth directions of F2-hybrids (Table 2).
In experiment III, the results of repeated backcrosses [(An. paraliae xAn. lesteri)F2 x An. lesteri and (An. lesteri x An. paraliae)F2 x An. paraliae]showed that the infection prevalence and mean intensity of F3-hybridsprogeny was 65% and 9.85 and 45% and 5.33, respectively. Infectionprevalence between parental and offspring progeny was not statisticallydifferent (Chi-square test, p>0.05) for either direction of the F3-hybridprogeny (Table 2).
3.2. The possible factor(s) influencing the degree of susceptibility
The average number of mf per infected midgut (Table 3) indicatedthat all of the mosquito species were successful in obtaining a con-siderable number of mf from infected blood. Some of these parasitessuccessfully penetrated the thoracic muscle cells and differentiated intoL1 intracellularly in all three species. The recovered normal and ab-normal L1 larvae (dead larvae) from thoraces are shown in Figs. 1A-F.Normal larvae had a sausage-like morphology, slow movement and
Table 2Infection prevalence and mean intensity in parental, reciprocal, back cross and repeated backcross progenies of An. paraliae (AP) and An. lesteri (AL) compared withcontrol, Ae. togoi (AT), after feeding on blood containing B. malayi microfilariae, with all mosquitoes dissected 14 days after feeding.
Mosquito species (Female x male)* Infection prevalence(No.)**
Mean intensity(range)+
L3- distribution
% head (No.) % thorax (No.) % abdomen (No.)
Experiment IAT 80% (16/20) 6.13 (1-20) 83.68 (82) 11.22 (11) 5.10 (5)ParentalAP 25% (5/20)a 4.8 (1-8)a 33.33 (8) 29.17 (7) 37.50 (9)AL 80% (16/20)b 7.56 (1-14)b 71.07 (86) 19.83 (24) 9.10 (11)Reciprocal crosses(AP x AL)F1 65% (13/20)g 4.69 (1-11)g 27.87 (17) 72.13 (44) 0 (0)(AL x AP)F1 80% (16/20)h 10.44 (1-18)h 41.32 (69) 42.51 (71) 16.17 (27)Experiment IIAT 85% (17/20) 4.41 (1-13) 92.00 (69) 2.67 (2) 5.33 (4)ParentalAP 50% (10-20)c 3.00 (1-5)c 60.00 (18) 33.33 (10) 6.67 (5)AL 85% (17/20)d 3.59 (1-5)d 75.41 (46) 13.11 (8) 11.48 (7)Back crosses(AP x AL)F1 x AL 60% (12-20)i 7.33 (1-18)i 73.86 (65) 20.46 (18) 5.68 (5)(AL x AP)F1 x AP 75% (15/20)j 5.87 (1-12)j 73.86 (65) 12.50 (11) 13.64 (12)
Experiment IIIAT 80% (16/20) 7.19 (1-20) 75.65 (87) 14.78 (17) 9.57 (11)ParentalAP 30% (6/20)e 4.17 (1-9)e 40.00 (10) 44.00 (11) 16.00 (4)AL 70% (14/20)f 13.86 (1-29)f 70.62 (137) 17.01 (33) 12.37 (24)Repeated backcrosses(AP x AL)F2 x AL 65% (13/20)k 9.85 (1-13)k 64.06 (82) 23.44 (30) 12.50 (16)(AL x AP)F2 x AP 45% (9/20)l 5.33 (1-8)l 68.75 (33) 27.08 (130) 4.17 (2)
*Dissected from 20 parental or hybrid progeny of each cohort.**Chi-square test: a, c, e vs. control (AT), p < 0.05; b, d, f, vs. control (AT), p > 0.05; g vs. parental (AP), p < 0.05; h, i, j, k, l vs. parental (AP or AL), p > 0.05.+ Student´s t-test (two-sided): a, b, c, d, e, f vs. control (AT), p > 0.05; g, h, i, j, k, l vs. parental (AP or AL), p > 0.05.
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
67
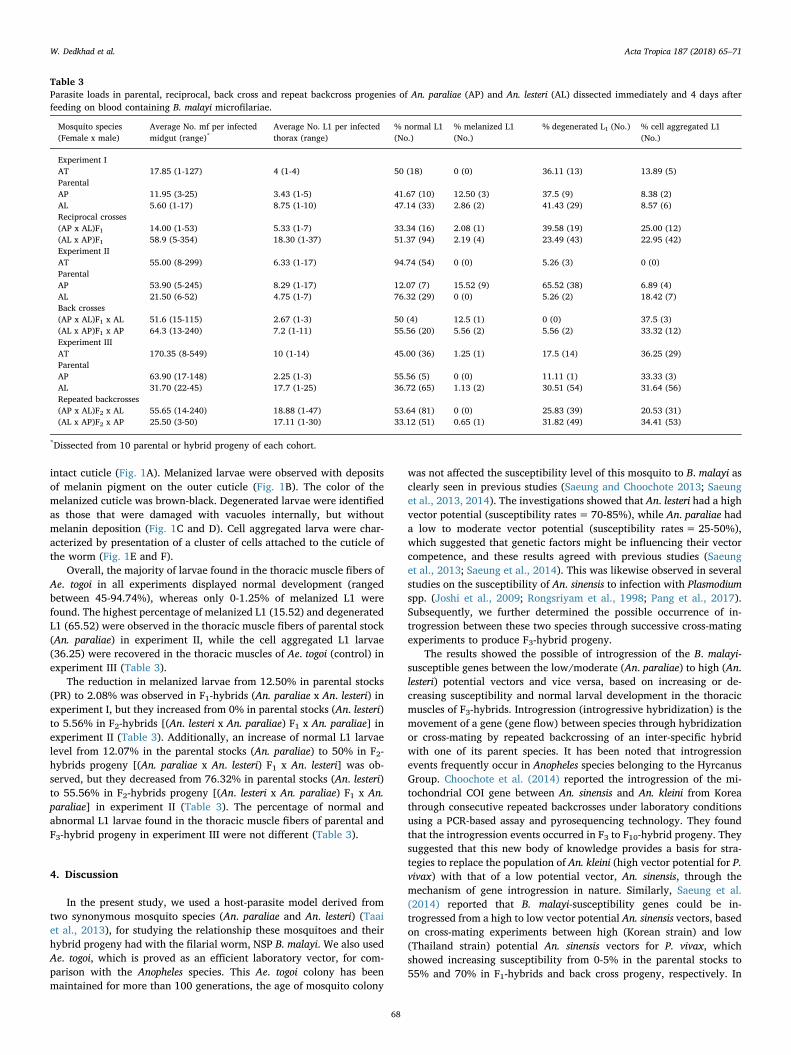
intact cuticle (Fig. 1A). Melanized larvae were observed with depositsof melanin pigment on the outer cuticle (Fig. 1B). The color of themelanized cuticle was brown-black. Degenerated larvae were identifiedas those that were damaged with vacuoles internally, but withoutmelanin deposition (Fig. 1C and D). Cell aggregated larva were char-acterized by presentation of a cluster of cells attached to the cuticle ofthe worm (Fig. 1E and F).
Overall, the majority of larvae found in the thoracic muscle fibers ofAe. togoi in all experiments displayed normal development (rangedbetween 45-94.74%), whereas only 0-1.25% of melanized L1 werefound. The highest percentage of melanized L1 (15.52) and degeneratedL1 (65.52) were observed in the thoracic muscle fibers of parental stock(An. paraliae) in experiment II, while the cell aggregated L1 larvae(36.25) were recovered in the thoracic muscles of Ae. togoi (control) inexperiment III (Table 3).
The reduction in melanized larvae from 12.50% in parental stocks(PR) to 2.08% was observed in F1-hybrids (An. paraliae x An. lesteri) inexperiment I, but they increased from 0% in parental stocks (An. lesteri)to 5.56% in F2-hybrids [(An. lesteri x An. paraliae) F1 x An. paraliae] inexperiment II (Table 3). Additionally, an increase of normal L1 larvaelevel from 12.07% in the parental stocks (An. paraliae) to 50% in F2-hybrids progeny [(An. paraliae x An. lesteri) F1 x An. lesteri] was ob-served, but they decreased from 76.32% in parental stocks (An. lesteri)to 55.56% in F2-hybrids progeny [(An. lesteri x An. paraliae) F1 x An.paraliae] in experiment II (Table 3). The percentage of normal andabnormal L1 larvae found in the thoracic muscle fibers of parental andF3-hybrid progeny in experiment III were not different (Table 3).
4. Discussion
In the present study, we used a host-parasite model derived fromtwo synonymous mosquito species (An. paraliae and An. lesteri) (Taaiet al., 2013), for studying the relationship these mosquitoes and theirhybrid progeny had with the filarial worm, NSP B. malayi. We also usedAe. togoi, which is proved as an efficient laboratory vector, for com-parison with the Anopheles species. This Ae. togoi colony has beenmaintained for more than 100 generations, the age of mosquito colony
was not affected the susceptibility level of this mosquito to B. malayi asclearly seen in previous studies (Saeung and Choochote 2013; Saeunget al., 2013, 2014). The investigations showed that An. lesteri had a highvector potential (susceptibility rates= 70-85%), while An. paraliae hada low to moderate vector potential (susceptibility rates= 25-50%),which suggested that genetic factors might be influencing their vectorcompetence, and these results agreed with previous studies (Saeunget al., 2013; Saeung et al., 2014). This was likewise observed in severalstudies on the susceptibility of An. sinensis to infection with Plasmodiumspp. (Joshi et al., 2009; Rongsriyam et al., 1998; Pang et al., 2017).Subsequently, we further determined the possible occurrence of in-trogression between these two species through successive cross-matingexperiments to produce F3-hybrid progeny.
The results showed the possible of introgression of the B. malayi-susceptible genes between the low/moderate (An. paraliae) to high (An.lesteri) potential vectors and vice versa, based on increasing or de-creasing susceptibility and normal larval development in the thoracicmuscles of F3-hybrids. Introgression (introgressive hybridization) is themovement of a gene (gene flow) between species through hybridizationor cross-mating by repeated backcrossing of an inter-specific hybridwith one of its parent species. It has been noted that introgressionevents frequently occur in Anopheles species belonging to the HyrcanusGroup. Choochote et al. (2014) reported the introgression of the mi-tochondrial COI gene between An. sinensis and An. kleini from Koreathrough consecutive repeated backcrosses under laboratory conditionsusing a PCR-based assay and pyrosequencing technology. They foundthat the introgression events occurred in F3 to F10-hybrid progeny. Theysuggested that this new body of knowledge provides a basis for stra-tegies to replace the population of An. kleini (high vector potential for P.vivax) with that of a low potential vector, An. sinensis, through themechanism of gene introgression in nature. Similarly, Saeung et al.(2014) reported that B. malayi-susceptibility genes could be in-trogressed from a high to low vector potential An. sinensis vectors, basedon cross-mating experiments between high (Korean strain) and low(Thailand strain) potential An. sinensis vectors for P. vivax, whichshowed increasing susceptibility from 0-5% in the parental stocks to55% and 70% in F1-hybrids and back cross progeny, respectively. In
Table 3Parasite loads in parental, reciprocal, back cross and repeat backcross progenies of An. paraliae (AP) and An. lesteri (AL) dissected immediately and 4 days afterfeeding on blood containing B. malayi microfilariae.
Mosquito species(Female x male)
Average No. mf per infectedmidgut (range)*
Average No. L1 per infectedthorax (range)
% normal L1(No.)
% melanized L1(No.)
% degenerated L1 (No.) % cell aggregated L1(No.)
Experiment IAT 17.85 (1-127) 4 (1-4) 50 (18) 0 (0) 36.11 (13) 13.89 (5)ParentalAP 11.95 (3-25) 3.43 (1-5) 41.67 (10) 12.50 (3) 37.5 (9) 8.38 (2)AL 5.60 (1-17) 8.75 (1-10) 47.14 (33) 2.86 (2) 41.43 (29) 8.57 (6)Reciprocal crosses(AP x AL)F1 14.00 (1-53) 5.33 (1-7) 33.34 (16) 2.08 (1) 39.58 (19) 25.00 (12)(AL x AP)F1 58.9 (5-354) 18.30 (1-37) 51.37 (94) 2.19 (4) 23.49 (43) 22.95 (42)Experiment IIAT 55.00 (8-299) 6.33 (1-17) 94.74 (54) 0 (0) 5.26 (3) 0 (0)ParentalAP 53.90 (5-245) 8.29 (1-17) 12.07 (7) 15.52 (9) 65.52 (38) 6.89 (4)AL 21.50 (6-52) 4.75 (1-7) 76.32 (29) 0 (0) 5.26 (2) 18.42 (7)Back crosses(AP x AL)F1 x AL 51.6 (15-115) 2.67 (1-3) 50 (4) 12.5 (1) 0 (0) 37.5 (3)(AL x AP)F1 x AP 64.3 (13-240) 7.2 (1-11) 55.56 (20) 5.56 (2) 5.56 (2) 33.32 (12)Experiment IIIAT 170.35 (8-549) 10 (1-14) 45.00 (36) 1.25 (1) 17.5 (14) 36.25 (29)ParentalAP 63.90 (17-148) 2.25 (1-3) 55.56 (5) 0 (0) 11.11 (1) 33.33 (3)AL 31.70 (22-45) 17.7 (1-25) 36.72 (65) 1.13 (2) 30.51 (54) 31.64 (56)Repeated backcrosses(AP x AL)F2 x AL 55.65 (14-240) 18.88 (1-47) 53.64 (81) 0 (0) 25.83 (39) 20.53 (31)(AL x AP)F2 x AP 25.50 (3-50) 17.11 (1-30) 33.12 (51) 0.65 (1) 31.82 (49) 34.41 (53)
*Dissected from 10 parental or hybrid progeny of each cohort.
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
68

contrast, the studies of the Aedes scutellaris complex to infection with B.pahangi using reciprocal and back crosses between susceptible Aedespolynesiensis and refractory Aedes malayensis, produced refractory hy-brid females which indicated that refractoriness may be dominant tosusceptibility (Macdonald, 1976). The genetic factors that influencevector competence of Ae. aegypti for B. malayi was first reported byMacdonald (1962). From cross-mating experiments between susceptibleand refractory strains, results demonstrated that susceptibility in the
thoracic muscles of Ae. aegypti is controlled by a sex-linked recessivegene (designated fm: filarial susceptibility, B. malayi). The gene fm alsocontrolled the susceptibility to periodic and subperiodic strains of B.malayi, B. pahangi and W. bancrofti (Macdonald and Ramachandran,1965). Trpis et al. (1981) demonstrated another mode of inheritance ofmosquito susceptibility to filarial parasites. They carried out crossingexperiments between susceptible and refractory species of the Ae. scu-tellaris complex, and the hybrid and backcross progeny were tested for
Fig. 1. First-stage larvae recovered from thoracic muscle fibers of Ae. togoi, An. lesteri and An. paraliae at 4 days post infections. (A) Normal live larva with intactcuticle and internal organs recovered from Ae. togoi. (B) Complete melanized larva obtained from An. paraliae. (C and D) Degenerated and vacuolated internal organslarvae recovered from An. paraliae and An. lesteri, respectively. (E and F) Cell aggregated larvae recovered from An. paraliae and An. lesteri, respectively. Scale bar:20 μm.
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
69

susceptibility to infection by B. malayi and B. pahangi. The results in-dicated that susceptibility was controlled by a non-Mendelian patternindicative of extrachromosomal factors inherited through the maternalparent. The use of molecular genetic linkage maps showed that thereare two quantitative trait loci (QTL) involved in B. malayi susceptibility.A quantitative trait locus on chromosome 1 (fsb[1, LF178] or fm locus)is a function that allows the worm to develop in the developmental site,and that on chromosome 2 (fsb[2, LF98]) is a function that expressessusceptibility of the phenotype (Severson et al., 1994). Although the fm
locus (LF178) is a conserved synteny gene between Anophelinae andCulicinae mosquitoes (Severson et al., 2004), it does not appear tofunction in an identical way in Anopheline mosquitoes, based on thedata presented here for hybrid progeny of An. paraliae and An. lesteri.Without knowing the specific gene or genes that drive this susceptibilitytrait in Anopheles species, it is not currently possible to specifically trackgene flow and introgression events in the hybrid progeny that wereproduced in our experiments. Thus, the combinations of phenotypicinformation obtained from this study with genomic evidence of geneflow are necessary to definitively prove the occurrence of introgression.Overall, our results indicate that the genetic control of susceptibility toB. malayi is complex and likely not identical between the subfamilies ofmosquitoes. Anopheles paraliae and An. lesteri are an interesting andviable model for understanding the relationship of B. malayi and ano-pheline mosquito hosts.
Phenotypically, susceptibility depends at least in part on the im-mune status of the mosquito. The first-line of defense against parasitesbegins with cells and/or cell products that are involved with externalcuticle, integument, intestinal mucosa and tracheal lining (Christensenet al., 2005). Organisms that breach these barriers and invade the hosthemocoel encounter constitutive and inducible defenses, includingphagocytosis and encapsulation by hemocytes, and humoral immunefactors produced by hemocytes, pericardial cells and fat body(Christensen et al., 2005; Hillyer, 2010; Choi et al., 2014). Melanizationis an important component of the cell-mediated immune response inmosquitoes (Christensen et al., 2005; Bartholomay, 2014). It involves acomplex series of reactions requiring tyrosine precursors and phenoloxidases to produce a capsule, consisting of melanin polymers cross-linked with proteins that sequester the filarial worm. In this study, wealso investigated the possible factor(s) influencing the degree of sus-ceptibility of these synonymous species to NSP B. malayi. Normal andabnormal L1 larvae from the thoracic muscle fibers of Ae. togoi, An.paraliae and An. lesteri were observed and counted at day 4 post in-fection. The results demonstrated that the percentage of normal andabnormal L1 larvae found in the thoracic muscle fibers of Ae. togoi wereconsistent with those of a previous study (Saeung et al., 2013). Thepercentage of melanized L1 found in the thoracic muscle fibers of Ae.togoi and An. lesteri were lower than those in An. paraliae, whereas thehigher rates of degenerated L1 and cell aggregated L1 were observed inthe two former species. We found that two defense reactions, directtoxicity (degeneration and cell aggregation) and melanization re-sponses were involved in the inhibition of L1 development in thoracicmuscle fibers. Indeed, it has been proposed that these responses may bedue to a physiological incompatibility. The larvae may simply die be-cause of an inappropriate environment for development and then theyare melanized and/or begin to degrade. Likewise, Gad et al. (1996)compared the defense reactions of the susceptible Culex pipiens and therefractory Aedes caspius against W. bancrofti. They found the vacuolatedlarvae in the thoracic muscle fibers of Ae. caspius, suggesting that thebarrier to parasite development is physiological in nature. In this study,the melanization responses to filarial worms are still unclear. We pro-posed that the process might be initiated by the adherance of hemocytesto the cuticle of L1 larvae, lysed hemocytes produce some factors (e.g.phenoloxidase, dopachrome conversion enzyme, C-type lectins)(Hillyer and Strand, 2014) at or near the surface of the parasite beforedeposition of melanin pigment to form a capsule. Surprisingly, the analpores of larvae were the most preferentially area in which these cells
attached firstly, and this observation agrees with the previous study byChristensen (1981). However, detailed ultrastructure by the aid ofelectron microscopy (SEM and TEM) of these cells must be performedbefore definite conclusions can be drawn about melanization. For de-generation processes, we assumed that the lytic factors, such as cyto-toxic reactive intermediates of oxygen (ROI) and nitrogen (RNI) andantimicrobial peptides, which can be generated by hemocytes duringmelanogenesis, might be a cause of vacuolation (Christensen et al.,2005; Hillyer and Strand, 2014). This process might be initiated by theaggregation of hemocytes, which then produced lytic factors that di-rectly kill these invading larvae. Furthermore, unknown factors mightcause tissue damage and death to developing of B. malayi in the thor-acic muscle fibers of these mosquitoes (Bartholomay, 2014).
5. Conclusions
The results demonstrated that An. lesteri had a higher susceptibilityto infection with NSP B. malayi than its synonymous species, An.paraliae. The introgression of the B. malayi-susceptible genes betweenthe low/moderate (An. paraliae) to high (An. lesteri) vector potentialvectors occurred after repeated backcrosses in the laboratory. Twopotential immune responses against filarial worm, direct toxicity andmelanization, by three mosquitoes species were observed in this study.This interesting host-parasite system provides a new body of knowledgeas well as a good model system for future investigations into the factorscontrolling susceptibility and refractoriness of mosquito vectors for fi-larial worms. However, the exact mechanism responsible for the in-hibition of development of these parasites and genetic evidence of geneflow has yet to be elucidated.
Disclosure of interest
The authors declare that they have no conflicts of interest con-cerning this article.
Acknowledgements
We are grateful to Professor Gi-Sik Min, Department of BiologicalSciences, Inha University, South Korea, who kindly provided the isolinecolony of An. lesteri used in this study. This research was supported bygrants from the Royal Golden Jubilee (RGJ) Ph.D. Program, ThailandResearch Fund (TRF) (grant number PHD/0066/2555 to A. Saeung andW. Dedkhad), the Diamond Research Grant (PAR-2560-04663, Contractnumber 120/2560) and the Faculty of Medicine Research Fund (PAR-2558-03170, contract number 011/2559) from the Faculty of Medicine,Chiang Mai University, and the Bio & Medical Technology DevelopmentProgram of the National Research Foundation (NRF) funded by theMinistry of Science & ICT, South Korea (grant number:2017M3A9E4070707) to A. Saeung.
References
Bartholomay, L.C., 2014. Infection barriers and responses in mosquito-filarial worm in-teractions. Current Opinion in Insect Science. 3, 37–42.
Bartholomay, L.C., Christensen, B.M., 2002. Vector-parasite interactions in mosquitoe-borne filariasis. In: In: Klei, T.R., Rajan, T.V., Black, S.J., Seed, J.R. (Eds.), WorldClass Parasites, vol. 5. The Filaria Kluwer Academic Publishers, pp. 9–19.
Beerntsen, B.T., James, A.A., Christensen, B.M., 2000. Genetics of mosquito vectorcompetence. Microbiol. Mol. Biol. Rev. 64, 115–137.
Choi, Y.J., Aliota, M.T., Mayhew, G.F., Erickson, S.M., Christensen, B.M., 2014. DualRNA-seq of parasite and host reveals gene expression dynamics during filarial worm-mosquito interactions. PLoS Negl. Trop. Dis. 8, e2905.
Choochote, W., Min, G.S., Intapan, P.M., Tantrawatpan, C., Saeung, A., Lulitanond, V.,2014. Evidence to support natural hybridization between Anopheles sinensis andAnopheles kleini (Diptera: Culicidae): possibly a significant mechanism for gene in-trogression in sympatric populations. Parasit Vectors. 7, 36.
Choochote, W., Saeung, A., 2013. Systematic techniques for the recognition of Anophelesspecies complexes. In: Manguin, S. (Ed.), Anopheles mosquitoes-New insights intomalaria vectors. In Tech., pp. 57–79.
Choochote, W., Sukhavat, K., Somboon, P., Khamboonruang, C., Maleewong, W.,
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
70

Suwanpanit, P., 1986. The susceptibility of small laboratory animals to nocturnallysuperiodic Brugia malayi in Thailand. J Parasitol Trop Med Assoc Thailand. 9, 35–37.
Christensen, B.M., 1981. Observations on the immune response of Aedes trivittatus againstDirofilaria immitis. Trans R Soc Trop Med Hyg. 75, 439–443.
Christensen, B.M., Li, J., Chen, C.C., Nappi, A.J., 2005. Melanization immune responses inmosquito vectors. Trends Parasitol. 21, 192–199.
Coluzzi, M., Cancrini, C., 1974. Genetica della suscettibilità di Aedes aegypti a Dirofilariarepens. Parassitologia. 16, 239–256.
Dickson, F.B., Graves, M.P., McBride, J.W., 2017. Lymphatic Filariasis in MainlandSoutheast Asia: a systematic review and meta-analysis of prevalence and diseaseburden. Trop. Med. Infect. 2, 32.
Gad, A.M., Farid, H.A., Hammad, R.E., Hussein, M.A., Kaschef, A.H., 1996. Host-parasiterelationships of Wuchereria bancrofti and mosquito hosts, Culex pipiens L. and Aedescaspius pallas. J. Egypt. Soc. Parasitol. 26, 93–104.
Gleave, K., Cook, D., Taylor, M.J., Reimer, L.J., 2016. Filarial infection influences mos-quito behaviour and fecundity. Sci. Rep. 6, 36319.
Harbach, R.E., 2018. Mosquito Taxonomic Inventory. (Accessed 15 May, 2018). http://mosquito-taxonomic-inventory.info/.
Hillyer, J.F., 2010. Mosquito immunity. Adv. Exp. Med. Biol. 708, 218–238.Hillyer, J.F., Strand, M.R., 2014. Mosquito hemocyte-mediated immune responses. Curr
Opin Insect Sci. 3, 14–21.Jariyapan, N., Saeung, A., Intakhan, N., Chanmol, W., Sor-Suwan, S., Phattanawiboon, B.,
Taai, K., Choochote, W., 2013. Peritrophic matrix formation and Brugia malayi mi-crofilaria invasion of the midgut of a susceptible vector, Ochlerotatus togoi (Diptera:Culicidae). Parasitol. Res. 112, 2431–2440.
Joshi, D., Choochote, W., Park, M.H., Kim, J.Y., Kim, T.S., Suwonkerd, W., Min, G.S.,2009. The susceptibility of Anopheles lesteri to infection with Korean strain ofPlasmodium vivax. Malar. J. 8, 42.
Joshi, D., Park, M.H., Saeung, A., Choochote, W., Min, G.S., 2010. Multiplex assay toidentify Korean vectors of malaria. Mol. Ecol. Resour. 10, 748–750.
Macdonald, W.W., 1962. The Selection of a Strain of Aedes aegypti susceptible to infectionwith semi-periodic Brugia malayi. Ann. Trop. Med. Parasitol. 56, 368–372.
Macdonald, W.W., 1976. In: genetic aspects of host-parasite relationship. Symposia of theBritish Society for Parasitology. Blackwell Scientific Publications, Oxford, pp. 1–24.
Macdonald, W.W., Ramachandran, C.P., 1965. The influence of the gene Fm (filarialsusceptibility, Brugia malayi) on the susceptibility of Aedes aegypti to seven strains ofBrugia, Wuchereria and Dirofilaria. Ann. Trop. Med. Parasitol. 59, 64–73.
McGreevy, P.B., McClelland, G.A., Lavoipierre, M.M., 1974. Inheritance of susceptibilityto Dirofilaria immitis infection in Aedes aegypti. Ann. Trop. Med. Parasitol. 68, 97–109.
Pang, S.C., Andolina, C., Malleret, B., Christensen, P.R., Lam-Phua, S.G., Razak, M.A.B.A.,Chong, C.S., Li, D., Chu, C.S., Russell, B., Rénia, L., Ng, L.C., Nosten, F., 2017.Singapore’s Anopheles sinensis Form A is susceptible to Plasmodium vivax isolates from
the western Thailand-Myanmar border. Malar J. 16, 465.Rattanarithikul, R., Harrison, B.A., Harbach, R.E., Panthusiri, P., Coleman, R.E., 2006.
Illustrated keys to the mosquitoes of Thailand IV. Anopheles. Southeast Asian J. Trop.Med. Public Health. 37, 1–128.
Rongsriyam, Y., Jitpakdi, A., Choochote, W., Somboon, P., Tookyang, B., Suwonkerd, W.,1998. Comparative susceptibility of two forms of Anopheles sinensis Wiedemann 1828(Diptera : Culicidae) to infection with Plasmodium falciparum, P. vivax, P. yoelii andthe determination of misleading factor for sporozoite identification. Southeast AsianJ Trop Med Public Health. 29, 159–167.
Saeung, A., Choochote, W., 2013. Development of a facile system for mass production ofBrugia malayi in a small-space laboratory. Parasitol. Res. 112, 3259–3265.
Saeung, A., Hempolchom, C., Baimai, V., Thongsahuan, S., Taai, K., Jariyapan, N.,Chaithong, U., Choochote, W., 2013. Susceptibility of eight species members in theAnopheles hyrcanus group to nocturnally subperiodic Brugia malayi. Parasit Vectors6, 5.
Saeung, A., Min, G.S., Thongsahuan, S., Taai, K., Songsawatkiat, S., Choochote, W., 2014.Susceptibility of five species members of the Korean Hyrcanus Group to Brugia ma-layi, and hybridization between B. malayi-susceptible and -refractory Anopheles si-nensis strains. Southeast Asian J. Trop. Med. Public Health 45, 588–597.
Severson, D.W., DeBruyn, B., Lovin, D.D., Brown, S.E., Knudson, D.L., Morlais, I., 2004.Comparative genome analysis of the yellow fever mosquito Aedes aegypti withDrosophila melanogaster and the malaria vector mosquito Anopheles gambiae. J Hered.95, 103–113.
Severson, D.W., Mori, A., Zhang, Y., Christensen, B.M., 1994. Chromosomal mapping oftwo loci affecting filarial worm susceptibility in Aedes aegypti. Insect Mol. Biol. 3,67–72.
Sulaiman, I., Townson, H., 1980. The genetic basis of susceptibility to infection withDirofilaria immitis in Aedes aegypti. Ann. Trop. Med. Parasitol. 74, 635–646.
Taai, K., Baimai, V., Saeung, A., Thongsahuan, S., Min, G.S., Otsuka, Y., Park, M.H.,Fukuda, M., Somboon, P., Choochote, W., 2013. Genetic compatibility betweenAnopheles lesteri from Korea and Anopheles paraliae from Thailand. Mem. Inst.Oswaldo Cruz. 108, 312–320.
Trpis, M., Duhrkopf, R.E., Parker, K.L., 1981. Non-Mendelian inheritance of mosquitosusceptibility to infection with Brugia malayi and Brugia pahangi. Science. 211,1435–1437.
WHO, 2017. Global programme to eliminate lymphatic filariasis: progress report, 201691. Wkly Epidemiol Rec., pp. 589–608.
WHO, 2013. Towards Eliminating Lymphatic Filariasis:Progress in the South-East Region.World Health Organization Regional Office for South-East Asia, New Delhi, India, pp.2001–2011.
Zielke, E., 1973. Studies on the heredity of susceptibility to the canine filaria, Dirofilariaimmitis, in Culex pipiens fatigans and Aedes aegypti. Z Tropenmed Parasitol. 24, 36–44.
W. Dedkhad et al. Acta Tropica 187 (2018) 65–71
71