effect of soil compaction on photosynthesis and carbon partitioning within a maize–soil system
TRANSCRIPT

Soil & Tillage Research 71 (2003) 151–161
Effect of soil compaction on photosynthesis and carbonpartitioning within a maize–soil system
Ashraf Tubeileha,∗, Virginie Groleau-Renaudb, Sylvain Plantureuxb,Armand Guckertb
a Department of Soil and Irrigation, Palestinian Ministry of Agriculture, P.O. Box 1217, Nablus, Palestineb Laboratoire Agronomie et Environnement, INPL (ENSAIA)-INRA, BP 172, 54505 Vandoeuvre-lès-Nancy, France
Received 21 August 2002; received in revised form 20 January 2003; accepted 26 January 2003
Abstract
Soil compaction is known to affect plant growth. However, most of the information regarding the effects of this factor oncarbon partitioning has been obtained on young plants while little is known about the evolution of these effects with plantage. The objective of this work was to investigate how soil compaction affects carbon assimilation, photosynthate partitioningand morphology of maize plants during vegetative growth up to tassel initiation. A pressure was applied on moist soil toobtain a bulk density of 1.45 g cm−3 (compacted soil (CS) treatment) while the loose soil (LS) treatment (bulk density of1.30 g cm−3) was obtained by gentle vibration of soil columns. Plants were grown in a growth chamber for 3–6 weeks andcarbon partitioning in the plant–soil system was evaluated using14C pulse-labelling techniques. Soil compaction greatlyhampered root elongation and delayed leaf appearance rate, thereby decreasing plant height, shoot and root dry weights andleaf area. The increase in soil bulk density decreased carbon assimilation rate especially in early growth stages. The main effectof soil compaction on assimilate partitioning occurred on carbon exudation, which increased considerably to the detrimentof root carbon. Furthermore, soil microbial biomass greatly increased in CS. Two hypotheses were formulated. The first wasthat increasing soil resistance to root penetration induced a sink limitation in roots and this increased carbon release intothe soil and resulted in a root feedback that regulated carbon assimilation rate. The second hypothesis relies on soil–plantwater relations since, due to compaction, the pore size distribution has to be considered. In a compacted soil, the peak of thepore size distribution curve is shifted towards the small pore size. The volume of small pores increases and the unsaturatedconductivity decreases substantially, when compared to non-compacted soil. Due to small hydraulic conductivity, the inflowinto the roots is well below optimum and the plant closes stomata thus reducing carbon assimilation rate. The effects of soilcompaction persisted with plant age although the difference between the two treatments, in terms of percentage, decreased atadvanced growth stages, especially in the case of root parameters.© 2003 Elsevier Science B.V. All rights reserved.
Keywords: Carbon; Morphology; Photosynthesis; Plant age; Root exudation; Soil compaction
∗ Corresponding author. Present address: Natural ResourcesManagement Program, International Center for Agricultural Re-search in the Dry Areas (ICARDA), P.O. Box 5466, Aleppo, Syria.Tel.: +963-21-2213477x551; fax:+963-21-2213490.E-mail address: [email protected] (A. Tubeileh).
1. Introduction
Compaction is a change in soil structure that de-creases its porosity and increases its penetrationresistance. This constraint results in restricted rootgrowth that in turn may affect the whole plant growth
0167-1987/03/$ – see front matter © 2003 Elsevier Science B.V. All rights reserved.doi:10.1016/S0167-1987(03)00061-8

152 A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161
and grain yield (Canarache et al., 1984). The mostpronounced effect of soil compaction is the reductionof root length. Several authors have observed suchan effect under different growth conditions.Logsdonet al. (1987)indicated that increasing soil bulk den-sity caused a linear decrease in maize root length withthe increase in soil bulk density. Similar results wereobtained with different crops (maize, barley, wheat,peas and beans) and various culture media (soil andnutrient solution with glass beads) (Abdalla et al.,1969; Asady and Smucker, 1989; Kayombo et al.,1991; Grath and Håkansson, 1992; Oussible et al.,1992; Groleau-Renaud et al., 1998). Meanwhile, theeffect of soil compaction on root biomass depends oncompactness degree, soil water status, and soil phys-ical properties. For instance, whileAtwell (1990a)observed that soil compaction increased root biomassof wheat, other authors have shown that increasingcompactive stress decreases root dry weight of maize(Panayiotopoulos et al., 1994). These morphologi-cal modifications probably affect water and nutrientuptake.
Soil compaction effects extend beyond root mor-phology, affecting both shoot morphology and gen-eral plant physiology. A reduction in leaf area andshoot biomass was observed in maize plants grownon compacted soil (Ekwue and Stone, 1995). Further-more, several authors have reported a decrease in pho-tosynthetic activity due to a drop in stomatal con-ductance in plants grown on compacted soils (Masleet al., 1990; Tardieu et al., 1992). This decrease wasattributed to a chemical message, mainly abscisic acid(ABA), produced in the stressed roots and travelling toshoots via the xylem sap (Masle et al., 1990; Tardieu,1994).
Carbon partitioning within the maize–soil systemwas the subject of several papers (Liljeroth et al.,1994; Qian et al., 1997). However, the effect of soilcompaction on carbon partitioning has rarely been in-vestigated. Studying carbon partitioning under differ-ent bulk densities is of major interest as it determinesroot:shoot ratio and grain yield.Sauerbeck and Helal(1986) labelled maize plants continuously from day7 to day 23 in order to study photosynthate distribu-tion. They reported increases with increasing soil bulkdensity in both the fraction of carbon recovered in thesoil and in that respired by roots and soil microorgan-isms. On the other hand,14C remaining in roots de-
creased with increasing soil strength. Similarly, rootexudation was stimulated when roots of young maizeplants were impeded in a glass bead nutrient solution(Boeuf-Tremblay et al., 1995). A main shortage in ex-isting data is that they were obtained using very youngplants (up to 3 weeks old). Moreover, some of the pre-vious works were conducted in hydroponic conditionsthat do not represent soil conditions.
The aim of the present paper is to track the evolu-tion of the effects of soil compaction on photosynthe-sis, carbon partitioning (including in the rhizosphere)and plant morphology of maize plants ageing from 21to 42 days (shortly before tassel initiation). We areparticularly interested in the effect of soil bulk densityon carbon budget in the maize–soil system and on soilmicrobial biomass.
2. Materials and methods
The culture medium used in this study was a mix-ture of sand and sandy loam soil. Soil was air-driedand sieved to pass a 5 mm mesh before being mixedwith 2 mm sieved sand (two-thirds to one-third). Themineral composition of the final mixture was 12.6%clay, 13.5% silt and 73.9% sand, while total carboncontent was 1.5%. The total porosity of the mixture(bulk density∼1.3 g cm−3) was around 50% whileits field capacity was 23% (w/w). The mixture wasthen packed into polyvinyl chloride (PVC) cylinders(40 cm× 15.5 cm inner diameter) and the cylinderswere closed at the bottom with a mesh to ensure goodaeration of the soil column. A 2 cm gravel layer sep-arated the soil from the mesh at the bottom.
The soil mixture was moistened and uniformlypacked in the cylinders with gentle vibration to obtainthe bulk density of 1.3 g cm−3, which represented theloose soil (LS). Meanwhile, the CS (bulk density of1.45 g cm−3) was prepared using a soft pressure ex-erted by hand. The height of the soil layer was 35 cmand that of headspace was 3 cm.
Maize seeds (Zea mays L., cv. Dea) were germi-nated on agar for 72 h in darkness at 25◦C and uni-form plantlets were selected. Thereafter, one plantletwas planted in each soil cylinder and placed in agrowth cabinet programmed to run at 16 h photope-riod, with a photosynthetic photon flux density (PPFD)of 300–400�mol m−2 s−1, and temperatures of 23◦C

A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161 153
by day and 18◦C by night. Relative humidity wasmaintained at 65–75% and soil water content was keptat 15% of soil dry weight throughout the growth pe-riod. Preliminary studies showed that this level is op-timum for maize growth under our conditions. Waterlosses were estimated and compensated by weighingevery 2 days and plants were irrigated withHoaglandand Arnon (1938)nutrient solution. For all the ex-periments detailed below, the plants were grown un-der these conditions from the first day after planting(DAP). However, for technical reasons the plants werenot planted at the same time.
2.1. Morphology experiment
Five plants were harvested at 21, 35 and 42 DAPin soil columns. Plant height, number of visibleleaves, root number and length of main root axeswere measured at harvest. Leaf area was determinedusing Analyra 1995 software (CIRAD, Montpellier,France). This software calculates leaf area by multi-plying the number of pixels by their size. Shoot androot dry weights were determined after being dried at60◦C for 3 days.
2.2. Carbon assimilation experiment
Photosynthetic activity of plants grown on com-pacted and loose soils was studied twice a week fromthe age of 21 DAP to the age of 42 DAP. This pa-rameter was measured in three plants for each dateand treatment. At each date, the root–soil compart-ment of an individual plant was air-tightly sealed inorder to separate soil and root respiration from that ofthe ambient atmosphere (Fig. 1). The sealing was ob-tained using liquid silicone (Rhodorsil, base+catalyst,Aventis, France). An individual plant was placed in aplexiglass growth chamber with the same environmen-tal conditions mentioned previously. Measurementswere taken around noon and carbon assimilation byall plant leaves was determined by tracing the lineardecrease curve of CO2 concentration inside the cham-ber. The slope of the straight line represents CO2 up-take per minute for the whole plant. This value wasthen divided by leaf area and calculated in terms of�mol CO2 m−2 s−1. CO2 concentration was measuredusing a Li-6200 (Li-Cor, Lincoln, NE, USA) infraredgas analyser.
2.3. Carbon partitioning experiment
At each date, six plants, three for each treatment,were selected to be labelled with14C at 21, 35 and42 DAP. These plants were placed in the plexiglasschamber 1 day prior to the date of experiment. Eachcylinder was placed in another PVC container andshoots were sealed at the soil surface in a way to en-sure air tightness of belowground compartments (rootsand soil) as shown inFig. 1. After hardening of thesealant, columns were transferred to the growth cabi-net and appropriate connections for water supply andsoil aeration were set up.
14C was generated by adding sulphuric acid toNa2H14CO3. Specific activity inside the chamberwas constant (14 MBq g−1 C) in all experiments butlabelling duration varied according to assimilationrate. Labelling started at a CO2 concentration of360�mol mol−1 and continued until a concentrationof 120�mol mol−1 was reached (two-thirds of carbonpresent was assimilated). Preliminary experimentsindicated that, for short periods, carbon assimilationwas constant in this range. The labelling process var-ied with plant age and lasted for 120, 26 and 19 minfor 21-, 35- and 42-day-old plants, respectively. Soilwas continuously aerated during the experiment andthe chase period in order to avoid root anoxia.
Fig. 1. A schematic diagram illustrating a cross-section of culturepot and the air-tight separation of underground compartments.

154 A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161
At harvest, shoots were cut at the soil surface andleaves were separated from the stem. The soil columnwas pushed out of the cylinder and roots were care-fully excavated and gently washed in tap water to re-move adherent rhizosphere soil. Plant components andsoil-containing solution were placed immediately at−25◦C. They were then freeze-dried and the biomassof each compartment (shoots, roots and soil) was de-termined.
Total carbon content of each compartment was de-termined using a Carlo Erba CNS Na 1500 analyser(Carlo Erba Instrumentazione, Milan, Italy).14C inshoots and roots was detected using a Flo-One/Beta(Packard, Meriden, CT, USA) solid scintillation anal-yser coupled with the Carlo Erba analyser. Soil ra-dioactivity was determined by burning a soil sample ina Carmograph (Wösthoff, Germany) and the resulting14CO2 was collected and counted with a Packard liq-uid scintillator (Packard Instrument, Downers Grove,IL, USA).
2.4. Soil microbial biomass
Soil microbial biomass was determined for threeplants per treatment and per date (21, 35 and 42 DAP).After removal of the remaining root fractions, the soilof each pot was homogenised and two samples werekept at −25◦C. Soil moisture content at analysiswas close to 50% of field capacity. A pre-extractionwith 0.05 M K2SO4 solution was performed in orderto eliminate remaining root fractions that were notpicked up manually (Mueller et al., 1992). Micro-bial biomass was determined using the chloroformfumigation–extraction method described byVanceet al. (1987). Organic carbon content in soil extractswas determined by a DC 190 carbon analyser (DC190, Dohrmann, Germany). The following formulawas used to determine soil microbial biomass carbon:
SMBC = CorgFS− CorgCS
KC
where CorgFS is the organic carbon resulting from fu-migated soil, CorgCS the organic carbon resulting fromcontrol (non-fumigated) soil, andKC the conversionfactor (0.45).
Since the amount of carbon released from fumigatedsoil does not account for the total carbon of soil mi-
crobial biomass, a proportionality coefficient (KC) of0.45 was used throughout (Jenkinson and Ladd, 1981).
The experiments were performed according to acompletely randomised design. All the results wereanalysed by ANOVA using Statistica 5 (1995) soft-ware. Thereafter, means were compared by means ofLSD at the level ofP < 0.05 unless otherwise indi-cated.
3. Results
As expected, increasing soil bulk density decreasedsoil porosity. At 15% moisture content, air-filledporosity occupied 29% of the total volume for CStreatment versus 35% for LS treatment. However, bothaeration levels were not growth limiting according toWolkowski (1990).
3.1. Plant growth
Soil compaction greatly hampered root elongation.For 21-day-old plants (DAP), the mean lengths ofnodal and seminal root axes were 30% less in CS thanin LS (Table 1). However, between the ages of 35 and42 days mean length of seminal root axes levelled offin the LS treatment while it continued to grow in CS.Consequently, the difference between the two treat-ments decreased to slightly more than 10% at the fi-nal harvest. In the case of nodal roots, elongation ratein the LS treatment slowed down sharply between theages of 21 and 35 DAP, so that the gap in mean lengthbetween the two treatments narrowed earlier than wasthe case for seminal roots. The decrease in root lengthof plants grown on CS was accompanied by a lowerroot biomass. Compared with LS plants, the decreasein root biomass of CS treatment plants ranged from21 to 37%.
Leaf appearance rate was slower in CS than in thecontrol. However, the difference in the number of visi-ble leaves between the two treatments started to appearonly at 35 DAP and became significant at 42 DAP.This slow down in leaf appearance was reflected inother morphological parameters. At the first harvest,shoot dry weight in the CS was already lower than thatin LS, but the difference between the treatments wasonly significant at the last harvest (Fig. 2). Similarly,plant height in CS was lower than that in LS. However,

A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161 155
Table 1Morphological parameters of maize plants grown in two different soil bulk densities: 1.30 and 1.45 g cm−3 representing LS and CStreatments, respectively
Plant age (DAP)
21 35 42
1.30 g cm−3 1.45 g cm−3 1.30 g cm−3 1.45 g cm−3 1.30 g cm−3 1.45 g cm−3
Number of visible leavesa 7.00 7.00 11.0 10.6 13.8 a 13.3 bPlant height (cm) 76.4 67.3 130 a 119 b 156 a 144 bLeaf area (m2) 0.037 0.025 0.208 0.179 0.345 a 0.288 bMean length of nodal axes (cm) 32.1 a 22.8 b 34.4 31.8 42.7 37.5Mean length of seminal axes (cm) 46.5 a 32.9 b 59.1 a 44.8 b 61.1 a 54.7 bShoot biomass (g) 1.31 1.05 13.0 10.3 27.6 a 23.5 bRoot biomass (g) 0.37 0.29 2.15 a 1.27 b 5.35 a 3.90 bRoot:shoot ratio 0.281 0.278 0.166 a 0.123 b 0.194 a 0.166 b
a Mean comparisons are performed for each parameter between plants of same age using LSD test. When used, different letters indicatea significant difference (P < 0.05).
the means of the two treatments became statisticallydifferent only at 35 and 42 DAP (Table 1).
In younger plants, high soil resistance to penetrationdid not affect root:shoot ratio for the two treatments atthe first harvest. However, at later harvests the effect ofsoil compaction was more severe on roots decreasingtherefore root:shoot ratio.
3.2. Carbon assimilation
Net carbon assimilation, as used here, refers to to-tal photosynthetically fixed carbon accumulated in the
Fig. 2. Shoot dry matter accumulation in maize plants grown at two soil bulk densities: 1.30 g cm−3 (LS) and 1.45 g cm−3 (CS).
plant system, excluding any carbon respired by shootsand roots. Assimilation rate is expressed in micro-moles of CO2 fixed per unit leaf area (m2) per second.
Net carbon assimilation rate of plants from the LStreatment increased with plant age and peaked at theage of 39 DAP after which it levelled off (Fig. 3). Thesame trend was observed with the CS treatment, buta slight decrease in carbon assimilation rate occurredafter the age of 35 DAP which represented the peakfor this treatment.
In general, soil compaction reduced carbon assim-ilation per unit leaf area. The greatest effect of soil
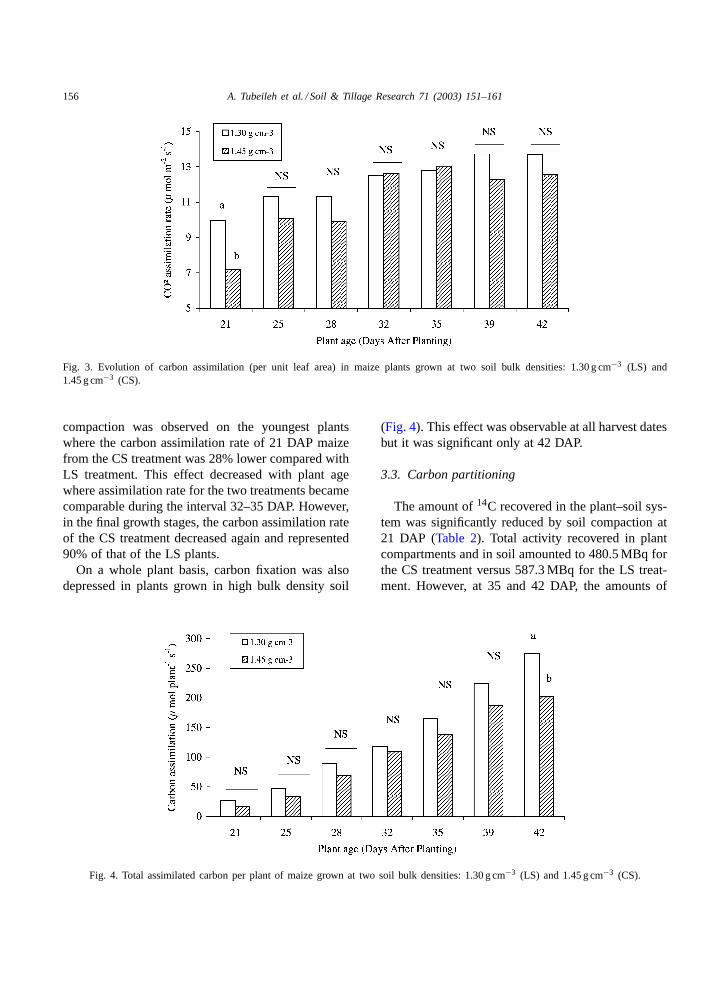
156 A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161
Fig. 3. Evolution of carbon assimilation (per unit leaf area) in maize plants grown at two soil bulk densities: 1.30 g cm−3 (LS) and1.45 g cm−3 (CS).
compaction was observed on the youngest plantswhere the carbon assimilation rate of 21 DAP maizefrom the CS treatment was 28% lower compared withLS treatment. This effect decreased with plant agewhere assimilation rate for the two treatments becamecomparable during the interval 32–35 DAP. However,in the final growth stages, the carbon assimilation rateof the CS treatment decreased again and represented90% of that of the LS plants.
On a whole plant basis, carbon fixation was alsodepressed in plants grown in high bulk density soil
Fig. 4. Total assimilated carbon per plant of maize grown at two soil bulk densities: 1.30 g cm−3 (LS) and 1.45 g cm−3 (CS).
(Fig. 4). This effect was observable at all harvest datesbut it was significant only at 42 DAP.
3.3. Carbon partitioning
The amount of14C recovered in the plant–soil sys-tem was significantly reduced by soil compaction at21 DAP (Table 2). Total activity recovered in plantcompartments and in soil amounted to 480.5 MBq forthe CS treatment versus 587.3 MBq for the LS treat-ment. However, at 35 and 42 DAP, the amounts of

A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161 157
Table 214C allocation (MBq per plant) among different compartments of a maize–soil system subjected to two different soil bulk densities: 1.30and 1.45 g cm−3 representing LS and CS treatments, respectively (percentage distributions are given in parentheses)
Plant age (DAP)
21 35 42
1.30 g cm−3 1.45 g cm−3 1.30 g cm−3 1.45 g cm−3 1.30 g cm−3 1.45 g cm−3
Shootsa 491 (83.4%) 387 (80.3%) 423 (76.4%) 465 (76.2%) 605 (81.6%) 559 (79.8%)Roots 62.8 (10.8%) a 29.2 (6.2%) b 93.0 (17.4%) a 46.4 (7.6%) b 77.8 (10.6%) 80.7 (11.5%)Total plant 553 (94.2%) 416 (86.5%) 516 (93.8%) 511 (83.8%) 683 (92.1%) 640 (91.3%)Soil 33.9 (5.8%) a 64.9 (13.5%) b 33.6 (6.2%) a 98.1 (16.2%) b 56.9 (7.9%) 60.4 (8.7%)
Total 587 a 481 b 550 609 739 700
a Mean comparisons are performed for each compartment between plants of same age using LSD test. When used, different lettersindicate a significant difference (P < 0.05) between percentage distributions.
14C accumulated in the plant–soil system showed nosignificant difference between treatments.
The proportion of14C retained in shoots did notvary, either with the increase in plant age or with in-creasing soil bulk density (Table 2). In the case of CS,this fraction accounted for 76–80% of total recoveredactivity versus 76–83% for LS. Meanwhile, increas-ing soil strength modified allocation patterns betweenbelowground compartments where we observed a de-crease in the proportion of recent assimilates in theroots. At the first and second harvests, the proportionof 14C retained in the roots of plants subjected to soilcompaction represented nearly half of that retained inroots of control plants. Nevertheless, at the last har-vest the proportion of root14C was the same for both
Fig. 5. Soil microbial biomass in pots grown with maize plants subjected to two soil bulk densities: 1.30 g cm−3 (LS) and 1.45 g cm−3 (CS).
treatments. This decrease in the proportion of labelledC remaining in the roots was accompanied by a sharpincrease in rhizodeposition at the same dates. At 21and 35 DAP, the proportion of14C recovered in thesoil of the CS treatment was more than doubled com-pared with LS. Once again, the difference between thetwo treatments decreased at the final harvest when theproportion of14C retained in the soil accounted for8.7 and 7.9% of total14C recovered in the CS and LStreatments, respectively.
3.4. Soil microbial biomass
Soil microbial biomass was significantly higher inCS than in LS (Fig. 5). This increase was observed

158 A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161
at all plant ages. Values for CS ranged from 79 to112�g C g−1 soil versus 47 to 66�g C g−1 soil in thecase of LS treatment. Moreover, it is interesting tonotice that this biomass increased with increasing plantage for both treatments.
4. Discussion
High soil bulk density decreased root length andbiomass and this effect persisted until the end of the ex-periment. However, the relative difference, in terms ofpercentage, between the two treatments decreased withplant age. Several authors have reported a 20–30% de-crease in root length of maize (Panayiotopoulos et al.,1994) and ryegrass (Houlbrooke et al., 1997) whenplants were grown on soil of high penetration resis-tance.
In addition to this effect on root growth, soil com-paction also affected aerial plant parts. The numberof visible leaves, plant height, leaf area and shootbiomass were all lower in the case of CS. However,plotting shoot height and shoot biomass against thenumber of visible leaves has shown that the reductionobserved in these two parameters is due, to a largeextent, to the delay in plant development estimatedfrom leaf appearance rate. Mechanical impedance ofroots has resulted in a slow down of leaf appearance inmaize (Groleau-Renaud et al., 1998), wheat and bar-ley (Young et al., 1997) grown in nutrient solution.Masle et al. (1990)suggested that there was a fac-tor that retarded shoot growth in plants encounteringcompact soil. More precisely,Young et al. (1997)at-tributed the reduction in leaf appearance rate to a hor-monal signal generated by impeded roots. This kindof message was also reported to affect the carbon as-similation rate, hence reducing the amount of assim-ilates available for plant growth (Tardieu, 1994). Soilcompaction induced a limitation in root growth thatwas reflected by a decrease in the root:shoot ratio. Itis now known that a limitation in root sink activity re-sults in the accumulation of carbohydrates in leaves,thus regulating the rate of carbon assimilation (Arp,1991). This root-to-shoot feedback is believed to oc-cur in the form of an increase in ABA concentrationin the shoot xylem sap (Tardieu et al., 1992; Turner,1997). Such an increase could raise stomatal resistancetherefore reducing carbon fixation.
Our results show that rhizodeposition was stimu-lated by the increase in soil bulk density. This is con-sistent with the results ofGroleau-Renaud et al. (1998)in hydroponic conditions andSauerbeck and Helal(1986)in soil. However, these latter authors obtainedtheir results using continuous labelling, which makesit difficult to quantify root exudation apart from rootturnover. Therefore, our study provides new informa-tion on the proportion of fixed carbon released intothe soil since root turnover may be considered mini-mal during the chase period. This increase in carboninput into the soil and its association with the sharpdecrease in root14C are noteworthy. A likely explana-tion is that soil compaction induced a sink limitationby hampering root growth leading to an accumulationof carbon already translocated to the roots. We there-fore suggest that unused carbon, i.e. that accumulatedin roots, will be released into the soil where it mayhave a beneficial role in the long term in improvinggrowth conditions and root penetration into the com-pacted layers. The stimulation of microbial biomassthat we observed in the CS provides further evidencesupporting this hypothesis. It has been demonstratedthat root exudates stimulate microbial activity and playan important role in improving soil stability and ag-gregation (Habib et al., 1990; Traore et al., 2000). Fur-thermore,Atwell (1993) suggested that sloughed-offcells and mucilaginous material covering roots facil-itated root progression by decreasing frictional resis-tance between soil and root tips. However, we couldnot detect a carbon accumulation in the roots due tothe relatively long chase period that permitted pho-tosynthates to be translocated from roots to the soil.Therefore, this hypothesis needs to be validated withvariable length chase periods in order to track the ex-act evolution of root and soil carbon. Nevertheless,this hypothesis may also be supported by the results ofAtwell (1990b)andMasle et al. (1990), who reportedthat high soil resistance induced a sink limitation thatresulted in the accumulation of soluble carbohydratesin roots.
After a chase period of 42 h, we could not ob-serve any modification in carbon partitioning betweenshoots on the one hand and belowground compart-ments on the other, although a reduction in root:shootratio was observed. This effect on the root:shoot ra-tio may be explained by the increase in carbon lossthrough exudation that we observed in the case of

A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161 159
CS, reducing therefore the quantity of carbon that re-mained in the roots.
The depressive effect of soil compaction on totalcarbon fixation became significant at 42 DAP. Indeed,this is mainly attributed to the significant decreasein leaf area observed at the same age (Table 1). Sur-prisingly, unlike these results, we did not observe asignificant effect of soil compaction on the total14Crecovered in the maize–soil system. This may be dueto the non-inclusion of rhizosphere respiration valuesthat could not be determined and which may be higherfor LS than CS. It should also be noted that the num-ber of replicates was probably not enough to show aneffect of soil compaction on carbon assimilation andpartitioning during labelling experiments given thehigh variability we observed. Detrimental effects ofsoil compaction on shoots and photosynthetic activityper unit leaf area diminished with plant age, as weobserved a decrease in the difference between the twotreatments in terms of percentage. It is important tonotice that this decrease with plant age was associatedwith a reduction in the gap in root length between the
Fig. 6. A conceptual model showing the relations between soil compaction and plant and soil parameters. Filled arrows (+ sign) indicatea positive effect of soil compaction on the concerned parameter while hollow arrows (− sign) indicate a negative effect.
LS and CS treatments. Although soil volume and soilwater content were not limiting, a differential wateruptake between the two treatments was likely. Com-pared to the roots of control plants, roots of plantsgrown on CS were shorter and less homogeneouslydistributed within the soil. This would affect wateraccessibility and the ability of roots to extract waterefficiently. A limitation in available soil water and/ora minor decrease in soil or root water potential couldhave incited roots to produce a signal of water short-age. This leads us to suppose that the reduction we ob-served in carbon assimilation rate and in shoot growthcould, at least partly, be due to water relations. Inother words, plants grown in CS were more sensitiveto water status than those grown in LS. This hypothe-sis, also suggested byPassioura (1988), is supportedby two points: the first is that in our study leavesof the CS treatment were visibly thicker and darkerthan those of the LS treatment. These symptoms usu-ally occur in plants subjected to dry conditions andimply a mechanism that reduces water loss throughtranspiration. However, further symptoms of water

160 A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161
shortage did not show up on our plants. Secondly,it has been shown that soil drying induced similareffects to those generated by high soil mechanicalresistance (Agnew and Carrow, 1985; Masle andPassioura, 1987; Bethenod et al., 1996). Similarly, ina field experiment,Tardieu et al. (1991)reported thatstomatal conductance was not affected by high soilresistance when plants were well-watered. Althoughthese authors did not provide information on root mor-phology under these conditions, they concluded thatthe observed increase in ABA concentration was dueto water relations rather than mechanical impedance.In spite of the fact that the information supplied byMasle and Passioura (1987)andTardieu et al. (1991)supports our second hypothesis about water relations,it is still necessary to study the water potential ofsoil, roots and leaves under our conditions in order toestablish the relation between water status and carbonassimilation.
5. Conclusion
Our experimental results may be synthesized inthe form of a conceptual model (Fig. 6) that illus-trates the relations among all the studied parametersin order to have a global approach regarding the ef-fects of soil compaction on plant and soil microbialbiomass.
Soil compaction reduced root and shoot growth andincreased carbon input into the soil and soil micro-bial biomass. Our results show that compaction effectspersist up to tassel initiation although they decreasein terms of percentage. Therefore, these results shouldnot be extrapolated for the whole growth period ofmaize before being validated on older plants. More-over, it seemed that the gap we observed in mean rootlength between the treatments at young plant ages nar-rowed just before tassel initiation, hence reducing po-tential negative effects of insufficient water supply onplant growth at this sensitive stage. Nevertheless, asshown inFig. 4, total carbon fixation per plant at laterstages was still lower in the CS plants as compared tothe LS plants. This indicates that plants grown at highsoil bulk density might not be able to compensate forthe delay in plant growth (particularly shoot) that tookplace at early stages, so a reduction in crop yield maybe expected.
Acknowledgements
The first author would like to thank Mrs. L. Zaidifor her useful help during carbon assimilation exper-iments. Thanks are also due to A. Gautier, P. Mar-chal, M. Philbert, and D. Thiery for their technicalassistance. The first author is grateful to the Frenchgovernment for granting him a Ph.D. scholarship inFrance.
References
Abdalla, A.M., Hettiaratchi, D.R.P., Reece, A.R., 1969. Themechanics of root growth in granular media. J. Agric. Eng.Res. 14, 236–248.
Agnew, M.L., Carrow, R.N., 1985. Soil compaction and moisturestress preconditioning in Kentucky bluegrass. II. Stomatalresistance, leaf water potential, and canopy temperature. Agron.J. 77, 878–884.
Arp, W.J., 1991. Effects of source–sink relations on photosyntheticacclimation to elevated CO2. Plant Cell Environ. 14, 869–875.
Asady, G.H., Smucker, J.M., 1989. Compaction and root modi-fications of soil aeration. Soil Sci. Soc. Am. J. 53, 251–254.
Atwell, B.J., 1990a. The effect of soil compaction on wheat duringearly tillering. I. Growth, development and root structure. NewPhytol. 115, 29–35.
Atwell, B.J., 1990b. The effect of soil compaction on wheatduring early tillering. III. Concentration of cell constituents.New Phytol. 115, 37–41.
Atwell, B.J., 1993. Response of roots to mechanical impedance.Environ. Exp. Bot. 33, 27–40.
Bethenod, O., Tardieu, F., Katerji, N., 1996. Relationship betweennet photosynthetic rate and stomatal conductance in leaves offield-grown maize subjected to soil compaction or soil drying.Photosynthetica 32, 367–379.
Boeuf-Tremblay, V., Plantureux, S., Guckert, A., 1995. Influenceof mechanical impedance on root exudation of maize seedlingsat two development stages. Plant Soil 172, 279–287.
Canarache, A., Colibas, I., Colibas, M., Horobeanu, I., Patru,V., Simota, H., Trandafirescu, T., 1984. Effect of inducedcompaction by wheel traffic on soil physical properties andyield of maize in Romania. Soil Till. Res. 4, 199–213.
Ekwue, E.I., Stone, R.J., 1995. Irrigation scheduling for sweetmaize relative to soil compaction conditions. J. Agric. Eng.Res. 62, 85–94.
Grath, T., Håkansson, I., 1992. Effects of soil compaction ondevelopment and nutrient uptake of peas. Swed. J. Agric. Res.22, 13–17.
Groleau-Renaud, V., Plantureux, S., Guckert, A., 1998. Influenceof plant morphology on root exudation of maize subjectedto mechanical impedance in hydroponic conditions. Plant Soil201, 231–239.
Habib, L., Morel, J.-L., Guckert, A., Plantureux, S., Chenu, C.,1990. Influence of root exudates on soil aggregation. Symbiosis9, 87–91.

A. Tubeileh et al. / Soil & Tillage Research 71 (2003) 151–161 161
Hoagland, D.R., Arnon, I.R., 1938. The water culture methodfor growing plants without soil. University of CaliforniaExperimental Station, Circular 347.
Houlbrooke, D.J., Thom, E.R., Chapman, R., McLay, C.D.A.,1997. A study of the effects of soil bulk density on root andshoot growth of different ryegrass lines. NZ J. Agric. Res. 40,429–435.
Jenkinson, D.S., Ladd, J.N., 1981. Microbial biomass in soil:measurement and turnover. In: Paul, E.A., Ladd, J.N.(Eds.), Soil Biochemistry, vol. 5. Dekker, New York, USA,pp. 415–471.
Kayombo, B., Lal, R., Mrema, G.C., Jensen, H.E., 1991.Characterizing compaction effects on soil properties and cropgrowth in southern Nigeria. Soil Till. Res. 21, 325–345.
Liljeroth, E., Kuikman, P., Van Veen, J.A., 1994. Carbontranslocation to the rhizosphere of maize and wheat andinfluence on the turnover of native soil organic matter atdifferent soil nitrogen levels. Plant Soil 161, 233–240.
Logsdon, S.D., Reneau Jr., R.B., Parker, J.C., 1987. Corn seedlingroot growth as influenced by soil physical properties. Agron.J. 79, 221–224.
Masle, J., Passioura, J.B., 1987. The effect of soil strength onthe growth of young wheat plants. Aust. J. Plant Physiol. 14,643–656.
Masle, J., Farquhar, G.D., Gifford, R.M., 1990. Growth and carboneconomy of wheat seedlings as affected by soil resistance topenetration and ambient partial pressure of CO2. Aust. J. PlantPhysiol. 17, 465–487.
Mueller, T., Joergensen, R.G., Meyer, B., 1992. Estimationof soil microbial biomass C in the presence of livingroots by fumigation–extraction. Soil Biol. Biochem. 24, 179–181.
Oussible, M., Crookston, R.K., Larson, W.E., 1992. Subsurfacecompaction reduces the root and shoot growth and grain yieldof wheat. Agron. J. 84, 34–38.
Panayiotopoulos, K.P., Papadopoulou, C.P., Hatajiionnidou, A.,1994. Compaction and penetration resistance of an Alfisol and
Entisol and their influence on root growth of maize seedlings.Soil Till. Res. 31, 323–337.
Passioura, J.B., 1988. Root signals control leaf expansion in wheatseedlings growing in dry soil. J. Plant Physiol. 15, 687–693.
Qian, J.H., Doran, J.W., Walters, D., 1997. Maize plantcontributions to root zone available carbon and microbialtransformations of nitrogen. Soil Biol. Biochem. 29, 1451–1462.
Sauerbeck, D.R., Helal, H.M., 1986. Plant root development andphotosynthate consumption depending on soil compaction. In:Proceedings of the Transactions of the XIII Congress of theInternational Society of Soil Science, pp. 948–949.
Tardieu, F., 1994. Growth and functioning of roots and of rootsystems subjected to soil compaction. Towards a system withmultiple signalling? Soil Till. Res. 30, 217–243.
Tardieu, F., Katerji, N., Bethenod, O., Zhang, J., Davies, W.J.,1991. Maize stomatal conductance in the field: its relationshipwith soil and plant water potentials, mechanical constraints andABA concentration in xylem sap. Plant Cell Environ. 14, 121–126.
Tardieu, F., Zhang, J., Katerji, N., Bethenod, O., Palmer, S., Davies,W.J., 1992. Xylem ABA controls the stomatal conductance offield-grown maize subjected to soil compaction or soil drying.Plant Cell Environ. 15, 193–197.
Traore, O., Groleau-Renaud, V., Plantureux, S., Tubeileh, A.,Boeuf-Tremblay, V., 2000. Effect of natural and modelled rootexudates on soil structure. Eur. J. Soil Sci. 51, 575–581.
Turner, N.C., 1997. Further progress in crop water relations. Adv.Agron. 58, 293–338.
Vance, E.D., Brookes, P.C., Jenkinson, D.S., 1987. An extractionmethod for measuring soil microbial biomass C. Soil Biol.Biochem. 19, 703–707.
Wolkowski, R.P., 1990. Relationship between wheel-traffic-inducedsoil compaction, nutrient availability, and crop growth: a review.J. Prod. Agric. 3, 460–469.
Young, I.M., Montagu, K., Conroy, J., Bengough, A.G., 1997.Mechanical impedance of root growth directly reduces leafelongation rates of cereals. New Phytol. 135, 613–619.