
UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE BIOLOGIA
CURSO DE PÓS-GRADUAÇÃO EM MEDICINA VETERINÁRIA
PARASITOLOGIA VETERINÁRIA
ALGUNS ASPECTOS BIOECOLÓGICOS DE Raillietia TROUESSART, 1902
(ACARI: GAMASIDA) PRINCIPAL AGENTE DA OTOCARÍASE
PARASITÁRIA DOS BOVINOS
ARLINDO LUIZ DA COSTA
ITAGUAÍ - RIO DE JANEIRO
JULHO/1990

ALGUNS ASPECTOS BIOECOLÓGICOS DE Raillietia TROUESSART, 1902
(ACARI: GAMASIDA) PRINCIPAL AGENTE DA OTOCARÍASE
PARASITÁRIA DOS BOVINOS
ARLINDO LUZ DA COSTA
1990

UNIVERSIDADE FEDERAL RURAL DO RIO DE JANEIRO
INSTITUTO DE BIOLOGIA
CURSO DE PÓS-GRADUAÇÃO EM MEDICINA VETERINÁRIA
PARASITOLOGIA VETERINÁRIA
ALGUNS ASPECTOS BIOECOLÓGICOS DE Raillietia TROUESSART, 1902
(ACARI: GAMASIDA) PRINCIPAL AGENTE DA OTOCARÍASE
PARASITÁRIA DOS BOVINOS
ARLINDO LUIZ DA COSTA
SOB A ORIENTAÇÃO DO PROFESSOR
DR. JOÃO LUIZ HORÁCIO FACCINI
Tese submetida à defesa como
requisito parcial para obten-
ção do Grau de Doutor em Ciên-
cias, Medicina Veterinária-
Parasitologia Veterinária.
ITAGUAÍ, RIO DE JANEIRO
JULHO, 1990

TÍTULO DA TESE
ALGUNS ASPECTOS BIOECOLÓGICOS DE Raillietia TROUESSART, 1902
(ACARI: GAMASIDA) PRINCIPAL AGENTE DA OTOCARÍASE
PARASITÁRIA DOS BOVINOS
AUTOR
ARLINDO LUIZ DA COSTA
TESE APROVADA EM: 27/07/1990
JOÃO LUIZ HORÁCIO FACCINI
DAISY WILWERTH DA CUNHA
ADIVALDO HENRIQUE DA FONSECA
RUBENS PINTO DE MELLO
RÔMULO CERQUEIRA LEITE

iv.
Dedico este trabalho
à minha mulher, Rosa
Maria, aos meus fi-
lhos Sandro Moretti
e Ayla Mércia e à mi-
nha sobrinha Carmi-
nha, pelo incentivo,
pelo apoio, pelo li-
nitivo às dificulda-
des, a mim proporci-
nadas por suas pre-
senças ao meu lado.

V.
À minha prima e amiga
D. Maria do Socorro
G. Gama.
e ao meu cunhado Ed-
son Varêda Guima-
rães, pela ajuda e
apoio nos momentos
em que deles preci-
s e i .

A G R A D E C I M E N T O S
Ao Prof. JOÃO LUIZ HORÁCIO FACCINI, professor adjunto
em Parasitologia Veterinária da Universidade Federal Rural do
Rio de Janeiro, orientador desta Tese, pela incansável ajuda,
estímulo e confiança em nós depositada.
Ao Prof. NICOLAU MAUÉS DA SERRA FREIRE, professor ad-
junto em Parasitologia Veterinária da UFRRJ, pela orientação,
amizade e colaboração durante a execução deste trabalho.
Aos Profs. CARLOS LUIZ MASSARD e NICOLAU MAUÉS DA SER-
RA FREIRE, que se sucederam na Coordenação do Curso de Pós-Gra-
duação em Medicina Veterinária: Parasitologia Veterinária da
UFRRJ pela acolhida e apoio durante o Curso.
Ao Prof. CARLOS WILSON GOMES LOPES, professor adjun-
to em Parasitologia Veterinária da UFRRJ pela amizade, ajuda
na execução das fotografias e pelos conhecimentos a nós por
ele repassados.
Ao Prof. RÔMULO CERQUEIRA LEITE, Prof. Adjunto do De-
partamento de Medicina Veterinária Preventiva da Escola de Ve-

vii.
terinária da Universidade Federal de Minas Gerais, que conosco
conviveu por um período, pela amizade, ajuda e apoio, fatores
decisivos para a condução deste trabalho.
Professora ELISA HELENA DE SOUZA FARIA do Departamen-
to de Matemática da UFRRJ pela orientação e execução da parte
estatística deste trabalho.
Ao Prof. ADIVALDO HENRIQUE DA FONSECA do Departamen-
to de Epidemiologia e Saúde Pública do Instituto de Veterinária
da UFRRJ pela confecção dos gráficos deste trabalho.
Aos Professores membros da Banca Examinadora desta Te-
se, pelas críticas valiosas, pelas orientações, cujo objetivo
principal é a afirmação científica deste trabalho.
A todos os Professores do Curso de Pós-Graduação em Me-
dicina Veterinária: Parasitologia Veterinária da UFRRJ, pelos
conhecimentos transmitidos.
Aos funcionários da UFRRJ que conosco conviveram neste
período, pelo apoio e colaboração a nós dispensada.
Aos colegas de curso, pela amizade solidificada nos mo-
mentos difíceis e nas vitórias que juntos partilhamos.
À Empresa Brasileira de Pesquisa Agropecuária-EMBRAPA,
através da área de Desenvolvimento de Pessoal do Departamento
de Recursos Humanos-DRH e da UEPAE de Rio Branco, pela oportuni-
dade ímpar a nós concedida e pelo apoio que jamais faltou.
Comissão de Aperfeiqoamento de Pessoal de Nível Su-
perior CAPES) do Ministério da Educação e Cultura pela Bolsa
de Estudos a nós concedida.
Ao Instituto de Zootecnia da UFRRJ, que através do seu

viii.
Setor de Gado de Corte nos cedeu os bovinos utilizados em par-
te dos experimentos.
Ao Núcleo de Pesquisa de Saúde Animal, NPSA, da EM-
BRAPA, Km 47, pelos ovinos e caprinos a nós cedidos para rea-
lização de parte dos experimentos.
Direção dos Matadouros de Rio Branco, Acre e de Ni-
lópolis, RJ, pela permissão e o nosso acesso à coleta de mate-
rial durante 24 meses.
A todos os animais que involuntariamente prestaram-se
realização deste trabalho.
Ao Sr. GILMAR FERREIRA VITA pelos serviços de datilo-
g r a f i a .
A todos, enfim, que nos ajudaram, o nosso sincero mui-
to obrigado.

BIOGRAFIA
ARLINDO LUIZ DA COSTA, filho de Raimundo Luiz da Costa
e Maria do Carmo Araújo, e natural de Mocambo, Ceará, onde nas-
ceu a 28 de março de 1945.
Cursou o primário em Mocambo, o ginasial e o colegial
em Fortaleza, no Colégio Estadual do Ceará.
Em 1967 ingressou no Curso de Medicina Veterinária da
Faculdade de Veterinária do Ceará, graduando-se em 12 de dezem-
bro de 1970.
Durante o período de março de 1971 a julho de 1972 tra-
balhou na Faculdade de Veterinária do Ceará na categoria de Co-
laborador de Ensino da Disciplina Fisiologia dos Animais Domés-
ticos.
Em julho de 1972 transferiu-se para a Secretaria de A-
gricultura, Indústria e Comércio do Estado do Acre onde perma-
neceu até maio de 1976, ocupando durante este período alguns
cargos como Chefe do Laboratório de Análises Clínicas, Diretor
do Departamento de Produção Animal (DPA), Executor do Programa

Nacional de Sanidade Animal (PRONASA) e Membro do Grupo Execu-
tivo de Saúde Animal (GESA).
Em dezembro de 1976 concluiu Licenciatura Plena em Pe-
dagogia na Universidade Federal do Acre-UFAC.
Em maio de 1976 ingressou na Empresa Brasileira de
Pesquisa Agropecuária - EMBRAPA, na categoria de Pesquisador
integrando-se à equipe de Produção Animal onde permaneceu até
1979, quando passou a integrar a linha de pesquisa em Sanida-
de Animal da UEPAE de Rio Branco.
Em 1980 foi admitido e se matriculou no Curso de Mes-
trado em Medicina Veterinária: Parasitologia Veterinária, na
Universidade Federal Rural do Rio de Janeiro - UFRRJ, tendo de-
fendido Tese em 04 de marco de 1982.
No período 1982-1986 desenvolveu atividades de pesqui-
sa em Produção e Sanidade Animal na EMBRAPA, UEPAE de Rio Bran-
co, tendo publicado vários trabalhos, com destaque para os re-
lativos às pesquisas iniciais sobre o Carrapato dos Bovinos e
Helmintos Gastrintestinais em Gado de Leite, apresentados em
1984 no I Simpósio Internacional do Trópico Úmido, Belém, Pa-
rá, e hoje plenamente integrados aos sistemas de produção em
uso na região.
Em 1987 foi admitido no Curso de Pós-Graduação em Me-
dicina Veterinária, a nível de Doutorado na UFRRJ, como inte-
grante do Projeto de pós-Graduação no País, do Departamento de
Recursos Humanos (DRH) da EMBRAPA.

ÍNDICE
INTRODUÇÃO
DINÂMICA DE PARASITISMO EM DUAS REGIÕES ECOLOGICAMENTE
DISTINTAS
Revisão da Literatura
Material e Métodos
Resultados e Discussão
ESPECIFICIDADE PARASITÁRIA
Revisão da Literatura
Material e Métodos
Resultados e Discussão
CICLO BIOLÓGICO "IN VIVO": CONSIDERAÇÕES PRELIMINARES
Revisão da Literatura
Material e Métodos
Resultados e Discussão
CONCLUSÕES
REFERÊNCIAS BIBLIOGRÁFICAS
APÊNDICE
Página
1
3
3
6
12
31
31
33
36
38
38
39
43
52
54
65

ÍNDICE DAS TABELAS
Dinâmica de população de Raillietia Troues-
sart, 1902 em bovinos azebuados naturalmen-
te infestados nas microrregiões Alto Purús,
AC e Vale do Rio Doce, MG, 1988
Página
13
Freqüência mensal de Raillietia Trouessart,
1902 em 30 bovinos azebuados da microrre-
gião Alto Purús, AC, Jan. 1987 a Dez. 1988. 15
Freqüência média mensal de Raillietia Troues-
sart, 1902 em 30 bovinos azebuados da mi-
crorregião Alto Purús, AC. Jan. 1987 a Dez.
1988 16
Freqüência mensal de Raillietia Trouessart,
1902 em 30 bovinos azebuados da microrre-
gião Vale do Rio Doce, MG, Nov. 1987 a Out.
1989 20
TABELA 1.
TABELA 2.
TABELA3.
TABELA 4.

xiii.
Página
21
26
27
29
30
44
Freqüência média mensal de Raillietia Troues-
sart, 1902 em 30 bovinos azebuados da micror-
região Vale do Rio Doce, MG. Nov. 1987 a Out.
1989
Freqüência média mensal de fêmeas de Raillie-
tia Trouessart, 1902 em 30 bovinos azebua-
dos da microrregião Alto Purús, AC, analisa-
da pelo teste de Tukey (P < 0,05)
Freqüência média mensal de larvas de Raillie-
tia Trouessart, 1902 em 30 bovinos azebua-
dos da microrregião Alto Purús, AC, analisa-
da pelo teste de Tukey (P < 0,05)
Freqüência média mensal de fêmeas de Raillie-
tia Trouessart, 1902 em 30 bovinos azebua-
dos da microrregião Vale do Rio Doce, MG,
analisada pelo teste de Tukey (P < 0,05)
Freqüência média mensal de larvas de Raillie-
tia Trouessart, 1902 em 30 bovinos azebua-
dos da microrregião Vale do Rio Doce, MG, a-
nalisada pelo teste de Tukey (P < 0,05)
Duração do ciclo biológico, de adulto jovem
até larva, de Raillietia Trouessart, 1902 "in
vivo" em bovinos susceptíveis. Itaguaí, RJ.
1990
TABELA 5.
TABELA 6.
TABELA 7.
TABELA 8.
TABELA 9.
TABELA 10.

xiv.
Página
49
66
67
68
69
70
Recuperação de formas de vida livre do ci-
clo biológico de Raillietia Trouessart,
1902 Itaguaí, RJ 1990 • •
Número de Raillietia Trouessart, 1902 em
30 bovinos azebuados naturalmente infesta-
dos. Alto Purús, AC, 1988
Número de Raillietia Trouessart, 1902 em
30 bovinos azebuados naturalmente infesta-
dos. Vale do Rio Doce, MG, 1989
Análise de variância da freqüência média
mensal de fêmeas de Raillietia Trouessart,
1902 em 30 bovinos azebuados da microrre-
gião Alto Purús, AC, no período de 24 me-
ses
Análise de variância da freqüência média
mensal de larvas de Raillietia Trouessart,
1902 em 30 bovinos azebuados da microrre-
gião Alto Purús, AC, no período de 24 me-
ses
Análise de variância da freqüência média
mensal de fêmeas de Raillietia Trouessart,
1902 em 30 bovinos azebuados da microrre-
gião Vale do Rio Doce, MG, no período de
24 meses
TABELA 11.
TABELA 12.
TABELA 13.
TABELA 14.
TABELA 15.
TABELA 16.

xv.
Página
71
72
73
Análise de variância da freqüência média men-
sal de larvas de Raillietia Trouessart, 1902
em 30 bovinos azebuados da microrregião Vale
do Rio Doce, MG no período de 24 meses
Dados climáticos da microrregião Alto Purús,
AC. Janeiro de 1987 a Dezembro de 1988
Dados climáticos da microrregião Vale do Rio
Doce, MG. Novembro de 1987 a Outubro de 1989
TABELA 17.
TABELA 18.
TABELA 19.

Página
7
8
10
17
18
ÍNDICE DAS FIGURAS
Localização geográfica das duas regiões cria-
tórias estudadas: Alto Purús, AC e Vale do
Rio Doce, MG
Dados climáticos da microrregião Alto Purús,
AC
Dados climáticos da microrregião Vale do Rio
Doce, MG
Freqüência média mensal de Raillietia Troues-
sart, 1902 em 30 bovinos azebuados da micror-
região Alto Purs, AC. 1987-1988
Incidência mensal de fêmeas e larvas de Boo-
philus microplus (Canestrini, 1887). Alto Pu-
rús, AC (COSTA, 1984)
FIGURA 1.
FIGURA 2.
FIGURA 3.
FIGURA 4.
FIGURA 5.

xvii.
Página
22
34
45
47
50
Freqüência média mensal de Raillietia Troues-
sart, 1902 em 30 bovinos azebuados da micror-
região Vale do Rio Doce, MG, Novembro de 1987
a Outubro de 1989
Localização geográfica das coletas em capri-
nos e ovinos, criados juntos com bovinos, pa-
ra estudo de especificidade parasitária de
Raillietia Trouessart, 1902
Estádios do ciclo biológico "in vivo" de Rai-
llietia Trouessart, 1902 obtidos a partir da
infestação de bovinos susceptíveis
Ciclo biológico de Raillietia Trouessart,
1902. Tentativa diagramática de representação
Formas de vida livre do ciclo biológico de
Raillietia Trouessart, 1902 recuperadas da
pastagem
FIGURA 6.
FIGURA 7.
FIGURA 8.
FIGURA 9.
FIGURA l0.

R E S U M O
Foram realizados estudos sobre a dinâmica do parasi-
tismo, especificidade e ciclo biológico "in vivo" de Raillie-
tia Trouessart, 1902, parasito do conduto auditivo de bovi-
nos. Na microrregião Alto Purús, Acre na Amazônia Ocidental
o parasitismo por Raillietia em bovinos foi de 100%, com in-
tensidade de infestação de 3 a 221 e intensidade média de 69
ácaros/animal. Dois piques definidos nos meses de maio e no-
vembro, limites entre os períodos chuvoso e seco, foram ob-
servados nesta região. Na microrregião Vale do Rio Doce, Mi-
nas Gerais no Sudeste Brasileiro, o parasitismo por Raillie-
tia em bovinos foi de 100%, com intensidade de infestação de
19 a 144 e intensidade média de 53 ácaros/animal. A infesta-
ção foi mais intensa no período de novembro a marco, verão
quente e chuvoso nesta região. Raillietia se comportou como
um parasito dotado de elevada especificidade com relação a bo-
vinos. O ciclo biológico, de adulto jovem a larva dura de 4-6
dias. A protoninfa e o adulto jovem foram as fases de vida li-

xix.
vre recuperadas das pastagens.

Research on the dynamics of parasitism by ear mites
of the genus Raillietia Trouessart, 1902 in cattle were
carried out in Alto Purús, State of Acre in the West Amazon and
Vale do Rio Doce, State of Minas Gerais in Southeastern Brazil.
Prevalence were 100% in both regions. Density of infestation
ranged from 3 to 221 mites/ear in Alto Purús and from 19 to
144 mites/ear in Vale do Rio Doce. The mean intensity of
infestation was, respectively, 69 and 53 mites/cattle. In both
regions the mites populations exhibited a seasonal pattern. In
Alto Purús, the mean intensity was higher in may and november
(limits of the rainy and day seasons). Only one peak of
infestation was seen from november to march (dry and hot
summer) in Vale do Rio Doce.
Experimental transfer of mites from cattle to goats
and sheep has not been successful.
The life cycle from teneral adults to larva took
SUMMARY

x x i .
4-6 days. Only the protonynph and teneral adult were collected
in the pastures.

Ácaros do gênero Raillietia Trouessart, 1902 são para-
sitos do conduto auditivo externo e da superfície externa da mem-
brana timpânica dos seus hospedeiros, são dotados de elevado po-
tencial de disseminação e considerados como os principais agen-
tes da otocaríase parasitária dos ruminantes domésticos.
Estes ácaros, como parasitos de bovinos, bubalinos e
caprinos, tem sido citados em estudos realizados em diferentes
continentes (LEIDY, 1872, OLSEN & BRACKEN, 1950, QUINTERO, 1978,
QUINTERO et alii, 1980, NUNES et alii, 1972, FONSECA, 1983, FON-
SECA et alii, 1983 nas Americas; TROUESSART, 1902a,b,c, FREUND,
1910 na Europa; ALWAR et alii, 1959, RAK & NAGHSHINEH, 1973 na
Ásia; FERGUSON & LAVOIPIERRE, 1962 na África; DONROW, 1963,
LADDS et alii, 1972 na Oceania), o que comprova sua ampla dis-
tribuição geográfica e, segundo FACCINI (1990) em dados ainda
não publicados, ao nível de Brasil os atuais conhecimentos per-
mitem afirmar que têm sido identificadas as seguintes espécies:
R. auris (Leidy, 1872) parasitando bovinos, Raillietia sp. n.
INTRODUÇÃO

parasitando bubalinos e bovinos e R. caprae Quintero, Bassols
& Acevedo, 1980 parasitando caprinos e ovinos.
A maioria dos estudos sobre Raillietia refere-se a
R. auris e tem ocorrido através de achados ocasionais de ne-
cropsias. A partir de resultados mais recentes obtidos sobre
sua patologia, R. auris é, atualmente considerada como um im-
portante parasito de rebanho, capaz de provocar lesões e seque-
las de caráter severo em seus hospedeiros, principalmente nas
infestações de grande porte, embora a importância econômica
destas lesões ainda seja desconhecida (MENZIES, 1957; NUNES,
1977; HEFFNER & HEFFNER, 1983a, LEITE et alii, 1987; McKEE et
alii, 1988; LEITE, 1989).
Devido a carência de informações sobre parâmetros
mais consistentes sobre a bioecologia de Raillietia, este es-
tudo enfoca alguns aspectos como prevalência, freqüência rela-
tiva, intensidade de infestação, estrutura de população e sa-
zonalidade em duas regiões ecologicamente distintas do Terri-
tório Brasileiro (Amazônia Ocidental e Sudeste), ciclo bioló-
gico "in vivo" e especificidade parasitária.
Durante a realização do presente trabalho, ficaram
evidenciadas as presenças, tanto de R. auris como Raillietia
sp. n. parasitando os bovinos por nós estudados. Visto que,
estudos detalhados de descrição desta nova espécie estão em
andamento, os dados aqui registrados se referem apenas a áca-
ros do gênero Raillietia, sem levar em conta a espécie.
2.

DINÂMICA DE PARASITISMO EM DUAS REGIÕES ECOLOGICAMERTE DISTINTAS
Revisão da Literatura
Os ácaros do gênero Raillietia (Gamasida = Mesostigmata)
são parasitos obrigatórios do conduto auditivo externo de bovi-
nos e/ou de outros ruminantes. O atual estágio de conhecimento
sobre este gênero permite-nos considerar que, no Brasil foram ob-
servadas as espécies R. auris parasitando bovinos; R. caprae pa-
rasitando ovinos e caprinos e Raillietia sp. n. parasitando buba-
linos e bovinos (FACCINI, 1990 comunicação pessoal).
Dados registrados na literatura pertinente indicam que
o parasitismo por Raillietia é capaz de produzir em bovinos fre-
qüentes e sérios casos de otites (FREUND, 1910; OLSEN & BRACKEN,
1950; MENZIES, 1957; LADDS et alii, 1972; NUNES et alii, 1975;
McKEE et alii, 1988); paralisia facial e bloqueio do conduto au-
ditivo (FERGUSON & LAVOIPIERE, 1982); perda completa de audição
(HEFFNER & HEFFNER, 1983a) ; abscedação com proliferação bacteria-
na e possível morte dos hospedeiros por encefalite (HEFFNER &

4.
HEFFNER, 1983b; LEITE et alii, 1987; LEITE, 1989).
Alguns aspectos da dinâmica de parasitismo em bovinos
têm sido estudados parcialmente por alguns autores. A ocorrên-
cia encontra-se registrada, quer em achados ocasionais de ne-
crópsias (LEIDY, 1872; RADFORD, 1938; MENZIES, 1957; SCHLOT-
THAUER, 1970; RAK & NAGHSHINEH, 1973) ou em observações de ca-
sos clínicos específicos de otites parasitárias provocadas pe-
lo ácaro (FREUND, 1910; OLSEN & BRACKEN, 1950; ALWAR et alii,
1959; FERGUSON & LAVOIPIERRE, 1962; LADDS et alii, 1972; NUNES
et alii, 1972; SHASTRI & DESHPANDE, 1983; McKEE et alii, 1988).
A prevalência e a intensidade de infestação sempre tiveram co-
mo base inquéritos epidemiológicos dirigidos de curta duração
em animais abatidos em matadouros (TROUESSART, 1902; NUNES et
alii, 1975; FACCINI et alii, 1976; OLIVEIRA, 1978; FONSECA & FON-
SECA, 1981; FONSECA, 1983; HEFFNER & HEFFNER, 1983a; JAVIER et
alii, 1983; BRUM et alii, 1986); a estrutura de população tam-
bém têm sido estudada em semelhantes condições, com resultados
que indicam a presença natural de ninfas no ciclo biológico do
parasito (MENZIES, 1957; NUNES et alii, 1975; OLIVEIRA, 1978;
QUINTERO, 1978; LARANJEIRA et alii, 1982; JAVIER et alii, 1983;
ROUX et alii, 1983) ou com dados que não registram em bovinos
naturalmente infestados a presença deste estádio de seu ciclo
evolutivo (FONSECA, 1983; BRUM et alii, 1986; FACCINI et alii,
1987; LEITE, 1989).
É importante ressaltar o estudo da prevalência de
Raillietia em diferentes categorias de animais de diversas ra-
Gas de bovinos efetuada por LEITE (1989) coma utilização de

5.
um método de coleta de parasitos em animais vivos adaptado da
técnica de FACCINI et alii (1987) em que revelou ser este áca-
ro protagonista de uma parasitose mais intensa nos bovinos de
raÇas azebuadas (Nelore, Guzera) criados de forma extensiva
e que vai se reduzindo à medida que se aproxima das raças de
aptidão leiteira (Holandês, Jersey) em regime semi-estabula-
d o .
Estudos específicos de sazonalidade restringem-se a-
penas as observações de TZYMBAL & LITVISHKO (1955) na Ucrâ-
nia, Rússia que efetuaram um estudo de ocorrência de R. auris
em bovinos levando em consideração o tempo em que foram proce-
didos as coletas de janeiro a dezembro com exeeção do mês de
agosto. Um total de 55 animais foram examinados em números ir-
regulares a cada mês sendo registradas as presenças de lar-
vas e adultos machos e fêmeas do parasito. Mesmo que de forma
inadequada para um estudo de variação sazonal estes dados re-
velam pequenos piques de freqüência nos meses de junho e se-
t e m b r o .
O objetivo deste trabalho foi registrar os dados so-
bre a dinâmica de parasitismo por Raillietia sobre bovinos
nas condições da microrregião Alto Purús, Estado do Acre na
Amazônia Ocidental Brasileira e da microrregião Vale do Rio
Doce, Estado de Minas Gerais no Sudeste Brasileiro.

6 .
Material e Métodos
Os estudos foram realizados através da colheita do pa-
rasito em matadouros realizados sobre bovinos procedentes de
duas regimes criatórias distintas (Figura 1).
A primeira região estudada foi o município de Senador
Guiomar, parte integrante da microrregião homogênea do Alto Pu-
rús e cujos animais foram abatidos no matadouro de Rio Branco,
Acre. A região apresenta uma topografia ligeiramente ondulada
com áreas de pastagens cultivadas de capim Colonião (Panicum
maximum), Braquiaria (Brachiaria decumbens) e Quicuio-da-Amazô-
nia (Brachiaria humidicola) (VALENTIM & COSTA, 1983) cortadas
por açudes e igarapés sendo sua latitude de 10°09'10" (S) e lon-
gitude 67044'28" (Wgr) (ANUÁRIO ESTATÍSTICO DO ACRE, 1982). O
clima é do tipo AWi de Köppen com índice pluviométrico elevado
e nítido período seco, sendo a época chuvosa de novembro a a-
bril e a época seca de maio a outubro, com os menores índices
de precipitação pluvial nos meses de junho a agosto. A umidade
relativa do ar tem uma média de 85% e a temperatura média é de
26°C (BASTOS, 1982).
O período de realização das colheitas foi de 24 me-
ses, de janeiro de 1987 a dezembro de 1988 e os dados meteoro-
lógicos registrados durante o experimento (Figura 2; Tabela 18
apêndice) foram fornecidos pela Estação Climatológica do Minis-
tério da Agricultura em Rio Branco, a cerca de 15 km das pro-
priedades fontes dos animais e integrante da mesma microrre-
gião homogênea.

7.
FIGURA 1. Localização geográfica das duas regiões criatórias es-
tudadas: = Microrregião Alto Purús, Acre; * = Mi-
crorregião Vale do Rio Doce, Minas Gerais. 1987-1989.

8.
FIGURA 2. Dados climáticos médios da microrregião Alto Purús,
Acre. janeiro/1987 - Dezembro/1988. Fonte: Delegacia
Federal da Agricultura, Acre.

9 .
Para o estudo de prevalência e intensidade de infesta-
tação e estrutura de população foram examinados 30 animais com
coletas individuais e lavagens nos condutos auditivos externos
direito e esquerdo. Para o estudo de sazonalidade foram reali-
zadas 24 coletas mensais de 30 animais abatidos perfazendo um
total de 720 bovinos, todos eles mestiços da raça Nelore.
Os animais da segunda região estudada procederam do
município de Governador Valadares, integrante da microrregião
Vale do Rio Doce, Minas Gerais e abatidos no matadouro de Niló-
polis, Rio de Janeiro. A região, segundo ARRUDA (1987), apre-
senta uma topografia ondulada e coberta por pastagens cultiva-
das onde predominam as gramíneas capins Colonião (Panicum ma-
ximum), Jaraguá (Hyparrhenia tufa) e Gordura (Melinis minuti-
flora). O seu clima é do tipo AW de Köppen, tropical úmido de
savana, com inverno seco e frio e verão quente e chuvoso. A tem-
peratura média dos meses mais frios é superior a 18°C e a pre-
cipitação dos meses mais secos é inferior a 60 mm. A altitude
é de 166 m e suas coordenadas são 18º51'0" de latitude sul e
41°56'18" de longitude WGr, a temperatura média compensada
de 24,1°C e a umidade relativa do ar tem uma média de 75°C (AN-
TUNES, 1986).
O período de realização das colheitas foi de novembro
de 1987 à outubro de 1989 e os dados climáticos registrados
durante o experimento (Figura 3; Tabela 19, apêndice) foram
fornecidos pelo 5º Distrito Meteorológico do Ministério da A-
gricultura sediado em Governador Valadares, Minas Gerais.
Para o estudo de prevalência, intensidade de infesta-

10.
FIGURA 3. Dados climáticos médios da microrregião Vale do Rio
Doce, Minas Gerais. Novembro/1987 - Outubro/1989.
Fonte: 5º Distrito Meteorológico, Governador Valada-
res, Minas Gerais.

1 1 .
ção e estrutura de populações foram examinados 30 animais com
coletas individuais e exames nos condutos auditivos externos di-
reito e esquerdo. Para o estudo de sazonalidade foram realiza-
das 24 coletas mensais de 30 animais abatidos, perfazendo um to-
tal de 720 bovinos todos mestiços das raças indubrasil, Guzerá,
Nelore e Gir.
O método de colheita utilizado consistiu de lavagem
dos condutos auditivos externos direito e esquerdo com aproxi-
madamente 50 ml de água para cada conduto através de pistola
dosificadora, munida de sonda mamária coma ponta cortada trans-
versalmente que ao injetar o líquido desaloja e retira os áca-
ros que estão no conduto auditivo e na membrana timpânica jun-
to aos processos infecciosos (FACCINI et alii, 1987).
O material coletado dos animais abatidos em matadouro
foi remetido ao Laboratório de Ácarologia da UFRRJ e os áca-
ros foram identificados e contados em sua totalidade utilizan-
do-se para isto o microscópio estereoscópico Wild M5. Os núme-
ros obtidos com relação a freqüência mensal principalmente de
larvas e fêmeas estádios presentes em maiores quantidades fo-
ram relacionadas corn os dados climáticos da região para defini-
ção da curva de freqüência mensal.
A análise estatística dos dados referentes à freqüên-
cia mensal de fêmeas e larvas foi efetuada através do método
da Diferença Minima Significativa (DMS) e Teste de F com os dados
transformados para Raiz Quadrada VX+0,5 e teste de Tukey 5% e por
meio da linguagem SISTANVA do microcomputador POLIMAX 201 DP1.
1 Prológica, São Paulo, Brasil.

12.
Resultados e Discussão
Na tabela 1, e nas tabelas 12 e 13 (apêndice) estão
sumarizados os dados relativos à prevalência, intensidade de
infestação e estrutura de população de Raillietia sobre 30 bo-
vinos azebuados coletados de forma individual e procedentes das
microrregiões Alto Purús, Acre e Vale do Rio Doce, Minas Ge-
rais. Em Alto Purús, AC, foram encontrados 2083 estádios do
ácaro assim distribuídos: 29 (1,4%) ovos, 808 (38,8%) larvas,
108 (5,2%) machos e 1138 (54,6%) fêmeas. A intensidade de in-
festação variou de 3 a 221 e a intensidade média foi de 69
ácaros/animal e a prevalência foi de 100%, de animais parasi-
tados. No Vale do Rio Doce, MG, foram encontrados 1583 está-
dios do ácaro assim distribudos: 35 (2,2%) ovos, 447 (28,3%)
larvas, 67 (4,2%) machos, 1034 (65,3%) fêmeas. A intensidadede
infestação variou de 19 a 144 e a intensidade média foi de 53
ácaros/animal e a prevalência foi de 100%. Estes dados estão de
acordo com aqueles obtidos para outras regiões do Brasil (NU-
NES et alii, 1975; OLIVEIRA, 1978; LARANJEIRA et alii, 1982;
FONSECA, 1983) inclusive com os dados obtidos no município de
Aimores no Vale do Rio Doce, MG por FONSECA (1983).
O pavilhão auricular, o conduto auditivo e a membra-
na timpânica parecem formar com o calor corporal e a umidade
um microclima favorável a evolução dos ácaros auriculares (DO-
RESTE, 1983). Nas espécies de ácaros que passam todo o ciclo
biológico no conduto auditivo, esse ambiente estável aparente-
mente permitiria um crescimento constante da população que po-

13.
TABELA 1. Dinâmica de população em infestações naturais de
Raillietia Trouessart, 1902 em bovinos azebuados
nas microrregiões Alto Purús, Acre e Vale do Rio
Doce, Minas Gerais, 1988.

14.
deria ser controlada somente em função da densidade parasitá-
ria e ou reações imunológicas por parte do hospedeiro. Nas es-
pécies que passam parte do ciclo biológico fora do hospedei-
ro como parece ser o caso dos ácaros do gênero Raillietia (ver
discussão no capítulo ciclo biológico) é de se esperar uma flu-
tuação de população de acordo com as condições climáticas lo-
cais. De fato é o que se observa, tanto na microrregião Alto
Purús, Acre como na microrregião Vale do Rio Doce, Minas Ge-
rais.
Na microrregião Alto Purús, Acre, observa-se dois
piques de incidência bem definidos nos meses de maio e novem-
bro (Tabelas 2 e 3; Figura 4) quando possivelmente ocorre um
equilíbrio entre os fatores integrantes das condições climá-
ticas (precipitação pluviométrica, umidade relativa do ar e
temperatura) pois correspondem respectivamente ao fim do pe-
ríodo chuvoso e início do período seco (maio) e fim do perío-
do seco e início do período chuvoso (novembro). Comportamen-
to semelhante foi observado para o carrapato Boophilus micro-
plus (Canestrini, 1887) em pastagens da região por COSTA
(1984) (Figura 5).
Nos meses de dezembro a abril, no rigor do período
chuvoso onde as pesadas e freqüentes chuvas são uma constan-
te na região elevando o índice pluviométrico anual para a fai-
xa de 1800 mm e tornando as pastagens excessivamente úmidas,
forma-se com isto um fator que contribui para uma redução na
população do ácaro que se torna impedido de completar seu ci-
clo com eficiência. Este fenômeno também tende a ocorrer na

TABELA 2. Freqüência mensal de Raillietia Trouessart, 1902 em 30 bovinos azebuados da microrregião
Alto Purús, Acre. 1987-1988.
Número antes da barra inclinada refere-se ao ano de 1987.
Número após a barra inclinada refere-se ao ano de 1988.

TABELA 3. Freqüência média mensal de Raillietia Trouessart, 1902 em 30 bovinos aze-
buados da microrregião Alto Purús, Acre, Jan. 1987 a Dez. 1988.

FIGURA 4. Freqüência média mensal de Raillietia Trouessart, 1902 em 30 bovinos azebuados da
microrregião Alto Purús, Acre. 1987-1988.

18.
FIGURA 5. Incidência mensal de fêmeas ( ) e larvas (----)
de Boophilus microplus (Canestrini, 1887). Alto Pu-
rús, Acre, 1982-1984 (COSTA, 1984).

19.
distribuição sazonal do carrapato dos bovinos B. microplus um
ectoparasito que apresenta formas de vida livre nas pastagens
cujo ciclo biológico tende a ser limitado pelas chuvas pesadas
e pelo excesso de água (WILKINSON, 1955; OLIVEIRA, 1976; COS-
TA, 1982).
Por outro lado, nos meses correspondentes ao período
seco em Alto Purús, Acre, junho a outubro, a população de Rai-
llietia se mantém reduzida. Possivelmente, o fator temperatu-
ra tenha alguma influência pois a variação desta no período
vem se manifestar desde a redução por tempo limitado (junho)
devido ao fenômeno da queda brusca de temperatura que é uma
conseqüência do degelo da Cordilheira dos Andes no País vizi-
nho Bolívia até o incremento da temperatura de agosto a outu-
bro provocado pelas queimadas da floresta amazônica, tornando
o ambiente quente a tal ponto que a umidade relativamente ele-
vada da região se torna incipiente e assim venha possivelmente
ocorrer a morte por desidratação e ressecamento de parte das
formas de vida livre que circulam nas pastagens visto que Rai-
llietia é um ácaro Gamasida portador de uma delicada e fina
cutícula protetora e por isto mesmo bastante sensível à eleva-
ção de temperatura e à redução da umidade relativa. Estas ob-
servações correspondem à descrição das condições ecológicas
ideais para a permanente evolução de ácaros e insetos onde a
umidade relativa dos períodos quentes e secos tendem a limitar
o ciclo destes ectoparasitos (DORESTE, 1983).
Os dados registrados nas tabelas 4 e 5 e na figura 6
sugerem que na microrregião Vale do Rio Doce, Minas Gerais a

TABELA 4. Freqüência mensal de Raillietia Trouessart, 1902 em 30 bovinos azebuados da microrre-
gião Vale do Rio Doce, Minas Gerais, Novembro de 1987 a Outubro de 1989.

TABELA 5. Freqüência média mensal de Raillietia Trouessart, 1902 em 30 bovinos
azebuados da microrregião Vale do Rio Doce, Minas Gerais, Nov. 1987 a
Out. 1989.

FIGURA 6. Freqüência média mensal de Raillietia Trouessart, 1902 em 30 bovinos azebuados da
microrregião Vale do Rio Doce, Minas Gerais. Novembro de 1987 a Outubro de 1989.

23.
infestação dos bovinos por Raillietia se eleva durante os me-
ses de novembro a março correspondente ao período de verão quen-
te e chuvoso com elevação da umidade relativa. Esta microrre-
gião por apresentar um índice de pluviosidade com variação de
700 a 1100 mm aproximadamente e pelo fato de que 50% destas chu-
vas ocorreram neste período (ANTUNES, 1986) caracteriza um índi-
ce pluviométrico de porte médio que forneceria aos ácaros a umi-
dade adequada do ambiente favorável ao seu ciclo biológico ape-
sar da temperatura elevada. Os resultados aqui obtidos são se-
melhantes com o estudo da sazonalidade de larvas de Dermatobia
hominis (Linnaeus, Jr., 1781) em bovinos daquela microrregião
realizado por MAIA & GUIMARÃES (1985) quando o verão quente e
chuvoso fez aumentar o número de insetos transportadores e como
conseqüência o número de larvas nos animais hospedeiros.
Insetos e ácaros completam seu desenvolvimento de for-
ma mais lenta nos períodos frios e secos (NETO et alii, 1976),
com conseqüente redução de população (DORESTE, 1983). Estes são
fatores que parecem ocorrer coma população de Raillietia que
se reduz bastante no período correspondente ao inverno seco e
frio da microrregião Vale do Rio Doce, Minas Gerais de junho a
agosto que, mesmo não apresentando o frio intenso das regiões
de clima temperado poderia possivelmente pelo tempo prolonga-
do de 3 meses, reduzir a atividade de disseminação das formas
livres do ácaro nas pastagens e com isto também a sua freqüên-
cia mensal.
Em ambas as regiões, apesar das variações climáticas
existentes, a presença do parasito se manifestou durante todo o

24.
ano, sugerindo um intenso potencial de disseminação e fácil a-
daptação entre bovinos hospedeiros confimando observações an-
teriormente registradas (NUNES et alii, 1975; FACCINI et alii,
1976; OLIVEIRA, 1978; FONSECA & FONSECA, 1981; JAVIER et alii,
1983; SHASTRI & DESHPANDE, 1983; McKEE et alii, 1978) e concordan-
do com os registros de LEITE (1989) que encontrou o parasito
se reproduzindo em bezerros, novilhas e vacas naturalmente in-
festadas e com mais ênfase naquelas raças azebuadas e criadas
de forma extensiva.
A ausência de ninfas foi uma constante observação em
todas as colheitas efetuadas nos animais procedentes das duas
microrregiões estudadas e localizadas em áreas distintas do
sistema criatório brasileiro, vindo com isto reforçar outras
observações com relação ao comportamento deste estádio do para-
sito em animais naturalmente infestados (FONSECA, 1983; BRUM
et alii, 1986; FACCINI et alii, 1987).
Os dados registrados nas tabelas 2 e 4 demonstram que
o número de ácaros presentes nas coletas mensais efetuadas nos
bovinos procedentes da microrregião Alto Purús, Acre é sempre
superior ao número de ácaros obtidos nas coletas feitas em bo-
vinos procedentes da microrregião Vale do Rio Doce, Minas Ge-
rais. Este aspecto poderia ser atribuído ao fato de que a re-
giào Amazônica apresenta condições climáticas tipo 1 com a tem-
peratura elevada e sempre favorável ao desenvolvimento dos pa-
rasitos durante todo o ano, enquanto o Sudeste Brasileiro apre-
senta condições climáticas tipo 2 com temperaturas mais redu-
zidas durante os meses de inverno o que limita em parte a evo-

25.
lução dos ácaros (EVANS, 1989). Por outro lado, deve também
ser levado em conta o fato de que os bovinos examinados em A1-
tos Purús, Acre serem exclusivamente da raça Nelore enquanto
que os bovinos do Vale do Rio Doce, Minas Gerais serem azebua-
dos das raças Indubrasil, Guzerá, Nelore e Gir simultaneamen-
te. Estudos anteriores demonstraram que os animais da raça Ne-
lore são os mais susceptíveis ao parasito dentro das raças de
origem zebu (LEITE, 1989).
Observando-se os dados referentes à Análise de Va-
riância da freqüência mensal de fêmeas e larvas de Raillietia
em bovinos da microrregião Alto Purús, Acre (Tabelas 6 e 7; Ta-
belas 14 e 15, apêndice) verifica-se que as médias mensais re-
ferentes ao número de fêmeas foram significativas pelo teste
de Tukey 5%, enquanto que para este teste as medias mensais
referentes ao número de larvas não foram significativas. En-
tretanto, segundo GOMES (1985), nos experimentos inteiramen-
te casualizados como no atual trabalho, onde as observações
são concentradas sobre animais com semelhantes condições (ida-
de de abate), os valores de F obtidos na Análise de Variância
não sendo significativas a 5% podem ser trabalhados em nível
menos exigente de probabilidade, geralmente o de 10% onde te-
rão significância estatística. Assim a Análise de Variância
dos dados da Tabela 15 (apêndice) relativos à freqüência mé-
dia mensal de larvas de Raillietia em Alto Purús, Acre com va-
lores de F de 2,75 para blocos e 3,68 para tratamentos que
não são significativas ao nível de 5% são significativos esta-
tisticamente ao nível de 10% de probabilidade.

26.
TABELA 6. Freqüência média mensal de fêmeas de Raillietia
Trouessart, 1902 em 30 bovinos azebuados da mi-
crorregião Alto Purús, AC, analisada pelo Teste
de Tukey (P < 0,05).
DMS = 742.47.
CV = 9.2319%.
* Tratamentos = meses do ano.

27.
TABELA 7. Freqüência média mensal de larvas de Raillietia
Trouessart, 1902 em 30 bovinos azebuados da mi-
crorregião Alto Purús, Acre analisada pelo tes-
te de Tukey (P < 0,05).
DMS = 849.655.
CV = 15.5276%.
* Tratamentos = meses do ano.

28.
Com relação aos dados referentes à Análise de Variân-
cia da freqüência mensal de fêmeas e larvas de Raillietia em
bovinos da microrregião Vale do Rio Doce, Minas Gerais (Tabe-
las 8 e 9; Tabelas 16 e 17, apêndice) observa-se que as médias
mensais referentes tanto ao número de fêmeas como ao número de
larvas foram significativas pelo teste de Tukey a 5% de proba-
bilidade.
Estas análises demonstram que as freqüências mensais
do parasito em ambas as regiões apresentam tendências defini-
das de acordo com as características climáticas de cada área,
tendências estas que se manifestam através de diferenças es-
tatísticas significantes para a intensidade média de infesta-
ção dos 30 animais coletados mensalmente e procedentes de áreas
criatórias da Amazônia Ocidental e do Sudeste Brasileiro.

29.
TABELA 8. Freqüência média mensal de fêmeas de Raillietia
Trouessart, 1902 em 30 bovinos azebuados da mi-
crorregião Vale do Rio Doce, Minas Gerais, ana-
lisada pelo teste de Tukey (P < 0,05).
DMS = 257.006.
CV = 6.5375%.
* Tratamentos = meses do ano.

30.
TABELA 9. Freqüência média mensal de larvas de Raillietia
Trouessart, 1902 em 30 bovinos azebuados da mi-
crorregião Vale do Rio Doce, Minas Gerais, ana-
lisada pelo teste de Tukey (P < 0,05).
DMS = 138.822.
CV = 5.7788%.
* Tratamentos = meses do ano.

ESPECIFICIDADE PARASITÁRIA
Revisão da Literatura
A maioria dos parasitos se acha distribuida em hospe-
deiros ou grupos de hospedeiros aparentados de forma ecológi-
ca e/ou filogenética, havendo assim, entre hospedeiro e parasi-
to um grau de intimidade denominado especificidade parasitária
que segundo FAIN (1962) pode ser de baixa, média ou de alta
intensidade.
Na especificidade ecológica os requerimentos essen-
ciais para se estabelecer uma alta especificidade parasitária
são: a) o parasito e o hospedeiro devem fazer contatos efeti-
vos um com o outro e isto implica em ação mútua quanto a méto-
dos de dispersão, comportamento, ecologia e habitat; b) o hos-
pedeiro deve prover ao parasito condições adequadas ao seu de-
senvolvimento e c) o parasito deve ser capaz de resistir a qual-
quer resposta orgânica do hospedeiro sobre ele. Por sua vez na
especificidade filogenética, a preferência de um parasito por

32.
um determinado hospedeiro seria determinada pela capacidade do
primeiro, de se adaptar ao longo do tempo à evolução do hospe-
deiro (KENNEDY, 1975).
Segundo FURMANN (1979) existe uma estreita correlação
entre a permanência de um parasito no hospedeiro e sua especi-
ficidade, aqueles que permanecem no hospedeiro durante todos
os estádios do seu ciclo biológico são mais específicos do que
aqueles que usam os hospedeiros de forma ocasional. Assim al-
guns insetos e piolhos são altamente específicos e outros para-
sitos semipermanentes como carrapatos e alguns ácaros mesostig-
matas tem especificidade baixa.
Com relação aos ácaros Mesostigmatas a especificida-
de varia desde a monoxenia até a polixenia. No primeiro grupo
encontram-se espécies parasitas, enquanto que, espécies nidico-
las como representantes da família Laelapidae tendem a des-
viar-se da monoxenia (RADOVSKY, 1985).
Considerando-se especificamente os ácaros do gênero
Raillietia os dados registrados na literatura ainda são escas-
sos e inconclusivos. De um total de cinco espécies descritas e
duas não descritas, quatro são conhecidas como parasitos de um
único hospedeiro. Das três espécies restantes R auris tem co-
mo hospedeiro tipo os bovinos, porém já foi assinalada em ovi-
nos (RAK & NAGHSHINEH, 1973; OLIVEIRA, 1979), caprinos (HAR-
RIS, 1966 citado por NUTTING, 1968) e búfalos (FONSECA, 1983).
Dados obtidos recentemente (LEITE, 1989) no entanto, sugerem
que a espécie parasita de búfalo é diferente de R. auris (FAC-
CINI, 1990 comunicação pessoal) mas pode parasitar bovinos. A

3 3 .
espécie R. caprae tem como hospedeiro tipo os caprinos, porém
já foi assinaladas em ovinos (FONSECA & FACCINI, 1985).
Este trabalho foi proposto para confirmar a importân-
cia dos caprinos e ovinos como hospedeiros das duas espécies
de Raillietia que habitam o conduto auditivo de bovinos no Bra-
sil.
Material e Métodos
O trabalho foi realizado através de duas etapas dis-
tintas. A primeira etapa consistiu de um levantamento atra-
vés da colheita de material em caprinos e ovinos que dividiam
o mesmo espaço físico de pastagem com bovinos.
Um total de 165 caprinos pertencentes as raças Anglo-
nubiana, SRD (Sem Raça Definida) e 302 ovinos pertencentes às
ragas Santa Inês (deslanada), Hampshire (lanada) e SRD (mis-
ta) em propriedades localizadas em diferentes regiões do país
(Alto Purús, Acre; Lages, Santa Catarina; Itaguaí e Magé, Rio
de Janeiro; Bagé, Rio Grande do Sul) foram utilizados durante
este levantamento efetuado no período entre janeiro de 1988 a
julho de 1989 (Figura 7).
A colheita de material foi realizada segundo a técnica
de lavagem dos condutos auditivos externos direito e esquer-
do idealizada por FACCINI et alii (1987) e adaptada para exe-
cução em animais vivos por LEITE (1989).
A segunda etapa consistiu da infestação experimental
de caprinos e ovinos com Raillietia procedente de bovinos, co-

34.
FIGURA 7. Localização geográfica das coletas em caprinos e ovi-
nos criados juntos com bovinos: 1 = Alto Purús, AC;
2 = Itaguaí e Magé, RJ; 3 = Lages, SC; 4 = Bagé, RS.
Janeiro 1988 a julho 1989.

35.
letada em matadouro e realizada em animais pertecentes ao reba-
nho da EMBRAPA - Unidade Nacional de Apoio ao Programa Nacional
de Pesquisa de Saúde Animal em Seropédica, Itaguaí, Rio de Ja-
neiro durante o período entre janeiro a julho de 1989.
Dois grupos de cinco caprinos e cinco ovinos SRD foram
infestados artificialmente, tendo para cada grupo um número cor-
respondente de animais como controle.
Cada animal foi infestado com aproximadamente 15 adul-
tos jovens tidos como a fase infestante do ciclo biológico do
ácaro (LEITE, 1989), no alto da cabeça entre os chifres (1ª in-
festação) e em um dos pavilhões auriculares (1ª infestação). Os
animais foram mantidos em baias e o material coletado para recu-
peração 120 horas após a infestação através de lavagem dos con-
dutos auditivos externos direito e esquerdo pela técnica de
FACCINI et alii (1987) e adaptada para animais vivos por LEITE
(1989).
Os adultos jovens utilizados no experimento foram ob-
tidos de larvas coletadas em bovinos naturalmente infestados e
abatidos para consumo mantidas em camara climatizada em tempera-
tufa de 30 ± l°C e umidade relativa de 90 ± 5% (FONSECA & FACCI-
NI, 1985).
O material obtido nas coletas feitas nos caprinos e o-
vinos, quer em condições naturais quer na infestação experimen-
tal foi acondicionado em frascos de 50 ml de capacidade conten-
do álcool a 70% e transferidos ao laboratório de Acarologia do
Curso de pós-Graduação de Parasitologia Veterinária para iden-
tificação através do microscópio estereoscópico Wild-M5.

3 6 .
Resultados e Discussão
Os exames dos condutos auditivos externos direito e
esquerdo de 165 caprinos e 302 ovinos que dividiam a mesma pas-
tagem com bovinos em propriedades localizadas em diferentes re-
giões do Brasil resultaram negativos em sua totalidade para
Raillietia.
Os dez ovinos infestados artificialmente cada um com
aproximadamente 15 adultos jovens de Raillietia, bem como os
dez animais controles, revelaram-se negativos ao exame de seus
condutos auditivos 120 horas após a infestação.
Dos dez caprinos infestados artificialmente cada um
com aproximadamente 15 adultos jovens, dois animais (20%) apre-
sentaram evolução do ácaro, sendo encontradas uma fêmea adulta
em um animal e duas fêmeas adultas em outro, perfazendo em re-
lação ao total de ácaros inoculados (150) a prevalência de 2%,
enquanto que os dez animais controles foram totalmente negati-
vos.
Os dados obtidos sob as condições do presente traba-
lho indicam que as espécies de Raillietia que parasitam bovi-
nos não são hospedeiros naturais de ovinos e caprinos.
FONSECA & FACCINI (1985) infestaram artificialmente
oito ovinos com R. auris procedentes de bovinos e examinaram
59 ovinos com todos os animais se revelando negativos. Os da-
dos observados no presente estudo confirmam estes resultados e
reforçam a sugestão de que os ovinos não podem set considera-
dos como hospedeiros naturais de Raillietia que tem bovinos co-

37.
mo hospedeiro tipo.
Os achados de R. auris em quantidade reduzida, duas
fêmeas e um macho sem larvas ou ovos que indicassem sinais de
reprodução em ovinos naturalmente infestados, registrados por
OLIVEIRA (1979) e a presença do ácaro em quantidade não regis-
trada (RAK & NAGHSHINEH, 1973) poderiam perfeitamente ser in-
terpretados como contaminação acidental, não implicando de for-
ma necessária na inclusão dos ovinos como hospedeiros natu-
rais do ácaro.
De forma semelhante, a comunicação pessoal de HARRIS
(1966) a NUTTING (1968) sobre a presença de R. auris no condu-
to auditivo de caprinos, sem explicitar quantidade bem como o
reduzido desenvolvimento do ácaro nas infestações experimen-
tais no presente trabalho (3 fêmeas adultas em 150 adultos jo-
vens inoculados) sugerem também, a impossibilidade da inclu-
são destes animais como hospedeiros naturais do ácaro.

CICLO BIOLÓGICO "IN VIVO": CONSIDERAÇÕES PRELIMINARES
Revisão da Literatura
O parasitismo de bovinos por ácaros identificados como
R. auris tem se caracterizado por uma elevada prevalência que
chega a se situar na faixa de 85 a 100% (NUNES et alii, 1975;
OLIVEIRA, 1978; FONSECA, 1983) em pecuária de corte, praticada
principalmente no Brasil em grandes áreas de pastagens e de for-
ma extensiva o que reforça, sobremodo, a capacidade de trans-
missão do parasito entre os rebanhos.
A estrutura de população de R. auris estudada através
de colheitas em animais naturalmente infestados têm sido regis-
trada inicialmente coma presença de ninfas (MENZIES, 1957; NU-
NES et alii, 1975; OLIVEIRA, 1978; QUINTERO, 1978).
Estudos mais recentes e bem mais amplos (FONSECA, 1983;
LEITE, 1989) não confirmaram no entanto a presença dos está-
dios ninfais em infestações naturais, embora, tanto a protonin-
fa como a deutoninfa estejam presentes no ciclo biológico "in

39.
vitro" de R. auris que se completa em torno de 4-5 dias (FONSE-
CA & FACCINI, 1985).
O registro de ninfas em condições naturais por alguns
autores provavelmente deve-se a uma identificação incorreta
da ninfa e do adulto jovem, adulto imediatamente após a muda a
partir do estádio de ninfa e sem se alimentar, que ao microscó-
pio estereoscópico são muito semelhantes.
Dados obtidos até o presente sobre o ciclo biológico
de Raillietia sugerem que a ausência de ninfas em condições na-
turais deve-se a ocorrência deste estádio fora do hospedeiro
(LEITE, 1989).
Este trabalho foi elaborado com o propósito de se am-
pliar os conhecimentos sobre o ciclo biológico "in vivo" de
Raillietia parasitos de bovinos.
Material e Métodos
O trabalho foi desenvolvido através de duas etapas dis-
tintas. A primeira etapa foi conduzida de forma experimental
na área de gado de corte do Instituto de Zootecnia da Universi-
dade Federal Rural do Rio de Janeiro durante o período janeiro
a março de 1990. Foram utilizados 12 bezerros desmamados da ra-
ça Nelore como animais experimentais e 08 bezerros também des-
mamados e da mesma raça como controles, todos pertencentes ao
plantel do Instituto de Zootecnia da UFRRJ.
Cada animal recebeu uma carga de infestação de apro-
ximadamente 20 adultos jovens de Raillietia em cada pavilhão

4 0 .
auricular. Esta fase do ciclo biológico foi escolhida porque
dados registrados na literatura sugerem ser o adulto jovem a
fase mais provável na transmissão do parasito (FONSECA & FAC-
CINI, 1985; LEITE, 1989).
Todos os animais infestados e controles tiveram seus
ouvidos lavados 24 hs antes da infestação e a colheita do ma-
terial inoculado foi realizada também por lavagem dos condu-
tos auditivo, externos direito e esquerdo segundo a técnica
de FACCINI et alii (1987) adaptada para execução em animais
vivos por LEITE (1989).
A colheitado material inoculado foi realizada por in-
tervalos de 12 horas, totalizando 12 coletas e 144 horas pós
infestação. Cada colheita consistiu de lavagem em ambos os ou-
vidos de um animal experimentalmente infestado. Os animais que
serviram de controle tiveram seus ouvidos lavados com interva-
io de 36 hs. A lavagem foi realizada em ambos os ouvidos em
todos os animais controles. Este procedimento permitiu moni-
torar a presenca de infestação acidental nos animais experi-
mentalmente infestados, já que tanto os animais controle como
os infestados permaneceram em um mesmo pasto de 3 ha de área
durante todo o experimento devido ao manejo da propriedade.
Os adultos jovens de Raillietia utilizados no expe-
rimento foram obtidos a partir de larvas colhidas em matadou-
ro de bovinos naturalmente infestados e mantidas em câmara
climatizada à temperatura de 30 ± 1ºC e umidade relativa de
90 ± 5% (FONSECA & FACCINI, 1985).
O material obtido nas colheitas, quer dos animais in-

41.
festados como dos animais controles foi recolhido e acondicio-
nado em frascos de 50 ml de capacidade contendo álcool a 70%
e transferidos ao Laboratório de Acarologia do Curso de Pós-
Graduação de Parasitologia Veterinária para identificação dos
estádios presentes através do uso do microscópio estereoscó-
pico Wild-M5.
A segunda etapa constou do levantamento ao nível de
campo visando a possível recuperação de formas livres do ci-
clo biológico de Raillietia diretamente da pastagem. Foi rea-
lizada na área de Gado de Corte do Instituto de Zootecnia da
UFRRJ também durante o período entre janeiro a março de 1990.
Com a finalidade de tornar possível a colheita do
ácaro na pastagem, foram utilizadas aproximadamente 150 cabe-
ças de bovinos pertencentes à raça Nelore que segundo LEITE
(1989) é altamente susceptível à presença de Raillietia. Es-
tes animais foram postos para pernoitar uma vez por semana du-
rante 10 semanas seguidas em um pasto com 02 ha de área loca-
lizado próximo ao centro de manejo, com pastagem formada à ba-
se de capim-de-burro (Cynodon dactilon) e capim Braquiaria
(Brachiaria decumbens) de tal forma que por volta das 05 ho-
ras da manhã, na hora da colheita, grande parte dos animais ain-
da estavam deitados sobre a pastagem ali existente.
O método de colheita utilizado foi uma adaptação da
técnica de DONALD (1967) para detectar a presença de larvas
de helmintos na pastagem e consistiu no corte das gramíneas a
partir do seu terço inferior após uma segura localização do
local onde os animais estavam deitados, onde teoricamente po-

42.
deria estar ocorrendo deslocamento das formas livres do para-
sito junto ao complexo pastagem-animal-pastagem. O corte era
realizado com o uso de uma faca amolada e as gramíneas colhi-
das eram imediatamente colocadas em um saco plástico que era
imediatamente fechado, levando-se em conta a grande motilida-
de das formas jovens (larvas, ninfas e adultos jovens) do áca-
ro e conduzidas ao Laboratório onde eram postas em baldes con-
tendo água com detergente para liberação dos parasitos coleta-
dos.
Uma vez no Laboratório de Acarologia do Curso de Pós-
Graduação em Parasitologia Veterinária da UFRRJ, o pasto cole-
tado era lavado em água corrente e seu conteúdo posto para re-
pousar em baldes por 02 horas. Em seguida, era tamisado em pe-
neira de malha fina (250 µ) e colocado em recipientes cujos
conteúdos eram submetidos a um novo repouso de 02 horas e a uma
decantação na qual a água em excesso era retirada, permanecen-
do o material coletado no fundo de cada recipiente. Por fim,
adicionava-se álcool 70% para conservação dos ácaros e peque-
nos insetos com eles coletados.
As formas de vida livre de Raillietia assim coleta-
das eram separadas, montadas entre lâmina e lamínula tendo o
meio de Hoyer's como clarificador e identificados através do
microscópio Wild M5.

43.
Resultados e Discussão
Os resultados do ciclo biológico de Raillietia estão
sumarizados na Tabela 10 e na Figura 8. Todos os animais infes-
tados experimentalmente tinham ácaros no interior dos condutos
auditivos.
Da 1ª à 5ª coleta, ou seja, entre o período de 12 a
60 horas após a inoculação (MAI), obteve-se como resultado áca-
ros adultos jovens em processo de alimentação progressiva com
ingurgitamente mas sem assumir em caráter definitivo a forma
de adultos plenos.
Nas 6ª e 7ª coletas (72 e 84 HAI) os ácaros recupera-
dos revelavam as formas plenas de adultos machos e fêmeas sem
apresentarem todavia ovos e ou larvas que indicassem o início
do processo reprodutivo. Estas observações correspondem aos
resultados obtidos por LEITE (1989) que não registraram repro-
dução do ácaro às 72 HAI.
A partir da 8ª e extendendo-se até a 12ª coleta, ou
seja, de 96 a 144 HAI registrou-se a presença de ácaros adul-
tos plenos machos e fêmeas e de ovos (2 vezes em número de 2)
e larvas (um mínimo de 3 e um máximo de 6).
Todos os bovinos usados como controle estavam negati-
vos comprovando que, mesmo mantidos na mesma pastagem com ou-
tros animais infestados, os ácaros coletados nos animais infes-
tados artificialmente originaram-se da própria infestação e
não de contaminação acidental.
Em nenhum dos animais foram encontrados os estádios

44.
TABELA 10. Duração do ciclo biológico, de adulto jovem até lar-
va de Raillietia Trouessart, 1902 "in vivo" em bo-
vinos susceptíveis. Itaguaí, Rio de Janeiro, 1990.
* = Sexagem difícil mesmo ao Microscópio Estereoscópico.
** = Sexagem fácil mesmo à vista desarmada.

45.
FIGURA 8. Estádios do ciclo biológico "in vivo" de Raillietia
Trouessart, 1902 obtidos a partir da infestacão experimental
de bovinos. Adulto jovem parcialmente alimentado (1) ; adul-
to jovem alimentado (2); adulto jovem ingurgitado (3); macho,
fêmea, larva e ovo (4). Escala, 1 mm.
2 1
3 4

46.
de proto e deutoninfa posteriores ao estádio larval. A ausên-
cia de ninfas após 24 horas do aparecimento do estádio larval
quando comparada com a presença da proto e deutoninfa num es-
paço de tempo menor que 24 horas para a ecdise das larvas "in
vitro" (FONSECA & FACCINI, 1985) somada à ausência de ninfas
nas infestações naturais (FONSECA, 1983; LEITE, 1989) corrobo-
ra a hipótese mantida por LEITE (1989) que sugere ser os es-
tádios ninfais de vida livre.
A presença de larvas a partir de 96 HAI observada nes-
te experimento somados com os dados de FONSECA & FACCINI
(1985) que observaram um período entre 4-5 dias para a larva
atingir o estádio de adulto jovem sugerem que o período do ci-
clo biológico "in vivo" é próximo do ciclo "in vitro" (FONSE-
CA & FACCINI, 1985), ou seja, entre 4 e 6 dias.
O ciclo biológico das duas espécies de Raillietia que
parasitam o conduto auditivo de bovinos se processaria da se-
guinte maneira: uma vez que tenha havido a eclosão dos ovos
dentro do conduto auditivo dos hospedeiros, a larva abandona
o hospedeiro apos alimentar-se. No meio ambiente ocorrem as
mudas para protoninfa, desta para deutoninfa e desta, final-
mente, para adulto jovem. Este estádio procura um outro hospe-
deiro para infestar e uma vez dentro do conduto auditivo ocor-
re o acasalamento e em seguida a oviposição (Figura 9).
A partir de 96 HAI (8ª coleta) quando se observou que
além da plena adaptação dos ácaros inoculados nos novos hospe-
deiros, já se iniciou o processo de reprodução, torna-se aumen-
tada a quantidade de cerúmen recuperado nas lavagens dos con-

47.
FIGURA 9. Ciclo biológico de Raillietia Trouessart, 1902. Ten-
tativa diagramática de representação. Ovos, larvas,
adultos machos e fêmeas no hospedeiro. Larvas deixam
o hospedeiro e no ambiente mudam para Protoninfas;
Deutoninfas e Adultos Jovens que infestam outros hos-
pedeiros. Itaguaí, RJ, 1990.

48.
dutos auditivos sugerindo que, mesmo estando sobre os hospedei-
ros em um reduzido período os ácaros já iniciaram os proces-
sos traumáticos nos condutos auditivos que estimulam a maior
produção e consequente liberação do cerúmen comumente observa-
do nas infestações naturais e que poderia ser considerado como
uma reação do organismo contra os traumatismos que ocorrem pe-
la presença dos ácaros.
Com relação à recuperação de formas livres do ciclo
biológico de Raillietia, de um total de 10 coletas, 05 foram
positivas (50%) tendo sido encontradas 03 formas de adulto jo-
vem, 01 forma de ninfa e 03 fêmeas adultas (Tabela 11 e Figura
10).
Quanto à viabilidade da técnica de colheita, observa-
se que, uma vez tenha se identificado com segurança o local
da pastagem onde o gado permaneceu a noite a possibilidade de
resultado positivo é elevada (50%) mesmo se considerando a mo-
tilidade das formas jovens do parasito.
As 03 formas de adulto jovem encontradas em 02 oportu-
nidades (02 na coleta 03 e 01 na coleta 05) e 01 forma de nin-
fa encontrada na coleta 08 eram esperadas, correspondendo as
expectativas e concordando com as hipóteses formuladas por FON-
SECA (1983) e LEITE (1989), de que realmente seriam estas as
formas de vida livre de Raillietia.
A forma de ninfa encontrada que tinha a cor branco
leitosa e trazia consigo vestígios recentes de muda poderia ca-
racterizar a evolução das fases de vida livre no meio ambiente,
enquanto que as formas de adulto jovem recuperadas poderia cor-

49.
TABELA 11. Recuperação de formas de vida livre do ciclo bioló-
gico de Raillietia Trouessart, 1902. Itaguaí, RJ,
1990.

50.
1
2
FIGURA 10. Estádios do ciclo biológico de Raillietia Troues-
sart, 1902 recuperados das pastagens. Adulto jovem (1); Nin-
fa (2). Escala, 1 mm.

51.
responder ao momento em que os ácaros circulantes na pastagem
se direcionam aos novos hospedeiros atraídos possivelmente por
quimiotropismo, tanto pelo odor (ARLIAN, 1989) como pelo Dióxi-
do de Carbono (FALCO & FISH, 1989) exalados de seus corpos.
Não foram encontradas, todavia, larvas na pastagem o
que corresponderia ao período que estas abandonariam seus hos-
pedeiros para a continuidade do ciclo biológico como forma de
vida livre no ambiente.
Por outro lado as 03 fêmeas adultas recuperadas em
duas oportunidades (02 na coleta 04 e 01 na coleta 07) poderiam
ser interpretadas como um comportamento exótico do parasito a-
bandonando seu nicho natural que é o conduto auditivo exter-
no.
A superlotação de parasitos poderia fazer com que al-
gumas fêmeas se desloquem para o pavilhão auricular e daí para
o exterior. O número excessivo de ácaros poderia ocorrer em
vacas Nelores utilizadas no trabalho, animais tidos como sen-
síveis à infestação por Raillietia (LEITE, 1989).
Por outro lado, a postura do ato de dormir dos bovi-
nos com o corpo deitado sobre os membros que são simetricamen-
te recolhidos sob ele (GURTLER et alii, 1987) tendo geralmente
como ponto de apoio a cabeça em contato com o solo através da
mandíbula, apoio mentoniano ou mandibular (DUKES, 1962), pode-
ria sugerir uma posição de declive que criaria as condições de
gravidade necessárias ao despreendimento e queda dos ácaros que
eventualmente venham circular pelo pavilhão auricular.

C O N C L U S Õ E S
Os resultados obtidos neste estudo permitem concluir
que a infestação por Raillietia em bovinos é mantida por uma
fase de vida livre no ambiente com distribuição e frequência
influenciadas pelas condições climáticas de cada região.
Em Altos Purús, Acre, Raillietia apresenta dois pi-
ques de infestações por ano nos meses de maio e novembro, o
que corresponde ao fim do período chuvoso e início do período
seco, e fim do período seco e início do período chuvoso, res-
pectivamente.
No Vale do Rio Doce, Minas Gerais, Raillietia apre-
senta maior carga de infestação no período de novembro a mar-
ço, correspondente ao verão quente e chuvoso.
O parasitismo por Raillietia em bovinos azebuados a-
tinge 100%, tanto na região do Alto Purús, Acre, onde variou
de 03 a 221 ácaros por animal, como no Vale do Rio Doce, Mi-
nas Gerais, onde a variação foi de 19 a 144 ácaros por ani-

53.
mal.
As espécies de Raillietia parasitas de bovinos apresen-
tam estreita e elevada especificidade parasitária.
Ovinos e caprinos não se comportaram como hospedeiros
naturais das espécies de Raillietia que tem bovinos como hos-
pedeiros tipos.
Raillietia é um parasito que apresenta em seu ciclo
biológico fases de vida livre que circulam na pastagem.
O ciclo biológico de Raillietia a partir da fase de
adulto jovem em animais susceptíveis ocorre entre 04 e 06 dias.
A hipótese provável sobre o ciclo biológico de Raillie-
tia é que os ovos eclodem no conduto auditivo do hospedeiro,
as larvas abandonam o hospedeiro, mudando no ambiente para pro-
toninfas, deutoninfas e adultos jovens que, por sua vez, pro-
curam e infestam outros hospedeiros, evoluem para adultos ple-
nose daí para o acasalamento e oviposição.

REFERÊNCIAS BIBLIOGRÁFICAS
ALWAR, V.S.; LALITHA, C.M. & ACUTHAN, H.N. RailIietia auris
(Leidy, 1872), Trouessart, 1902 in the ear of cattle (Bos
indicus) in Madras. Indian Vet. J., 36:276-80, 1959.
ANTUNES, F.Z. Caracterização climática do Estado de Minas Ge-
rais. Inf. Agropec., 12(138):9-13, 1986.
ANUÁRIO ESTATÍSTICO DO ACRE. Rio Branco, Acre, v. 20, 1982.
357 p.
ARLIAN, L.G.; VYSZENSKI-MOHER, D.L. & CORDOVA, D. Hos Speci-
ficity of Sarcoptes scabiei var. canis (Acari: Sarcopti-
dae) and the role of host odor. J. Med. Entomol., 25:52-56,
1988.
ARRUDA, M.L.R. Estabelecimento e recuperação de pastagens no
Vale do Rio Doce. Inf. Agropec., 13(153) :23-25, 1987.

5 5 .
BASTOS, T.X. 0 clima da Amazônia Brasileira segundo Köppen. Be-
lém, EMBRAPA-CPATU, 1982. 4 f. (EMBRAPA, CPATU. Pesquisa em
Andamento, 87).
BRUM, J.G.W.; RIBEIRO, P.B. & FACCINI, J.L. Raillietia auris
(Acari: Gamasida) em bovinos do Rio Grande do Sul. In: CON-
GRESSO BRASILEIRO DE MEDICINA VETERINÁRIA, 20, Cuiabá, 1986.
Cuiabá, 1986. p. 55 (Resumo).
COSTA, A.L. da. Bioecologia de Boophilus microplus (Canestrini,
1887) (Acarina: Ixodidae) no Estado do Rio de Janeiro: ovipo-
sição e sazonalidade. Considerações preliminares. Seropédi-
ca, RJ, Universidade Federal Rural do Rio de Janeiro, 1982.
37 p. (Tese, Mestrado).
COSTA, A.L. da. Incidância mensal do carrapato dos bovinos no
Estado do Acre. In: SIMPÓSIO DO TRÓPICO ÚMIDO, 1, Belém, PA,
1984. Anais. Belém, EMBRAPA-CPATU, 1984. p. 366 (Resumo).
DONALD, A.D. A technique for the recovery os Strongyloid infec-
tive larvae from small units of pasture. J. Helmintol., 12
(1):2-10, 1967.
DONROW, R. New records and species of Austromalayan Laelapid mi-
tes. Proc. Linn. Soc. New South Wales, 88:199-220, 1963.

56.
DORESTE, E.S. Los ácaros como plagas y sus reaciones ecológicas.
In: Acarologia. San Jose, Instituto Interamericano de Coope-
racion para la Agricultura, 1983. p. 323-39 (Série Investiga-
cion y Desarrollo, 11).
DUKES, H.H. Fisiologia de los Animales Domesticos. 2 ed. Madrid,
Aguilar, 1962. 462 p.
EVANS, D.E. Epidemiologia do carrapato dos bovinos, Boophilus mi-
croplus no Brasil. In: CURSO DE PARASITOLOGIA ANIMAL, 2. Ba-
gé, RS, 1988. Anais ... Bagé, Colégio Brasileiro de Parasito-
logia Veterinária, 1989. p. 72-88.
FACCINI, J.L.H.; CONFALONIERI, U.E.C.; MASSARD, C.L. & SERRA
FREIRE, N.M. Situação do parasitismo por Raillietia auris
(Leidy, 1872) e referência do encontro de Raillietia sp. em
caprinos no Brasil. In: CONGRESSO BRASILEIRO DE MEDICINA VE-
TERINÁRIA, 15, Rio de Janeiro, 1976. Anais ... Rio de Janei-
ro, 1976. p. 149-50 (Resumo).
FACCINI, J.L.H.; LIGNON, G.B. & LEITE, R.C. Evaluation of an ear
flushing technique as a post-mortem measure of infestation of
Raillietia auris (Leidy) (Acari) in cattle. Exp. Appl. Aca-
rol., 3:175-9, 1987.

57.
FAIN, A. Les acariens Mesostigmatiques ectoparasites des ser-
pents. Bullo Inst. Roy. Nat. Belgique, 33(25):1-49, 1962.
FALCO, R.C. & FISH, D. The use of Carbon Dioxide-Baited tick
traps for sampling Ixodes dammini (Acari: Ixodidae). Aca-
rologia, 30(1):29-33, 1989.
FERGUSON, W. & LAVOIPIERRE, M.M.J. The occurrence of Raillie-
tia auris in zebu cattle in Nigeria. Vet. Rec., 74:678,
1962.
FONSECA, A.H. Biologia e ecologia de Raillietia auris (Leidy,
1872) (Acari, Mesostigmata). Seropédica, RJ, Universidade
Federal Rural do Rio de Janeiro, 1983. 54 p. (Tese, Douto-
rado).
FONSECA, A.H. & FACCINI, J.L.H. In vitro development of Rai-
llietia auris (Leidy) (Acarina: Mesostigmata). Acarologia,
26:211-14, 1985.
FONSECA, A.H.; FACCINI, J.L.H. & MASSARD, C.L. Raillietia ca-
prae (Acari: Mesostigmata) em caprinos e ovinos no Brasil.
Pesq. Vet. Bras., 3(1):29-31, 1983.

58.
FONSECA, A.H. & FONSECA, A.H. Prevalência de Raillietia auris
(Leidy, 1872) em bubalinos e bovinos na região amazônica.
In: CONGRESSO BRASILEIRO DE PARASITOLOGIA, 6, Belo Hori-
zonte, 1981. Anais .. Belo Horizonte, 1981. p. 203 (Resu-
mo) .
FREUND, L. Zur kenntnis der ohrmilbe des Remides. Zool. Jahrb.
Abt. Syst. Geogr. Biol. Tiere, 29:313-32, 1910.
FURMANN, D.P. Specificity, adaptation and paralled evolution
in the endoparasitic mesostigmata of mammals. In: RODRI-
GUES, J.C. Recent Advances in Acarology. 2 ed. New York,
Academic Press, 1979. p. 329-37.
GOMES, F.P. A Estatística moderna na pesquisa agropecuária.
Piracicaba. Associação Brasileira para Pesquisa da Potassa
e do Fosfato, 1985. 159 p.
GURTLER, H.; KEITZ, A.; KOLB, E.; SCHRODER, L. & SEIDEL, H.
Fisiologia Veterinária. 4 ed. Rio de Janeiro, Guanabara Koo-
gan, 1987. p. 504-33.
HEFFNER, R.S. & HEFFNER, H.E. Effect of cattle ear mite infes-
tation on hering in a cow. J. Am. Vet. Med. Assoc., 182:
612-14, 1983.

59.
HEFFNER, R.S. & HEFFNER, H.E. Occurrence of the cattle ear mite
(Raillietia auris) in South Eastern Kansas. Cornell Vet., 73:
193-99, 1983.
JAVIER, F.R.A.; QUINTERO, M.T.M. & ACEVEDO, A.H. Frequência de
Raillietia auris en bovinos sacrificados en la Empacadora y
Frigorifico de la Ciudad de Villa Hermosa, Tambasco. Vet.
Mex., 14:204-5, 1983.
KENNEDY, C.R. Ecological animal parasitology. Oxford. Blackwell,
1975. 163 p.
LADDS, P.W.; COPEMAN, D.B.; DANIELS, P. & TRUEMAN, K.F. Raillie-
tia auris and otitis média in Northern Queensland. Aust.
Vet. J., 48:532-33, 1972.
LARANGEIRA, N.L.; LIMA, M.M.; PAIVA, F. & RIBEIRO, H.S. Preva-
lência de Raillietia sp. (Leidy, 1792) Trouessart, 1902 (Aca-
ri: Mesostigmata) em bovinos de Mato Grosso do Sul. In: CON-
GRESSO BRASILEIRO DE PARASITOLOGIA, 7, Porto Alegre, 1982.
Anais ... Porto Alegre, 1982. p. 104 (Resumo).
LEIDY, J. On a mite in the ear of the ox. Proc. Acad. Nat. Sci.
Philadelphia, 24:872-3, 1872.
LEITE, R.C. Raillietiose bovina. Seropédica, RJ, Universidade Fe-
deral Rural do Rio de Janeiro, 1989. 24 p. (Tese, Doutorado).

60.
LEITE, R.C.; NUNES, V.A.; COELHO, A.M.B.; CHIQUILOFF, M.A.G. &
NUNES, I.J. Patologia da infecção do ouvido de bovinos por
Raillietia auris (Leidy, 1872) Trouessart, 1902 (Acari - Me-
sostigmata). II. Achados bacteriologicos. Arq. Bras. Med.
Vet. Zoot., 39:325-32, 1987.
MAIA, A.A.M. & GUIMARÃES, M.P. Distribuição sazonal de larvas
da Dermatobia hominis (Linnaeus Jr., 1871) (Diptera: Cutere-
bridae) em bovinos de corte da Região de Governador Valada-
res - Minas Gerais. Arq. Bras. Med. Vet. Zoot., 37(5): 469-
75, 1985.
McKEE, T.J.; COUVILLION, C.E. & RANDOLPH, T.C. Ear mites in ca-
ttle: More prevalent than previously thought. Vet. Med., 83:
731-32, 1988.
MENZIES, G.C. The cattle ear mite, Raillietia auris (Leidy,
1872) in Texas. J. Parasitol., 43:200, 1957.
NETO, S.S.; NAKANO, 0.; BARBIN, D. & VILLA NOVA, N.A. Manual
de Ecologia dos Insetos. São Paulo, Editora Agronômica Ce-
res, 1976. 419 p.
NUNES, V.A. Patologia da infecção do ouvido de bovinos por Rai-
llietia auris (Leidy, 1872) Trouessart, 1902 (Acari - Mesos-
tigmata). Belo Horizonte, Escola de Veterinária da UFMG,
1977. 52 p. (Tese, Mestrado).

61.
NUNES, I.J.; MARTINS Jr., W.; LEITE, R.C.; NUNES, V.A. & RIBEI-
RAL, L.A. Da presença de Raillietia sp. (Mesostigmata - Rai-
llietidae) no conduto auditivo e na membrana timpânica de bo-
vinos da região de Brasília - DF. In: CONGRESSO BRASILEIRO
DE MEDICINA VETERINÁRIA, 13, Brasília, 1972. Anais ... Brasí-
lia, Sociedade de Medicina Veterinária de Brasília. 1972. p.
252 (Nota Prévia).
NUNES, I.J.; MARTINS Jr., W.; NUNES, V.A. & LEITE, R.C. Ocorrên-
cia de Raillietia auris (Leidy, 1872) Trouessart, 1902 em bo-
vinos da região geo-econômica de Brasília, DF. Arq. Esc. Vet.F. Arq. Esc. Vet.
UFMG, 27(2):375-83, 1975.
NUTTING, W.B. Host specificity in parasitic acarines. Acarolo-
gia, 10(2):165-180, 1968.
OLIVEIRA, G.P. Estudo do desenvolvimento de ovos e larvas de
Boophilus microplus (Canestrini, 1887) (Acarina: Ixodidae)
em condições de imersão e de ambiente. Seropédica, RJ, Uni-
versidade Federal Rural do Rio de Janeiro, 1976, 69 p. (Te-
se, Mestrado).
OLIVEIRA, G.P. Raillietia auris (Leidy, 1872) Trouessart, 1902
(Acari: Mesostigmata) em bovinos no Estado de São Paulo. Arqs.
Esc. Vet. UFMG, Belo Horizonte, 30(3):307-310, 1978.

62.
OLIVEIRA, G.P. Ocorrência de Raillietia auris (Leidy, 1872)
Trouessart, 1902 em Ovis aries no Brasil. Científica, 7:149-
51, 1979.
OLSEN, O.W. & BRACKEN, F.K. Occurrence of ear mite, Raillietia
auris (Leidy, 1872) of cattle in Colorado. Vet. Med., 45:320-
21, 1950.
QUINTERO, M.T.M. Importância de Raillietia auris en bovinos. In:
CURSO DE ACTUALIZACION. ENFERMIDADES PARASITARIAS DEL GANADO
BOVINO, México, DF, 1978. México, DF, Faculdad de Medicina
Veterinaria y Zootecnia, UNAM, 1978. 13 p.
QUINTERO, M.T.C.; BASSOLS, I.B. & ACEVEDO, A.H. Hallazgo y des-
cription de Raillietia caprae sp. n. (Acari: Mesostigmata)
en caprinos de Sinaloa, Mexico. Vet. Med., 11(2):17-20, 1980.
RADFORD, C.D. Notes on some new species of parasitic mites. Pa-
rasitology, 30:427-40, 1938.
RADOVSKY, F.J. Evolution of mammalian mesostigmate mites. In:
CHUNG, K. ed. Coevolution of Parasitic Arthropods and ani-
mals. New York, 1985, p. 441-504.
RAK, H. & NAGHSHINEH, R. First report and redescription of Rai-
llietia auris (Trouessart, 1902) (Acari: Gamasidae) in Iran.
Entomol. Mon. Mag., 109:59, 1973.

6 3 .
ROUX, J.P.; MANCEBO, D.A. & BULMAN, G.M. Primera descripcion
en la Republica Argentina de Raillietia auris del conduto
auditivo externo del bovino. In: JORNADAS INTERNACIONALES.
FACULTAD CIENCIAS VETERINARIAS. UNIVERSIDADE NACIONAL LA
PLATA. 6, Formosa, 1983, p. 30.
SCHLOTTHAUER, J.C. Cattle en mite (Raillietia auris) in Minne-
sota. J. Am. Vet. Med. Assoc,, 157:1193-94, 1970.
SHASTRI, U.V. & DESHPANDE, P.D. Raillietia auris (Leidy,
1872) Trouessart, 1902 in Marathwada cattle (Maharashtra)
India. Trop. Vet. Anim. Sci. Res., 1:91-6, 1983.
TROUESSART, M.E. Existence de laparthenogenese chezle Gamasus
auris (Leidy), de l'oreille du boeuf domestique. C.R. Hebd.
Seances Mem. Soc. Biol., 54:806-8, 1902.
TROUESSART, M.E. Note sur le Gamasus auris (Leidy, 1872). Ty-
pe d'un genre nouveau (Raillietia). Bull. Soc. Zool. Ft.,
27:231-33, 1902.
TROUESSART, M.E. Sur le Gamasus auris, type d'un genre nou-
veau (Raillielia). C.R. Seances Soc. Biol. Fil., 54:1335-
37, 1902.
TSYMBAL, T.G. & LITVISHKO, N.T. [Otocariasis in cattle]. Zool.
Zh., 34:1229-41, 1955 (In Russian).

64.
VALENTIM, J.F. & COSTA, A.L. da. Efeito da pressão de pastejo
na persistência de pastagens cultivadas no Acre. In: SEMI-
NÁRIO AGROPECUÁRIO DO ACRE, 1, Rio Branco, 1983. Anais ...
Rio Branco, 1983, p. 383-94.
WILKINSON, P.R. Observations on infestation of undipped cattle
of british breeds with the cattle tick, Boophilus microplus
(Canestrini). Aust J. Agric. Res., 6(4):655-665, 1955.


66.
TABELA 12. Número de Raillietia Trouessart, 1902 em 30 bovinos
azebuados naturalmente infestados. Alto Purús, Acre,
1988.

67.
TABELA 13. Número de Raillietia Trouessart, 1902 em 30 bovinos
azebuados naturalmente infestados. Vale do Rio Do-
ce, Minas Gerais, 1989.
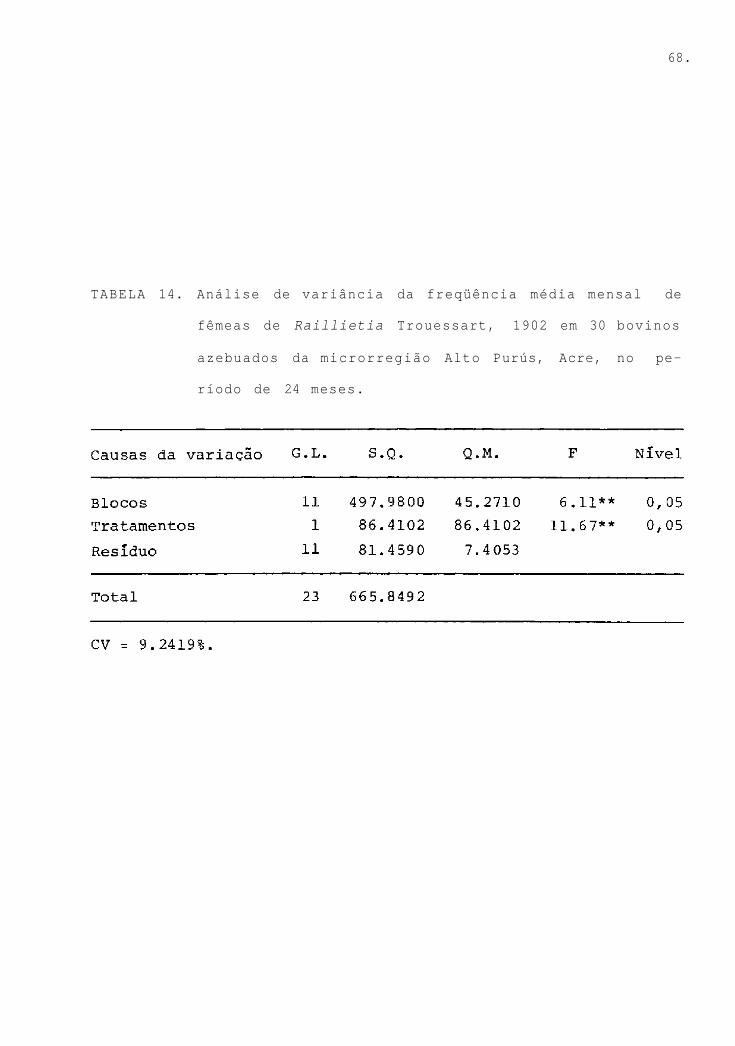
68.
TABELA 14. Análise de variância da freqüência média mensal de
fêmeas de Raillietia Trouessart, 1902 em 30 bovinos
azebuados da microrregião Alto Purús, Acre, no pe-
ríodo de 24 meses.

69.
TABELA 15. Análise de variância da freqüência média mensal de
larvas de Raillietia Trouessart, 1902 em 30 bovinos
azebuados da microrregião Alto Purús, Acre, no perío-
do de 24 meses.

70.
TABELA 16. Análise de variância da freqüência média mensal de
fêmeas de Raillietia Trouessart, 1902 em 30 bovinos
azebuados da microrregião Vale do Rio Doce, Minas
Gerais, no período de 24 meses.

71.
TABELA 17. Análise de variância da freqüência média mensal de
larvas de Raillietia Trouessart, 1902 em 30 bovinos
azebuados da microrregião Vale do Rio Doce, Minas Ge-
rais, no período de 24 meses.
CV = 5.7788%.

72.
TABELA 18. Dados climáticos da microrregião Alto Purús, Acre,
1987-1988. Fonte: Delegacia Federal da Agricultu-
ra, Acre.
Ano 1 = Jan-Dez. 1987.
Ano 2 = Jan-Dez. 1988.

73.
TABELA 19. Dados climáticos da microrregião Vale do Rio Doce,
Minas Gerais. Novembro 1987 - Outubro 1989. Fonte:
5º Distrito Meteorológico, Governador Valadares, Mi-
nas Gerais.
Ano 1 = Nov. 87-Out. 88.
Ano 2 = Nov. 88-Out. 89.







