early larval shell development in mytilid bivalve mytilus
TRANSCRIPT

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
VENUS (Jap, jour, Mulac.) nkVol. 58, No.3 <1999): 119-127
Early Larval Shell Development in
Mytilid Bivalve Mytilus gaUoprovincialis
Emiko HAyAKAzE and Kazushige TANABE
Geological institute, University of lbkyo, 7iokyo 113-O033, Japan
Abstract: The early shcll rnorphogenesis of a mytilid bivalve M),tilus gattoprovincialis
Lamarck was studied by scanning electron microscopy, Larvue were taken at 1 - 1,5 hour inter-
vals after hatching until the early veliger stage. Trochophore larvae appeared within 16 hours
after fertilization at incubating temperatures between 15.0 - 18.2 OC.
They have the same pat-tern in the mode of formation er the prodissoconch 1 (Pd I). The Pd I began to build in the mid-
dlc trochophore stage. It was initially made of an ovul-shaped, fu11y organic univalved shell
(pellicle). The univalved organie shell developed further into a bivalved shape with a median
hinge line in the late trochophore stage. Calcification of the larval shell occurred in the early
veliger stage within 70 h()urs after fertilization. At this stage of development, the shell gland on
the dersal side secreted irregularly orientcd, needle-like spherulites undcrneath the pellicle. An
oval or low, cap-shaped, univulved shell observed in the eariy veliger larvae of the Bivalvia is
known to occur in the early stages of other molluscan groups such as the Monoplacophora,
Cephalopoda and Gastropoda, suggesting its synapomorphic nature amongst the Conchifera.
Keywords: M.vtilus galXoprovincialis, larval shell, development
Introduction
Embryonic dcvelopment and larval morphology of Bivalvia have been described in some edi-
ble and economically important spccies such as Ostrea edutis (Waller, 1981), Chlamys hastata
(Hodgson & Burke, 1988), Ai;gopecten pur;p"ratus (Bellolio et al,, 1993), and others (e.g. Rees,
1950; Loosanoff & Davis, 1963; Yoshida, 1964; Sastry, 1979; Cragg & Crisp, 1991). However, lit-
tle is known on the early development of the larval shell of Bivalvia, especially concerning onto-
genetic changes of shell structures and mineralogical cemposition. Most papers dealing with
bivalve larval shells have been fbcussed on the ecological and taxonomic aspects of the late veliger
stage (Loesanoff et al., 1966; Lutz & Hidu, 1979; Chanley & Diamani, 1980; Fuller & Lutz, 1989).
Only a few workers have described the morphology of living bivalves at early stages of devel-
opment (Ansell. 1962; LaBarbera, 1974; Waller, 1981; Bandel, 1988; Togo et al., 1993; Moueza et
at., 1999). These explained how the shell structure and mineralogical composition changed from
larval to post-larval stagcs. However, microstrucrural features of the shell gland and the primary
shell have not yet been sufficiently described.
in this study, these features were examined in a Japanese common mussel species by means of
optical and scanning electron microscopy and their phylogenetic implications were discussed.
Material and Methods
Sampting ofliving larvae:
Mytilus gallQprovinciatis Lamarck (Mytiliidae) that is
around the Japanese Islands, was used in this study. About
on February 22, 1996 at the intertidal zone of Nojima
distributed cornmonly in the waters
1OO mature individuals were collected
Canal, Yokohama City, Kanagawa

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
120 VENUS: VoL 58, No.3 (l999)
Prefecture, Gametes were samp]ed by the inducement method to abrupt changes in water tempera-
ture, Oocytcs and sperms were mixed in a small tank (20 liters in volume) and kept at daity meantemperatures of 17.4
"C for the first 20 hours after fertilization (temperature range: 17.0 - 18.2
OC)
and of 15.5 OC
thereat'ter (tcmperature range: 15.0 - 16,O OC).
Sixteen hours after fertilization liv-ing larvae were taken at 1 - 1.5 hour intervals for about 96 heurs.Scanni.ng electron nticroscopy:
Some living larvae of Ml gattoprovincialis were anacsthetised using 2-phenoxyethanol (WakoPure Chemicals Industries, Ltd.) buffered with sea water. The anuesthctised specimens could still
swim but did not retract the velum. This suggcsts that they swam using ciliary currents which were
unaffected by the nEurcotic agent.
All specimens were fixed with 6% glutaraldehyde-paraformaldehyde in O.2M cacody]ate bufferwith sea water at pH7.5, They were obscrved and photographed using an Olympus model AHBS-515 optica[ microscope. For scanning electron microscopy, specimens were cleaned twice with dis-tilled water and dehydrated through a graded series of ethanol. They wcre freeze-dried with t-butyl-
alcohol and mounted onto SEM stubs using a double-sided carbon tape. Specimens were coated
with p]atinum and then observed using scanning electron microscopes (Hitachi medel S-2400S and
model S-4500).
To ebserve the internal structure of the 1arval shell, selected fixed specimens of M. gatlo-provi.ncialis wcre cut using a razor blade. These fractured specimens were coated with platinumand observed with the SEM.
Laryal Development
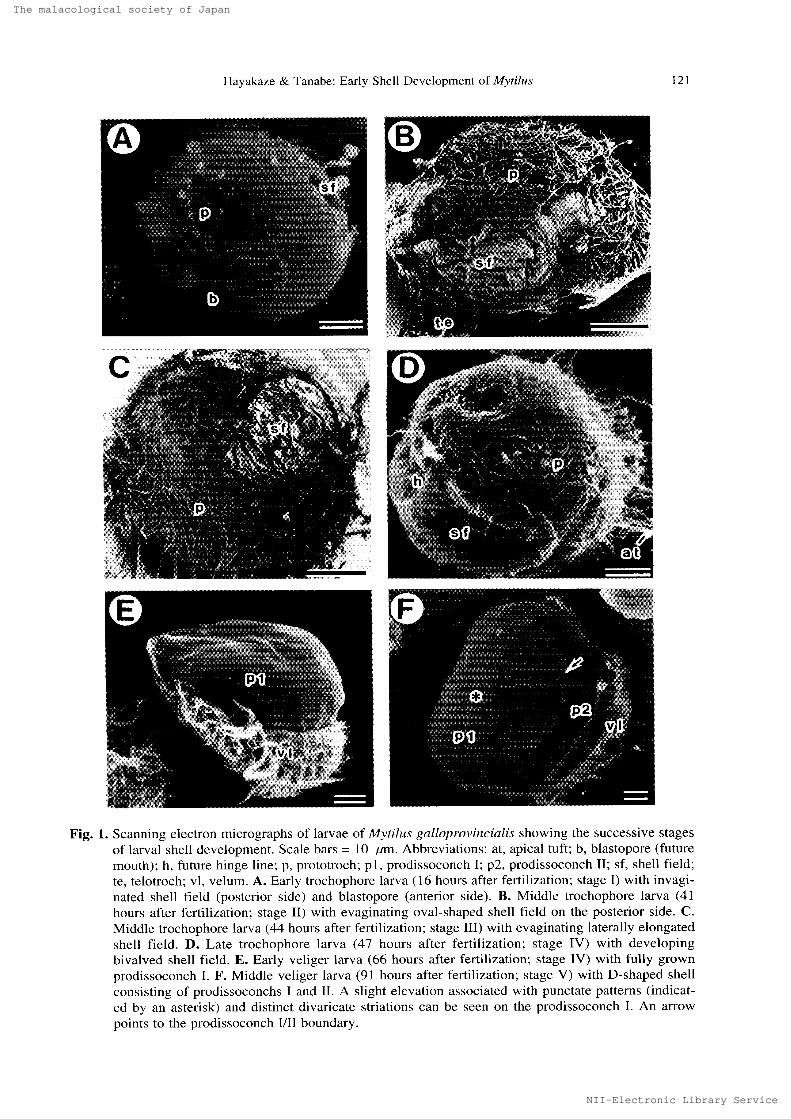
Spawned oocytes develeped into trochophere larvae 16 hours after fertilization, and their mean
diumeter attained 60 pt rn (Fig. 1A), Two shallow invaginations on the opposite sidcs were observed
on each trochophore larva, one was the mouth primordium (blastopore) on the anterior side and theother was the shell field on thc posterior side. Each trochophore larva also possessed a prototroch,apical tuft, and telotroch. The apical tuft was about 30 um in length, The prototroch developedfrom the primary trochoblasts and encircled the mid-region of the trochophore larvae (Fig. IA-D).The telotroch occurrcd on the posterior apex of the trochophorc that beared a small tuft of cilia
(Fig, IB).
The trochophore larvae began to develop into the vc]iger Iarvae 66 hours aftcr fertilization (Fig.IE), Each veliger larva, about 70 ltm length, had a D-shaped bivalved shell in which the larvalbody was housed. The shell in the early veliger stage lacked apparent surface sculpture such as
gruwth lines, except for irregular elevations on thc apical portion. At 80 hours after fertilization, the
larva began to secrete an another type of the shell with commarginat growth lines termed the
prodissoconch II (Jablonski & Lutz, 1980) on the anterior side of the prodissoconch I (Fig. IF).
Mode of Laryal Shell Formation
As a result of scanning electron microscopy of 1arvae at different growth stagcs, the larval shelldeveiopment in Ml gaUopn)vincialis may be divided into the fo11owing five stagcs. The relationship
between developmental stages of the 1arval organs and the shell is summarized in Fig. 2.Stage I:
Prior to shell secretion. two invaginations began to appear on oppesite sides of the 1arval bodyin the early trochophore stage (Fig. IA). One of them had a crcscent shape with a relatively deepdepression, and the other was crater shaped with a shallow dcprcssion. Following the criteria of
Kniprath (1980), they are identified as the posterior shell field and the anterior blastopore (mouth
The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
Hayakaze & Tunabe: Early Sheli Devclepmcnt of "dytil"s 121
Fig. 1. Seanning electron micrographs of 1arvae of Mr tilt{s galloprovincialis showing the successive stages
of larval shell development. Scalc bars = 1O um. Abbreviations: at. apical tuft; b, blastopore (future
mouth); h, future hinge line; p, prototroch; p1, prodissoconch I; p2, prodissocench II; sf, shell field;
tc, Lclotroch; vl, velum. A. Early trochQphore larva (16 hours after fertilization; stage I) with invagi-
nuted shell field (postcrior sidc) and blastopore (anterior side). B. Middle trochophere larva (41 hours after fertilization; stagc II) with evaginuting eval-shaped shell field on thc posterior side. C. Middlc trochophore 1arva (44 hours after fortilization; stage III) with evaginating latcral]y elongated
shell ficld. D. Late trochophore larva (47 hours after fertilization; stage IV) with developing
bivalved shell field, E. Early veliger larva (66 hours after fertilization; stage IV) with fully grown prodisseconch I, F. Middle veliger larva <91 huurs after fertilization: stage V) with D-shaped $hell
consisting of prodissoconchs I and II. A slight elevation associated with punctatc patterns (indicat- cd by an asterisk) and distinct divaricate striations can be seen on the prodissoconch I. An arrow
points to the prodissoconch IllI boundary.

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
l22 VENUS: VeL 58, No.3 (1999)
HOURS AFTERFERTILIZATION
24 48 72 96
Trochophore
------
Veliger
llIHIV
wwwwvakXXXXXXXNXXXXN""N"XtXNXXXXXXNXX)tXXXS
---------ny----------s-mt-flf-!/.iZV--------
c --
sasrmceww Early trochophore stage (prototroch, apical tuft)
t,N,,"",,N Late trochophore stage (telotroch, two invaginations)x4(ncrno Early veliger stage (vetum, prodissoconch 1)
- Middle-late veliger stage (prodissoconch l, prodissoconch lt)
jl: Stage 11 (an oval univalved shell secreted)
ltl; Stage lll (univalved shell developed to bivalves)
IV: Stage IV (hinge line occurred; prodissoconch l)
V: Stage V (prodissoconch ll appeared), C: Calcification started
Fig. 2. Relatienship between developmental stages and shell formation in larvae of Mytitus gaUny)rovin- ciatis examined.
primordia) respectively. The shell gland primordium did not secrete a shell in this stage.Stage IJ:
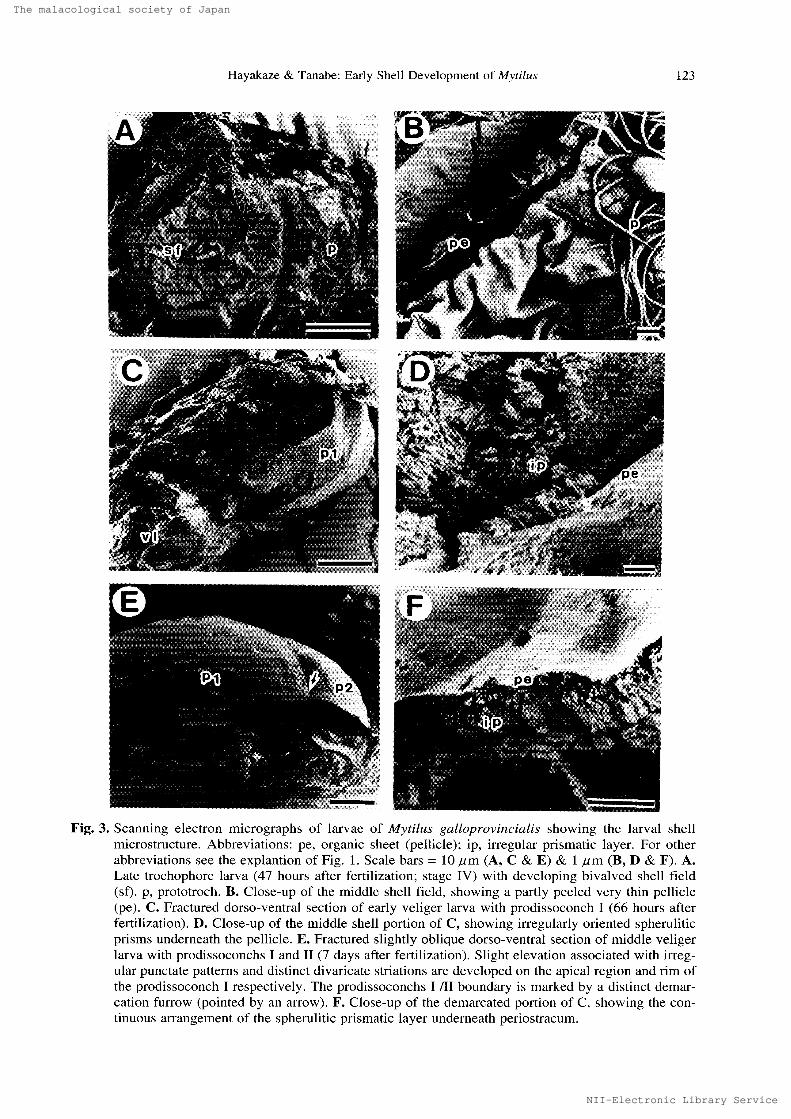
In this stagc of development, the central ponion of the shell field was distinctly evaginated and
covered with a primary shell consisting of a very thin organic sheet (pellicle), less than O.2umthick (Fig. 3B). The pellicle appeared 41 hours after fertilization. It was univalved with an oval out-
]ine, measuring about 10 - 15 um in lenger diameter.
Stage M:
Soon after stage II, the oval organic shell enlarged and covered about 30% of the larval body
(Fig. IC). The longer diameter attained about 30 um. There was no trace of a ca]cified structure
under the pellicle,Stage IV (prodissoconch I):
In the middle to late trochophore stage, a shallow t'urrow began to appear along the mid-line of
the elongated organic shell (Fig. ID). Along this 1'urrow, the shell was distinctly partitioned intotwo valves that were gently curved inward. The furrow was changed into the hinge line in the sub-
sequent early veliger stage. Hinge tccth and ligament were still undeveloped in the trochophore
stage. Thc bivalved larval shell in this stage of development is defined as the prodissoconch 1. Asdevelopmcnt proeeeded, the hinge area rapidly enlarged laterally, and the prodissocench I became
a typical D shape in the early veliger stage (Fig. IE). The full-grown prodissoconch I was 70 um
in length.
Threughout the trochophore and earliest veliger stages, the prodissoconch I lackcd apparent
growth lines (Fig, 1D) and censisted only of a very thin petlicle (Fig. 3A-B). Calcification of the
prodissoconch I started in the early veliger stage. Name}y, a thin calcified layer, about O.5 - 1,O ptmthick, consisting of aggregations of irregularly oriented, needle-likc spherulitic prisms, began to
The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
Hayakaze & Tanabe: Early Shell Deyelupment of M.ytilus l23
Fig.3. Scanning electron micrographs of larvae of Mytilus galloprovineiatis showing thc larval shcll
microstructure. Abbreviations: pe, organic sheet (pellicle); ip. irregular prismatic layer. For other
abbreviations see the explantion of Fig. 1. Scale bars = 10 ptm (A, C & E) & 1 "m (B, D & F). A.
Late trochephure larva (47 hours after fertilization; stage IV) with dcvcloping bivalved shell field
(bf). p, prototroch. B. Close-up of the middle sheil field, showing a partly peeled very thin pellicLe (pe). C. Fractured dorso-ventral section of early veliger larva with prodisgoconch I (66 hours after
ferti]ization). D. Close-up of the middle shell portion or C, showing irregularly oriented spherulitic
prisms underneath the pellicle. E. Fractured slightly oblique dorso-ventral section of middle veliger
larva with prodissoconchs 1 and tl (7 days after fertilization). Slight elcvation associatcd with irreg-
ular punctate patterns and distinct divaricate striations are developed on thc apical rcgion and rim of
the prodissoconch I respectively. The prodissoconchs I III boundary is murked by a distinct demur-
cation furrow (pointed by an arrow). F. Close-up of the demarcated portion of C, showing the con-
tinuous arrangement of the spherulitic prismatic layer undcrncath pcriostracum.

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
124 VENUS: VoL 58, No.3 (1999)
appear undcrneath the preformed pellicle in the dorsal region of each valve (Fig. 3C-D). At this
stage of development, the upical shell portion had a slight elevation (Fig. IE). Distinct divaricate
striations were present at the rim ef the apical shell elevation and were cxtendcd te the margin ef
the prodissoconch I (Fig, 3E). These surface sculptural pattcms of the prodissoconch I were
described previously us "punctate-stellate
puttem" (Carricker & Palmer, 1979) or "pitted
and stel-
late-radial zones" (Waller, 1981).
Stage V (Prodissoconch ll):
After stage IV, the bivalved shell grew rapidly, and the two D-shttped valves became to contact
each other at the ventral margin. Then, the prodissoconch II with commiarginal growth lines began
to appear at the shell margin of the prodissoconch I. The boundary between prodissoconchs I and
II is marked by thc prcscncc of a shallow demarcation furrow (indicated by arrows in Figs. IF &
3E). As the shell growth proceeded, the calcified layer broadly covered the inner surft!ce of prodis-soconch Il (Fig. 3E). There was no discontinuity in the shell microstructurc at the boundary (Fig.3F).
Discussion
EarL>' development qf the larvat shelt
As a result uf this study, it was realized that in M. galloprevincialis, trochophore larvae devel-
oped two invaginations on the anterior and posterior sides of the body, They correspond respec-
tively to the mouth primordium (blastopore) and shell field, The invaginated shell field and mouth
have been described in trochophore larvae of the same species from southern France (Kniprath,1980), S()isLfta sotidissin'ta from the north Atlantic coast in Massachusetts (Eyster, 1986) and
Anom.alocaidia brasiliana from the tropical mangrovc swamps of western Atlantic (Moueza et al,,
1999). These lines of evidence indicate that the development of an invaginated shell field and
mouth in the trochophore larvae is a common feature in the Bivalvia.
Our SEM observations revealed that the trochophore ]arvac of M. galloprovinc'iatis first secret-
ed a univalved, elliptical-shaped orgttnic shell (Fig. IB). A similar oval-shaped univalved organic
shell wus figured in larvae of the tridinid c]am, 71eredora malleotus from the Mediterranean Sea
(Bandel, 1988) and the tropical mangrove clam A. brasiliana (Moueza et aL, 1999), Bande] (1988)regarded those of the fbrmer species as abnormal larval shells, but they apparently correspond in
the degree of development to the univalved organic shells secreted in the late trochophore stage
(stage tl) of M. gattoprovinciati,g described herein. Hayakaze (1997) has also recognized an oval,
univalved organic shell in the late trochophore larvae of Pinctada fLtcata martensii and Ruditapes
philillpinarum. Sccretion of an organic shcll prior te the calcification in the larval and/or embryon-
ic stage has been knowri in other molluscan groups such as gastropods (Iwata, 1980; Togo,1984;
Togo et aL, 1993) and cephalepods (Tanabe & Uchiyama, 1997).
Larval shclls secrcted in thc early veligcr stagc have been described in the spiny scallop,
Chlamys hastata from the west coast of Canada (Hodgson & Birke, 1988), the European oyster,
Ostrea edulis t'rom Long Islund, northeast ot' the United States (Waller, 1981), and the south Pacific
scallop, Aigopecten purpiiratus from Chile (Bellolio et al., 1993), They are dorsovcntrally elongat-
ed, with a pair of notches on the mid-lateral sides of the shclls. A shallow and straight furrow con-
nects the notches, by which the larval shel] is distinctly partitioned into two equal-sized "valves".
At that stage of development, the bivalved larval shell in these species is made up of a single thin
organic membrane, without any trace of hinge teeth and provinculum, These larval shells may be
compared to the larval shells at stage IV in our definition,
Calcification of bivalve larval shells was previously considered to start at the central part of
each valve (MacBride, 1904). This study has revealed that in M. gatloprovincialis, calcification

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
Hayakaze & Tanabe: Early Shell Devcloprnent of M.vtilus 125
first occurred underneath the erganic shell in the regien of thc retractor muscles in the early veliger
stagc (Fig. 2C), LaBarbera (1974), however, realized by means of polarized microscopy and X-ray
diffraction anatysis that in larvae of 7-7'idacna squamosa, calcification began along the hinge line.
Kniprath (1980) also demonstrated with a polarized microscopc that in larvae of M, gatlru)rovin-
cialis fi'om southern France, the region covered with the retracter muscles was first calcified. These
lines of cvidence suggest that in rnany bivalve species, culcification appears to occur in the early
veliger stage.
Thc size of prodissoconch [ and the relatiye length of prodissoconch II have been regarded as
important criteria to infer the modes of development and capability of dispersal in the Bivalvia
(Jablonski & Lutz, 1980). According to these authors, it has been generally interpreted that the
prodissoconch I and II boundary was fbrmed when the shell gland developed into the mantle, since
the abrupt appearance uf grewth lines at the boundary refiected secretion of CaC03 at the shell
margin, However, our study makes clear that in M. gatl(provinciatis, the shell gland in the poste-
rior region secreted spherulitic prisms undemeath the pellicle in thc early veliger stage (Fig. 3D).
There is no major break in the shell microstructure at thc boundary between prodissoconchs I and
II (Fig. 3F). Waller (1981) concluded that in O, edulis, the shell gland gradually changed into the
mant]e prior to the appearance of Pd II. Our observations on M. galloprovinciali.s corre]ate well
with those on O. edulis by Waller (1981), both suggesting that the prodissoconchs I and II de not
reflect the ditTerentiation of the mantle.
Comparison (lf larval shell moilpholegy with other molluscs
The pre$encc of calcified bivalves aniculated by the ligament is now regarded as one ef the
important autapomorphies in the class Bivalvia (Salvini-Plawen & Steiner, 1996), Howcver, our
work makes clear that bivalves first sccrete a univalved, oval-shaped organic shell in the middle
trochophore stage. A similar eval-shaped or low cap-shaped shell was known to occur in the early
stage of other molluscan classes such as extunt menoplacophorans Laevipilina and Micropilina
(Waren & Hain, 1992) and the cephalopod Ntit{titus (Tanabe & Uchiyama, 1997), The early 1arval
shell development of the Gastrepoda has not yet been well docuTnented, though a flat organic pri-
mary shell (pellicle) secreted by a shell gland was describcd in trochophore larvae of Limnaea
stangnalis by Raven (1958, fig. 43). Furthermore, Sasaki (1998) has shown that the primitive gas-
tropods (Patellogastropoda) share an uncoiled, high conical protoconch with a bilatera] symmetry.
These lines ef evidence seems to suggest that the ancestor of thc Bivalvia had a univalved calcified
shell.
According to the schemc of early molluscan evolution by Runnegar (1983) and Runnegar &
Pojeta (1985), the Bivalvia branched off from the Rostroconchia clade with a laterally compressed
univalved shcll by the development of paircd adductor muscles and decalcification of the pos-terodorsal shell margin, The prodissoconch I of M. gattoprovirtcialis observed in this study was
developed into an elongated form with a median furrow in stagc IV. It was distinctly partitioned
into two "valves",
but the "valves"
were not articulated by a ligament ner hinge teeth. These fea-
tures are very similar to the conditions of adult shells in the Rostroconchia and support Runnegar's
(1983) hypothesis that the ancestors of the Bivalvia probably had a hatchet-shaped foot and well-
diversified pallial and adductor muscles, both of which were suitable for collecting food.
The phylogenetic relationships and origin of mollusean classes have not yet been sufficiently
clucidated, This subject shouLd be considered by combining different approachcs such as molecu-
lar systematics, molcculur developmental biology, and comparative anatomy and embryology of
extant forms us well as paleobiological studies on Cambro-Ordoyician fussils. Further cemparative
analysis focusing on organogenesis and biomineralization of growing larvae will provide a lot of
important infbrmation te understund the morphological evolution of molluscs.

The malacological society of Japan
NII-Electronic Library Service
The malaoologioal sooiety of Japan
】26 VENUS :Vol.58, No .3 (1999)
Acknowledgn 昭 nts ;一
We thank Drs. Neil H 、 Landman and John D . Taylor for critical reading of a firstdra丘 of this manuscript , alld Ms . Keiko Shimizu lbr her asslstance in the SEM , and Messrs. Yuji Uchimura,Yurimasa Terao and Dr. Shin’ichi SaIo f{〕r lheirhelp in collccting growing bivalve larvae. This manuscriptwas much improved by critical comments of two anonymous referees . This work was supported by [heGrant−In−Aid of the Japanese Ministry of Education , Science, Culture and Sports(No ,07304042 f{〕r 1996 ;
No .09304049 fbr 1997 − 1999),
ム ラサ キ イガ イ の 初 期幼生殻の 発 達過 程
早風 恵美.了 ・棚 部
・成
(.東京大学地 質学教室)
要 約
ム ラ サ キ イガ イ (Mytilus galloprovinciati.s Lanlarck) の 幼生殻形態 を走査 型電子 顕微鏡を用 い て 観察
し,そ の 発達過 程 を記載 した 。 水温 15.O − 182 ℃ の 条件下で ,受精後 16 時間 で トロ コ フ ォ ア 幼 生が
出現 し,さ らに 約 40 時 問を経過 し て ベ リ ジ ャー幼生 に 変化 した 。 トロ コ フ ォ ア幼生中期に 楕円形 の
一
枚 の 有機 質薄層 か ら な る 原 殻 1 が 現 れ た 。 時 問 と と もに原殻 1は ひ ょ うた ん 型 に 変 わ り, 中央の くび れ
に沿 っ て ヒ ン ジが 出現 し て 2 枚 の 殻が形 成 された。受精 後約 70 時間を経過 し た ベ リ ジ ャー幼生 初 期 に
原 殻 1 の 石 灰 化 が 確 認 さ れ, さ ら に 受精後約 80 時間で ,原殻 1の 周囲 に 明 瞭な くびれ を伴 っ て 原殻 II
が 発達 し始め た 。 有 機 質で で きた 1 枚の 楕円形の 殻は ,他 の 二 枚貝類 ばか りで な く単板類 ,腹 足類,
頭 足類 の 発 生初期 に も確認 さ れ る こ とか ら,貝殻亜 門 の 共有派生 形質 の ひ とつ と解釈さ れ る D
References
Ansell, A . D .1962. The functional morphology of the Iarva and pc}stlarval devclopment of Venus striatuta
(da Costa).ノl Mar . Biθ1. Ass θ c. U . K,42 : 419 − 443 ,Bande1 , K .1988. Stages in the ontc 》geny and a model of the evolution of bivalves(Mollusca). Pal伽 砿 Z.. 62 : 217 − 254 .Bellolio, G . Lohrma 冂 n , K .& Dupr6, E.1993 . Larval m 【〕rphc〕logy of the scallop Argopecten purpurat“ s as
revealed by scanning electron microscopy . Vetiger,36 : 332 − 342 .Carricker. M 、 R .& Palmer , R . E .1979a , UltrastructUral morphogenesis of prodissoconch and early disso−
conch valvcs of the oyster Cr α・s’・9t爪 t厂 ea vi厂ginica. Proc . Natn . Shelltfisheries A ∬ oc .,69 : 103 − 128 .Chanley, P.& Diamani, P.1980 . Comparative descriptions of some oyster larvac from New Zealand and
Chile, and a description of a new genus of oyster , Tioslrea. N . Z .」. Ma r. Fre,yhwater Res .,14 :103 −
120.Cragg . S . M .& Crisp, D .」.1991,The bi{}bgy of scallop larvae, pp.75 − 132.ノη :Shulnway, S. E.(ed .) Devetopments in Aquaculture 硼 d Fishe 厂ies ∫cien (:e,21 ’ Sca〃ops .’Biolo8y, Ec θ 1θ gy and
Aquaculture . Elsevier, New York,Eyster, L . S ,1986. Shell inorganic composit .ion and onset Qf shell mineralization during bivalve and gas−
tr〔〕pod embryogenesis . Biθ 1. Bu 〃..170:211 − 231.Fuller, S, C.& Lutz, R . A .1989 . Shell morpho 豆ogy of larval and post−larval mytilids from the North −
Western Atlantic..1.〜瞼 臥 Biot, A ∬ oc .σ, K .,69 : 181 − 218 .Hyayakazc . E.1997. Larval developmcnt and shell morphology of five species of Bivalvia.42pp, Mastcr Thesls, GeoL Inst., Univ. TokyQ .Hodgsun . C , A .& Burke , R . S.1988. Development and larval morphology of the spiny scallop , C〃 αm メs
hastata. Biθ1. Bull.,174 :303 − 318.Iwata, K .1980. Minera ]ization and architecture of the larval shell of Haliotis di,ycus hannai Ino (Archaeogastropoda).ノ, Fac . Sci., H θkkaido しJniv,, Ser.ハ/ ‘6eol.& Mineral .丿,
19 : 305 − 320 .Jablonski. D .& Lutz, R . A .1980 . Molluscan larval 8hell morphology , Ecoiogical and paleontological appli 一
N 工工一Eleotronio Library

The malacological society of Japan
NII-Electronic Library Service
The malacologicalsociety of Japan
Hayakaze & Tanabe; Early Shell Development of dytilus 127
cations, pp. 323 - 377. In: Rhoads, D. C. & Lutz, R. A. (eds) Skeletal Growth ofAquatic Organisms.
Plenurn Press, New York.Kniprath, E. 19RO. Larval development of the shell ttnd the shcll g}and in Mytitus (Bivalvia). RouxFs
Archives Devel(w. Biot,, 188: 201 - 204.
LaBarbera, M. 1974. Calcification of the first larva] shell of 1:ridacna squamosa (Tridacnidae: Bivalvia).
Man Biol,, 25: 233 J 238.
Loosanoff, V. L, & Davis, H. C. 1963. Rearing of bivalve motluscs, pp. 1 - 136. In: F. S. Russel (ed.)
Advances of Marine Biolog.),, 1. Academic Press, London,Loosanoff, V. L., Davis, H. C. & Chanley, P. E. 1966. Dimensions and shapes of larvae ef seme marine
bivalve mollusks. Malacologia. 4: 351 - 435,
Lutz, R, A, & Hidu, H. I979. Hinge merphogenesis in the shells of larval and early post-larval mussels
(M>,tilzas edulis L. and Modiolus mediolus (L.)). J. Mar, Biol. Assoc. U. K., 59: 1 1 1 - 121.
MacBride, E. W. 1904. The Canadian oyster. Canad. Rec. Sci., 9: 145 - 156.
Moueza, M., Gros, O. & Frenkiel, L. 1999. Ernbryonic, larval and postlarval developrnent of the tropical
clam Anomalocardia hrasitiana (Bivalvia, Veneridae). J. MotL Stud., 65: 73 - 88.
Raven, C. P. I958. Morphogenesis; 71he Analysis of Moltuscan Development. 311pp. Pergamon Press,
London.Rees, C. B. 1950. The idcntification and classification of lamellibranch larvae. HuU ButL Mar. EcoL, 3: 73
- 104,
Runnegar, B., 1983: Molluscan phylogeny revisited. Mem. Assoc. Australasian Palaeont., 1: 121 - 144.
Runnegar, B. & Pojeta, J. Jr. 1985. 0rigin and diversification of the Mollusca, pp, 1 - 57. ltu Trueman,
E.
R. & Clarke, M. R, (eds) 7)Pie Molluh'ca, 10: Evolution. Academic Press, Orlando.
Sastry, A. N. 1979. Pelecypoda, pp., 113-292. In: Giese, A. C. & Pearse, J. S. (eds.) Reproduction of Marine Invertehrates. 5. Academic Press, New York.
Sa]vini-Plawen, L. v. & Steiner, G, 1996. Synapomorphics and plesiomorphies in higher classification of
Mollusca, pp, 29 - 52. In: J. D. Taylor (ed.) Origin and Evol"tionarly Radiation of the Moliusca,
Oxford Univ. Press, Oxford.Sasaki, T. 1998. Comparative anatomy and phylogeny of the Recent Archtteogastropoda <Mollusca:
Gastropoda). U}iiv. Mus., Univ. 71}kyo, Bull., (38), 223pp.Tanabe, K. & Uchiyama, K. 1997. Development of the embryonic shell stmcturc in Nautilas. Veliger, 40:
2U3 -
215.Togo, Y. 1984, Scanning electron microscepy ef larval and early postlarval shells in the freshwater snail,
Cipangopaludina .iaponica (V. Martens) (Mesogastropoda, Viviparidae). I GeoL Soc. Jap., 90: 565
-
576.Togo, Y., Suzuki, S, & Hikida, Y. 1993. 0ntogenic changes in the shell structurc of the freshwater bivalve
SPhaerium J'aponicum (Heterodonta), pp. 55 - 63. bi: Kobayashi, T., Mutvei, H. & Sahni, A. (eds.)
Structure, Formation and Evolution of Fossil Hard 7'issues. Tokai University Press, Tokyo.
Waller, T. R. 1981. Functional morphology and 1arval development of veliger larvae of the European oys-
ter, Ostrea edulis Linn6. Smithson. Contr. Zbol., 328, 1 -70.
War6n, A. & Hain, S. 1992. Laevipitina antarctica and Micropolina artzi, two new monoplacophorans from
the Antarctic. Vketiger, 5: 165 - 176.
Yoshida, H. 1964. Seedings of Shelij}sh. Hokuryu-kan Beok Company, Tokyo 221pp. (in Japanesc).
[Received: April 9, 1999; Accepted; June 25, 1999]