ecological costs of feral predator control: foxes and rabbits
TRANSCRIPT

Allen Press
Ecological Costs of Feral Predator Control: Foxes and RabbitsAuthor(s): Peter B. Banks, Christopher R. Dickman and Alan E. NewsomeSource: The Journal of Wildlife Management, Vol. 62, No. 2 (Apr., 1998), pp. 766-772Published by: Wiley on behalf of the Wildlife SocietyStable URL: http://www.jstor.org/stable/3802353 .
Accessed: 16/09/2013 18:56
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
Wiley, Wildlife Society, Allen Press are collaborating with JSTOR to digitize, preserve and extend access toThe Journal of Wildlife Management.
http://www.jstor.org
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

766 OVERFLIGHTS AND CARIBOU * Maier et al. J. Wildl. Manage. 62(2):1998
versity of Alaska Fairbanks, Fairbanks, Alaska, USA.
RUSSELL, D. E., A. M. MARTELL, AND W. A. C. NIx- ON. 1993. Range ecology of the Porcupine Car- ibou Herd in Canada. Rangifer, Special Issue 8: 1-167.
SAS INSTITUTE. 1985. SAS user's guide: statistics. Version 5 edition. SAS Institute, Cary, North Car- olina, USA.
SIH, A. 1984. The behavioral response race between predator and prey. American Naturalist 123:143- 150.
SKOGLAND, T. 1984. Wild reindeer foraging-niche or- ganization. Holarctic Ecology 7:345-379.
STOCKWELL, C. A., G. C. BATEMAN, AND J. BERGER. 1991. Conflicts in national parks: a case study of helicopters and bighorn sheep time budgets at the Grand Canyon. Biological Conservation 56: 317-328.
VALKENBURG, P. 1994. Investigation of regulating and limiting factors in the Delta caribou herd. Alaska Department of Fish and Game, Federal Aid in Wildlife Restoration, Project W-24-2, Progress Report.
, AND J. L. DAVIS. 1985. The reaction of car- ibou to aircraft: a comparison of two herds. Pro- ceedings of the North American Caribou Work- shop 1:7-9.
WEISENBERGER, M. E., P. R. KRAUSMAN, M. C. WAL-
LACE, D. W. DEYOUNG, AND O. E. MAUGHAN. 1996. Effects of simulated jet aircraft noise on heart rate and behavior of desert ungulates. Jour- nal of Wildlife Management 60:52-61.
WHITE, R. G. 1983. Foraging patterns and their mul- tiplier effects on productivity of northern ungu- lates. Oikos 40:377-384.
- , B. R. THOMSON, T. SKOGLAND, S. J. PERSON, D. E. RUSSELL, D. F. HOLLEMAN, AND J. R. LUICK. 1975. Ecology of caribou at Prudhoe Bay, Alaska. Pages 151-201 in J. Brown, editor. Ecolog- ical investigations of the tundra biome in the Prud- hoe Bay region. Biological Papers of the University of Alaska Fairbanks, Fairbanks, Alaska, USA.
Received 6 June 1996. Accepted 13 October 1997. Associate Editor: Hobbs.
ECOLOGICAL COSTS OF FERAL PREDATOR CONTROL: FOXES AND RABBITS
PETER B. BANKS,' Institute of Wildlife Research, School of Biological Sciences, A08, University of Sydney, NSW 2006, Australia CHRISTOPHER R. DICKMAN, Institute of Wildlife Research, School of Biological Sciences, A08, University of Sydney, NSW
2006, Australia ALAN E. NEWSOME, Commonwealth Scientific and Industrial Research Organization Division of Wildlife and Ecology, P.O. Box
84 Lyneham, ACT 2602, Australia
Abstract: We used a predator removal experiment to examine the role of red fox (Vulpes vulpes) predation in suppressing rabbit (Oryctolagus cuniculus) population growth in Namadgi National Park in southeastern Australia. At 2 sites, fox abundance was reduced with a 1080 poisoning campaign maintained over 18 months. The responses of rabbit populations in these fox-reduced sites were compared to 2 other sites where fox
populations remained intact. In the 2 removal sites, rabbit populations grew to 6.5 and 12.0 times their initial
population size within 18 months. In the untreated sites, rabbit populations showed very small population increases over the same period. The experiment demonstrated that 1 introduced pest species suppressed the
population growth of another pest species. As fox removal was initially planned to protect native fauna threat- ened by fox predation, the response of the rabbits represents a serious ecological cost of fox control.
JOURNAL OF WILDLIFE MANAGEMENT 62(2):766-772
Key words: Australia, European rabbit, Oryctolagus cuniculus, predator effect, predator-prey, red fox, re- moval experiment, vertebrate pests, Vulpes vulpes.
The red fox (hereafter, fox) was introduced into Australia in the 1850s and quickly became a major pest species across the southern half of the continent (Rolls 1969). This species is im-
plicated in the decline or extinction of 23 spe- cies of native small- and medium-sized mam- mals and currently is listed as a significant threat to 30 species of endangered or vulnera- ble mammals and birds (Saunders et al. 1995). The fox also is a significant predator of domestic stock, in particular sheep and lambs, and can 1 E-mail: [email protected]
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

J. Wildl. Manage. 62(2):1998 IMPACT OF FOX PREDATION ON RABBITS * Banks et al. 767
kill up to 30% of the lambs produced in some areas (Lugton 1993). Hence, control of foxes has become a priority for nature conservation
agencies and the agricultural industry in Aus- tralia.
There is a close relation between the fox and another introduced vertebrate pest in Australia, the European rabbit (hereafter, rabbit). Where rabbits are available, they are consistently the
major prey item of foxes, composing up to 100% of the fox diet in some areas (Newsome et al. 1997). The spread of the rabbit less than a decade before the introduction of the fox no doubt assisted spread of the latter species (Rolls 1969); today, the distributions of these 2 pests closely overlap (Wilson et al. 1992). Rabbits ap- parently support fox populations at elevated lev- els, which then affect native species via oppor- tunistic predation (Christensen 1980, Lundie-
Jenkins et al. 1993). Direct environmental dam-
age caused by the rabbit includes competition with many native species of herbivores as well as domestic stock, and severe land degradation via overgrazing and construction of warrens (Williams et al. 1995). Indeed, the rabbit is rec-
ognized as Australia's number 1 vertebrate pest. There is abundant anecdotal evidence that
fox predation suppresses growth of rabbit pop- ulations, but there is limited experimental sup- port. In arid New South Wales, Wood (1981) estimated that foxes took up to 75% of newly emergent juvenile rabbits and caused a mortal-
ity rate of >85% for rabbits <9 months old. Foxes also prey heavily on adult rabbits in arid environments (Martenz 1971, Bayly 1978, Cat-
ling 1988), and Wood (1981) suggested that
density-dependent predation by foxes may limit rabbit population growth. This hypothesis was confirmed by Newsome et al. (1989) in a fox removal experiment demonstrating predator suppression of rabbit populations, and Pech et al. (1992) showed further that predators regu- lated rabbit numbers at low densities through density-dependent predation. In arid environ- ments, the presence of foxes mitigates the ef- fects of rabbits because foxes slow the recovery of rabbit populations after drought (Newsome et al. 1989).
In temperate areas where environments are less variable, the role of predation in suppres- sion of rabbit numbers is less clear. Rabbit pop- ulation dynamics in cooler climates are strongly influenced by rainfall (Gilbert et al. 1987) and also by nutrient levels (Andrewartha and Birch
1984), but populations do not fluctuate as dra-
matically as in arid areas. Mortality of juveniles in temperate areas is around 30%, which is low- er than arid areas, but appears to be higher in areas with predators (Richardson and Wood 1982). Rabbits remain the staple prey for foxes (Newsome et al. 1997), but the effect of this
predation is not known, and it is not clear if this
predation suppresses rabbit population growth or is compensatory.
In this paper, we report the responses of rab- bit populations to a fox control program de-
signed to protect native fauna in a subalpine area. Hence, we measure the responses of rab- bit populations to fox control as a by-product of efforts to protect native wildlife, and discuss the role of fox predation in the suppression of this
significant vertebrate pest.
STUDY AREA The study was conducted in Namadgi Na-
tional Park, an area of subalpine forest and re-
possessed open grassy farmland in the Brinda- bella Ranges, 50 km south of Canberra, Austra- lia (35040'S, 14905'E). We chose 4 valleys, each
approximately 10 km2 in area and separated by 7-22 km of mostly sclerophyllous forest. We as- sumed these distances provided some barrier to fox movement, and thus independence among sites. Rabbits were most abundant in the eco- tone between the cleared valley and the rem- nant timbered areas; foxes were prevalent throughout. The main mammalian prey of foxes is rabbits (40% by occurrence) and kangaroos (Macropus giganteus; <545%), with native small mammals, invertebrates, and vegetation com-
posing the remainder (P. Banks, unpublished data).
METHODS Reducing Fox Numbers
We reduced fox numbers at the 2 removal sites via 1080 baiting. We buried approximately 60 commercial FOXOFF baits (35 g each) in and around the 2 valleys; each bait contained 0.3 mg of 1080 (sodium monofluoroacetate). To
target transient and reinvading animals, we
placed baits at 200-m intervals on the main ac- cess roads through the sites and on all roads in a 5-km radius around the sites. The baits were buried 10 cm beneath a pile of loose soil that attracted passing foxes.
We initially checked baits over a 10-day pe- riod and replaced any baits taken. This schedule
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

768 IMPACT OF FOX PREDATION ON RABBITS * Banks et al. J. Wildl. Manage. 62(2):1998
was designed to reduce fox numbers as quickly as possible. After this period, fresh baits were
placed monthly, checked after 7 days, and re- moved if not taken. Baiting began in July 1993 and continued to January 1995. Despite a clear decline in local fox activity, baits were continu-
ally taken by reinvading foxes, and individual foxes likely took multiple baits before they died.
Monitoring Fox Populations To ensure that fox numbers were reduced in
the removal sites, we used a combination of
spotlighting, uptake of nontoxic baits, and scat counts to index the abundance of foxes in each of the 4 areas. We conducted spotlighting counts of foxes (animals/km) for 3-4 consecu- tive nights each month along 5-km transects at each site. Any foxes seen while spotlighting in the removal sites were shot in the head with a low velocity 0.22-calibre rifle that made mini- mal noise. Less than 25% of the animals seen were shot, because many animals were too far
away or too difficult to shoot. Overall, however, 1080 baiting was the most effective control
technique. Once fox numbers had been sub-
stantially reduced (late 1993), baiting was used as the primary method of fox control.
We also used nontoxic baits to monitor fox numbers. Prior to using poison baits, we buried 30 nontoxic baits at 200-m intervals along roads at each site. The number of baits removed after 2 days was then used as an index of fox abun- dance. We conducted this monitoring twice at all 4 sites, 12 and 18 months after fox removal
began. The techniques used to monitor fox numbers at the removal sites were identical to those used in the nonremoval sites and are di-
rectly compared to nonremoval sites in the re- sults. We also counted scats and removed them from 3-km transects established along roadsides at each site as another index of fox abundance. We made collections bimonthly because few scats were present in the removal sites after the removal campaign began.
Monitoring Rabbit Populations We monitored changes in rabbit abundance
associated with fox removal via spotlight counts (animals/km) on 5-km transects established at each site (Newsome et al. 1989, Pech et al. 1992). Spotlighting is an acceptable and reliable index of rabbit abundance compared to other census methods such as counts of active warren entrances (Pech et al. 1992), and was consid-
ered the technique best suited for our local con- ditions and requirements. Although spotlighting may miss juvenile animals that remain close to the warren (Newsome et al. 1989), counts of active warren entrances would have underesti- mated the growth in rabbit numbers at the re- moval sites because many animals apparently were living on the surface.
To spot rabbits, we used a 100 W spotlight from the top of a vehicle travelling <5 km/hr. We drove along spotlight transects on 3-4 con- secutive nights/month at or near the new moon. In preliminary surveys, we saw more rabbits on dark, moonless nights compared to moonlit
nights (Kolb 1992). Surveys were not conducted on rainy or on windy nights, which reduced weather-related variability in the data (Williams et al. 1995). Spotlighting commenced 1 hr after dusk to coincide with the peak in activity of rab- bits (Williams et al. 1995). We conducted sur-
veys for foxes and rabbits on the same nights, but if foxes were shot at the removal sites, we omitted from the dataset the survey for rabbit numbers on that night, due to potential distur- bance. We conducted spotlight censuses of rab- bit numbers every month from May 1993 to
February 1995.
RESULTS Fox Removal
The removal campaign began in July 1993 and in 6 months resulted in a decline of foxes from 2.8 and 3.4/km to <0.5/km at the 2 re- moval sites (Fig. 1A). After 12 months, this den-
sity was reduced to near zero. Over the same
period, numbers in the untreated sites re- mained relatively steady, with a large peak as- sociated with the appearance of cubs at 1 site, followed by some declines during the second winter (Fig. 1B). Initial fox densities at the con- trol sites were 2.0/km and less than at the re- moval sites (2.5-3.0 km). However, at 12 and 18 months of fox control, the nontoxic bait indices of fox activity showed substantially more baits taken in the nonremoval sites compared to re- moval sites. Between 90 and 94% of baits were taken in 2 nights at the nonremoval sites com- pared to 0 and 5% at the removal sites. Simi- larly, signs of foxes were substantially reduced at the removal sites, where <3 scats/km were found along transects compared to about 20 scats/km of transect at the nonremoval sites. These data confirm the efficacy of the removal
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions
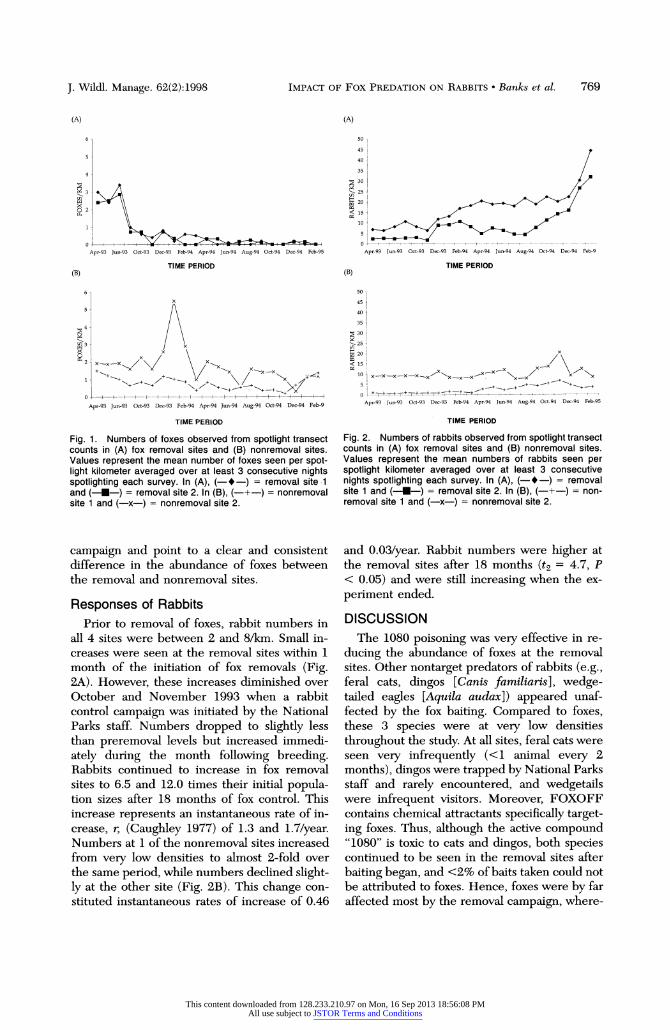
J. Wildl. Manage. 62(2):1998 IMPACT OF FOX PREDATION ON RABBITS * Banks et al. 769
(A)
02
4
Apr-93 Jun-93 Oct-93 Dec-93 Feb-94 Apr-94 Jun-94 Aug-94 Oct-94 Dec-94 Feb-95
TIME PERIOD (B)
6
x3
0 x
. ?/ +? ?
- I I I I
Apr-93 Jun-93 Oct-93 Dec-93 Feb-94 Apr-94 Jun-94 Aug-94 Oct-94 Dec-94 Feb-9
TIME PERIOD
Fig. 1. Numbers of foxes observed from spotlight transect counts in (A) fox removal sites and (B) nonremoval sites. Values represent the mean number of foxes seen per spot- light kilometer averaged over at least 3 consecutive nights spotlighting each survey. In (A), (-*-) = removal site 1 and (----) = removal site 2. In (B), (-+-) = nonremoval site 1 and (-x-) = nonremoval site 2.
campaign and point to a clear and consistent difference in the abundance of foxes between the removal and nonremoval sites.
Responses of Rabbits Prior to removal of foxes, rabbit numbers in
all 4 sites were between 2 and 8/km. Small in- creases were seen at the removal sites within 1 month of the initiation of fox removals (Fig. 2A). However, these increases diminished over October and November 1993 when a rabbit control campaign was initiated by the National Parks staff. Numbers dropped to slightly less than preremoval levels but increased immedi-
ately during the month following breeding. Rabbits continued to increase in fox removal sites to 6.5 and 12.0 times their initial popula- tion sizes after 18 months of fox control. This increase represents an instantaneous rate of in- crease, r, (Caughley 1977) of 1.3 and 1.7/year. Numbers at 1 of the nonremoval sites increased from very low densities to almost 2-fold over the same period, while numbers declined slight- ly at the other site (Fig. 2B). This change con- stituted instantaneous rates of increase of 0.46
(A)
50 45 i
35
S30 S25
S20i
5
0
Apr-93 Jun-93 Oct-93 Dec-93 Feb-94 Apr-94 Jun-94 Aug-94 Oct-94 Dec-94 Feb-9
TIME PERIOD (B)
50 1
45
40
35 30 .25 S20 x
t i ,-X-X-.X-
X-X X - - X X 5
0 + -?
" -'--+-7--+-r - -- i-+ ---7 --' +-''+-t- -•-i '• .......--- • ... --- • - ... ...• - -
Apr-93 Jun-93 Oct-93 Dec-93 Feb-94 Apr-94 Jun-94 Aug-94 Oct-94 Dec-94 Feb-95
TIME PERIOD
Fig. 2. Numbers of rabbits observed from spotlight transect counts in (A) fox removal sites and (B) nonremoval sites. Values represent the mean numbers of rabbits seen per spotlight kilometer averaged over at least 3 consecutive nights spotlighting each survey. In (A), (-*-) = removal site 1 and (--U--) = removal site 2. In (B), (-+-) = non- removal site 1 and (-x-) = nonremoval site 2.
and 0.03/year. Rabbit numbers were higher at the removal sites after 18 months (t2 = 4.7, P < 0.05) and were still increasing when the ex-
periment ended.
DISCUSSION The 1080 poisoning was very effective in re-
ducing the abundance of foxes at the removal sites. Other nontarget predators of rabbits (e.g., feral cats, dingos [Canis familiaris], wedge- tailed eagles [Aquila audax]) appeared unaf- fected by the fox baiting. Compared to foxes, these 3 species were at very low densities
throughout the study. At all sites, feral cats were seen very infrequently (<1 animal every 2 months), dingos were trapped by National Parks staff and rarely encountered, and wedgetails were infrequent visitors. Moreover, FOXOFF contains chemical attractants specifically target- ing foxes. Thus, although the active compound "1080" is toxic to cats and dingos, both species continued to be seen in the removal sites after
baiting began, and <2% of baits taken could not be attributed to foxes. Hence, foxes were by far affected most by the removal campaign, where-
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

770 IMPACT OF FOX PREDATION ON RABBITS * Banks et al. J. Wildl. Manage. 62(2):1998
by the abundance of foxes was the only clear difference between removal and nonremoval sites that was attributable to the poisoning.
We conclude that fox predation suppressed population growth of rabbits in this subalpine environment. In the removal sites, rabbit pop- ulations increased at least 6.5 times within 18 months following fox control. Further, if rabbits had not been reduced by National Parks Service control efforts, the population growth could have been significantly larger. In the areas where foxes remained, the numbers of rabbits remained steady and grew only 10% over the same period. Much of the population growth occurred in the latter half of the experiment, which was during the second breeding season. After pasture improvement in arid New South Wales, rabbit populations that were reduced by drought increased 11.7 times after 14 months of predator control at a fox-removal site com-
pared to 2.8 times at the nonremoval site (New- some et al. 1989). This initial response was much quicker than occurred in the present study where populations increased 1.5 times within just 6 weeks. At Namadgi, where there were no major fluctuations in environmental conditions during the study, foxes appeared to
strongly suppress rabbit population growth. There has been little other experimental
work investigating the role of predator sup- pression on rabbit populations. Trout and Tit- tensor (1989) reviewed an abundance of an- ecdotal evidence, analyzing patterns of rabbit abundance under varying degrees of predator control in rural England. They concluded that
predators did play a role in the suppression of rabbit numbers. However, rabbit populations continued to fluctuate and, in some instances, only the magnitude of fluctuation was affected
by predation. Experimental evidence provided by Newsome et al. (1989) and the present study confirms these predictions. At Namadgi, rabbit numbers remained stable over the study period at the sites with foxes, and there were
only small and rapidly subsiding rises during the breeding season. However, it was not pos- sible to confirm that this small decline was due to fox predation.
MANAGEMENT IMPLICATIONS
The fox control program outlined here was initiated primarily to quantify the direct effect of fox predation on native mammal species. However, a by-product of this control was an
increase in another vertebrate pest species and creation of potentially greater environmental
problems than in the areas where foxes re- mained. The environmental damage caused by rabbits can be severe (see reviews by Sumption and Flowerdew 1985, Williams et al. 1995). In
particular, overgrazing can lead to increased
competitive pressure upon the very native her- bivores (Cooke 1993, Lundie-Jenkins et al. 1993) that the fox control program is designed to protect.
Foxes may have a role to play in management of rabbit populations, but this role needs quan- tification (Williams et al. 1995). Rabbit recovery following control programs might be signifi- cantly faster if predators are removed. There- fore, control techniques with less potential for
secondary poisoning of predators may be more effective in the long-term management of rab- bits. Clearly, if foxes had not been removed, the rabbit population would not have increased to the observed levels in such a short period. In the absence of foxes, the rabbit control program initiated in November 1993 was essentially re- dundant within 3 months; after 18 months, the rabbit problem was 5 times worse. This re-
sponse represents a clear and potentially great ecological cost of fox control.
This scenario is likely to be repeated in many areas where rabbits and foxes coexist, in partic- ular where foxes are controlled to relieve pre- dation pressure on alternate or opportunistically eaten native species of prey (Lundie-Jenkins et al. 1993). Where rabbits are present, no native
prey species has been taken more frequently than rabbits, and there is some evidence that foxes may take rabbits selectively (Brunner et al. 1975, Seebeck 1978). Although native prey species that are very infrequent prey items for foxes may still be limited by this low level of
predation, any significant population growth re-
sulting from fox control will likely be slower and less obvious than the response by rabbits. Re-
covery times for native species are likely longer than for rabbits because native species generally have lower initial densities (e.g., Kinnear et al. 1988) and lower intrinsic rates of increase com-
pared to rabbits (Dickman 1996), and there is a greater likelihood that predation pressure will be much higher and potentially more limiting for rabbits than native species. As a result, con- trol of foxes will result in a rabbit problem long before native species show signs of significant
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

J. Wildl. Manage. 62(2):1998 IMPACT OF Fox PREDATION ON RABBITS * Banks et al. 771
recovery. This inevitable result is an important consideration for land managers.
Overall, this study shows that foxes can sup- press populations of other vertebrate pests and have a role to play in effective management of rabbits (Newsome 1990). Protection of native
species threatened directly by fox predation via control of foxes may therefore result in signifi- cant and potentially large increases in rabbit numbers. This unwarranted ecological cost of fox control must be weighed against the benefits to native prey species. Where fox control is im-
perative, integrated programs that control foxes and rabbits simultaneously will be necessary, and the economic costs of rabbit control must be included in costs of removing foxes to pro- tect native fauna. Alternatively, where potential native prey for foxes are not threatened, long- term control of both rabbits and foxes might best be achieved with rabbit control techniques that do not immediately affect foxes (e.g., war- ren destruction). This approach would allow for
persistence of some foxes to maintain predation pressure upon rabbit populations and suppress rabbit population growth for an extended peri- od (King et al. 1981).
ACKNOWLEDGMENTS This project was funded by the Australian
Alps Liaison Committee, Namadgi National Park, and the Institute of Wildlife Research to whom we express much gratitude. Many people assisted in this project and endured cold nights spotlighting under trying conditions to collect this data, and we offer many thanks to all. Spe- cial thanks for multiple trips to J. M. Tipping, W. E. Maitz, A. N. Banks, B. Tamayo, A. Ried, R. J. Spencer, J. Walker, P. Ostrowski, L. McLachlan, and S. Parker. D. F Hochuli, and G. H. MacNaught offered constructive com- ments on earlier drafts. Thanks also to the staff of Namadgi for their support and encourage- ment, and to D. Moore, G. Toddkill, A. Huxley, W. Sayer, R. Watchorn, C. Richardson, and B. Terril for their advice and assistance with fox control.
LITERATURE CITED ANDREWARTHA, H. G., AND L. C. BIRCH. 1984. The
ecological web. Chicago University Press, Chica- go, Illinois, USA.
BAYLY, C. P 1978. A comparison of the diets of red fox and feral cats in an arid environment. South Australian Naturalist 53:20-28.
BRUNNER, H., J. W. LLOYD, AND B. J. COMAN. 1975.
Fox scat analysis in a forest park in southeastern Australia. Australian Wildlife Research 2:147- 157.
CATLING, P. C. 1988. Similarities and contrasts in the diets of foxes, Vulpes vulpes, and cats, Felis catus, relative to fluctuating prey population and drought. Australian Wildlife Research 15:307- 317.
CAUGHLEY, G. 1977. Analysis of vertebrate popula- tions. John Wiley & Sons, London, United King- dom.
CHRISTENSEN, P. E. S. 1980. A sad day for native fauna. Forest Focus 23:3-12.
COOKE, B. D. 1993. Recent research on rabbit con- trol in South Australia. Xanthopus 149:7-8.
DICKMAN, C. R. 1996. Impact of feral cats in Aus- tralia. Australian Nature Conservation Agency Publishers, Canberra, Australia.
GILBERT, N., K. MYERS, B. D. COOKE, J. D. DUNS- MORE, P. J. FULLAGAR, J. A. GIBB, D. R. KING, I. PARER, S. H. WHEELER, AND D. H. WOOD. 1987. Comparative dynamics of Australasian rab- bit populations. Australian Wildlife Research 14: 491-503.
KING, D. R., A. J. OLIVER, AND R. J. MEAD. 1981. Bettongia and flouroacetate: a role for 1080 in fauna management. Australian Wildlife Research 8:529-536.
KINNEAR, J. E., M. L. ONUS, AND R. N. BROMILOW.
1988. Fox control and rock wallaby population dynamics. Australian Wildlife Research 15:435- 450.
KOLB, H. H. 1992. The effects of moonlight on ac- tivity in the wild rabbit (Oryctolagus cuniculus). Journal of Zoology, London 228:661-665.
LUGTON, I. 1993. Fox predation on lambs. Pages 17- 26 in D. A. Hucker, editor. Proceedings of the Australian Sheep Veterinary Society. Australian Veterinary Association, Gold Coast, Australia.
LUNDIE-JENKINS, G., L. K. CORBETT, AND C. M.
PHILLIPS. 1993. Ecology of the rufous hare-wal- laby, Lagorchestes hirsutus Gould (Marsupialia: Macropodidae) in the Tanami Desert, Northern Territory, III. Interactions with introduced mam- mal species. Wildlife Research 20:495-511.
MARTENZ, P. N. 1971. Observations of the food of the fox (Vulpes vulpes L.) in an arid environment. Commonwealth Scientific and Industrial Re- search Organisation Wildlife Research 16:15-17.
NEWSOME, A. E. 1990. The control of vertebrate pests by vertebrate predators. Trends in Ecology and Evolution 5:187-191.
, . PARER, AND P. C. CATLING. 1989. Pro-
longed prey suppression by carnivores-predator- removal experiments. Oecologia 78:458-467.
- R. P. PECH, P. B. BANKS, C. R. DICKMAN, AND R. SMYTHE. 1997. Potential impacts on Aus- tralian native fauna of rabbit calicivirus disease. Australian Nature Conservation Agency, Canber- ra, Australia.
PECH, R. P., A. R. E. SINCLAIR, A. E. NEWSOME, AND P. C. CATLING. 1992. Limits to predator regu- lation of rabbits in Australia: evidence from pred- ator-removal experiments. Oecologia 89:102-112.
RICHARDSON, B. J., AND D. H. WOOD. 1982. Exper- imental ecological studies on a subalpine rabbit
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions

772 IMPACT OF FOX PREDATION ON RABBITS * Banks et al. J. Wildl. Manage. 62(2):1998
population I. Mortality factors acting on emer-
gent kittens. Australian Wildlife Research 9:443- 450.
ROLLS, E. C. 1969. And they all ran wild. Angus and Robertson, Sydney, Australia.
SAUNDERS, G., B. COMAN, J. KINNEAR, AND M. BRAYSHER. 1995. Managing vertebrate pests: foxes. Australian Government Publishing Service, Canberra, Australia.
SEEBECK, J. H. 1978. Diet of the red fox Vulpes vul- pes in a western Victorian forest. Australian Jour- nal of Zoology 3:105-108.
SUMPTION, K. J., AND J. R. FLOWERDEW. 1985. The ecological effects of the decline in rabbits (Oryc- tolagus cuniculus L.) due to myxomatosis. Mam- mal Review 15:151-186.
TROUT, R. C., AND A. M. TITTENSOR. 1989. Can predators regulate wild rabbit Oryctolagus cuni-
culus population density in England and Wales? Mammal Review 19:153-173.
WILLIAMS, K., I. PARER, B. J. COMAN, J. BURLEY, AND M. L. BRAYSHER. 1995. Managing verte- brate pests: rabbits. Australian Government Pub- lishing Service, Canberra, Australia.
WILSON, G. R., N. DEXTER, P. O'BRIEN, AND M. BOM- FORD. 1992. Pest animals in Australia: a survey of introduced wild animals. Bureau of Resource Sci- ences, and Kangaroo Press, Sydney, Australia.
WOOD, D. H. 1981. The demography of a rabbit population in an arid region of New South Wales. Journal of Animal Ecology 49:55-79.
Received 16 September 1996. Accepted 25 July 1997. Associate Editor: O'Connor.
SURVIVAL RATES IN A NONHARVESTED BROWN HARE POPULATION
ERIC MARBOUTIN,' Office National de la Chasse, 163 Boulevard Charcot, 63100 Clermont-Ferrand, France KURT HANSEN, National Environmental Research Institute, Grenivej 14, 8410 Ronde, Denmark
Abstract: Survival in the brown hare (Lepus europaeus) is poorly documented because only life tables and other nonrobust methods have been used to estimate constant annual survival rates. We used recent devel- opments in mark-recapture analysis to model survival patterns in a Danish hare population monitored from 1957 to 1970. Goodness-of-fit tests revealed that the Cormack-Jolly-Seber (CJS) model was an adequate starting point for adults, and age-dependence was considered in modeling survival of yearlings. We found no differences in annual survival rates (4) from 1957 to 1967 in adults, but males survived better than females
(4males = 0.55,
95% CI = 0.50-0.61; Ofemales = 0.50, 95% CI = 0.44-0.56). Yearling survival rates were time dependent and varied with winter severity, sex, and mass. Indeed, body mass strongly influenced the survival of yearling hares: larger animals
(-3 kg) had higher survival rates than smaller animals, in both males and females (4 = 0.40-
0.68 for heavy males and 4 = 0.20-0.44 for lighter males; 4 = 0.31-0.52 for heavy females and 4 = 0.22- 0.40 for lighter females). These variations in survival rates were parallel between body mass classes over time (modeled as a function of winter temperature) within each sex.
JOURNAL OF WILDLIFE MANAGEMENT 62(2):772-779
Key words: brown hare, Denmark, Lepus europaeus, mark-recapture analysis, survival rates.
Over 5 million brown hares (Lepus euro-
paeus) are harvested each year in Western Eu-
rope (Chapman and Flux 1990), which makes this animal an important game species. In ad- dition to short-term oscillations in hare num- bers (Tapper 1987), population abundance has been declining over the last 20 years, as evi- denced by a decrease in hunter harvest (Tapper 1992). Understanding such patterns is difficult because local population densities are highly time and space dependent (Frylestam 1979,
Kovacs and Heltay 1981), and demographic de- terminants of population trends are poorly doc- umented (but see Frylestam 1980, Hansen 1992). Hare populations often exhibit marked fluctuations that are linked to changing survival (Keith 1981), particularly of young (Keith 1990). Rough estimates of adult survival rates
range between 0.35 and 0.52 (Broekhuizen 1979, Kovacs 1983, Pepin 1987). However, these estimates were calculated using ad hoc methods that assumed age- and sex-indepen- dent survival rates. A recent study reports esti- mates of constant annual survival rates that are 1 E-mail: [email protected]
This content downloaded from 128.233.210.97 on Mon, 16 Sep 2013 18:56:08 PMAll use subject to JSTOR Terms and Conditions