dynorphin: important mediator for electroacupuncture analgesia in the spinal cord of the rabbit
TRANSCRIPT

Pain, 18 (1984) 367-376
Elsevier
367
PA1 00615
Dynorphin: Important Mediator for Electroacupuncture Analgesia in the Spinal Cord of
the Rabbit
Ji-Sheng Han ’ and Guo-Xi Xie Department of Physiology, Beijing (Peking) Medical College, Beijing 300083 (People’s Republic of China)
(Received 18 July 1983, accepted 14 October 1983)
Summary
Intrathecal injection of 12 nmol of dynorphin elicited marked analgesia as
measured by tail flick latency, the effect being about 20 times more potent than with morphine. This analgesic effect could be reversed by naloxone at a dose 1.5fold higher than that needed to reverse morphine analgesia. Intrathecal injection of anti-dynorphin antibody blocked electroacupuncture (EA) analgesia by 77%, the
effect lasting for at least 4 h. In rabbits made tolerant to EA analgesia by long-term EA stimulation, intrathecal injection of dynorphin no longer exhibited an analgesic effect. No analgesia was noticed when dynorphin (10 nmol) was injected into the
periaqueductal grey (PAG) of the rabbit, nor was EA analgesia blocked by anti-dy-
norphin antibody injected into PAG. These results suggest that dynorphin reduces
nocifensive responses in the spinal cord and may play an important role in mediating EA analgesia at the spinal level.
Introduction
Although dynorphin (DYN) has been characterized as the most potent endoge- nous opioid peptide assessed in guinea pig ileum preparation [7], little is known about its physiological role in intact animals or humans. Intracerebroventricular (icv) injection of DYN caused an increased rate of feeding, excessive grooming [20],
hypothermia [14] and, when administered in a large dose, catatonia [13]. In contrast
’ To whom correspondence should be sent.
0304-3959/84/$03.00 0 1984 Elsevier Science Publishers B.V.

to morphine and most of the endogenous opioids, DYN elicited little or ver! ucah
analgesia on icv administration [3,13.14,20,22].
In a previous paper we reported that DYN showed a potent analgesic effect after
intrathecal injection into the rat [9]. This observation was substantiated in the
present study in the rabbit. Further evidence is presented to show that DYN may
serve as one of the neurochemical substrates for mediating electroacupuncture (EA) analgesia in the spinal cord of the rabbit.
Methods
Intracerebral and intrathecal injection
Male rabbits weighing 2.0-2.4 kg were used. Stainless steel cannulae of outer
diameter 0.7 mm, were permanently fixed on the skull directed bilaterally to the
periaqueductal gray (PAG) at coordinates P9.5, L1.0, H1O.O mm according to
Sawyer et al. [20]. During the experiment, an injection tube of outer diameter 0.3 mm was inserted into and protruded 1.5 mm beyond the cannula to reach the PAG.
The volume to be injected within 8 min was 4 ~1.
For intrathecal injection, a polyethylene tube (PE-10) was introduced under
pentobarbital anesthesia through the foramen magnum to the subarachnoid space of the lumbar segments and fixed in situ leaving 8 cm outside the skin. The injection volume was 50 ~1 followed by 20 ~1 of normal saline for flushing.
Experiments were commenced after a recovery period of 1 week and were conducted once a week, DYN being injected only once into each animal.
The sites of injection were verified by cryostat examination on completion of the experiment.
Nociceptive test and EA stimulation
The animal was blindfolded and fixed in a hammock. Two stainless steel needles
of diameter 0.15 mm were inserted into the hind leg points, Zusanli and Quenlun, as
described elsewhere [12]. Nociception was tested by radiant heat applied to the snout or the tail and the latency of escape response (ERL) was measured to the accuracy of 0.1 sec. The average value of 3 measurements at the beginning of the experiment was
taken as the basal ERL. EA of 1 V, 2-15 Hz was then given for 10 min, and the nociceptive test repeated every 10 min, 5 times in all. The latencies measured after EA were expressed as percentages of the basal ERL with + 200% as the cut-off limit
WI. The data are presented as mean k S.E.M. Student’s t test (two-tailed) was used to
determine the significance of the difference between the two groups.
Chemicals DYN (full sequence of 17 amino acids) and affinity purified IgG from antiserum
against DYN,,_,,, were gifts from Professor Avram Goldstein (Addiction Research Foundation, Palo Alto, CA, U.S.A.). This antiserum product, designated as ap-D,,-4, was dissolved in a vehicle of 1 ml containing 150 mM sodium phosphate and 3%
369
bovine serum albumin and gave a titer of 1 : 2#,~0 in the radioimmunoassay for
DYNfl-13) or DYN. Its cross-immunoreactivity with a-neo-endorphin was less than 0.01%. and it had no cross-reactivity with P-endorphin or Leu-enkephalin.
Normal rabbit serum was taken from a non-immunized rabbit.
Naloxone-HCl was obtained from Endo Laboratories (U.S.A.). Morphine HCl is a product of Shenyang Drug House (China).
Results
The effect of intratheca~ ~~jecti~~ of D YN on EM
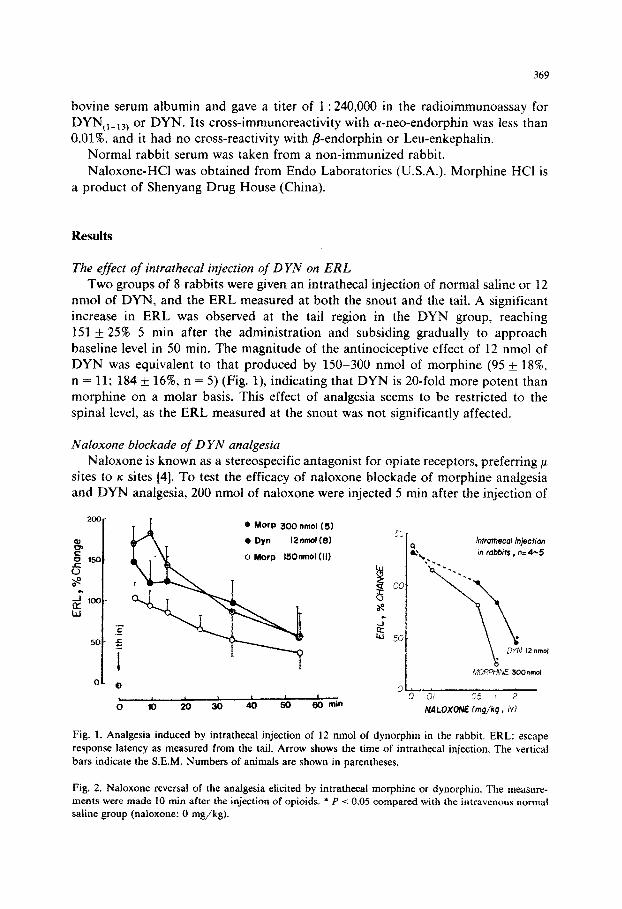
Two groups of 8 rabbits were given an intrathecal injection of normal saline or 12 nmol of DYN, and the ERL measured at both the snout and the tail. A significant increase in ERL was observed at the tail region in the DYN group, reaching
151 rf: 25% 5 min after the administration and subsiding gradually to approach baseline level in 50 min. The magnitude of the antinociceptive effect of 12 nmol of DYN was equivalent to that produced by 150-300 nmol of morphine (95 + 18%,
n = 11; 184 _C 16% n = 5) (Fig. l), indicating that DYN is 20-fold more potent than morphine on a molar basis. This effect of analgesia seems to be restricted to the
spinal level, as the ERL measured at the snout was not significantly affected.
Kaloxone stockade of D YAJ analgesia
Naloxone is known as a stereospecific antagonist for opiate receptors, preferring p
sites to K sites [4]. To test the efficacy of naloxone blockade of morphine analgesia and DYN analgesia, 200 nmol of naloxone were injected 5 min after the injection of
200.. @ MOrP 300 nmol(5)
e, P CJ so- 0 Morp 150nmol (II)
6
8
2=_,00-
w
._
.c
=i
o* 0 , t olo7io30~~ 60 min
htmhecol Injection in robbits , n=4-5
05 I 2
NALOXONE fmg/kg , id
Fig. 1. Analgesia induced by intrathecal injection of 12 nmol of dynorphin in the rabbit. ERL: escape
response latency as measured from the tail. Arrow shows the time of intrathecal injection. The verticai
bars indicate the S.E.M. Numbers of animals are shown in parentheses.
Fig. 2. Naloxone reversal of the analgesia elicited by intrathecal morphine or dynorphin. The measure- ments were made 10 min after the injection of opioids. * P -C 0.05 compared with the intravenous normal saline group (naloxone: 0 n-&kg).
TA
BL
E
I
RE
VE
RSA
L
OF
AN
AL
GE
SIA
IN
DU
CE
D
BY
IN
TR
AT
HE
CA
L
MO
RPH
INE
O
R
DY
NO
RPH
IN
BY
IN
TR
AT
HE
CA
L
NA
LO
XO
NE
Dyn
orph
in
or
mor
phin
e w
as
inje
cted
at
0
min
; na
loxo
ne
(Nal
) w
as
inje
cted
at
5.
5 m
in.
N
Bas
al
Cha
nge
in
ER
L
(Rb)
ER
L
(W
5 m
in
10 m
in
15 m
in
35
min
55
m
in
15
min
MO
T
300
Inn0
1 5
5.9k
O.8
17
1 it
23
184+
16
14
3+24
86
+37
55+3
8 52
k40
Mor
p 30
0 M
l01
+NaI
20
01~1
~1
5 5.
6 *0
.8
176+
16
78+3
8 *
45*2
5 *
27+3
4 74
*39
60&
29
DY
N
12 n
mol
4
6.5
+ 1.
1 14
2+46
12
4k43
12
5*47
91
+38
48+3
6 16
+17
DY
N
12 n
mol
+NaI
20
0 nm
ol
5 6.
5 f
0.9
169k
31
135*
40
79*3
2 43
+17
* 83
k34
67+4
4 _.
__
I_
* P
< 0
.05
com
pare
d w
ith
the
corr
espo
ndin
g gr
oup
with
out
nalo
xone
.

371
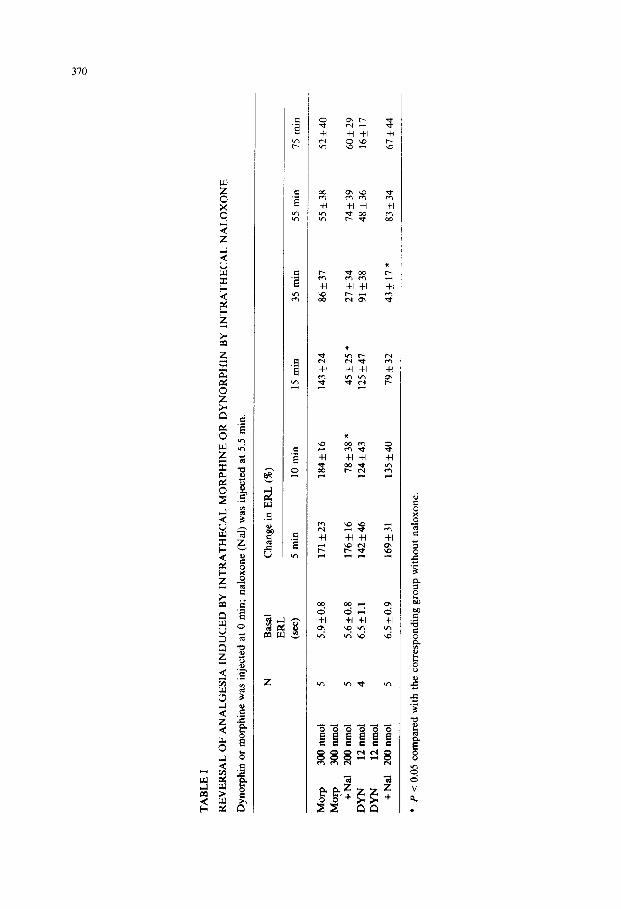
morphine (300 nmol) or DYN (12 nmol) via the same intrathecal cannula. Naloxone
produced a prompt (within 5 min) and complete (at 30 min) reversal for morphine analgesia, whereas for DYN analgesia, only a weak and delayed antagonism was
encountered (Table I). In another experiment, naloxone or normal saline was intravenously injected 5
min prior to the intrathecal injection of morphine (300 nmol) or DYN (12 nmol), and ERL was measured 10 min after the opiate injection. Fig. 2 shows a dose-depen- dent naloxone reversal of morphine analgesia or DYN analgesia. Intrathecal injec-
tion of 300 nmol of morphine or 12 nmol of DYN elicited a similar degree of analgesia (130-1408). Whereas the effect of morphine was almost completely
reversed by 1 mg/kg of naloxone (P < 0.05), 2 mg/kg of naloxone was needed to
reverse the effect of DYN by 64% (P < 0.05). The doses of naloxone required for
50% reversal of morphine and DYN analgesia were 0.6 and 1.5 mg/kg, respectively. The results indicate that naloxone is more effective in blocking the analgesic effect
of morphine than that of DYN.
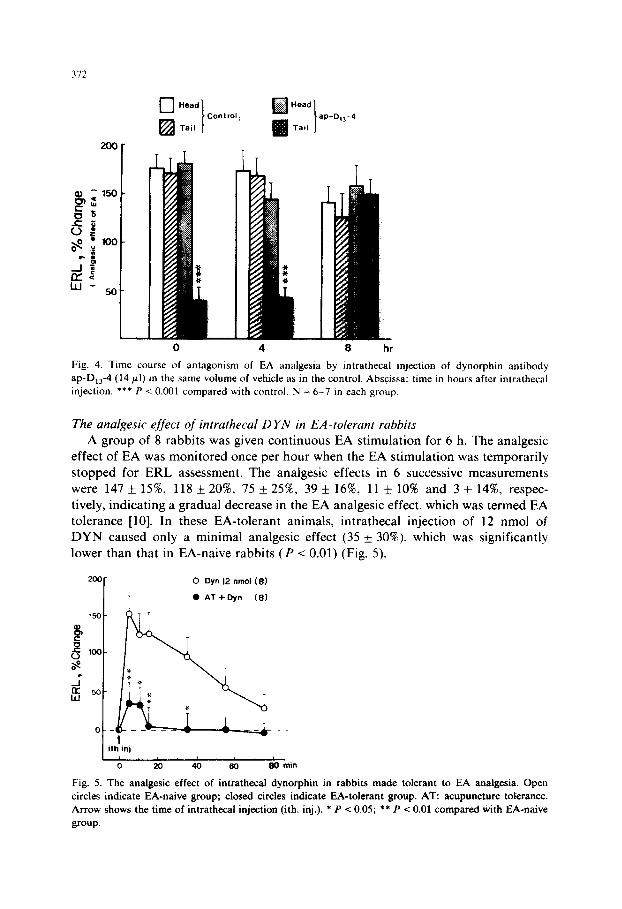
The effect of intrathecal injection of anti-DYN IgG on EA analgesia Three groups of 7 rabbits were given an intrathecal injection of 14 ~1 of DYN
antibody (ap-D,,4), the vehicle or normal rabbit serum, diluted with normal saline
to a final volume of 50 ~1. EA was given 10 min after the intrathecal injection and ERL measured both at the snout and tail regions. EA analgesia was markedly
attenuated in the ap-D,,4 group as measured at the tail (39 f 15’%), leaving the analgesic effect of EA intact in the head region (Fig. 3). The antagonistic effect of
anti-DYN IgG to EA analgesia lasted for at least 4 h and disappeared within 8 h (Fig. 4). The analgesic effect obtained in the normal serum group (139 f 24’S, not
shown in Fig. 3) was apparently of the same order of magnitude as that of the vehicle group (170 &- 15%).
0 Control (7)
0 aP-chp4 (7)
-Head
I 1
-lo 0 lo 20 30 40 50 min
Fig. 3. Antagonism of electroacupuncture @A) analgesia by intrathecal injection of anti-dynorphin IgG (ap-D,,-4). EA was applied at the hind leg points. Significant attenuation of EA analgesia was seen only in the tail region. *** P < 0.001 compared with control (vehicle) group.
372
-
cl Head
q Tail I Con,ro,, q Head
Tail I
0 4
aP-D,3-4
8 hr
Fig. 4. Time course of antagonism of EA analgesia by intrathecal injection of dynorphin antibody
ap-D,,4 (14 pl) in the same volume of vehicle as in the control. Abscissa: time in hours after intrathecal
injection. *** P c 0.001 compared with control. N = 6-7 in each group.
The analgesic effect of intrathecal D YN in EA-tolerant rabbits
A group of 8 rabbits was given continuous EA stimulation for 6 h. The analgesic effect of EA was monitored once per hour when the EA stimulation was temporarily
stopped for ERL assessment. The analgesic effects in 6 successive measurements
were 147 of: 15%, 118 f 20%, 75 f 25%, 39 k 16%, 11 f 10% and 3 + 14W, respec-
tively, indicating a gradual decrease in the EA analgesic effect, which was termed EA tolerance [lo]. In these EA-tolerant animals, intrathecal injection of 12 nmol of DYN caused only a minimal analgesic effect (35 + 30%). which was significantly lower than that in EA-naive rabbits (P < 0.01) (Fig. 5).
200 0 Dyn 12 nmol(8)
, 0 AT +Dyn (8)
ith’inj
0 20 40 60 80 min
Fig. 5. The analgesic effect of intrathecal dynorphin in rabbits made tolerant to EA analgesia. Open
circles indicate EA-naive group; closed circles indicate EA-tolerant group. AT: acupuncture tolerance.
Arrow shows the time of intrathecal injection (ith. inj.). * P < 0.05; ** P < 0.01 compared with ISA-naive
group.
313
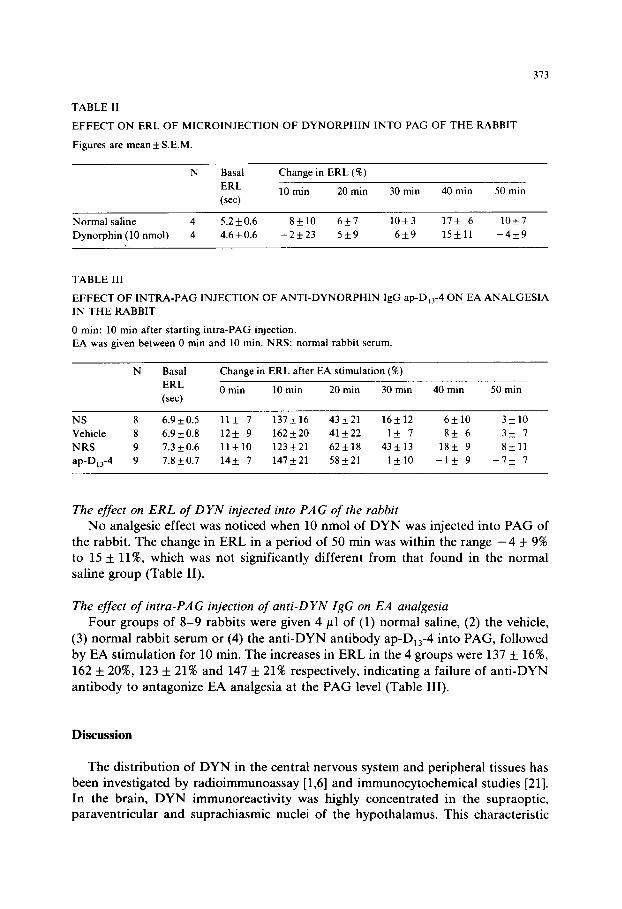
TABLE II
EFFECT ON ERL OF MICROINJECTION OF DYNORPHIN INTO PAG OF THE RABBIT
Figures are mean + S.E.M.
N Basal Change in ERL (‘K)
ERL 10 min 20 min 30 min 40 min 50 min (set)
Normal saline 4 5.2 + 0.6 S&10 6k7 10*3 17* 6 10+7
Dynorphin (10 nmol) 4 4.6 * 0.6 -2k23 5+9 6+9 15*11 -4*9
TABLE III
EFFECT OF INTRA-PAG INJECTION OF ANTI-DYNORPHIN IgG ap-D,,-4 ON EA ANALGESIA
IN THE RABBIT
0 min: 10 min after starting intra-PAG injection.
EA was given between 0 min and 10 min. NRS: normal rabbit serum
N Basal Change in ERL after EA stimulation (X)
ERL 0 min 10 min 20 min 30 min 40 min 50 min (set)
NS 8 6.9 k 0.5 11* 7 137*16 43+21 16+12 6klO 3+10
Vehicle 8 6.9 * 0.8 12+ 9 162 + 20 41k22 1* 7 8k 6 3+ 7
NRS 9 1.3 k 0.6 11*10 123*21 62k18 43+13 18+ 9 8+11
ap-D,,-4 9 7.8 + 0.7 14+ 7 147+21 58+21 1*10 -1* 9 -7* I
The effect on ERL of DYN injected into PAG of the rabbit
No analgesic effect was noticed when 10 nmol of DYN was injected into PAG of the rabbit. The change in ERL in a period of 50 min was within the range -4 + 9% to 15 + ll%, which was not significantly different from that found in the normal saline group (Table II).
The effect of intra-PAG injection of anti-DYN IgG on EA analgesia
Four groups of 8-9 rabbits were given 4 ~1 of (1) normal saline, (2) the vehicle,
(3) normal rabbit serum or (4) the anti-DYN antibody ap-D,,-4 into PAG, followed by EA stimulation for 10 min. The increases in ERL in the 4 groups were 137 + 1696,
162 f 20%, 123 f 21% and 147 f 21% respectively, indicating a failure of anti-DYN antibody to antagonize EA analgesia at the PAG level (Table III).
Discussion
The distribution of DYN in the central nervous system and peripheral tissues has been investigated by radioimmunoassay [1,6] and immunocytochemical studies [21]. In the brain, DYN immunoreactivity was highly concentrated in the supraoptic, paraventricular and suprachiasmic nuclei of the hypothalamus. This characteristic

374
distribution, together with the finding that injection of anti-DYN antiserum into the arcuate nucleus of the hypothalamus increased the level of luteinizing hormone in
the blood of the immature female rat [18], seem to attribute to cerebral DYN a regulatory role in endocrine secretion. There is much controversy concerning the
analgesic effect of DYN [3,13,14,20,22]. Results from this laboratory have shown
that no significant analgesia was observed when as much as 40 pg (about 20 nmol) of DYN was injected into the lateral ventricle of the rat [9] or 10 nmol into PAG of
the rabbit, nor was the effect of EA analgesia blocked by antibody against DYN administered into PAG of the rabbit. None of these data support DYN as a potent
analgesic at the brain level.
In the spinal cord, DYN was found mainly in the dorsal gray matter. The fact that the content of this peptide was not seriously affected by spinal transection and
multiple dorsal rhizotomy seems to indicate dynorphinergic neurons as short-axoned interneurons intrinsic to the spinal cord [l]. Evidence revealed in this study has
shown that exogenously injected DYN in the spinal cord produced prompt analgesia which lasted for nearly 1 h. The effect produced by 12 nmol of DYN was bracketed by 150 and 300 nmol of morphine, indicating that DYN was about 20 times more potent than morphine in producing analgesia. This may prove to be the first
evidence ascribing a potent analgesic effect to DYN in the spinal cord of rabbit. Similar results have been obtained in rats [9,16] and mice [15].
An important issue which should be mentioned here is that, in contrast to findings in the rabbit, where there was no difficulty in showing DYN analgesia by injecting through a chronically implanted cannula, a full analgesic effect could only
be elicited in the rat when DYN was injected within 24-48 h of placement of intrathecal cannula [l]. In rats bearing a cannula for more than 1 week, DYN was
no longer effective. We reported this finding at the 1982 International Narcotic
Research Conference, and it has since been verified by Herz’s group [16] and Goldstein’s group [5]. The reason for this curious decrease in potency of DYN with
time after cannulation in rats is not clear. This technical problem may well account
for the inability of Tung and Yaksh [19] to obtain analgesia with DYN,_,, after its intrathecal injection in rats, although morphine and EKC were still effective in these
chronic preparations. The implication of endogenous opioids in mediating acupuncture analgesia has
been well documented [8], although exactly which member of the opioid family is involved has not yet been clarified. The participation of DYN in acupuncture analgesia was suggested by the following experimental evidence: (1) a cross-tolerance
existed between EA analgesia and DYN analgesia; (2) the effect of EA analgesia was markedly decreased for at least 4 h by intrathecal injection of IgG against DYN. By adopting the antibody microinjection technique it has become possible to ‘dissect’
the action of different endogenous opioids more clearly than the conventional method of blocking opiate receptors by naloxone. Results obtained in previous [ll] and present studies suggest that EA utilizes enkephalins and /3-endorphin, but not DYN, in the PAG area for mediating its analgesic effect, while in the spinal cord it is DYN and enkephalin, but not /3-endorphin, that are important for channeling the
acupuncture effect.

DYN has been suggested as the endogenous ligand for the K opioid receptors [2] and one of the criteria for characterizing K receptor was its low sensitivity to naloxone blockade. Data obtained in this study indicate that DYN analgesia is much less sensitive to naloxone antagonism than morphine. Besides, rats made tolerant to intrathecal morphine showed no significant change in the analgesic response to intrathecal DYN while those tolerant to intrathecal EKC showed a di~nished analgesic response to DYN (unpublished~_ It, therefore, seems reasona- bte to postulate that the K receptor is the most plausible candidate for mediating DYN analgesia in the spinal cord of the rabbit and rat.
Acknowkdgements
We thank Dr. A. Goldstein for the generous gifts of dynorphin and dynorphin antiserum used in this study. The purification of IgG from dynorphin antiserum was kindly performed by Dr. W. Fischli, Addiction Research Foundation, Palo Alto, CA, U.S.A. The gift of naloxone from Endo Laboratories is sincerely acknowledged.
References
1 Bottieelli, L.J., Cox, B.M. and Goldstein, A., Immunoreactive dynorphin in mammalian spinal cord and dorsal root ganglia, Proc. nat. Acad. Sci. (Wash.), 78 (1981) 7783-7786.
2 Chavkin, C., James, I.F. and Goldstein, A., Dynorphin is a specific endogenous &and of the I( opioid receptor, Science, 215 (1982) 413-415.
3 Friedman, H.J., Jen, M.F.. Chang, J.K., Lee, N.M. and Loh, H.H., Dynorphin: a possible modulatory peptide on morphine or ~~ndo~hin analgesia in mouse, Europ. J. Ph~acol., 69 (1981) 357-360.
4 Gilbert, P.E. and Martin, W.R., The effects of morphine- and nalorphine-like drugs in the nondepen- dent, morphine-dependent and cyclazocine-dependent chronic spinal dog, J. Pharmacol. exp. Ther., 198 (1976) 66-82.
5 Goldstein, A., Biology and chemistry of the dynorphin peptides. In: J. Meienhofer and S. Udenfriend (Eds.), The Peptides: Analysis, Synthesis, Biology, Vol. 7, Academic Press, New York, in press.
6 Goldstein, A. and Ghazarossian, V.E., Immunoreactive dynorphin in pituitary and brain, Proc. nat. Acad. Sci. (Wash.), 77 (1980) 6207-6210.
7 Goldstein, A., Tachibana, S., Lowney, L.I., Hunkapiller, M. and Hood, L., Dynorphin-(l-13), an extraordinary potent opioid peptide, Proe. nat. Acad. Sci. (Wash.), 76 (1979) 6666-6670.
8 Han, J.S. and Terenius, L., Nemochemical basis of acupuncture analgesia, Ann. Rev. Pharmacol. Toxicol., 22 (1982) 193-220.
9 Han, J.S. and Xie, C.W., Dynorphin: potent analgesic effect in spinal cord of the rat, Life Sci., 31 (1982) 1781-1784.
10 Han, J.S., Li, S.J. and Tang, J., Tolerance to electroacupuncture and its cross tolerance to morphine, Neuropharmacology, 20 (1981) 593-596.
11 Han, J.S., Xie, G.X., Zhou, Z.F., Folkesson, R. and Terenius, L., Enkephalin and &endorphin as mediators of ele-ctro-acupuncture analgesia in rabbits: an antiserum microinjection study. In: E. Costa and M. Trabucchi (Eds.), Regulatory Peptides: from Molecuiar Biology to Function, Raven Press, New York, 1982, pp. 369-377.
12 Han, J.S., Zhou, Z.F. and Xuan, Y.T., Acupuncture has an analgesic effect in rabbits, Pain. 15 (1982) 83-91.
13 Herman, H.J., Leslie, F. and Goldstein, A., Behavioral effects and in vivo degradation of intraventric- uIar administered d~o~hin~l-13) and D-Ala*-dyno~~n~l-11) in rats, Life Sci., 27 (1980) 883-892.

376
14 Petrie, EC, Tiffany, S.T., Baker, T.B. and Dahl, J.L., Dynorphin(l-13): analgesia. hypothermia. cross
tolerance with morphine and &endorphin, Peptides, 3 (1982) 41-47.
15 Piercey, M.F., Varner, K. and Schroeder, L.A., Analgesic activity of intraspinally administered
dynorphin and ethylketocyclazocine, Europ. J. Pharmacol., 80 (1982) 283-284.
16 Przewlocki, R., Shearman, G.T. and Herz. A., Mixed opioid/nonopioid effects of dynorphin and
dynorphin related peptides after their intrathecal injection in rats, Neuropeptides. 3 (1983) 233-240.
17 Sawyer, C.H., Everett, J.W. and Green, J.D.. The rabbit diencephalon in stereotaxic coordinates, J.
camp. Neurol., 101 (1954) 801-824.
18 Schulz, R., Wilhelm, A., Pirke, K.M., Gramsch, C. and Hem, A., P-Endorphin and dynorphin control
serum luteinizing hormone level in immature female rats, Nature (Lond.), 294 (1981) 757-759.
19 Tung, AS. and Yaksh, T.L., In viva evidence for multiple opiate receptors mediati. g analgesia in the
rat spinal cord, Brain Res., 247 (1982) 75-83.
20 Walker, J.M., Katz, R.J. and Akil, H., Behavioral effect of dyno~hin,_,~ in the mo se and rat: initial
observations, Peptides, 1 (1980) 341-345.
21 Watson, S.J.. AkiI, H., Ghazarossian, V.E. and Goldstein, A., Dynorphin imt mocytochemicai
localization in brain and peripheral nervous system: preliminary studies, Proc. nat. A ad. Sci. (Wash.),
78 (1981) 1260-1263.
22 Wtister, M., Schulz, R. and Hem, A., Opiate activity and receptor selectivity of dyr >rphin,_,, and
related peptides, Neurosci. Lett., 20 (1980) 79-83.