drug resistance updates - aalto...cle, we examined the recent crystal structures of abc proteins to...
TRANSCRIPT

I
OM
WDa
b
c
d
e
f
g
a
ARRA
KMCAPNNN
C
h1
Drug Resistance Updates 27 (2016) 14–29
Contents lists available at ScienceDirect
Drug Resistance Updates
jo ur nal homep age: www.elsev ier .com/ locate /drup
nvited review
vercoming ABC transporter-mediated multidrug resistance:olecular mechanisms and novel therapeutic drug strategies
en Lia, Han Zhangb, Yehuda G. Assaraf c, Kun Zhaod, Xiaojun Xue, Jinbing Xie f,ong-Hua Yangg,∗∗, Zhe-Sheng Cheng,∗
Department of Hematology and Oncology, Winship Cancer Institute, Emory University, Atlanta, GA 30322, USADepartment of Biology, Georgia State University, Atlanta, GA 30303, USAThe Fred Wyszkowski Cancer Research Laboratory, Department of Biology, Technion-Israel Institute of Technology, Haifa 32000, IsraelDepartment of Mathematics and Statistics, Georgia State University, Atlanta, GA 30303, USADivision of Vaccine Discovery, La Jolla Institute for Allergy and Immunology, La Jolla, CA 92037, USADepartment of Physics, Nanjing University, Nanjing, ChinaDepartment of Pharmaceutical Sciences, St. John’s University, Queens, NY 11439, USA
r t i c l e i n f o
rticle history:eceived 17 October 2015eceived in revised form 24 April 2016ccepted 6 May 2016
eywords:ultidrug resistance
ancer chemotherapyBC transporter-glycoproteinanoparticle
a b s t r a c t
Multidrug resistance is a key determinant of cancer chemotherapy failure. One of the major causes ofmultidrug resistance is the enhanced efflux of drugs by membrane ABC transporters. Targeting ABC trans-porters projects a promising approach to eliminating or suppressing drug resistance in cancer treatment.To reveal the functional mechanisms of ABC transporters in drug resistance, extensive studies have beenconducted from identifying drug binding sites to elucidating structural dynamics. In this review arti-cle, we examined the recent crystal structures of ABC proteins to depict the functionally importantstructural elements, such as domains, conserved motifs, and critical amino acids that are involved inATP-binding and drug efflux. We inspected the drug-binding sites on ABC proteins and the molecularmechanisms of various substrate interactions with the drug binding pocket. While our continuous battleagainst drug resistance is far from over, new approaches and technologies have emerged to push forward
ext generation sequencingovel drug target
our frontier. Most recent developments in anti-MDR strategies include P-gp inhibitors, RNA-interference,nano-medicines, and delivering combination strategies. With the advent of the ‘Omics’ era – genomics,epigenomics, transcriptomics, proteomics, and metabolomics – these disciplines play an important rolein fighting the battle against chemoresistance by further unraveling the molecular mechanisms of drugresistance and shed light on medical therapies that specifically target MDR.
© 2016 Elsevier Ltd. All rights reserved.
ontents
1. Background . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152. MDR and ABC drug transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 15
2.1. ABC drug transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 152.2. Structures of ABC transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 162.3. Energy transduction of drug efflux mediated by ABC transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 172.4. Drug binding to P-gp . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
3. MDR transport inhibitors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.1. First generation MDR transporter inhibitors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183.2. Second generation MDR transporter inhibitors. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .18
3.3. Third generation MDR transporter inhibitors . . . . . . . . . . . . . . . . . . . . . .3.4. Natural compounds with a MDR reversal effect . . . . . . . . . . . . . . . . . . .3.5. Small molecules from versatile scaffolds . . . . . . . . . . . . . . . . . . . . . . . . . . .
∗ Corresponding author at: Department of Pharmaceutical Sciences, St. John’s Universit∗∗ Corresponding author at: Department of Pharmaceutical Sciences, St. John’s Universit
E-mail addresses: [email protected] (D.-H. Yang), [email protected] (Z.-S. Chen).
ttp://dx.doi.org/10.1016/j.drup.2016.05.001368-7646/© 2016 Elsevier Ltd. All rights reserved.
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
y, Queens, NY 11439, USA. Tel.: +1 718 990 1432; fax: +1 718 990 1877.y, Queens, NY 11439, USA. Tel.: +1 718 990 6468; fax: +1 718 990 1877.

W. Li et al. / Drug Resistance Updates 27 (2016) 14–29 15
4. Novel strategies to overcome MDR in cancer cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214.1. Inactivation of MDR-associated genes by microRNA and RNA interference . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214.2. Monoclonal antibodies targeting P-gp . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214.3. Development of novel anticancer drugs that are not P-gp substrates . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214.4. Nanotechnology-based approaches to overcome MDR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .21
4.4.1. Polymeric nanoparticles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214.4.2. Liposome nanoparticles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224.4.3. Micelles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 224.4.4. Mesoporous silica nanoparticles (MSNPs) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 234.4.5. Gold nanoparticles . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 23
4.5. Combinational strategies to overcome MDR. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .235. Identification of novel MDR biomarkers using ‘Omics’ technologies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 236. Conclusions and future perspectives . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
Conflicts of interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24Acknowledgement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
. . . . . .
1
idsttepliClca
uaaiGAAZtdhtGs1tKfc2(eeafitAhdgs
References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. Background
Multidrug resistance (MDR) is one of the major clinical imped-ments in cancer chemotherapy and treatment of infectiousiseases. Nearly 80% of patients develop drug resistance in hospitalettings, leading healthcare cost to skyrocket (Wang, 2014). Cen-er for Disease Control and Prevention (CDC) reported that morehan two million people are diagnosed with serious infectious dis-ases every year due to antibiotic resistance and about 20,000eople die due to failure of treatment (Prevention, 2015). Some
ife-threatening pathogens such as HIV, herpes simplex virus, andnfluenza are becoming increasingly resistant and difficult to treat.ancer cells that are originally sensitive to a single anti-cancer drug
ater become resistant to multiple anti-cancer drugs. Although aombination of drugs may effectively direct cancer cells towardpoptosis, this approach frequently results in undesired toxicity.
MDR can be caused by several mechanisms, such as reducedptake of drugs, altered cell cycle checkpoint and cell cycle arrest,ltered drug target, increased efflux of drugs by drug transporters,nd sequestration of anticancer drugs in lysosomes as well as inntracellular organelles and intercellular vesicles (Chai et al., 2010;oler-Baron and Assaraf, 2011; Goler-Baron et al., 2012; Gonen andssaraf, 2012; Ifergan et al., 2005; Lebedeva et al., 2012; Livney andssaraf, 2013; Raz et al., 2014; Zhitomirsky and Assaraf, 2015, 2016;hou et al., 2015). In cancer cells, the predominant cause of MDR ishe increased efflux of anti-cancer drugs by membrane-embeddedrug transporters. The most well-studied drug transporters inumans is P-glycoprotein (P-gp/MDR1/ABCB1), which belongs tohe ATP-Binding Cassette (ABC) superfamily (Ferry et al., 1992;ottesman and Pastan, 1993; Rosenberg et al., 1997). Other welltudied ABC drug transporters include multidrug resistance protein
(MRP1/ABCC1) (Muller et al., 1996), breast cancer resistant pro-ein (BCRP/MXR/ABCP/ABCG2) (Bram et al., 2007; Bram et al., 2009;awabata et al., 2001; Lage and Dietel, 2000; Shafran et al., 2005)
rom human and bacterial homologues such as LmrA from Lacto-occus lactis (Federici et al., 2007; van den Berg van Saparoea et al.,005), DrrAB doxorubicin transporter from Streptomyces peucetiusLi, 2014; Li et al., 2013a, 2014b; Pradhan et al., 2009; Zhangt al., 2015b), macrolide antibiotics transporter MacB (Kobayashit al., 2003; Piao et al., 2008), and MsbA from E. coli (Doerrlernd Raetz, 2002; Schmitt, 2002). In the present review article, werst present a systematic overview of the crystal structure, func-ional mechanism, and dynamic drug binding features of severalBC drug transporters. Using different models, we also describe
ow the energy transduction, triggered by ATP binding, promotesrug efflux to the extracellular milieu. Several advanced technolo-ies to combat MDR are presented and discussed with real casetudies. The current knowledge of MDR in cancer that we present. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 24
in the current review article will benefit the clinical therapies anddrug development in the future.
2. MDR and ABC drug transporters
2.1. ABC drug transporters
To date, at least 15 human ABC proteins, such as P-gp, MRP1,and BCRP, have been found to mediate MDR as drug efflux pumps.However, several recent studies reported that, besides drug efflux,these ABC proteins may further extend their role through othermechanisms (Breier et al., 2013; Videira et al., 2014). For example,an observed decrease in sensitivity to therapeutic substances (suchas cisplatin) that are not P-gp substrates in P-gp-overexpressingtumor cells suggested that ABC transporters may play additionalroles in regulatory pathways (Shen et al., 2012). Newly discoveredroles of ABC transporters include: (1) modifying the distributionof ABC transporters to intracellular (e.g., cytoplasmic vesicles) orextracellular (e.g., exosomes which enable the transformation ofdrug-sensitive cells to drug-resistant cells) compartments to fur-ther improve the efficiency of drug sequestration (Goler-Baron andAssaraf, 2011; Horio et al., 1988); (2) involvement in tumor cellproliferation, invasion, and defense against anti-tumor regulatorypathways, such as deregulating and inhibiting the apoptotic path-ways and complement-mediated cytotoxicity (Pallis and Russell,2000; Weisburg et al., 1999; Zhang et al., 2014). Both prokaryotesand eukaryotes share the minimal structure requirement of highlyconserved nucleotide binding domains (NBDs) and variable trans-membrane domains (TMDs) of ABC proteins. Some ABC proteinsexhibit these domains within a single polypeptide chain (P-gp)(Ambudkar et al., 1999), while others may be assembled from twohomodimeric (DrrAB) (Kaur et al., 2005) or heterodimeric polypep-tide chains (LmrCD) (Kast et al., 1995; Lubelski et al., 2006; Tusnadyet al., 1997).
P-glycoprotein, also known as ABCB1, is encoded by a singlepolypeptide chain with two homologous NBDs and two homolo-gous TMDs (Ambudkar et al., 1999; Ferry et al., 1992). The two NBDscontain conserved motifs such as Walker A, Walker B, ABC signa-ture, a glutamine loop (Q-loop), and a switch motif (Davidson et al.,2008; Sharom, 2011). These motifs form a large pocket for ATP-binding and hydrolysis, and it is generally believed that both NBDsare required for normal ATPase activity and drug transport function.However, it has been reported that NBD2 is partially dispensablefor channel activity in mammalian CFTR (Cui et al., 2007), while
ATP binding and hydrolysis may occur at two NBDs alternatively(Senior et al., 1995).P-gp consists of a single polypeptide with two NBDs and twoTMDs. Each TMD contains six transmembrane helices (TMH).

16 W. Li et al. / Drug Resistance Updates 27 (2016) 14–29
Fig. 1. Catalytic cycle of P-glycoprotein drug efflux. (1) “TMDs Open Inward/NBDs Open”: P-glycoprotein stays in the ground state, substrate binds at the cytoplasmic sideof the TMDs; (2) “TMDs Closed Inward/NBDs Dimerization”: two Mg-ATP molecules bind to the ATP binding pocket of the NBDs, which triggers the dimerization of twoN ATP stc olysise ns, 20
TTsc(aNAhbpbbeifbasatbcewa
2
fet3camospe
t
BDs as a ‘sandwich’ dimer; (3) “TMDs Open Outward/Drug Efflux”: binding of Mg-onformation, substrate is released to the extracellular environment; (4) “ATP Hydrnergy were provided for the NBD dissociation and reset to the ground state (Wilke
herefore, two TMDs form one large and flexible channel with 12MHs for drug binding and efflux (Jones and George, 1998). Aseen in Fig. 1, the drug efflux process is associated with dynamiconformational changes of the ABC drug transporter, including:1) dimerization of NBDs, (2) drug efflux, (3) ATP hydrolysis,nd (4) return to original state. Three highly conserved motifs inBDs, including Walker A, Walker B, and Signature C, form theTP-binding pocket and play a significant role in ATP-binding,ydrolysis, and energy transduction (Frelet and Klein, 2006). It iselieved that ATP-binding, not hydrolysis, drives substrate trans-ort from intracellular to the extracellular space. The linked regionsetween NBDs and TMDs are critical for the coupling between ATP-inding and drug efflux, as well as energy transduction (Callaghant al., 2006). Drugs, initially binding to the high-affinity drug bind-ng sites on the internal surface of the inward facing chamberormed by 12 TMHs, were subsequently moved to the low-affinityinding sites and released into the extracellular aqueous space (Liund Sharom, 1996). Unlike NBDs that share high homology irre-pective of substrate specificities in the ABC family, the sequencesnd structures of TMDs may vary significantly. TMDs appear to behe main determinants of specificity (Locher and Borths, 2004). Toetter understand the mechanism of drug resistance, it is critical toharacterize the crystal structure, the exact drug binding sites, andnergy transduction during drug binding and efflux. These studiesill help develop new chemotherapeutic inhibitors, modulators,
nd non-substrate anti-cancer drugs for cancer chemotherapy.
.2. Structures of ABC transporters
The first high resolution structure of ABC proteins was obtainedrom the NBD domain of HisP from Salmonella typhimurium (Hungt al., 1998). Subsequently, the crystal structure of the vitamin B12ransporter BtuCD from E.coli was determined at a resolution of.2 A (Locher and Borths, 2004; Locher et al., 2002). The BtuCDomplex (90 A × 60 A × 30 A) contains two copies of NBDs (BtuD)nd two copies of TMDs (BtuC). The NBDs sit directly below theembrane connecting to the TMDs. The TMDs with 10 �-helices
n each subunit form a flexible cavity structure and translocate theubstrate by alternatively opening or closing its cavity to either the
eriplasmic or cytoplasmic sides (Locher and Borths, 2004; Lochert al., 2002).MsbA is a half transporter with lipid flippase activity. It con-ains a single TMD fused to an NBD. The crystal structure of MsbA
imulates the conformational change of TMDs by switching from inward to outward”: after ATP hydrolysis, 2 ADP and 2 phosphates are released from the NBDs and the15).
was initially obtained in 2001 (Chang and Roth, 2001), although amore accurate structure with higher resolution was not obtaineduntil 2007 (Ward et al., 2007). Fig. 2A represents the updated struc-ture of MsbA from Salmonella typhimurium in a closed-apo form. Inthis structure, the two pairs of TMD/NBD subunits are connectedwith each other through a �-helix linker. The whole complex formsan open ‘V’ shape structure with a distance of approximately 50 A.Cross-linking experiments and Electron Paramagnetic Resonance(EPR) spectroscopy studies indicated that MsbA can form a largeand flexible chamber facing the cytoplasmic region to allow for theflipping of large molecules such as lipid A or lipopolysaccharide(LPS) from the cytoplasm to the exterior (Ward et al., 2007).
In 1997, Rosenberg et al., reported a low resolution (2.5 nm)structure of P-gp using electron microscopy (EM) single particleimage analysis of a purified protein (Rosenberg et al., 1997). Theprotein was found to form a six-fold symmetric toroid with a diam-eter of 10 nm. However, a higher resolution crystal structure of P-gpwas subsequently obtained by Aller et al. (2009). This structurewas based on nucleotide-free inward facing conformation with aresolution of 3.8 A. The two NBDs with 6 transmembrane heliceson each domain were separated by ∼30 A. They formed a largeinternal cavity of ∼6000 A3 that can accommodate at least two sub-strates simultaneously (Loo et al., 2003a). The drug binding pocketmainly consists of hydrophobic and aromatic residues, indicatingits preferential binding to polyspecific substrates and lipids (Alleret al., 2009). A more refined crystal structure of P-gp was laterobtained by the same group in 2014 using de novo model build-ing of selected regions (Li et al., 2014a). With a resolution of 3.8 A(Fig. 3A), the new model exhibited a more refined protein geometry,with more residues observed in favorable Ramachandran regionas well as corrected amino acid residues in drug binding sites. Byusing Fluorescence Resonance Energy Transfer (FRET) analysis, itwas shown that the cysteine residue in the signature sequence(427GNSGCGKS434) of one Walker A motif could cross-link withthe cysteine on the other Walker A motif (1070GSSGCGKS1077) inclose proximity with the residues of a downstream 1176LSGGQ1180
motif (Loo et al., 2002; Qu and Sharom, 2001; Urbatsch et al., 2001).This interaction indicated that the two ATP-binding sites partici-pate in formation of the ATP-binding pocket, and their interactions
are essential for coupling ATP hydrolysis to drug transport. Theclose view of these conserved motifs in both MsbA (PDB code: 3B60,AMPPNP-bound) and P-gp (PDB code: 4M1M, open-apo form) arepresented in Fig. 3B and C.
W. Li et al. / Drug Resistance Updates 27 (2016) 14–29 17
Fig. 2. Structural representations of MsbA from Salmonella typhimurium in close-apo form. (A) Overall structure with nucleotide-analog (AMPPNP) bound (PDB code:3B60, AMPPNP-bound); one subunit of the dimer is shown in magenta while theother subunit is in gray; the nucleotide analogs (AMPPNP) are in stick representa-tion; (B) close-up view of the AMPPNP-bound NBD domains, with one subunit inmagenta and the other in silver; the Walker A motifs (also known as P-loops) are incyan, the Walker B motifs are in yellow, the Q-loops are in blue, and the ABC signa-ture motifs are in green; the AMPPNPs are in orange stick. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web version oft
tdtmppatuMidwd2
Fig. 3. Structural representations of mouse P-glycoprotein in open-apo form. (A)Overall structure (PDB code: 4M1M) with the N-terminal half (residue 30–650) ofthe monomer is shown in magenta while the C-terminal half (residue 651–1270) isin gray; (B) close-up view of the substrate-free NBD domains; the Walker A motifs(also known as P-loops) are in cyan, the Walker B motifs are in yellow, the Q-loops
to the physical locations of subsequent events.
he article.)
With the advances of structural biology and the wider applica-ion of electron crystallography, 3D structure and conformationalynamics of several ABC proteins were updated. For example,he mechanism of maltotriose binding to the apo-open state of
altose-binding protein was recently updated by a new dockingrotocol-based 3D-RISM-KH theory. This result not only confirmedrediction of ligand-binding model from NMR spectroscopy, butlso provided new molecular details that were not observed inraditional X-ray crystallography (Huang et al., 2015). Using molec-lar dynamic simulations, open to close conformational changes ofcjD and CFTR during ATP-binding or ligand translocation were
llustrated (Gu et al., 2015b; Mornon et al., 2014). Moreover, threeimensional quantitative structure–activity relationship studiesere applied to quantify binding and stability of either anti-cancer
rugs or MDR inhibitors (Kakarla et al., 2016; Srivastava et al.,008).are in blue, and the ABC signature motifs are in green. (For interpretation of thereferences to color in this figure legend, the reader is referred to the web version ofthe article.)
2.3. Energy transduction of drug efflux mediated by ABCtransporters
The biochemical and biophysical bases of energy transductionof drug efflux mediated by ABC transporters remain controver-sial subjects, mainly due to the discrepancies between the twomajor mechanistic models proposed to describe these processes:the “ATP switch” model, and the “reciprocating twin-channel”model. Energy transduction takes place during the overall drugefflux, including ATP catalysis (which occurs in NBDs and can befurther dissected into ATP-binding and occlusion, ATP-hydrolysis,and ADP-Pi release) and the drug transport cycle (which occurs inTMDs where substrate binding, translocation from inner to outerleaflet, and drug release takes place). Therefore, the objective ofthese models is to explain how drug translocation is delicatelycoupled with ATP hydrolysis, by addressing the following key ques-tions: (1) the sequence of these steps; (2) conformational changesinduced by each step; and (3) how these signals are communicated
In the ATP switch model, it has been proposed thatdrug/substrate binding to the inward facing side of the TMD pro-motes the switch of two NBDs from the “open” to the “closed” dimer

1 nce U
cepihtfHfttthitAtco(taGak
2
csa2wts3abitbsumdLriRm1(
ivsctasiuMit
8 W. Li et al. / Drug Resista
onformation and increases the affinity to ATP (Linton, 2007; Reest al., 2009). This switch serves as the power stroke of drug trans-ort. The model also suggests that the “open” to “closed” transition
s driven by ATP-binding. Therefore, ATP-binding rather than ATP-ydrolysis creates the necessary conformational changes to inducehe repacking of TMDs and the translocation of drug/substraterom an inward facing state to an outward facing state (Linton andiggins, 2007). The role of ATP hydrolysis is to facilitate the switch
rom “closed” NBDs to its “open” state and restore the transportero resting state. This model highlights the close cross-talk betweenhe two halves of ABC transporters that are essential for forminghe ATP hydrolysis sites and drug translocation pathways. It alsoelps explain how substrate binding can stimulate ATP-binding
nstead of ATP hydrolysis in some ABC exporters. Conversely, inhe reciprocating twin-channel model, it has been proposed thatTP catalysis and substrate translocation will alternate between
he two halves of transporters, implying that the ATP hydrolysisycle at each NBD is coupled to a complete set of events in onef the two functionally separate substrate translocation pathwaysJones and George, 2014). Each NBD functions in a constant con-act scheme in which nucleotide triphosphate remains occluded in
closed site at all times until released from the NBD (Jones andeorge, 2009). This model provides insightful explanations for thellosteric effect of certain substrates on the binding and transportinetics of other substrates.
.4. Drug binding to P-gp
P-gp is able to interact with hundreds of structurally diversehemical compounds, including anticancer drugs, fluorescent dyes,teroid hormones, hydrophobic toxic peptides and lipids (Assarafnd Borgnia, 1994; Borgnia et al., 1996; Eytan et al., 1994; Ma et al.,015; Shapiro and Ling, 1998; Sharma et al., 1992). It is still unclearhether or not P-gp contains a single large drug binding site or mul-
iple sites. In 1997, Shapiro and Ling identified two drug bindingites on P-gp, an H-site that preferentially interacts with Hoechst3342 and the R-site observed to interact with Rhodamine 123nd anthracyclines (Shapiro and Ling, 1997). Later, a third drug-inding site with a positive allosteric effect on H and R sites was
dentified (Shapiro et al., 1999). In a subsequent related study, Mar-in et al., reported the use of equilibrium and kinetic radio-ligandinding assays to identify at least four distinct drug interactionites on P-gp (Martin et al., 2000). Additionally, Loo and Clarkesed a combination of cysteine-scanning mutagenesis and thiol-odification techniques to identify specific residues involved in
rug binding on P-gp (Loo and Clarke, 2001, 1999a, 1997, 2000;oo and Clarke, 2002). A list of residues within the transmembraneegion of membrane-embedded P-gp which affect substrate- bind-ng and transport were highlighted in Fig. 4B and listed in Fig. 4C.esidues in TM 4, 5, 6 in TMD1 and TM 9, 10, 11, 12 in TMD2 areainly involved in the formation of the drug binding pocket. Nearly
8% of amino acids involved in drug-binding are aromatic residuesLoo and Clarke, 1999b).
With a highly flexible drug-binding pocket, multiple chem-cal compounds can bind to P-gp simultaneously. For instance,erapamil and TMEA can bind to the different regions of thehared drug-binding pocket of P-gp (Loo et al., 2003a). Fluores-ence studies indicated that both LDS-751 and R123 can bind tohe R-site of P-gp simultaneously and interact with each other in
non-competitive manner (Lugo and Sharom, 2005). In anothertudy, mutation of residues that form hydrogen bonds with P-gpnhibitors caused reversal of the inhibitors, resulting in their stim-
latory effect on ATP binding and hydrolysis (Chufan et al., 2016).oreover, cysteine cross-linking assays revealed that the pack-ng of TM segments of P-gp was different for each substrate, andhis “induced-fit” type mechanism can explain how the binding
pdates 27 (2016) 14–29
pocket accommodates such a broad range of structurally diversecompounds (Loo et al., 2003b).
3. MDR transport inhibitors
To date, P-gp has proven to be able to interact with >200structurally unrelated chemical compounds that are classifiedas substrates and modulators (Didziapetris et al., 2003). Vari-ous substrates can be directly transported by P-gp, includinganthracyclines, Vinca alkaloids, and fluorescent lipids (Higgins andGottesman, 1992; van Helvoort et al., 1996). Modulators suchas verapamil and cyclosporine A can bind and block the trans-port function of the transporter, which presents significant clinicalapplications in cancer chemotherapy (Dong et al., 1996; Shuklaet al., 2012; Tinsley et al., 2010). P-gp contains a large flexibledrug-binding pocket with low specificity; development of morespecific, potent inhibitors/modulators are urgently needed to over-come MDR. Generally, inhibitors/modulators capable of reversingMDR can be divided into three generations (Kathawala et al., 2015),as summarized below, and a list of frequently used anti-cancerdrugs are summarized in Table 1.
3.1. First generation MDR transporter inhibitors
In 1981, Tsuruo’s group discovered that verapamil could reversethe profile of drug resistance in leukemia cells (Tsuruo et al.,1981). However, to acquire effective inhibition of MDR, patientsreceived high dosages that could result in serious cardiovasculartoxicity (Pennock et al., 1991). Other examples of early discoveryof inhibitors/modulators include cyclosporine A, trifluoperazine,quinidine, and progesterone (Kelly et al., 2011). Due to low thera-peutic response and high cell toxicity, these inhibitors were quicklyreplaced by a second generation of inhibitors (Bechtel et al., 2008;Koski et al., 2012; Thomas and Coley, 2003).
3.2. Second generation MDR transporter inhibitors
The second generation inhibitors were designed by structurallymodifying the first generation inhibitors in order to acquire higherpotency, specificity, and low cell toxicity. Examples include dexver-apamil (Wilson et al., 1995), valspodar (PSC 833) (Tidefelt et al.,2000), and biricodar citrate (VX-710) (Minderman et al., 2004).PSC833 is the most frequently used inhibitor with a potency5–10 times that of cyclosporine A (Friche et al., 1992). However,these transport inhibitors of P-gp are also frequently inhibitorsof cytochrome P450, and therefore, may cause drug-drug interac-tions with concomitant anticancer drugs (Gottesman et al., 2006;Kathawala et al., 2015; Leonard et al., 2003).
3.3. Third generation MDR transporter inhibitors
Continuous problems with MDR have driven the development ofthird generation MDR transporter inhibitors. At nanomolar concen-trations, these inhibitors may be more than 200-fold more potentat reversing MDR than first and second generation inhibitors,while exhibiting almost no pharmacokinetic interaction with otherchemotherapeutic drugs. Third generation inhibitors that havereached the stage of clinical trials include Tariquidar (XR9576)(Fox and Bates, 2007), Zosuquidar (LY335979) (Kemper et al.,2004), Laniquidar (R101933) (Luurtsema et al., 2009), Elacridar(F12091) (Dorner et al., 2009), Mitotane (NSC-38721) (Bateset al., 1991), Annamycin (Consoli et al., 1996), and R101933 (van
Zuylen et al., 2000). Tariquidar (XY9576) binds to P-gp with highaffinity and inhibits its ATPase activity at very low concentra-tions (25–80 nM) (Lubelski et al., 2006). Zosuquidar, an oral P-gpinhibitor, can significantly increase the uptake of daunorubicin,
W. Li et al. / Drug Resistance Updates 27 (2016) 14–29 19
Table 1List of some frequently used anti-cancer drugs.
Class Example Mechanism Trade name Target diseases Side effect Reference
Anthracyclines Doxorubicin Topoisomerase IIinhibitor
Adriamycin Leukemias, breast,stomach, lung, ovaries,bladder cancers,Hodgkin’s lymphoma
Cardiomyopathy,typhlitis
(Tacar et al., 2013)
Daunorubicin Topoisomerase IIinhibitor
Cerubidine Acute myeloidleukemia and acutelymphocytic leukemia
Cough or hoarseness,fever or chills,irregular heartbeat,lower back or sidepain
(Tan et al., 1967)
Epirubicin Topoisomerase IIinhibitor
Ellence Ovarian cancer, gastriccancer, lung cancer andlymphoma
Chills, fever, shock,urticaria, less sideeffect thanDoxorubicin
(Bonfante et al.,1980)
Alkylating agents Cisplatin DNA alkylation Platinol Sarcomas, somecarcinomas, bladdercancer, cervical cancerand germ cell tumors
Nephrotoxicity,neurotoxicity,ototoxicity
(Einhorn, 1990;Rosenberg et al.,1969)
Cyclophosphamide DNA alkylation Lyophilizedcytoxan Cancers, autoimmunedisorders, and ALamyloidosis
Acute myeloidleukemia, bladdercancer, hemorrhagiccystitis, andpermanent infertility
(Huttunen et al.,2011)
Antimetabolites Methotrexate(MTX)
Inhibit themetabolism of folicacid
Trexall Cancers, autoimmunediseases, ectopicpregnancy
Hepatotoxicity,ulcerative stomatitis,low white blood cellcount
(Goodsell, 1999)
Vinca alkaloids Vinblastine Inhibit mitosis N/A Hodgkin’s lymphoma,lung cancer, bladdercancer, brain cancer,and testicular cancer
Change in sensation,constipation,weakness, loss ofappetite, andheadaches
(Gobbi et al., 2003)
Vincristine Inhibit mitosis Oncovin Cancers Peripheralneuropathy,hyponatremia,constipation, and hairloss.
(Jordan, 2002)
Topoisomerase inhibitor Etoposide Topoisomerase IIinhibitor
Etopophos Kaposi’s sarcoma,Ewing’s sarcoma, lungcancer, testicularcancer, lymphoma,nonlymphocyti-cleukemia, andglioblastoma
Low blood pressure,hair loss, constipationand diarrhea, bonemarrow suppression
(Hande, 1998)
Taxanes Paclitaxel Microtubuleassembly inhibitor
Abraxane, Taxol Ovarian cancer, breastcancer, lung cancer andpancreatic cancer
Hair loss, muscle andjoint pains, anddiarrhea
(Saville et al., 1995)
Phenylalkylamine Verapamil L-type calciumchannel blocker
Isoptin, Verelan,Verelan PM,
Hypertension, anginapectoris, cardiacarrhythmia, and mostrecently, clusterheadaches
Constipation,headache, sinus pain,sore throat
(Bellamy, 1996;Martin et al., 1987;Srinivasan et al.,2011)
Class I antiarrhythmicagent
Quinidine Blocking the fastinward sodiumcurrent
Quinaglute, Quinidex Amyotrophic lateralsclerosis and multiplesclerosis. Plasmodiumfalciparum malaria.
Increased bloodlevels of lidocaine,beta blockers, opioids
(Brooks et al.,2004; Sadequeet al., 2000)
Tariquidar InhibitP-glycoprotein
XR9576 Phase III trials (Fox and Bates,2007)
Dactinomycin(Actinomycin D)
Inhibittranscription
Cosmegen Gestationaltrophoblasticneoplasia, Wilms’tumor,Rhabdomyosarcoma,Ewing’s sarcoma
Bone marrowsuppression, fatigue,hair loss, mouthulcer, loss of appetite
(D’Angio et al.,1981; Sobell, 1985)
Zosuquidar InhibitP-glycoprotein
LY335979 Solid tumors,leukemias, and othermalignancies
Reversible grade 1 or2 neurologic toxicity
(Morschhauseret al., 2007; Rubinet al., 2002)
Mitotane Alters steroidperipheralmetabolism
Lysodren Adrenocorticalcarcinoma
Anorexia and nausea,diarrhea, vomiting,decreased memoryand ability toconcentrate
(Gagliano et al.,2014)

20 W. Li et al. / Drug Resistance Updates 27 (2016) 14–29
Fig. 4. Drug binding site of the P-gp in open apo form. (A) Inward-facing conformation of P-gp with drug binding residues highlighted with red or blue color. Green representsa ts druga emb( rred to
i(ydstwndtiaepur
3
oa
nd N-terminal half and yellow represents the C terminal half of P-gp, red represent the C-terminal half of P-gp; (B) closer view of the drug binding sites at the transmFor interpretation of the references to color in this figure legend, the reader is refe
darubicin, mitoxantrone, and mylotarg in acute myeloid leukemiaTang et al., 2008). Recently, studies using QSAR and 3D-QSAR anal-ses have found that the structures of modulators are the maineterminants of their inhibitory function. Results of a comprehen-ive study of 32 anthranilamide derivatives of tariquidar indicatedhat a hydrophobic domain is required for the inhibitory function,hich includes a large aromatic ring system and a heteroatomear the anthranilamide nucleus at the other end of the tetrahy-roquinoline group (Pajeva and Wiese, 2009). However, althoughariquidar has been evaluated in clinical trials as a potent P-gpnhibitor, recent studies showed that tariquidar is both a substratend an inhibitor of P-gp depending on its in vivo dosage (Kannant al., 2011). Several tariquidar analogs, aiming to have higherotency and better stability than tariquidar, was characterizedsing 3D-QSAR (three dimensional quantitative structure-activityelationship) and contour map analysis (Kakarla et al., 2016).
.4. Natural compounds with a MDR reversal effect
Natural compounds or plant extracts can be potent inhibitorsf P-gp and they exhibit less cytotoxicity and better oral bioavail-bility. Polyphenolic compounds such as flavonoids or stilbenes
binding residues at the N-terminal half and blue represents drug binding residuesrane domain of P-gp; (C) list of drug binding residues and their binding substrates.
the web version of the article.)
extracted from Chinese herbal plants can reverse MDR causedby ABC drug transporters in vitro (Michalak and Wesolowska,2012). Neochamaejasmin B (NCB) extracted from the root of Steller-achamaejasme L. has been widely used as a traditional Chinesemedicine due to its antitumor, antivirus, and antibacterial activi-ties. RT-PCR and Western blot analyses have shown that NCB couldsuppress the expression of P-gp and inhibit P-gp-mediated drugefflux in a dose-dependent manner (Pan et al., 2015). Some naturalcompounds such as cytarabine, trabectedin, and halaven have alsobeen approved for clinical use, based on their strong MDR rever-sal activity (Abraham et al., 2010; Huang et al., 2014; Lopez andMartinez-Luis, 2014; Shi et al., 2007).
3.5. Small molecules from versatile scaffolds
A library of small molecules containing different scaffolds,such as tetralin, furoxane, and biphenyl, were designed to inhibitP-gp efflux activity. In vitro assays and in vivo PET studies
proved that co-administration of these novel molecules with anti-cancer drugs could increase the pharmacological activities ofanticancer drugs (Zinzi et al., 2014). Although co-administration ofP-gp inhibitors/modulators with anti-cancer drugs may effectively
nce U
rtPia
4
cm2
4i
oldcsfieseNmcNrf
sA2rric
4
1reew1wt(gOoMevc
4s
g
W. Li et al. / Drug Resista
everse drug efflux activity, no direct evidence has been presented,o date, that these inhibitors/modulators can alter the expression of-gp in normal tissue or tumor cells. Improved cellular permeabil-ty, bioavailability, and metabolism of these inhibitors/modulatorsre also major obstacles to clinical application.
. Novel strategies to overcome MDR in cancer cells
To solve these problems, novel strategies to overcome MDR inancer cells are being developed using biochemical and physicalethods as well as nanotechnologies (2014; Livney and Assaraf,
013; Shapira et al., 2012; Tiram et al., 2016).
.1. Inactivation of MDR-associated genes by microRNA and RNAnterference
MicroRNAs are small non-coding RNAs that bind to the 3′UTRf mRNA and inhibit the expression of protein at the translationalevel (Li et al., 2012b; Shen et al., 2013). MicroRNAs are generallyysregulated in cancer cells, and alterations in miRNA levels mayontribute to the development of MDR (To, 2013). For instance, aeries of miRNAs (miR-27a,-451, -296, -298, -1253) were identi-ed to inhibit the expression of P-gp, and their chemotherapeuticffects were evaluated in MCF-7 breast cancer cells and esophagealquamous carcinoma cells (Bao et al., 2012; Hong et al., 2010; Zhut al., 2008). To systematically understand how regulation of miR-As could contribute to the development of drug resistance, deepiRNAs sequencing of vincristine sensitive and -resistant colon
ancer cells were performed (Dong et al., 2015). Out of 24 miR-As showing significant differences, 17 were upregulated in drug
esistant cells. These miRNAs could be used as unique biomarkersor cancer diagnosis and treatment.
Synthetic siRNAs are also extensively used to reverse chemore-istance by inhibiting the expression of MDR genes, such asBCB1 (MDR1), ABCB4 (MDR3), BCRP (Fisher et al., 2007; Li et al.,012a; Perez et al., 2011; Wu et al., 2003). An example is theeduced expression level of ABCC4 using RNA interference in drug-esistant gastric cancer cells. This inhibition resulted in an effectivenhibition of cell proliferation, and increased cell sensitivity tohemotherapeutic drugs (Zhang et al., 2015a).
.2. Monoclonal antibodies targeting P-gp
In the 1980s, two monoclonal antibodies targeting P-gp, MRK-6 and MRK-17, were developed to effectively reverse drugesistance mediated by P-gp both in vivo and in vitro (Broxtermant al., 1988; Hamada and Tsuruo, 1986; Pearson et al., 1991; Tsuruot al., 1989). MRK-16 blocked actinomycin D and vincristine efflux,hile MRK-17 inhibited the proliferation of MDR cells. Later in
992, Roninson et al., designed a mouse monoclonal antibody UIC2hich specifically binds to a cell surface epitope of P-gp. It inhibited
he drug efflux activity of P-gp and enhanced cellular cytotoxicityMechetner and Roninson, 1992). Monoclonal antibodies in conju-ation with P-gp-reversing agents can enhance anti-cancer activity.ne example is the study from the Naito’s group; the combinationf MRK-16 and cyclosporine A increased the chemosensitivity ofDR K562/ADM cells to vincristine and doxorubicin, leading to an
nhanced anti-tumor activity than monotreatment of the cells withincristine and doxorubicin alone (Naito et al., 1993). MRK-16 isurrently being evaluated in clinical phase I trials.
.3. Development of novel anticancer drugs that are not P-gp
ubstratesAs many anti-tumor agents including the first and the secondeneration inhibitors of P-gp, are also substrates of the cytochrome
pdates 27 (2016) 14–29 21
P450 isoenzyme 3A4, competition between these P-gp modulatorsand this cytochrome P450 isoenzyme 3A4 can cause seriously toxicpharmacokinetic interactions (Thomas and Coley, 2003). Therefore,new anticancer drugs that are not recognized by P-gp and/or otherABC transporters are being developed. For example, taxane analogsDJ-927 (Shionoya et al., 2003), BMS-184476 (Altstadt et al., 2001),and ortataxel (Cassinelli et al., 2002) are not substrates of P-gp.These drugs have been evaluated in clinical trials for their MDRevasion activity in both sensitive and MDR tumor cell lines. Like-wise, indirubin, a Chinese traditional medicine, is not transportedby P-gp, but may significantly inhibit the efflux of doxorubicin andvincristine by P-gp (Su et al., 2012).
4.4. Nanotechnology-based approaches to overcome MDR
Nanoparticles are used extensively to deliver anti-tumor,anti-infection, or anti-inflammation chemicals to specific targetcells/tissues of patients. The size of NPs varies between 1 to sev-eral hundred nm. Examples of NPs include polymers, solid lipids,metals, quantum dots, dendrimers, liposomes, and micelles (Bar-Zeev et al., 2016; Livney and Assaraf, 2013; Shapira et al., 2012;Sun et al., 2014; Yang et al., 2016; Zhang and Akbulut, 2011). TheseNPs can be assembled in multiple layers, and the surface coating isalways the most crucial step for the specificity, stability, and solu-bility of these NPs (Cho et al., 2008; Zhang et al., 2016). NPs loadedwith anti-cancer drugs, P-gp inhibitors or RNAi molecules avoidproblems associated with macromolecules, such as high dosage,cell toxicity, low specificity, cellular uptake, and intracellular deliv-ery (Kapse-Mistry et al., 2014; Li et al., 2013b; Wen and Meng,2014). A schematic overview of the internalization and delivery ofdrugs or miRNA loaded-NPs into drug-resistant cells is presented inFig. 5. Compared to freely diffusing hydrophobic drugs, drug-loadedNPs have higher solubility, in vivo stability, increased specific bio-distribution and better control of drug release (Kim et al., 2009;Zhang et al., 2011). Moreover, combination of multiple anti-tumorcomponents (such as anti-tumor drug and miRNA), as shown inFig. 5, was shown to be a more efficient strategy in overcomingMDR (Gandhi et al., 2014; Kang et al., 2015).
4.4.1. Polymeric nanoparticlesBio-degradable polymeric NPs are the most commonly used
nanovehicles for drug delivery. Polymers may be natural or syn-thetic. Frequently used natural polymers include gelatin, chitosan,and albumin. Commonly used bio-degradable synthetic poly-mers include poly (d,l-lactic acid) (PLA), poly(d,l-lactic-co-glycolicacid) (PLGA), and poly (�-caprolactone) (PLC) (Devulapally andPaulmurugan, 2014; Li et al., 2015). Various techniques have beenused to form polymeric NPs, such as salting-out, dialysis, micro-emulsion, interfacial polymerization, solvent evaporation, andsupercritical fluid technology (SCF) (Harsha, 2012). Co-polymerscan be formed by combining different polymers, such as PLA-block-PEG co-polymers (Ben-Shabat et al., 2006).
Albumin is a plasma protein used in drug delivery systems.Abraxane, also known as Nab-paclitaxel, is formed through con-jugation of the anti-microtubule drug paclitaxel to human albuminNPs. It has been approved for clinical use by the US Food and DrugAdministration. Abraxane1 has many advantages over paclitaxel,including longer blood circulation time and lower hypersensitiv-ity (Gradishar, 2006). Clinical trials have shown that Abraxane hashigher efficacy than paclitaxel for metastatic breast cancer and pan-creatic cancer (Gradishar, 2006; Von Hoff et al., 2013).
Cisplatin has been frequently used in chemotherapy for prostate
cancers. However, its clinical utility has been hampered by drugresistance phenomena and toxicity to healthy tissues. A PLGA-b-PEG NP with cell surface anchored aptamers (Apt) targetingprostate-specific membrane antigen (PSMA) has been used to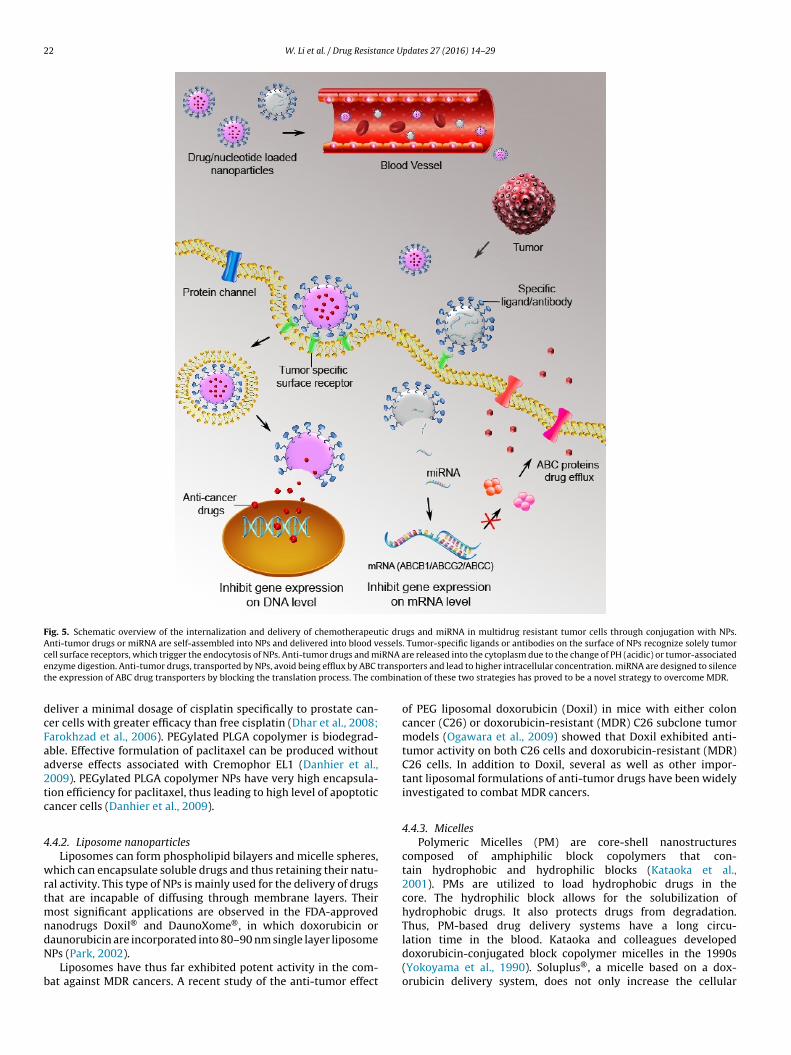
22 W. Li et al. / Drug Resistance Updates 27 (2016) 14–29
Fig. 5. Schematic overview of the internalization and delivery of chemotherapeutic drugs and miRNA in multidrug resistant tumor cells through conjugation with NPs.Anti-tumor drugs or miRNA are self-assembled into NPs and delivered into blood vessels. Tumor-specific ligands or antibodies on the surface of NPs recognize solely tumorc iRNA ae ranspt mbina
dcFaa2tc
4
wrtmndN
b
ell surface receptors, which trigger the endocytosis of NPs. Anti-tumor drugs and mnzyme digestion. Anti-tumor drugs, transported by NPs, avoid being efflux by ABC the expression of ABC drug transporters by blocking the translation process. The co
eliver a minimal dosage of cisplatin specifically to prostate can-er cells with greater efficacy than free cisplatin (Dhar et al., 2008;arokhzad et al., 2006). PEGylated PLGA copolymer is biodegrad-ble. Effective formulation of paclitaxel can be produced withoutdverse effects associated with Cremophor EL1 (Danhier et al.,009). PEGylated PLGA copolymer NPs have very high encapsula-ion efficiency for paclitaxel, thus leading to high level of apoptoticancer cells (Danhier et al., 2009).
.4.2. Liposome nanoparticlesLiposomes can form phospholipid bilayers and micelle spheres,
hich can encapsulate soluble drugs and thus retaining their natu-al activity. This type of NPs is mainly used for the delivery of drugshat are incapable of diffusing through membrane layers. Their
ost significant applications are observed in the FDA-approvedanodrugs Doxil® and DaunoXome®, in which doxorubicin or
aunorubicin are incorporated into 80–90 nm single layer liposomePs (Park, 2002).Liposomes have thus far exhibited potent activity in the com-at against MDR cancers. A recent study of the anti-tumor effect
re released into the cytoplasm due to the change of PH (acidic) or tumor-associatedorters and lead to higher intracellular concentration. miRNA are designed to silencetion of these two strategies has proved to be a novel strategy to overcome MDR.
of PEG liposomal doxorubicin (Doxil) in mice with either coloncancer (C26) or doxorubicin-resistant (MDR) C26 subclone tumormodels (Ogawara et al., 2009) showed that Doxil exhibited anti-tumor activity on both C26 cells and doxorubicin-resistant (MDR)C26 cells. In addition to Doxil, several as well as other impor-tant liposomal formulations of anti-tumor drugs have been widelyinvestigated to combat MDR cancers.
4.4.3. MicellesPolymeric Micelles (PM) are core-shell nanostructures
composed of amphiphilic block copolymers that con-tain hydrophobic and hydrophilic blocks (Kataoka et al.,2001). PMs are utilized to load hydrophobic drugs in thecore. The hydrophilic block allows for the solubilization ofhydrophobic drugs. It also protects drugs from degradation.Thus, PM-based drug delivery systems have a long circu-
lation time in the blood. Kataoka and colleagues developeddoxorubicin-conjugated block copolymer micelles in the 1990s(Yokoyama et al., 1990). Soluplus®, a micelle based on a dox-orubicin delivery system, does not only increase the cellular
nce U
cd
4
gdasCle
4
tfaaethgnthdc
4
ochRNfpltswM(
rtTnbeptNawa(cnobg(
W. Li et al. / Drug Resista
oncentration of doxorubicin, but also reverses P-gp-mediatedrug efflux (Jin et al., 2015).
.4.4. Mesoporous silica nanoparticles (MSNPs)Recently, mesoporous silica materials (MSNPS) have gained a
reat deal of attention due to their superior advantages for drugelivery, such as biocompatibility, large pore volume, high surfacerea, and tunable pore size. MSNPS have been used to load bothiRNA and anticancer drugs simultaneously (Chen et al., 2009).o-delivery of two types of anti-tumor molecules to cancer cell
ines could effectively inhibit the expression of P-gp, thus greatlynhancing intracellular drug concentrations (Sheng et al., 2015).
.4.5. Gold nanoparticlesGold NPs (AuNPs) have been investigated as suitable bioma-
erials for drug delivery. They can be easily synthesized andunctionalized (Wu et al., 2010). In addition, they are non-toxicnd biocompatible. The surface modification of gold NPs providesdvantages of tumor targeting and controllable drug release (Wut al., 2010; Yang et al., 2011). In addition, they can overcomehe problem of MDR (Xu et al., 2006). For example, Lee et al.,ave designed Dox-PLGA-Au NPs with anti-DR4-antibody conju-ated onto the Au surface. NPs-loaded tumor cells treated withear infrared (NIR) light for 10 min could reach an intracellularemperature of 45 ◦C. Highly effective reduction of tumor growthas been observed after hyperthermia. In addition, much lowerosage of doxorubicin is required to achieve the same effect of thehemotherapy (Lee et al., 2014).
.5. Combinational strategies to overcome MDR
Even though RNAi is capable of reversing MDR, effective deliveryf RNA-interfering agents to the targeted cancer cells still remainshallenging. To overcome this hurdle, Lage’s group delivered a shortairpin RNA (shRNA) into target cancer cells by TransKingdomNAi (tkRNAi) (Kruhn et al., 2009; Lage and Kruhn, 2010). In tkR-Ai, an expression vector TRIP is incorporated with the following
our essential elements: the shRNA of interest, bacteriophage T7romoter, Inv gene encoding invasion, and HlyA gene expressing
isteriolysin O. TRIP is then incorporated into non-pathogenic bac-eria and delivered into target cells to induce RNAi. This treatmenthowed approximately 45% inhibition of ABCB1 mRNA expressionith ∼11-fold increase in drug accumulation in P-gp-dependentDR cancer cells, compared to treatment with the drug alone
Kruhn et al., 2009).In another novel gene delivery system, RNAi was incorpo-
ated into NPs with anti-CD44 monoclonal antibody coated athe surface, forming a nano-complex named anti-CD44-P123-PPI.he CD44 monoclonal antibody, anchored at the surface of thisano-complex, could not only increase the transfection efficiency,ut also lower the cytotoxicity. In vitro studies reported that annhanced inhibition of MDR1/P-gp expression at both mRNA androtein levels was observed for this nano-complex than with tradi-ional PPI/pDNA NPs (Gu et al., 2015a). Similarly, CD44-targetedPs based on cholesteryl-hyaluronic acid (HA) conjugates havelso been designed to deliver siRNA into MDR ovarian cancer cellsith significant inhibitory effect on tumor growth, P-gp expression,
nd increased apoptosis in mice harboring MDR ovarian cancersYang et al., 2015). Rachur’s group designed a protamine andarboxymethyl cellulose polyelectrolyte multi-layered magneticanocapsules with Fe3O4 anchored at the surface. In the presence
f a magnetic field, this magnetic nanocapsule loaded with doxoru-icin showed enhanced cellular uptake of anti-cancer drugs withreater probability of inducing apoptosis than other cytotoxic drugsElumalai et al., 2015).pdates 27 (2016) 14–29 23
Peptide self-assembly system is an emerging technology for theefficient delivery and retention of anti-cancer drugs or antibodiesinto cancer cells (Wen et al., 2013, 2014a,b; Zheng et al., 2011). Byforming an amphiphilic environment by the peptides, hydrophobicanticancer drugs or antibodies were loaded onto the core througharomatic hydrophobic interactions (Sundar et al., 2014). The speci-ficity of peptide-assembled nanostructures could be modulated byaltering the peptide sequence on the surface; therefore efficientdelivery and more than 120 h retention of immune-antibody intumor cells were observed (Wen et al., 2013).
In recent years, some basic structural components of plasmamembranes, such as ceramide, sphingosines and caveolin, wereshown to be involved in cancer MDR (Mitra et al., 2006; van Vlerkenet al., 2007). For example, caveolin-1 is the major component ofcaveolae membrane coat which has been found to be essential forthe ATPase activity and functionality of P-gp. Studies by Zhu et al.showed that overexpression of caveolin-1 results in the reductionof plasma membrane cholesterol levels, which further increasesplasma membrane fluidity, thus blocking P-gp transport activity(Cai et al., 2004; Zhu et al., 2004).
5. Identification of novel MDR biomarkers using ‘Omics’technologies
The development of advanced genomics and proteomicstechnologies in the last decade significantly revolutionized theparadigm of drug target discovery in cancer chemotherapy (Li,2015). Nowadays, deciphering the heterogeneous responses ofeach individual patient to anticancer drugs can be a major obsta-cle in clinical diagnostics and therapeutics (Liu et al., 2015b; Shenget al., 2015). Precision oncology data from individual patients arerequired to determine the cause of cancer (Morrison et al., 2014).Rather than treating all the patients harboring a given malignancywith the same statistics-based protocol, personalized treatmentmodalities are required for each individual. This will not onlyincrease the therapeutic efficacy, but also bring down the medicalcost.
Next generation sequencing (NGS) techniques allow the deter-mination of the complete genomic profile of each individual orquantitatively sequencing hundreds of cancer-related genes in afew hours. The most commonly used NGS platforms include Illu-mina (Bentley et al., 2008; Fedurco et al., 2006; Turcatti et al., 2008),SMRT (Chhibber and Schroeder, 2008), SOLiD (Shendure et al.,2005), and Roche 454 (Margulies et al., 2005; Turcatti et al., 2008).Third generation sequencing platforms, aiming to avoid error ofbase-pairing during PCR, are currently under development. Exam-ples of these include nanopore DNA sequencing (Stoddart et al.,2009) and mass spectrometry sequencing (Edwards et al., 2005).
Novel anti-cancer drug targets or MDR biomarkers could bepredicted by comparing the genetic mutation rate of high through-put genomic data from cancer patients. A set of small moleculesand synthetic peptides or antibodies could be selected by com-putational analyses, such as SVM (Support Vector Machines), andthe results can be validated experimentally (Jeon et al., 2014). Forexample, massive parallel sequencing was performed with hun-dreds of breast cancer samples in order to study the correlationbetween somatic mutations and breast cancer. Stephens et al. (143)reported somatic mutations in at least 40 cancer genes, includingseveral new genes such as AKT2, ARID1B, CASP8, CDKN1B, MAP3K1,MAP3K13, NCOR1, SMARCD1, and TBX3 (Stephens et al., 2012).In the study of upregulated and down-regulated chemoresistance
genes in ovarian clear cell carcinoma, NAPSA, GPX3 and ALDH1A1were found to be remarkably overexpressed in tumor cells, and theyare now considered to be new biomarker candidates (Kobayashiet al., 2015). In another comprehensive data mining study, reliable
2 nce U
bcaatAe(mdtmticm
cagtsnttTpp(ce
tsTtpliruadIttcan
6
fetctrrsduow
4 W. Li et al. / Drug Resista
iomarkers for both chemosensitive and chemoresistant tumorells were identified by screening human tumor samples against
large range of drug responses that correlated with genomic vari-tions (Sadanandam and Covell, 2015). Sensitivity to cisplatin inumor cells is associated with mutations in EWS FLI1, PTEN, ERBB2,PC, XRCC1 and KRAS (Goel et al., 2012; Niedner et al., 2001; Xut al., 2014). They also reported the global analysis of the CGPSanger Cancer Genomic Project) data, which described the genetic
utations, copy number variations and gene fusions related to 41rugs’ responses. From the heatmap of drug-mutation combina-ions across all the compounds tested in CGP database, CDKN2a.p14
utation was found in a 24-drug response test, while a single muta-ion was found in SMAD4, MDM2, EGFR and CDH1. These datandicated that genetic mutations may be shared across distinct anti-ancer drugs from different classes, and multiple genetic mutationsay contribute to the variable response to drug treatment.While powerful sequencing technologies allow us to explore
ancer biology from a genetic standpoint with better timeline andccuracy, they also have some challenges. For example, wholeenome sequencing data have insufficient clinical annotation dueo the existence of long non-coding RNA, intron–exon, repetitiveequences, and pseudogenes. Moreover, synonymous substitutioneither results in a change of amino acid sequence of a protein, norhe structure of the protein. Epigenetic modifications may also con-ribute to the alteration of drug response pathways (Bird, 2007).herefore, proteomics studies, aiming to identify the function ofroteins at large scale both in quantity and quality, is crucial forharmacological studies and personalized drug target discoveryJain, 2002; Koomen et al., 2008). In the study of MDR in can-er cells, proteomics analysis can identify the difference in proteinxpression between drug sensitive and drug resistant tumor cells.
In the comparative proteomics study of colon cancer stem cells,otal proteins extracted from cells were digested into peptides andubjected to liquid chromatography mass spectrometry (LS–MS).he LS–MS/MS spectral data was searched against human interna-ional protein index database using Sequest software. Out of 3048roteins identified, 32 proteins were found to be up-regulated by at
east 2-fold in cancer cells (Van Houdt et al., 2011); these proteinsncluded glutathione peroxidase 2 (GPX2), ferritin (FTL), and RRS2ibosome biogenesis regulator (RRS1). One of the most interestingp-regulated proteins was BIRC6. The latter belongs to the class ofpoptotic protein inhibitors, and is critical in conferring anticancerrug resistance to cisplatin and oxaliplatin (Van Houdt et al., 2011).
n another study on human ovarian cancer, isotope-coded affinityag mass spectrometry (ICAT-MS) identified a total of 1117 pro-eins, from which 58 proteins were overexpressed in drug resistantells (Van Houdt et al., 2011). Some proteins such as CASPR3, S100,nd CDC42-binding protein kinase � may be involved in mecha-isms of drug resistance (Stewart et al., 2006).
. Conclusions and future perspectives
Clinical evidence reveals that >90% of metastatic cancer patientsace treatment failure due to drug resistance phenomena (Longleyt al., 2006). Hence, MDR poses a major challenge to effec-ive/successful chemotherapy in cancer treatment. As significantontributors that render cancer cell drug-resistance, ABC pro-eins belong to one of the largest protein superfamily that giveesearchers hope to unravel the molecular mechanism of drugesistance. Despite concentrated efforts made in better under-tanding MDR, it is still hampered by several impediments. Firstly,
ue to the instability of membrane protein in vitro, it is unfeasible tose a single type of purification method to extract all the subunitsf the protein with native conformation. Most membrane proteinsill be misfolded, degraded or aggregated without appropriatepdates 27 (2016) 14–29
supply of detergents during the purification process. Secondly, theexpression levels of MDR-related proteins may vary significantly;some proteins are expressed at extremely low levels while oth-ers are overexpressed. As traditional biochemical approaches (suchas SDS-PAGE and 2-DE) are capable of detecting/quantifying pro-teins at a certain quantity, they are generally not sensitive enoughfor extremely low abundance membrane proteins. Finally, whenpotent MDR inhibitors, such as tyrosine kinase inhibitors, tariq-uidar, were used to induce apoptosis in tumor cells, severe drugtoxicity to normal tissues occurred. All of these factors remain trueobstacles; nevertheless, researchers started taking advantages ofnewly emerging technologies to explore bypasses or alternatives.
Besides genetic mutations, MDR in cancer cells could also beinduced by epigenetic factors, such as protein expression vari-ations, post-translational modifications, or changes in signalingpathways. Whole exome and bisulfite sequencing help identifynovel genetic and epigenetic biomarkers related to the devel-opment of drug resistance (Chang et al., 2010; Glasspool et al.,2006; Henrique et al., 2013). Pharmacogenomics combined withgenomics and epigenetics, transcriptomics, proteomics as well asmetabolic data, are currently used to identify genetic diversities ofdrug metabolic pathways (Huang and Ratain, 2009; Weng et al.,2013). The unique and novel biomarkers identified through thisapproach could be used for cancer diagnostics and prediction ofpharmacotherapy. Recent studies have found that several signalingpathways are involved in drug resistance, including protein tyro-sine kinase (Koch et al., 2015; Rexer et al., 2011; Stebbing et al.,2015), mTOR (Jiang and Liu, 2008; Vo et al., 2014), NF-kB (Bentires-Alj et al., 2003; Mongre et al., 2015), EGFR (Porcelli et al., 2014;Song et al., 2015), MAPK (mitogen-activated protein kinase) (Liuet al., 2015a; Wang et al., 2015), and Notch signaling pathways(Cho et al., 2011; Kajiyama et al., 2007; Li et al., 2009). To trace howthe ABC superfamily of proteins evolved, Miao’s group performedcomparative genomics and phylogenetic analysis based on wholegenome sequencing data of different eukaryotic species. The puta-tive evolutionary pathway of ABC transporters showed how genemutations, duplications, and gene fusions triggered the generationof new functions of ABC transporter proteins (Xiong et al., 2015).
In this review article, we summarized the most current studiesof ABC drug transporters and their contribution to MDR in cancer.The crystal structures and dynamic nature of ABC proteins duringATP binding, hydrolysis, and drug efflux were discussed in detail.Conserved domains, motifs, and amino acid residues involved inthe catalytic function were highlighted. Questions remain on howdifferent drug substrates bind to the drug binding pocket and howthey synergistically perform drug efflux function. By integratingexperimental data emerging from OMICs technologies into clinicalpractice, these studies should greatly benefit clinical therapies anddrug development toward overcoming cancer MDR.
Conflicts of interest
The authors have declared no potential conflicts of interest.
Acknowledgement
This work was partially supported by funds from the RayBiotechINC (Norcross, GA).
References
Abraham, I., Jain, S., Wu, C.-P., Khanfar, M.A., Kuang, Y., Dai, C.-L., Shi, Z., Chen, X., Fu, L.,Ambudkar, S.V., Sayed, K.E., Chen, Z.-S., 2010. Marine sponge-derived sipholanetriterpenoids reverse P-glycoprotein (ABCB1)-mediated multidrug resistance incancer cells. Biochem. Pharmacol. 80, 1497–1506.

nce U
A
A
A
A
B
B
B
B
B
B
B
B
BB
B
B
B
B
B
W. Li et al. / Drug Resista
ller, S.G., Yu, J., Ward, A., Weng, Y., Chittaboina, S., Zhuo, R., Harrell, P.M., Trinh, Y.T.,Zhang, Q., Urbatsch, I.L., Chang, G., 2009. Structure of P-glycoprotein reveals amolecular basis for poly-specific drug binding. Science 323, 1718–1722.
ltstadt, T.J., Fairchild, C.R., Golik, J., Johnston, K.A., Kadow, J.F., Lee, F.Y., Long, B.H.,Rose, W.C., Vyas, D.M., Wong, H., Wu, M.J., Wittman, M.D., 2001. Synthesis andantitumor activity of novel C-7 paclitaxel ethers: discovery of BMS-184476. J.Med. Chem. 44, 4577–4583.
mbudkar, S.V., Dey, S., Hrycyna, C.A., Ramachandra, M., Pastan, I., Gottesman, M.M.,1999. Biochemical, cellular, and pharmacological aspects of the multidrug trans-porter. Annu. Rev. Pharmacol. Toxicol. 39, 361–398.
ssaraf, Y.G., Borgnia, M.J., 1994. Probing the interaction of the multidrug-resistancephenotype with the polypeptide ionophore gramicidin D via functional channelformation. Eur. J. Biochem./FEBS 222, 813–824.
ao, L., Hazari, S., Mehra, S., Kaushal, D., Moroz, K., Dash, S., 2012. Increased expres-sion of P-glycoprotein and doxorubicin chemoresistance of metastatic breastcancer is regulated by miR-298. Am. J. Pathol. 180, 2490–2503.
ar-Zeev, M., Assaraf, Y.G., Livney, Y.D., 2016. Beta-casein nanovehicles for oraldelivery of chemotherapeutic drug combinations overcoming P-glycoprotein-mediated multidrug resistance in human gastric cancer cells. Oncotarget, http://dx.doi.org/10.18632/oncotarget.8019.
ates, S.E., Shieh, C.Y., Mickley, L.A., Dichek, H.L., Gazdar, A., Loriaux, D.L., Fojo, A.T.,1991. Mitotane enhances cytotoxicity of chemotherapy in cell lines expressinga multidrug resistance gene (mdr-1/P-glycoprotein) which is also expressed byadrenocortical carcinomas. J. Clin. Endocrinol. Metab. 73, 18–29.
echtel, L.K., Haverstick, D.M., Holstege, C.P., 2008. Verapamil toxicity dysregulatesthe phosphatidylinositol 3-kinase pathway. Acad. Emerg. Med. 15, 368–374.
ellamy, W.T., 1996. P-glycoproteins and multidrug resistance. Annu. Rev. Pharma-col. Toxicol. 36, 161–183.
en-Shabat, S., Kumar, N., Domb, A.J., 2006. PEG-PLA block copolymer as potentialdrug carrier: preparation and characterization. Macromol. Biosci. 6, 1019–1025.
entires-Alj, M., Barbu, V., Fillet, M., Chariot, A., Relic, B., Jacobs, N., Gielen, J.,Merville, M.-P., Bours, V., 2003. NF-�B transcription factor induces drug resis-tance through MDR1 expression in cancer cells. Oncogene 22, 90–97.
entley, D.R., Balasubramanian, S., Swerdlow, H.P., Smith, G.P., Milton, J., Brown, C.G.,Hall, K.P., Evers, D.J., Barnes, C.L., Bignell, H.R., Boutell, J.M., Bryant, J., Carter, R.J.,Keira Cheetham, R., Cox, A.J., Ellis, D.J., Flatbush, M.R., Gormley, N.A., Humphray,S.J., Irving, L.J., Karbelashvili, M.S., Kirk, S.M., Li, H., Liu, X., Maisinger, K.S., Mur-ray, L.J., Obradovic, B., Ost, T., Parkinson, M.L., Pratt, M.R., Rasolonjatovo, I.M.,Reed, M.T., Rigatti, R., Rodighiero, C., Ross, M.T., Sabot, A., Sankar, S.V., Scally,A., Schroth, G.P., Smith, M.E., Smith, V.P., Spiridou, A., Torrance, P.E., Tzonev,S.S., Vermaas, E.H., Walter, K., Wu, X., Zhang, L., Alam, M.D., Anastasi, C., Aniebo,I.C., Bailey, D.M., Bancarz, I.R., Banerjee, S., Barbour, S.G., Baybayan, P.A., Benoit,V.A., Benson, K.F., Bevis, C., Black, P.J., Boodhun, A., Brennan, J.S., Bridgham, J.A.,Brown, R.C., Brown, A.A., Buermann, D.H., Bundu, A.A., Burrows, J.C., Carter, N.P.,Castillo, N., Chiara, E.C.M., Chang, S., Neil Cooley, R., Crake, N.R., Dada, O.O., Diak-oumakos, K.D., Dominguez-Fernandez, B., Earnshaw, D.J., Egbujor, U.C., Elmore,D.W., Etchin, S.S., Ewan, M.R., Fedurco, M., Fraser, L.J., Fuentes Fajardo, K.V., ScottFurey, W., George, D., Gietzen, K.J., Goddard, C.P., Golda, G.S., Granieri, P.A., Green,D.E., Gustafson, D.L., Hansen, N.F., Harnish, K., Haudenschild, C.D., Heyer, N.I.,Hims, M.M., Ho, J.T., Horgan, A.M., Hoschler, K., Hurwitz, S., Ivanov, D.V., Johnson,M.Q., James, T., Huw Jones, T.A., Kang, G.D., Kerelska, T.H., Kersey, A.D., Khreb-tukova, I., Kindwall, A.P., Kingsbury, Z., Kokko-Gonzales, P.I., Kumar, A., Laurent,M.A., Lawley, C.T., Lee, S.E., Lee, X., Liao, A.K., Loch, J.A., Lok, M., Luo, S., Mammen,R.M., Martin, J.W., McCauley, P.G., McNitt, P., Mehta, P., Moon, K.W., Mullens,J.W., Newington, T., Ning, Z., Ling Ng, B., Novo, S.M., O’Neill, M.J., Osborne, M.A.,Osnowski, A., Ostadan, O., Paraschos, L.L., Pickering, L., Pike, A.C., Pike, A.C., ChrisPinkard, D., Pliskin, D.P., Podhasky, J., Quijano, V.J., Raczy, C., Rae, V.H., Rawlings,S.R., Chiva Rodriguez, A., Roe, P.M., Rogers, J., Rogert Bacigalupo, M.C., Romanov,N., Romieu, A., Roth, R.K., Rourke, N.J., Ruediger, S.T., Rusman, E., Sanches-Kuiper,R.M., Schenker, M.R., Seoane, J.M., Shaw, R.J., Shiver, M.K., Short, S.W., Sizto, N.L.,Sluis, J.P., Smith, M.A., Ernest Sohna Sohna, J., Spence, E.J., Stevens, K., Sutton, N.,Szajkowski, L., Tregidgo, C.L., Turcatti, G., Vandevondele, S., Verhovsky, Y., Virk,S.M., Wakelin, S., Walcott, G.C., Wang, J., Worsley, G.J., Yan, J., Yau, L., Zuerlein, M.,Rogers, J., Mullikin, J.C., Hurles, M.E., McCooke, N.J., West, J.S., Oaks, F.L., Lund-berg, P.L., Klenerman, D., Durbin, R., Smith, A.J., 2008. Accurate whole humangenome sequencing using reversible terminator chemistry. Nature 456, 53–59.
ird, A., 2007. Perceptions of epigenetics. Nature 447, 396–398.onfante, V., Bonadonna, G., Villani, F., Martini, A., 1980. Preliminary clinical expe-
rience with 4-epidoxorubicin in advanced human neoplasia. Recent ResultsCancer Res. 74, 192–199.
orgnia, M.J., Eytan, G.D., Assaraf, Y.G., 1996. Competition of hydrophobic peptides,cytotoxic drugs, and chemosensitizers on a common P-glycoprotein pharma-cophore as revealed by its ATPase activity. J. Biol. Chem. 271, 3163–3171.
ram, E.E., Ifergan, I., Grimberg, M., Lemke, K., Skladanowski, A., Assaraf, Y.G., 2007.C421 allele-specific ABCG2 gene amplification confers resistance to the antitu-mor triazoloacridone C-1305 in human lung cancer cells. Biochem. Pharmacol.74, 41–53.
ram, E.E., Stark, M., Raz, S., Assaraf, Y.G., 2009. Chemotherapeutic drug-inducedABCG2 promoter demethylation as a novel mechanism of acquired multidrugresistance. Neoplasia 11, 1359–1370.
reier, A., Gibalova, L., Seres, M., Barancik, M., Sulova, Z., 2013. New insight into
p-glycoprotein as a drug target. Anticancer Agents Med. Chem. 13, 159–170.rooks, B.R., Thisted, R.A., Appel, S.H., Bradley, W.G., Olney, R.K., Berg, J.E., Pope, L.E.,Smith, R.A., Group, A.-A.S., 2004. Treatment of pseudobulbar affect in ALS withdextromethorphan/quinidine: a randomized trial. Neurology 63, 1364–1370.
pdates 27 (2016) 14–29 25
Broxterman, H.J., Kuiper, C.M., Schuurhuis, G.J., Tsuruo, T., Pinedo, H.M., Lankelma,J., 1988. Increase of daunorubicin and vincristine accumulation in multidrugresistant human ovarian carcinoma cells by a monoclonal antibody reacting withP-glycoprotein. Biochem. Pharmacol. 37, 2389–2393.
Cai, C., Zhu, H., Chen, J., 2004. Overexpression of caveolin-1 increases plasma mem-brane fluidity and reduces P-glycoprotein function in Hs578 T/Dox. Biochem.Biophys. Res. Commun. 320, 868–874.
Callaghan, R., Ford, R.C., Kerr, I.D., 2006. The translocation mechanism of P-glycoprotein. FEBS Lett. 580, 1056–1063.
Cassinelli, G., Lanzi, C., Supino, R., Pratesi, G., Zuco, V., Laccabue, D., Cuccuru, G.,Bombardelli, E., Zunino, F., 2002. Cellular bases of the antitumor activity of thenovel taxane IDN 5109 (BAY59-8862) on hormone-refractory prostate cancer.Clin. Cancer Res. 8, 2647–2654.
Chai, S., To, K.K., Lin, G., 2010. Circumvention of multi-drug resistance of cancer cellsby Chinese herbal medicines. Chin. Med. 5, 26.
Chang, G., Roth, C.B., 2001. Structure of MsbA from E. coli: a homolog of the multidrugresistance ATP binding cassette (ABC) transporters. Science 293, 1793–1800.
Chang, X., Monitto, C.L., Demokan, S., Kim, M.S., Chang, S.S., Zhong, X., Califano, J.A.,Sidransky, D., 2010. Identification of hypermethylated genes associated withcisplatin resistance in human cancers. Cancer Res. 70, 2870–2879.
Chen, A.M., Zhang, M., Wei, D., Stueber, D., Taratula, O., Minko, T., He, H., 2009.Co-delivery of doxorubicin and Bcl-2 siRNA by mesoporous silica nanoparticlesenhances the efficacy of chemotherapy in multidrug-resistant cancer cells. Small5, 2673–2677.
Chhibber, A., Schroeder, B.G., 2008. Single-molecule polymerase chain reactionreduces bias: application to DNA methylation analysis by bisulfite sequencing.Anal. Biochem. 377, 46–54.
Cho, K., Wang, X., Nie, S., Chen, Z.G., Shin, D.M., 2008. Therapeutic nanoparticles fordrug delivery in cancer. Clin. Cancer Res. 14, 1310–1316.
Cho, S., Lu, M., He, X., Ee, P.L., Bhat, U., Schneider, E., Miele, L., Beck, W.T., 2011.Notch1 regulates the expression of the multidrug resistance gene ABCC1/MRP1in cultured cancer cells. Proc. Natl. Acad. Sci. U. S. A. 108, 20778–20783.
Chufan, E.E., Kapoor, K., Ambudkar, S.V., 2016. Drug-protein hydrogen bonds governthe inhibition of the ATP hydrolysis of the multidrug transporter P-glycoprotein.Biochem. Pharmacol. 101, 40–53.
Consoli, U., Priebe, W., Ling, Y.H., Mahadevia, R., Griffin, M., Zhao, S., Perez-Soler,R., Andreeff, M., 1996. The novel anthracycline annamycin is not affected byP-glycoprotein-related multidrug resistance: comparison with idarubicin anddoxorubicin in HL-60 leukemia cell lines. Blood 88, 633–644.
Cui, L., Aleksandrov, L., Chang, X.B., Hou, Y.X., He, L., Hegedus, T., Gentzsch, M., Alek-sandrov, A., Balch, W.E., Riordan, J.R., 2007. Domain interdependence in thebiosynthetic assembly of CFTR. J. Mol. Biol. 365, 981–994.
D’Angio, G.J., Evans, A., Breslow, N., Beckwith, B., Bishop, H., Farewell, V., Goodwin,W., Leape, L., Palmer, N., Sinks, L., Sutow, W., Tefft, M., Wolff, J., 1981. The treat-ment of Wilms’ tumor: results of the Second National Wilms’ Tumor Study.Cancer 47, 2302–2311.
Danhier, F., Lecouturier, N., Vroman, B., Jerome, C., Marchand-Brynaert, J., Feron, O.,Preat, V., 2009. Paclitaxel-loaded PEGylated PLGA-based nanoparticles: in vitroand in vivo evaluation. J. Control. Release 133, 11–17.
Davidson, A.L., Dassa, E., Orelle, C., Chen, J., 2008. Structure, function, and evolutionof bacterial ATP-binding cassette systems. Microbiol. Mol. Biol. Rev. 72, 317–364(table of contents).
Devulapally, R., Paulmurugan, R., 2014. Polymer nanoparticles for drug and smallsilencing RNA delivery to treat cancers of different phenotypes. Wiley Interdis-cip. Rev. Nanomed. Nanobiotechnol. 6, 40–60.
Dhar, S., Gu, F.X., Langer, R., Farokhzad, O.C., Lippard, S.J., 2008. Targeted deliv-ery of cisplatin to prostate cancer cells by aptamer functionalized Pt(IV)prodrug-PLGA-PEG nanoparticles. Proc. Natl. Acad. Sci. U. S. A. 105, 17356–17361.
Didziapetris, R., Japertas, P., Avdeef, A., Petrauskas, A., 2003. Classification analysisof P-glycoprotein substrate specificity. J. Drug Target. 11, 391–406.
Doerrler, W.T., Raetz, C.R., 2002. ATPase activity of the MsbA lipid flippase ofEscherichia coli. J. Biol. Chem. 277, 36697–36705.
Dong, M., Penin, F., Baggetto, L.G., 1996. Efficient purification and reconstitutionof P-glycoprotein for functional and structural studies. J. Biol. Chem. 271,28875–28883.
Dong, W.-H., Li, Q., Zhang, X.-Y., Guo, Q., Li, H., Wang, T.-Y., 2015. Deep sequencingidentifies deregulation of microRNAs involved with vincristine drug-resistanceof colon cancer cells. Int. J. Clin. Exp. Pathol. 8, 11524–11530.
Dorner, B., Kuntner, C., Bankstahl, J.P., Bankstahl, M., Stanek, J., Wanek, T., Stundner,G., Mairinger, S., Loscher, W., Muller, M., Langer, O., Erker, T., 2009. Synthesisand small-animal positron emission tomography evaluation of [11C]-elacridaras a radiotracer to assess the distribution of P-glycoprotein at the blood-brainbarrier. J. Med. Chem. 52, 6073–6082.
Edwards, J.R., Ruparel, H., Ju, J., 2005. Mass-spectrometry DNA sequencing. Mutat.Res. 573, 3–12.
Einhorn, L.H., 1990. Treatment of testicular cancer: a new and improved model. J.Clin. Oncol. 8, 1777–1781.
Elumalai, R., Patil, S., Maliyakkal, N., Rangarajan, A., Kondaiah, P., Raichur, A.M.,2015. Protamine-carboxymethyl cellulose magnetic nanocapsules for enhanceddelivery of anticancer drugs against drug resistant cancers. Nanomedicine 11,
969–981.Eytan, G.D., Borgnia, M.J., Regev, R., Assaraf, Y.G., 1994. Transport of polypeptideionophores into proteoliposomes reconstituted with rat liver P-glycoprotein. J.Biol. Chem. 269, 26058–26065.

2 nce U
F
F
F
F
F
F
F
F
G
G
G
G
G
G
G
G
G
G
G
G
G
G
H
H
H
H
H
H
H
H
H
6 W. Li et al. / Drug Resista
arokhzad, O.C., Cheng, J., Teply, B.A., Sherifi, I., Jon, S., Kantoff, P.W., Richie,J.P., Langer, R., 2006. Targeted nanoparticle–aptamer bioconjugates for cancerchemotherapy in vivo. Proc. Natl. Acad. Sci. U. S. A. 103, 6315–6320.
ederici, L., Woebking, B., Velamakanni, S., Shilling, R.A., Luisi, B., van Veen, H.W.,2007. New structure model for the ATP-binding cassette multidrug transporterLmrA. Biochem. Pharmacol. 74, 672–678.
edurco, M., Romieu, A., Williams, S., Lawrence, I., Turcatti, G., 2006. BTA, a novelreagent for DNA attachment on glass and efficient generation of solid-phaseamplified DNA colonies. Nucleic Acids Res. 34, e22.
erry, D.R., Russell, M.A., Cullen, M.H., 1992. P-glycoprotein possesses a 1,4-dihydropyridine-selective drug acceptor site which is alloserically coupled toa vinca-alkaloid-selective binding site. Biochem. Biophys. Res. Commun. 188,440–445.
isher, M., Abramov, M., Van Aerschot, A., Xu, D., Juliano, R.L., Herdewijn, P., 2007.Inhibition of MDR1 expression with altritol-modified siRNAs. Nucleic Acids Res.35, 1064–1074.
ox, E., Bates, S.E., 2007. Tariquidar (XR9576): a P-glycoprotein drug efflux pumpinhibitor. Expert Rev. Anticancer Ther. 7, 447–459.
relet, A., Klein, M., 2006. Insight in eukaryotic ABC transporter function by mutationanalysis. FEBS Lett. 580, 1064–1084.
riche, E., Jensen, P.B., Nissen, N.I., 1992. Comparison of cyclosporin A and SDZPSC833 as multidrug-resistance modulators in a daunorubicin-resistant Ehrlichascites tumor. Cancer Chemother. Pharmacol. 30, 235–237.
agliano, T., Gentilin, E., Benfini, K., Di Pasquale, C., Tassinari, M., Falletta, S., Feo,C., Tagliati, F., Uberti, E.D., Zatelli, M.C., 2014. Mitotane enhances doxorubicincytotoxic activity by inhibiting P-gp in human adrenocortical carcinoma cells.Endocrine 47, 943–951.
andhi, N.S., Tekade, R.K., Chougule, M.B., 2014. Nanocarrier mediated delivery ofsiRNA/miRNA in combination with chemotherapeutic agents for cancer therapy:current progress and advances. J. Control. Release 194, 238–256.
lasspool, R.M., Teodoridis, J.M., Brown, R., 2006. Epigenetics as a mechanism drivingpolygenic clinical drug resistance. Br. J. Cancer 94, 1087–1092.
obbi, P.G., Broglia, C., Merli, F., Dell’Olio, M., Stelitano, C., Iannitto, E., Federico, M.,Berte, R., Luisi, D., Molica, S., Cavalli, C., Dezza, L., Ascari, E., 2003. Vinblastine,bleomycin, and methotrexate chemotherapy plus irradiation for patients withearly-stage, favorable Hodgkin lymphoma: the experience of the Gruppo ItalianoStudio Linfomi. Cancer 98, 2393–2401.
oel, A., Lin, Y.-L., Liau, J.-Y., Yu, S.-C., Ou, D.-L., Lin, L.-I., Tseng, L.-H., Chang, Y.-L., Yeh,K.-H., Cheng, A.-L., 2012. KRAS mutation is a predictor of oxaliplatin sensitivityin colon cancer cells. PLoS ONE 7, e50701.
oler-Baron, V., Assaraf, Y.G., 2011. Structure and function of ABCG2-rich extracel-lular vesicles mediating multidrug resistance. PLoS ONE 6, e16007.
oler-Baron, V., Sladkevich, I., Assaraf, Y.G., 2012. Inhibition of the PI3K-Akt signalingpathway disrupts ABCG2-rich extracellular vesicles and overcomes multidrugresistance in breast cancer cells. Biochem. Pharmacol. 83, 1340–1348.
onen, N., Assaraf, Y.G., 2012. Antifolates in cancer therapy: structure, activity andmechanisms of drug resistance. Drug Resist. Updat. 15, 183–210.
oodsell, D.S., 1999. The molecular perspective: methotrexate. Oncologist 4,340–341.
ottesman, M.M., Ludwig, J., Xia, D., Szakacs, G., 2006. Defeating drug resistance incancer. Discov. Med. 6, 18–23.
ottesman, M.M., Pastan, I., 1993. Biochemistry of multidrug resistance mediatedby the multidrug transporter. Annu. Rev. Biochem. 62, 385–427.
radishar, W.J., 2006. Albumin-bound paclitaxel: a next-generation taxane. ExpertOpin. Pharmacother. 7, 1041–1053.
u, J., Fang, X., Hao, J., Sha, X., 2015a. Reversal of P-glycoprotein-mediated multidrugresistance by CD44 antibody-targeted nanocomplexes for short hairpin RNA-encoding plasmid DNA delivery. Biomaterials 45, 99–114.
u, R.-X., Corradi, V., Singh, G., Choudhury, H.G., Beis, K., Tieleman, D.P., 2015b.Conformational changes of the antibacterial peptide ATP binding cassette trans-porter McjD revealed by molecular dynamics simulations. Biochemistry 54,5989–5998.
amada, H., Tsuruo, T., 1986. Functional role for the 170- to 180-kDa glycoproteinspecific to drug-resistant tumor cells as revealed by monoclonal antibodies. Proc.Natl. Acad. Sci. U. S. A. 83, 7785–7789.
ande, K.R., 1998. Etoposide: four decades of development of a topoisomerase IIinhibitor. Eur. J. Cancer 34, 1514–1521.
arsha, S., 2012. Dual drug delivery system for targeting H. pylori in the stom-ach: preparation and in vitro characterization of amoxicillin-loaded Carbopol(R)nanospheres. Int. J. Nanomed. 7, 4787–4796.
enrique, R., Oliveira, A., Costa, V.L., Baptista, T., Martins, A., Morais, A., Oliveira,J., Jerónimo, C., 2013. Epigenetic regulation of MDR1 gene through post-translational histone modifications in prostate cancer. BMC Genomics 14, 898.
iggins, C.F., Gottesman, M.M., 1992. Is the multidrug transporter a flippase? TrendsBiochem. Sci. 17, 18–21.
ong, L., Han, Y., Zhang, H., Li, M., Gong, T., Sun, L., Wu, K., Zhao, Q., Fan, D., 2010. Theprognostic and chemotherapeutic value of miR-296 in esophageal squamouscell carcinoma. Ann. Surg. 251, 1056–1063.
orio, M., Gottesman, M.M., Pastan, I., 1988. ATP-dependent transport of vinblastinein vesicles from human multidrug-resistant cells. Proc. Natl. Acad. Sci. U. S. A.85, 3580–3584.
uang, R.S., Ratain, M.J., 2009. Pharmacogenetics and pharmacogenomics of anti-cancer agents. CA. Cancer J. Clin. 59, 42–55.
uang, W., Blinov, N., Wishart, D.S., Kovalenko, A., 2015. Role of water in ligand bind-ing to maltose-binding protein: insight from a new docking protocol based onthe 3D-RISM-KH molecular theory of solvation. J. Chem. Inf. Model. 55, 317–328.
pdates 27 (2016) 14–29
Huang, X.-C., Xiao, X., Zhang, Y.-K., Talele, T.T., Salim, A.A., Chen, Z.-S., Capon,R.J., 2014. Lamellarin O, a pyrrole alkaloid from an Australian Marine Sponge,Ianthella sp., reverses BCRP mediated drug resistance in cancer cells. Mar. Drugs12, 3818–3837.
Hung, L.W., Wang, I.X., Nikaido, K., Liu, P.Q., Ames, G.F., Kim, S.H., 1998. Crystal struc-ture of the ATP-binding subunit of an ABC transporter. Nature 396, 703–707.
Huttunen, K.M., Raunio, H., Rautio, J., 2011. Prodrugs – from serendipity to rationaldesign. Pharmacol. Rev. 63, 750–771.
Ifergan, I., Scheffer, G.L., Assaraf, Y.G., 2005. Novel extracellular vesicles mediate anABCG2-dependent anticancer drug sequestration and resistance. Cancer Res. 65,10952–10958.
Jain, K.K., 2002. Proteomics-based anticancer drug discovery and development.Technol. Cancer Res. Treat. 1, 231–236.
Jeon, J., Nim, S., Teyra, J., Datti, A., Wrana, J.L., Sidhu, S.S., Moffat, J., Kim, P.M., 2014. Asystematic approach to identify novel cancer drug targets using machine learn-ing, inhibitor design and high-throughput screening. Genome Med. 6, 57.
Jiang, B.-H., Liu, L.-Z., 2008. Role of mTOR in anticancer drug resistance: perspectivesfor improved drug treatment. Drug Resist. Updat. 11, 63–76.
Jin, X., Zhou, B., Xue, L., San, W., 2015. Soluplus® micelles as a potential drug deliverysystem for reversal of resistant tumor. Biomed. Pharmacother. 69, 388–395.
Jones, P.M., George, A.M., 1998. A new structural model for P-glycoprotein. J. Membr.Biol. 166, 133–147.
Jones, P.M., George, A.M., 2009. Opening of the ADP-bound active site in the ABCtransporter ATPase dimer: evidence for a constant contact, alternating sitesmodel for the catalytic cycle. Proteins 75, 387–396.
Jones, P.M., George, A.M., 2014. A reciprocating twin-channel model for ABC trans-porters. Q. Rev. Biophys. 47, 189–220.
Jordan, M.A., 2002. Mechanism of action of antitumor drugs that interact with micro-tubules and tubulin. Curr. Med. Chem. Anticancer Agents 2, 1–17.
Kajiyama, H., Shibata, K., Terauchi, M., Yamashita, M., Ino, K., Nawa, A., Kikkawa, F.,2007. Chemoresistance to paclitaxel induces epithelial-mesenchymal transitionand enhances metastatic potential for epithelial ovarian carcinoma cells. Int. J.Oncol. 31, 277–283.
Kakarla, P., Inupakutika, M., Devireddy, A.R., Gunda, S.K., Willmon, T.M., Ranjana,K.C., Shrestha, U., Ranaweera, I., Hernandez, A.J., Barr, S., Varela, M.F., 2016. 3D-QSAR and contour map analysis of tariquidar analogues as multidrug resistanceprotein-1 (MRP1) inhibitors. Int. J. Pharm. Sci. Res. 7, 554–572.
Kang, L., Gao, Z., Huang, W., Jin, M., Wang, Q., 2015. Nanocarrier-mediated co-delivery of chemotherapeutic drugs and gene agents for cancer treatment. ActaPharm. Sinica B 5, 169–175.
Kannan, P., Telu, S., Shukla, S., Ambudkar, S.V., Pike, V.W., Halldin, C., Gottesman,M.M., Innis, R.B., Hall, M.D., 2011. The “specific” P-glycoprotein inhibitor Tariq-uidar is also a substrate and an inhibitor for breast cancer resistance protein(BCRP/ABCG2). ACS Chem. Neurosci. 2, 82–89.
Kapse-Mistry, S., Govender, T., Srivastava, R., Yergeri, M., 2014. Nanodrug deliveryin reversing multidrug resistance in cancer cells. Front. Pharmacol. 5, 159.
Kast, C., Canfield, V., Levenson, R., Gros, P., 1995. Membrane topology of P-glycoprotein as determined by epitope insertion: transmembrane organizationof the N-terminal domain of MDR3. Biochemistry 34, 4402–4411.
Kataoka, K., Harada, A., Nagasaki, Y., 2001. Block copolymer micelles for drug deliv-ery: design, characterization and biological significance. Adv. Drug Deliv. Rev.47, 113–131.
Kathawala, R.J., Gupta, P., Ashby Jr., C.R., Chen, Z.-S., 2015. The modulation ofABC transporter-mediated multidrug resistance in cancer: a review of the pastdecade. Drug Resist. Updat. 18, 1–17.
Kaur, P., Rao, D.K., Gandlur, S.M., 2005. Biochemical characterization of domains inthe membrane subunit DrrB that interact with the ABC subunit DrrA: identifi-cation of a conserved motif. Biochemistry 44, 2661–2670.
Kawabata, S., Oka, M., Shiozawa, K., Tsukamoto, K., Nakatomi, K., Soda, H., Fukuda, M.,Ikegami, Y., Sugahara, K., Yamada, Y., Kamihira, S., Doyle, L.A., Ross, D.D., Kohno,S., 2001. Breast cancer resistance protein directly confers SN-38 resistance oflung cancer cells. Biochem. Biophys. Res. Commun. 280, 1216–1223.
Kelly, R.J., Draper, D., Chen, C.C., Robey, R.W., Figg, W.D., Piekarz, R.L., Chen, X., Gard-ner, E.R., Balis, F.M., Venkatesan, A.M., Steinberg, S.M., Fojo, T., Bates, S.E., 2011.A pharmacodynamic study of docetaxel in combination with the P-glycoproteinantagonist tariquidar (XR9576) in patients with lung, ovarian, and cervical can-cer. Clin. Cancer Res. 17, 569–580.
Kemper, E.M., Cleypool, C., Boogerd, W., Beijnen, J.H., van Tellingen, O., 2004.The influence of the P-glycoprotein inhibitor zosuquidar trihydrochloride(LY335979) on the brain penetration of paclitaxel in mice. Cancer Chemother.Pharmacol. 53, 173–178.
Kim, C.K., Ghosh, P., Pagliuca, C., Zhu, Z.J., Menichetti, S., Rotello, V.M., 2009. Entrap-ment of hydrophobic drugs in nanoparticle monolayers with efficient releaseinto cancer cells. J. Am. Chem. Soc. 131, 1360–1361.
Kobayashi, H., Sugimoto, H., Onishi, S., Nakano, K., 2015. Novel biomarker candidatesfor the diagnosis of ovarian clear cell carcinoma. Oncol. Lett. (Review).
Kobayashi, N., Nishino, K., Hirata, T., Yamaguchi, A., 2003. Membrane topology ofABC-type macrolide antibiotic exporter MacB in Escherichia coli. FEBS Lett. 546,241–246.
Koch, H., Busto, M.E.D.C., Kramer, K., Médard, G., Kuster, B., 2015. Chemical pro-teomics uncovers EPHA2 as a mechanism of acquired resistance to small
molecule EGFR kinase inhibition. J. Proteome Res. 14, 2617–2625.Koomen, J.M., Haura, E.B., Bepler, G., Sutphen, R., Remily-Wood, E.R., Benson, K.,Hussein, M., Hazlehurst, L.A., Yeatman, T.J., Hildreth, L.T., Sellers, T.A., Jacob-sen, P.B., Fenstermacher, D.A., Dalton, W.S., 2008. Proteomic contributions topersonalized cancer care. Mol. Cell. Proteomics 7, 1780–1794.

nce U
K
K
L
L
L
L
L
L
L
L
LL
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
L
W. Li et al. / Drug Resista
oski, A., Raki, M., Nokisalmi, P., Liikanen, I., Kangasniemi, L., Joensuu, T., Kanerva,A., Pesonen, S., Alemany, R., Hemminki, A., 2012. Verapamil results in increasedblood levels of oncolytic adenovirus in treatment of patients with advancedcancer. Mol. Ther. 20, 221–229.
ruhn, A., Wang, A., Fruehauf, J.H., Lage, H., 2009. Delivery of short hairpin RNAsby transkingdom RNA interference modulates the classical ABCB1-mediatedmultidrug-resistant phenotype of cancer cells. Cell Cycle 8, 3349–3354.
age, H., Dietel, M., 2000. Effect of the breast-cancer resistance protein on atypicalmultidrug resistance. Lancet Oncol. 1, 169–175.
age, H., Kruhn, A., 2010. Bacterial delivery of RNAi effectors: transkingdom RNAi. J.Vis. Exp.
ebedeva, I.V., Goler-Baron, V., Assaraf, Y.G., 2012. Overcoming multidrug resis-tance via photodestruction of ABCG2-rich extracellular vesicles sequesteringphotosensitive chemotherapeutics. PLoS ONE 7, e35487.
ee, S.M., Kim, H.J., Kim, S.Y., Kwon, M.K., Kim, S., Cho, A., Yun, M., Shin, J.S., Yoo, K.H.,2014. Drug-loaded gold plasmonic nanoparticles for treatment of multidrugresistance in cancer. Biomaterials 35, 2272–2282.
eonard, G.D., Fojo, T., Bates, S.E., 2003. The role of ABC transporters in clinicalpractice. Oncologist 8, 411–424.
i, H., Zhou, S., Li, T., Liu, Z., Wu, J., Zeng, G., Liu, C., Gong, J., 2012a. Suppres-sion of BCRP expression and restoration of sensitivity to chemotherapy inmultidrug-resistant HCC cell line HEPG2/ADM by RNA interference. Hepatogas-troenterology. 59, 2238–2242.
i, J., Jaimes, K.F., Aller, S.G., 2014a. Refined structures of mouse P-glycoprotein.Protein Sci. 23, 34–46.
i, W., 2014. Characterization of DrrAB Complex from Streptomyces peucetius as aMultidrug Transporter.
i, W., 2015. The Era of Nanotechnology and Omics Sciences, vol. 1., pp. 2.i, W., Rao, D.K., Kaur, P., 2013a. Dual role of the metalloprotease FtsH in biogenesis
of the DrrAB drug transporter. J. Biol. Chem. 288, 11854–11864.i, W., Sharma, M., Kaur, P., 2014b. The DrrAB efflux system of Streptomyces peucetius
is a multidrug transporter of broad substrate specificity. J. Biol. Chem. 289,12633–12646.
i, X., Gao, L., Cui, Q., Gary, B.D., Dyess, D.L., Taylor, W., Shevde-Samant, L.R., Samant,R.S., Dean-Colomb, W., Piazza, G.A., Xi, Y., 2012b. Sulindac inhibits tumor cellinvasion by suppressing NF-�B mediated transcription of microRNAs. Oncogene31, 4979–4986.
i, Y., Guessous, F., Zhang, Y., Dipierro, C., Kefas, B., Johnson, E., Marcinkiewicz, L.,Jiang, J., Yang, Y., Schmittgen, T.D., Lopes, B., Schiff, D., Purow, B., Abounader, R.,2009. MicroRNA-34a inhibits glioblastoma growth by targeting multiple onco-genes. Cancer Res. 69, 7569–7576.
i, Z., Liu, K., Sun, P., Mei, L., Hao, T., Tian, Y., Tang, Z., Li, L., Chen, D., 2013b. Poly(d,l-lactide-co-glycolide)/montmorillonite nanoparticles for improved oral deliveryof exemestane. J. Microencapsulation 30, 432–440.
i, Z., Qiu, L., Chen, Q., Hao, T., Qiao, M., Zhao, H., Zhang, J., Hu, H., Zhao, X., Chen,D., Mei, L., 2015. pH-sensitive nanoparticles of poly(l-histidine)-poly(lactide-co-glycolide)-tocopheryl polyethylene glycol succinate for anti-tumor drugdelivery. Acta Biomater. 11, 137–150.
inton, K.J., 2007. Structure and function of ABC transporters. Physiology (Bethesda)22, 122–130.
inton, K.J., Higgins, C.F., 2007. Structure and function of ABC transporters: the ATPswitch provides flexible control. Pflugers Arch. 453, 555–567.
iu, H., Huang, J., Peng, J., Wu, X., Zhang, Y., Zhu, W., Guo, L., 2015a. Upregulationof the inwardly rectifying potassium channel Kir2.1 (KCNJ2) modulates mul-tidrug resistance of small-cell lung cancer under the regulation of miR-7 andthe Ras/MAPK pathway. Mol. Cancer 14.
iu, R., Sharom, F.J., 1996. Site-directed fluorescence labeling of P-glycoproteinon cysteine residues in the nucleotide binding domains. Biochemistry 35,11865–11873.
iu, S., Wang, H., Zhang, L., Tang, C., Jones, L., Ye, H., Ban, L., Wang, A., Liu, Z., Lou,F., Zhang, D., Sun, H., Dong, H., Zhang, G., Dong, Z., Guo, B., Yan, H., Yan, C.,Wang, L., Su, Z., Li, Y., Huang, X.F., Chen, S.Y., Zhou, T., 2015b. Rapid detection ofgenetic mutations in individual breast cancer patients by next-generation DNAsequencing. Hum. Genomics 9, 2.
ivney, Y.D., Assaraf, Y.G., 2013. Rationally designed nanovehicles to overcome can-cer chemoresistance. Adv. Drug Deliv. Rev. 65, 1716–1730.
ocher, K.P., Borths, E., 2004. ABC transporter architecture and mechanism: impli-cations from the crystal structures of BtuCD and BtuF. FEBS Lett. 564, 264–268.
ocher, K.P., Lee, A.T., Rees, D.C., 2002. The E. coli BtuCD structure: a framework forABC transporter architecture and mechanism. Science 296, 1091–1098.
ongley, D.B., Allen, W.L., Johnston, P.G., 2006. Drug resistance, predictive mark-ers and pharmacogenomics in colorectal cancer. Biochim. Biophys. Acta 1766,184–196.
oo, T.W., Bartlett, M.C., Clarke, D.M., 2002. The “LSGGQ” motif in each nucleotide-binding domain of human P-glycoprotein is adjacent to the opposing walker Asequence. J. Biol. Chem. 277, 41303–41306.
oo, T.W., Bartlett, M.C., Clarke, D.M., 2003a. Simultaneous binding of two differentdrugs in the binding pocket of the human multidrug resistance P-glycoprotein.J. Biol. Chem. 278, 39706–39710.
oo, T.W., Bartlett, M.C., Clarke, D.M., 2003b. Substrate-induced conformationalchanges in the transmembrane segments of human P-glycoprotein. Direct evi-
dence for the substrate-induced fit mechanism for drug binding. J. Biol. Chem.278, 13603–13606.oo, T.W., Clarke, D.M., 2001. Defining the drug-binding site in the human multidrugresistance P-glycoprotein using a methanethiosulfonate analog of verapamil,MTS-verapamil. J. Biol. Chem. 276, 14972–14979.
pdates 27 (2016) 14–29 27
Loo, T.W., Clarke, D.M., 1999a. Identification of residues in the drug-binding domainof human P-glycoprotein. Analysis of transmembrane segment 11 by cysteine-scanning mutagenesis and inhibition by dibromobimane. J. Biol. Chem. 274,35388–35392.
Loo, T.W., Clarke, D.M., 1997. Identification of residues in the drug-binding siteof human P-glycoprotein using a thiol-reactive substrate. J. Biol. Chem. 272,31945–31948.
Loo, T.W., Clarke, D.M., 2000. Identification of residues within the drug-bindingdomain of the human multidrug resistance P-glycoprotein by cysteine-scanning mutagenesis and reaction with dibromobimane. J. Biol. Chem. 275,39272–39278.
Loo, T.W., Clarke, D.M., 2002. Location of the rhodamine-binding site in the humanmultidrug resistance P-glycoprotein. J. Biol. Chem. 277, 44332–44338.
Loo, T.W., Clarke, D.M., 1999b. Merck Frosst Award Lecture 1998. Molecular dissec-tion of the human multidrug resistance P-glycoprotein. Biochem. Cell Biol. 77,11–23.
Lopez, D., Martinez-Luis, S., 2014. Marine natural products with P-glycoproteininhibitor properties. Mar Drugs 12, 525–546.
Lubelski, J., van Merkerk, R., Konings, W.N., Driessen, A.J., 2006. Nucleotide-bindingsites of the heterodimeric LmrCD ABC-multidrug transporter of Lactococcus lactisare asymmetric. Biochemistry 45, 648–656.
Lugo, M.R., Sharom, F.J., 2005. Interaction of LDS-751 and rhodamine 123 with P-glycoprotein: evidence for simultaneous binding of both drugs. Biochemistry44, 14020–14029.
Luurtsema, G., Schuit, R.C., Klok, R.P., Verbeek, J., Leysen, J.E., Lammertsma,A.A., Windhorst, A.D., 2009. Evaluation of [11C]laniquidar as a tracer of P-glycoprotein: radiosynthesis and biodistribution in rats. Nucl. Med. Biol. 36,643–649.
Ma, L.-Y., Wang, B., Pang, L.-P., Zhang, M., Wang, S.-Q., Zheng, Y.-C., Shao,K.-P., Xue, D.-Q., Liu, H.-M., 2015. Design and synthesis of novel 1,2,3-triazole–pyrimidine–urea hybrids as potential anticancer agents. Bioorg. Med.Chem. Lett. 25, 1124–1128.
Margulies, M., Egholm, M., Altman, W.E., Attiya, S., Bader, J.S., Bemben, L.A., Berka, J.,Braverman, M.S., Chen, Y.J., Chen, Z., Dewell, S.B., Du, L., Fierro, J.M., Gomes, X.V.,Godwin, B.C., He, W., Helgesen, S., Ho, C.H., Irzyk, G.P., Jando, S.C., Alenquer, M.L.,Jarvie, T.P., Jirage, K.B., Kim, J.B., Knight, J.R., Lanza, J.R., Leamon, J.H., Lefkowitz,S.M., Lei, M., Li, J., Lohman, K.L., Lu, H., Makhijani, V.B., McDade, K.E., McKenna,M.P., Myers, E.W., Nickerson, E., Nobile, J.R., Plant, R., Puc, B.P., Ronan, M.T., Roth,G.T., Sarkis, G.J., Simons, J.F., Simpson, J.W., Srinivasan, M., Tartaro, K.R., Tomasz,A., Vogt, K.A., Volkmer, G.A., Wang, S.H., Wang, Y., Weiner, M.P., Yu, P., Begley,R.F., Rothberg, J.M., 2005. Genome sequencing in microfabricated high-densitypicolitre reactors. Nature 437, 376–380.
Martin, C., Berridge, G., Higgins, C.F., Mistry, P., Charlton, P., Callaghan, R., 2000.Communication between multiple drug binding sites on P-glycoprotein. Mol.Pharmacol. 58, 624–632.
Martin, S.K., Oduola, A.M., Milhous, W.K., 1987. Reversal of chloroquine resistancein Plasmodium falciparum by verapamil. Science 235, 899–901.
Mechetner, E.B., Roninson, I.B., 1992. Efficient inhibition of P-glycoprotein-mediatedmultidrug resistance with a monoclonal antibody. Proc. Natl. Acad. Sci. U. S. A.89, 5824–5828.
Michalak, K., Wesolowska, O., 2012. Polyphenols counteract tumor cell chemoresis-tance conferred by multidrug resistance proteins. Anticancer Agents Med. Chem.12, 880–890.
Minderman, H., O’Loughlin, K.L., Pendyala, L., Baer, M.R., 2004. VX-710 (birico-dar) increases drug retention and enhances chemosensitivity in resistant cellsoverexpressing P-glycoprotein, multidrug resistance protein, and breast cancerresistance protein. Clin. Cancer Res. 10, 1826–1834.
Mitra, P., Oskeritzian, C.A., Payne, S.G., Beaven, M.A., Milstien, S., Spiegel, S., 2006.Role of ABCC1 in export of sphingosine-1-phosphate from mast cells. Proc. Natl.Acad. Sci. U. S. A. 103, 16394–16399.
Mongre, R., Sodhi, S., Ghosh, M., Kim, J., Kim, N., Park, Y., Kim, S., Heo, Y., Sharma,N., Jeong, D., 2015. The novel inhibitor BRM270 downregulates tumorigenesisby suppression of NF-�B signaling cascade in MDR-induced stem like cancer-initiating cells. Int. J. Oncol.
Mornon, J.-P., Hoffmann, B., Jonic, S., Lehn, P., Callebaut, I., 2014. Full-open and closedCFTR channels, with lateral tunnels from the cytoplasm and an alternative posi-tion of the F508 region, as revealed by molecular dynamics. Cell. Mol. Life Sci.72, 1377–1403.
Morrison, C.D., Liu, P., Woloszynska-Read, A., Zhang, J., Luo, W., Qin, M., Bshara, W.,Conroy, J.M., Sabatini, L., Vedell, P., Xiong, D., Liu, S., Wang, J., Shen, H., Li, Y.,Omilian, A.R., Hill, A., Head, K., Guru, K., Kunnev, D., Leach, R., Eng, K.H., Darlak,C., Hoeflich, C., Veeranki, S., Glenn, S., You, M., Pruitt, S.C., Johnson, C.S., Trump,D.L., 2014. Whole-genome sequencing identifies genomic heterogeneity at anucleotide and chromosomal level in bladder cancer. Proc. Natl. Acad. Sci. U. S.A. 111, E672–E681.
Morschhauser, F., Zinzani, P.L., Burgess, M., Sloots, L., Bouafia, F., Dumontet, C., 2007.Phase I/II trial of a P-glycoprotein inhibitor, Zosuquidar.3HCl trihydrochloride(LY335979), given orally in combination with the CHOP regimen in patients withnon-Hodgkin’s lymphoma. Leuk. Lymphoma 48, 708–715.
Muller, M., de Vries, E.G., Jansen, P.L., 1996. Role of multidrug resistance protein(MRP) in glutathione S-conjugate transport in mammalian cells. J. Hepatol. 24
(Suppl 1), 100–108.Naito, M., Tsuge, H., Kuroko, C., Koyama, T., Tomida, A., Tatsuta, T., Heike, Y.,Tsuruo, T., 1993. Enhancement of cellular accumulation of cyclosporine by anti-P-glycoprotein monoclonal antibody MRK-16 and synergistic modulation ofmultidrug resistance. J. Natl. Cancer Inst. 85, 311–316.

2 nce U
N
O
P
P
P
P
P
P
P
P
P
P
C
Q
R
R
R
R
R
R
S
S
S
S
S
S
S
S
8 W. Li et al. / Drug Resista
iedner, H., Christen, R., Lin, X., Kondo, A., Howell, S.B., 2001. Identification of genesthat mediate sensitivity to cisplatin. Mol. Pharmacol. 60, 1153–1160.
gawara, K., Un, K., Tanaka, K., Higaki, K., Kimura, T., 2009. In vivo anti-tumor effectof PEG liposomal doxorubicin (DOX) in DOX-resistant tumor-bearing mice:involvement of cytotoxic effect on vascular endothelial cells. J. Control. Release133, 4–10.
ajeva, I.K., Wiese, M., 2009. Structure–activity relationships of tariquidar analogsas multidrug resistance modulators. AAPS J. 11, 435–444.
allis, M., Russell, N., 2000. P-glycoprotein plays a drug-efflux-independent rolein augmenting cell survival in acute myeloblastic leukemia and is associatedwith modulation of a sphingomyelin-ceramide apoptotic pathway. Blood 95,2897–2904.
an, L., Hu, H., Wang, X., Yu, L., Jiang, H., Chen, J., Lou, Y., Zeng, S., 2015. Inhibitoryeffects of neochamaejasmin B on P-glycoprotein in MDCK-hMDR1 cells andmolecular docking of NCB binding in P-glycoprotein. Molecules 20, 2931–2948.
ark, J.W., 2002. Liposome-based drug delivery in breast cancer treatment. BreastCancer Res. 4, 95–99.
earson, J.W., Fogler, W.E., Volker, K., Usui, N., Goldenberg, S.K., Gruys, E., Riggs, C.W.,Komschlies, K., Wiltrout, R.H., Tsuruo, T., et al., 1991. Reversal of drug resistancein a human colon cancer xenograft expressing MDR1 complementary DNA byin vivo administration of MRK-16 monoclonal antibody. J. Natl. Cancer Inst. 83,1386–1391.
ennock, G.D., Dalton, W.S., Roeske, W.R., Appleton, C.P., Mosley, K., Plezia, P., Miller,T.P., Salmon, S.E., 1991. Systemic toxic effects associated with high-dose vera-pamil infusion and chemotherapy administration. JNCI J. Natl. Cancer Inst. 83,105–110.
erez, J., Bardin, C., Rigal, C., Anthony, B., Rousseau, R., Dutour, A., 2011. Anti-MDR1 siRNA restores chemosensitivity in chemoresistant breast carcinoma andosteosarcoma cell lines. Anticancer Res. 31, 2813–2820.
iao, S., Xu, Y., Ha, N.C., 2008. Crystallization and preliminary X-ray crystallographicanalysis of MacA from Actinobacillus actinomycetemcomitans. Acta Crystallogr.Sect. F: Struct. Biol. Cryst. Commun. 64, 391–393.
orcelli, L., Giovannetti, E., Assaraf, Y.G., Jansen, G., Scheffer, G.L., Kathman, I.,Azzariti, A., Paradiso, A., Peters, G.J., 2014. The EGFR pathway regulates BCRPexpression in NSCLC cells: role of erlotinib. Curr. Drug Targets 15, 1322–1330.
radhan, P., Li, W., Kaur, P., 2009. Translational coupling controls expression andfunction of the DrrAB drug efflux pump. J. Mol. Biol. 385, 831–842.
.f.D.C.a. Prevention, 2015. Antibiotic Resistance Solutions Initiative $264M: A Com-prehensive Response. Center for Disease Control and Prevention.
u, Q., Sharom, F.J., 2001. FRET analysis indicates that the two ATPase active sites ofthe P-glycoprotein multidrug transporter are closely associated. Biochemistry40, 1413–1422.
az, S., Sheban, D., Gonen, N., Stark, M., Berman, B., Assaraf, Y.G., 2014. Severe hypoxiainduces complete antifolate resistance in carcinoma cells due to cell cycle arrest.Cell Death Dis. 5, e1067.
ees, D.C., Johnson, E., Lewinson, O., 2009. ABC transporters: the power to change.Nat. Rev. Mol. Cell Biol. 10, 218–227.
exer, B.N., Ham, A.J.L., Rinehart, C., Hill, S., de Matos Granja-Ingram, N., González-Angulo, A.M., Mills, G.B., Dave, B., Chang, J.C., Liebler, D.C., Arteaga, C.L., 2011.Phosphoproteomic mass spectrometry profiling links Src family kinases toescape from HER2 tyrosine kinase inhibition. Oncogene 30, 4163–4174.
osenberg, B., VanCamp, L., Trosko, J.E., Mansour, V.H., 1969. Platinum compounds:a new class of potent antitumour agents. Nature 222, 385–386.
osenberg, M.F., Callaghan, R., Ford, R.C., Higgins, C.F., 1997. Structure of the mul-tidrug resistance P-glycoprotein to 2.5 nm resolution determined by electronmicroscopy and image analysis. J. Biol. Chem. 272, 10685–10694.
ubin, E.H., de Alwis, D.P., Pouliquen, I., Green, L., Marder, P., Lin, Y., Musanti, R.,Grospe, S.L., Smith, S.L., Toppmeyer, D.L., Much, J., Kane, M., Chaudhary, A., Jor-dan, C., Burgess, M., Slapak, C.A., 2002. A phase I trial of a potent P-glycoproteininhibitor, Zosuquidar.3HCl trihydrochloride (LY335979), administered orally incombination with doxorubicin in patients with advanced malignancies. Clin.Cancer Res. 8, 3710–3717.
adanandam, A., Covell, D.G., 2015. Data mining approaches for genomic biomarkerdevelopment: applications using drug screening data from the cancer genomeproject and the cancer cell line encyclopedia. PLOS ONE 10, e0127433.
adeque, A.J., Wandel, C., He, H., Shah, S., Wood, A.J., 2000. Increased drug deliveryto the brain by P-glycoprotein inhibition. Clin. Pharmacol. Ther. 68, 231–237.
aville, M.W., Lietzau, J., Pluda, J.M., Feuerstein, I., Odom, J., Wilson, W.H., Humphrey,R.W., Feigal, E., Steinberg, S.M., Broder, S., et al., 1995. Treatment of HIV-associated Kaposi’s sarcoma with paclitaxel. Lancet 346, 26–28.
chmitt, L., 2002. The first view of an ABC transporter: the X-ray crystal structure ofMsbA from E. coli. Chembiochem 3, 161–165.
enior, A.E., al-Shawi, M.K., Urbatsch, I.L., 1995. ATP hydrolysis by multidrug-resistance protein from Chinese hamster ovary cells. J. Bioenerg. Biomembr. 27,31–36.
hafran, A., Ifergan, I., Bram, E., Jansen, G., Kathmann, I., Peters, G.J., Robey, R.W.,Bates, S.E., Assaraf, Y.G., 2005. ABCG2 harboring the Gly482 mutation con-fers high-level resistance to various hydrophilic antifolates. Cancer Res. 65,8414–8422.
hapira, A., Davidson, I., Avni, N., Assaraf, Y.G., Livney, Y.D., 2012. �-Caseinnanoparticle-based oral drug delivery system for potential treatment of gas-
tric carcinoma: stability, target-activated release and cytotoxicity. Eur. J. Pharm.Biopharm. 80, 298–305.hapiro, A.B., Fox, K., Lam, P., Ling, V., 1999. Stimulation of P-glycoprotein-mediateddrug transport by prazosin and progesterone. Evidence for a third drug-bindingsite. Eur. J. Biochem./FEBS 259, 841–850.
pdates 27 (2016) 14–29
Shapiro, A.B., Ling, V., 1998. The mechanism of ATP-dependent multidrug transportby P-glycoprotein. Acta Physiol. Scand. Suppl. 643, 227–234.
Shapiro, A.B., Ling, V., 1997. Positively cooperative sites for drug transport by P-glycoprotein with distinct drug specificities. Eur. J. Biochem./FEBS 250, 130–137.
Sharma, R.C., Inoue, S., Roitelman, J., Schimke, R.T., Simoni, R.D., 1992. Peptide trans-port by the multidrug resistance pump. J. Biol. Chem. 267, 5731–5734.
Sharom, F.J., 2011. The P-glycoprotein multidrug transporter. Essays Biochem. 50,161–178.
Shen, D.W., Pouliot, L.M., Hall, M.D., Gottesman, M.M., 2012. Cisplatin resistance: acellular self-defense mechanism resulting from multiple epigenetic and geneticchanges. Pharmacol. Rev. 64, 706–721.
Shen, H., Liu, T., Fu, L., Zhao, S., Fan, B., Cao, J., Li, X., 2013. Identification of microRNAsinvolved in dexamethasone-induced muscle atrophy. Mol. Cell. Biochem. 381,105–113.
Shendure, J., Porreca, G.J., Reppas, N.B., Lin, X., McCutcheon, J.P., Rosenbaum, A.M.,Wang, M.D., Zhang, K., Mitra, R.D., Church, G.M., 2005. Accurate multiplex polonysequencing of an evolved bacterial genome. Science 309, 1728–1732.
Sheng, J., Li, F., Wong, S., 2015. Optimal drug prediction from personal genomicsprofiles. IEEE J. Biomed. Health Inform.
Shi, Z., Jain, S., Kim, I.W., Peng, X.X., Abraham, I., Youssef, D.T., Fu, L.W., El Sayed, K.,Ambudkar, S.V., Chen, Z.S., 2007. Sipholenol A, a marine-derived sipholane triter-pene, potently reverses P-glycoprotein (ABCB1)-mediated multidrug resistancein cancer cells. Cancer Sci. 98, 1373–1380.
Shionoya, M., Jimbo, T., Kitagawa, M., Soga, T., Tohgo, A., 2003. DJ-927, a novel oraltaxane, overcomes P-glycoprotein-mediated multidrug resistance in vitro andin vivo. Cancer Sci. 94, 459–466.
Shukla, S., Chen, Z.-S., Ambudkar, S.V., 2012. Tyrosine kinase inhibitors as modulatorsof ABC transporter-mediated drug resistance. Drug Resist. Updat. 15, 70–80.
Sobell, H.M., 1985. Actinomycin and DNA transcription. Proc. Natl. Acad. Sci. U. S. A.82, 5328–5331.
Song, S., Honjo, S., Jin, J., Chang, S.S., Scott, A.W., Chen, Q., Kalhor, N., Correa, A.M.,Hofstetter, W.L., Albarracin, C.T., Wu, T.T., Johnson, R.L., Hung, M.C., Ajani, J.A.,2015. The Hippo coactivator YAP1 mediates EGFR overexpression and conferschemoresistance in esophageal cancer. Clin. Cancer Res. 21, 2580–2590.
Srinivasan, V., Sivaramakrishnan, H., Karthikeyan, B., 2011. Detection, isolation andcharacterization of principal synthetic route indicative impurities in verapamilhydrochloride. Sci. Pharm. 79, 555–568.
Srivastava, V., Kumar, A., Mishra, B.N., Siddiqi, M.I., 2008. CoMFA and CoMSIA 3D-QSAR analysis of DMDP derivatives as anti-cancer agents. Bioinformation 2,384–391.
Stebbing, J., Zhang, H., Xu, Y., Grothey, A., Ajuh, P., Angelopoulos, N., Giamas, G., 2015.Characterization of the tyrosine kinase-regulated proteome in breast cancer bycombined use of RNA interference (RNAi) and stable isotope labeling with aminoacids in cell culture (SILAC) quantitative proteomics. Mol. Cell. Proteomics 14,2479–2492.
Stephens, P.J., Tarpey, P.S., Davies, H., Van Loo, P., Greenman, C., Wedge, D.C., Nik-Zainal, S., Martin, S., Varela, I., Bignell, G.R., Yates, L.R., Papaemmanuil, E., Beare,D., Butler, A., Cheverton, A., Gamble, J., Hinton, J., Jia, M., Jayakumar, A., Jones, D.,Latimer, C., Lau, K.W., McLaren, S., McBride, D.J., Menzies, A., Mudie, L., Raine, K.,Rad, R., Chapman, M.S., Teague, J., Easton, D., Langerod, A., Oslo Breast Cancer, C.,Lee, M.T., Shen, C.Y., Tee, B.T., Huimin, B.W., Broeks, A., Vargas, A.C., Turashvili,G., Martens, J., Fatima, A., Miron, P., Chin, S.F., Thomas, G., Boyault, S., Mariani, O.,Lakhani, S.R., van de Vijver, M., van’t Veer, L., Foekens, J., Desmedt, C., Sotiriou,C., Tutt, A., Caldas, C., Reis-Filho, J.S., Aparicio, S.A., Salomon, A.V., Borresen-Dale, A.L., Richardson, A.L., Campbell, P.J., Futreal, P.A., Stratton, M.R., 2012. Thelandscape of cancer genes and mutational processes in breast cancer. Nature486, 400–404.
Stewart, J.J., White, J.T., Yan, X., Collins, S., Drescher, C.W., Urban, N.D., Hood, L.,Lin, B., 2006. Proteins associated with cisplatin resistance in ovarian cancercells identified by quantitative proteomic technology and integrated with mRNAexpression levels. Mol. Cell. Proteomics 5, 433–443.
Stoddart, D., Heron, A.J., Mikhailova, E., Maglia, G., Bayley, H., 2009. Single-nucleotidediscrimination in immobilized DNA oligonucleotides with a biological nanopore.Proc. Natl. Acad. Sci. U. S. A. 106, 7702–7707.
Su, Y., Cheng, X., Tan, Y., Hu, Y., Zhou, Y., Liu, J., Xu, Y., Xie, Y., Wang, C., Gao, Y.,Wang, J., Cheng, T., Yang, C., Xiong, D., Miao, H., 2012. Synthesis of a dual func-tional anti-MDR tumor agent PH II-7 with elucidations of anti-tumor effects andmechanisms. PLoS ONE 7, e32782.
Sun, T., Zhang, Y.S., Pang, B., Hyun, D.C., Yang, M., Xia, Y., 2014. Engineered nanopar-ticles for drug delivery in cancer therapy. Angew. Chem. Int. Ed. Engl. 53,12320–12364.
Sundar, S., Chen, Y., Tong, Y.W., 2014. Delivery of therapeutics and molecules usingself-assembled peptides. Curr. Med. Chem. 21, 2469–2479.
Tacar, O., Sriamornsak, P., Dass, C.R., 2013. Doxorubicin: an update on anticancermolecular action, toxicity and novel drug delivery systems. J. Pharm. Pharmacol.65, 157–170.
Tan, C., Tasaka, H., Yu, K.P., Murphy, M.L., Karnofsky, D.A., 1967. Daunomycin, anantitumor antibiotic, in the treatment of neoplastic disease. Clinical evaluationwith special reference to childhood leukemia. Cancer 20, 333–353.
Tang, R., Faussat, A.M., Perrot, J.Y., Marjanovic, Z., Cohen, S., Storme, T., Morjani,H., Legrand, O., Marie, J.P., 2008. Zosuquidar restores drug sensitivity in P-
glycoprotein expressing acute myeloid leukemia (AML). BMC Cancer 8, 51.Thomas, H., Coley, H.M., 2003. Overcoming multidrug resistance in cancer: an updateon the clinical strategy of inhibiting p-glycoprotein. Cancer Control 10, 159–165.
Tidefelt, U., Liliemark, J., Gruber, A., Liliemark, E., Sundman-Engberg, B., Julius-son, G., Stenke, L., Elmhorn-Rosenborg, A., Mollgard, L., Lehman, S., Xu, D.,

nce U
T
T
T
T
T
T
T
U
v
v
V
v
v
V
V
V
W
W
W
W
W
W
W. Li et al. / Drug Resista
Covelli, A., Gustavsson, B., Paul, C., 2000. P-glycoprotein inhibitor valspodar(PSC 833) increases the intracellular concentrations of daunorubicin in vivo inpatients with P-glycoprotein-positive acute myeloid leukemia. J. Clin. Oncol. 18,1837–1844.
insley, H.N., Gary, B.D., Thaiparambil, J., Li, N., Lu, W., Li, Y., Maxuitenko, Y.Y., Keeton,A.B., Piazza, G.A., 2010. Colon tumor cell growth inhibitory activity of sulindacsulfide and other NSAIDs is associated with PDE5 inhibition. Cancer Prev. Res.(Philadelphia, PA) 3, 1303–1313.
iram, G., Segal, E., Krivitsky, A., Shreberk-Hassidim, R., Ferber, S., Ofek, P., Uda-gawa, T., Edry, L., Shomron, N., Roniger, M., Kerem, B., Shaked, Y., Aviel-Ronen,S., Barshack, I., Calderón, M., Haag, R., Satchi-Fainaro, R., 2016. Identificationof dormancy-associated MicroRNAs for the design of osteosarcoma-targeteddendritic polyglycerol nanopolyplexes. ACS Nano 10, 2028–2045.
o, K.K., 2013. MicroRNA: a prognostic biomarker and a possible druggable targetfor circumventing multidrug resistance in cancer chemotherapy. J. Biomed. Sci.20, 99.
suruo, T., Hamada, H., Sato, S., Heike, Y., 1989. Inhibition of multidrug-resistanthuman tumor growth in athymic mice by anti-P-glycoprotein monoclonal anti-bodies. Jpn. J. Cancer Res. 80, 627–631.
suruo, T., Iida, H., Tsukagoshi, S., Sakurai, Y., 1981. Overcoming of vincristine resis-tance in P388 leukemia in vivo and in vitro through enhanced cytotoxicity ofvincristine and vinblastine by verapamil. Cancer Res. 41, 1967–1972.
urcatti, G., Romieu, A., Fedurco, M., Tairi, A.P., 2008. A new class of cleavable flu-orescent nucleotides: synthesis and optimization as reversible terminators forDNA sequencing by synthesis. Nucleic Acids Res. 36, e25.
usnady, G.E., Bakos, E., Varadi, A., Sarkadi, B., 1997. Membrane topology distin-guishes a subfamily of the ATP-binding cassette (ABC) transporters. FEBS Lett.402, 1–3.
rbatsch, I.L., Gimi, K., Wilke-Mounts, S., Lerner-Marmarosh, N., Rousseau, M.E.,Gros, P., Senior, A.E., 2001. Cysteines 431 and 1074 are responsible for inhibitorydisulfide cross-linking between the two nucleotide-binding sites in human P-glycoprotein. J. Biol. Chem. 276, 26980–26987.
an den Berg van Saparoea, H.B., Lubelski, J., van Merkerk, R., Mazurkiewicz, P.S.,Driessen, A.J., 2005. Proton motive force-dependent Hoechst 33342 transport bythe ABC transporter LmrA of Lactococcus lactis. Biochemistry 44, 16931–16938.
an Helvoort, A., Smith, A.J., Sprong, H., Fritzsche, I., Schinkel, A.H., Borst, P., vanMeer, G., 1996. MDR1 P-glycoprotein is a lipid translocase of broad specificity,while MDR3 P-glycoprotein specifically translocates phosphatidylcholine. Cell87, 507–517.
an Houdt, W.J., Emmink, B.L., Pham, T.V., Piersma, S.R., Verheem, A., Vries, R.G.,Fratantoni, S.A., Pronk, A., Clevers, H., Borel Rinkes, I.H., Jimenez, C.R., Kra-nenburg, O., 2011. Comparative proteomics of colon cancer stem cells anddifferentiated tumor cells identifies BIRC6 as a potential therapeutic target. Mol.Cell. Proteomics 10, M111.011353.
an Vlerken, L.E., Duan, Z., Seiden, M.V., Amiji, M.M., 2007. Modulation of intracellu-lar ceramide using polymeric nanoparticles to overcome multidrug resistancein cancer. Cancer Res. 67, 4843–4850.
an Zuylen, L., Sparreboom, A., van der Gaast, A., van der Burg, M.E., van Beur-den, V., Bol, C.J., Woestenborghs, R., Palmer, P.A., Verweij, J., 2000. The orallyadministered P-glycoprotein inhibitor R101933 does not alter the plasma phar-macokinetics of docetaxel. Clin. Cancer Res. 6, 1365–1371.
ideira, M., Reis, R.L., Brito, M.A., 2014. Deconstructing breast cancer cell biology andthe mechanisms of multidrug resistance. Biochim. Biophys. Acta 1846, 312–325.
o, T.-T., Lu, M., Nguyen, D., Lui, B., Maylla, S., Fruman, D., 2014. Abstract B24: inhi-bition of mTOR induces multidrug resistance in B-progenitor acute lymphocyticleukemia. Cancer Res. 74, B24.
on Hoff, D.D., Ervin, T., Arena, F.P., Chiorean, E.G., Infante, J., Moore, M., Seay, T., Tju-landin, S.A., Ma, W.W., Saleh, M.N., Harris, M., Reni, M., Dowden, S., Laheru, D.,Bahary, N., Ramanathan, R.K., Tabernero, J., Hidalgo, M., Goldstein, D., Van Cut-sem, E., Wei, X., Iglesias, J., Renschler, M.F., 2013. Increased survival in pancreaticcancer with nab-paclitaxel plus gemcitabine. N. Engl. J. Med. 369, 1691–1703.
ang, W., Zou, L., Zhou, D., Zhou, Z., Tang, F., Xu, Z., Liu, X., 2015. Overexpres-sion of ubiquitin carboxyl terminal hydrolase-L1 enhances multidrug resistanceand invasion/metastasis in breast cancer by activating the MAPK/Erk signalingpathway. Mol. Carcinog.
ang, Y.L., 2014. Tiny Mutation Triggers Drug Resistance for Patients With One Typeof Leukemia. The University of Chicago Medicine – Communications, http://www.uchospitals.edu/news/2014/20140528-leukemia.html.
ard, A., Reyes, C.L., Yu, J., Roth, C.B., Chang, G., 2007. Flexibility in the ABC trans-porter MsbA: alternating access with a twist. Proc. Natl. Acad. Sci. U. S. A. 104,19005–19010.
eisburg, J.H., Roepe, P.D., Dzekunov, S., Scheinberg, D.A., 1999. Intracellular pH andmultidrug resistance regulate complement-mediated cytotoxicity of nucleatedhuman cells. J. Biol. Chem. 274, 10877–10888.
en, Y., Kolonich, H.R., Kruszewski, K.M., Giannoukakis, N., Gawalt, E.S., Meng, W.S.,2013. Retaining antibodies in tumors with a self-assembling injectable system.Mol. Pharm. 10, 1035–1044.
en, Y., Liu, W., Bagia, C., Zhang, S., Bai, M., Janjic, J.M., Giannoukakis, N.,Gawalt, E.S., Meng, W.S., 2014a. Antibody-functionalized peptidic membranes
pdates 27 (2016) 14–29 29
for neutralization of allogeneic skin antigen-presenting cells. Acta Biomater. 10,4759–4767.
Wen, Y., Meng, W., 2014. Recent in vivo evidences of particle-based delivery of small-interfering RNA (siRNA) into solid tumors. J. Pharm. Innov. 9, 158–173.
Wen, Y., Roudebush, S.L., Buckholtz, G.A., Goehring, T.R., Giannoukakis, N., Gawalt,E.S., Meng, W.S., 2014b. Coassembly of amphiphilic peptide EAK16-II with his-tidinylated analogues and implications for functionalization of �-sheet fibrils invivo. Biomaterials 35, 5196–5205.
Weng, L., Zhang, L., Peng, Y., Huang, R.S., 2013. Pharmacogenetics and pharma-cogenomics: a bridge to individualized cancer therapy. Pharmacogenomics 14,315–324.
Wilkens, S., 2015. Structure and mechanism of ABC transporters. F1000Prime Rep.7, 14.
Wilson, W.H., Jamis-Dow, C., Bryant, G., Balis, F.M., Klecker, R.W., Bates, S.E., Chabner,B.A., Steinberg, S.M., Kohler, D.R., Wittes, R.E., 1995. Phase I and pharmacoki-netic study of the multidrug resistance modulator dexverapamil with EPOCHchemotherapy. J. Clin. Oncol. 13, 1985–1994.
Wu, H., Hait, W.N., Yang, J.-M., 2003. Small interfering RNA-induced suppression ofMDR1 (P-glycoprotein) restores sensitivity to multidrug-resistant cancer cells.Cancer Res. 63, 1515–1519.
Wu, X., He, X., Wang, K., Xie, C., Zhou, B., Qing, Z., 2010. Ultrasmall near-infrared goldnanoclusters for tumor fluorescence imaging in vivo. Nanoscale 2, 2244–2249.
Xiong, J., Feng, J., Yuan, D., Zhou, J., Miao, W., 2015. Tracing the structural evolutionof eukaryotic ATP binding cassette transporter superfamily. Sci. Rep. 5, 16724.
Xu, Z.P., Lu, G.Q., Yu, A.B., 2006. Inorganic nanoparticles as carriers for efficientcellular delivery. Chem. Eng. Sci. 61, 1027–1040.
Xu, W., Wang, S., Chen, Q., Zhang, Y., Ni, P., Wu, X., Zhang, J., Qiang, F., Li, A., Røe,O.D., Xu, S., Wang, M., Zhang, R., Zhou, J., 2014. TXNL1-XRCC1 pathway regu-lates cisplatin-induced cell death and contributes to resistance in human gastriccancer. Cell Death Dis. 5, e1055.
Yang, L., Li, W., Kirberger, M., Liao, W., Ren, J., 2016. Design of nanomaterial basedsystems for novel vaccine development. Biomater. Sci.
Yang, L., Zhang, X., Ye, M., Jiang, J., Yang, R., Fu, T., Chen, Y., Wang, K., Tan, W., 2011.Aptamer-conjugated nanomaterials and their applications. Adv. Drug Deliv. Rev.63, 1361–1370.
Yang, X., Iyer, A.K., Singh, A., Choy, E., Hornicek, F.J., Amiji, M.M., Duan, Z., 2015.MDR1 siRNA loaded hyaluronic acid-based CD44 targeted nanoparticle systemscircumvent paclitaxel resistance in ovarian cancer. Sci. Rep. 5, 8509.
Yokoyama, M., Miyauchi, M., Yamada, N., Okano, T., Sakurai, Y., Kataoka, K., Inoue, S.,1990. Characterization and anticancer activity of the micelle-forming polymericanticancer drug adriamycin-conjugated poly(ethylene glycol)-poly(asparticacid) block copolymer. Cancer Res. 50, 1693–1700.
Zhang, F., Zhang, H., Wang, Z., Yu, M., Tian, R., Ji, W., Yang, Y., Niu, R., 2014.P-glycoprotein associates with Anxa2 and promotes invasion in multidrug resis-tant breast cancer cells. Biochem. Pharmacol. 87, 292–302.
Zhang, G., Wang, Z., Qian, F., Zhao, C., Sun, C., 2015a. Silencing of the ABCC4 gene byRNA interference reverses multidrug resistance in human gastric cancer. Oncol.Rep. 33, 1147–1154.
Zhang, H., Rahman, S., Li, W., Fu, G., Kaur, P., 2015b. Characterization of a noveldomain ‘GATE’ in the ABC protein DrrA and its role in drug efflux by the DrrABcomplex. Biochem. Biophys. Res. Commun. 459, 148–153.
Zhang, M., Akbulut, M., 2011. Adsorption, desorption, and removal of polymericnanomedicine on and from cellulose surfaces: effect of size. Langmuir 27,12550–12559.
Zhang, T.-T., Li, W., Meng, G., Wang, P., Liao, W., 2016. Strategies for transportingnanoparticles across the blood–brain barrier. Biomater. Sci. 4, 219–229.
Zhang, X.-Q., Xu, X., Lam, R., Giljohann, D., Ho, D., Mirkin, C.A., 2011. Strategy forincreasing drug solubility and efficacy through covalent attachment to polyva-lent DNA–nanoparticle conjugates. ACS Nano 5, 6962–6970.
Zheng, Y., Wen, Y., George, A.M., Steinbach, A.M., Phillips, B.E., Giannoukakis, N.,Gawalt, E.S., Meng, W.S., 2011. A peptide-based material platform for displayingantibodies to engage T cells. Biomaterials 32, 249–257.
Zhitomirsky, B., Assaraf, Y.G., 2015. Lysosomal sequestration of hydrophobicweak base chemotherapeutics triggers lysosomal biogenesis and lysosome-dependent cancer multidrug resistance. Oncotarget 6, 1143–1156.
Zhitomirsky, B., Assaraf, Y.G., 2016. Lysosomes as mediators of drug resistance incancer. Drug Resist. Updat. 24, 23–33.
Zhou, X., Li, D., Wang, X., Zhang, B., Zhu, H., Zhao, J., 2015. Galectin-1 is overexpressedin CD133(+) human lung adenocarcinoma cells and promotes their growth andinvasiveness. Oncotarget 6, 3111–3122.
Zhu, H., Cai, C., Chen, J., 2004. Suppression of P-glycoprotein gene expres-sion in Hs578T/Dox by the overexpression of caveolin-1. FEBS Lett. 576,369–374.
Zhu, H., Wu, H., Liu, X., Evans, B.R., Medina, D.J., Liu, C.G., Yang, J.M., 2008. Role
of MicroRNA miR-27a and miR-451 in the regulation of MDR1/P-glycoproteinexpression in human cancer cells. Biochem. Pharmacol. 76, 582–588.Zinzi, L., Capparelli, E., Cantore, M., Contino, M., Leopoldo, M., Colabufo, N.A., 2014.Small and innovative molecules as new strategy to revert MDR. Front. Oncol. 4,2.