effect of split doses of n-methyl-n-nitrosourea on dna repair synthesis in cultured mammalian cells
TRANSCRIPT

Cancer Letters, 3 (1977) 183--188 193 © Elsevidr/North-Holland Scientific Publishers, Ltd.
E F F E C T OF SPLIT DOSES OF N-METtIYL-N-NITROSOUREA ON DNA REPAIR SYNTHESIS IN CULTURED MAMMALIAN CELLS
LUCIANO ZARDI*, LEONE ST. VINCENT, ALAIN BARBIN, RUGGERO MONTESANO and G E O F F R E Y P. MARGISON**
International Agency for Research on Cancer, 150 cours Albert Thomas, 69008 Lyon (France)
(Received 4 April 1977) (Accepted 25 May 1977)
SUMMARY
DNA repair was measured in human fibroblasts, mouse C3H 10 T 1~ flbro- blasts and rat hepatocytes by the non-semi-conservative incorporation of [3H]- TdR during DNA repair synthesis using liquid scintillation techniques. Con- f luent monolayers of these cells grown on cover slips were exposed to split doses (125 or 250 pg/ml) of the mutagenic and carcinogenic alkylating agent MNU and DNA repair synthesis compared with that produced by a single dose (500 ~g/ml). No significant difference in DNA repair capacity was detected in the three cell lines treated with a single dose or split doses of MNU.
INTRODUCTION
It has been suggested that DNA repair processes could be an important factor in the mechanism of carcinogenesis [13] . A reduction in the activity of enzymes which excise abnormal methyla ted bases from DNA has been shown in the livers of rats t reated with consecutive doses of MNU in vivo [7] . Recently, it has been reported that a reduct ion in the capacity of DNA repair synthesis occurs in cultured fibroblasts after exposure to split doses of 4-nitroquinoline-l-oxide [14] .
In order to determine whether this is a general phenomenon, we have inves- tigated whether similar reductions in DNA repair synthesis capacity take place
*L.Z. was on leave of absence from the Ist i tuto di Oncologia, Universit~ di Genova and was suppor ted by the Consorzio Antineoplast ico Regione Liguria, Viale Benedetto XV, 10, Genova, Italy. **Present address: Paterson Laboratories, Christie Hospital, Manchester M20 9BX, England. Abbreviations: HU, hydroxyurea ; [3H]TdR, [Me-3H]thymidine; MNU, N-methyl-N-nitro- sourea.

184
in three different cell lines when they are exposed to split doses of MNU. In all the cell lines tested, no significant reductions in DNA repair activity were detected.
MATERIALS AND METHODS
The cells used in these studies were: (a) epithelial-like liver cells (IAR-20) originating from 10-day-old BD-VI rats, (b) mesenchymal cells (IAR-28) originating from the liver of a 40-year-old man and (c) C3H mouse 10 T 1A clone 8 fibroblasts kindly provided by Prof. C. Heidelberger. The rat cells were cultured on cover slips in Williams D medium, the human cells in MEM and the mouse cells in BME medium, containing 10% foetal calf serum. Unscheduled [3H]TdR incorporation was rout inely measured by scintillation counting of a hot acid extract of the cells. In some instances the cover slips were coated with Kodak NTB3 emulsion and held in light-free boxes for approx. 10 days at 4°C, developed and stained with toluidine.
The protocol was as follows. To the cultured cells was added hydroxyurea (HU) (Sigma, London) to a final concentrat ion of 700 pg/ml. After 15 min the medium was removed and the cells washed once with phosphate-buffered saline. Fresh medium containing 0.5% foetal calf serum, 700 pg/ml HU and 10 pg/ml [3H]TdR (Radiochemical Centre, Amersham, England) (50 Ci/mmol) were added. Immediately afterwards, N-nitrosourea (MNU) (Schuchardt, Munich , West Germany) dissolved in 3 mM citrate 0.9% NaC1, pH 6.0, was added to give the required final concentration. In control experiments only the buffer was added. In the split dose experiments the first dose of MNU was added directly to the medium and after incubation for the required time, t rea tment was carried out as above. In all cases, incubation with [3H]TdR was for 4 h. The cells were then washed on the cover slips with ice-cold saline (3 ml) and incubated at 4°C for 10 min with 3.0 ml of 0.5 N HC104 before being transferred to glass vials. After washing 3 times with 5 ml of ice-cold 0.5 N HC104 and twice with 10 ml ethanol, the cells were extracted with 1 ml of 0.5 HC104 at 80°C for 20 min. Radioactivity was determined in a Packard 3375 spectrometer after the addition of 10 ml of a standard scintillation cocktail containing Triton X-100. The results are expressed as cpm per cover slip. The values are the average + S.D. of the counts of six cover slips treated with MNU in pairs.
RESULTS AND DISCUSSION
Confluent monolayers of the three cell lines examined were treated with either single or split doses of MNU and DNA repair was measured as unscheduled DNA synthesis, i.e. by [3H]TdR incorporation in the presence of HU [9]. Under the conditions used, the semi-conservative incorporation of [3H]TdR was inhibited by more than 95% in the presence of HU (Table 1). Radioactivity in the perchloric acid extract of the carcinogen-treated cells above that in control g~oups (cells treated only with HU) was taken as a measure of the

185
T A B L E 1
I N C O R P O R A T I O N OF [3H]THYMIDINE INTO C U L T U R E D CELLS T R E A T E D WITH V A R I O U S D O S E S OF M N U IN THE P R E S E N C E OF H Y D R O X Y U R E A (700 ttg/ml)
MNU Hydroxyurea cpm/Cover slip (ttl/ml) (700 ttg/ml)
C3H 10 T ½ IAR-28 IAR-20 cells cells cells
0 0 5 5 6 9 0 • 7 0 0 3 1 2 5 8 0 0 ± 1 1 7 2 0 1 3 2 4 2 0 ± 9 7 4 0 0 + 8 3 6 ± 276 2 4 7 6 ± 827 2 2 0 6 ± 630
50 + 2372 ± 663 3 0 4 9 ± 386 2 8 3 0 ± 410 100 + 2 8 5 2 ± 500 3 5 0 6 ± 970 3 4 0 0 ± 930 200 + 3 5 7 9 ± 361 5487 • 1 1 4 6 5 7 0 0 ± 820 4 0 0 + 5 2 1 1 e 993 6 8 0 5 ± 1103 6 7 5 0 ± 940 800 + 5 1 1 6 e 1275 5 3 6 2 ± 1207 5 4 0 0 ± 750
The results are expressed as cpm per cover slip. Exper imenta l details are described in Materials and Methods.
Q~
ss
0
p- i
lO0
50 A B C
T
? I C
-F
RAT LIVER CELLS HUMAN LIVER CELLS IOT½ CELLS
Fig. 1. Effect of consecutive doses of MNU on [SH]TdR incorporat ion in the presence of HU in three dif ferent cell l ines. The first dose was 125 (B) or 250 (C) #g of MNU]ml and this was fo l lowed 3 h later by a second dose of 250 #g of MNU/ml . The results were compared wi th [aH]TdR incorporat ion after a single dose of 500 ttg of M N U / m l (A) taken as 100%.
186
unscheduled incorporation of [~H]TdR. Examination by autoradiography showed a random distribution of grains over the nuclei which occurred in more than 95% of the cells, which is characteristic of DNA repair replication.
The extent of [3H] TdR incorporation after exposure to different doses of MNU is shown in Table 1. The maximum inc'orporation occurred at a dose between 200 and 400 gg/ml in all three cell lines. The slight decrease of [3H]- TdR incorporation at very high doses of MNU (800 pg/ml) was probably due to toxic effects. [ 3H] TdR incorporation into the MNU-treated cells was 2--3 times higher than that in control cells, in the human and rat cell lines, but 6 times higher in the mouse C3H 10 T 1/~ cell line. We have observed, however, that the relative increase in [3H]TdR incorporation could change with the proliferative activity of the cells, possibly due to a higher level in [3H]TdR kinase activity or to a change in the size of the thymidine pool.
The induction of DNA repair synthesis by a single or split dose of MNU in the three cell lines is shown in Fig. 1. [3H]TdR incorporation was measured
o
g
0 e l .
0
I - - I
i00
50
I
I i , I I , i I I I J 4 8 12 16 20 24
Pretreatment interval (hours)
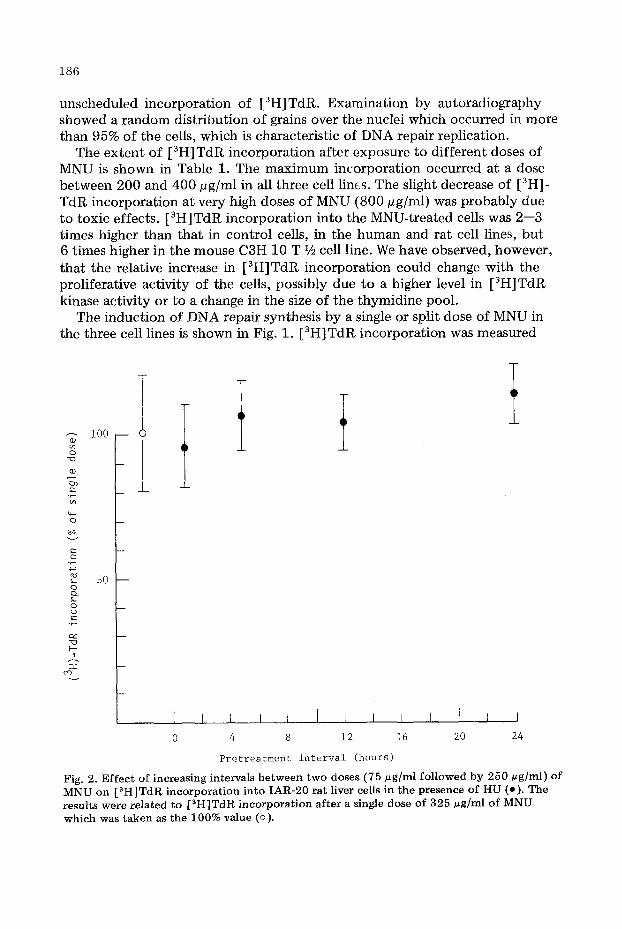
Fig. 2. E f f e c t o f increasing intervals b e t w e e n t w o doses (75 ug /ml f o l l o w e d by 250 ug/ml) o f M N U on [ 3 H ] T d R incorporat ion in to I A R - 2 0 rat l iver cel ls in the presence o f H U (o). The results were related to [ a H ] T d R incorporat ion after a s ingle dose o f 3 2 5 # g ] m l o f M N U w h i c h was taken as the 100% value (o).

187
after a dose of 250 pg/ml which was preceded by 3 h by a dose of either 125 or 250 pg/ml; these results are compared with the effect of a single dose of 500 pg/ml. Although there was an apparent slight reduction in repair synthesis after split doses of MNU, this was not statistically significant even though doses of MNU employed were very high.
In rat liver cells, increasing the interval be tween split doses of MNU (75 pg fol lowed by 250 pg) f rom 1 to 26 h, again did not produce a significant altera- tion in the extent of repair incorporation compared with that induced by a single dose of 325 pg/ml (Fig. 2).
The repair of DNA damaged by mutagenic or oncogenic agents may be an important mechanism by which cells pro tec t themselves against transformation and mutat ion. The inhibition of DNA repair following administration of cocar- cinogens, DNA binding agents and steroids was suggested to be the basis of the biological activity of these agents [2--4] . However, it has now been shown that biological activity is no t confined to effects on DNA repair synthesis bu t also affects normal DNA synthesis and is therefore a non-specific phenomenon [1] . On the other hand, in the Xeroderma pigmentosum syndrome, defective repair of UV damaged DNA does correlate with a high incidence of skin tumours. Furthermore, the low activity of alkylated base excision enzymes in certain tissues has been correlated with the organ specific effect of certain carcinogenic N-nitroso compounds [5--10] .
In human fibroblasts, split doses of 4-nitroquinoline-l-oxide were found to inhibit DNA repair [14] ; however, our results show a lack of effect of split doses of MNU on DNA repair synthesis capacity in the three cell lines examined. The discrepancy between the present results with MNU and those after split doses of 4-nitroquinoline-l-oxide in human fibroblasts ~ a y be related to the nature of the DNA damage and consequent ly the types of repair produced by these different agents [11] .
It remains to be seen if these observations on the DNA repair capacities in these cells treated with split doses of a carcinogen are reflected in differences in the in vitro transformation frequencies.
ACKNOWLEDGEMENTS
This work was partially supported by NCI contract No. ICP-55630.
REFERENCES
1 Cleaver, J.E. and Painter, R.B. (1975) Absence of specificity in inhibition of DNA repair replication by DNA-binding agents, cocarcinogens and steroids in human cells. Cancer Res., 35, 1773--1778.
2 Gaudin, D., Gregg, R.S. and Yielding, K.L. (1971) Repair inhibition: a possible mecha- nism of action of co-carcinogens. Biochem. Biophys. Res. Commun., 45,630--636.
3 Gaudin, D. and Gregg, R.A. (1972) Inhibition of DNA repair by cocarcinogens. Biochem. Biophys. Res. Commun., 48,945--949.
4 Gaudin, D., Guthrie, L. and Yielding, K.L. (1974) DNA repair inhibition: a new mecha- nism of action of steroids with possible implication for tumor therapy. Proc. Soc. Exp. Biol. Med., 146,401--405.

188
5 Goth, R. and Rajewsky, M.F. (1974) Persistence of 06-ethylguanina in rat-brain DNA: correlation with nervous system - - specific carcinogenesis by ethylnitrosourea. Proc. Natl. Acad. Sci. USA, 71 ,639- -643 .
6 Kleihues, P. and Margison, G.P. (1974) Carcinogenity of N-methyl-N-nitrosourea: possible role of excision repair of 06-methylguanine from DNA. J. Natl. Cancer Inst., 53, 1839.
7 Kleihues, P. and Margison, G.P. (1976) Exhaustion and recovery of repair excision of 06-methylguanina from rat liver DNA. Nature, 259, 153--155.
8 Kuroki, T., Drevon, C., Saint Vincent, L., Tomatis, L. and Montesano, R. (1977) Studies on the use of liver parenchymol cells in vitro carcinogenesis. In: Proceedings of C.N.R.S. Internat ional Colloquium, in press.
9 Lieberman, M.W., Baney, R.N., Lee, R.E., Sell, S. and Farber, E. (1971) Studies on DNA repair in human lymphocytes treated with proximate carcinogens and alkylating agents. Cancer Res., 31, 1297--1306.
10 Nicoll, J.W., Swann, P.F. and Pegg, A.E. (1975) Effect of dimethylni t rosoamine on persistence of methyla ted guanines in rat liver and kidney DNA. Nature, 254 ,261- -262 .
11 Regan, J.D. and Setlow, R.B. (1974) Two forms of repair in the DNA of human cells damaged by chemical carcinogens and mutagens. Cancer Res., 34, 3318--3325.
12 San, R.H.C. and Stich, H.F. (1975) DNA repair synthesis of cultured human cells as a rapid bioassay for chemical carcinogens. Int. J. Cancer, 16, 284--291.
13 Van Lanker, J.L. (1974) Carcinogenesis and DNA repair. In: Chemical Carcinogenesis, p. 427. Editors: Paul O.P. Ts'O and J.A. Di Paolo. Marcel Dekker, New York.
14 Warren, P.M. and Stich, H.F. (1975) Reduced DNA repair capacity and increased cyto- toxic i ty following split doses of the mutagen 4-ni t roquinidine-l-oxide in cultured human cells. Mutat. Res., 28 ,285- -293 .