effects of caffeine on amino acid transport in the brain
TRANSCRIPT

Neurochem. Int. Vol. 14, No. 1, pp. 55-60, 1989 0197-0186/89 $3.00+0.00 Printed in Great Britain. All rights reserved Copyright © 1989 Pergamon Press plc
EFFECTS OF CAFFEINE ON AMINO ACID TRANSPORT IN THE BRAIN
EDMUND A. DEBLER,* ISABEL WAJDA, ISAAC MANIGAULT, ALESSANDRO P. BURLINAt and ABEL LAJTHA
The Nathan S. Kline Institute for Psychiatric Research, Center for Neurochemistry, Ward's Island, New York, NY 10035, U.S.A.
(Received 21 June 1988; accepted 16 August 1988)
Abstract--Chronic administration of caffeine to mice did not alter cellular (low-affinity) transport as measured in brain slices of the amino acids y-aminobutyric acid, glutamic acid, and glycine. Chronic caffeine administration did, however, increase the long-term (60-min) uptake of a-aminoisobutyric acid and valine into brain slices. A similar tendency, although not statistically significant, towards increased amino acid uptake was also seen in the transport of phenylalanine and lysine across the blood-brain barrier in chronically treated rats. The increase in neutral amino acid uptake seems unrelated to changes in brain protein metabolism, since chronic caffeine administration did not significantly alter brain protein synthesis rates. In vitro, caffeine (0.01-1 mM) did not alter brain slice transport of the amino acids 3'-aminobutyric acid, glutamic acid, glycine, valine, and the amino acid analog a-aminoisobutyric acid. Caffeine (1-100#M) was also ineffective in altering the synaptosomal (high-affinity) transport of the neurotransmitter amino acids 3'-aminobutyric acid, glutamic acid, and glycine. These data support the idea that caffeine is a mildly stimulating drug with no significant effect on neurotransmitter amino acid transport systems in rodent brain.
Current theories on the central action of caffeine in the brain implicate interactions with specific mem- brane functions and thereby membrane components such as transport carriers or membrane bound recep- tors. For example, Schlosberg et aL (1981) demon- strated that caffeine injected i.p. (10 mg/kg) into male rats acutely increased brain levels of tryptophan, serotonin (5-HT), and 5-hydroxyindoleacetic acid (5-HIAA). The elevated levels of 5-HT and 5-HIAA in the brain may be attributed to the enhanced up- take of the precursor amino acid tryptophan from the blood (Schlosberg et al., 1981; Fernstrom and Wurtman, 1971; Dickerson and Pao, 1975) via a direct effect of caffeine on the transport of neutral amino acids.
The role of caffeine in increasing the effectiveness of neutral amino acid uptake by the brain may also be indirect. For instance, insulin, which is secreted as a response to the caffeine analog theophyiline (Turtle et al., 1967), has been shown to enhance the uptake
*Address all correspondence to: Dr Edmund A. Debler, Center for Neurochemistry, Ward's Island, New York, NY 10035, U.S.A.
tOn leave from the Istituto delle Malattie Nervose e Mentali, Universitfi di Padova, Italy.
of L-[3H]tryptophan into the brain (Fernstrom and Wurtman, 1971) and to increase brain tryptophan levels (Schlosberg et al., 1981). The mechanism be- hind this insulin-induced increase in brain tryptophan transport is not fully defined, but it appears to be related to an insulin-induced decrease of amino acids competing with tryptophan for transport from plasma (Fernstrom and Wurtman, 1972).
Of specific membrane components potentially in- volved in the central action of caffeine and other methylxanthines, an impressive body of evidence specifically implicates adenosine receptors (Daly et al., 1981). In studies in which adenosine receptors were labeled with the nonmetabolizable iigand N6-cyclohexyl [3H]adenosine ([3H]CHA) (Bruns et al., 1980) it was observed that chronic caffeine treatment enhanced the number of [3H]CHA receptors in the reticular formation (Chou et aL, 1985). Chronic administration of the methylxanthine theophylline also results in an upregulation of adenosine receptors (in the rat cortex) (Murray, 1982). This upregulation appears to be a response to the competitive blockage of adenosine receptors by the antagonistic methyl- xanthines (Snyder et al., 1981). The blocking action by methylxanthines was also observed in studies on [3H]norepinephrine release (Hams et al., 1978;
55

56 EDMUND A. DEBLER et al.
Jackisch et al., 1985). These effects of caffeine may involve changes in the metabol ism of the receptor protein. To our knowledge, few studies have been conducted on the effects of caffeine on brain protein synthesis. In developmental studies, caffeine aug- mented bra in protein synthesis in newborn rats tha t were malnour ished, but not in those normal ly nour ished (Mori et al., 1984). Using post-mito- chondr ia l superna tan ts from adult rats to measure cell-free prote in synthesis, it was shown tha t caffeine in the micromolar range in the incubat ion medium caused a small increase in prote in synthesis (Felipo et al., 1986). High concent ra t ions (0.2-1 mM), however, proved to be inhibitory.
In the present study we evaluated the effects of caffeine, adminis tered bo th in rico and in vitro, on the t r anspor t of amino acids into bra in slices (low-affinity cellular t ranspor t ) and synaptosomal fract ions (high- affinity terminal t ransport) . The effect of caffeine adminis tered in vivo was also-evaluated with respect to amino acid t r anspor t across the b l ood - b r a i n bar- rier and with respect to bra in prote in synthesis.
EXPERIMENTAL PROCEDURES
Caffeine administration in vivo In the brain slice transport studies and in the evaluation
of protein synthesis rates, the mice (BALB/cBy) were housed in groups of 6, and caffeine was administered for 3 weeks in the drinking water, 0.5 mg/ml during the first week and 1.0 mg/ml during the following 2 weeks. The weight and the fluid consumption were measured twice a week. Calcu- lated from those values, the average caffeine intake was 80 mgtkg/day in the first week, 260 mg/kg/day in the second, and 160 mg/kg/day in the third, respectively.
In the second group of experiments in the study of transport across the blood-brain barrier, rats (Wistar) were also exposed to caffeine in the drinking water for 3 weeks. Body weight and fluid consumption were measured twice a week and caffeine concentration in the water was adjusted to result in caffeine intake values of 90, 230, and 160mg/kg/day during the first, second, and third week respectively.
Amino acid uptake into brain slices Cerebral cortical hemispheres from adult BALB/cBy mice
were sliced with a Mcllwain tissue chopper. Slices of 0.42-mm thickness were incubated in a HEPES-buffered medium as described previously (Sershen and Lajtha, 1979). After a 15-min pre-incubation, ~4C-labeled and unlabeled amino acid were added to the incubation media to obtain a final concentration of 0.1 mM. Incubation was then continued for either 5 or 60 rain. In studies concerned with effects/n vitro, caffeine was added immediately before the labeled substrate. After the uptake period, the medium was filtered from the slices, which were weighed, then homo- genized in perchloric acid (5% w/v), and centrifuged. A portion of the perchloric acid extract was added to scintillation fluid and the radioactivity of the tissue was determined.
Amino acid uptake into synaptosomal fractions Synaptosomal P: fractions were prepared from the cere-
bral cortex of adult BALB/cBy mice. Brain tissue was homogenized in 0.32 M sucrose (Tris-HC1) buffer as de- scribed previously (Debler et al., 1986) and centrifuged at 1000g for 10min. The supernatant was centrifuged at 14,000g for 10 rain and the P2 pellet was resuspended in 0.32 M sucrose (Tris-HCl). Uptake was measured at 32°C for 1 min for L-[14C]glutamic acid (4 #M), and for 2 min for )'-aminobutyric acid (GABA) (2pM) and glycine (7/tM) (Debler and Lajtha, 1987). Incubation was conducted in HEPES buffer as described above, and was terminated by filtration on Whatman glass fiber filters (GF/B) under vacuum, rinsing once with 5 ml of ice-cold buffer. The uptake values presented represent the specific uptake, i.e. total uptake minus the uptake in the presence of 100-fold excess unlabeled amino acid, to correct for nonspecific synaptosomal binding or diffusion.
Amino acid transport across the blood--brain barrier The transport of amino acids across the blood-brain
barrier was measured by comparing the passage of ~4C-labeled amino acid to a 3H-labeled water reference from the capillary circulation (Oldendorf, 1971). Briefly, adult rats (250-350g) were anesthetized with chloral hydrate (380mg/kg), and the left carotid artery was exposed. A mixture of 0.1-0.5 pCi of ~4C-labeled amino acid and 2/~Ci of 3H-labeled water in 0.2 ml of HEPES-buffered medium was injected with a 27-gauge needle into the left carotid artery. The rat was decapitated 15 s after the injection. The left cerebrum was removed and homogenized in 2 ml of perchloric acid. After centrifugation, the clear supernatant (0.5 mg) was added to scintillation fluid and radioactivity was determined. The brain uptake index (% uptake with water uptake as 100) was calculated as tissue t4CpH ratio divided by the ~4C/3H ratio of the solution used for injection and multiplied by 100 (Oldendorf, 1971).
Protein synthesis To measure average protein synthesis rates, mice were
injected with 0.2pCi of L-[U-CI4]valine (ICN Radio- chemical, Irvine, Calif.; sp. act. 225 mCi/mmol) per g body weight in 0.5 ml of 500 mM valine. This large dose of amino acid was used in order to flood the pool of amino acid, which otherwise might equilibrate slowly. This method, which we used previously (Dunlop et al., 1975), results in constant specific activity of the free amino acid pool and measures protein synthesis rates reliably. After 1 h the mice were decapitated and the brain was dissected on ice.
The hemispheres were cut into 0.26-ram thick slices with a Mcllwain tissue chopper, and the tissue was homogenized in 5ml of 6% perchloric acid, followed by chloro- form:methanol (1:1) mixture, and finally by ether. After drying, the protein was weighed, digested with i N NaOH, and acidified; the protein content was determined and its label was counted at 85% etficiency in Triton-X-toluene scintillation fluid.
RESULTS
Uptake o f amino acids into brain slices
Glycine, L-glutamic acid, G A B A , valine, or ~-aminoisobutyr ic acid (AIB) uptake in slices of
0
°1 ° o ~
-![ I -
O ~ O C
+1 +1 -H +
+1 +) +1 + ~
i
+1 +1 +t + N
+l +1 +) + ~
,..~ ,...: ,..~ ,...~ o
~ + 1 +
0 0 0 0
+1~+1+1
0 0 0 0
+)+1+1+1
+1 +1 +1 +1
+1 +[ +1
+1 +1 +1 +1
Caffeine and amino acid transport
il
57
E .=_
"8
:a. I
.=.
:a.
~ m
).~'~
.~~ ~
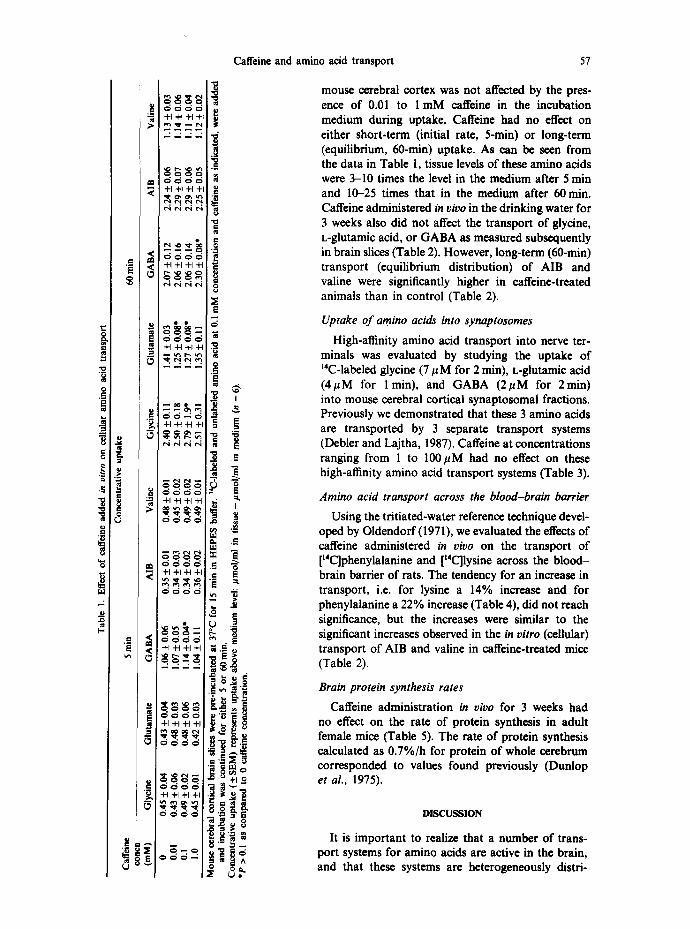
mouse cerebral cortex was not affected by the pres- ence of 0.01 to 1 mM caffeine in the incubation medium during uptake. Caffeine had no effect on either short-term (initial rate, 5-rain) or long-term (equilibrium, 60-min) uptake. As can be seen from the data in Table 1, tissue levels of these amino acids were 3-10 times the level in the medium after 5 min and 10-25 times that in the medium after 60 min. Caffeine administered in vivo in the drinking water for 3 weeks also did not affect the transport of glycine, L-glutamic acid, or GABA as measured subsequently in brain slices (Table 2). However, long-term (60-min) transport (equilibrium distribution) of AIB and valine were significantly higher in caffeine-treated animals than in control (Table 2).
Uptake o f amino aciab into synaptosomes
High-affinity amino acid transport into nerve ter- minals was evaluated by studying the uptake of 14C-labeled glycine (7/~ M for 2 min), L-glutamic acid (4#M for l min), and GABA (2/zM for 2rain) into mouse cerebral cortical synaptosomal fractions. Previously we demonstrated that these 3 amino acids are transported by 3 separate transport systems (Debler and Lajtha, 1987). Caffeine at concentrations ranging from 1 to 100 tzM had no effect on these high-affinity amino acid transport systems (Table 3).
Amino acid transport across the blood-brain barrier
Using the tritiated-water reference technique devel- oped by Oldendorf (1971), we evaluated the effects of caffeine administered /n vivo on the transport of [14C]phenylalanine and [14C]lysine across the blood- brain barrier of rats. The tendency for an increase in transport, i.e. for lysine a 14% increase and for phenylalanine a 22% increase (Table 4), did not reach significance, but the increases were similar to the significant increases observed in the in vitro (cellular) transport of AIB and valine in caffeine-treated mice (Table 2).
Brain protein synthesis rates
Caffeine administration in vivo for 3 weeks had no effect on the rate of protein synthesis in adult female mice (Table 5). The rate of protein synthesis calculated as 0.7%/h for protein of whole cerebrum corresponded to values found previously (Dunlop et al., 1975).
D I S C U S S I O N
It is important to realize that a number of trans- port systems for amino acids are active in the brain, and that these systems are heterogeneously distil-

58 EDMUND A. DEBLER et al.
Table 2. Effect of caffeine administered in vivo on cellular amino acid transport
['4C]compound 5 min
Control Concentrative uptake
Caffeine treated
60 min 5 min 60 min
AIB n = 9 0.12+0.01 0.97_+0.11 0 .15+0 .02 1.27_+0.07" Valine n = 6 0.20 _+ 0.01 0.73 + 0.03 0.21 _+ 0.02 0.82 -+ 0.07* Glutamate n = 12 1.11 +0.04 1.93_+0.07 1.17_+0.05 1.89_+0.04 GABA n = 12 1.34-+0.06 2.01 -+0.07 1.28_+0.07 1.84-+0.09 Glycine n = 12 0.61 + 0.03 2.49 -+ 0.08 0.62 +_ 0.05 2.57 + 0.10
Caffeine was administered for 3 weeks in the drinking water. Caffeine intake was 80 mg/kg/day during the first week, 260 mg/kg/day during the second week, and 160 mg/kg/day during the third week. Mouse cerebral cortical slices were pre-incubated at 37°C for 15rain in HEPES buffer. 14C-labeled and unlabeled amino acid was added at 0.1 mM concentration and incubation was continued for either 5 or 60min. Concentrative uptake (_+SEM) represents uptake above medium levels: #mol/ml t i s sue - #mol/ml in medium.
AIB = :t-aminoisobutyric acid; GABA = ),-aminobutyric acid. *P < 0.05, as compared to corresponding control values.
Table 3. Effect of caffeine in vitro on synaptosomal amino acid transport
Uptake (nmol/mg protein/min) Caffeine concentration (uM)
[]4C]compound 0 1.0 I 0 100
Glycine (7 p M) 0.145 + 0.004 0.144 _+ 0.004 0.140 _+ 0.004 0.142 + 0.002 Glutamate (4 pM) 1.26 + 0.03 1.23 _+ 0.01 1,21 + 0.02 1.21 + 0.03 GABA (2/zM) 0.36 + 0.02 0.37 + 0.03 0.37 + 0.01 0.37 + 0.01
Uptake was monitored in HEPES buffer for 2 rain at 32"C. Values are averages of 3 experiments + SEM.
buted; i.e. any membrane contains only some of the systems. In cellular uptake with brain slices we could distinguish at least 10 separate systems for amino acids (Sershen and Lajtha, 1979); only a few of these are present in brain capillaries. There are indications that systems present in synaptosomal fractions and synaptic storage vesicles represent separate systems with significantly different properties. For example, not only do GABA and glutamate represent two different transport systems, but several systems exist for each, and their regional, neuronal, glial, and vesicular uptake is mediated by systems that differ in substrate specificity (competing amino acids), energy dependence (Na, ATP dependence), and kinetic constants (Allen et al. , 1986; Ferkany and Coyle, 1986; Debler and Lajtha, 1987). The direction of
Table 4. Effect of caffeine in vivo on amino acid transport across the blood-brain barrier
Uptake (BUI)t [L4C]compound Control Treated
Phenylalanine 26.4 _+ 1.7 33.8 + 3.5* Lysine 16.0 + l.I 18.6 + 1.6
Rats were exposed to caffeine in the drinking water for 3 weeks (see Experimental Procedures).
tBU1, brain uptake index: percentage uptake of 14C-labeled compound compared to the uptake of 3H-labeled water in brain (rat) after injection into the carotid artery.
*P = 0.08, as compared to control.
transport by these systems may also differ, some primarily active inward, others outward (Betz and Goldstein, 1978). Although short-term uptake is gov- erned by initial rates of inward transport, equilibrium will be governed by two-directional movement, the properties about which we know considerably less. Since it is likely that compounds do not affect all transport systems in similar fashion, substrates for several systems have to be examined. No direct effect of caffeine administered in vitro was noted on the transport of glycine, glutamate, GABA, valine, or AIB in brain slices (low-affinity cellular transport) or of glycine, glutamate, or GABA in synaptosomes (high-affinity nerve terminal transport) (Tables 1 and 3).
In vivo administration also did not alter the trans- port of the putative amino acid neurotransmitters,
Table 5. Effect of caffeine in vivo on brain protein synthesis rates
dpm/mg protein + SEM
Control (n = 6) 228 + 12 Treated (n = 7) 208 + 8*
Female BALB/cBy mice were exposed to caffeine in the drinking water for 3 weeks. One hour incorporation of a flooding dose of labeled valine was measured (see Experimental Procedures).
*P >0.1.

Caffeine and amino acid transport 59
i.e. glycine, glutamate, and GABA, into brain slices (Table 2). Thus, in contrast to its effects on scrotonin/serotonin precursors (Schlosberg et al.,
1981; Fernstrom and Wurtman, 1971, 1972; Dickerson and Pao, 1975; Turtle et al., 1967), caffeine proved to be ineffective in altering the transport of neurotransmitter amino acids. However, caffeine administered in vivo did alter the long-term (60-min) transport of AIB and valine. AIB is a non- metabolizable neutral amino acid analog used as a model substrate for low-affinity amino acid uptake processes (Blasberg and Lajtha, 1966). Valine, like AIB, is transported by a low-affinity transport system, specifically the L (leucine-preferring) system (Sershen and Lajtha, 1979). For both AIB and valine, caffeine treatment resulted in a significant increase in uptake, 30 and 13%, respectively (Table 2). These results are in agreement with the observed caffeine-promoted increase of neutral amino acid transport from the blood into the brain (Schlosberg et al., 1981; Fernstrom and Wurtman, 1971; Dickerson and Pao, 1975). The caffeine treatment also showed a tendency, although not a significant one, to increase the transport of lysine and phenyl- alanine into the brain across the blood-brain barrier as determined by the tritiated-water reference technique. These changes in the low-affinity transport of neutral amino acids appear not to be related to changes in protein content, since in rico caffeine administration did not significantly alter brain protein synthesis rates (Table 4). These data, though not statistically significant, do demonstrate a tendency towards a reduction in the rate of brain protein synthesis (in rico) that is consonant with the in vitro studies of Felipo et al. (1986) in which they demonstrated that high concentrations of caffeine (0.2-1 mM) inhibited protein synthesis in post-mitochondrial supernatants.
The effect on long-term uptake of neutral amino acids indicates a selectivity of caffeine effects on cerebral amino acid transport. The presence of caffeine does not influence all amino acid transport systems, but only one or a few, and it seems to influence exit processes more than uptake. This suggests some small alteration of membrane proper- ties or processes that may influence neural function. Changes in the level of neutral amino acids are not likely to influence protein metabolism but may influence neurotransmitter formation from tyrosine or tryptophan.
In a previous study we demonstrated that caffeine had little or no effect on monoaminergic systems in mouse brain (Reith et al., 1987). In agreement with
that work, we at present also find little or no effect of caffeine on neurotransmitter amino acid transport systems in rodent brain.
Acknowledgement--The grant support of the International Life Sciences Institute is gratefully acknowledged.
REFERENCES
Allen I. C., Schousboe A. and Griffiths R. 0986) Effect of L-homocysteine and derivatives on the high-affinity up* take of taurine and GABA into synaptosomes and cul- tured neurons and astrocytes. Neurochem. Res. 11, 1487-1496.
Betz A. L and Goldstein G. W. (1978) Polarity of the blood-brain barrier: neutral amino add transport into isolated brain capillaries. Science 202, 225-227.
Blasberg R. and Lajtha A. (1966) Heterogeneity of the mediated transport systems of amino acid uptake in brain. Brain Res. 1, 86-104.
Bruns R. F., Daly J. W. and Snyder S. H. (1980) Adenosine receptors in brain membranes: binding of Ne-cyclohexyl [3H]adenosine and 1,3-diethyl-8-[3H]phenylxanthine. Proc. natn. Acad. Sci. U.S.A. 77, 5547-5551.
Chou D. T., Khan S., Forde J. and Hirsh K. R. (1985) Caffeine tolerance: behavioral, electrophysiological and neurocbemical evidence. Life Sci. 36, 2347-2358.
Daly J. W., Bruns R. F. and Snyder S. H. (1981) Adenosine receptors in the central nervous system: relationship to the central actions of methylxanthines. Life Sci. 28, 2083-2097.
Debler E. A. and Lajtha A. (1987) High-affinity transport of ~,-aminobutyric acid, glycine, taurine, L-aspartic acid, and L-glutamic acid in synaptosomal (P2) tissue: a kinetic and substrate specificity analysis. J. Neurochem. 48, 1851-1856.
Debler E. A., Sershen H., Lajtha A. and Gennaro J. F. jr (1986) Superoxide radical-mediated alteration of syn- aptosome membrane structure and high-affinity ),-[t4C]aminobutyric acid uptake. J. Neurochem. 47, 1804-1813.
Dickerson J. W. T. and Pao S. K. (1975) The effect of a low protein diet and exogenous insulin on brain tryptophan and its metabolites in the weanling rat. J. Neurochem. 25, 559-564.
Dunlop D. S., van Elden W. and Lajtha A. (1975) A method for measuring brain protein synthesis rates in young and adult rats. J. Neurochem. 24, 337-344.
Felipo V., Portoles M., Minana M. D. and Grisolia S. (1986) Rats that consume caffeine show decreased brain protein synthesis. Neurochem. Res. 11, 63-69.
Ferkany J. and Coyle T. (1986) Heterogeneity of sodium- dependent excitatory amino acid uptake mechanisms in rat brain. J. Neurosci. Res. 16, 491-503.
Fernstrom J. D. and Wurtman R. J. (1971) Brain serotonin content: increase following ingestion of carbohydrate diet. Science 174, 1023-1025.
Fernstrom J. D. and Wurtman R. J. (1972) Brain serotonin content: physiological regulation by plasma neutral amino acids. Science 178, 414-416.
Hams H. H., Wardel G. and Mulder A. H. (1978) Ade- nosine modulates depolarization-induced release of 3H-noradrenaline from slices of rat brain neocortex. Fur. J. Pharmac. 49, 305-308.

60 EDMUND A. DEBLER et al.
Jackisch R., Fehr R. and Hertting G. (1985) Adenosine: an endogenous modulator of hippocampal noradrenaline release. Neuropharmacology 24, 499-507.
Mori M., Wilber J. F. and Nakamoto T. (1984) Protein- energy malnutrition during pregnancy alters caffeine's effect on brain tissue of neonate rats. Life Sci. 35, 2553-2560.
Murray T. F. (1982) Up-regulation of rat cortical adeno- sine receptors following chronic administration of theophylline. Fur. J. Pharmac. 82, 113-114.
Oldendorf W. H. (1971) Brain uptake of radiolabeled amino acids, amines and hexoses after arterial injection. Am. J. Physiol. 22, 1629-1639.
Reith M. E. A., Sershen H. and Lajtha A. (1987) Effects of caffeine on monoarninergic systems in mouse brain. Acta biochim, biophys, hung. 22, 149-163.
Schlosberg A. J., Fernstrom J. D., Kopczynski M. C.. Cusack B. M. and Gillis M. A. (1981) Acute effects of caffeine injection on neutral amino acids and brain mono- amine levels in rats. Life Sci. 29, 173-183.
Sershen H. and Lajtha A. (1979) Inhibition pattern by analogs indicates the presence of ten or more transport systems for amino acids in brain cell. J. Neurochem. 32, 719-726.
Snyder S. H., Katims J. J., Annan Z., Bruns R. F. and Daly J. W. (1981) Adenosine receptors and behavioral actions of methylxanthines [caffeine/theophylline/N6-cyclohexyl - adenosine/N6(phenylisopropyl)adenosine]. Proc. nam. Acad. Sci. U.S.A. 78, 3260-3264.
Turtle J. R., Littleton G. K. and Kipnis D. M. (1967) Stimulation of insulin secretion by theophylline. Nature 213, 727-728.