early season damage to birch foliage affects the performance of a late season herbivore
TRANSCRIPT

J. Appl. Ent. 105 (1988), 182-189 0 1988 Verlag Paul Parey, Hamburg und Berlin ISSN 0044-2240
Laboratory of Ecological Zoology, Department of Biology, and Kevo Subarctic Research Institute, University of Turku, Finland
Early season damage to birch foliage affects the performance of a late season herbivore
By S. NEUVONEN, S. HANHIMAKI, J. SUOMELA and E. HAUKIOJA
Abstract Early season damage to mountain birch foliage decreased the performance of a late season herbivore, Dineuru virididorsatu (Hym., Tenthredinidae). D . virididorsutu larvae grew significantly slower on trees whose leaves were partially damaged early in the previous or the same growing season than on control trees. Birches refoliating after an early season complete defoliation supported the growth of D . virididorsatu less well than control trees with mature folia e, but another species of insect (Deporuus betulue) (Col., Attelabidae) preferred refoliated to mature ?eaves. Hence, a seasonal niche segregation between species of birch herbivores is not sufficient to prevent interspecific interactions.
1 Introduction
Mechanical damage or insect feeding on leaves can cause adverse chemical and physical changes in foliage quality. Such inducible resistance in the foliage of several deciduous trees is well documented (e.g. BENZ 1974; HAUKIOJA and NIEMELA 1977; EDWARDS and WRATTEN 1985; VALENTINE et al. 1983; RAUPD and DENNO 1984; HAUKIOJA and NEUVO- NEN 1985; but see FOWLER and LAWTON 1985). It is also a potential mechanism by which herbivores may compete with each other. Recent field studies have shown that even temporally separated species may interact via induced changes in host quality (WEST 1985; FAETH 1986a, b; FOWLER and MCGARVIN 1986; HARRISON and KARBAN 1986). In addition, induced changes in foliage quality may determine the propensity of pest populations to fluctuate (HAUKIOJA and NEUVONEN 1987).
In the mountain birch (Betulu pubescens ssp. to~tuosu) zone of Fennoscandia geometrid caterpillars (e.g. Epiwita autumnuta [Bkh.], Operophteru spp.) fluctuate strongly in numbers and they periodically defoliate birches on vast areas (TENOW 1972). All of them feed on young foliage and complete their larval stage by no later than mid season. A tenthredinid sawfly, Dineuru virididorsutu (Retz.), whose larval stage is in late summer, is the most abundant foliage feeding insect on mountain birch during many years but it seldom reaches outbreak densities (HAUKIOJA et al. 1973; HAUKIOJA and ISO-IIVARI 1976; KOPONEN 1981).
Induced resistance in birch foliage may play a prominent role in the population dynamics of E. autumnutu; especially since it prevents rapid growth of the population after a peak density (HAUKIOJA 1980; HAUKIOJA et al. 1985, 1987). Induced changes in foliage quality may also have incidental effects on other insect species feeding on the mountain birch. If early season foliage damage reduces the food quality for late season herbivores, this may partially explain why outbreaks by D. virididorsutu (or late season herbivores in general) are rare.
In this paper we report experiments designed to test whether simulated earlier damage to mountain birch foliage affects the performance of a late season herbivore. Specifically we
U.S. Copyright Clearance Center Code Statement: 0044-2240/88/0502-0182 .$ 02.50/0

Early season damage affects performance of lute season herbivore 183
addressed the following questions: How does D. virididorsutu grow on foliage partially damaged earlier in the season? Does that effect extend to the next growing season? How does D. virididorsutu grow on leaves from refoliated birches?
2 Materials and methods
The experiments were conducted in a mixed pine/birch forest in the immediate vicinity of the Kevo Subarctic Research Station (69"45'N, 2PE), in Finnish Lapland. The quality of mature leaves of the mountain birch was bioassayed by using larvae of D. virididorsata (Hym., Tenthredinidae). Since the effects of our experimental damage may depend more on the phenolo 'cal stage than on calendar date, we calculated the accumulated temperature sums (degree-days above t8e base +5"C = dd5) to facilitate comparison among years.
2.1 D. virididorsata performance in relation to damage to early season foliage in previous and current year
We had three different treatments: 1. untreated controls (C; 11 trees); 2. trees with leaves manually damaged during the previous year (1982) (D-1; 10 trees); and 3. trees with leaves manually damaged earlier in the same season (1983) (D-0; 4 trees).
tearing about 45 % of each leaf. The damage was spread over three occasions (within a week) so t la t at each time a third of the leaves were torn. Treatments were scheduled to coincide with the main feeding eriod of E. auturnnata larvae. In 1982 the treatment began on July 16 (188 dd5) and in 1983 on JUG 12 (275 dd5).
D. virididorsata larvae were collected from the field before the onset of the ex eriment (August 9). Ten larvae with a mean weight of 3.3 mg (range 2-5 mg) were put into each enclkure. Each tree had two enclosures. At the end of the experiment (August 26), larvae were weighed to the nearest mg.
The manual damage was done b
2.2 D. virididorsata performance on leaves refoliated after a total defoliation in early season
Four trees were defoliated on June 19 1985 (68 dds), and six more on June 24 1985 (111 dd5) by cutting, with scissors, the petioles of all leaves. Ten trees remained as untreated controls. The defoliated trees reflushed two to three weeks after the treatment.
Identical (with respect to progeny and size) batches of ten haploid (male) larvae were enclosed in meshba s on experimental trees (one bag per tree) durin August 1-27. The enclosures were changed to new kranches in the same tree on Au ust 23 to avoid food deprivation. Performance was measured by the mean weight of larvae at the encfof the experiment.
2.3 D. virididorsata performance on branches with foliage damaged in mid season The effect of mid season foliage damage on the growth of D. vin'dzdorsata larvae was tested in 1985. Leaves on single branches in 15 trees were damaged by tearing about half of the lamina off on July 23 (350 dd5). One branch on each tree remained as control. In addition, branches from 15 undamaged trees were used. Identical batches of eight larvae (each from the progeny of a different female) were enclosed on these branches from August 8 to August 26. Larval weight at the end of the bioassay was used as a measure of performance.
2.4 Statistical analyses We used tree or branch specific means as individual observations to avoid pseudoreplication (HURLBERT 1984; NEUVONEN and HAUKIOJA 1985). The data was analysed by ANOVA. If there were more than two groups to be compared, the differences between means were tested by the Student- Newman-Keuls test.
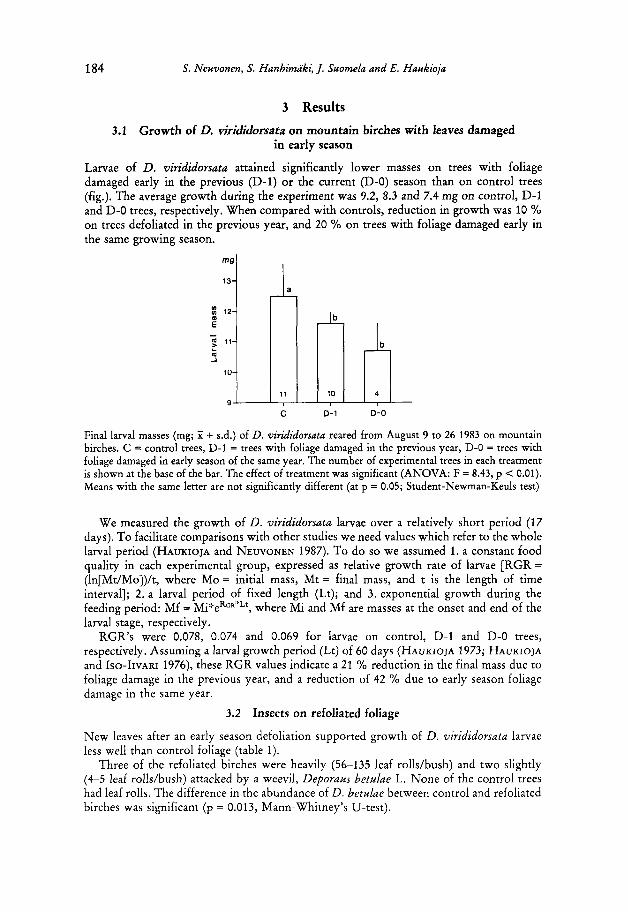
184 S. Neuvonen, S. Hanhimaki, J. Suomela and E . Haukioja
3 Results
in early season
Larvae of D. virididorsata attained significantly lower masses on trees with foliage damaged early in the previous (D-1) or the current (D-0) season than on control trees (fig.). The average growth during the experiment was 9.2, 8.3 and 7.4 mg on control, D-1 and D-0 trees, respectively. When compared with controls, reduction in growth was 10 % on trees defoliated in the previous year, and 20 % on trees with foliage damaged early in the same growing season.
3.1 Growth of D. virididorsata on mountain birches with leaves damaged
C 0-1 D-0
Final larval masses (mg; X + s.d.) of D. virididorsata reared from August 9 to 26 1983 on mountain birches. C = control trees, D-1 = trees with foliage damaged in the previous year, D-0 = trees with foliage damaged in early season of the same year. The number of experimental trees in each treatment is shown at the base of the bar. The effect of treatment was significant (ANOVA: F = 8.43, p < 0.01). Means with the same letter are not significantly different (at p = 0.05; Student-Newman-Keuls test)
We measured the growth of D. virididorsata larvae over a relatively short period (17 days). To facilitate comparisons with other studies we need values which refer to the whole larval period (HAUKIOJA and NEUVONEN 1987). To do so we assumed 1. a constant food quality in each experimental group, expressed as relative growth rate of larvae [RGR = (ln[Mt/Mo])/t, where Mo = initial mass, Mt = final mass, and t is the length of time interval]; 2. a larval period of fixed length (Lt); and 3. exponential growth during the feeding period: Mf = Mi:~eRcR'iLr, where Mi and Mf are masses at the onset and end of the larval stage, respectively.
RGR's were 0.078, 0.074 and 0.069 for larvae on control, D-l and D-0 trees, respectively. Assuming a larval growth period (Lt) of 60 days (HAUKIOJA 1973; HAUKIOJA and ISO-IIVARI 1976), these RGR values indicate a 21 % reduction in the final mass due to foliage damage in the previous year, and a reduction of 42 % due to early season foliage damage in the same year.
3.2 Insects on refoliated foliage
New leaves after an early season defoliation supported growth of D. wirididovsata larvae less well than control foliage (table 1).
Three of the refoliated birches were heavily (56-135 leaf rolldbush) and two slightly (4-5 leaf rolls/bush) attacked by a weevil, Deporaxs betulae L. None of the control trees had leaf rolls. The difference in the abundance of D . betulae between control and refoliated birches was significant (p = 0.013, Mann-Whitney's U-test).

Early season damage affects performance of late season herbivore 185
Table 1. Final larval masses (mg) of D. virididorsata reared from August 1 to 27 1985 on leaves of control or defoliated, and thereafter reflushed, mountain birches
T h e number of trees was 10 in both groups. Treatment means differ significantly (ANOVA: F = 6.97, p < 0.02)
I Treatment Mean SD
Control (mature leaves) Defoliation (reflushed leaves)
12.6 10.2
2.31 1.66
3.3 D. virididorsata performance on branches with leaves damaged in mid season
There were no significant differences between the masses of D. virididorsutu larvae on branches damaged at mid season and on control branches in 1985 (table 2).
Table 2. Final larval masses (mg) of D. viriddorsata reared from August 8 to 26 1985 on branches of mountain birches whose leaves were either damaged or left intact
Damage was done in mid season. The number of experimental branches was 15 in each group. Treatment means do not differ significantly (ANOVA: F = 0.31, p > 0.70)
Treatment Mean SD
Control trees Experimental trees
Control branches Branches with damaged leaves
10.2
10.3 10.6
1.32
0.96 1.19
4 Discussion
4.1 Effects of previous foliage damage on the performance of D. virididorsata
Growth of D. virididorsutu larvae was not impaired in earlier experiments where larvae received mountain birch leaves in late summer, two days after experimental damage (HAUKIOJA and NIEMELA 1979). In our study damage to foliage in late July (accumulated temperature sum 350 dd5) did not affect the growth of D. virididorsutu two to four weeks later. There are at least two potential explanations to these results: 1. birches do not respond to damage to mature foliage, or 2. D. virididorsutu is insensitive to damage- induced reactions in the birch.
Our study demonstrated that D. virididorsutu (or sawflies in general, cf. NIEMELA et al. 1984) was not immune to reactions induced by foliage damage done early in the season (accumulated temperature sum less than 280 dd5). The reduction in the growth of D. virididorsutu (when estimated for the whole larval period) was at least of similar magnitude as in E. uutumnutu (HAUKIOJA and NEUVONEN 1987). We also found an interannual carry- over effect: after being triggered by early season damage, foliage quality remained poor late in the next season. The effects of defoliation in the previous year were apparently not as strong on D. virididorsutu as on E. uutumnuta (HAUKIOJA and NEUVONEN 1987).
Our results parallel those by WRATTEN et al. (1984), which showed that damage to Bet& pubescens foliage in southern England in June reduced significantly the palatability of damaged and adjacent leaves. This effect lasted at least two months while late season damage did not decrease palatability. The main differences between our results and theirs

186 S . Neuvonen, S . Hanhimaki, J. Suomela and E . Haukioja
are that we studied the effects on a specialist herbivore, measured growth rather than palatability, and found that the induced response lasted over 12 months.
Heavy defoliation early in the season may cause a reflush of new foliage even in subarctic birches. It has been suggested that the refoliated leaves - presumably because they are young - are of better quality for insects than mature leaves that occur simultane- ously on undefoliated trees (ROCKWOOD 1974; WILLIAMS and MYERS 1984; FAETH 1986a, 1987). High densities of the weevil (0. betulae) on some of our refoliated trees fitted this expectation but the reverse was true for D. virididorsata: larvae grew better on control than on refoliated trees. FOWLER and LAWTON (1984) found that sawflies (including D. virididorsata) were more abundant on experimentally advanced birch saplings (i.e. with more mature foliage) than on their controls. These results suggest that D. virididorsata larvae are specialized in feeding on mature foliage.
4.2 Consequences of outbreaks by early season defoliators on late season herbivores
Induced resistance in foliage of the mountain birch, triggered by early season herbivory, may contribute to outbreaks by late season herbivores being rare (see also FAETH 1987). From the viewpoint of a late season herbivore it is a density-independent factor in population dynamics and may prevent rapid multiplication of the population. The same applies to the generally low nutritive value of mature birch leaves (HAUKIOJA et al. 1978). Also natural enemies may create difficulties for late season herbivores. Predation on larvae by birds may be heavier in mid and late season than in early summer (HOLMES et al. 1979). Furthermore, insects having their larval periods in late summer generally overwinter as pupae or prepupae (sawflies), and thus they are exposed to small mammal predators, like shrews, throughout the winter (HANSKI and OTRONEN 1985; HANSXI and PARVIAINEN 1985).
Obviously late season herbivores may have properties mitigating the harmful effects of early season herbivory. D. virididorsata is one of the very few sawfly species that oviposits into the petioles, not into laminas, of birch leaves. During outbreaks of an early season species even a partial defoliation may destroy eggs laid into leaf laminas, but there are still the petioles left for oviposition and egg development.
If D. virididorsata larvae can compensate for reduced growth rates by growing a longer period, we may have overestimated the negative effects of early season foliage damage on their performance. However, in the short and capricious subarctic summer a slow growth rate may be detrimental for D. virididorsata because frosts may occur before larvae attain the weight necessary for pupation. In fact this seems to be what happened in the cold summer 1975 (the temperature sum for the season was 444 dd5, i.e. 28 YO below the average for 1962-1985). In that autumn D. virididorsata larvae tried to continue feeding on abscissed (yellow) birch leaves (L. ISO-IIVARI and P. NIEMELA, pers. observation). Peak E. autumnata larval density for the decade at Kevo occurred in the previous year, 1974, (E. HAUKIOJA, pers. observation) and may have influenced growth rates of D. virididorsata larvae in 1975 via food quality. These factors together may explain why larval densities of D. virididorsata near Kevo were about 99 YO lower in 1976 than in 1975 (P. NIEMELA and H. OJALA, unpubl. data).
4.3 Indirect interactions via host quality - general considerations
Although indirect effects between coexisting herbivores via host quality have been studied for a short time only, work done to date shows that these phenomena may be widespread (LOVIS 1975; CROFT and HOYING 1977; KARBAN and CAREY 1984; WEST 1985; FAETH 198613; FOWLER and MCGARVIN 1986; HARRISON and KARBAN 1986). This casts doubts on

Early seuson damage affects performance of late seuson herbivore 187
earlier hypotheses claiming competition to be unimportant in structuring phytophagous insect communities (LAWTON and STRONG 1981; PRICE 1983; STRONG et al. 1984).
Our results show that although two herbivore species have seasonally separated niches they still may interfere via changes in their host - an idea proposed by JANZEN (1973). This finding is interesting when considered in the framework of the classical competition and niche theory. Niches are generally delineated along some measurable dimensions, and niche overlap is considered as a prerequisite to competition (PIANKA 1981). If damage- induced changes in food quality do not relax rapidly, seasonal overlap in feeding time is not a prerequisite to competition or amensalism (which may be more common than actual symmetrical competition in insect communities, LAWTON and HASSELL 1981). Conse- quently, seasonal niche segregation of herbivorous insects probably has other explanations than those based on competition theory. Our results even suggest that despite of the apparent ‘niche segregation’ indirect effects of early season herbivores may be stronger on late season species than the effects of other late season herbivores (FAETH 1987).
Acknowledgements We thank all persons who helped in the experiments, and the personnel of the Kevo Station for their generous help. R. KARBAN, S. KOPONEN, P. NIEMELA, J. SENN and C. TOBIN provided constructive comments on earlier versions of the manuscri t. T. LAURIKAINEN kindly drew the figure. Financially the study was supported by grants from Acaiemy of Finland.
Zusammenfassung EinJuJ.? von Fr~hjuhrs-FruJ.?schaden auf die Entwicklung von spatjressenden Insekten un der Birke
Blattbeschadigungen an Betula pubescens ss tortuosu zu Beginn der Vegetationsperiode beeintrachti- gen die Entwicklung der spadressenden Bl%twespe, Dineura wirididorsutu. Die Larven wuchsen im Vergleich zu Kontrollbaumen signifikant langsamer auf solchen Birken, deren Blatter im Friihjahr desselben oder des vorhergehenden Jahres teilweise beschadigt wurden. Birken, die nach einer vollstandigen Entlaubung im Friihjahr erneut wieder austrieben, waren fur das Wachstum von D . virididorsutu weniger gut geeignet als die Kontrollbaume mit dteren Blattern. Jedoch bevorzugte Deporuus betulae solche Baume, die nach Entlaubun erneut austrieben. Eine interspezifische Konkurrenz wird durch diese Art von Nischenbildung aherdings nicht verhindert.
References BENZ, G., 1974: Negative Riickkoppelung durch Raum- und Nahrungskonkurrenz sowie zyklische
Veranderung der Nahrungsgrundlage als Regelprinzip in der Populationsdynamik des Grauen Larchenwicklers, Zeiraphera diniana (Guenie) (Lep., Tortricidae). Z. ang. Ent. 76, 196-228.
CROFT, B. A.; HOYING, S. A., 1977: Competitive displacement of Panonychus ulmi (Acarina: Tetranychidae) by Aculus schlechtenduli (Acarina: Eriophyidae) in apple orchards. Can. Entomol.
EDWARDS, P. J.; WRAITEN, S. D., 1985: Induced plant defences against insect grazing: fact or artefact? Oikos 44, 70-74.
FAETH, S. H., 1986a: Indirect interactions between seasonal herbivores via leaf chemistry and structure. In: Chemical Mediation of Coevolution. Ed. by K. SPENCER. AIBS Symposium.
- 1986b: Indirect interactions between temporally-separated herbivores mediated by the host plant. Ecology 67, 479494.
- 1987: Community structure and folivorous insect outbreaks: The roles of vertical and horizontal interactions. In: Pest Outbreaks. Ed. by P. BARBOSA and J. C. SCHULTZ. New York: Academic Press.
FOWLER, S. V.; LAWTON, J. H., 1984: Foliage preferences of birch herbivores: a field manipulation experiment. Oikos 42, 239-248.
_ _ 1985: Rapidly induced defenses and talking trees: the devil’s advocate position. Am. Nat. 126,
FOWLER, S. V.; MCGARVIN, M., 1986: The effects of leaf damage on the performance of insect
HANSKI, I.; OTRONEN, M., 1985: Food quality induced variances in larval performance: comparison
109, 1025-1034.
18 1-195.
herbivores on birch, Betula pubescens. J. Anim. Ecol. 55, 565-573.
between rare and common pine-feeding sawflies (Diprionidae). Oikos 44, 165-174.

188 S. Neuvonen, S . Hanhimaki, J . Suomela and E . Haukioja
HANSKI, I., PARVIAINEN, P., 1985: Cocoon predation by small mammals, and pine sawfly population dynamics. Oikos 45, 125-136.
HARRISON, S.; KARBAN, R., 1986: Effects of an early-season folivorous moth on the success of a later- season species, mediated by a change in the quality of the shared host, Lupinus arboreus Sims. Oecologia (Berlin) 69, 354-359.
HAUKIOJA, E., 1973: Weight development, consumption and egestion of Dineura virididorsata (Hym., Tenthredinidae) larvae. Rep. Kevo Subarctic Res. Stat. 10, 9-13.
- 1980: O n the role of plant defences in the fluctuation of herbivore populations. Oikos 35,202-213. HAUKIOJA, E.; ISO-IIVARI, L., 1976: Local and annual variation in secondary production by Dinewra
virididorsatu (Hym., Tenthredinidae). Rep. Kevo Subarctic Res. Stat. 13, 26-32. HAUKIOJA, E.; KOPONEN, S.; OJALA, H., 1973: Local differences in birch leaf consumption by
invertebrates in northern Norway and Finland. Rep. Kevo Subarctic Res. Stat. 10, 29-33. HAUKIOJA, E.; NEUVONEN, S., 1985: Induced long-term resistance of birch foliage against defoliators:
defensive or incidental? Ecology 66, 1303-1308. _ _ 1987: Insect population dynamics and induction of plant resistance: the testing of hypotheses.
In: Pest Outbreaks. Ed. by P. BARBOSA and J. C. SCHULTZ. New York: Academic Press. HAUKIOJA, E.; NEUVONEN, S.; HANHIMAKI, S.; NIEMELA, P., 1987: The autumnal moth in Fennos-
candia. In: Dynamics of Forest Insect Populations: Patterns, Causes, and Management Strategies. Ed. by A. A. BERRYMAN. New York: Plenum Press.
HAUKIOJA, E.; N I E M E ~ , P., 1977: Retarded growth of a geometrid larva after mechanical damage to leaves of its host tree. Ann. Zool. Fennici 14, 48-52.
_ _ 1979: Birch leaves as a resource for herbivores: Seasonal occurrence of increased resistance in foliage after mechanical damage of adjacent leaves. Oecologia, Berlin, 39, 151-159.
HAUKIOJA, E.; N I E M E ~ , P.; ISO-IIVARI, L.; OJALA, H.; ARO, E.-M., 1978: Birch leaves as a resource for herbivores. I. Variation in the suitability of leaves. Rep. Kevo Subarctic Res. Stat. 14, 5-12.
HAUKIOJA, E.; SUOMELA, J.; NEUVONEN, S., 1985: Long-term inducible resistance in birch foliage: triggering cues and efficacy on a defoliator. Oecologia, (Berlin), 65, 363-369.
HOLMES, R. T.; SCHULTZ, J. C.; NOTHNAGLE, P., 1979: Bird predation on forest insects: an exclosure experiment. Science 206,462-463.
HURLBERT, S. H., 1984: Pseudoreplication and the design of ecological field experiments. Ecol. Monogr. 54, 187-211.
JANZEN, D. H., 1973: Host plants as islands. 11. Competition in evolutionary and contemporary time. Am. Nat. 107, 786-790.
KARBAN, R.; CAREY, J. R., 1984: Induced resistance of cotton seedlings to mites. Science 225, 53-54. KOPONEN, S., 1981 : Outbreaks of Dineura virididorsata (Hymenoptera) and Eriocrania (Lepidop-
LAWTON, J. H.; HASSELL, M. P., 1981: Asymmetrical competition in insects. Nature 289, 793-796. LAWTON, J. H.; STRONG, D. R. JR., 1981: Community patterns and competition in folivorous insects.
Am. Nat. 118, 317-338. LOVIS, C., 1975: Contribution a l'etude des tenthredes du meleze (Hymenoptera: Symphyta) en
relation avec l'evolution dynamique des populations de Zeiraphera diniana GuenCe (Lepidoptera: Tortricidae) en Haute-Engadine. Mitt. Schweiz. Entomol. Ges. 48, 181-192.
NEUVONEN, S.; HAUKIOJA, E., 1985: How to study induced plant resistance? Oecologia, Berlin, 66, 456-457.
NIEMELA, P., TUOMI, J.; MANNILA, R.; OJALA, P., 1984: The effect of previous damage on the quality of Scots pine foliage as food for Diprionid sawflies. Z. ang. Ent. 98, 33-43.
PIANKA, E., 1981: Competition and niche theory. In: Theoretical Ecology. Principles and Applica- tions. Ed. by R. M. MAY. 2nd edn. Oxford: Blackwell Scientific Publications. Pp. 167-196.
PRICE, P. W., 1983: Hypotheses on organization and evolution in herbivorous insect communities. In: Variable Plants and Herbivores in Natural and Managed Systems. Ed. by R. F. DENNO and M. S. MCCLURE. New York: Academic Press. Pp. 559-596.
RAUPP, M. J.; DENNO, R. F., 1984: The suitability of damaged willow leaves as food for the leaf beetle, Plagiodera versicolora. Ecol. Entomol. 9, 443448.
ROCKWOOD, L. L., 1974: Seasonal changes in the susceptibility of Crescentia alata leaves to the flea beetle Oedionychus sp. Ecology 55, 142-148.
STRONG, D. R.; LAWTON, J. H.; SOUTHWOOD, R., 1984: Insect on plants. Community patterns and mechanisms. Oxford: Blackwell Scientific Publications.
TENOW, O., 1972: The outbreaks of Oporinia awt~rnnata Bkh. and Operophthera spp. (Lep., Geometridae) in the Scandinavian mountain chain and northern Finland 1862-1968. Zool. Bidrag Uppsala, Suppl. 2, 1-107.
VALENTINE, H. T.; WALLNER, W. E.; WARGO, P. M., 1983: Nutritional changes in host foliage during and after defoliation and their relation to the weight of gypsy moth pupae. Oecologia (Berlin) 57,
WEST, C., 1985: Factors underlying the late seasonal appearance of the lepidopterous leaf-mining
tera) on mountain birch in northernmost Norway. Not. Entomol. 61, 41-44.
298-302.
guild on oak. Ecol. Entomol. 10, 111-120.

Early season damage affects performance of late season herbivore 189
WILLIAMS, K. S.; MYERS, J. H., 1984: Previous herbivore attack of red alder may improve food quality
WRATTEN, S. D.; EDWARDS, P. J.; DUNN, I., 1984: Wound-induced changes in the palatability of for fall webworm larvae. Oecologia, Berlin, 63, 166-170.
Betula pubescens and B. pendula. Oecologia, Berlin, 61, 372-375.
University of Turku, SF-20500 Turku 50, Finland Author’s address: SEPPO NEUVONEN, Laboratory of Ecological Zoology, Department of Biology,