draft · draft chronic intermittent hypobaric hypoxia provides vascular protection in aorta of...
TRANSCRIPT

Draft
Chronic intermittent hypobaric hypoxia provides vascular
protection in aorta of two-kidney one clip rat model of
hypertension
Journal: Canadian Journal of Physiology and Pharmacology
Manuscript ID cjpp-2017-0356.R2
Manuscript Type: Article
Date Submitted by the Author: 30-Sep-2017
Complete List of Authors: Congrui, Fu; Hebei Medical University
Na, Li; Medical College, Hebei University, Department of Physiology Yujia, Yuan; Hebei Medical University, Department of Physiology Ri, Wang; Hebei Medical University, Department of Physiology Jinting, Chen; Hebei Medical University, Department of Physiology Jing, Yang; Hebei Medical University, Department of Physiology Zan, Guo; Hebei Medical University, Department of Physiology Sheng, Wang; Hebei Medical University, Department of Physiology; Hebei Collaborative Innovation Center for Cardio-cerebrovascular Disease Yi, Zhang; Hebei Medical University, Department of Physiology; Hebei Collaborative Innovation Center for Cardio-cerebrovascular Disease yixian, Liu; Hebei Medical University; Hebei Collaborative Innovation Center for Cardio-cerebrovascular Disease
Jinghui, Dong; Hebei Medical University, Department of Physiology
Keyword: 2K1C rats, chronic intermittent hypobaric hypoxia, endothelium-dependent relaxation, p38, BMP-4
Is the invited manuscript for consideration in a Special
Issue?: N/A
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Title page
Chronic intermittent hypobaric hypoxia provides
vascular protection in aorta of two-kidney one clip rat
model of hypertension
Congrui Fu1, Na Li
2, Yujia Yuan
1, Ri Wang
1, Jinting Chen
1, Jing Yang
1 ,
Zan Guo 1
, Sheng Wang
1, 3, Yi Zhang
1, 3, Yixian Liu
1, 3*, Jinghui Dong
1*
1Department of Physiology, Hebei Medical University, Shijiazhuang, Hebei, China,
2Department of Physiology, Medical College, Hebei University, Baoding, Hebei, Chi-
na and 3Hebei Collaborative Innovation Center for Cardio-Cerebrovascular Disease,
Shijiazhuang, Hebei, China.
*Corresponding author: Yixian Liu, Department of Physiology, Hebei Medical Uni-
versity, 361 East Zhongshan Road, Shijiazhuang, Hebei, 050017, China. Email: phy-
[email protected]. Tel: +86-0311-86265643; Fax: +86-0311-86261183
*Jinghui Dong, Department of Physiology, Hebei Medical University, 361 East
Zhongshan Road, Shijiazhuang, Hebei, 050017, China. Email: [email protected].
Tel: +86-0311-86265643; Fax: +86-0311-86261183
Page 1 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
1
1
Abstract
Many studies have demonstrated that chronic intermittent hypobaric
hypoxia (CIHH) can reduce blood pressure in spontaneously hypertensive
rats and renovascular hypertensive (RVH) rats in which endothelial
dysfunction is determined as a critical factor. However, whether CIHH
can regulate vasodilation of the aorta in RVH rats remains unknown. The
purpose of this study was to investigate the effect of CIHH on impaired
relaxation of the aorta in the two-kidney one-clip (2K1C) RVH rat model.
The results showed CIHH improved the impaired endothelium-dependent
relaxation in the 2K1C rat aorta. The endothelial dysfunction was
prevented by the p38 antagonist SB203580, but not by the ERK1/2
antagonist PD98059 or JNK antagonist SP600125. Furthermore, the
expression of p-eNOS, HIF-1ɑ and HIF-2ɑ increased while that of p-p38
and BMP-4 decreased in CIHH-treated aortas from 2K1C rats. Finally,
the p-eNOS expression was up-regulated and the p-p38 expression was
down-regulated by pre-incubation of SB203580 or the BMP-4 antagonist
Noggin with the aorta. In conclusion, CIHH ameliorated the impairment
of endothelium-dependent relaxation through up-regulating the
expression of p-eNOS which may be mediated by the inhibition of BMP-
4 / p-p38 MAPK and up-regulating the expression of HIFs in the 2K1C
rat aorta.
Page 2 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
2
2
Key words: 2K1C rats, chronic intermittent hypobaric hypoxia,
endothelium-dependent relaxation, p38, BMP-4
Page 3 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
3
3
Introduction
Hypertension is caused by pathological changes in renal and vascular
structures (Curb et al. 1996). Accumulated studies have shown that
endothelial dysfunction could contribute to elevated blood pressure in
hypertension (Carlisle et al. 2016; Statsenko et al. 2015).
CIHH is a programmed hypobaric hypoxia created by simulating
altitude in a hypobaric chamber, which yields numerous beneficial effects
on the body. Previous studies demonstrated that CIHH could protect the
cardiovascular system and brain against ischemia/reperfusion injury
(Zhang et al. 2016; Zhang et al. 2012). Importantly, CIHH was shown to
decrease the arterial blood pressure in patients with essential hypertension
and in rats with spontaneously hypertension (Aleshin et al. 1993;
Manukhina et al. 2000). Recent research suggested that CIHH treatment
improved the reduced vasorelaxation of mesenteric arteries through
activating BKCa channels in rats with renovascular hypertension (RVH)
(Guan et al. 2016). However, whether CIHH modulates vascular tension
in the aorta of RVH rats remains unclear.
Hypoxia inducible factors (HIFs) are oxygen-sensitive transcriptional
activators related to hypoxia. HIFs are comprised of an O2-regulated
HIF-1α or HIF-2α subunit and form a complex with HIF-1β (Prabhakar et
Page 4 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
4
4
al. 2012). Many of the above beneficial effects of CIHH are mediated by
HIFs. However, whether CIHH provides effective vascular protection in
hypertension through the HIF pathway is still unknown.
Conventional mitogen-activated protein kinases (MAPKs) include the
extracellular signal-regulated kinases (ERKs), c-Jun amino (N)-terminal
kinases (JNKs) and p38 isoforms (α, β, γ and δ) (Hadwiger et al. 2011).
Many studies have shown changes in the activity of MAPKs along with
hypoxia. Hypoxia-induced pulmonary arterial hypertension was found to
be improved by reducing the activity of the p38 MAPK signaling
pathway (Awad et al. 2016; Yan et al. 2016). The expression of ERK
increased after the rats were exposed to chronic intermittent hypoxia for 5
weeks (Micova et al. 2016). Chronic intermittent hypoxia-activated
MAPK family members, ERK, JNK and p38, rely on the O2
concentration in the rat obstructive sleep apnea model (Wang et al. 2016).
However, whether MAPKs participate in the effect of CIHH on vascular
tension in RVH rats remains elusive. In this study, antagonists of
ERK1/2, JNK and p38 were used to verify their roles in the protective
effect of CIHH.
Page 5 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
5
5
Bone morphogenic protein 4 (BMP4), belonging to the BMP family,
was originally discovered to participate in embryonic development and
bone and cartilage formation (Li et al. 2001; Massague 2000). Up-
regulation of BMP-4 has been linked to hypertension in the systemic
circulation, whereas the disruption of BMP-4 signaling is associated with
the development of pulmonary hypertension (Csiszar et al. 2008). BMP4
stimulation was shown to lead to endothelial dysfunction through reactive
oxygen species (ROS)-dependent p38 MAPK activation (Kamali et al.
2016).
Thus, the current study investigated the potential protective effects of
CIHH against endothelial dysfunction in the two-kidney one-clip (2K1C)
hypertensive rat model and discussed the mechanism involved.
Materials and methods
Animals and group
Fifty-two young male Sprague-Dawley rats (170–190 g, supplied by
the Experimental Animal Center of Hebei Medical University) were
randomly divided into eight groups: (1) Sham group: rats in this group
were treated with a sham operation; (2) CIHH group: rats in this group
were subjected to a sham operation and then treated with CIHH, which
was performed to simulate a 5000 m altitude (Pb = 404 mmHg) for 6 h
Page 6 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
6
6
daily over 28 days in a hypobaric chamber, while a normal oxygen
environment was provided the rest of the time; (3) 2K1C group: rats
received a 2K1C operation but without CIHH treatment. In the 2K1C
operation, the left renal artery was clipped with a small silver clip to
produce hypertension; (4) 2K1C + CIHH group: rats received a 2K1C
operation and CIHH treatment; (5) 2K1C + SB203580 group: aortas were
isolated from 2K1C rats and then incubated with SB203580 for 30 min
before use; (6) 2K1C + PD98059 group: the protocol was the same as that
in group 5, except that PD98059 was used for the incubation; (7) 2K1C +
SP600125 group: the protocol was the same as at for group 5, except that
SP600125 was used for the incubation; (8) 2K1C + Noggin group: the
protocol was the same as that for group 5, except that Noggin was used
for the incubation. The animals were housed at constant temperature (21
± 1°C) under a 12-h light/dark cycle and had free access to chow diet and
water. The animals were cared for in accordance with the Guide to the
Care and Use of Experimental Animals (Vol. 1, 2nd ed., 1993, and Vol. 2,
1984, available from the Canadian Council on Animal Care). All animal
procedures were approved by the Institutional Animal Care and Use
Committee of Hebei Medical University.
Chemicals and antibodies
Page 7 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
7
7
Protease inhibitor and phosphoric acid protease inhibitors were
purchased from Roche Mannheim (Mannheim, Germany). Mouse
polyclonal anti-HIF-1α and anti-HIF-2α primary antibodies, rabbit
polyclonal primary antibodies to p-eNOS, eNOS, p-p38, p38, p-ERK,
ERK, p-JNK, JNK and BMP-4 were from Abcam (Cambridge, UK). The
rabbit polyclonal primary antibody to β-actin primary antibody was from
Cell Signaling Technology (Danvers, MA, USA). The horseradish
peroxidase labeled anti-rabbit and anti-mouse secondary antibodies for
Western blot were from KPL Scaffold Inc (La Mirada, CA, USA) and
Earthox Life Sciences (Millbrae, California, USA), respectively. All
secondary antibodies for immunofluorescence staining were from
Jackson Laboratories Inc (West Grove, PA, USA). All general chemicals
were purchased from Sigma (St. Louis, MO, USA) unless stated
otherwise. Polyvinylidene difluoride (PVDF) membranes and the
enhanced chemiluminescence detecting reagents were from Millipore
Corporation (Billerica, MA, USA).
Preparation of renovascular hypertension rat model
Renovascular hypertension was induced by the Goldblatt 2K1C
method as described in a previous study (Guo et al. 2015). Briefly, the
rats were anesthetized with sodium pentobarbital (35 mg / kg, ip), and a
retroperitoneal flank incision was performed. The left renal artery was
Page 8 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
8
8
exposed and partially occluded by a U-shaped silver clip with an internal
diameter of 0.20 mm. After the operation, all animals received penicillin
(100000 units) and an analgesic ketorolac (0.6 mg / kg, ip) injection to
prevent infection and pain. The rats were maintained four weeks for full
recovery from the operation. Blood pressure was recorded by using the
tail-cuff electrosphygmomanometer system (AD Instruments, Sydney,
Australia) after the rats were stabilized and remained quiescent. An
average of three readings was taken as the systolic blood pressure for
each rat. Only 2K1C rats with a systolic arterial blood pressure > 150
mmHg were used in the experiment.
Thoracic aortic rings preparation and measurement of vascular
relaxation
The rats were killed by anesthetization with sodium pentobarbital (50
mg / kg, ip). The thoracic aorta was dissected, and surrounding
connective tissues were carefully removed. The arterial segment was cut
into 3–4 mm rings. The rings were then suspended between stainless steel
triangles in an organ bath filled with Krebs-Henseleit (K-H) solution of
the following composition (mmol / L): NaCl 119.0; KCl 4.7; CaCl2 2.5;
MgCl2 1.0; NaHCO3 25.0; KH2PO4 1.2; glucose 11.0; pH 7.4. The bath
solution was constantly gassed with a mixture of 95% O2 and 5% CO2
and maintained at 37°C. Before the experiment, the aortic rings were
Page 9 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
9
9
allowed to equilibrate for 60 min under a resting tension of 2.5 g, during
which the K-H solution was replaced every 15 min. The aortic ring was
connected to a force transducer, and the isometric force was recorded
using a biological signal recording system. After equilibration for 1 h,
phenylephrine (PE, 1 µmol / L) was given to induce contractions. When
the contraction was steady, acetylcholine (ACh, from 1 nmol / L to 10
µmol / L) or sodium nitroprusside (SNP, 1 nmol / L to 1 µmol / L) was
added in a cumulative manner to record the endothelial-dependent
relaxation or endothelial-independent relaxation response curve. The
rings with Ach (10 µmol / L)-induced relaxation of more than 80% was
used as the intact endothelium specimen. The extent of relaxation was
expressed as the percentage of relaxation from a submaximal
phenylephrine-induced constriction. Vasocontraction was determined in
relative values as the percentage of 60 mmol / L KCl contraction. Each
of the antagonists, when used, was applied for a 30-min incubation period
prior to the addition of Ach. The experiments were designed to determine
whether p38, ERK1/2, JNK and BMP-4 had effects on Ach-induced
vasorelaxation in the rat aorta by exposure to the p38 antagonist
SB203580 (10 µmol / L), ERK1/2 antagonist PD98059 (10 µmol / L),
JNK antagonist SP600125 (10 µmol / L) or BMP-4 antagonist Noggin
(100 ng / ml) before Ach treatment.
Page 10 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
10
10
Western blotting
The thoracic aortic rings were homogenized in ice-cold RIPA lysis
buffer containing protease inhibitor and phosphoric acid protease
inhibitors. The homogenates were incubated on ice for 20 min and then
centrifuged. The supernatant was collected, and the protein concentration
was determined using the bicinchoninic acid (BCA) method (Generay
biotechnology, Shanghai, China) according to the manufacturer's
instructions. Briefly, 20 µl of different concentrations of standard protein
solutions, samples to be measured or blank control were added into the
wells of 96-well plates, and then 200 µl of working fluid was added to
each well and mixed slightly. The reagents were incubated for 30 min at
37°C, cooled to room temperature and then read at 562 nm in a
spectrophotometer. A standard protein curve was drawn, and the
concentrations of the samples were calculated through the standard curve
regression formula. Equal amounts of protein samples (10 µg) were
electrophoresed through a 10% SDS-polyacrylamide gel and then
transferred onto a PVDF membrane. Non-specific binding sites were
blocked by 5% non-fat milk or 1% BSA in 0.05% Tween-20 phosphate-
buffered saline (PBST), and then incubated overnight at 4°C with primary
antibodies against p-eNOS and eNOS, HIF-1α and HIF-2α, p-p38, p38, p-
ERK1/2, ERK1/2, p-JNK, JNK, BMP-4 and β-actin. After being washed
Page 11 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
11
11
three times with Tris buffered saline with Tween 20 (TBST), the
membranes were incubated with appropriate horseradish peroxidase-
labeled secondary antibodies for 1 h at room temperature. After washing
again, the signals were developed with an enhanced chemiluminescence
system. Band images were quantified, and the ratio of the integrated
optical density (IOD) of the target band to that of β-actin or of the
phosphorylated protein to that of the total protein was determined.
Immunofluorescence staining
The thoracic aortic rings were dissected and stored in 4%
paraformaldehyde at 4°C for 48 h. The ring was embedded in paraffin
and sectioned (5 µm). After deparaffinization and hydration, the sections
were treated with 3% H2O2, heated in citrate buffer and permeabilized
with 0.25% Triton X-100 in turn. After incubation with normal goat
serum for 1 h at 37°C, the sections were incubated at 4°C overnight with
primary antibody against HIF-1α and HIF-2α. After rinsing, the sections
were incubated with matched secondary antibodies tagged with
fluorescence reporters for 1 h at 37°C. Finally sections were washed, air-
dried and mounted on slides with Vectashield Antifade Mounting
Medium. Phosphate-buffered saline was used as negative controls in
place of primary antibodies. Sections were imaged using a DM6000B
fluorescence microscope (Leica Microsystems , Germany).
Page 12 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
12
12
Statistical analysis
Results are presented as means ± SEM from different groups.
Concentration-response curves were analyzed by nonlinear regression
curve fitting using GraphPad Prism software. Statistical significance was
determined by one-way ANOVA or two-way ANOVA followed by the
Tukey post hoc test when two treatments were compared (GraphPad
Software, San Diego, CA, USA). P < 0.05 indicated a statistically
significant difference.
Results
Differences in body weight and blood pressure between different
groups
The body weight was significantly lower in the 2K1C group than in
the Sham, CIHH and 2K1C + CIHH groups from the fourth week after
the 2K1C operation (Table 1).
The systolic blood pressure was significantly higher in the 2K1C
group than in the Sham group from the first week after the 2K1C
operation. CIHH treatment for 4 weeks significantly decreased the
systolic blood pressure in the 2K1C + CIHH group from the fifth week
compared with the 2K1C group (Table 2).
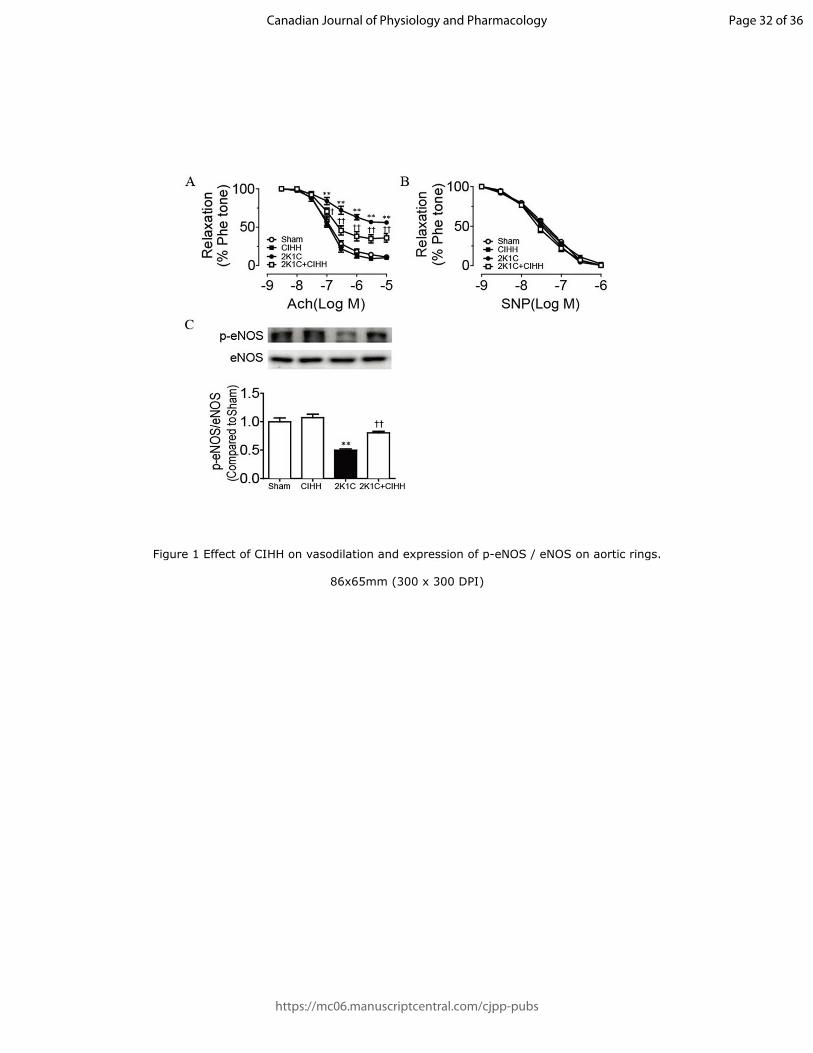
Relaxation effects of Ach and SNP on aortic rings
Page 13 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
13
13
Ach induced relaxation of aortic rings in a concentration-dependent
manner (1 nmol / L-10 µmol / L). The relaxation was obviously impaired
in 2K1C rats compared with Sham rats. CIHH treatment for 4 weeks had
no obvious effect on the relaxation of aortic rings compared with the
Sham group, but CIHH treatment significantly improved the relaxation in
the 2K1C+CIHH group compared with the 2K1C group (Figure 1A). In
addition, Western blot results demonstrated that the expression of p-
eNOS was decreased in the 2K1C group compared with the Sham group,
and strikingly increased in the 2K1C + CIHH group compared with the
2K1C group (Figure 1C). However, no significant change was observed
in the expression of eNOS in each group. On the other hand, the
relaxation induced by SNP showed no difference among these groups
(Figure 1B). These results demonstrated that CIHH improved the
impaired endothelium-dependent relaxation in aortas isolated from 2K1C
rats by promoting the expression of p-eNOS.
Comparison of HIF-1α and HIF-2α expression between groups
The expression of HIF-1α (green) and HIF-2α (red) was examined by
an immunofluorescence assay. The results showed that HIF-1α and HIF-
2α expressed in the layers of blood vessels, especially in endothelial cells
after CIHH treatment. Compared with the Sham group, the fluorescent
intensities of HIF-1α and HIF-2α in endothelial cells were significantly
Page 14 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
14
14
increased in the CIHH group. Similarly, they were also higher in the
2K1C + CIHH group than in the 2K1C group (Figure 2A). The
expression levels of these proteins were also studied by Western blotting.
After the CIHH treatment, the expression levels of both HIF-1α and HIF-
2α were increased clearly compared with the Sham group. No significant
change was observed in the expression of the two HIF subunits in the
2K1C group compared with the Sham group, while CIHH treatment up-
regulated their expression levels in the 2K1C + CIHH group compared
with the 2K1C group (Figure 2B). The results suggested CIHH could up-
regulate the expression of HIF-1α and HIF-2α in the aortic ring with or
without hypertension.
Effects of SB203580, PD98059 and SP600125 on the impaired
endothelium-dependent relaxation in 2K1C rat aorta
In order to examine whether MAPKs participated in the impaired
endothelium-dependent relaxation in the 2K1C rat aorta, antagonists of
the three MAPKs were used. The impaired endothelium-dependent
relaxation in the 2K1C aortic rings was rescued in the presence of p38
antagonist SB203580, but not by ERK1/2 antagonist PD98059 or JNK
antagonist SP600125 (Figure 3A). The results indicated that the impaired
endothelium-dependent relaxation in the 2K1C rat aorta was mediated by
p38, but not by ERK1/2 or JNK.
Page 15 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
15
15
Comparisons of expression of p-p38 / p38, p-ERK1/2 / ERK1/2 and
p-JNK / JNK between groups
Whether CIHH treatment could affect the expression of various
MAPK members was investigated by observing their protein levels. As
shown in Figure 3B and 3C, minimal changes were seen in the expression
of p-ERK1/2/ ERK1/2 and p-JNK/JNK among the different groups.
Similarly, no obvious change of the p38 expression was found among
these groups. CIHH treatment itself had no obvious effect on the
expression of p-p38 compared with the Sham group. However, the
expression of the p-p38 protein was strikingly augmented in the 2K1C
group compared with the Sham group. Furthermore, CIHH treatment
reduced the augmented expression of p-p38 in the 2K1C + CIHH group
compared with the 2K1C group. The results suggested that CIHH
treatment might rescue the impaired endothelium-dependent relaxation in
aortic rings of 2K1C rats by inhibiting the augmented expression of p-p38
(Figure 3A).
Effect of Noggin on impaired endothelium-dependent relaxation of
aortic rings isolated from 2K1C group
We observed the effect of Noggin, an antagonist of BMP-4, on the
impaired endothelium-dependent relaxation in the 2K1C rat aorta. The
Page 16 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
16
16
results showed Noggin ameliorated the impaired endothelium-dependent
relaxation in 2K1C aortic rings (Figure 4A), which suggested that BMP-4
mediated the endothelium dysfunction in renovascular hypertension.
Effect of CIHH on expression of BMP-4
In order to examine whether CIHH treatment could regulate the
expression of BMP-4, we observed changes in the level of this protein in
different groups. As with the expression of p-p38, the expression of
BMP-4 in the 2K1C group was higher than that in the Sham group, and it
was obviously declined in 2K1C + CIHH rats compared with 2K1C rats
(Figure 4B, C). The changes of these proteins, together with the rescuing
effects of Noggin on endothelium-dependent relaxation of aortic rings of
2K1C rats indicated a relationship of endothelial dysfunction with
enhanced BMP-4.
Effects of SB203580 and Noggin on expression of p-eNOS / eNOS and
p-p38 / p38
The expression of p-eNOS in the aorta of 2K1C rats was less than that
in the Sham group. After preincubation of the p38 antagonist SB203580
or BMP-4 antagonist Noggin, the expression of p-eNOS was remarkably
increased in the 2K1C + SB203580 or 2K1C + Noggin group compared
Page 17 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
17
17
with the 2K1C group (Figure 5A, B). No significant change was
observed in the expression of eNOS in each group.
By contrast, the expression of p-p38 in 2K1C rats was much greater
than that in the Sham group. After application of the p38 antagonist
SB203580 or BMP-4 antagonist Noggin, the amount of p-p38 was
reduced compared with the 2K1C group. No obvious change was found
in the expression of p38 in each group (Figure 5A, B). The results
illustrated that the decreased expression of p-eNOS was concomitant with
the increased expression of p-p38 in aortas isolated from 2K1C rats.
Furthermore, the expression of p-p38 could be inhibited by giving the
BMP-4 antagonist Noggin, suggesting that BMP-4 is part of the upstream
cascade of p-p38.
Discussion
The present study tested the effects of CIHH on endothelial
dysfunction in the 2K1C rat aorta and explored the underlying
mechanisms. The findings showed that CIHH could improve the impaired
endothelium-dependent relaxation in the 2K1C rat aorta and increase the
expression of p-eNOS, HIF-1ɑ and HIF-2ɑ. The blockage of p-p38/p38
and BMP-4 rescued the impaired endothelium-dependent relaxation in the
2K1C rat aorta. Furthermore, CIHH reduced the increased expression of
Page 18 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
18
18
p-p38/p38 and BMP-4 in the 2K1C + CIHH group compared with the
2K1C group. Finally, we showed that the blockage of p38 or BMP-4 was
concomitant with the increased expression of p-eNOS. Thus, we can
conclude that CIHH improved the impaired endothelium-dependent
relaxation in the 2K1C rat aorta through inhibiting BMP4 and p-p38 and
up-regulating the expression of HIFs, consequently resulting in higher
eNOS activity.
Different protocols for inducing intermittent hypoxia produce diverse
responses. For example, a long-cycle intermittent hypobaric hypoxia (6 h
a day for 28–42 days) showed a cardiac protective effect (Peng et al. 2012)
and also an anti-hypertensive effect (Li et al. 2016). However,
normobaric hypoxia (20–50 s hypoxia, 20–50 s normoxia alternately,
10–12 h a day) was shown to cause an increase in arterial blood pressure
and deleterious myocardial infarction induced by cardiac
ischemia/reperfusion (Ramond et al. 2007). The short-cycle intermittent
normobaric hypoxia in the obstructive sleep apnea patients may cause
hypertension (Lesske et al. 1997). CIHH as used in our study is a
programmed intermittent hypoxia with a long cycle which has many
beneficial effects on the body. This work represents our first efforts
focusing on the protective effect of CIHH on aortic endothelial
dysfunction in renovascular hypertension. Results of this study confirmed
Page 19 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
19
19
our hypothesis that CIHH could rescue the dysfunction of the
endothelium in 2K1C rats.
Under hypoxia, HIF-α subunits are stabilized and translocated to the
nucleus where they heterodimerize with the arylhydrocarbon receptor
nuclear translocator and bind to hypoxia response elements (HREs)
located within regulatory elements of HIF target genes. There are three
types of HIF-α subunits, HIF-1α, HIF-2α, and HIF-3α. At present, HIF-
1α and HIF-2α have gained increased attention (Liu et al. 2016).
Interestingly, these two subunits show different patterns of tissue
distributions. HIF-1α is ubiquitously expressed in the body, but HIF-2α
expression is restricted to specific tissues, such as blood vessels (Ema et
al. 1997). Many studies have identified NOS as one of the target proteins
of HIFs with HREs in its regulatory elements (Coulet et al. 2003; Yuan et
al. 2016). In the present study, when the rats were exposed to CIHH, the
expression levels of HIF-1α and HIF-2α in endothelial cells were both
increased, so do the protein expressions. Correspondingly, the expression
of p-eNOS was also increased. The results suggested that CIHH treatment
increased the expression of HIF-1α and HIF-2α in normal and
hypertensive rats, which counterbalanced the decrease in eNOS activity
in hypertensive rats.
Page 20 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
20
20
MAPKs are relevant to the expression of eNOS. For example,
advanced glycation end-products is known to cause endothelial
dysfunction by a mechanism associated with decreased eNOS expression
in human coronary artery endothelial cells through activation of p38 and
ERK1/2 (Ren et al. 2017). Adenosine was shown to be required for
ERK1/2 activation by statins, which resulted in Akt and
eNOS phosphorylation (Merla et al. 2007). Thus, we examined whether
CIHH could increase eNOS by modulating the MAPK pathway. In this
study, the impaired endothelium-dependent relaxation in 2K1C aortic
rings was restored by the p38 antagonist SB203580, but it was not altered
by the ERK1/2 antagonist PD98059 or JNK antagonist SP600125. The
results suggested that p38, but not ERK1/2 or JNK mediated the
endothelial dysfunction in the aorta in hypertension. Accordingly, the
expression of p-p38 was also remarkably increased in the 2K1C rat aorta.
CIHH treatment reversed the abnormal expression of p-p38 MAPK in the
2K1C rat aorta, while the expression of p-ERK and p-JNK basically
remained in a stable state in each group. Thus, we can draw a conclusion
that CIHH improved the relaxation of aortas in 2K1C rats partly through
inhibiting the activity of p38.
Previous studies demonstrated that BMP4 increased phosphorylation
of p38 MAPK to promote chondrogenesis and exert proinflammatory
Page 21 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
21
21
effects on endothelial dysfunction (Kamali et al. 2016; Kim et al. 2016).
Therefore, we investigated whether CIHH could inhibit the activity of p-
p38 via down-regulating the expression of BMP-4 to exert a protective
effect. In our study, expression levels of p-p38 and BMP-4 both
decreased by CIHH treatment in the 2K1C + CIHH group compared with
the 2K1C group. In addition, BMP-4 was determined to be in the
upstream cascade of p-p38, as the expression of p-p38 was inhibited by
giving the BMP-4 antagonist Noggin. Furthermore, the expression of p-
eNOS was strikingly augmented and p-p38 remarkably reduced by the
incubation of SB203580 or Noggin with 2K1C rat aortas. These results
indicated that CIHH treatment restored p-eNOS expression in aortas
isolated from 2K1C rats through inhibiting BMP-4 and in turn inhibiting
the p38 MAPK activity.
In this study, the observed vascular protection was explained by
“short-term” mechanisms such as phosphorylation of eNOS and p38
proteins. Longer-term mechanisms for the anti-hypertensive effect of
CIHH have also been described, such as the potentiating effect on
baroreflex (Gao et al. 2012) or enhancement of the relaxation of
mesenteric arteries (Guan et al. 2016). Furthermore, CIHH has been
demonstrated to induce a cardiovascular protective effect for at least two
weeks and a neuroprotective effect for at least one week (Wang et al.
Page 22 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
22
22
2017; Zhang et al. 2000). Determining the length of time that the
antihypertensive effect can be maintained will require further observation.
Together, results of our study demonstrated that CIHH could improve
the impaired endothelium-dependent relaxation in 2K1C hypertensive rat
aortic rings. The mechanism may be that CIHH inhibits the expression of
BMP-4 and the activity of p38, thus increasing the expression of p-eNOS.
Furthermore, CIHH may up-regulate the expression of HIF-1α and HIF-
2α in endothelial cells, and then increase the expression of the product of
the target genes, p-eNOS. Thus, CIHH may potentially be developed as a
therapeutic to ameliorate endothelial dysfunction in hypertension.
Acknowledgements
This work was supported by the University Science and Technology Re-
search Project of Hebei Province, China (ZD2017052).
Disclosure: The authors declare no conflict of interest in this study.
References
Aleshin IA, Kots Ia I, Tverdokhlib VP, Galiautdinov GS, Vdovenko LG, Zabirov MR, et al. 1993. The nondrug treatment of hypertension patients by their adaptation to periodic hypoxia in a barochamber. Ter Arkh. 65(8): 23-29 Awad KS, West JD, de Jesus Perez V, MacLean M. 2016. Novel signaling pathways in pulmonary arterial hypertension (2015 Grover Conference Series). Pulm Circ. 6(3): 285-294. doi:10.1086/688034.
Page 23 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
23
23
Carlisle RE, Werner KE, Yum V, Lu C, Tat V, Memon M, et al. 2016. Endoplasmic reticulum stress inhibition reduces hypertension through the preservation of
resistance blood vessel structure and function. J Hypertens. 34(8): 1556-1569. doi:10.1097/HJH.0000000000000943.
Coulet F, Nadaud S, Agrapart M, Soubrier F. 2003. Identification of hypoxia-response element in the human endothelial nitric-oxide synthase gene promoter. J Biol Chem. 278(47): 46230-46240. doi:10.1074/jbc.M305420200.
Csiszar A, Labinskyy N, Jo H, Ballabh P, Ungvari Z. 2008. Differential proinflammatory and prooxidant effects of bone morphogenetic protein-4 in coronary and pulmonary arterial endothelial cells. Am J Physiol Heart Circ Physiol. 295(2): H569-577. doi:10.1152/ajpheart.00180.2008. Curb JD, Pressel SL, Cutler JA, Savage PJ, Applegate WB, Black H, et al. 1996. Effect of diuretic-based antihypertensive treatment on cardiovascular disease risk in older
diabetic patients with isolated systolic hypertension. Systolic Hypertension in the Elderly Program Cooperative Research Group. JAMA. 276(23): 1886-1892 Ema M, Taya S, Yokotani N, Sogawa K, Matsuda Y, Fujii-Kuriyama Y. 1997. A novel bHLH-PAS factor with close sequence similarity to hypoxia-inducible factor 1alpha regulates the VEGF expression and is potentially involved in lung and vascular
development. Proc Natl Acad Sci U S A. 94(9): 4273-4278 Gao L, Guan Y, Cui F, Liu YX, Zhou ZN, Zhang Y. 2012. Facilitation of chronic intermittent hypobaric hypoxia on carotid sinus baroreflex in anesthetized rats. Chin J
Physiol. 55(1): 62-70. doi:10.4077/CJP.2012.AMM076. Guan Y, Li N, Tian YM, Zhang L, Ma HJ, Maslov LN, et al. 2016. Chronic intermittent hypobaric hypoxia antagonizes renal vascular hypertension by enhancement of
vasorelaxation via activating BKCa. Life Sci. 157: 74-81. doi:10.1016/j.lfs.2016.05.028.
Guo Z, Liu YX, Yuan F, Ma HJ, Maslov L, Zhang Y. 2015. Enhanced vasorelaxation effect of endogenous anandamide on thoracic aorta in renal vascular hypertension rats. Clin Exp Pharmacol Physiol. doi:10.1111/1440-1681.12450.
Hadwiger JA, Nguyen HN. 2011. MAPKs in development: insights from Dictyostelium signaling pathways. Biomol Concepts. 2(1-2): 39-46. doi:10.1515/BMC.2011.004. Kamali M, Khodadoost M, Tavakoli H, Kamalinejad M, Gachkar L, Adibi P, et al. 2016.
The Role of Syndrome Differentiation in the Clinical Efficacy of Punica Granatum on Patients with Ulcerative Colitis. Iran J Med Sci. 41(3 Suppl): S15 Kim H, Sonn JK. 2016. Rac1 promotes chondrogenesis by regulating STAT3 signaling
pathway. Cell Biol Int. 40(9): 976-983. doi:10.1002/cbin.10635. Lesske J, Fletcher EC, Bao G, Unger T. 1997. Hypertension caused by chronic intermittent hypoxia--influence of chemoreceptors and sympathetic nervous system. J Hypertens. 15(12 Pt 2): 1593-1603 Li N, Guan Y, Zhang L, Tian Y, Zhang Y, Wang S. 2016. Depressive Effects of Chronic
Intermittent Hypobaric Hypoxia on Renal Vascular Hypertension through Enhancing Baroreflex. Chin J Physiol. 59(4): 210-217. doi:10.4077/CJP.2016.BAF444. Li RH, Wozney JM. 2001. Delivering on the promise of bone morphogenetic proteins.
Trends Biotechnol. 19(7): 255-265 Liu YM, Ying SP, Huang YR, Pan Y, Chen WJ, Ni LQ, et al. 2016. Expression of HIF-1alpha and HIF-2alpha correlates to biological and clinical significance in papillary
thyroid carcinoma. World J Surg Oncol. 14(1): 30. doi:10.1186/s12957-016-0785-9. Manukhina EB, Mashina S, Smirin BV, Lyamina NP, Senchikhin VN, Vanin AF, et al. 2000. Role of nitric oxide in adaptation to hypoxia and adaptive defense. Physiol Res. 49(1): 89-97
Page 24 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
24
24
Massague J. 2000. How cells read TGF-beta signals. Nat Rev Mol Cell Biol. 1(3): 169-178. doi:10.1038/35043051.
Merla R, Ye Y, Lin Y, Manickavasagam S, Huang MH, Perez-Polo RJ, et al. 2007. The central role of adenosine in statin-induced ERK1/2, Akt, and eNOS phosphorylation.
Am J Physiol Heart Circ Physiol. 293(3): H1918-1928. doi:10.1152/ajpheart.00416.2007. Micova P, Hahnova K, Hlavackova M, Elsnicova B, Chytilova A, Holzerova K, et al.
2016. Chronic intermittent hypoxia affects the cytosolic phospholipase A2alpha/cyclooxygenase 2 pathway via beta2-adrenoceptor-mediated ERK/p38 stimulation. Mol Cell Biochem. 423(1-2): 151-163. doi:10.1007/s11010-016-2833-8. Peng YJ, Nanduri J, Zhang X, Wang N, Raghuraman G, Seagard J, et al. 2012. Endothelin-1 mediates attenuated carotid baroreceptor activity by intermittent hypoxia. J Appl Physiol (1985). 112(1): 187-196.
doi:10.1152/japplphysiol.00529.2011. Prabhakar NR, Semenza GL. 2012. Adaptive and maladaptive cardiorespiratory
responses to continuous and intermittent hypoxia mediated by hypoxia-inducible factors 1 and 2. Physiol Rev. 92(3): 967-1003. doi:10.1152/physrev.00030.2011. Ramond A, Ribuot C, Levy P, Joyeux-Faure M. 2007. Deleterious myocardial
consequences induced by intermittent hypoxia are reversed by erythropoietin. Respir Physiol Neurobiol. 156(3): 362-369. doi:10.1016/j.resp.2006.10.011. Ren X, Ren L, Wei Q, Shao H, Chen L, Liu N. 2017. Advanced glycation end-products
decreases expression of endothelial nitric oxide synthase through oxidative stress in human coronary artery endothelial cells. Cardiovasc Diabetol. 16(1): 52. doi:10.1186/s12933-017-0531-9.
Statsenko ME, Derevianchenko MV. 2015. Possibilities of Correction of Endothelial Dysfunction at the Background of Combined Antihypertensive Therapy in Patients
With Arterial Hypertension and Type 2 Diabetes. Kardiologiia. 55(3): 17-20 Wang J, Zhang S, Ma H, Yang S, Liu Z, Wu X, et al. 2017. Chronic Intermittent Hypobaric Hypoxia Pretreatment Ameliorates Ischemia-Induced Cognitive
Dysfunction Through Activation of ERK1/2-CREB-BDNF Pathway in Anesthetized Mice. Neurochem Res. 42(2): 501-512. doi:10.1007/s11064-016-2097-4. Wang Y, Hai B, Niu X, Ai L, Cao Y, Li R, et al. 2016. Chronic intermittent hypoxia
disturbs insulin secretion and causes pancreatic injury via MAPK signaling pathway. Biochem Cell Biol. doi:10.1139/bcb-2016-0167. Yan S, Wang Y, Liu P, Chen A, Chen M, Yao D, et al. 2016. Baicalin Attenuates
Hypoxia-Induced Pulmonary Arterial Hypertension to Improve Hypoxic Cor Pulmonale by Reducing the Activity of the p38 MAPK Signaling Pathway and MMP-9. Evid Based Complement Alternat Med. 2016: 2546402. doi:10.1155/2016/2546402. Yuan XH, Fan YY, Yang CR, Gao XR, Zhang LL, Hu Y, et al. 2016. Progesterone amplifies oxidative stress signal and promotes NO production via H2O2 in mouse
kidney arterial endothelial cells. J Steroid Biochem Mol Biol. 155(Pt A): 104-111. doi:10.1016/j.jsbmb.2015.09.029. Zhang S, Guo Z, Yang S, Ma H, Fu C, Wang S, et al. 2016. Chronic intermittent
hybobaric hypoxia protects against cerebral ischemia via modulation of mitoKATP. Neurosci Lett. 635: 8-16. doi:10.1016/j.neulet.2016.10.025. Zhang Y, Zhong N, Zhu HF, Zhou ZN. 2000. Antiarrhythmic and antioxidative effects
of intermittent hypoxia exposure on rat myocardium. Acta Physiol. Sin. . 52: 4 Zhang Y, Zhou ZN. 2012. [Beneficial effects of intermittent hypobaric hypoxia on the body]. Zhongguo Ying Yong Sheng Li Xue Za Zhi. 28(6): 504-509
Page 25 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
25
25
Figure captions
Figure 1 Effect of CIHH on vasodilation and expression of p-eNOS /
eNOS on aortic rings. A, CIHH improved the impaired Ach (1 nmol / L
to 10 µmol / L)-induced relaxation on aortic rings in the 2K1C group. B,
No change of SNP (1 nmol / L to 1 µmol / L)-induced relaxation was
observed on aortic rings in the 2K1C group compared with the Sham
group, and CIHH had no protective effect on the endothelial relaxation. C,
CIHH reversed the decreased expression of p-eNOS / eNOS on aortic
rings in the 2K1C group. All data are expressed as the mean ± SEM, n =
6 for Sham, CIHH and 2K1C+ CIHH groups, n = 5 for 2K1C group. **P
< 0.01 vs. Sham group; †P < 0.05 and
††P < 0.01 vs. 2K1C group.
Figure 2 Effects of CIHH treatment on expression of HIF-1α and HIF-2α
in aortic rings. A, Immunofluorescence staining showed the fluorescent
intensities of HIF-1α (green) and HIF-2α (red) in endothelial cells were
significantly increased in the CIHH group, compared with the Sham
group. Similarly, they were also higher in the 2K1C + CIHH group
compared with 2K1C group. B, Western blot showed that HIF-1α and
HIF-2α were both up-regulated in CIHH and 2K1C + CIHH groups. All
data are expressed as the mean ± SEM, n = 5 for each group. Scale bar =
Page 26 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
26
26
50 μm. **P < 0.01 vs. Sham group;
†P < 0.05 and
††P < 0.01 vs. 2K1C
group.
Figure 3 Effects of antagonists of MAPKs on the endothelial vasodilation
of aortic rings in the 2K1C group and effect of CIHH on expression of
MAPKs. A, Impaired vasodilation was rescued by preincubation of p38
antagonist SB203580 (10 µmol / L), but not by ERK1/2 antagonist
PD98059 (10 µmol / L) or JNK antagonist SP600125 (10 µmol / L). B, C,
Expression of p-p38 / p38 was increased obviously in the 2K1C group,
but it was decreased by CIHH treatment in the 2K1C + CIHH group. The
expression of p-ERK1/2 / ERK1/2 and p-JNK / JNK showed no obvious
changes with or without CIHH treatment in the 2K1C group. All data are
expressed as the mean ± SEM, n = 5 for each group. **P < 0.01 vs. Sham
group; ††P < 0.01 vs. 2K1C group.
Figure 4 Effects of BMP-4 antagonist Noggin on the impaired
endothelium-dependent relaxation in 2K1C aortic rings and effect of
CIHH on BMP4 expression. A, The impaired Ach-induced vasodilation
was rescued by incubation of BMP4 antagonist Noggin (100 ng / mL). B,
CIHH treatment inhibited BMP4 expression in the 2K1C + CIHH group.
All data are expressed as the mean ± SEM, n = 4 for 2K1C + Noggin
Page 27 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
27
27
group, n = 5 for other groups. **P < 0.01 vs. Sham group;
††P < 0.01 vs.
2K1C group.
Figure 5 Effects of p38 antagonist SB203580 and BMP-4 antagonist
Noggin on expression of p-eNOS / eNOS and p-p38 / p38. The
expression of p-eNOS was up-regulated and the expression of p-p38 was
down-regulated by preincubation of the two antagonists compared with
the 2K1C group. All data are expressed as the mean ± SEM, n = 5 for
each group. **P < 0.01 vs. Sham group;
††P < 0.01 vs. 2K1C group.
Page 28 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Table 1. The changes of bodyweight in rats on CIHH and/or 2K1C treatment
0 week(g) 1 week(g) 2 week(g) 3 week(g) 4 week(g) 5 week(g) 6 week(g) 7 week(g) 8 week(g)
Sham 182.5±2.0 184.3±3.1 193.7±3.6 206.8±5.2 229.3±5.8 250.0±7.0 277.8±8.1 319.2±9.2 354.8±9.3
CIHH 182.3±2.3 188.2±2.8 193.8±3.7 204.8±4.9 222.0±6.3† 247.0±5.3
†† 271.8±7.6
†† 318.2±8.9
†† 345.5±9.4
††
2K1C 178.7±1.9 185.3±2.8 190.3±3.7 196.7±4.8 212.3±5.6** 216.2±5.9** 226.7±6.1** 255.0±7.5** 278.0±9.5**
2K1C+CIHH 181.2±1.8 187.2±3.0 191.7±3.8 201.3±5.0 220.8±5.5† 249.7±6.0
† 265.0±7.7
†† 298.2±8.0
†† 328.8±10.3
††
All data are expressed as mean ± SEM, n=6 for each group. *P < 0.05 and
**P < 0.01 vs. Sham group;
†P < 0.05 and
††P <
0.01 vs. 2K1C group. CIHH treatment was given from the fifth week in CIHH and 2K1C+CIHH group.
Page 29 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Table 2. The changes of systolic blood pressure in rats on CIHH and/or 2K1C treatment
Group
0 week
(mmHg)
1 week
(mmHg)
2 week
(mmHg)
3 week
(mmHg)
4 week
(mmHg)
5 week
(mmHg)
6 week
(mmHg)
7 week
(mmHg)
8 week
(mmHg)
Sham 107.5±2.3 110.0±2.0 112.3±2.5 110.0±3.2 116.2±2.9 111.0±3.4 111.3±4.6 114.7±4.1 115.0±5.4
CIHH 108.5±1.9 108.3±2.2†† 112.7±2.6
†† 114.2±3.7
†† 112.8±3.3
†† 115.2±5.2
†† 113.5±4.1
†† 115.8±4.2
†† 116.0±4.7
††
2K1C 108.2±3.0 123.3±2.8** 166.8±3.5** 169.2±2.4** 169.0±2.9** 168.2±2.5** 172.2±6.4** 168.2±5.5** 169.0±4.7**
2K1C+CIHH 108.2±4.3 117.7±3.2 166.3±3.6 164.3±3.0 164.2±5.4 147.7±3.2†† 143.7±3.1
†† 139.3±4.6
†† 140.3±3.9
††
All data are expressed as mean ± SEM, n=6 for each group. *P < 0.05 and
**P < 0.01 vs. Sham group;
†P < 0.05 and
††P <
0.01 vs. 2K1C group. CIHH treatment was given from the fifth week in CIHH and 2K1C+CIHH group.
Page 30 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
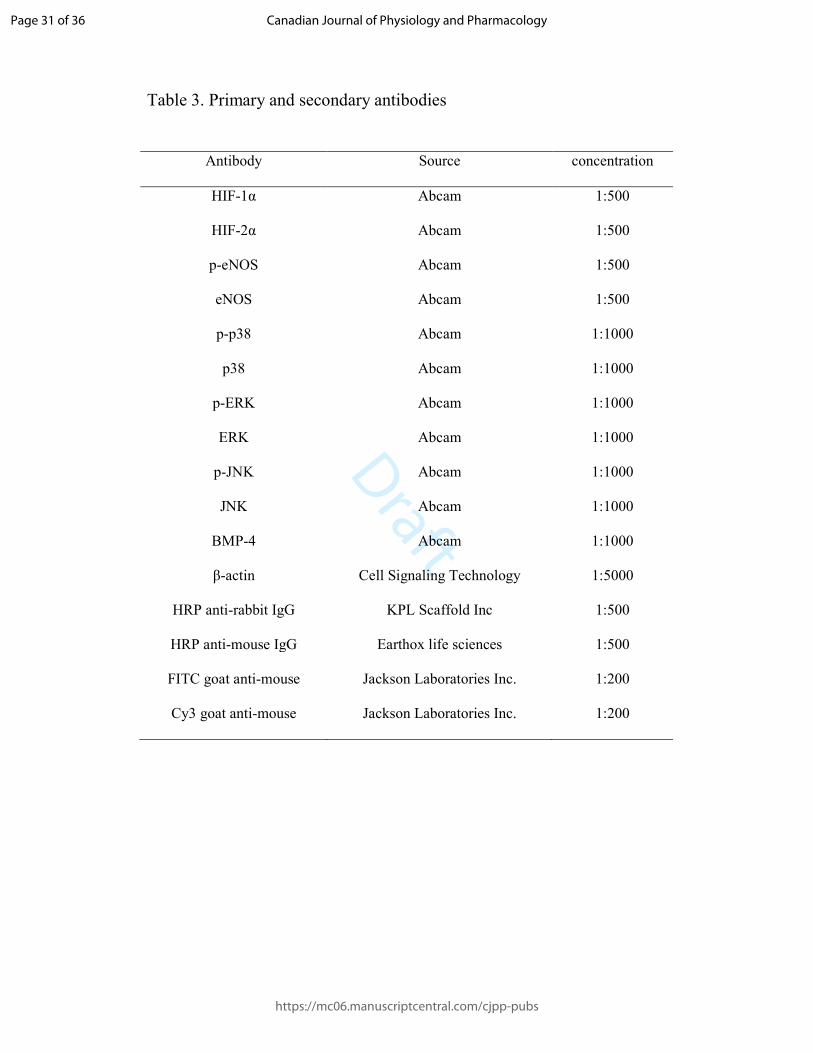
Table 3. Primary and secondary antibodies
Antibody Source concentration
HIF-1α Abcam 1:500
HIF-2α Abcam 1:500
p-eNOS Abcam 1:500
eNOS Abcam 1:500
p-p38 Abcam 1:1000
p38 Abcam 1:1000
p-ERK Abcam 1:1000
ERK Abcam 1:1000
p-JNK Abcam 1:1000
JNK Abcam 1:1000
BMP-4 Abcam 1:1000
β-actin Cell Signaling Technology 1:5000
HRP anti-rabbit IgG KPL Scaffold Inc 1:500
HRP anti-mouse IgG Earthox life sciences 1:500
FITC goat anti-mouse Jackson Laboratories Inc. 1:200
Cy3 goat anti-mouse Jackson Laboratories Inc. 1:200
Page 31 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology
Draft
Figure 1 Effect of CIHH on vasodilation and expression of p-eNOS / eNOS on aortic rings.
86x65mm (300 x 300 DPI)
Page 32 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Figure 2 Effects of CIHH treatment on expression of HIF-1α and HIF-2α in aortic rings.
86x58mm (300 x 300 DPI)
Page 33 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Figure 3 Effects of antagonists of MAPKs on the endothelial vasodilation of aortic rings in the 2K1C group and effect of CIHH on expression of MAPKs.
86x60mm (300 x 300 DPI)
Page 34 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Figure 4 Effects of BMP-4 antagonist Noggin on the impaired endothelium-dependent relaxation in 2K1C aortic rings and effect of CIHH on BMP4 expression.
86x60mm (300 x 300 DPI)
Page 35 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology

Draft
Figure 5 Effects of p38 antagonist SB203580 and BMP-4 antagonist Noggin on expression of p-eNOS / eNOS and p-p38 / p38.
86x66mm (300 x 300 DPI)
Page 36 of 36
https://mc06.manuscriptcentral.com/cjpp-pubs
Canadian Journal of Physiology and Pharmacology