
Tierärztliche Hochschule Hannover
Einfluss von Haltezeiten während der Frühphase der
Konservierung auf den Energiemetabolismus und
die Qualität von Eberspermien
INAUGURAL – DISSERTATION
zur Erlangung des Grades einer
Doktorin der Veterinärmedizin
- Doctor medicinae veterinariae –
( Dr. med. vet. )
vorgelegt von
Ulrike Wallner
Heidenheim a. d. Brenz
Hannover 2015

Wissenschaftliche Betreuung: Apl. Prof. Dr. med. vet. Dagmar Waberski
Reproduktionsmedizinische Einheit der Kliniken/
Klinik für kleine Klauentiere und forensische
Medizin und Ambulatorik
Tierärztliche Hochschule Hannover
1. Gutachterin: Apl. Prof. Dr. med. vet. Dagmar Waberski
2. Gutachterin: Prof. Dr. Martina Hoedemaker, PhD
Tag der mündlichen Prüfung: 02.11.2015
Eine Arbeit aus dem Virtuellen Zentrum für Reproduktionsmedizin Niedersachsen

Meiner Familie
und Thomas

Auszüge aus der vorliegenden Arbeit wurden bereits der Öffentlichkeit in Form einer
Veröffentlichung und zwei Vorträgen vorgestellt:
M. Schulze, H. Henning, K. Rüdiger, U. Wallner, D. Waberski (2013); Temperature management during semen processing: Impact on boar sperm quality under laboratory and field condition Theriogenology, 80, 990-998
„Effects of holding times during boar semen processing on energy metabolism
and bacterial growth“
U. Wallner, H. Henning, Q. T. Nguyen, D. Waberski
(47. Jahrestagung Physiologie und Pathologie der Fortpflanzung, Gießen,
27.02.2014)
„Revision of current protocols for boar semen processing:Retarded cooling of
diluted semen enhances sperm quality“
U. Wallner, S. Kastens, A. Riesenbeck, H. Henning, D. Waberski (48th Annual Conference on Physiology and Pathology of Reproduction, Zürich, 11.02.2015)

Inhaltsverzeichnis
1 Einleitung ..................................................................................................................... 1
2 Literaturübersicht ........................................................................................................ 2
2.1 Kälteschock bei Eberspermatozoen ........................................................................ 2
2.1.1 Einfluss von Kälteschock auf Plasmamembranen ............................................ 2
2.1.2 Einfluss von Haltezeiten auf die Kälteschocksensibilität ................................... 4
2.2 Samenverarbeitungsverfahren in der Praxis ............................................................ 6
2.3 Energiemetabolismus bei Spermatozoen ................................................................ 7
2.3.1 ATP-Produktion und Energieladung ................................................................. 7
2.3.2 Einfluss der Lagerung auf den Energiemetabolismus ......................................11
3 Material und Methoden ...............................................................................................16
3.1 Versuchsübersicht ..................................................................................................16
3.2 Verbrauchsmaterialien und Geräte .........................................................................17
3.3 Lösungen und Chemikalien ....................................................................................18
3.4 Tiere .......................................................................................................................18
3.5 Samengewinnung ..................................................................................................19
3.6 Untersuchung des nativen Samens ........................................................................19
3.7 Monitoring der Probentemperatur ...........................................................................20
3.8 Experimentelle Designs ..........................................................................................21
3.8.1 Experiment 1: Vergleich zweistufig isothermer und zweistufig hypothermer
Verdünnung bei zwei Verdünnungsgraden ......................................................21
3.8.2 Experiment 2: Auswirkungen von Haltezeiten und Haltetemperaturen des
vorverdünnten Samens bei zweistufig isothermer Verdünnung .......................24
3.8.3 Experiment 3: Vergleich zweistufig isothermer mit zweistufig hyperthermer
Verdünnung ....................................................................................................27
3.8.4 Experiment 4: Auswirkungen von Haltezeiten des ausverdünnten Samens nach
zweistufig hypothermer (Exp. 4 a) oder einstufig isothermer Verdünnung
(Exp. 4 b) ........................................................................................................29
3.8.5 Experiment 5: Auswirkungen einer beschleunigten Abkühlrate des
ausverdünnten Samens nach einstufig isothermer Verdünnung ......................31
3.8.6 Experiment 6: Temperaturverlauf in der Prozesskette der Samenverarbeitung
auf einer Besamungsstation und Vergleich mit Laborstandards ......................33
3.8.6.1 Temperaturverlauf im Sperma während der Verarbeitung auf einer
Besamungsstation bei Samenverarbeitung ..............................................33

3.8.6.2 Vergleich des Verdünnungsverfahrens einer Besamungsstation mit dem
Laborstandard ..........................................................................................35
3.9 Computerassistierte Spermienanalyse (CASA) ......................................................37
3.9.1 Testprinzip ......................................................................................................37
3.9.2 Gerät und Software .........................................................................................38
3.9.3 Messung .........................................................................................................39
3.10 Membranintegrität in der Durchflusszytometrie .......................................................40
3.10.1 Testprinzip ......................................................................................................40
3.10.2 Gerät und Farbstoffe .......................................................................................41
3.10.3 Grundeinstellungen .........................................................................................41
3.10.4 Messung und Auswertung ...............................................................................43
3.11 Mitochondrienmembranpotential in der Durchflusszytometrie .................................44
3.11.1 Testprinzip ......................................................................................................44
3.11.2 Gerät und Farbstoffe .......................................................................................44
3.11.3 Grundeinstellungen .........................................................................................45
3.11.4 Messung und Auswertung ...............................................................................46
3.12 Kinetik des Calcium-Influx ......................................................................................47
3.12.1 Testprinzip ......................................................................................................47
3.12.2 Gerät und Farbstoffe .......................................................................................47
3.12.3 Vorbereitung der Medien und Spermien ..........................................................48
3.12.3.1 Messung und Auswertung ........................................................................49
3.13 Bestimmung des ATP-Gehalts und der Energieladung ...........................................51
3.13.1 Testprinzip ......................................................................................................51
3.13.2 Geräte und Vorbereitung .................................................................................52
3.13.3 Messung und Auswertung ...............................................................................53
3.13.3.1 Präparation der Spermien für die Lagerung bei -20 °C .............................54
3.13.3.2 Erstellen der Standardkurve für die ATP-Messung ...................................54
3.13.3.3 Extraktion von ATP aus der gefrorenen Probe .........................................55
3.13.3.4 Messung der Lumineszenz und Berechnung der ATP-Konzentrationen ...55
3.13.3.5 Konvertieren der Nukleotide und deren Bestimmung ...............................56
3.13.3.6 Berechnung der Energieladung ................................................................57
3.14 Bestimmung der Gesamtkeimzahl ..........................................................................57
3.15 Statistische Auswertung .........................................................................................57

4 Ergebnisse ..................................................................................................................59
4.1 Experiment 1: Vergleich von zweistufig isothermer und zweistufig hypothermer
Verdünnung bei zwei Verdünnungsgraden ................................................................59
4.1.1 Temperaturverlauf während der Samenverarbeitung.......................................59
4.1.2 CASA-Motilitätsparameter ...............................................................................61
4.1.3 Membranintegrität ...........................................................................................63
4.2 Experiment 2: Auswirkungen von Haltezeiten und -temperaturen des vorverdünnten
Samens bei zweistufig isothermer Verdünnung .........................................................65
4.2.1 Temperaturverlauf während der Samenverarbeitung.......................................65
4.2.2 CASA-Motilitätsparameter ...............................................................................67
4.2.3 Membranintegrität ...........................................................................................70
4.2.4 Mitochondrienmembranpotential .....................................................................71
4.2.5 Energiestoffwechsel ........................................................................................73
4.2.6 Gesamtkeimzahlbestimmung ..........................................................................75
4.3 Experiment 3: Vergleich zweistufig isothermer mit zweistufig hyperthermer
Verdünnung ..............................................................................................................79
4.3.1 Temperaturverlauf während der Samenverarbeitung.......................................79
4.3.2 Casa-Motilitätsparameter ................................................................................81
4.3.3 Membranintegrität ...........................................................................................83
4.4 Experiment 4 a: Auswirkungen von Haltezeiten des ausverdünnten Samens nach
zweistufig hypothermer Verdünnung .........................................................................84
4.4.1 Temperaturverlauf während der Samenverarbeitung.......................................84
4.4.2 Casa-Motilitätsparameter ................................................................................86
4.4.3 Membranintegriät ............................................................................................87
4.5 Experiment 4 b: Auswirkungen von Haltezeiten des ausverdünnten Samens nach
einstufig isothermer Verdünnung...............................................................................88
4.5.1 Temperaturverlauf während der Samenverarbeitung.......................................88
4.5.2 CASA-Motilitätsparameter ...............................................................................90
4.5.3 Membranintegrität ...........................................................................................91
4.6 Experiment 5: Auswirkungen einer beschleunigten Abkühlung des ausverdünnten
Samens nach einstufig isothermer Verdünnung ........................................................92
4.6.1 Temperaturverlauf während der Samenverarbeitung.......................................92
4.6.2 Casa-Motilitätsparameter ................................................................................94
4.6.3 Membranintegrität ...........................................................................................95

4.7 Experiment 6: Temperaturverlauf in der Prozesskette der Samenverarbeitung auf
einer Besamungsstation und Vergleich mit Laborstandards ......................................96
4.7.1 Temperaturverlauf während der Samenverarbeitung.......................................96
4.7.2 CASA-Motilitätsparameter ...............................................................................98
4.7.3 Membranintegrität ...........................................................................................99
4.7.4 Spezifische Reaktivität gegenüber Bikarbonat (Calcium-Influx) ..................... 100
5 Diskussion ................................................................................................................ 101
6 Zusammenfassung ................................................................................................... 111
7 Summary ................................................................................................................... 113
8 Literaturverzeichnis .................................................................................................. 115
9 Anhang ...................................................................................................................... 122
9.1 Tabellen und Grafiken .......................................................................................... 122
9.2 Chemikalien ......................................................................................................... 146
9.3 Rezepte ................................................................................................................ 147
9.3.1 Formolcitrat ................................................................................................... 147
9.3.2 HBS (HEPES®- buffered saline) .................................................................... 147
9.3.3 Tyrode basierte Medien ................................................................................. 148
9.3.3.1 Herstellung der Stammlösungen ............................................................ 148
9.3.3.2 Herstellung der Tyr BikCa-Vormischung .................................................... 148
9.3.3.3 Herstellung der TyrCa-Vormischung ........................................................ 149
9.3.3.4 Komplettierung der Medien .................................................................... 149
9.4 Geräte und Verbrauchsmaterialien ....................................................................... 150
10 Abbildungsverzeichnis ............................................................................................. 153
11 Tabellenverzeichnis .................................................................................................. 159
Danksagung ...................................................................................................................... 161
Erklärung ............................................................................ Fehler! Textmarke nicht definiert.

Abkürzungsverzeichnis
°C Grad Celsius
µg Mikrogramm
µl Mikroliter
Abb. Abbildung
ADP Adenosindiphosphat
AK Adenylatkinase
ALH gemittelte maximale seitliche Kopfauslenkung von der mittleren Bahn
(amplitude of lateral head displacement)
AMP Adenosinmonophosphat
ATP Adenosintriphosphat
BCF Frequenz des Kreuzens des gewundenen Bewegungspfades mit der
gemittelten Bewegungsbahn (beat cross frequency)
BSA bovines Serumalbumin
BTS Beltsville Thawing Solution
bzw. beziehungsweise
Ca Calzium
ca. circa
CASA computer-assistierte Spermienanalyse
cm Zentimeter
D Tag (die)
DAP Länge der gemittelten Bahn (distance average path)
DCL Länge der gewundenen Bahn (distance curved line)
DNS Desoxiribonuleinsäure (desoxy ribonucleic acid)
DSL Länge der direkten Strecke (distance straight line)
EC Energieladung (energy charge)
EDTA Ethylendiamintetraacetat
et al. et alii
Exp. Experiment
Fa. Firma
FITC Fluoreszeinisothiocyanat

FL 1-4 Filter Nummer 1 – 4
FSC Vorwärtsstreulicht (forward scatter)
g Erdbeschleunigung (gravitational acceleration)
G Gate
h Stunde (hora)
HBS Hepes-gepufferte Salzlösung (hepes buffered saline)
Hz Hertz
Jagg Agglutinierte Form des Farbstoffs JC-1
JC-1 5,5',6,6'-Tetrachloro-1,1',3,3'-tetraethylbenzimidazolylcarbo-cyanine iodide
l Liter
LIN Linearität (linearity)
M Mol
MAS Morphologisch abweichende Spermien
mg Milligramm
min Minute
Mio Millionen
ml Milliliter
mM millimol
MMP Mitochondrienmembranpotential
Mrd Milliarden
ms Millisekunde
MW Mittelwert
n Probandenzahl (numerus)
neg. negativ
nm nanometer
p Irrtumswahrscheinlichkeit (probability)
PEP Phosphoenolpyruvat
PI Propidiumiodid
PK Pyruvatkinase
pmol picomol
PNA peanut agglutinin, Lektin der Erdnuss (arachis hypogea)

pos. positiv
PPi Pyrophosphat
prog. progressiv
PVA Polyvinylalkohol
PVP Polyvinylpyrrolidon
R Reaktivität
RLU relative light units
RN Range
RT Raumtemperatur
s Sekunde
s. siehe
SD Standardabweichung (standard deviation)
Sp. Spermien
SSC Seitwärtsstreulicht (sideward scatter)
STR straightness
Tab. Tabelle
VAP Geschwindigkeit der gemittelten Bahn (velocity average path)
VCL Geschwindigkeit der gewundenen Bahn (velocity curved line)
VSL Geschwindigkeit der direkten Strecke (velocity straight line)
WOB wobble
z.B. zum Beispiel
ZDS Zentralverband der deutschen Schweineproduktion e.V.


Einleitung
1
1 Einleitung
Da Eberspermien sehr kältesensibel sind, ist die Konservierung noch immer mit
Schwierigkeiten verbunden. Die Verarbeitung von Ebersperma zu
flüssigkonservierten Samenportionen hat in der Schweinezucht eine hohe Relevanz.
Inzwischen werden über 90% der Sauen künstlich besamt; mehr als 95 % der
Besamungen werden mit flüssig konserviertem Samen durchgeführt, der bei + 17 °C
gelagert wird (JOHNSON et al. 2000, RIESENBECK 2011). Eine hygienische und
temperaturoptimierte Verarbeitung des Spermas gilt als maßgeblich für die Qualität
des bis zu fünf Tagen gelagerten Spermas.
Es ist seit langem bekannt, dass Haltezeiten des vorverdünnten Samens bei höheren
Temperaturen (20 – 32 °C) während der Frühphase der Verarbeitung positive
Auswirkungen auf die Kältetoleranz der Zellen bei nachfolgender Abkühlung auf die
Lagerungstemperatur haben (PURSEL et al. 1973 b, CASAS et al. 2013). Bei
welcher Temperatur und innerhalb welcher Zeitspanne die bestmögliche Adaption
der Zellen stattfindet, und ob Haltezeiten bei höheren Temperaturen möglicherweise
negative Effekte auf Energiestoffwechsel und Bakterienwachstum in den
Spermaportionen haben, ist noch weitestgehend unbekannt. Zudem sind Haltezeiten
bis zu sechs Stunden während der Prozesskette der Spermaverarbeitung häufig
unvermeidlich, ohne dass deren Auswirkung auf Spermaqualität und mikrobiellen
Status bekannt ist.
Ziel der vorliegenden Studie war es daher, den Einfluss von Haltezeiten zwischen
30 min und 6 h bei 21 °C sowie 32 °C während der Frühphase der Konservierung auf
die Qualität gelagerten Eberspermas unter Anwendung gängiger Praxisverfahren der
Spermaverdünnung zu untersuchen. Dabei wurden neben spermatologischen
Parametern, wie der Motilität und Membranintegrität, mögliche Einflüsse auf den
Energiestoffwechsel und das bakterielle Wachstum betrachtet. Zusätzlich sollten die
praxisüblichen Verarbeitungsmethoden überprüft und Temperaturprofile während der
Verarbeitungskette erhoben werden.

Literaturübersicht
2
2 Literaturübersicht
2.1 Kälteschock bei Eberspermatozoen
2.1.1 Einfluss von Kälteschock auf Plasmamembranen
Eberspermien gelten im Gegensatz zu anderen Säugetierspermien als besonders
sensibel gegenüber Abkühlung. Dabei spielt die Struktur der Plasmamembran der
Samenzellen eine große Rolle. Wie alle biologischen Membranen besteht diese aus
Lipiden (hauptsächlich Phospholipide), die sich in einer Doppelschicht anordnen, und
Proteinen. Membranen sind fluide Strukturen, was bedeutet, dass sich Moleküle
innerhalb der Ebene der Doppelschicht bewegen können. Sie sind asymmetrisch
aufgebaut, das heißt die Lipid- und Proteinzusammensetzung der beiden Schichten
ist unterschiedlich (DE LEEUW et al., 1991). Die Fähigkeit von Spermien, niedrige
Temperaturen zu tolerieren, scheint vor allem von der Lipidzusammensetzung der
Membran abzuhängen, welche tierartlich sehr verschieden ist (DE LEEUW et al.,
1990; WATERHOUSE et al., 2006).
Ein relativ geringer Cholesteringehalt in der Plasmamembran unterscheidet die
Samenzellen von temperaturempfindlichen Spezies wie z.B. dem Eber von denen
temperaturresistenter Spezies (z.B. Mensch, Kaninchen). Der Cholesteringehalt von
Eberspermien beträgt ca. 0,034 µmol/ 109 Zellen, Bullenspermien enthalten ca.
0,893 µmol/ 109 Zellen, während Spermien von Menschen und Kaninchen einen
Cholesteringehalt von ca. 1,4 µmol/ 109 Zellen haben (DARIN-BENNETT u. WHITE,
1977; NIKOLOPOULOU et al., 1985). Beim Eber ergibt sich daraus ein, im Vergleich
zu anderen Arten, niedriges molares Cholesterin/ Phospholipid-Verhältnis von 0,12,
welches die Temperaturempfindlichkeit der Zellen verstärkt. Beim resistenteren
Bullen beträgt das Verhältnis 0,45, beim Menschen sogar 0,99 (DARIN-BENNETT u.
WHITE 1977, NIKOLOPOULOU et al. 1985; DE LEEUW et al., 1990). PARKS und
LYNCH (1992) berichten von einem Verhältnis von 0,26 beim Eber.
Die Kombination aus niedrigem Gehalt an Cholesterin und dessen asymmetrischen
Verteilung in der Membran - es befindet sich mehr in der äußeren Schicht als in der
inneren - macht vor allem die innere Schicht sehr anfällig. PARKS und LYNCH
(1992) fanden außerdem eine Beziehung zwischen der Kältesensitivität und dem

Literaturübersicht
3
Proteingehalt der Plasmamembran. Eberspermien zeigten einen höheren Gehalt an
Membranproteinen als Spermien anderer Säugetierspezies.
Zudem besteht ein ungünstigeres Verhältnis von ungesättigten zu gesättigten
Fettsäuren, welches bei Eberspermien > 2,5 im Vergleich zu ungefähr 1 bei
humanen Spermien beträgt. Der relativ hohe Anteil an ungesättigten Fettsäuren
macht die Samenzellen anfällig für Schäden durch Peroxidation, welche sich negativ
auf Motilität, Metabolismus, Zellstruktur und Fertilität auswirken (WHITE, 1993). Ein
hoher Anteil an ungesättigten Fettsäuren zusammen mit dem niedrigen
Cholesteringehalt verursacht zusätzlich eine höhere Fluidität der Spermienmembran,
und dadurch eine erhöhte Kälteschocksensibilität (LADBROOKE u. CHAPMAN,
1969; WATERHOUSE et al., 2006).
Kälteschock führt zu Schäden in der Membran der Samenzellen. Einige
Veränderungen der Membran sind dabei reversibel, während andere fatal für die
Zelle sind (GUTHRIE u. WELCH, 2005). Das offensichtlichste Zeichen eines
ausgeprägten Kälteschocks ist ein irreversibler Motilitätsverlust. Dieser wird begleitet
von erhöhter Permeabilität der Plasmamembran, welche zum Verlust von Kationen,
ATP und Enzymen führt (PARKS u. LYNCH, 1992; DROBNIS et al., 1993).
Kälteschock bei Eberspermien entsteht, wenn frisch ejakulierte Samenzellen
entweder schnell, mit einer Abkühlrate von 1 - 2 °C/ min oder schneller, abgekühlt
werden (JOHNSON et al., 2000) und/ oder unter 15 °C gekühlt werden (DE LEEUW
et al., 1991). Bei Temperaturen von < 15 °C treten bei einer Subpopulation von
Spermien bereits Motilitätsverluste, verringerte Membranintegrität sowie eine erhöhte
Permeabilität der Membran auf (DROBNIS et al., 1993; JOHNSON et al., 2000). Der
Zusammenhang von Kälteschockverhalten und der Lipidzusammensetzung der
Spermienmembran lässt darauf schließen, dass die Schädigung mit einem
Phasenübergang (Phasentransition) der Membranlipide zusammenhängt (DROBNIS
et al., 1993, reviewed in WATSON, 1996). Die Membranlipide wechseln bei der
Abkühlung vom flüssigen in einen gelförmigen Zustand. Die verschiedenen
Lipidgruppen innerhalb der Membran erreichen diesen Zustand der Phasentransition
bei unterschiedlichen Temperaturen. Wenn dabei feste und flüssige Bereiche direkt
nebeneinander auftreten, kann es zur Phasenseparation kommen, wobei

Literaturübersicht
4
Membranproteine irreversibel agglutinieren und dadurch ihre Funktion verlieren (DE
LEEUW et al., 1990; DE LEEUW et. al., 1991). Diese Defekte sind geringer, wenn
sich die Membran entweder in der flüssigen oder in der Gelphase befindet. Die
Permeabilität erhöht sich aber stark durch die Reorganisation während der
Phasentransition (DROBNIS et al., 1993). Da die Transition für jede
Phospholipidgruppe bei einer anderen Temperatur stattfindet, kann nur ein
Temperaturbereich für die Reorganisation der Membran angegeben werden. Bei
Ebersamenzellen liegt der Bereich des Phasenübergangs bei 30 – 10 °C (DROBNIS
et al., 1993; SCHMID et al., 2013).
In praktischer Hinsicht ist es wichtig, dass die Kälteschocksensibilität weniger von der
Lagerungsdauer abhängig ist, sondern vielmehr von der Zieltemperatur und davon,
in welcher Geschwindigkeit diese erreicht wird (JOHNSON et al., 2000; SCHMID et
al. 2014).
2.1.2 Einfluss von Haltezeiten auf die Kälteschocksensibilität
Das Halten bzw. die Inkubation von verdünntem und frischem unverdünnten
Eberspermien bei 24 – 26 °C (PURSEL et al., 1973 b) sowie bei 30 °C (PURSEL et
al., 1972) für bis zu 7,5 Stunden vor der Endverdünnung führt zu einer graduellen
Kälteschockresistenz für die weitere Abkühlung und Lagerung bei 5 °C. Die Autoren
zeigten, dass vor allem die Spermien, die 4,5 h und 6 h gehalten wurden, gegenüber
den direkt abgekühlten Proben bessere Motilität und Membranintegrität aufwiesen.
CASAS et al. (2013) zeigten, dass Proben, die einer Haltezeit von 24 h bei 17 °C
ausgesetzt waren, bessere Motilität und höhere Membranintegrität bei der Abkühlung
und während der 24-stündigen Lagerung bei 5 °C aufwiesen, als Proben, die ohne
Haltezeit abgekühlt wurden. Dagegen beschrieben GUTHRIE und WELCH (2005),
dass Haltezeiten von 24 h bei 15 °C von verdünntem Ebersamen vor dem Einfrieren
der Proben zu verringerten Wurfgrößen führte.
Vor allem in der Kryokonservierung der Eberspermien werden mittlerweile die
positiven Effekte einer Haltezeit bei Temperaturen über 15 °C beobachtet und
genutzt. In den meisten Kryoprotokollen werden Haltezeiten von mindestens zwei bis
vier Stunden nach der Verdünnung und vor der weiteren Abkühlung integriert, um die

Literaturübersicht
5
Schäden des Einfrierens und Auftauens zu minimieren (WATSON, 1996; ERIKSSON
et al., 2001). Die Zeitspanne hat dabei größeren Einfluss als die Haltetemperatur.
Verlängerte Haltezeiten bei 17 °C von bis zu 10 h oder 20 h führen zu einer höheren
Membranintegrität nach dem Auftauen (ERIKSSON et al., 2001). TAMULI und
WATSON (1994) berichteten von erhöhter Kälteresistenz von Eberspermien nach
Haltezeiten bis zu 16 h bei Raumtemperatur.
Die genauen Mechanismen, durch welche die Haltezeiten zu einer besseren
Kältetoleranz der Membran führen, werden noch diskutiert. Vermutlich wird durch die
Haltezeiten den Samenzellen die Möglichkeit zur Reorganisation der Membranlipide
und somit zur Adaption an niedrige Temperaturbereiche gegeben. Während der
Zeitspanne binden Komponenten aus dem Seminalplasma bzw. Verdünner an der
Zellmembran und tragen zur Stabilisierung bei (PURSEL et. al., 1973 a; CASAS u.
ALTHOUSE, 2013; ERIKSSON et al., 2001). Substanzen, wie beispielsweise
Phosphatidylserin, schützen dabei die Plasmamembran von Eberspermien
gegenüber Kälteeinflüssen (BUTLER u. ROBERTS, 1975; NIKOLOPOULOU et al.,
1985).
Im Gegensatz dazu stellen PETRUNKINA et al. (2005) die Verarbeitung von
Spermien bei höheren Temperaturen in Frage. Spermien, die mit 32 °C warmen
Verdünner verdünnt wurden, zeigten eine erhöhte Destabilisierung der Membran im
Gegensatz zu mit 21 °C verdünnten Proben. Die Autoren nahmen an, dass die
Spermien, die bei Temperaturen nahe der Körpertemperatur verarbeitet werden,
nicht, wie bei niedrigen Temperaturen, ihren Stoffwechsel verringern. Dies könnte
beispielsweise in Form von Membrandestabilisierungen zu Beeinträchtigungen der
Spermienqualität führen, die die Kapazitation beeinflussen. Die genauen Vorgänge,
wie bzw. ob sich die Haltezeiten, vor allem nahe der Körpertemperatur,
beispielsweise auf Vorgänge wie den Energiestoffwechsel auswirken, sind nach
Kenntnis der Autorin nicht beschrieben.
Nicht nur die Abkühlung, sondern auch der Verdünnungsgrad des Ejakulats während
der Haltezeit beeinflusst die Kältesensibilität der Eberspermien. Proben, die 1:2
vorverdünnt einer Haltezeit für 1 h, 3 h oder 5 h bei 30 °C ausgesetzt und
anschließend ausverdünnt wurden, zeigten höhere Motilität nach dem Kälteschock

Literaturübersicht
6
(5 °C), als die Proben, die vor der Haltezeit bereits ausverdünnt wurden (PURSEL et
al., 1973 a).
2.2 Samenverarbeitungsverfahren in der Praxis
Grundsätzlich werden derzeit in der Praxis zwei unterschiedliche Methoden für die
Verdünnung von flüssigkonservierten Eberspermien durchgeführt: Die einstufige und
die zweistufige Verdünnung. Bei der einstufigen Verdünnung werden die Spermien
isotherm, d.h. mit einem auf 32 °C (± 1 °C) temperierten Verdünner innerhalb von
30 min nach der Samengewinnung in einem Schritt auf die Endkonzentration
ausverdünnt. Bei der zweistufigen Verdünnung wird das Ejakulat zunächst mit auf
32 °C (± 1 °C) angewärmten Verdünner im Verhältnis 1:1 vorverdünnt, und in einem
zweiten Schritt entweder mit ebenfalls 32 °C (± 1 °C) warmen Verdünner (zweistufig
isotherm) oder mit einem hypothermen, auf 21 °C (± 1 °C) temperierten Verdünner,
ausverdünnt (zweistufig hypotherm) (WABERSKI, 2009). Vor allem bei der zweistufig
hypothermen Verdünnung werden die Proben häufig vor der zweiten Verdünnung
einer Haltezeit von bis zu 20 min bei Raumtemperatur ausgesetzt. Die
Vorverdünnung ist ein kritischer Schritt in der Samenverarbeitung und sollte nicht in
einem niedrigeren Verhältnis als 1:1 durchgeführt werden, da sich sonst der
stabilisierende Effekt auf die Samenzellen verringert (PURSEL et al., 1972; STÄHR
et al., 2009; WABERSKI, 2009).
Die in der Praxis am häufigsten durchgeführte Verdünnung ist das zweistufig
hypotherme Verfahren (LÓPEZ RODRÍGUEZ et a., 2012). Diese ist für den Ablauf
der Samenverarbeitung in Besamungsstationen praktikabler, da an
produktionsintensiven Tagen oder bei Transport von vorverdünnten Ejakulaten aus
entfernten Eberställen in zentrale Labore zur Weiterverarbeitung ohnehin Haltezeiten
bei Raumtemperatur auftreten, deren Dauer variabel ist. Aus wirtschaftlicher Sicht
spielt außerdem eine Rolle, dass bei hypothermer Ausverdünnung weniger Energie
für das Erwärmen des Verdünners benötigt wird. Ein weiterer Grund für die
hypotherme Verdünnung ist das Ziel, die Samenproben schnellstmöglich auf die
Lagerungstemperatur abzukühlen, da so ein zeitnaher Versand bei konstanten

Literaturübersicht
7
Temperaturen möglich ist (LÓPEZ RODRÍGUEZ et al., 2012). Weiterhin könnte
durch beschleunigtes Abkühlen mögliches Bakterienwachstum verringert werden.
Festzustellen ist, dass Haltezeiten derzeit in der Praxis wenig standardisiert sind und
keine gesicherten Informationen über das Abkühlverhalten (Temperaturprofile) in den
verdünnten Samenproben während der Anwendung der unterschiedlichen
Verdünnungsregime vorliegen.
Ob die Spermienqualität durch die zweistufig hypotherme Verdünnung und den damit
verbundenen schnellen Temperaturabfall der Samenproben beeinträchtigt wird, wird
noch diskutiert. Bei einem direkten Vergleich von zweistufig isothermer Verdünnung,
mit zweistufig hypothermer Verdünnung, (Verdünnertemperatur 20 – 23 °C, nach
isothermer Vorverdünnung im Verhältnis 1:1), konnte kein Einfluss des
Verdünnungsregimes auf CASA-Motilitätsparameter festgestellt werden
(PETRUNKINA et al., 2005; LÓPEZ RODRÍGUEZ et al., 2012). PETRUNKINA et al.
(2005) konnten zusätzlich keine negativen Effekte der zweistufig hypothermen
Verdünnung auf die Membranintegrität, sowie keine Veränderung der
Reaktionsfähigkeit auf kapazitierende Stimuli zeigen.
In einem parallel zur vorliegenden Studie erfolgten Praxisversuch von mehreren
europäischen Besamungsstationen (n = 23) wurden Verdünnungsmethoden und
Spermaqualität untersucht. 30 % der Besamungsstationen verarbeiteten den Samen
einstufig isotherm, bei 70 % erfolgte die Verdünnung zweistufig hypotherm
(24 ± 3 °C). Hier konnte ein Einfluss des Verdünnungsregimes ermittelt werden. Die
zweistufig hypotherm verdünnten Proben wiesen eine niedrigere Membranintegrität
und niedrigere Aktivität der Mitochondrien im Vergleich zu den einstufig verarbeiteten
Proben. Die Autoren führen dies auf den schnellen Temperaturabfall in den für
Eberspermien kritischen Temperaturbereich zurück (SCHULZE et al., 2013).
2.3 Energiemetabolismus bei Spermatozoen
2.3.1 ATP-Produktion und Energieladung
Samenzellen von Säugetieren sind hochspezialisierte und funktionelle Zellen, deren
Hauptaufgabe es ist, das paternale Genom zu der Eizelle zu bringen. Um diese

Literaturübersicht
8
Aufgabe zu erfüllen, ist zunächst die Entwicklung und Reifung der Spermien im
Hoden notwendig. Während dieser Zeit sind die Spermien in der Regel immotil. Ein
wichtiger Schritt während der Passage durch den Nebenhoden ist der Erwerb der
progressiven Motilität, die in erster Linie energieabhängig ist. Diese wird aber
gleichzeitig durch das Milieu der viskösen epididymalen Flüssigkeit gehemmt. In
dieser verbleiben die Samenzellen bis zur Ejakulation (MISRO u. RAMYA, 2012).
Eine kontinuierliche Energiebereitstellung für energieverbrauchende Prozesse wie
Motilität, Biosynthese, aktiver Ionentransport, Kapazitation und Akrosomenreaktion
wird wie bei allen tierischen Zellen durch die Synthese von ATP ermöglicht. Der
größte Teil der bereitstehenden Energie wird allerdings für die Motilität aufgewendet,
die durch das Schlagen der langen Geißel erzeugt wird (BOHNENSACK u.
HALANGK, 1986). Die Geißel macht ca. 90 % der Länge des Spermiums bei
Säugetieren aus. Energie in Form von ATP gilt als Notwendigkeit, damit die effiziente
Geißelbewegung zur Vorwärtsbeweglichkeit führen kann. Veränderte
Bewegungsmuster, aufgebrauchte Energieressourcen oder beides führen dazu,
dass die Zellen die Fähigkeit der Vorwärtsbeweglichkeit verlieren und damit auch ihre
Befruchtungsfähigkeit. Es ist daher wichtig, den Energiestatus der Spermien zu
kennen, um deren Potential für Motilität und Überleben einzuschätzen.
Zum Verständnis des Energiestoffwechsels muss der Aufbau eines Spermiums bzw.
der Geißel betrachtet werden. Die Geißel untergliedert sich in drei Teile; dem
Mittelstück, dem Hauptstück (ca. ¾ der Gesamtlänge) sowie dem Endstück. Sie
besteht aus einem Axonem, welches aus neun Mikrotubuli-Paaren gebildet wird, die
sich um ein zentrales Mikrotubulus-Paar gruppieren. Das Axonem erstreckt sich über
die gesamte Länge des Flagellums (FAWCETT, 1975). Mitochondrien sind nur im
Mittelstück der Geißel zu finden, wo sie das Axonem umhüllen. Hier kann unter
aeroben Bedingungen Energiestoffwechsel stattfinden (TRAVIS et al., 1998).
In der fibrinösen Hülle der Geißel, die für die Stabilität und für die optimale
Krümmung zuständig ist, wurden vor allem im Hauptstück Enzyme der Glykolyse
(z.B. Hexokinase und Phosphofruktokinase) nachgewiesen.
Die benötigte Energie in Form von intrazellulärem Adenosintriphosphat (ATP) kann
durch zwei verschiedene Stoffwechselwege generiert werden: Durch die Glykolyse

Literaturübersicht
9
und die oxidative Phosphorylierung, wobei die oxidative Phosphorylierung eine
größere Energieausbeute vorweist. Bei der Glykolyse wird Glukose zu Pyruvat
umgewandelt, wobei nur ein kleiner Teil des energetischen Potentials von Glukose
freigesetzt wird: 2 Mol ATP pro Mol Glukose. Wenn Pyruvat in den Mitochondrien
durch die oxidative Phosphorylierung weiter oxidiert wird, werden pro Mol Glukose
30 Mol ATP freigesetzt (FORD, 2006). Welche Art des Energiestoffwechsels in
überwiegendem Maße in Spermien stattfindet, und welche Substrate in welchem
Verhältnis dafür herangezogen werden, ist nach bisherigen Erkenntnissen abhängig
von der Tierart (FORD, 2006; MISRO u. RAMYA, 2012).
ATP, welches in den Mitochondrien erzeugt wird, müsste auch bis in die distalen
Segmente des Flagellums transportiert werden, um den Energiebedarf des Axonems
zu decken. Die Distanz von Bildungs- zu Verbrauchsstätte erscheint allerdings zu
groß, um ATP auf ganzer Länge der Geißel zur Verfügung zu stellen (TURNER,
2003). Die Mitochondrien von Eberspermien enthalten nur wenige innere Cristae, an
welchen wichtige Schritte der ATP-Synthese stattfinden, im Gegensatz zu
beispielsweise Mitochondrien von Leberzellen. Deshalb wird angenommen, dass
diese wenig effizient und damit für die Hauptenergieversorgung nicht geeignet sind
(RODRIGUEZ-GIL, 2013).
Es wird von einigen Forschern angenommen, dass die Glykolyse bei Eberspermien
den wichtigsten Weg für die Energiegewinnung darstellt (MARIN et al., 2003; MISRO
u. RAMYA, 2012). Obwohl die Ausbeute bei der Glykolyse wesentlich geringer ist
als bei der oxidativen Phosphorylierung zeigten MARIN et al. (2003), dass nur 5 %
der Energie von Eberspermien durch letztere erzeugt werden. Eine Erklärung dafür
ist, dass sich die Samenzellen nach der Ejakulation im weiblichen Genitaltrakt unter
hauptsächlich anaeroben Bedingungen fortbewegen müssen. Unter diesen
Umständen ist die Glykolyse der effizientere Stoffwechselweg (RODRIGUEZ-GIL,
2013). Studien mit Mäusen zeigten, dass die Glykolyse unverzichtbar für die
Funktionen der Spermien ist, aber die oxidative Phosphorylierung für die männliche
Fruchtbarkeit nicht essentiell ist (MISRO u. RAMYA, 2012). Vor allem scheint die
Glykolyse für die Kapazitation und speziell für die Hyperaktivierung wesentlich zu
sein. Dies legt nahe, dass die Glykolyse den primären Stoffwechselweg darstellt und

Literaturübersicht
10
oxidative Phosphorylierung eine Reservefunktion erfüllt. Eine weitere Theorie ist,
dass sich die Spermien ihrer Umwelt anpassen, also je nach verfügbarem Substrat
den entsprechenden Syntheseweg wählen. Beispielsweise würde bei Mangel an
Glukose und Fruktose mehr Laktat und Pyruvat in den Mitochondrien verarbeitet
werden. Außerdem scheinen verschiedene Vorgänge in der Zelle unterschiedliche
Energiequellen zu nutzen. So erfordert die Akrosomenreaktion bei Eberspermien
wahrscheinlich eher Laktat oder Pyruvat für die ATP-Produktion in den
Mitochondrien, während Energie für die Motilität wie oben erwähnt hauptsächlich
über die Glykolyse gewonnen wird (MIKI, 2017; RODRIGUEZ-GIL, 2013).
FORD (2006) dagegen berichtet, dass der Austausch von ATP, Adenosindiphosphat
(ADP) und Phosphat (Pi) zwischen dem Flagellum und den Mitochondrien mit Hilfe
der Adenylatkinase, die als Shuttle fungiert, durchaus möglich ist und dadurch die
Aufrechterhaltung der Motilität über den mitochondrialen Stoffwechsel gewährleistet
werden kann. Außerdem zeigen seine Experimente, dass Spermien von Mäusen, bei
denen die Glykolyse mit dem GAPDH-Inhibitor α-Chlorohydrin geblockt wurde,
dennoch für lange Zeit motil waren. Weiterhin argumentiert der Autor, dass die
Spermien der meisten Spezies auch in zuckerfreien Medien vollständig motil bleiben.
Wie oben erwähnt, ist Energie in Form von ATP wichtig, um die für die Zellen
lebenswichtigen Synthesereaktionen durchzuführen. Das Maß der Energieladung
(„Energy charge“, EC) ermöglicht eine Einschätzung der Wachstumsphasen, der
Vitalität der Zellen und einen Vergleich von Zellpopulationen (FORD u. LEACH,
1998). Die Menge von metabolisch verfügbarer Energie, die vorübergehend im
Adenylat-System gespeichert ist, hängt linear mit dem Stoffmengenanteil von ATP
plus der Hälfte des Stoffmengenanteils von ADP zusammen (CHAPMAN et al.,
1971). Dieser Parameter, definiert als Energieladung, und die Formel zur
Berechnung wurden von ATKINSON (1967; 1968) wie folgt beschrieben:
Energy charge (EC) =[ATP] + 0,5[ADP]
[ATP] + [ADP] + [AMP]

Literaturübersicht
11
Die Grundvoraussetzung ist, dass Zellen/ Organismen versuchen, ein bestimmtes
Verhältnis von ATP zu ADP und Adenosinmonophosphat (AMP) aufrechtzuerhalten,
und dieses von den Wachstumsphasen der Zellen abhängt (WIEBE u. BANCROFT,
1975). CHAPMAN et al. (1971) zeigten, dass eine große Auswahl von Organismen,
sowohl Eukarioten als auch Prokarioten, während derselben Wachstumsphase
dieselbe Energieladung haben. Aktiv wachsende und sich teilende Zellen haben ein
EC-Verhältnis von 0,8 - 0,95; Zellen, die sich in einer stationären Wachstumsphase
befinden, zeigen ein Verhältnis von 0,6 und ruhende bzw. alternde Zellen haben ein
Verhältnis von unter 0,5 (WIEBE u. BANCROFT, 1975).
CHULAVATNATOL et al. (1977) analysierten den Nukleotid-Gehalt von humanen
Samenzellen mittels Biolumineszenz-Assay und zeigten, dass die physiologische
Energieladung von humanen Samenzellen zwischen 0,8 und 0,9 liegt. Damit
bestätigten sie ATKINSONS Aussage, dass jede normale, homöostatische Zelle eine
physiologische Energieladung zwischen 0,8 und 0,95 hat (ATKINSON, 1968). Mittels
Erhöhung des pH-Werts auf über 9 verursachten CHULAVATNATOL et al. (1977)
einen drastischen Abfall der Motilität und des Nukleotid-Gehalts. Dennoch blieb die
Energieladung stabil bei ca. 0,8. Bei einer pH-Wert-Erniedrigung auf unter 8 sank die
Motilität ebenfalls drastisch ab, der Nukleotid-Gehalt und die Energieladung sanken
aber nur geringgradig. Auch längere Inkubation bei 30 °C des Spermas verursachte
einen Rückgang der Motilität und des Nukleotid-Gehalts, die Energieladung
allerdings blieb bei über 0,6. Die Autoren schlossen aus den Ergebnissen, dass die
Energieladung nicht direkt mit dem ATP-Gehalt gekoppelt ist und dass sie in
humanen Samenzellen auch unter Stress stabil ist.
2.3.2 Einfluss der Lagerung auf den Energiemetabolismus
Mit zunehmender Lagerungsdauer nimmt die Befruchtungsfähigkeit der
Eberspermien ab. Dies wird als Folge eines natürlichen, unvermeidbaren
Alterungsprozesses angesehen. Neben der sinkenden Befruchtungsfähigkeit finden
während der Lagerung auch strukturelle und funktionelle Veränderungen statt. Die
Motilität wird hierbei häufig als Maßstab für einen intakten Metabolismus
herangezogen. Für die Motilität wird ein Grenzwert von 60 – 70 % für die

Literaturübersicht
12
Verwendung in der künstlichen Besamung gesetzt (JOHNSON et al., 2000). Mit
zunehmender Lagerungsdauer sinkt die Motilität und der Grenzwert wird im mit BTS
verdünnten Samen teilweise sogar schon nach 48 h (KUMARESAN et al., 2009) oder
nach 96 h (CEROLINI et al., 2000; WAERHOUSE et al., 2004) Lagerungsdauer
unterschritten. Andererseits werden in einigen Studien auch nach einer Lagerung
von über 100 h noch mehr als 60 % motile Spermien gefunden (WABERSKI et al.,
1994, HENNING et al., 2012).
Die Abnahme der Motilität während der Lagerungsdauer wird unter anderem mit der
zunehmenden Produktion von reaktiven Sauerstoffspezies („reactive oxygen
species“, ROS), oder von der sich anschließenden Lipidperoxidation in Verbindung
gebracht (CEROLINI et al., 2000; GUTHRIE et al., 2008; KUMARESAN et al., 2009).
Eberspermien gelten wegen ihres hohen Gehalts an mehrfach ungesättigten
Fettsäuren als sehr anfällig, da diese als Substrat für die ROS dienen. Diese wirken
sich nicht nur auf die Motilität sondern vor allem auf den Energiestoffwechsel der
Zellen aus. Der Angriff von freien Radikalen auf die ungesättigten Fettsäuren der
Samenzellmembranen führt zu irreversibler Reduktion der Membranfluidität und zur
Schädigung membranverbundener ATPasen, die für die Regulation der
intrazellulären Ionenkonzentration verantwortlich sind, um die normale
Spermienbeweglichkeit aufrechtzuerhalten. Es wurde gezeigt, dass niedrige
Konzentrationen dieser Substanzen zahlreiche zelluläre Enzyme und metabolische
Prozesse wie die Glykolyse hemmen, und dadurch die ATP-Produktion der Zelle
limitieren (GOGOL et al. 2009).
Die Mitochondrien gelten als Hauptbildungsstätte der intrazellulären ROS. Ihr
vermehrtes Auftreten führt zu einer Unterbrechung des Elektronentransports in den
Mitochondrien. Da der Elektronentransport für die oxidative Phosphorylierung
essentiell ist, führt eine Störung zur Reduktion des Mitochondrienmembranpotentials
und damit zu einer verringerten mitochondrialen ATP-Produktion und in Folge dessen
zu verringerter Motilität (CRAMER u. KNAFF, 1990; GUTHRIE et al., 2008).
ARMSTRONG et al. (1999) zeigten, dass ROS zu Membranschäden führen; dadurch
wird der ATP-Gehalt reduziert und die Motilität der Samenzellen verringert sich. Sie
wiesen nach, dass H2O2 die für humane Samenzellen aggressivste ROS ist. Der

Literaturübersicht
13
Zusatz von H2O2 (0,3 mM – 1,5 mM) in der Samenprobe führte zu einer Reduktion
der Motilität und dosisabhängig zur Verringerung des ATP-Gehalts der Spermien.
Zudem konnten die Autoren bei hohen Konzentrationen von H2O2 einen Effekt auf
das Mitochondrienmembranpotential nachweisen. Allerdings zeigten GUTRIE et al.
(2008) bei Eberspermien, dass der Zusatz von 300 µM H2O2 eine abrupte Reduktion
der Motilität verursachte, aber keinerlei Veränderungen im ATP-Gehalt der Zelle
festzustellen waren. Die Autoren vermuten, dass das H2O2 nicht zu einer
Unterbrechung der Funktion der Mitochondrien geführt hat, sondern stattdessen die
Nutzung von ATP im Axonem unterbrochen oder die Kontraktionsfähigkeit gestört
wurde.
GOGOL et al. (2008) ermittelten mit Hilfe der Chemilumineszenzmessung den
oxidativen Schaden von Eberspermien während der Lagerung und die Auswirkungen
auf den ATP-Gehalt. In vorherigen Studien konnten die Autoren bei humanen
Spermien zeigen, dass die induzierte Photonenemmission eng mit der
Lipidperoxidation der Samenzellen zusammenhängt und somit als Indikator für
oxidativen Stress dienen kann. Nach sieben Tagen Lagerung der mit BTS
verdünnten Samenproben bei 15 °C erhöhten sich die Werte der Chemilumineszenz
signifikant, parallel dazu sanken die Motilität und der ATP-Gehalt. Die Motilität sank
innerhalb der ersten vier Tage um 8 %, der ATP-Gehalt um 25 %. Nach sieben
Tagen Lagerung war die Motilität um 11 % gesunken, der ATP-Gehalt um 33 %. Die
Autoren vermuten, dass die erniedrigte Motilität die Folge der herabgesetzten ATP-
Produktion ist. LONG und GUTRHRIE (2006) analysierten den ATP-Gehalt mit Hilfe
eines Biolumineszenz Assays und zeigten, dass keine signifikanten Veränderungen
bei in mit BTS konservierten Ebersamenproben während der Lagerung über fünf
Tage bei 17 °C auftraten. DZIEKONSKA et al. (2009) testeten den ATP-Gehalt von
Ebersamenproben, die mit dem Verdünner Kortowo 3 (K3) verdünnt wurden, über
eine Lagerungsdauer von drei Tagen bei 16 °C. Dem Verdünner wurde bei einem
Teil der Proben Eigelb zugesetzt. Der ATP-Gehalt wurde ebenfalls mit einem
Biolumineszenz-Assay bestimmt. Bei den Proben ohne Eigelb sank der ATP-Gehalt
signifikant von 12,9 nmol/ 108 Spermien an Tag 0 auf 9,6 nmol/ 108 Spermien an

Literaturübersicht
14
Tag 3. Mit Zusatz von Eigelb waren keine signifikanten Veränderungen während der
Lagerungsdauer zu messen.
Der Nukleotid-Gehalt wurde von KAMP et al. (2003) im frischen Eberejakulat, sowie
in 1:2 mit BTS verdünnten Ejakulat nach einer Haltezeit von 60 min bei 35 °C mittels
eines Spektrofluorometers gemessen. Für dieselbe Zeit wurde unverdünntes Ejakulat
bei 15 °C gehalten. Außerdem wurden die Messungen mit neunfach verdünnten
Samenproben nach einer Lagerungsdauer von 48 h bei 15 °C durchgeführt. Die
Energieladung (EC) wurde nach der in Kapitel 2.3.1 genannten Formel berechnet. Im
frischen Ejakulat wurde eine EC von 0,82 ermittelt. Nach 60 min bei 35 °C sank bei
den 1:2 verdünnten Proben die EC auf 0,33. Dagegen betrug bei den unverdünnten
Proben, die 60 min bei 15 °C gehalten wurden, die EC 0,58; nach 48 h Lagerung
wurde bei den neunfach verdünnten Proben mit 0,59 ein sehr ähnlicher Wert
ermittelt. Die Autoren vermuten, dass diese großen Unterschiede vom
Sauerstoffverbrauch der Samenzellen abhängen. Ihre Berechnungen ergaben, dass
beim Halten der Ebersamenprobe in einem geschlossenen Gefäß bei 35 °C nach
10 min der Sauerstoff aufgebraucht ist. Durch die Abkühlung werden die
Samenzellen immotil und der Verbrauch reduziert sich um 98,6%. Deshalb kann eine
höherer EC aufrechterhalten werden.
Dass die Auswirkungen der Lagerung auf die Spermien auch von dem Einzeltier
abhängen können, zeigten YI et al. (2008). Sie verglichen die Samenproben eines
kältschockresistenten Ebers mit den Samenproben eines kälteschockempfindlichen
Ebers bezüglich der Lagerungsfähigkeit bei 17°C. Als kälteschockresistent galt ein
Eber, wenn nach zwei Tagen Lagerung bei 4 °C noch mehr als 70 % motile
Spermien vorhanden waren. Als Parameter wurde unter anderem der ATP-Gehalt
der Samenzellen mittels Biolumineszenzmessung analysiert. Während der Lagerung
sank der ATP-Gehalt bei dem kälteschockresistenten Eber von
2,26 µM/ 2 x 107 Spermien auf 0,74 µM/ 2 x 107 Spermien, beim
kälteschockempfindlichen Eber von 1,57 µM/ 2 x 107 Spermien auf
0,11 µM/ 2 x 107 Spermien nach vier Tagen Lagerung bei 17 °C. Die für die
Befruchtungsfähigkeit kritische Motilität von 60 % wurde vom kälteschockresistenten

Literaturübersicht
15
Eber nach vier Tagen, vom kälteschockempfindlichen Eber nach drei Tagen
Lagerung bei 17 °C erreicht.

Material und Methoden
16
3 Material und Methoden
3.1 Versuchsübersicht
Das übergeordnete Ziel war es, die Einflüsse der Verdünnertemperatur, der
Abkühlgeschwindigkeit der Samenproben und von verschiedenen
Verarbeitungsregimes, wie zum Beispiel dem Halten des 1:1 (v:v) verdünnten oder
ausverdünnten Spermas bei 32 °C und 21 °C, auf die Qualität flüssig konservierter
Eberspermien während der Lagerung bei 17 °C zu untersuchen.
Zunächst wurden im Experiment 1 zwei im ZDS-Handbuch für Besamungsstationen
(STÄHR et al., 2009) beschriebene und in der Praxis gängige
Verarbeitungsverfahren verglichen. Im Anschluss wurden Ansätze zur Optimierung
der Prozesskette getestet, die auf verschiedenen Ebenen der Spermaverdünnung
ansetzten.
In den Experimenten 2 bis 4 wurden die Auswirkungen von Haltezeiten bei 21 °C und
32 °C des vorverdünnten (Exp. 2 und 3) bzw. ausverdünnten (Exp. 4) Samens auf
die Qualität der gelagerten Samenprobe untersucht. In Experiment 3 wurden
zusätzlich zwei Verdünnungsverfahren - zweistufig isotherm vs. zweistufig
hypertherm - vergleichend untersucht.
Experiment 5 diente zur Untersuchung der Auswirkungen einer beschleunigten
Abkühlrate des ausverdünnten Samens nach einstufig isothermer Verdünnung.
In Experiment 6 wurde der Temperaturverlauf während der Prozesskette in einer
Besamungsstation dokumentiert und in Beziehung zu Laborstandards gesetzt.
Zusätzlich wurde die Verarbeitungsmethode auf der Besamungsstation im Labor
nachgestellt und die Spermaqualität mit der nach im Laborstandard verarbeiteten
Samenproben verglichen.
Die Inhalte der Experimente sind als Übersicht in Abb. 1 und in Kapitel 3.8 detailliert
dargestellt.

Material und Methoden
17
Abb. 1: Übersicht aller durchgeführten Experimente
3.2 Verbrauchsmaterialien und Geräte
Alle verwendeten Materialien und Geräten sind im Anhang aufgelistet (Kapitel 9.4).
Experiment 6 Temperaturverlauf in der Prozesskette der Samenverarbeitung auf einer
Besamungsstation und Vergleich mit Laborstandards
Experiment 2 Auswirkungen von Haltezeiten und Haltetemperaturen des vorverdünnten Samens bei
zweistufig isothermer Verdünnung
Experiment 3
Vergleich zweistufig isothermer und zweistufig hyperthermer Verdünnung nach
Haltezeiten bei 21 °C
Experiment 5 Auswirkungen einer beschleunigten Abkühlrate des ausverdünnten Samens nach
einstufig isothermer Verdünnung
Experiment 4 a Auswirkungen von Haltezeiten des
ausverdünnten Samens nach
zweistufig hypothermer Verdünnung
Experiment 4 b
Auswirkungen von Haltezeiten des
ausverdünnten Samens nach
einstufig isothermer Verdünnung
Experiment 1
Vergleich zweistufig isothermer und zweistufig hypothermer Verdünnung bei zwei
Verdünnungsgraden

Material und Methoden
18
3.3 Lösungen und Chemikalien
Die Rezepte für die verwendeten Lösungen sowie eine Liste der dazugehörigen
Chemikalien mit deren Bezugsquellen befinden sich im Anhang dieser Arbeit (Kapitel
9.2 und 9.3).
Das Wasser wurde über eine Ionenaustauschersäule (Fa. Landgraf, Langenhagen)
aufbereitet und danach durch eine Milipore Express PLUS Membran
(Polyethersulfon, hydrophil, 0,22 µm Porendurchmesser, 47 mm Filterdurchmesser)
sterilfiltriert (Druckfiltrationseinheit Sartobran 300, Fa. Landgraf, Langenhagen).
3.4 Tiere
Für die Versuche im Labor der Reproduktionsmedizinischen Einheit der Kliniken
standen zwölf institutseigene, klinisch gesunde und im Sinne der
Gewährschaftsbestimmungen des Zentralverbandes der deutschen
Schweineproduktion (ZDS, 2005) normosperme Eber zur Verfügung. Voraussetzung
für die Normospermie waren mindestens 70 % motile Spermien und weniger als
25 % morphologisch abweichende Spermien (MAS) im Ejakulat.
Es handelte sich um fertile Eber der Rassen Pietrain (n=8), Deutsche Landrasse
(n=2) und Duroc (n=2) im Alter zwischen einem und fünf Jahren.
Für jeden Versuch wurden, sofern nicht anders beschrieben, sechs Ejakulate von
verschiedenen normospermen Ebern verwendet.
Für die Untersuchungen auf einer Besamungsstation standen elf klinisch gesunde,
fertile Eber zur Verfügung. Für diese Messungen wurden teilweise Ejakulate von
Ebern verwendet, die zu diesem Zeitpunkt nicht für den Verkauf bestimmt waren.
Fünf von diesen elf Ejakulaten waren nicht normosperm.

Material und Methoden
19
3.5 Samengewinnung
Die Samengewinnung fand mit der sogenannten „Handmethode“ mit Hilfe eines
kunststoffüberzogenen Stahlphantoms statt. Nach Abgang des Vorsekrets wurde die
spermienreiche und spermienarme Phase in einem auf ca. 36 °C vorgewärmten
Samenauffangbecher (Fa. Minitüb, Tiefenbach) aufgefangen, der mit einem
Samenauffangbeutel (US BagTM, Fa. Minitüb, Tiefenbach) versehen war. Eine im
Beutel integrierte Gaze separierte das Bulbourethraldrüsensekret vom übrigen
Ejakulat. Die Gaze wurde nach der Samenentnahme abgetrennt und der
verschlossene Beutel in einer ebenfalls auf ca. 36 °C vorgewärmten Styroporkiste in
das naheliegende Labor transportiert.
Die Samengewinnung auf der Besamungsstation fand unter ähnlichen Bedingungen
statt. Die spermienreiche und spermienarme Phase wurden in einem vorgewärmten
500 ml Schraubglas aufgefangen und mit Rohrpost direkt in das Labor verbracht.
Das Bulbourethraldrüsensekret wurde hier mittels eines auf das Glas aufgesetzten
Milchfilters getrennt.
3.6 Untersuchung des nativen Samens
Nach Ankunft im Labor wurde die Ejakulate in einen auf 36 °C vorgewärmten 500 ml
Glaszylinder umgefüllt und standardspermatologisch untersucht. Es wurden das
Volumen, die Farbe, die Konsistenz, die Spermienkonzentration, die
Spermiengesamtzahl, der pH-Wert, der Anteil motiler Samenzellen nach subjektiver
Schätzung und der Anteil der morphologisch abweichenden Spermien bestimmt.
An einem Phasenkontrastmikroskop mit Heiztisch (Axiostar Plus, Fa. Zeiss, Jena)
wurde bei 38 °C und 200-facher Vergrößerung (Phase 2) der Anteil motiler
Samenzellen geschätzt, indem mindestens drei Gesichtsfelder in drei
unterschiedlichen Tropfen (1,5 – 5 µl) ausgewertet wurden. Die hierfür verwendeten
Objektträger, Deckgläschen und Pipettenspitzen wurden auf einer Heizplatte
(HT 200, Minitüb GmbH, Tiefenbach) auf 38 °C vorgewärmt.

Material und Methoden
20
Die Spermienkonzentration wurde mit der Zählkammer „Thoma-neu“ (Fa. Jürgens,
Hannover) im Phasenkontrastmikroskop (Axiostar Plus, Zeiss, Jena) ermittelt (400-
fache Vergrößerung, Phase 2). In Anlehnung an die Methodik nach KRAUSE (1966)
erfolgte die Auswertung, wobei in jeder Kammerhälfte alle 16 Felder ausgezählt
wurden. Für ein valides Ergebnis mussten in jeder Hälfte mindestens 100
Spermienköpfe vorhanden sein, der Toleranzbereich zwischen den Hälften lag bei
einer Differenz von 10 %. Die Spermiengesamtzahl errechnete sich aus dem Produkt
von Volumen und Spermienkonzentration des Ejakulates. Der pH-Wert wurde mit
einem pH-Meter (Multiplex 3000/ pMX, Fa. WTW, Weilheim) gemessen.
Die Bestimmung des Anteils morphologisch abweichender Spermien wurde in
Anlehnung an die Klassifikation nach KRAUSE (1966) durchgeführt. Abhängig von
der Spermienkonzentration wurden zwischen 5 – 30 µl des nativen Samens in 300 µl
4 %-iges Formolcitrat pipettiert. Je 200 Zellen wurden bei 1000-facher Vergrößerung
(Ölimmersion, Phase 3) mikroskopisch beurteilt.
3.7 Monitoring der Probentemperatur
Das Abkühlverhalten der Samenproben bei der Anwendung der verschiedenen
Verdünnungsarten wurde durch den Temperaturlogger Micromec® multisens (Fa.
Technetics, Freiburg) dokumentiert. Dieser verfügt über acht flexible, zwei bis fünf
Meter lange Temperatursonden (TK700C, Messbereich -200 bis +600 °C,
Messgenauigkeit 0,1 °C, Technetics, Freiburg), die auch in Flüssigkeiten zur
Temperaturmessung eingesetzt werden können. Da während der Versuche die
Temperaturmessung nicht möglich war, erfolgten die Messungen in einem separaten
Ansatz mit wassergefüllten Ebersamenflaschen. In einem Vorversuch konnte gezeigt
werden, dass das Abkühlverhalten einer Ebersamenprobe mit 20 Mio Spermien/ ml
dem Abkühlverhalten von Wasser entspricht. Für jedes Experiment wurde die
Temperatur in je 3 Probengefäßen (500 ml Glaszylinder bzw. aufrechtstehende
Ebersamenflaschen) mit einer Doppelmessung (je zwei Sonden pro Probengefäß,
mittig angebracht) aufgezeichnet. Über 6 Stunden erfolgte einmal pro Minute die
Aufzeichnung der Probentemperatur, zeitgleich wurde mit einer weiteren Sonde die

Material und Methoden
21
jeweilige Umgebungstemperatur dokumentiert. Das Auslesen der Daten am PC
erfolgte mit der Software MM-grafix (Version 7.1, Technetics, Freiburg). Die Daten
wurden in Excel exportiert, wo die weitere Auswertung und Erstellung der Grafiken
stattfand.
3.8 Experimentelle Designs
3.8.1 Experiment 1: Vergleich zweistufig isothermer und zweistufig
hypothermer Verdünnung bei zwei Verdünnungsgraden
Hypothese:
Bei zweistufigem Verdünnungsverfahren weisen Samenproben, die im zweiten
Verdünnungsschritt isotherm verdünnt wurden, eine bessere Qualität während der
Lagerung bei 17 °C auf, als solche die hypotherm verdünnt wurden. Der negative
Effekt einer zweistufig hypothermen Verdünnung ist stärker ausgeprägt bei einer
Samenprobe mit höherem Verdünnungsgrad (10 x 106 Spermien/ ml) als bei einem
Verdünnungsgrad von 20 x 106 Spermien/ ml.
Versuchsdurchführung:
Nach der standardspermatologischen Untersuchung und Dichtebestimmung erfolgte
die isotherme Verdünnung des Samens mit auf 32 °C vorgewärmten BTS (Beltsville
Thawing Solution, ca. 330 mOsmol/ kg, pH 7,05-7,11 bei Raumtemperatur; Fa.
Minitüb, Tiefenbach) im Verhältnis 1:1 (v:v). Um ein vergleichbares
Temperaturverhalten der Proben zu erreichen wurden stets konstante Volumina von
200 ml Ejakulat und 200 ml BTS in einem auf 32 °C vorgewärmten 500 ml
Glaszylinder vermischt. Nach einer erneuten Dichtebestimmung wurde die
Samenprobe zu gleichen Teilen in zwei auf 32 °C vorgewärmte 250 ml Glaszylinder
überführt und die Zylinder mit Parafilm verschlossen.
Für die zweistufig isotherme Verdünnung die nach den Vorgaben für die
„zweiphasige Verdünnung“ des ZDS-Handbuchs (2009) durchgeführt wurde, wurde
einer der Zylinder während der zweiten Dichtebestimmung (für ca. 15 min) bei 32 °C
im Wasserbad gehalten. Anschließend erfolgte die Verdünnung in

Material und Methoden
22
Ebersamenflaschen (100 ml, Fa. Minitüb, Tiefenbach) mit auf 32 °C temperiertem
Verdünner auf eine Endkonzentration von 20 x 106 Spermien/ ml bzw.
10 x 106 Spermien/ ml. Hierzu wurde die entsprechende Menge des vorverdünnten
Samens in die Flaschen vorgelegt und mit BTS auf 100 ml aufgefüllt. Nachdem
verbleibende Luft so weit wie möglich entfernt wurde, wurden die Flaschen
verschlossen, für 90 min bei 21 °C gehalten und daraufhin bei 17 °C im Klimaschrank
gelagert. Die Abkühlung bzw. Lagerung der Flaschen erfolgte immer stehend und mit
ca. 2 cm Abstand zueinander, um für alle Proben eine möglichst gleiche
Abkühlungsrate zu gewährleisten.
Der zweite Glaszylinder wurde für die zweistufig hypotherme Verdünnung, die nach
den Vorgaben für die „zweiphasige Verdünnung“ des ZDS-Handbuchs (2009)
durchgeführt wurde, für 20 min in einem auf 21 °C temperierten Klimaschrank (Fa.
Minitüb, Tiefenbach) gehalten. Danach fand die Ausverdünnung wie oben
beschrieben statt, mit dem Unterschied, dass ein auf 21 °C temperierter Verdünner
verwendet wurde. Wie bei der isothermen Variante kühlten die Proben 90 min bei
21 °C ab. Die Lagerung erfolgte ebenfalls wie oben beschrieben bei 17 °C.
Die Messung der Motilitätsparameter mittels computerassistierter Spermienanalyse
(CASA; siehe Kapitel 3.9) und der Integrität der Plasma- und Akrosommembran
(siehe Kapitel 3.10) der zweistufig isothermen verdünnten Probe fand unmittelbar
nach der Verdünnung der Proben auf die finale Konzentration statt. Die weitere
Analyse aller Proben erfolgte nach 24 h, 72 h und 144 h.
Der Ablauf von Experiment 1 ist in Abb. 2 dargestellt.
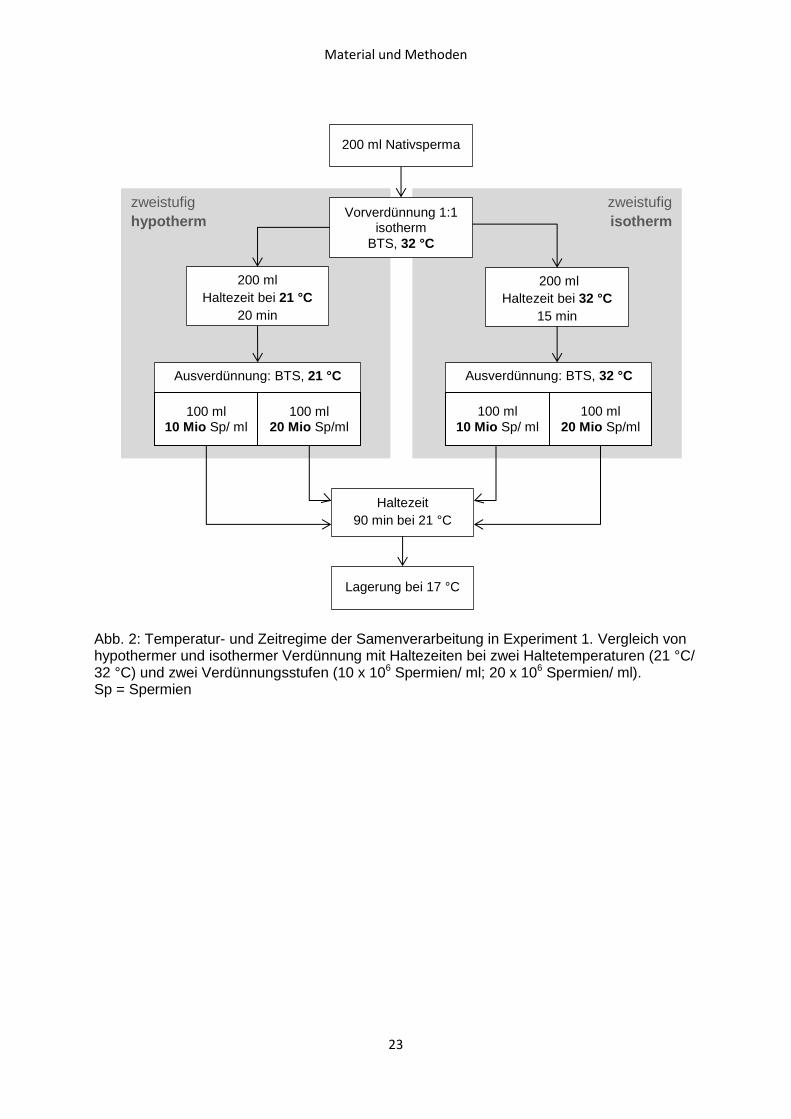
Material und Methoden
23
Abb. 2: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 1. Vergleich von hypothermer und isothermer Verdünnung mit Haltezeiten bei zwei Haltetemperaturen (21 °C/ 32 °C) und zwei Verdünnungsstufen (10 x 106 Spermien/ ml; 20 x 106 Spermien/ ml). Sp = Spermien
zweistufig
hypotherm
zweistufig
isotherm
Vorverdünnung 1:1 isotherm
BTS, 32 °C
200 ml
Haltezeit bei 21 °C
20 min
200 ml
Haltezeit bei 32 °C
15 min
Haltezeit
90 min bei 21 °C
Lagerung bei 17 °C
200 ml Nativsperma
Ausverdünnung: BTS, 21 °C
100 ml 10 Mio Sp/ ml
100 ml 20 Mio Sp/ml
Ausverdünnung: BTS, 32 °C
100 ml 10 Mio Sp/ ml
100 ml 20 Mio Sp/ml

Material und Methoden
24
3.8.2 Experiment 2: Auswirkungen von Haltezeiten und Haltetemperaturen
des vorverdünnten Samens bei zweistufig isothermer Verdünnung
Hypothese:
Haltezeiten des 1:1 (v:v) verdünnten Spermas bis 60 Minuten bei 32 °C haben im
Gegensatz zu Haltezeiten bei 21 °C einen positiven Einfluss auf die Qualität der bei
17°C gelagerten Samenproben.
Längere Haltezeiten (> 60 min bis 6 h) bei 32 °C haben dagegen einen negativen
Einfluss auf den Energiestoffwechsel und das Keimwachstum der später bei 17 °C
gelagerten Samenproben.
Versuchsdurchführung:
Da in der Prozesskette der Samenverarbeitung in Besamungsstationen
Verzögerungen während der Verarbeitung und damit Haltezeiten der Ejakulate
unterschiedlicher Länge auftreten können, wurde in diesem Versuch das
Hauptaugenmerk auf die Auswirkungen von Haltezeiten (30 min, 60 min, 3 h und 6 h)
von 1:1 (v:v) verdünntem Samen bei 32 °C auf die Qualität der gelagerten
Samenproben gelegt. Hierbei wurden neben den Motilitätsparametern und der
Membranintegrität zusätzlich der ATP-Gehalt und die Energieladung der Spermien,
der Anteil der Spermien mit hohem Mitochondrienmembranpotential, sowie der Effekt
auf die mikrobiologische Qualität analysiert. Die Verdünnung des Samens erfolgte
zweistufig isotherm.
Für diesen Versuch wurden insgesamt zwölf Ejakulate von neun verschiedenen
Ebern verwendet. Der Versuch wurde zweimal mit jeweils sechs Ejakulaten
durchgeführt. Da der zeitliche Abstand der Versuche bei ca. einem Jahr lag, wurde
jedes Ejakulat trotz der Verwendung der gleichen Tiere eigenständig gewertet.
Die Vorverdünnung, die zweistufig isotherme Verdünnung sowie die Abkühlung und
Lagerung der Samenproben wurde wie in Kapitel 3.8.1 beschrieben, durchgeführt.
Die vorverdünnten Samenproben wurden vor der Ausverdünnung Haltezeiten
ausgesetzt, mit Ausnahme der Kontrollproben, die direkt ausverdünnt wurden. Eine
Hälfte der 1:1 (v:v) vorverdünnten Samenproben (Zylinder 1) wurde für 30 min, 60
min, 3 h und 6 h Haltezeiten bei 32 °C gehalten. Die Ausverdünnung fand direkt nach
jeder Haltezeit statt.

Material und Methoden
25
Der zweite Zylinder wurde für dieselben Haltezeiten im Klimaschrank bei 21 °C
gehalten. Um auch hier eine zweistufig isotherme Verdünnung gewährleisten zu
können, wurde ein 500 ml Glaszylinder mit einem auf 32 °C vorgewärmtem
Verdünner zeitgleich in den Klimaschrank verbracht und kühlte parallel mit der Probe
ab. Die isotherme Ausverdünnung fand ebenfalls nach jeder Haltezeit statt. Die
Abkühlung und Lagerung der Proben entsprachen den bei 32 °C verarbeiteten
Proben.
Die Messung der Membranintegrität (s. Kapitel 3.10) und der Motilitätsparameter
(s. Kapitel 3.9) wurde bei allen zwölf Ejakulaten direkt nach jeder Haltezeit sowie
nach 24 h und 72 h durchgeführt. Sechs Ejakulate wurden einer weiteren
Untersuchung nach 144 h Lagerung unterzogen.
Das Mitochondrienmembranpotential wurde bei sechs Ejakulaten zu denselben
Zeitpunkten wie die Membranintegrität und die Motilität analysiert (s. Kapitel 3.11).
Die Probenentnahme für die Bestimmung der ATP-Konzentration und der
Energieladung im nativen Samen erfolgte zunächst direkt nach der
standardspermatologischen Untersuchung (s. Kapitel 3.13). Dazu wurde eine
zusätzliche Samenprobe (100 ml, 20 x 106 Spermien) direkt nach der
standardspermatologischen Untersuchung isotherm mit vorgewärmtem BTS
verdünnt, um eine vergleichbare Menge Samenzellen für die Messung eines
Kontrollwerts der ATP-Konzentration zur Verfügung zu haben. Die weitere
Probenentnahme fand zu den oben genannten Messzeitpunkten, also nach jeder
Haltezeit sowie nach 24 h, 72 h, und 144 h Lagerung statt.
Der Einfluss der Haltezeiten auf die mikrobiologische Qualität der Proben wurde
durch eine Gesamtkeimzahlbestimmung ermittelt. Von 4 Ejakulaten wurde je 1 ml
des nativen Samens entnommen. Am Tag der Verdünnung erfolgte des Weiteren die
Beprobung des 1:1 (v:v) verdünnten Samen der Kontrolle (nach ZDS-Handbuch
verarbeitete Probe) sowie nach 60 min und 6 h Inkubation bei 32 °C sowie 21 °C.
Nach 72 h Lagerungsdauer wurde in den ausverdünnten Proben erneut eine
Gesamtkeimzahlbestimmung durchgeführt. Alle Proben für die mikrobiologische
Untersuchung wurden nach der Entnahme gekühlt (< 10°C, Styroporbox mit
Material und Methoden
26
Kühlakkus) ins Institut für Mikrobiologie der Stiftung Tierärztliche Hochschule
Hannover verbracht und dort untersucht (s. Kapitel 3.14).
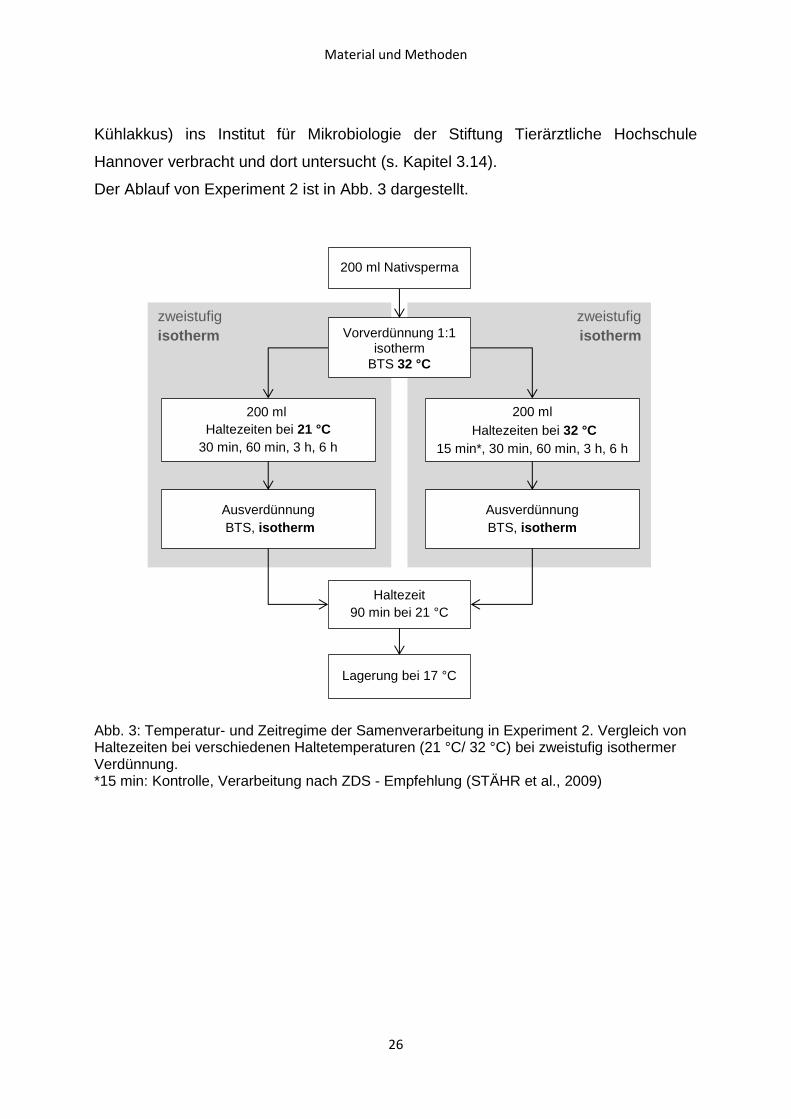
Der Ablauf von Experiment 2 ist in Abb. 3 dargestellt.
Abb. 3: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 2. Vergleich von Haltezeiten bei verschiedenen Haltetemperaturen (21 °C/ 32 °C) bei zweistufig isothermer Verdünnung. *15 min: Kontrolle, Verarbeitung nach ZDS - Empfehlung (STÄHR et al., 2009)
zweistufig
isotherm
zweistufig
isotherm
Vorverdünnung 1:1 isotherm
BTS 32 °C
200 ml
Haltezeiten bei 32 °C
15 min*, 30 min, 60 min, 3 h, 6 h
Ausverdünnung
BTS, isotherm
Haltezeit
90 min bei 21 °C
Lagerung bei 17 °C
200 ml Nativsperma
Ausverdünnung
BTS, isotherm
200 ml
Haltezeiten bei 21 °C
30 min, 60 min, 3 h, 6 h

Material und Methoden
27
3.8.3 Experiment 3: Vergleich zweistufig isothermer mit zweistufig
hyperthermer Verdünnung
Hypothese:
Die Ausverdünnung mit im Vergleich zur Samenprobe hyperthermen Verdünner hat
einen negativen Einfluss auf die Qualität des bei 17 °C gelagerten Samens.
Versuchsdurchführung:
Während des Betriebsablaufs in einer Besamungsstation kann es zu Verzögerungen
kommen und damit zu Haltezeiten bei Raumtemperatur, so dass der vorverdünnte
Samen allmählich abkühlt. Der Verdünner wird hingegen in der Regel auf einer
konstanten Temperatur von 32 °C gehalten. Daraus kann sich unter
Praxisbedingungen eine hypertherme Ausverdünnung ergeben. Im Experiment
wurde die zweistufig hypertherme Verdünnung der im Kapitel 3.8.1 untersuchten
zweistufig isothermen Verdünnung als Kontrolle gegenübergestellt.
Die Vorverdünnung sowie die Abkühlung und Lagerung erfolgten wie in Kapitel 3.8.1
beschrieben, die zweistufig isotherme Verdünnung mit Haltezeiten analog zu der
Beschreibung in Kapitel 3.8.2.
Für die zweistufig hypertherme Verdünnung wurde ebenfalls ein 250 ml Glaszylinder
mit 200 ml vorverdünntem Sperma für die angegebenen Haltezeiten, mit Ausnahme
der Kontrollprobe, bei 21 °C gehalten. Ein 500 ml Zylinder mit BTS wurde während
des Versuchs bei 32 °C gehalten. Die hypertherme Ausverdünnung erfolgte nach
jeder der Haltezeiten mit dem auf 32 °C temperierten Verdünner.
Die untersuchten spermatologischen Parameter waren auch in diesem Experiment
die Membranintegrität (s. Kapitel 3.10) sowie die Messung CASA –
Motilitätsparameter (s. Kapitel 3.9). Die Messungen wurden jeweils direkt nach der
Verdünnung auf die Endkonzentration und nach 24 h und 72 h Lagerungsdauer
durchgeführt.
Der Ablauf von Experiment 3 ist in Abb. 4 dargestellt.

Material und Methoden
28
Abb. 4: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 3. Vergleich zweistufig isothermer und zweistufig hyperthermer Verdünnung nach Haltezeiten bei 21 °C. *15 min: Kontrolle, Verarbeitung nach ZDS - Empfehlung (STÄHR et al., 2009)
zweistufig
isotherm
zweistufig
hypertherm
Vorverdünnung 1:1 isotherm
BTS 32 °C
200 ml
Haltezeiten bei 21 °C
30 min, 60 min, 3 h, 6 h
Ausverdünnung
BTS, 32 °C
Ausverdünnung
BTS, isotherm
Haltezeit
90 min bei 21 °C
Lagerung bei 17 °C
200 ml Nativsperma
200ml
Haltezeiten bei 21 °C
15min*, 30 min, 60 min, 3 h, 6 h

Material und Methoden
29
3.8.4 Experiment 4: Auswirkungen von Haltezeiten des ausverdünnten
Samens nach zweistufig hypothermer (Exp. 4 a) oder einstufig isothermer
Verdünnung (Exp. 4 b)
Hypothese:
Haltezeiten des zweistufig hypotherm oder einstufig isotherm ausverdünnten
Spermas haben einen positiven Einfluss auf die Qualität der bei 17 °C gelagerten
Samenprobe.
Versuchsdurchführung Exp. 4 a:
Isotherm 1:1 (v:v) vorverdünntes Sperma (Volumen: 400 ml) wurde 20 min bei 21 °C
gehalten und anschließend mit auf 21 °C temperierten BTS in 100 ml
Ebersamenflaschen ausverdünnt. Die Kontrolle wurde gemäß der ZDS-
Empfehlungen für 90 min bei 21 °C gehalten und dann bei 17 °C gelagert. Eine
Versuchsvariante wurde bis zu 6 h bei 21 °C gehalten, während die zweite
Versuchsvariante bei 32 °C gehalten wurde. Nach Ende der Haltezeiten (30 min,
60 min, 3 h bzw. 6 h) wurden alle Proben für 90 min bei 21 °C gehalten und
anschließend bei 17 °C gelagert.
Die Analyse der Membranintegrität (s. Kapitel 3.10) sowie der Motilität (s. Kapitel 3.9)
fand am Tag der Samenverarbeitung (d 0) sofort nach den initialen Haltezeiten
(30 min – 6 h) und nach 24 h und 72 h Lagerung statt.
Der Ablauf von Experiment 4 a ist in Abb. 5 dargestellt.
Versuchsdurchführung Exp. 4 b:
Dieses Experiment untersuchte ebenfalls den Effekt von Haltezeiten des
ausverdünnten Spermas auf die Qualität des gelagerten Spermas. Der Unterschied
zu Experiment 4 a lag darin, dass in diesem Versuch eine einstufig isotherme
Verdünnung verwendet wurde, die nach den Empfehlungen des ZDS-Handbuchs für
„einphasige Verdünnung“ durchgeführt wurde.
Die Ejakulate wurden nach der standardspermatologischen Untersuchung mit auf
32 °C temperierter, isothermer BTS in einem Schritt zur Endkonzentration
20 x 106 Spermien/ ml ausverdünnt. Hierfür wurde die entsprechende Menge

Material und Methoden
30
Ejakulat in der Ebersamenflasche vorgelegt und langsam mit dem Verdünner auf
100 ml aufgefüllt. Die Kontrolle (Verarbeitung ohne Haltezeiten, nach ZDS-
Handbuch) wurde wie bei den vorausgegangenen Versuchen erstellt, während in den
Versuchsvarianten Samenproben für 30 min, 60 min, 3 h bzw. 6 h bei 32 °C bzw.
21 °C gehalten wurden. Die weitere Abkühlung, Lagerung und Analyse erfolgte
analog zu Experiment 4 a.
Der Ablauf von Experiment 4 b ist in Abb. 6 dargestellt.
Abb. 5: Temperatur- und Zeitregime der zweistufig hypothermen Samenverarbeitung in Experiment 4 a. Vergleich von Haltezeiten bei verschiedenen Haltetemperaturen (21 °C/ 32 °C) des ausverdünnten Samens nach zweistufig hypothermer Verdünnung.
zw
eis
tufi
g
hyp
oth
erm
Ausverdünnung mit BTS 21 °C
Haltezeiten bei 21 °C
30 min, 60 min, 3 h, 6 h
Haltezeiten bei 32 °C
30 min, 60 min, 3 h, 6 h
Haltezeit
90 min bei 21 °C
Lagerung bei 17 °C
Vorverdünnung 1:1
isotherm
BTS 32 °C
Haltezeit bei 21 °C
20 min
0 min Haltezeit
(Kontrolle)
200 ml Nativsperma

Material und Methoden
31
Abb. 6: Temperatur- und Zeitregime der einstufig isothermen Samenverarbeitung in Experiment 4 b. Vergleich von Haltezeiten bei verschiedenen Haltetemperaturen (21 °C/ 32 °C) des ausverdünnten Samens nach einstufig isothermer Verdünnung.
3.8.5 Experiment 5: Auswirkungen einer beschleunigten Abkühlrate des
ausverdünnten Samens nach einstufig isothermer Verdünnung
Hypothese:
Eine schnelle Abkühlrate der Samenprobe während der Frühphase der
Konservierung hat einen negativen Einfluss auf die Qualität der gelagerten
Samenprobe.
Versuchsdurchführung:
Die Folgen einer erhöhten Abkühlrate nach der Samenverarbeitung auf die Qualität
des bei 17°C gelagerten Samens wurden mit diesem Versuch untersucht.
Ausverdünnung einstufig isotherm
mit BTS, 32 °C
Haltezeiten bei 21 °C
30 min, 60 min, 3 h, 6 h
Haltezeiten bei 32 °C
30 min, 60 min, 3 h, 6 h
Haltezeit
90 min bei 21 °C
200 ml Nativsperma
Lagerung bei 17 °C
0 min Haltezeit
(Kontrolle)

Material und Methoden
32
Zu diesem Zweck wurden die Ejakulate wie in Kapitel 3.8.4 (Exp. 4 b) beschrieben
einstufig isotherm mit auf 32 °C temperierter BTS verdünnt. Anschließend wurde je
ein Teil der Proben in 100 ml Flaschen wie bei den vorausgegangenen Versuchen
90 min bei 21 °C gehalten und danach bei 17 °C gelagert. Der andere Teil der
Samenproben wurde direkt nach der einstufig isothermen Verdünnung in die
Lagerung bei 17 °C überführt.
Die Analyse von Membranintegrität (s. Kapitel 3.10) und Motilität (s. Kapitel 3.9) fand
nach 24 h, 72 h und 144 h Lagerungsdauer statt.
Der Ablauf von Experiment 5 ist in Abb. 7 dargestellt.
Abb. 7: Temperatur- und Zeitregime der einstufig isothermen Samenverarbeitung in Experiment 5. Vergleich von einer beschleunigten Abkühlrate ohne Haltezeit zur Standardabkühlung mit 90 min Haltezeit nach einstufig isothermer Verdünnung.
Ausverdünnung einstufig isotherm
mit BTS 32 °C
Haltezeit
90 min bei 21 °C
(Kontrolle)
Nativsperma
Lagerung bei 17 °C

Material und Methoden
33
3.8.6 Experiment 6: Temperaturverlauf in der Prozesskette der
Samenverarbeitung auf einer Besamungsstation und Vergleich mit
Laborstandards
3.8.6.1 Temperaturverlauf im Sperma während der Verarbeitung auf einer
Besamungsstation bei Samenverarbeitung
Dieser Versuch wurde in Zusammenarbeit mit dem Labor einer Besamungsstation
durchgeführt. Ziel des Experimentes war es, die Haltezeiten des vorverdünnten
Samens während der Samenverarbeitung und den Temperaturverlauf der Proben in
der Prozesskette, von Ankunft der Ejakulate im Labor bis zur Umlagerung der
Spermaportionen für den Versand, in Relation zur Qualität des gelagerten Samens
setzen.
Die frisch gewonnenen Ejakulate erreichten das Labor via Rohrpost in einem 500 ml
Schraubglas. Als erstes wurde die Spermienkonzentration des Ejakulats gemessen
und direkt eine erste Vorverdünnung im Glas durchgeführt. Die Vorverdünnung
erfolgt mit auf 32 °C angewärmter BTS, indem jede Probe auf 500 ml aufgefüllt
wurde. Im Anschluss wurden die Motilität und der Anteil morphologisch
abweichender Spermien mikroskopisch geschätzt. Das Sperma wurde mit auf 26 °C
temperierter BTS ausverdünnt und anschließend in 80 ml Tuben abgefüllt. Die
Samenportionen wurden nach Ebern getrennt in Tüten gefüllt und in oben offenen
Plastikboxen im anliegenden Versandraum (21 °C Raumtemperatur) bis zum
Versand gelagert. Die am Samengewinnungstag nicht verkauften Proben wurden in
einem verschließbaren auf 17 °C temperierten Kühlregal bis zum nächsten Tag
gelagert.
Um die Haltezeiten vor der Abfüllung unter Praxisbedingungen zu erfassen, wurde
bei sieben Ejakulaten die Dauer der Verzögerungen während der Prozesskette
notiert.
Anhand von vier Ejakulaten einer Tagesproduktion wurde die Abkühlung während
der gesamten Verarbeitungskette gemessen. Direkt nach der Ankunft im Labor
wurden die Ejakulate wie oben beschrieben vorverdünnt. Zwei Ejakulate wurden
direkt weiterverarbeitet, die anderen Beiden wurden erst nach einer Stunde abgefüllt,

Material und Methoden
34
um eine Haltezeit zu simulieren. Während dieser Zeitspanne wurde die Temperatur
des vorverdünnten Spermas zwölfmal, im Abstand von drei bis sechs Minuten, mit
einem Glasthermometer gemessen und notiert. Bei der anschließenden
Untersuchung und zweiten Verdünnung mit dem auf 26 °C temperierten BTS wurde
ebenfalls die Temperatur des Samens gemessen. Unmittelbar nach der Abfüllung
wurden die Samentuben je eines Ebers in eine Plastikbox gelegt und in den
Versandraum verbracht. Dort wurden jeweils zwei Tuben eines verarbeiteten
Ejakulates (n = 4) mit einer Temperatursonde (mikromec® multisens) bestückt. Um
die Temperatur in der Tube zu messen, wurde die Tube an der Tülle aufgeschnitten
und die Sonde bis zur Mitte der Tube eingeführt. Mit Hilfe eines vulkanisierenden
Klebebandes wurde die Sonde an der Tülle fixiert. Anschließend wurde eine Tube
mittig, zwischen die anderen Tuben in die Box gelegt, so dass sie von allen Seiten
von der gleichen Menge Tuben umgeben war. Die andere Tube wurde oben auf den
anderen Tuben platziert. Die Temperatur wurde einmal pro Minute dokumentiert.
Während der gesamten Verarbeitung betrug die Raumtemperatur am Versuchstag
zwischen 21 °C und 22 °C. Nachdem der Versand der Tuben für diesen Tag beendet
war (ca. 5 h nach Beginn des Versuchs), wurden die Boxen in ein Klimaregal mit
17 °C umgelagert, welches nach ca. 1,5 h mit einem Rollo verschlossen wurde. 6,5 h
nach Platzierung der Messsonden in den Tuben wurde die Messung beendet.
Die Prozesskette in der Besamungsstation ist in Abb. 8 dargestellt.

Material und Methoden
35
Abb. 8: Prozesskette der Samenverarbeitung in einer Besamungsstation. Die grauen Flächen stellen Zeitpunkte für mögliche Verzögerungen in der Verarbeitung dar, aus denen sich Haltezeiten unterschiedlicher Länge ergeben.
3.8.6.2 Vergleich des Verdünnungsverfahrens einer Besamungsstation mit dem
Laborstandard
Im Labor der Reproduktionsmedizinischen Einheit der Kliniken der Tierärztlichen
Hochschule Hannover wurde das Temperaturregime der Besamungsstation
nachgestellt und mit dem Laborstandard (Verarbeitung nach ZDS-Empfehlungen für
„einphasige Verdünnung“) verglichen. Die wesentlichen Unterschiede in der
Vorgehensweise auf Besamungsstationen und im Forschungslabor liegen darin,
dass in Besamungsstationen zumeist zweistufig hypotherm verdünnt wird und die
vorverdünnten Ejakulate Haltezeiten unterschiedlicher Länge bei Raumtemperatur
Dichtebestimmung
Ankunft des Samens im Labor
(500 ml Schraubglas, Rohrpost)
mikroskopische
Motilitätsschätzung
Ausverdünnung (BTS 26 °C)
Abfüllung
Versandraum (21 °C)
Vorverdünnung im Glas (Auffüllen auf 500 ml)
BTS 32 °C
Haltezeit bei RT
Haltezeit bei RT
Haltezeit bei RT

Material und Methoden
36
ausgesetzt sind. Um die Samenqualität während der Lagerung zu bestimmen,
wurden CASA-Motilitätsparameter (Kapitel 3.9), Membranintegrität (Kapitel 3.10) und
Calcium-Influx (Kapitel 3.12) gemessen.
Versuchsdurchführung:
Bei dem Laborstandard wird direkt nach der standardspermatologischen
Untersuchung das Ejakulat in der Ebersamenflasche auf 20 x 106 Spermien/ ml
isotherm mit auf 32 °C vorgewärmter BTS ausverdünnt. Die Samenproben werden
dann für 90 min bei 21 °C gehalten und anschließend zur weiteren Abkühlung und
Lagerung bei 17 °C gelagert.
Bei der Simulation der Samenverarbeitung der Besamungsstation fand nach der
standardspermatologischen Untersuchung eine isotherme Vorverdünnung (BTS,
32 °C) des Nativspermas im Verhältnis 1:1 (250 ml: 250 ml) statt. Danach wurde der
Samen 30 min bei 21 °C gehalten. Diese Zeitspanne entsprach der vorher ermittelten
mittleren Standzeit der Besamungsstation. Daraufhin wurde mit auf 26 °C
temperierten Verdünner die Ausverdünnung in der Ebersamenflasche durchgeführt.
Für die weitere Abkühlung wurde die Flasche in einer Plastikbox mittig zwischen 40
Tuben gelegt, die auf dieselbe Temperatur vorgewärmt waren. Analog zu den
Messungen in der Besamungsstation verblieb die Box für 2,5 h bei 21 °C.
Anschließend wurde diese in den auf 17 °C temperierten Klimaschrank verbracht.
Die Messung der Motilitätsparameter (s. Kapitel 3.9) und der Membranintegrität (s.
Kapitel 3.10) fanden nach 24 h und 144 h statt. Die Motilitätsparameter wurden
zusätzlich nach 72 h bestimmt. Die Analyse des Calcium-Influx (s. Kapitel 3.12) fand
nach 24 h Lagerungsdauer statt.
Der Ablauf des Vergleichs ist in Abb. 9 dargestellt.

Material und Methoden
37
Abb. 9: Vergleich von Temperatur- und Zeitregime der Samenverarbeitung in der
Besamungsstation mit Laborstandard in Experiment 6. Sp = Spermien
3.9 Computerassistierte Spermienanalyse (CASA)
3.9.1 Testprinzip
Die Untersuchung der Probe fand im negativen Phasenkontrast statt, so dass die
Spermienköpfe als weiße Fläche auf grauem Hintergrund erschienen. Die Spermien
wurden von einer Kamera erfasst und die Bilder an den angeschlossenen Computer
weitergeleitet. Die weißen Flächen wurden von der Software als Samenzellen
erkannt und diese ermittelte daraus den Anteil motiler Spermien. Aus den Daten
wurden zusätzlich verschiedene Motilitätsparameter wie zurückgelegte Distanz,
Geschwindigkeit der Spermien und deren Bewegungsmuster errechnet.
Station Standard Vorverdünnung 1:1
isotherm, BTS 32 °
Haltezeit bei 21 °C
30 min
Ausverdünnung
20 x 106 Sp/ ml
hypotherm, BTS 26 °C
Ausverdünnung
20 x 106 Sp/ ml
isotherm, BTS 32 °C
Haltezeit in Flaschen
einzeln
90 min bei 21 °C
Lagerung bei 17 °C
Nativsperma
Haltezeit in Tuben in
Box mittig zwischen 40 auf
26 °C temperierten Tuben
Lagerung bei 17 °C

Material und Methoden
38
3.9.2 Gerät und Software
Für die Messung der Motilitätsparameter stand das SpermVision®-System (Fa.
Minitüb, Tiefenbach) zur Verfügung. Dieses bestand aus einem Mikroskop (Okular
10, Objektiv 20, BX41TF, Fa. Olympus, Hamburg) mit einem beheizbaren Objekttisch
und einem automatisch fahrbaren Kreuztisch, sowie einer Heizplatte (HT 300, Fa.
Minitüb, Tiefenbach) zum Vorwärmen der Messkammern und Pipettenspitzen, die
gleichzeitig die Temperatur des Objekttisches steuerte. Für die Messung wurde die
Probe in eine Leja-Messkammer mit 20 µm Schichtdicke (SC 20-01-04-B, Leja
Products B.V., Nieuw-Vennep, Niederlande) pipettiert. Die mit Hilfe eines TV-
Adapters (U-TV 0.63XC; Olympus, Hamburg) am Mikroskop angebrachte
Digitalkamera (AccuPiXEL TM6760 CL, Auflösung 600 x 800 Pixel, JAI A/S,
Glostrup, Dänemark), nahm die Bilder des Präparates auf, die direkt an den
Computer weitergeleitet wurden. Für die Analyse der Bilder und die Steuerung des
automatischen Kreuztisches wurde die Software SpermVision® (Version 3.7, Fa.
Minitüb, Tiefenbach) verwendet. Für die Bewegungsparameter und die Motilität
wurden 10 aufeinanderfolgende Positionen, die in der zentralen Längsachse der
Kammer lagen, angesteuert. Pro Messposition wurde in 0,5 Sekunden eine Serie von
30 Einzelbildern aufgenommen. Als Spermienkopf wurde vom System jede weiße
Fläche zwischen 23 µm2 und 120 µm2 detektiert. Anhand deren Positionsänderung
während der 0,5 Sekunden errechnete die Software die folgenden
Motilitätsparameter:
Zurückgelegte Wegstrecken [µm]: DSL = distance straight line
DCL = distance curvilinear line
DAP = distance average path
Geschwindigkeiten [µm/ s]: VSL = velocity straight line
VCL = velocity curvilinear line
VAP = velocity average path

Material und Methoden
39
Gerichtetheit der Bewegung: WOB = VAP / VCL = wobble
STR = VSL / VAP = straightness
LIN = VSL / VCL = linearity
gemittelte maximale seitliche Kopf-: ALH = amplitude of lateral head-
auslenkung von der mittleren Bahn displacement
[µm]
Frequenz des Kreuzens des tat-: BCF = beat cross frequency
sächlichen Bewegungspfads mit
der gemittelten Bahn [Hz]
durchschnittlicher Richtungswechsel: AOC = average orientation change
des Spermienkopfes [°]
Die Motilitätsklassen der einzelnen Samenzellen wurden anhand der berechneten
Parameter folgendermaßen definiert:
Immotil: AOC < 2,5
lokal motil: AOC > 2,5 und DSL < 4,5
progressiv motil: Spermien, die nicht diesen Kriterien entsprachen.
Als Ergebnis der Motilitätsmessung wurden die Gesamtmotilität in Prozent und der
prozentuale Anteil der progressiv motilen Spermien dokumentiert. Die
Motilitätsparameter wurden für jede einzelne Samenzelle gespeichert und ein
Mittelwert von jedem Parameter für die Gruppe der progressiv motilen Spermien
berechnet.
3.9.3 Messung
Als Vorbereitung für die Messungen wurden die Heizplatte und der Objekttisch
genauso wie die Leja-Messkammern und die Pipettenspitzen auf 38 °C erwärmt.

Material und Methoden
40
Zwei Milliliter des verdünnten Samens wurden für 15 min in einem offenen
Reagenzglas bei 38 °C im Wasserbad inkubiert. Nach Ablauf der Inkubationsdauer
wurde das Reagenzglas mit Parafilm verschlossen und zur Durchmischung evtl.
abgesetzter Samenzellen mehrfach um 180° geschwenkt. Die Messkammer wurde
dann mit 2,7 µl der Probe befüllt, so dass eine vollständige Befüllung ohne
Lufteinschlüsse vorlag. Dazu wurde die Pipette genau senkrecht über der
Einfüllmulde positioniert, und der Inhalt gleichmäßig freigegeben, so dass der
Tropfen durch Kapillarkräfte in die Kammer gesogen wurde. Die erfolgreich befüllte
Kammer wurde mit dem Kreuztisch automatisch auf die erste Messposition gefahren
und, nach fokussieren des Bildes mit dem Feintrieb des Mikroskops und Einstellen
der Lichtintensität für einen optimalen Kontrast, die Analyse gestartet.
Alle Messungen wurden innerhalb von 60 Sekunden nach Befüllen der Kammer
abgeschlossen und am Ende des Versuchstags die Messdaten als Excel-Datei
exportiert.
3.10 Membranintegrität in der Durchflusszytometrie
3.10.1 Testprinzip
Diese Analyse beruht auf der unterschiedlichen Permeabilität für
Fluoreszenzfarbstoffe von intakten und defekten Membranen der Samenzelle.
Anhand von Fluoreszenzfarbstoffen mit verschiedenem Bindungsverhalten sowie
unterschiedlichem Emissionsspektrum können die Defekte der Plasma- und
Akrosommembranen gleichzeitig sichtbar gemacht und unterschieden werden. Der
rot fluoreszierende Farbstoff Propidiumiodid dringt bei defekter Plasmamembran in
die Zelle ein und bindet an die DNA; das grün fluoreszierende FITC-PNA zeigt
Schäden am Akrosom durch das Binden an die Innenseite der äußeren
Akrosomenmembran auf.

Material und Methoden
41
3.10.2 Gerät und Farbstoffe
Die Analyse der Membranintegrität der Samenzellen fand am Durchflusszytometer
„Dako Galaxy“ (Fa. Dako, Hamburg) statt. Die Steuerung erfolgte durch die Software
„FloMax“ 2.0 (Fa. Partec, Münster). Zur Ausstattung gehörten ein Argonionenlaser
(488 nm; 20 mW) zur Anregung der Farbstoffe, sowie drei verschiedene Filter: Die
Detektion von grünem Fluoreszenzlicht fand im Kanal FL-1 (537,5/22,5 nm) und
oranges Licht im Kanal FL-2 (590/25 nm) statt. Rotes Licht wurde im Kanal FL-3
durch einen Longpassfilter (630 nm LP) detektiert, blaues Licht im Kanal FL-4
(465 nm BP).
Eine weitere Anregung erfolgt durch eine HBO Quecksilberlampe (300 bis 400 nm,
100 W). Im Strahlengang sind zur Selektion der Wellenlängen zwei Filter
hintereinander geschaltet. Der erste Filter lässt Licht mit einer Wellenlänge zwischen
300 und 700 nm passieren, der zweite selektiert auf eine Wellenlänge zwischen
270 nm und 405 nm, der Hauptemissionspeak liegt hier bei 365 nm.
Die Differenzierung von DNA-haltigen Partikeln (Spermien) und Detritus wurde mit
dem membranpermeablen Fluoreszenzfarbstoff Hoechst 33342 erreicht, welcher
durch Bindung an die DNA alle in der Probe enthaltenen Zellen blau färbte. Das nicht
membrangängige, rot fluoreszierende Propidiumiodid (PI, Fa. Sigma-Aldrich,
Steinheim) dagegen kann nur bei defekter Plasmamembran in die Zelle eindringen
und an die DNA binden. Zur Darstellung von Samenzellen mit geschädigter äußerer
Akrosomenmembran wurde Peanut-Agglutinin eingesetzt, welches an
Fluoreszeinisothiocyanat (FITC-PNA, Axxora GmbH, Lörrach), gekoppelt ist. FITC-
PNA bindet spezifisch an β-Galaktosereste an der Innenseite der äußeren
Akrosomenmembran. Im Falle eines grünen Fluoreszenzsignals ist damit von einer
Schädigung der äußeren Akrosomenmembran auszugehen.
3.10.3 Grundeinstellungen
Um die Ausrichtung des Lasers, der UV-Lampe und der optischen Bank zu
überprüfen und ggf. einzustellen, wurde zu Beginn des Versuchstags mit

Material und Methoden
42
fluoreszierenden Beads (Calibration Bead 3 µm, Fa. Partec, Münster) und UV-Beads
(DNA control UV, Fa. Partec, Münster) das Durchflusszytometer kalibriert. Als
Sheathlösung diente in 5 l Aqua dest. gelöstes Sodium Azide (500 mg, Partec
GmbH, Münster) und Tween® 20 (500 µl, Partec GmbH, Münster) (3 - 4 mOsmol,
pH Wert: 6,0 – 6,5; Raumtemperatur). Die Sheathlösung wurde bei Raumtemperatur
gelagert und verwendet.
Vor der ersten Messung wurden außerdem die Grundeinstellungen der
Signalverstärkungen festgelegt, die dann für alle Messungen des Versuchstages
beibehalten wurden.
Für die Grundeinstellungen sowie die späteren Messungen setzten sich die Proben
folgendermaßen zusammen:
980 µl HBS
5 µl Propidiumiodid (1 mg/ml)
5 µl FITC PNA (600 µg/ ml)
5 µl Hoechst 33342 (150 µl/ ml)
5 µl Samenprobe (20 x 106 Spermien/ ml)
Die Messungen erfolgten nach 5 min Inkubation der Probe auf einer Wärmeplatte bei
38 °C (Heizplatte, HT 300, Fa. Minitüb, Tiefenbach).
Als Maß für die Partikelgröße wurde die Signalverstärkung der Vorwärtsstreuung
(FSC = forward scatter) und die Seitwärtsstreuung (SSC = sideward scatter) jeweils
auf linearen Skalen dargestellt. Die im Histogramm des FSC entstandene
glockenförmige Kurve repräsentierte die Partikelgröße. Ereignisse, die links davon
lagen, wurden von der Analyse ausgeschlossen, da es sich hierbei um Debris und
Schmutzpartikel handelte. In einem Punktewolkendiagramm (dotplot) wurden FSC
(Abszisse) und SSC (Ordinate) gegeneinander aufgetragen. Die Signale der
Spermien stellten sich als eine Dreiecks- bzw. „L“-Form dar.
Um die Verstärkung der Fluoreszenzkanäle festlegen zu können, wurden die
Signalverstärkungen während der Messungen so eingestellt, dass bei Kanal FL-1
und FL-3 zwei Populationen zu differenzieren waren. Hierfür wurde auf der
logarithmischen Abszisse die durchschnittliche Fluoreszenzintensität der PI-
negativen bzw. FITC-PNA-negativen Spermien so verschoben, dass sie mittig über

Material und Methoden
43
dem Wert 1 lag. Die PI- bzw. FITC-PNA-positive Population zeigte in der Regel eine
um den Faktor 10 erhöhte Fluoreszenzintensität. Im Kanal FL-4 wurde das Signal auf
der logarithmischen Skala so verstärkt, dass die Hoechst 33342-positiven Ereignisse
zwischen der zweiten und dritten Dekade dargestellt wurden. An jedem Versuchstag
wurden die Grundeinstellungen überprüft und, im Falle einer Abweichung der
Signaldarstellung, die Signalverstärkungen neu justiert.
3.10.4 Messung und Auswertung
Alle Messungen wurden mit der oben genannten Dreifachfärbung in einem
abgedunkelten Raum durchgeführt. Jeweils 880 µl eines Mastermix aus HBS und
Farbstoffen wurden in 3,5 ml Reagenzröhrchen (Fa. Sarstedt, Nümbrecht) aliquotiert,
verschlossen und im Wärmeschrank auf 38 °C angewärmt. Zum Zeitpunkt der
Messung wurden 5 µl der zu messenden Spermaprobe in die angewärmte Lösung
pipettiert und mit dem Vortex gemischt. Nach 5 min Inkubation auf einer Wärmeplatte
bei 38 °C wurde, nach erneutem Mischen, die Analyse gestartet. Je Spermienprobe
wurden 10.000 Ereignisse bei einer Flussrate von 400 – 600 Ereignissen/ Sekunde
erfasst.
Für die Auswertung wurden zunächst verschiedene Grenzlinien („Ranges“)
verwendet, um die Messereignisse zu selektieren, die für eine Auswertung relevant
sind. Die erste Range (RN1) im FSC-Kanal grenzte die Ereignisse ein, die der Größe
von Eberspermien entsprachen (Abszisse, lineare Skala, Bereich von 0 – 150). Die
zweite Range (RN2) definierte im Kanal FL-4- die DNA-haltigen Messereignisse. Aus
RN1 und RN2 wurde ein logisches Gate G1 (RN1 and RN2 = G1) gebildet. Dieses
wurde auf alle Histogramme gelegt, so dass nur noch Messereignisse angezeigt
wurden, die der Größe von Spermien entsprachen und DNA enthielten.
Für die Auswertung wurden Messergebnisse aus den Kanälen FL-1 (PNA-FITC) und
FL-3 (PI) in einem Histogramm als Dotplot gegenübergestellt. Um eine Überlagerung
der Emmissionsspektren der Farbstoffe zu korrigieren, wurde eine nachträgliche
mathematische Kompensation durchgeführt.
Durch eine vertikale und horizontale Grenzlinie (Gate) wurde das Dotplot in vier
Quadranten (Q1 – Q4) aufgeteilt. Q1 entsprach dem prozentualen Anteil PI-positiver

Material und Methoden
44
und FITC-PNA-negativer, Q2 definierte PI- und FITC-PNA positive Ereignisse. In Q3
wurden Zellen mit intakter Plasma- und Akrosomenmembran (PI- und FITC-PNA
negative Zellen) bestimmt und im vierten Quadranten (Q4) PI-negative und FITC-
PNA-positive Zellen. Soweit möglich, wurden für den gesamten Versuchszeitraum
ein einheitliches „Gate“ und identische Einstellungen für die Kompensation
beibehalten.
3.11 Mitochondrienmembranpotential in der Durchflusszytometrie
3.11.1 Testprinzip
Das Prinzip dieser Messung beruht auf den Eigenschaften des Farbstoffes 5,5',6,6'-
Tetrachloro-1,1',3,3'-tetraethylbenzimidazolylcarbo-cyanine iodide (JC-1, Fa. Enzo
Lifescience, Lörrach; Stammlösung 1,53 mM), der sich, abhängig vom
elektrochemischen Gradient der Mitochondrienmembran, reversibel vom Monomer
zu einer aggregierten Form transformiert und dabei seine Fluoreszenzeigenschaften
verändert. Als grün fluoreszierendes Monomer färbt er das Zellplasma grün, bei
steigendem Mitochondrienmembranpotential lagert der Farbstoff sich dort an und
bildet orange fluoreszierende Aggregate. Eine zusätzliche Färbung mit dem Farbstoff
Propidiumiodid (PI, Kapitel 3.8.2) ermöglicht die zusätzliche Differenzierung von
lebenden und toten Spermien.
3.11.2 Gerät und Farbstoffe
Für die Analyse wurde das unter 3.10.2 beschriebene Gerät verwendet.
Die Abgrenzung von Zellen zu anderen Partikeln erfolgte mit Hoechst 33342
(Stammlösung 150 µl/ ml), die Differenzierung in plasmamembranintakte
und -defekte Samenzellen erfolgte mit Propidiumiodid (PI, Stammlösung 1 mg/ ml).
Die detaillierten Eigenschaften dieser Farbstoffe wurden bereits in Kapitel 3.10.2
beschrieben. Der lipophile, kationische Farbstoff 5,5',6,6'-Tetrachloro-1,1',3,3'-
tetraethylbenzimidazolylcarbo-cyanine iodide (JC-1, Fa. Enzo Lifescience, Lörrach;
Stammlösung in DMSO: 1,53 mM/ l) passiert sowohl die Plasma- als auch

Material und Methoden
45
Mitochondrienmembran. Je nach dem vorliegenden Gradienten der
Mitochondrienmembran liegt JC-1 als Monomer (bei niedrigem Gradienten) oder in
aggregierter Form (bei hohem Mitochondrienmembranpotential) vor. Die beiden
Formen haben unterschiedliche Emmisionsspektra: das Monomer fluoresziert grün
(Emissionsmaximum: 527 nm), während die aggregierte Form eine
Orangefluoreszenz (590 nm) aufweist.
3.11.3 Grundeinstellungen
Am ersten Versuchstag wurden die Grundeinstellungen für die Verstärkung von FSC
und SSC sowie der Fluoreszenzsignale von PI (Rotfluoreszenz, Kanal FL-3) und
Hoechst 33342 (Blaufluoreszenz, Kanal FL-4) wie in Kapitel 3.10.3 beschrieben
vorgenommen. Für das Einstellen der Verstärkung von JC-1, wurde zunächst 1 ml
einer Spermaprobe bei -20 °C eingefroren und wieder aufgetaut, um Spermien mit
einen niedrigen Mitochondrienmembranpotential zu erhalten. Diese Probe wurde mit
1 µl JC-1 vermischt und 15 Min. bei 38 °C im Wasserbad inkubiert. Nach erneutem
Mischen wurden 5 µl der Probe in 995 µl auf 38 °C angewärmtes HBS pipettiert,
vermischt und die Messung gestartet. Die Verstärkung für das FL-2-Signal wurde so
reguliert, dass es in der ersten Dekade einer logarithmischen Skala abgebildet
wurde, das Signal von FL-1 in der zweiten Dekade. Danach wurde 1 ml einer frischen
Samenprobe mit 1 µl JC-1 vermischt und nach der Inkubationszeit wieder 5 µl in HBS
überführt und gemessen. Die Fluoreszenzsignale sollten hier weiterhin deutlich
innerhalb der Grenzen der Histogramme liegen. Für die Feinjustierung der
Signalverstärkungen wurde eine Färbung mit allen drei Farbstoffen verwendet.
Neben den Histogrammen für die einzelnen Fluoreszenzkanäle wurde zusätzlich ein
Dotplot verwendet, in dem FL-1 gegen FL-2 (Grün- gegen Orangefluoreszenz)
aufgetragen wurden. Die mathematische Kompensation für die Überlagerung der
Farbspektren wurde vor Beginn des Versuchs festgelegt. Diese Grundeinstellungen
wurden gespeichert und für die gesamte Versuchsdauer verwendet.

Material und Methoden
46
3.11.4 Messung und Auswertung
Alle Arbeitsschritte wurden in einem abgedunkelten Raum durchgeführt. Im Vorfeld
der Messungen wurden entsprechend der Anzahl Proben je ein 3,5 ml
Reagenzröhrchen mit 995 µl HBS vorbereitet. Diese wurden verschlossen in einem
Metallblock im Wärmeschrank auf 38 °C erwärmt. Zu Beginn der Untersuchung
wurde 1 ml der Samenprobe mit 1 µl JC-1, 10 µl Hoechst 33342 und 20 µl PI
vermischt und offen für 15 min bei 38 °C inkubiert. Anschließend, nach erneutem
Mischen mit dem Vortex, wurden 5 µl des gefärbten Spermas in die vorgewärmte
HBS Lösung pipettiert, gemischt und die Messung gestartet. Pro Messung wurden
10.000 Ereignisse erfasst.
Für die Auswertung wurden wie in Kapitel 3.8.4 Ranges gebildet, um die
auszuwertenden Messereignisse zu definieren. Im Histogramm des FL-4 Kanals
wurde die erste Range (RN1) gebildet, um die DNA-haltigen Ereignisse darzustellen.
Die zweite Range (RN1) im FSC-Kanal grenzte die Ereignisse ein, die der Größe von
Eberspermien entspricht (Abszisse, lineare Skala, Bereich von 0 – 150). Aus RN1
und RN2 wurde ein logisches Gate G1 (RN1 AND RN2 = G1) gebildet, und auf das
Histogramm des FL-3 Kanal gelegt. So wurden dort nur noch Messereignisse
gezeigt, die der Größe von Spermien entsprachen und DNA enthielten. PI – positive
Messereignisse wurden mit der dritten Range definiert (RN3).
Der prozentuale Anteil der Samenzellen mit hohem Mitochondrienmembranpotential
konnte im Dotplot FL-1/ FL-2 abgelesen werden. Für die Auswertung wurde mittels
einer horizontalen Linie in diesem Histogramm eine Trennung der Samenzellen mit
mehrheitlich hohem Mitochondrienmembranpotential (Q2), diese lagen über der
Linie, von denen mit niedrigem Potential ermöglicht (Q4). Durch ein zweites
logisches Gate G2 (RN1 AND RN2 AND RN3 = G2) wurden hier nur Ereignisse
berücksichtigt, die der Größe von Spermien entsprachen, DNA enthielten und eine
intakte Plasmamembran aufwiesen.
Außerdem konnte die mittlere Fluoreszenzintensität aller Spermien ermittelt werden.
Dafür wurden im Histogramm des Kanals FL-2 die Messwerte für DNA-haltige
Partikel mit der Größe von Spermien (G1) dargestellt. Eine weitere Range (RN4)

Material und Methoden
47
wurde über die gesamte Breite der Abszisse des FL-2 Histogramm gesetzt und das
arithmetische Mittel der Fluoreszenzintensität abgelesen.
Das beschriebene Gating wurde für alle Proben verwendet.
3.12 Kinetik des Calcium-Influx
3.12.1 Testprinzip
Der Assay beruht auf der durchflusszytometrischen Erfassung der Veränderungen
des intrazellulären Calciumgehaltes von Spermien unter in vitro-
Kapazitationsbedingungen. Der Anstieg der intrazellulären Calciumionen-
konzentration dient hier als Indikator für die Reaktion auf kapazitationsinduzierende
Stimuli. Die Veränderung der intrazellulären Calciumionenkonzentration wird mit Hilfe
eines Farbstoffs ermittelt, der an Calciumionen bindet. Die Fluoreszenzintensität der
Zellen ist dabei direkt mit dem freien Calciumgehalt der Zellen korreliert (HENNING
et al., submitted).
3.12.2 Gerät und Farbstoffe
Für die Analyse wurde das in 3.10.2 beschriebene Gerät verwendet.
Der intrazelluläre Calciumgehalt wurde mit Hilfe des Farbstoffs Fluo-3/ AM (Fa.
Biomol, Hamburg) ermittelt. Dieser Farbstoff bindet selektiv an Calciumionen. Durch
den Acetoxymethylester (AM) ist er auch bei Zellen mit intakter Plasmamembran
membrangängig. Intrazelluläre Esterasen spalten den Ester ab, so dass das
verbleibende Fluo-3 nicht mehr membrangängig ist und in der Zelle verbleibt. Fluo-3
bildet mit Calcium einen Komplex, der durch den Laser des Durchflusszytometers
angeregt grünes Fluoreszenzlicht emittiert (Emissionsmaximum bei 526 nm). Im
Kanal FL-1 wurde das Signal detektiert und auf einer logarithmischen Skala
dargestellt. Dabei wird die Fluoreszenzintensität von der Menge des
aufgenommenen Calciums bestimmt.

Material und Methoden
48
Die Abgrenzung von Zellen zu anderen Partikeln erfolgte mit Hoechst 33342
(Stammlösung 300 µl/ ml), die Differenzierung in plasmamembranintakte
und -defekte Samenzellen erfolgte mit Propidiumiodid (PI, Stammlösung 1 mg/ ml).
Die Eigenschaften dieser Farbstoffe wurden bereits detailliert in Kapitel 3.10.2
beschrieben.
3.12.3 Vorbereitung der Medien und Spermien
Am Versuchstag wurden von den unter Kapitel 9.3 beschriebenen Tyrodemedien
jeweils 100 ml hergestellt. Dafür wurden den eingefrorenen Tyrode-Stammlösungen
Na-Laktat, Na-Pyruvat, Gentamycinsulfat und Phenolrot frisch hinzugefügt und das
Volumen auf 100 ml mit destilliertem Wasser ergänzt. Der pH-Wert wurde auf 7,6 bei
Raumtemperatur für das Kapazitationsmedium (TyrBikCa) und auf 7,55 für das nicht
kapazitierende Medium (TyrCa) eingestellt und die Osmolarität überprüft
(300 ± 5 mOsmol/ kg). Schließlich wurden die Lösungen über Spritzenfilter (PES
Membran, Porendurchmesser 0,22 µm, Fa. Roth, Karlsruhe) filtriert. In beiden
Medien wurde nun 3 mg/ ml BSA (Albumin bovine Fraction V, Serva, Heidelberg)
gelöst.
In je 20 ml der beiden Medien wurde 5 µl/ ml der PI-Stammlösung zugesetzt,
vermischt und jeweils 995 µl in 3,5 ml Reagenzröhrchen aliquotiert. Anschließend
wurde die bikarbonathaltige Lösung (TyrBikCa) in einem CO2-Inkubator (100 %
Luftfeuchtigkeit, 5 % CO2, 38 °C) auf 38 °C erwärmt. Das nicht kapazitierende
Medium (TyrCa) wurde mit Parafilm luftdicht verschlossen und im Wärmeschrank auf
dieselbe Temperatur erhitzt. Als Sheath-Lösung wurden zwei Liter HBS-Lösung
vorbereitet (300 ± 5 mOsmol, pH 7,55, Raumtemperatur) und im Sheath-Kanister auf
einer Heizplatte auf 38 °C vorgeheizt.
Um die Spermien mit Fluo-3/AM zu beladen, wurden jeweils 2 ml der verdünnten
Samenprobe mit 2 µl der Fluo-3/AM-Stammlösung (1 mM in DMSO) und
5 µl Hoechst 33342 vermischt und 15 min unter Lichtauschluss inkubiert. Erneutes
Mischen und weitere 15 min Inkubation folgten.

Material und Methoden
49
3.12.3.1 Messung und Auswertung
Die Grundeinstellungen für die Verstärkungen wurden am ersten Versuchstag
festgelegt. An jedem Untersuchungstag wurden diese überprüft und falls notwendig
angepasst. Für die Einstellungen wurden die mit Farbstoff beladenen Spermien
verwendet und dabei auch die erfolgreiche Beladung mit Fluo-3/ AM überprüft.
Vorbereitet wurden folgende Ansätze:
995 µl TyrCa (mit PI) + 5 µl verdünntes Sperma
993,5 µl TyrCa (ohne PI) + 5 µl verdünntes Sperma
993,5 µl TyrCa (ohne PI) + 5 µl verdünntes Sperma + 1,5 µl A23187-Stammlösung
993,5 µl TyrCa (mit PI) + 5 µl verdünntes Sperma + 1,5 µl A23187-Stammlösung
Die Konzentration der Stammlösung des Calciumionophors A23187 (Fa. Alexis,
Lausen, Schweiz) betrug 500 mM/ ml, die Endkonzentration in den Ansätzen lag
damit bei 2 µM/ ml. Für den zweiten Ansatz wurde TyrCa verwendet, die kein
Propidiumiodid enthielt. Ein Aliquot für die Grundeinstellungen wurde vor der oben
genannten Färbung separiert und bei 38 °C erwärmt.
Die Verstärkungen für FSC, SSC, FL-3 und FL-4 wurden mit dem ersten Ansatz
festgelegt. Mit dem zweiten Ansatz wurde die Verstärkung für FL-1 ermittelt. Die
Festlegung der Verstärkung erfolgte wie im Kapitel 3.10.3 angegeben. Der dritte
Ansatz diente der Prüfung, ob die Beladung mit Fluo-3/ AM erfolgreich war. Das
Calciumionophor im Ansatz Nummer drei bewirkte einen Permeabilitätsanstieg der
Plasmamembran. Das Calcium aus dem Medium kann so ins Zytosol einströmen und
verursacht bei erfolgreicher Beladung mit Fluo-3/ AM eine deutliche Verschiebung
der Fluoreszenzintensität nach rechts. Der vierte Ansatz diente zur Feinjustierung der
Einstellungen.
5 µl der vorbehandelten Spermiensuspension wurden pro Ejakulat, Medium und
Inkubationszeitpunkt in die vorbereiten Reagenzröhrchen pipettiert. Die Messungen
erfolgten bei beiden Medien (TyrBikCa und TyrCa) nach 3 und nach 60 min. Von jeder
Probe wurden 10.000 Ereignisse mit einer Flussrate von 400 – 600 Ereignissen/ s

Material und Methoden
50
gemessen. Als Sheath-Lösung und Spüllösung zwischen den Messungen diente das
auf 38 °C erwärmte HBS.
Im FSC-Histogramm wurde mit Grenzlinien („range“) Agglutinationen und Debris
abgetrennt, die in den Histogrammen nach drei Minuten Inkubationsdauer festgelegt
wurden. Eine zweite Range definierte im Kanal FL-4 die DNA-haltigen Ereignisse. In
die Berechnungen wurden damit nur die Ereignisse der Spermienpopulation
miteinbezogen. Anhand der Diagramme nach 3 min Inkubation wurden die Grenzen
zwischen PI-negativen und –positiven sowie Spermien mit geringem intrazellulären
Calciumgehalt (Fluo-3-negativ) und hohem intrazellulärem Calciumgehalt (Fluo-3–
positiv) festgelegt. Die Grenzen wurden mit vertikalen und horizontalen Linien in den
FL-3/ FL-1 Dotplot eingezogen und so die prozentuale Verteilung auf die einzelnen
Quadranten ermöglicht. Q1 entsprach dem prozentualen Anteil PI-positiver und Fluo-
3-negativer, Q2 definierte PI- und Fluo3-positive Ereignisse. In Q3 wurden PI- und
Fluo-3-negative Zellen bestimmt und im vierten Quadranten (Q4) PI-negative und
Fluo3-positive Zellen.
Die Kompensation wurde anhand der Proben für die Grundeinstellung und
Überprüfung der Indikator-Beladung vorgenommen.
Die Reaktivität auf die Inkubationsbedingungen wurde nach der Methode von
HENNING et al. (2012) bestimmt. Es wurde in der Population der lebenden Zellen mit
basalem intrazellulären Calciumgehalt (PI-neg./ Fluo-3 neg.) die Differenz
zwischen 60 min und dem Anfangszeitpunkt der Inkubation (3 min) bestimmt. Die
Reaktivität im nicht kapazitierenden Medium wurde ebenso berechnet. Um die
spezifische Reakivität zu ermitteln, wurde die Differenz aus der Reaktivität im
Tyrode- und im Kontrollmedium in der Population der lebenden Spermien mit
niedrigem intrazellulären Calciumgehalt (PI-neg./ Fluo-3 neg.) gebildet.

Material und Methoden
51
3.13 Bestimmung des ATP-Gehalts und der Energieladung
3.13.1 Testprinzip
Die chemische Energie, die für die Motilität der Spermien erforderlich ist, wird in
Form von Adenosintriphosphat (ATP) bereitgestellt.
Der Test ermöglicht mittels der Firefly-Luciferin-Luciferase-Reaktion die qualitative
Bestimmung von ATP. Die Analyse beruht auf der Eigenschaft des Enzyms
Luciferase bei der Reaktion mit ATP, Luciferin und Sauerstoff, Licht zu emittieren,
welches photometrisch detektiert werden kann.
Luciferin + ATP + O2 Luciferase→ Oxyluciferin + AMP + PPi + CO2 + Licht
Unter definierten Bedingungen ist die Lichtintensität proportional zur ATP
Konzentration.
Nach der Asservierung der Proben (Kapitel 3.13.3.1) wurde deren ATP-Gehalt
gemessen, dann wurden mit Hilfe von Enzymen ADP und AMP in mehreren Schritten
in ATP umgewandelt, um sie dann mit dem oben genannten ATP-Assay analysieren
zu können. Durch Subtraktion des zuerst gemessenen ATP-Gehalts von diesen
Ergebnissen war es möglich die Menge jedes Nukleotids in der Probe zu bestimmen.
Die einzelnen Schritte der Umwandlung sind in den folgenden Gleichungen
dargestellt:
ADP + Phosphoenolpyruvat Pyruvatkinase→ ATP + Pyruvat
AMP + ATP Adenylatkinase→ 2 ADP
Das Konzept für die Ermittlung der Energieladung („Energy Charge“, EC) der
Samenzelle beruht auf dem Vergleich des Nukleotidgehalts (ATP, ADP, AMP) einer
Zelle, mit einem Akku bzw. einer Batterie. Die gemeinsamen Elemente umfassen: (1)
Die Gesamtmenge an Material bleibt in der Regel konstant, und (2) chemische
Energie kann durch Veränderung der Verhältnisse der Komponenten gespeichert
und wiedergewonnen werden (FORD u. LEACH, 1998).

Material und Methoden
52
Die Energieladung errechnet sich durch die Summe von ATP (mol) plus die Hälfte
der ADP-Fraktion. Dies wird dividiert durch die Gesamtsumme aller Nukleotide. Die
Formel wurde in Kapitel 2.3.1 beschrieben.
Es handelt sich hier um ein lineares Maß der Stoffwechselenergie, die im Adenin-
Nukleotidsystem gespeichert ist. Wenn nur AMP vorhanden ist, ist die Energieladung
Null. Ist ATP als einziges Nukleotid verfügbar, entspricht das der Energieladung 1.
Bei Werten > 0,6 wird von vitalen Zellen gesprochen. Während des Zellwachstums
und der Zellteilung beträgt die Energieladung 0,8 – 0,95, in einer stationären
Wachstumsphase ist die Ladung 0,6. Ruhende bzw. alternde Zellen zeigen eine
Energieladung von < 0,5 (WIEBE u. BANCROFT, 1975).
3.13.2 Geräte und Vorbereitung
Die Bestimmung des ATP-Gehalts der Probe wurde modifiziert nach LONG und
GUTHRIE (2006) mittels ATP-Bioluminescent- Assay-Kit (FLAA, Fa. Sigma Aldrich,
Steinheim) durchgeführt. Dieser beinhaltete den „Dilution buffer“, „Assay-Mix“ sowie
den „ATP-Standard“. Die Lösungen wurden nach Herstellerangaben aufbereitet. Der
„Dilution buffer“ ist ein portioniertes Lyophilisat aus MgSO4, DTT, EDTA, bovinem
Serumalbumin (BSA) und Tricin, das mit 50 ml sterilem Wasser (Ampuwa®, Fa.
Fresenius, Bad Homburg) in Lösung gebracht wird. Diese war bis zu zwei Wochen
bei 0 - 5 °C stabil. Beim „Assay-Mix“ handelt es sich ebenfalls um ein portioniertes
gefriergetrocknetes Pulver; es enthält Luciferase, Luciferin, MgSO4, DTT, EDTA,
BSA und Tricin und wurde mit 5 ml sterilem Wasser gelöst. Die Wasserqualität ist
hier von hoher Bedeutung, da schon eine minimale Kontamination eine hohe
Hintergrundlumineszenz verursachen kann. Diese Lösung ist unter Lichtauschluss
ebenfalls mindestens zwei Wochen bei 0 - 5 °C stabil. Da eine lagerungsbedingte
Abnahme der Lumineszenz nicht auszuschließen ist, wurde an jedem Versuchstag
eine neue Standardkurve erstellt. Der „Assay-Mix“ wurde 1:25 (v:v) mit dem „Dilution
buffer“ verdünnt und dunkel bei 4 °C gelagert.
Für die Konvertierung der Nukleotide wurden Adenylatkinase (AK; M 3003, Sigma-
Aldrich, Steinheim) und Pyruvat-Kinase (PK; P1506, Sigma-Aldrich, Steinheim)
verwendet. Beide lagen als Suspension in (NH4)2SO4 vor. Um das Ammoniumsulfat

Material und Methoden
53
zu eliminieren, wurden die Suspensionen für 5 min bei 1000 g zentrifugiert. Das
dabei entstehende Pellet wurde in 20 mM Tricin, pH 7,5, incl. 0,1% BSA (Albumin
bovine Fraction V, Serva, Heidelberg) gelöst. Es wurde jeweils nur die für den
Versuchstag benötigte Menge Enzym vorbereitet.
Für den Reaktionspuffer A wurde zunächst ein zehnfach konzentrierter Puffer
hergestellt (750 mM Tricin, 50 mM MgCl2 und 0,125 mM KCl, pH 7,5). Eine 1:10
Verdünnung mit sterilem Wasser ergab den Puffer A (75 mM Tricin, 5 mM MgCl2
und 0,0125 mM KCl, pH 7.5). Puffer B enthielt neben dem Inhalt von Puffer A 0,5 mM
Phosphoenolpyruvat (PEP) (P7022, Sigma- Aldrich, Steinheim) und 0,4 µg/ µl
Pyruvatkinase. Schließlich wurde aus dem Puffer B durch Zugabe von 0,5 µg/ µl
Adenylatkinase der Reaktionspuffer C hergestellt. Wie oben beschrieben, konvertiert
Adenylatkinase AMP um zu ADP; Phosphoenolpyruvat sowie Pyruvatkinase sind für
die Umwandlung von ADP zu ATP zuständig.
Für die Detektion der Lumineszenz stand ein Tecan GENios Pro plate reader (Tecan
Group Ltd, Männedorf, Schweiz) in der Abteilung Endokrinologie der Klinik für Rinder
zur Verfügung. Die Verarbeitung der Daten fand mit der Software MagellanTM
(Version V5.03, Tecan Group Ltd, Männedorf, Schweiz) statt.
3.13.3 Messung und Auswertung
Die Analyse der Nukleotide erfolgte in sechs Schritten:
Präparation der Spermien für die Lagerung bei -20°C
Erstellen der Standardkurve für die ATP - Messung
Extraktion von ATP aus der gefrorenen Probe
Messung der Lumineszenz und Berechnung der ATP - Konzentrationen
Konvertieren der Nukleotide und deren Bestimmung
- ATP - Bestimmung
- Konvertierung von ADP in ATP und Messung des ATP - Gehaltes
- Konvertierung von AMP in ATP und Messung des ATP - Gehaltes
Berechnung der Energieladung

Material und Methoden
54
3.13.3.1 Präparation der Spermien für die Lagerung bei -20 °C
Um die Vergleichbarkeit der Ergebnisse des ATP-Assays zu gewährleisten, wurde
angestrebt alle Proben eines Versuchdurchgangs an einem Tag zu messen. Um dies
zu gewährleisten, wurden die Proben bis zur ATP-Analyse bei -20 °C gelagert. Um
den Abbau von ATP während der Lagerung zu verhindern, wurden 100 µl jeder zu
untersuchenden Probe (entspricht 2 x 106 Spermien) mit 1 µl Phosphatase Inhibitor
Cocktail 2 (P5725, Fa. Sigma Aldrich, Steinheim) versetzt, gemischt und 30 min auf
Eis inkubiert. Anschließend wurden die Proben bei -20 °C eingefroren und gelagert.
Erstellung und Verifizierung des Protokolls erfolgte in einem parallel laufenden
Projekt (NGUYEN et al., unveröffentlicht).
3.13.3.2 Erstellen der Standardkurve für die ATP-Messung
Zunächst wurde eine Serienverdünnung des ATP-Standards mit sterilem Wasser
hergestellt, um folgende Konzentrationen zu erhalten: 62,5, 125, 250, 500, 1000 und
2000 pmol ATP/ ml. Für die Ermittlung der Standardkurve wurden je 25 µl der
Standardkonzentrationen sowie eine Kontrolle (steriles Wasser) in die Wells einer 96
Well Mikrotiter Platte (white wall, clear bottom, Fa. Greiner Bio-One, Frickenhausen)
pipettiert. Alle Proben wurden im Doppelansatz gemessen. Um die Reaktion zu
starten, wurde mit Hilfe einer elektronischen Pipette (Biohit eLine E 1000, Biohit,
Rosbach, Germany) in jedes Well 100 µl des verdünnten „Assay-Mix“ pipettiert. Die
Detektion der Lumineszenz im Plattenphotometer (100 ms/ well) schloss sich
unmittelbar an. Die Ergebnisse der Doppelansätze wurden jeweils gemittelt und die
relativen Lichteinheiten (relative light units, RLU) der Kontrolle wurden von den
Ergebnissen der Standards subtrahiert. Die so ermittelten Ergebnisse entsprachen
dem ATP-Gehalt der Standards. Es wurde eine lineare Regressionslinie erstellt und
die dazugehörige Gleichung zur Berechnung der Menge an ATP in den Proben
verwendet (Abb. 10).

Material und Methoden
55
Abb. 10: Beispiel einer Standardkurve des ATP-Assays
3.13.3.3 Extraktion von ATP aus der gefrorenen Probe
Um die Nukleotide aus den Proben zu extrahieren, wurden diese, ohne vorher
aufzutauen, mit 900 µl eines auf 95 °C erhitzten „boiling buffer“ (50 mM Tricin, 10 mM
MgSO4 und 2 mM EDTA, pH 7,80) versetzt und für 10 min bei 95°C in einem
Thermomixer 5436 (Eppendorf, Hamburg) inkubiert. Danach kühlten die Proben
10 min auf Eis ab und wurden anschließend für 30 min bei 4 °C und 5000 g
zentrifugiert. Der Überstand wurde für die weitere Bestimmung des Nukleotidgehalts
verwendet.
3.13.3.4 Messung der Lumineszenz und Berechnung der ATP-Konzentrationen
Die ATP-Messung wurde, als Doppelansatz durchgeführt. Als Null-Kontrolle diente
steriles Wasser, um die Hintergrundluminiszenz zu bestimmen und später korrigieren
zu können. Es wurden jeweils 25 µl des Überstandes einer Probe in zwei Wells
pipettiert. Zusätzlich wurde als Positivkontrolle in jedem Messdurchgang die
Lichtintensität eines ATP-Standards gemessen. Um eine zu lange Verzögerung der
photometrischen Messung zu vermeiden, die zu einer reduzierten Lichtintensität
y = 213,22x - 8890,3 R² = 0,9995
0
50000
100000
150000
200000
250000
300000
350000
400000
450000
0 500 1000 1500 2000 2500
rela
tive lig
ht
units
Standardprobe
RLU results
Linear (RLU results)

Material und Methoden
56
führen kann, wurde der verdünnte „Assay-Mix“ mit Hilfe einer elektronischen Pipette
hinzugefügt. Unmittelbar danach wurde die bei der Reaktion entstehende
Lichtintensität photometrisch detektiert. Die Ergebnisse der doppelt gemessenen
Proben wurden gemittelt und anschließend die Hintergrundlumineszenz durch
Subtraktion des Wertes der Nullkontrolle vom Probenwert korrigiert. Mit dem
Ergebnis der Standardprobe wurde ein Korrekturfaktor berechnet. Dieser ergab sich
aus der Division der RLUs der Standardprobe eines Messdurchgangs durch den
entsprechenden Messwert der Standardprobe, der der Standardkurve des
Messtages zugrunde lag. Die RLUs der Samenproben wurden dann mit dem
Korrekturfaktor multipliziert. Die ATP-Konzentrationen errechneten sich aus den
korrigierten RLUs der Proben mit Hilfe der Gleichung der linearen
Regressionsgeraden (y = ax + b) aus der Standardkurve, wobei „y“ den relative light
units und „x“ der ATP-Konzentration entsprach.
3.13.3.5 Konvertieren der Nukleotide und deren Bestimmung
Für die Ermittlung der Konzentration der einzelnen Nukleotide muss zunächst erneut
ATP in den Proben bestimmt werden (Messung 1). Zu diesem Zweck wurden je 25 µl
des Puffer A mit je 100 µl des Überstandes der ATP-Extraktion (Kapitel 3.13.3.3) in
einem Eppendorfgefäß vermischt und für 30 min bei 30°C inkubiert. Um die Reaktion
zu stoppen, wurden danach die Proben für 3 min bei 95°C erhitzt und anschließend
auf Eis abgekühlt.
Um ADP zu ATP umzuwandeln (Messung 2) wurden 25 µl Puffer B mit 100 µl des
Überstands vermischt. Das in dem Puffer enthaltene Phosphoenolpyruvat und die
Pyruvatkinase konvertierten das vorhandene ADP zu ATP. Die so vorbereitete Probe
wurde 30 Minuten bei 30 °C inkubiert, darauf ebenfalls drei Minuten bei 95 °C erhitzt
und anschließend auf Eis gekühlt, um die Reaktion zu stoppen.
25 µl Puffer C wurde mit 100 µl Probe vermischt, dieser enthielt zusätzlich zu
Phosphoenolpyruvat und Pyruvatkinase das Enzym Adenylatkinase, welches AMP
zu ADP wandelt (Messung 3). Die Inkubationszeit betrug hier 90 Minuten bei 30 °C.

Material und Methoden
57
Die Reaktion wurde ebenso durch Erhitzen der Probe für drei Minuten bei 95 °C
induziert und nachfolgendes Abkühlen gestoppt.
Nach der jeweiligen Abkühlzeit wurde die ATP-Konzentration aller Proben nach der
unter 3.13.3.4 beschriebenen Methode analysiert.
3.13.3.6 Berechnung der Energieladung
Die AMP-Konzentration wird durch Subtraktion des Ergebnisses der Messung 2 von
den Werten der Messung 3 errechnet. ADP kann analog durch Subtraktion des
Ergebnisses der Messung 1 mit dem Ergebnis von Messung 2 (ATP-Konzentration)
ermittelt werden. Die Ergebnisse wurden zur Berechnung der Energieladung
herangezogen, die mittels der unter Kapitel 2.3.1 genannten Formel durchgeführt
wurde.
3.14 Bestimmung der Gesamtkeimzahl
Die Gesamtkeimzahlen wurden kulturell, nach einer Verdünnungsreihe im
anschließenden Spatelverfahren und Bebrütung auf Blutagar im Institut für
Mikrobiologie der Tierärztlichen Hochschule Hannover bestimmt.
3.15 Statistische Auswertung
Die statistische Auswertung fand mit SAS (Version 9.2; SAS Inst. Inc., Cary, NC,
USA) und IBM SPSS Statistics 2.0 statt. Grafisch wurden, wenn nicht anders
angegeben, die Mittelwerte und Standardabweichungen dargestellt.
Nach der Prüfung der Merkmale auf Normalverteilung mit dem Shapiro-Wilk-Test
(PROC UNIVARIATE) war trotz Logarithmierung oder Quadrierung der Originalwerte
die große Mehrzahl der Daten nicht normalverteilt. Um den Einfluss der Haltezeiten,
Verdünnungsart und Lagerungsdauer überprüfen zu können, wurde der Friedman-
Test verwendet. Paarvergleiche zwischen Haltezeiten, Verdünnungsarten und

Material und Methoden
58
Lagerungszeiten wurden mittels des Wilcoxon signed rank Test für nicht
normalverteilte Daten durchgeführt. Bei einer Irrtumswahrscheinlichkeit (p) von
p < 0,05 wurden die Unterschiede zwischen den verglichenen Haltetemperaturen und
–zeiten bzw. Lagerungszeitpunkten als signifikant erachtet. Korrelationskoeffizienten
nach Spearman wurden zwischen Parametern mit der Prozedur CORR ermittelt.
Um Spermiensubpopulationen darstellen zu können, wurden die Motilitätsparameter
mit SPSS klassiert. Hierfür wurden die Ergebnisse jedes Parameters in Klassen
eingeteilt und der prozentuale Anteil jeder Klasse ermittelt.

Ergebnisse
59
4 Ergebnisse
4.1 Experiment 1: Vergleich von zweistufig isothermer und
zweistufig hypothermer Verdünnung bei zwei Verdünnungsgraden
4.1.1 Temperaturverlauf während der Samenverarbeitung
Nach der Vorverdünnung des Samens im Verhältnis 1:1 (v:v) wurde eine Hälfte
(200 ml) für die isotherme Verarbeitung im Glaszylinder für 15 min bei 32 °C
gehalten. Während der Ausverdünnung in 100 ml Ebersamenflaschen mit isothermen
Verdünner (32 °C) fiel die Temperatur von 31,6 °C auf 31,3 °C ab.
Bei der hypothermen Verarbeitung wurden die andere Hälfte der vorverdünnten
Proben (200 ml) zunächst 20 min bei 21 °C gehalten. Währenddessen sank die
Temperatur von 31,3 °C auf 29,0 °C. Bei der Ausverdünnung mit dem hypothermen
Verdünner (21 °C) sank die Temperatur der Proben um 5,6 °C von 29 °C auf 23,4 °C.
Während der folgenden 90 min wurden die Samenproben bei 21 °C
Umgebungstemperatur gehalten. Beide Kurven zeigen eine asymptotische
Annäherung an die Umgebungstemperatur. Während der ersten 30 min nach der
Verdünnung zeigten die isotherm verdünnten Proben eine durchschnittliche
Abkühlrate von 0,13 ± 0,05 °C/ min, dann 0,08 ± 0,04 °C/ min (31 - 60 min) und
schließlich 0,05 ± 0,04 °C/ min (61 - 90 min). Die Abkühlrate der hypotherm
verdünnten Proben bei 21 °C betrug 0,03 ± 0,04 °C/ min (0 – 30 min nach der
Verdünnung), 0,01 ± 0,04 °C/ min (31 – 60 min), und 0,01 ± 0,04 °C/ min (61 –
90 min). Nach 90 Minuten bei 21 °C erreichten die isotherm verdünnten Proben
23,3 °C während die hypotherm verarbeiteten Proben 21,9 °C aufwies. Nach 150 min
hatten beide Varianten die gleiche Temperatur erreicht (20,3 °C). Zwischen dem
Zeitpunkt des Umlagerns bis zur 150. Minute betrug die Abkühlrate der isotherm
verdünnten Proben 0,08 ± 0,05 °C/ min, die der hypothermen dagegen
0,05 ± 0,04 °C/ min. Nach 262 Minuten (isotherm) bzw. 292 Minuten (hypotherm)
nach der ersten Verdünnung erreichten die Proben die Lagerungstemperatur von
17,0 ± 0,5 °C.
Der Temperaturverlauf ist in Abb. 11 dargestellt

Ergebnisse
60
Abb. 11: Temperaturverlauf während der zweistufig hypothermen (blaue Linie) und
isothermen (rote Linie) Samenverarbeitung bei Experiment 1. Mittelwerte von je 3
Samenproben. Die Samenproben wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville
Thawing Solution (BTS, 32 °C) verdünnt. Für die zweistufig isotherme Verdünnung wurde
eine Hälfte des vorverdünnten Samen für ca. 15 min bei 32 °C (Wasserbad) gehalten um
dann mit BTS (32 °C) auf die Endkonzentration in Ebersamenflaschen verdünnt zu werden.
Abkühlung der Proben 90 min bei 21 °C anschließend weitere Abkühlung und Lagerung in
einen auf 17 °C temperierten Klimaschrank verbracht. Für zweistufig hypotherme
Verarbeitung wurde nach der Vorverdünnung die andere Hälfte der Probe für 20 min bei
21 °C gehalten. Die weitere Verdünnung erfolgte mit auf 21 °C temperierter BTS, wodurch
der schnelle Abfall der Probentemperatur zu erklären ist. Die weitere Verarbeitung entsprach
den zweistufig isotherm verarbeiteten Proben.
*: Wechsel der Messsonde zwischen zwei Gefäßen
10
15
20
25
30
35
0 30 60 90 120 150 180 210 240
Tem
per
atu
r °C
Minuten
zweistufig hypotherm zweistufig isotherm
verdünnt, gehalten bei 17°C verdünnt, gehalten bei 21°C 1:1 bei 32 °C
verdünnt, gehalten bei 21°C verdünnt, gehalten bei 17°C 1:1 bei 21 °C
*
9

Ergebnisse
61
4.1.2 CASA-Motilitätsparameter
Abb. 12: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Art der zweistufigen Verdünnung (isotherm vs. hypotherm), Verdünnungsgrad (20 x 10
6 und 10 x 10
6 Spermien/ ml) und
Lagerungsdauer (d 1, d 3, d 6) bei 17 °C; n = 6 Ejakulate. #: Unterschiede zw. Verdünnungsarten bei gleicher Lagerungsdauer und Verdünnungsgrad (p < 0,05) *: Unterschiede zw. Verdünnungsgraden bei gleicher Lagerungsdauer und Verdünnungsart (p < 0,05) a, b: Unterschiede zw. Lagerungstagen (d1 – d6) für eine Verarbeitungsvariante (p < 0,05)
Von der Verdünnungsart beeinflusst wurden die Gesamtmotilität, progressive
Motilität, Linearität (LIN) und Wobble (WOB). Signifikante Unterschiede konnten aber
nur bei der Gesamtmotilität ermittelt werden. Innerhalb der Samenproben mit
10 x 106 Spermien/ ml zeigte sich an d 1 ein signifikanter höherer Wert von 4,4 % bei
der Gruppe, die isotherm verdünnt wurde, im Vergleich zur hypothermen Gruppe.
Der Verdünnungsgrad hatte Einfluss auf die Gesamtmotilität, progressive Motilität,
Velocity average path (VAP), Velocity straight line (VSL), Wobble (WOB) und
Amplitude of lateral head displacement (ALH). Allerdings gab es bei VAP, VSL und
WOB keine signifikanten Unterschiede zwischen den analysierten Proben (Tabelle 1).
0
10
20
30
40
50
60
70
80
90
100
isotherm hypotherm isotherm hypotherm
motile
Sperm
ien in %
Verdünnungsart/ -stufe
d1
d3
d6
20 x 106 10 x 106
a,b b b b
b
a a,b
# * *
a a a a
b

Ergebnisse
62
Bei zweistufig hypothermer Verdünnung zeigte sich bei den Proben mit
20 x 106 Spermien/ ml an d 3 mit 86,2 ± 6,9 % eine signifikant höhere Gesamtmotilität
als bei den auf 10 x 106 Spermien/ ml Proben mit 76,5 ± 10,3 %. An d 6 gab es
signifikante Veränderungen zwischen den Verdünnungsgraden der zweistufig
isotherm verdünnten Proben. Die Gesamtmotilität betrug 77,1 ± 12,1 % bei
20 x 106 Spermien/ ml und 61,9 ± 15,0 % bei der Spermienkonzentration von
10 x 106 Spermien/ ml.
Die progressive Motilität (Tabelle 1) zeigte ebenfalls an d 6 der zweistufig isotherm
verdünnten Proben signifikante Unterschiede zwischen den Vedünnungsgraden.
Samenproben mit einer Konzentration von 20 x 106 Spermien/ ml wiesen eine
progressive Motilität von 62,4 ± 18,2 % auf, während bei den Samenproben mit
10 x 106 Spermien/ ml nur 45,8 ± 19,9 % der Samenzellen progressiv motil waren.
Alle Motilitätsparameter, mit Ausnahme von der velocity curvilinear (VCL), wurden
von der Lagerungsdauer beeinflusst (p < 0,05). Die Gesamtmotilität und progressive
Motilität verringerten sich bei allen Verarbeitungsvarianten zwischen d 0 bzw. d 1 und
d 6 (Tabelle 1). Bei den zweistufig isotherm verdünnten Samenproben sank die
Gesamtmotilität von 91,1 ± 2,2 % nach 24 h auf 86,4 ± 5,1 % bereits nach 72 h.
Innerhalb beider Verdünnungsgrade verringerte sich auch die progressive Motilität
der zweistufig isotherm verdünnten Samenproben bereits zwischen 24 h und 72 h.
Bei Proben mit einer Konzentration von 20 x 106 Spermien/ ml sank sie zwischen
Lagerungstag 1 und 3 um 11,6 %, bei den auf 10 x 106 Spermien/ ml verdünnten
Proben verringerte sie sich im gleichen Zeitraum um 16,1 %. Bei den weiteren
Motilitätsparametern war der Einfluss der Lagerung weniger deutlich ausgeprägt.
In der grafischen Darstellung (Abb. 12) wurden die Ergebnisse des
Verdünnungstages (d 0) nicht mit abgebildet, da hier nur die zweistufig isothermen
Proben analysiert wurden und dadurch kein Vergleich der Verdünnungsarten möglich
ist.

Ergebnisse
63
4.1.3 Membranintegrität
Abb. 13: Prozentualer Anteil membranintakter (PI und FITC-PNA negativer) Spermien (MW + SD) in Abhängigkeit von der Art der zweistufigen Verdünnung (isotherm vs. hypotherm), Verdünnungsgrad (20 x 10
6 und 10 x 10
6 Spermien/ ml) und Lagerungsdauer (d 1, d3, d 6) bei 17 °C; n = 6 Ejakulate.
#: Unterschiede zwischen Verdünnungsarten bei gleicher Lagerungsdauer und Verdünnungsgrad
(p < 0,05)
a, b: Unterschiede zwischen Lagerungstagen (d1 –d6) für eine Verarbeitungsvariante (p < 0,05)
Der Anteil der PI- sowie FITC-PNA-negativen (plasma- und
akrosomenmembranintakte) Spermien wurde von der Art der Verdünnung (isotherm,
bzw. hypotherm) und der Lagerungsdauer beeinflusst (p < 0,05).
In der auf 20 x 106 Spermien/ ml verdünnten Gruppe wiesen hypotherm verdünnte
Samenproben am Lagerungstag 3 signifikant geringere Prozentsätze
membranintakter Spermien als isotherm verdünnte Proben auf, wobei die mittlere
Differenz mit 1,8 % gering war. In der Versuchsgruppe mit 10 x 106 Spermien/ ml
waren zu allen Lagerungszeitpunkten signifikant geringere Prozentsätze
membranintakter Spermien in den hypotherm verdünnten Proben, verglichen mit
isotherm aufbereiteten Proben festzustellen.
0
10
20
30
40
50
60
70
80
90
100
isotherm hypotherm isotherm hypotherm
Mem
bra
nin
takt
e Sp
erm
ine
in %
Verdünnungsart/ -grad
d1
d3
d6
20 x 106 10 x 106
a a,b b a
b b a a b a a b
# #
# #

Ergebnisse
64
Während der Lagerung sanken die Werte der isotherm auf 20 x 106 Spermien/ ml
verdünnten Proben zwischen Lagerungstag 1 und 6 signifikant um 4 %. Bei der
hypothermen Verdünnung konnte ein signifikanter Abfall von 3,1 % der
membranintakten Spermien zwischen d 1 und d 3 festgestellt werden.
Bei der auf 10 x 106 Spermien/ ml verdünnten Gruppe sanken die Werte der isotherm
verdünnten Proben zwischen Lagerungstag 1 und 6 signifikant um 5,4 %. Die
hypotherm verdünnen Proben zeigten im gleichen Zeitraum einen signifikanten Abfall
von 6,5 %.
In der grafischen Darstellung (Abb. 13) wurden die Ergebnisse des
Verdünnungstages (d 0) nicht mit abgebildet, da hier nur die zweistufig isothermen
Proben analysiert wurden und dadurch kein Vergleich der Verdünnungsarten möglich
ist.

Ergebnisse
65
4.2 Experiment 2: Auswirkungen von Haltezeiten
und -temperaturen des vorverdünnten Samens bei zweistufig
isothermer Verdünnung
4.2.1 Temperaturverlauf während der Samenverarbeitung
Der Zylinder mit der 1:1 (v:v) vorverdünnten Samenprobe wurde bis zu 6 h bei 21 °C
gehalten; hier verringerte sich die Temperatur von 31,1 °C auf 23,9 °C innerhalb der
ersten 90 Minuten. Die Abkühlrate betrug 0,10 ± 0,08 °C/min (0 – 30 min), dann
0,09 ± 0,08 °C/ min (31 – 60 min) und schließlich 0,05 ± 0,06 °C/ min (61 – 90 min).
Nach der Ausverdünnung der Kontrollproben mit isothermem Verdünner wurden
diese ebenfalls für 90 min bei 21 °C Umgebungstemperatur gehalten. In diesem
Zeitraum sank die Temperatur der Samenprobe von 30,2 °C auf 23,8 °C. Die
durchschnittliche Abkühlrate der Kontrollprobe bei 21 °C während der ersten 30 min
nach der Verdünnung betrug 0,10 ± 0,07 °C/ min, dann 0,07 ± 0,06 °C/ min (31 –
60 min) und schließlich 0,04 ± 0,06 °C/min (61 – 90 min).
Beide Kurven zeigen eine asymptotische Annäherung an die Umgebungstemperatur.
Die vorverdünnte Probe im Zylinder erreichte nach 269 min 21,0 ± 0,5 °C.
Die Temperatur der gehaltenen Samenproben zum Zeitpunkt der Ausverdünnung
belief sich auf 28,2 °C (Probe mit 30 min Haltezeit), 25,3 °C (60 min Haltezeit),
22,1 °C (3 h Haltezeit) und 21,5 °C (6 h Haltezeit). Die Abkühlraten in den ersten
30 min nach der jeweiligen Verdünnung betrug 0,08 ± 0,07 °C/ min (30 min Haltezeit)
und 0,05 ± 0,05 °C/ min (60 min Haltezeit). Bei den Samenproben, welche länger als
drei Stunden gehalten wurden, war die Abkühlrate in derselben Zeitspanne
< 0,01 °C/ min. Bei der Umlagerung in den auf 17 °C temperierten Klimaschrank
konnten folgende Temperaturen gemessen werden: 23,3 °C (30 min Haltezeit),
22,4 °C (60 min Haltezeit), 21,7 °C (3 h Haltezeit) und 21,3 °C bei der für 6 h
gehaltenen Samenprobe. Die Temperatur des Klimaschranks (17,0 ± 0,5 °C) wurde
von den Kontrollproben nach 273 min erreicht. Die für 30 min und 60 min gehaltenen
Proben erreichten sie fast zeitgleich (294 bzw. 297 min). Die für 3 h gehaltenen
Proben erreichten sie nach 423 min und die Probe mit der längsten Haltezeit (6 h)
erreichte die Lagerungstemperatur nach 576 min.
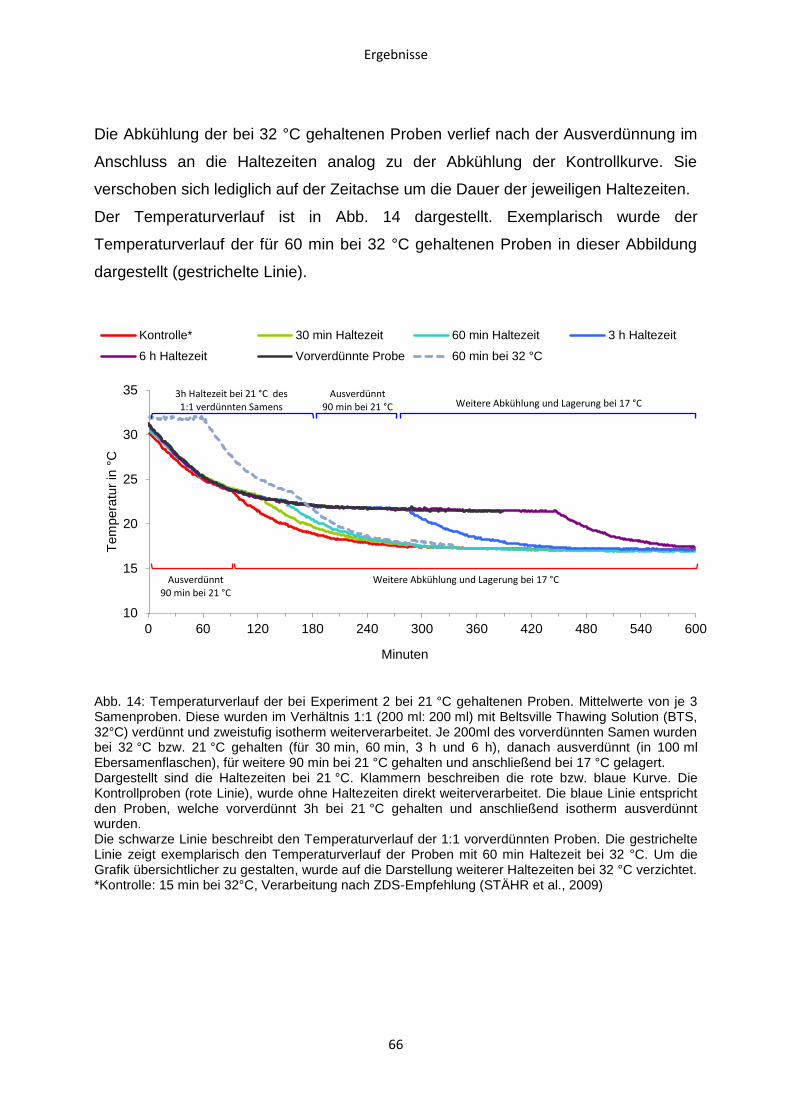
Ergebnisse
66
Die Abkühlung der bei 32 °C gehaltenen Proben verlief nach der Ausverdünnung im
Anschluss an die Haltezeiten analog zu der Abkühlung der Kontrollkurve. Sie
verschoben sich lediglich auf der Zeitachse um die Dauer der jeweiligen Haltezeiten.
Der Temperaturverlauf ist in Abb. 14 dargestellt. Exemplarisch wurde der
Temperaturverlauf der für 60 min bei 32 °C gehaltenen Proben in dieser Abbildung
dargestellt (gestrichelte Linie).
Abb. 14: Temperaturverlauf der bei Experiment 2 bei 21 °C gehaltenen Proben. Mittelwerte von je 3 Samenproben. Diese wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville Thawing Solution (BTS, 32°C) verdünnt und zweistufig isotherm weiterverarbeitet. Je 200ml des vorverdünnten Samen wurden bei 32 °C bzw. 21 °C gehalten (für 30 min, 60 min, 3 h und 6 h), danach ausverdünnt (in 100 ml Ebersamenflaschen), für weitere 90 min bei 21 °C gehalten und anschließend bei 17 °C gelagert. Dargestellt sind die Haltezeiten bei 21 °C. Klammern beschreiben die rote bzw. blaue Kurve. Die Kontrollproben (rote Linie), wurde ohne Haltezeiten direkt weiterverarbeitet. Die blaue Linie entspricht den Proben, welche vorverdünnt 3h bei 21 °C gehalten und anschließend isotherm ausverdünnt wurden. Die schwarze Linie beschreibt den Temperaturverlauf der 1:1 vorverdünnten Proben. Die gestrichelte Linie zeigt exemplarisch den Temperaturverlauf der Proben mit 60 min Haltezeit bei 32 °C. Um die Grafik übersichtlicher zu gestalten, wurde auf die Darstellung weiterer Haltezeiten bei 32 °C verzichtet. *Kontrolle: 15 min bei 32°C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
10
15
20
25
30
35
0 60 120 180 240 300 360 420 480 540 600
Tem
pera
tur
in °
C
Minuten
Kontrolle* 30 min Haltezeit 60 min Haltezeit 3 h Haltezeit
6 h Haltezeit Vorverdünnte Probe 60 min bei 32 °C
Ausverdünnt 90 min bei 21 °C
Weitere Abkühlung und Lagerung bei 17 °C
3h Haltezeit bei 21 °C des 1:1 verdünnten Samens
Ausverdünnt 90 min bei 21 °C Weitere Abkühlung und Lagerung bei 17 °C

Ergebnisse
67
4.2.2 CASA-Motilitätsparameter
Abb. 15: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=12 Ejakulate für d0 – d3; n=6 Ejakulate für d6. Klammer: Unterschiede zwischen den Haltetemperaturen bei gleicher Lagerungsdauer und Haltezeit
(p > 0,05)
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die Haltezeit hatte auf die Motilität der Spermien keinen Einfluss.
Die Haltetemperatur hatte auf die Gesamtmotilität (Abb. 15) und progressive Motilität
bei n = 12 (d0 – d3) Einfluss (Tabelle 3). Signifikante Unterschiede zeigten sich bei
der Gesamtmotilität nur an d 1: Die Motilität bei den für 30 min gehaltenen Proben
war bei einer Haltetemperatur von 32 °C signifikant höher als bei 21 °C, wobei die
mittlere Differenz mit 0,8 % gering war. Bei den für 6 h bei 32 °C gehaltenen Proben
zeigten die Proben eine signifikant niedrigere Motilität als bei 21 °C, wobei die
mittlere Differenz von 0,2 % auch hier gering war.
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3 d6
mo
tile
Sp
erm
ien
in
%
Lagerungsdauer
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
n=12 n=6
*

Ergebnisse
68
Mit Ausnahme von der Gesamtmotilität und der progressiven Motilität wurden alle
erhobenen Motilitätsparameter durch die Lagerungsdauer beeinflusst (p < 0,05),
allerdings konnten nur wenige signifikante Unterschiede ermittelt werden (Tabelle 3).
In der folgenden Grafik (Abb. 16) sowie in den Abbildungen 41 – 47 (Anhang)
wurden Spermiensubpopulationen einiger Motilitätsparameter dargestellt. Hierfür
wurden jeweils die Kontrollproben und die für 6 h bei 32 °C bzw. 21 °C gehaltenen
Proben analysiert. Bei allen Parametern waren Unterschiede hauptsächlich an Tag 0
zu sehen, ab d 1 zeigten die Kurven der verschiedenen Behandlungsgruppen jeweils
sehr ähnliche Geschwindigkeitsprofile. Am meisten ausgeprägt waren diese bei den
Geschwindigkeitsparametern VCL (velocity curvilinear) und VSL (velocity straight
line, Abb. 41). Bei der Analyse wurde die Geschwindigkeit in einzelne Klassen à 20
µm/ s (VCL) bzw. 10 µm/ s (VSL) aufgeteilt und der Anteil der Spermien pro Klasse
ermittelt.
Bei VCL ergab die Analyse von d 0 bei den Kontrollproben mit 8,2 % den größten
Anteil der Spermien bei 61 – 80 µm/ s. Die für 6 h bei 32 °C gehaltenen Proben hatte
einen Peak mit 8,6 % der Spermien bei 81 – 100 µm/ s, bei den bei 21 °C gehaltenen
Proben waren mit 6,7 % die meisten Spermien in der Geschwindigkeitsklasse 141 –
160 µm/ s vertreten. Nach 24 h Lagerungsdauer (d 1) wiesen die Spermaproben der
unterschiedlichen Temperaturgruppen sehr ähnliche Geschwindigkeitsprofile auf.
Die Peaks bei VSL befanden sich in niedrigeren Geschwindigkeitsklassen.
Samenproben, die für 6 h bei 32 °C gehalten wurden, erreichten mit 8,6 % der
Spermien bei 41 – 50 µm/ s ihren Höhepunkt. Sowohl die für 6 h bei 21 °C
gehaltenen Proben, als auch die Kontrollproben waren mit den meisten Spermien,
15,28 % (Kontrolle) und 18,72 % (21 °C), in der Geschwindigkeitsklasse
51 - 60 µm/ s vertreten.
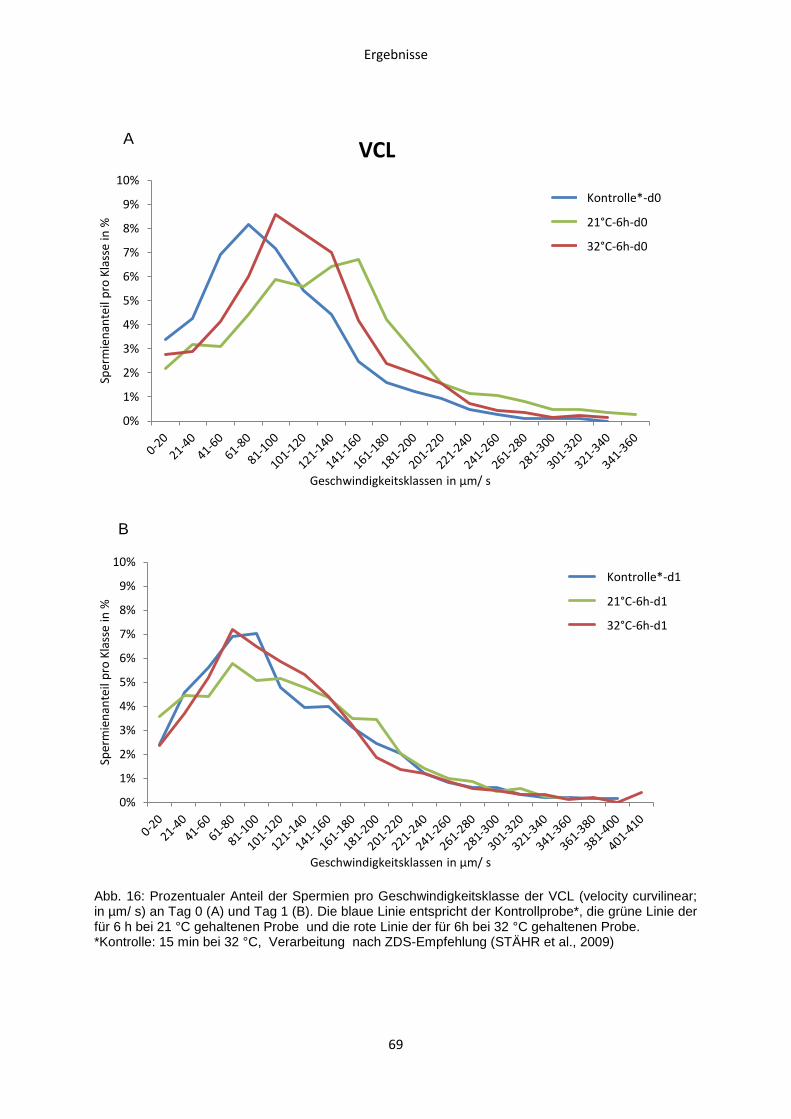
Ergebnisse
69
Abb. 16: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VCL (velocity curvilinear; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
10%
Sper
mie
nan
teil
pro
Kla
sse
in %
Geschwindigkeitsklassen in µm/ s
VCL
K d0
21-6 d0
32-6 d0
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
10%
Sper
mie
nan
teil
pro
Kla
sse
in %
Geschwindigkeitsklassen in µm/ s
K d1
21-6 d1
32-6 d1
A
B
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1

Ergebnisse
70
4.2.3 Membranintegrität
Abb. 17: Prozentualer Anteil membranintakter (PI und FITC-PNA negative) Spermien (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Die Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=12 Ejakulate für d0 – d3; n=6 Ejakulate für d6). Klammer: Unterschiede zwischen den Haltezeiten bei gleicher Haltetemperatur und Lagerungsdauer
(p > 0,05).
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die statistische Auswertung zeigte Einflüsse der Haltezeit und der Lagerungsdauer
auf den Anteil membranintakter Spermien (p < 0,05).
Proben, die bei 32 °C gehalten wurden, zeigten signifikante Veränderungen durch
die Haltezeit nach Lagerungstag 1 und 3. An d 1 zeigten sich signifikante
Unterschiede zwischen 30 min Haltezeit und 60 min, wobei die mittlere Differenz mit
0,3 % gering war. An d 3 zeigten die Proben nach 3 h und 6 h Haltezeit signifikant
geringere Werte im Vergleich zu 30 min, wobei auch hier die mittlere Differenz mit
0,8 % nach 3 h und 1,1 % nach 6 h gering war.
Die Proben, die bei 21 °C gehalten wurden, zeigten haltezeitabhängig signifikante
Veränderungen an d 1. Nach 30 min Haltezeit waren noch 89,4 ± 2,9 % Spermien
membranintakt, dieser Wert sank um 0,3 % nach 3 h Haltezeit. Ein weiterer
Unterschied ergab sich zwischen den für 60 min und 3 h gehaltenen Proben. Der
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3 d6
Mem
bra
nin
takt
e Sp
erm
ien
in %
Lagerungsdauer
Kontrolle(=0min)32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
n=12 n=6
*

Ergebnisse
71
Wert verringerte sich ebenfalls um 0,3 % von 89,5 ± 3,1 % (60 min) auf 89,1 ± 3,1 %
(3 h).
Signifikante Unterschiede der Lagerungsdauer konnten im Paarvergleich nicht
ermittelt werden.
Die Ergebnisse sind in Abb. 17 dargestellt.
4.2.4 Mitochondrienmembranpotential
Abb. 18: Prozentualer Anteil der PI-negativen (Plasmamembran-intakten) Spermien mit mehrheitlich hohem Mitochondrienmembranpotential (MMP) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Die Farbabstufungen: unterschiedlichen Haltezeiten; n=6 Ejakulate. Klammer: Unterschied zw. Lagerungstagen (d0 – d6) innerhalb einer Verarbeitungsvariante (p < 0,05)
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Das Mitochondrienmembranpotential (Abb. 18) wurde durch die Lagerungsdauer
(p < 0,05) beeinflusst. Signifikante Unterschiede zeigten sich nur innerhalb der für
30 min bei 32 °C gehaltenen Proben an d 6. Hier sank das
Mitochondrienmembranpotential von 87,2 ± 7,6 % an d 3 auf 76,6 ± 18,2 % an d 6.
0
10
20
30
40
50
60
70
80
90
100
110
d0 d1 d3 d6
MM
P in
%
Lagerungsdauer
Kontrolle
32-30
32-60
32-3
32-6
21-30
21-60
21-3
21-6
n=6
*

Ergebnisse
72
Abb. 19: Mittlere Fluoreszenzintensität (orange Jagg) aller Spermien (MW + SD) in Abhängigkeit von der Lagerungsdauer, Haltetemperatur und der Haltezeit des vorverdünnten Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen Haltezeiten;n=6 Ejakulate. Klammer: Unterschied zwischen Lagerungstagen (d0 – d6) für eine Verarbeitungsvariante. (p < 0,05) *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die mittlere Fluoreszenzintensität (Orangefluoreszenz, Jagg) wurde nur von der
Lagerungsdauer beeinflusst (p < 0,05, Abb. 19).
Bei für 3 h bei 32 °C gehaltenen Samenproben sank der Wert signifikant von
15,5 ± 5,9 an d 0 auf 11,6 ± 4,7 nach 3 Tagen Lagerung. Die Fluoreszenzintensität
der Kontrollproben sank signifikant von 14,0 ± 4,8 an d 1 auf 10,9 ± 5,1 nach 6 Tagen
Lagerung. Bei den bei 21 °C gehaltenen Proben zeigten sich signifikante
Veränderungen der Fluoreszenzintensität; bei der für 30 min gehaltenen Probe
sank der Wert von 16,4 ± 6,6 (d 1) auf 11,2 ± 4,6 (d 6) und nach 60 min Haltezeit von
15,81 ± 7,2 (d 1) auf 11,9 ± 5,1 (d 6).
0
5
10
15
20
25
30
d0 d1 d3 d6
arb
itra
ry u
nit
s
Lagerungsdauer
Kontrolle
32-30
32-60
32-3
32-6
21-30
21-60
21-3
21-6
n=6
*

Ergebnisse
73
4.2.5 Energiestoffwechsel
Abb. 20: ATP – Gehalt der Spermien in pmol/ 50000 Spermien (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die grüne Säule entspricht dem ATP – Gehalt der unbehandelten Samenprobe, die gestreifte Säule der Kontrolle*. Rote Säulen entsprechen den Proben mit einer Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. Klammer: Unterschied zwischen Lagerungstagen (d0 – d6) für eine Verarbeitungsvariante. (p < 0,05)
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die Haltezeit und Haltetemperatur hatten keinen Einfluss auf den ATP-Gehalt der
Spermien. Nur ein Einfluss der Lagerungsdauer konnte ermittelt werden. In der
grafischen Darstellung (Abb. 20) wird deutlich, dass alle ATP-Werte an d 1
tendenziell niedriger sind als am Verdünnungstag (d 0) und anschließend über die
Lagerung stabil bleiben. Signifikante Unterschiede bestehen nur zwischen der für
60 Minuten bei 21 °C gehaltenen Samenproben an d 0 mit 306 ± 94 pmol/ 50000
Spermien im Vergleich zu d 1 mit 231 ± 60 pmol/ 50000 Spermien, bzw. zu d 6 mit
254 ± 66 pmol/ 50000 Spermien.
Der ADP - und AMP - Gehalt der Spermien zeigte sich stabil während der Lagerung
und wurde weder von der Haltezeit und -temperatur noch von der Lagerungsdauer
beeinflusst (Tabelle 5).
0
50
100
150
200
250
300
350
400
450
d0 d1 d3 d6
ATP
in p
mo
l/ 5
0.0
00
Sp
erm
ien
Lagerungsdauer
ATP nativ
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
*

Ergebnisse
74
Abb. 21: Energieladung (EC) der Spermien in Abhängigkeit von der Lagerungsdauer, Halte-temperatur und der Haltezeit des vorverdünnten Samens. Die grüne Säule entspricht der EC der unbehandelten Samenproben, die gestreifte Säule der Kontrollen*. Rote Säulen entsprechen den Proben mit einer Haltetemperatur von 32 °C und die blauen Säulen Proben mit einer Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS Empfehlung (STÄHR et al., 2009)
Die aus den Nukleotiden errechnete Energieladung („Energy Charge“, EC) der Zellen
kann Null betragen bei Zellen, die nur AMP enthalten und kann einen Wert bis Eins
erreichen bei Zellen, die nur ATP enthalten. Bei Werten > 0,6 ist von vitalen Zellen
auszugehen (s. Kapitel 3.13.1). Der Wert von 0,6 wurde im Mittel während des
Versuchs nicht unterschritten.
Beeinflusst wurde die EC nur von der Haltetemperatur. Allerdings wurden im
statistischen Test keine signifikanten Unterschiede im Paarvergleich ermittelt
(Abb. 21).
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
d0 d1 d3 d6
Ener
giel
adu
ng
Lagerungsdauer
ATP nativ
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
*
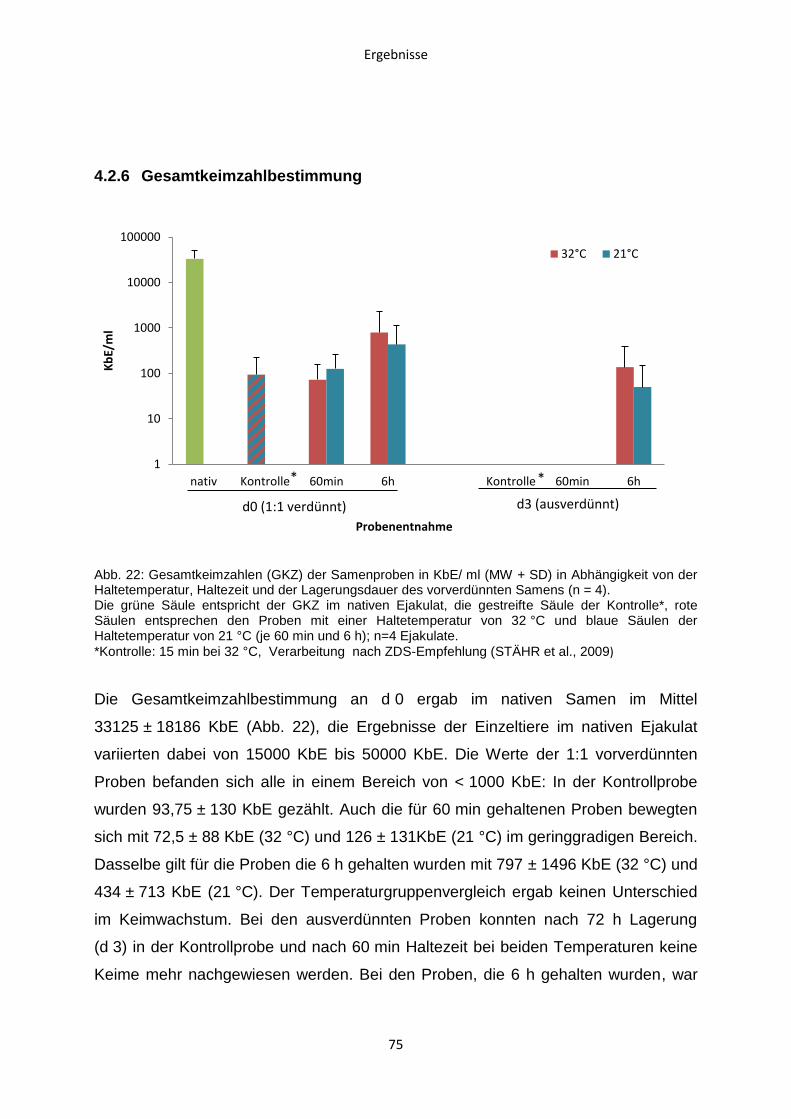
Ergebnisse
75
4.2.6 Gesamtkeimzahlbestimmung
Abb. 22: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens (n = 4). Die grüne Säule entspricht der GKZ im nativen Ejakulat, die gestreifte Säule der Kontrolle*, rote Säulen entsprechen den Proben mit einer Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C (je 60 min und 6 h); n=4 Ejakulate.
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die Gesamtkeimzahlbestimmung an d 0 ergab im nativen Samen im Mittel
33125 ± 18186 KbE (Abb. 22), die Ergebnisse der Einzeltiere im nativen Ejakulat
variierten dabei von 15000 KbE bis 50000 KbE. Die Werte der 1:1 vorverdünnten
Proben befanden sich alle in einem Bereich von < 1000 KbE: In der Kontrollprobe
wurden 93,75 ± 130 KbE gezählt. Auch die für 60 min gehaltenen Proben bewegten
sich mit 72,5 ± 88 KbE (32 °C) und 126 ± 131KbE (21 °C) im geringgradigen Bereich.
Dasselbe gilt für die Proben die 6 h gehalten wurden mit 797 ± 1496 KbE (32 °C) und
434 ± 713 KbE (21 °C). Der Temperaturgruppenvergleich ergab keinen Unterschied
im Keimwachstum. Bei den ausverdünnten Proben konnten nach 72 h Lagerung
(d 3) in der Kontrollprobe und nach 60 min Haltezeit bei beiden Temperaturen keine
Keime mehr nachgewiesen werden. Bei den Proben, die 6 h gehalten wurden, war
1
10
100
1000
10000
100000
nativ Kontrolle 60min 6h Kontrolle 60min 6h
Kb
E/m
l
Probenentnahme
32°C 21°C
d0 (1:1 verdünnt) d3 (ausverdünnt)
* *

Ergebnisse
76
mit 138 ± 243 KbE (32 °C) und 50 ± 100 KbE (21 °C) noch Keimwachstum
vorhanden, allerdings im geringgradigen Bereich.
In der vergleichenden Darstellung der Eber (Abb. 23) ist zu sehen, dass die Werte
aller Proben an d 0 im geringgradigen Bereich von 102 KbE liegen, mit Ausnahme
von Eber 2, der unmittelbar nach 6 h Haltezeit bei beiden Temperaturen mehr
Keimwachstum im verdünnten Sperma aufweist, d.h.: 3000 KbE bei 32 °C und 1500
KbE bei 21 °C Haltetemperatur. Dieses reduzierte sich in der ausverdünnten,
gelagerten Probe. Nach 72 h Lagerung (Abb. 24) zeigte bei der in 32 °C für 6 h
gehaltenen Probe nur noch Eber 1 mit 500 KbE und Eber 4 mit 50 KbE
Keimwachstum. Bei den bei 21 °C gehaltenen Proben wurden nur bei Eber 1 mit
200 KbE noch Keime nachgewiesen.
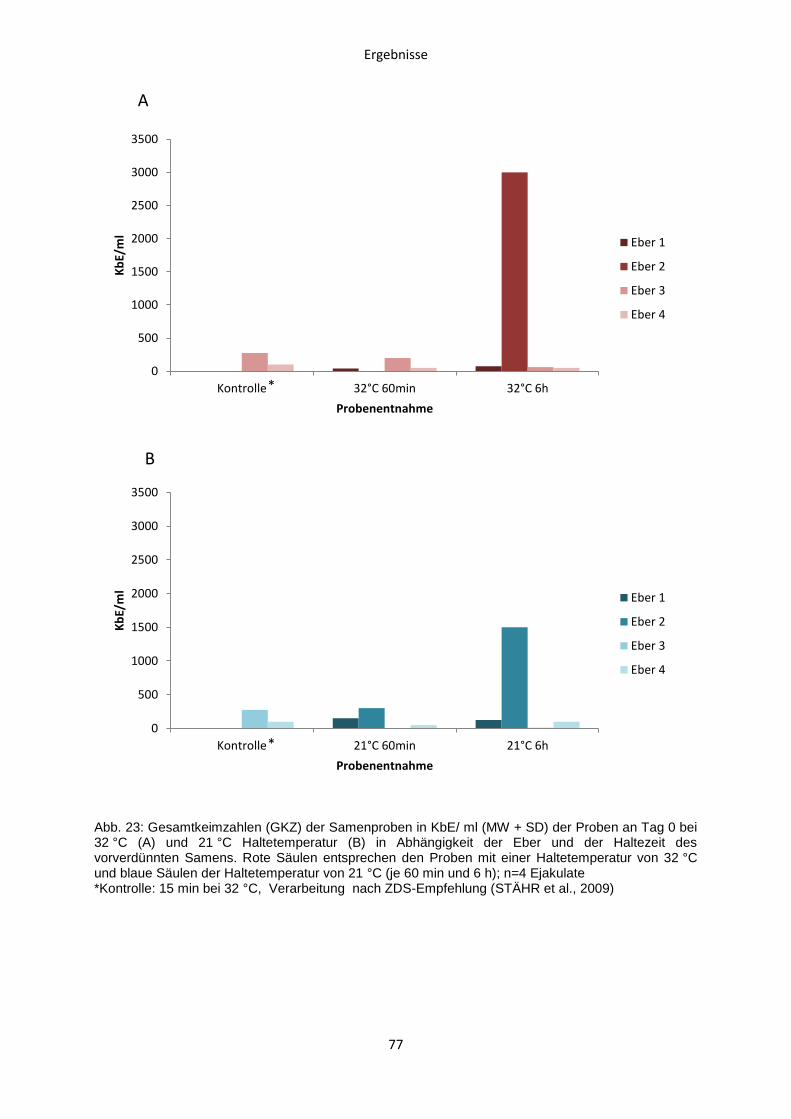
Ergebnisse
77
Abb. 23: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW + SD) der Proben an Tag 0 bei 32 °C (A) und 21 °C Haltetemperatur (B) in Abhängigkeit der Eber und der Haltezeit des vorverdünnten Samens. Rote Säulen entsprechen den Proben mit einer Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C (je 60 min und 6 h); n=4 Ejakulate *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
0
500
1000
1500
2000
2500
3000
3500
Kontrolle 32°C 60min 32°C 6h
Kb
E/m
l
Probenentnahme
Eber 1
Eber 2
Eber 3
Eber 4
0
500
1000
1500
2000
2500
3000
3500
Kontrolle 21°C 60min 21°C 6h
Kb
E/m
l
Probenentnahme
Eber 1
Eber 2
Eber 3
Eber 4
A
B
*
*

Ergebnisse
78
Abb. 24: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW * SD) der Proben an Tag 3 bei 32 °C (A) und 21 °C Haltetemperatur (B) in Abhängigkeit der Eber und der Haltezeit des vorverdünnten Samens. Rote Säulen entsprechen den Proben mit einer Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C (je 60 min und 6 h); n=4 Ejakulate. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS- Empfehlung (STÄHR et al., 2009)
0
500
1000
1500
2000
2500
3000
3500
Kontrolle 32°C 60min 32°C 6h
Kb
E/m
l
Probenentnahme
Eber 1
Eber 2
Eber 3
Eber 4
*
0
500
1000
1500
2000
2500
3000
3500
Kontrolle 21°C 60min 21°C 6h
Kb
E/m
l
Probenentnahme
Eber 1
Eber 2
Eber 3
Eber 4
*
A
B

Ergebnisse
79
4.3 Experiment 3: Vergleich zweistufig isothermer mit zweistufig
hyperthermer Verdünnung
4.3.1 Temperaturverlauf während der Samenverarbeitung
Nach der Vorverdünnung im Verhältnis 1:1 (v:v) wurden die Ejakulate in zwei
Glaszylinder aufgeteilt und jeweils bis zu 6 h bei 21 °C gehalten. Innerhalb der ersten
90 min verringerte sich die Temperatur hier von 31,1 °C auf 23,9 °C. Während der
ersten 30 min bei 21 °C betrug die durchschnittliche Abkühlrate 0,09 ± 0,09 °C/ min,
dann 0,10 ± 0,05 °C/ min (31 – 60 min) und schließlich 0,05 ± 0,05 °C/ min
(61 - 90 min).
Die Kontrollprobe wurde nach der Ausverdünnung mit isothermem Verdünner für
90 min bei 21 °C Umgebungstemperatur abgekühlt. Während dieser Zeitspanne sank
die Temperatur der Samenproben im Mittel von 31,6 °C auf 24,1 °C. Sie zeigten die
gleichen Abkühlraten wie die vorverdünnten Samenproben.
Nach jeder Haltezeit wurden die unterschiedlich abgekühlten Proben mit isothermem
bzw. auf 32 °C erwärmten Verdünner ausverdünnt. Nach 30 min Haltezeit bei 21 °C
waren die vorverdünnten Proben auf 28,2 °C abgekühlt, die hypertherme
Ausverdünnung erwärmte diese auf 31,6 °C. Nach 60 min Haltezeit erwärmten sich
die Proben von 25,4 °C auf 30,4 °C, von 22,1 °C auf 29,8 °C (3 h Haltezeit) und nach
6 h Haltezeit von 21,5 °C auf 29,6 °C. Im Anschluss an die Ausverdünnung wurden
die Proben für 90 min erneut bei 21 °C gehalten. Die Abkühlraten bei in den ersten
30 min nach der jeweiligen Verdünnung betrug 0,13 ± 0,04 °C/ min (30 min
Haltezeit), 0,12 ± 0,06 °C/ min (60 min) und 0,10 ± 0,06 °C/ min nach 3 h und 6 h
Haltezeit. Nach 90 min bei 21 °C hatten alle Proben eine Temperatur von 23,7 °C.
Die Lagerungstemperatur (17,0 ± 0,5 °C) erreichten die Kontrollproben 255 min nach
Messbeginn, die für 30 min bei 21 °C gehaltenen Proben nach 273 min, die für
60 min gehaltenen Proben erreichten sie nach 305 min, die Proben mit 3 h Haltezeit
nach 532 min und die für 6 h gehaltenen Proben nach 635 min.
Der Temperaturverlauf ist in Abb. 25 dargestellt.

Ergebnisse
80
Abb. 25: Temperaturverlauf der bei Experiment 3 bei 21 °C gehaltenen und anschließend hypertherm verdünnten Proben. Mittelwerte von je 3 Samenproben. Diese wurden im Verhältnis 1:1 (200ml: 200 ml) mit Beltsville Thawing Solution (BTS, 32 °C) verdünnt und zweistufig hypertherm bzw. isotherm weiterverarbeitet. Beide vorverdünnte Proben wurden bei 21 °C gehalten (für 30 min, 60 min 3 h und 6 h), danach hypertherm (BTS, 32 °C) bzw. isotherm ausverdünnt, für weitere 90 min bei 21 °C gehalten und anschließend bei 17 °C gelagert. Dargestellt sind die hypertherm verdünnten Proben, sowie die Kontrollprobe. Klammern beschreiben den Verlauf der roten bzw. blauen Kurve. Die Kontrollproben (rote Linie), wurde ohne Haltezeiten direkt isotherm weiterverarbeitet. Die blaue Linie entspricht den Proben, welche vorverdünnt 3h bei 21 °C gehalten und anschließend hypertherm (BTS 32 °C) ausverdünnt wurden. Um die Grafik übersichtlicher zu gestalten, wurde auf die Darstellung der isothermen Verdünnung verzichtet.
10
15
20
25
30
35
0 60 120 180 240 300 360 420 480 540 600
Tem
per
atu
r in
°C
Minuten
30 min Haltezeit 60 min Haltezeit 3 h Haltezeit
6 h Haltezeit Kontrolle (zweist. isotherm)
Ausverdünnt 90 min bei 21 °C
Weitere Abkühlung und Lagerung bei 17 °C
3h Haltezeit bei 21 °C des 1:1 verdünnten Samens
Ausverdünnt 32 °C BTS 90 min bei 21 °C Weitere Abkühlung und Lagerung bei 17 °C

Ergebnisse
81
4.3.2 Casa-Motilitätsparameter
Abb. 26: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Verdünnertemperatur im zweiten Verdünnungsschritt, der Haltezeit des vorverdünnten Samens und der Lagerungsdauer. Dargestellt sind die Kontrolle* (gestreifte Säulen), hypertherm ausverdünnte Proben (rote Säulen) und isotherm ausverdünnte Proben (blaue Säulen). Farbabstufungen: unterschiedliche Haltezeiten; n=6 Ejakulate. Klammer: Unterschiede zwischen Lagerungstagen (d0 – d3) für eine Verarbeitungsvariante (p > 0,05)
*Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (STÄHR et al., 2009)
Die Temperatur des Verdünners beeinflusste von den Motilitätsparameter nur die
Geschwindigkeitsparameter VCL, VSL und VAP (p < 0,05). Statistisch signifikante
Unterschiede konnten bei den für 6 h gehaltenen Proben nach 3 Tagen Lagerung
ermittelt werden (Tabelle 6). Die für 6 h gehaltenen Proben zeigten nach 3 Tagen
Lagerungsdauer eine velocity average path (VAP) der hypertherm ausverdünnten
Proben von 63,97 ± 11,55 µm/ s, wohingegen der Wert der isotherm ausverdünnten
Probe mit 60,35 ± 9,97 µm/ s signifikant niedriger war. Die velocity straight-line (VSL)
an Lagerungstag 3 der hypertherm ausverdünnten Proben betrug 50,11 ± 4,81 µm/ s,
die Werte der isotherm verdünnten Proben dagegen 46,18 ± 4,94 µm/ s.
Von den Haltezeiten beeinflusst wurden nur die Parameter straightness (STR),
linearity (LIN), wobble (WOB) und amplitude of lateral head displacement (ALH).
Signifikante Unterschiede zeigte sich vor allem am Tag der Verdünnung (d 0).
Während der Lagerung verminderten sich diese (Tabelle 6).
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3
Mo
tile
Sp
erm
ien
in %
Lagerungsdauer
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
Kontrolle*

Ergebnisse
82
Alle Motilitätsparameter wurden durch die Lagerungsdauer beeinflusst (p < 0,05).
Signifikante Unterschiede konnten nur bei der isothermen Verdünnung festgestellt
werden. Hier sank die Gesamtmotilität bei der für 6 h gehaltenen Probe von
93,0 ± 2,2 % an d 0 auf 84,3 ± 15,4 % an d 3. Die für 30 min gehaltenen Probe
verringerte sich von 92,8 ± 3,0 % an d 1 auf 89,5 ± 5,2 % an d 3.
Bei der progressiven Motilität zeigten sich signifikante Unterschiede nach 72 h
Lagerung ebenfalls hauptsächlich bei den isotherm verdünnten Proben (nach 3 h und
6 h Haltezeit, Tabelle 6). Bei den für 30 min gehaltenen Proben zeigten sowohl die
isotherm verdünnte, als auch die hypertherm verdünnte Probe signifikante
Unterschiede. Die progressive Motilität der hypertherm ausverdünnten Probe sank
von 85,6 ± 4,0 % an d 0 auf 80,3 ± 6,47 % nach 3 Tagen Lagerung. Der Wert der
isotherm ausverdünnten Probe sank von 85,22 ± 4,2 % (d 0) auf 78,6 ± 10,8 % an
d 3.
Die Gesamtmotilität in Abhängigkeit von der Verdünnertemperatur bei der
Ausverdünnung ist in Abb. 26 dargestellt.

Ergebnisse
83
4.3.3 Membranintegrität
Abb. 27: Prozentualer Anteil membranintakter (PI und FITC-PNA negative) Spermien (MW + SD) in Abhängigkeit von der Verdünnertemperatur im zweiten Verdünnungsschritt, der Haltezeit des vorverdünnten Samens und der Lagerungsdauer; n=6 Ejakulate Dargestellt sind die Kontrolle* (gestreifte Säulen), hypertherm ausverdünnte Proben (rote Säulen) und isotherm ausverdünnte Proben (blaue Säulen). Farbabstufungen: unterschiedliche Haltezeiten. #: Unterschied zwischen den Verdünnertemperaturen innerhalb eines Lagerungstages. (p > 0,05) Klammer: Unterschiede zwischen Lagerungstagen für eine Verarbeitungsvariante. (p < 0,05) *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS Empfehlung (STÄHR et al., 2009)
Der Anteil der PI- und FITC-PNA-negativen (plasma- und
akrosomenmembranintakter) Spermien wurde von der Temperatur des zweiten
Verdünners und von der Lagerungsdauer beeinflusst (p < 0,05) (Abb. 27).
Der Einfluss der Temperatur des zweiten Verdünners zeigte sich statistisch
signifikant im Paarvergleich nur bei den für 60 min gehaltenen Proben an
Lagerungstag 3. Die hypertherm verdünnten Samenproben zeigten geringere
Prozentsätze membranintakter Spermien als isotherm verdünnte Proben auf, wobei
die mittlere Differenz mit 0,8 % gering war.
Die Werte bewegten sich bei der Lagerungsdauer von 72 h in einem sehr engen
Bereich. Mit Ausnahme eines Messzeitpunktes lagen die Mittelwerte der
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3
Mem
bra
nin
tegr
ität
in %
Lagerungsdauer
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
#
*

Ergebnisse
84
membranintakten Samenzellen bei allen Proben während der gesamten
Lagerungsdauer bei über 90 %.
Signifikante Unterschiede zeigten sich nur bei den isotherm verdünnten Proben:
Die für 60 min gehaltenen Proben wiesen nach Lagerungstag 1 und 3 signifikant
niedrigere Werte auf verglichen mit d 0, wobei die mittleren Differenzen zwischen den
Lagerungstagen mit 1 % bis 2,2 % gering waren. Nach 30 min Haltezeit ergab die
Messung an d 0 einen Anteil von 92,7 ± 2,3 % membranintakten Spermien; dieser
Wert erniedrigte sich auf 92,3 ± 2,0 an (d 1) und auf 90,6 ± 1,6 % an d 3.
Bei den für 3 h gehaltenen Proben reduzierte sich der Wert von 92,4 ± 1,6 % (d 0)
auf 90,7 ± 0,7 % (d 3).
4.4 Experiment 4 a: Auswirkungen von Haltezeiten des
ausverdünnten Samens nach zweistufig hypothermer Verdünnung
4.4.1 Temperaturverlauf während der Samenverarbeitung
Nach der isothermen Vorverdünnung wurde das Sperma zunächst 20 min bei 21 °C
gehalten. Die Temperatur nach der Verdünnung betrug 31,6 °C; während der
Haltezeit kühlten die Proben auf 29,0 °C ab. Die anschließende Ausverdünnung in
100 ml Ebersamenflaschen wurde hypotherm mit einem auf 21 °C temperierten BTS
durchgeführt; dadurch verringerte sich die Probentemperatur auf 23,8 °C. Die
Kontrollproben wurden anschließend 90 min bei 21 °C gehalten, wobei die
Temperatur auf 21,9 °C sank. Die durchschnittliche Abkühlrate der Kontrollproben
während der ersten 30 min (0 – 30 min) war 0,03 ± 0,04 °C/ min.
Die anderen ausverdünnten Proben wurden bei 21 °C oder 32 °C für 30 min, 60 min,
3 h bzw. 6 h gehalten. Die bei 32 °C gehaltenen Proben erreichten die
Haltetemperatur nach 27 min. Die Temperatur erhöhte sich in diesem Zeitraum von
23,4 °C auf 32 °C mit einer Rate von 0,27 ± 0,42 °C/ min. Nach der jeweiligen
Haltezeit wurden die Proben 90 min bei 21 °C abgekühlt. Die Abkühlrate verringerte
sich von 0,13 ± 0,05 °C (0 – 30 min), auf 0,08 ± 0,05 °C/ min (31 – 60 min) und
0,05 ± 0,04 °C/ min (61 – 90 min). Beim anschließenden Umlagern auf 17 °C war die

Ergebnisse
85
Probentemperatur 23,9 °C. Die Lagerungstemperatur von 17,0 ± 0,5 °C wurde nach
291 min (Kontrolle), 303 min (30 min Haltezeit), 334 min (60 min Haltezeit), 454 min
(3 h Haltezeit) und 642 min (6 h Haltezeit) erreicht.
Der Temperaturverlauf ist in Abb. 28 dargestellt.
Abb. 28: Temperaturverlauf der bei Experiment 4a zweistufig hypotherm verdünnten und anschließend bei 32 °C gehaltenen Proben. Mittelwerte von je 3 Samenproben. Diese wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville Thawing Solution (BTS 32 °C) verdünnt 20 min bei 21 °C gehalten und schließlich mit BTS (21 °C) hypotherm ausverdünnt. Die ausverdünnten Proben (in 100 ml Ebersamenflaschen) wurden bei 32 °C bzw. 21 °C gehalten (für 30 min, 60 min, 3 h und 6 h), und danach für weitere 90 min bei 21 °C gehalten und anschließend bei 17 °C gelagert. Dargestellt sind der Temperaturverlauf der bei 32 °C gehaltenen Proben und die Kontrollproben. Die Klammern beschreiben den Verlauf der roten bzw. blauen Kurve. Die rote Kurve entspricht den Kontrollproben, die ohne Haltezeiten direkt weiterverarbeitet wurden. Die blaue Kurve entspricht den Proben, die nach der Ausverdünnung 3 h bei 32 °C gehalten wurden. Um die Grafik übersichtlicher zu gestalten wurde auf die Darstellung der Haltezeiten bei 21 °C
verzichtet.
*Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
10
15
20
25
30
35
0 60 120 180 240 300 360 420 480 540 600 660
Tem
pera
tur
in °
C
Minuten
6 h Haltezeit 3 h Haltezeit 60 min Haltezeit 30 min Haltezeit Kontrolle*
zweistufig hypotherm ausverdünnt,
90 min Abkühlung bei 21 °C
Weitere Abkühlung und Lagerung bei 17 °C
3 h Halten bei 32 °C 90 min bei 21 °C Abkühlung und Lagerung bei 17 °C
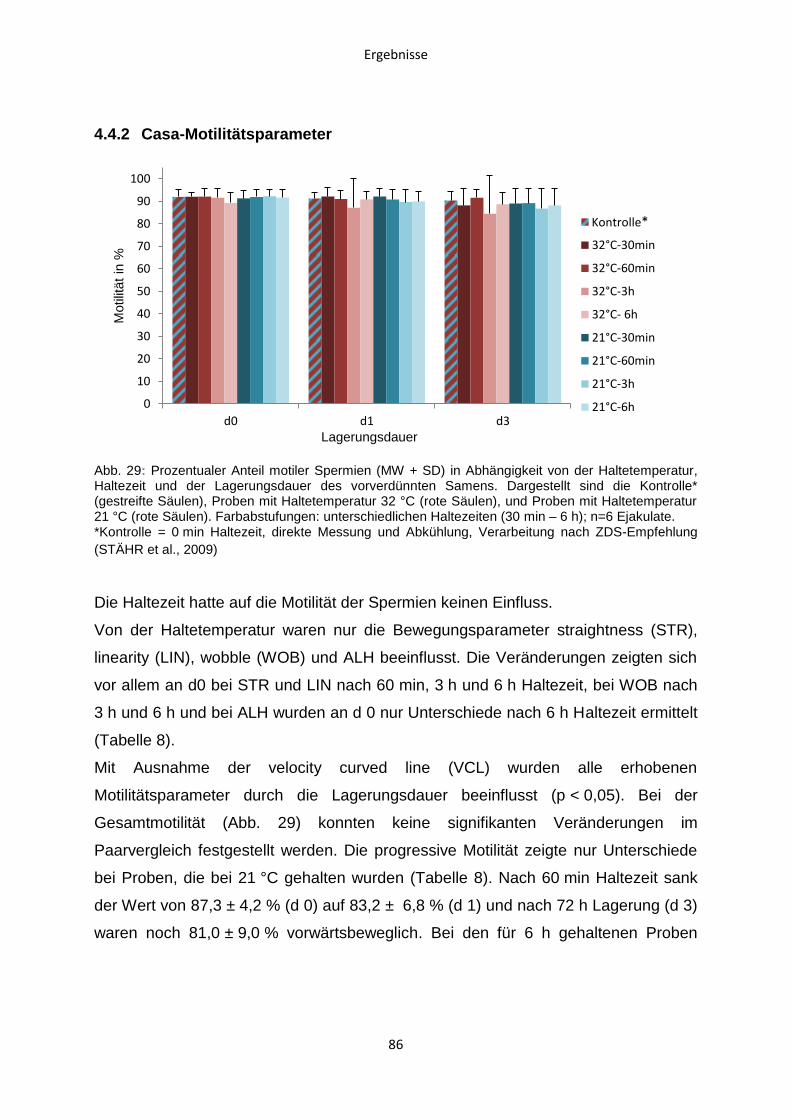
Ergebnisse
86
4.4.2 Casa-Motilitätsparameter
Abb. 29: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen). Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. *Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
Die Haltezeit hatte auf die Motilität der Spermien keinen Einfluss.
Von der Haltetemperatur waren nur die Bewegungsparameter straightness (STR),
linearity (LIN), wobble (WOB) und ALH beeinflusst. Die Veränderungen zeigten sich
vor allem an d0 bei STR und LIN nach 60 min, 3 h und 6 h Haltezeit, bei WOB nach
3 h und 6 h und bei ALH wurden an d 0 nur Unterschiede nach 6 h Haltezeit ermittelt
(Tabelle 8).
Mit Ausnahme der velocity curved line (VCL) wurden alle erhobenen
Motilitätsparameter durch die Lagerungsdauer beeinflusst (p < 0,05). Bei der
Gesamtmotilität (Abb. 29) konnten keine signifikanten Veränderungen im
Paarvergleich festgestellt werden. Die progressive Motilität zeigte nur Unterschiede
bei Proben, die bei 21 °C gehalten wurden (Tabelle 8). Nach 60 min Haltezeit sank
der Wert von 87,3 ± 4,2 % (d 0) auf 83,2 ± 6,8 % (d 1) und nach 72 h Lagerung (d 3)
waren noch 81,0 ± 9,0 % vorwärtsbeweglich. Bei den für 6 h gehaltenen Proben
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3
Motilit
ät
in %
Lagerungsdauer
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
Kontrolle*
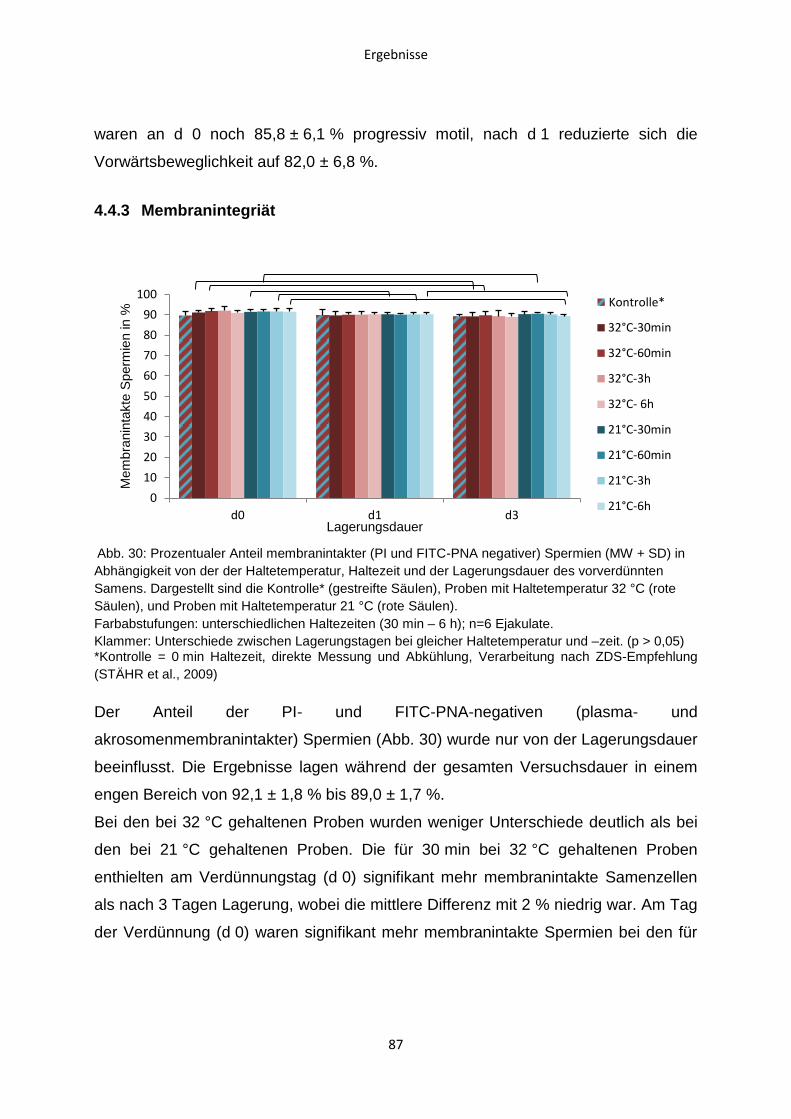
Ergebnisse
87
waren an d 0 noch 85,8 ± 6,1 % progressiv motil, nach d 1 reduzierte sich die
Vorwärtsbeweglichkeit auf 82,0 ± 6,8 %.
4.4.3 Membranintegriät
Abb. 30: Prozentualer Anteil membranintakter (PI und FITC-PNA negativer) Spermien (MW + SD) in
Abhängigkeit von der der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten
Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote
Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen).
Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate.
Klammer: Unterschiede zwischen Lagerungstagen bei gleicher Haltetemperatur und –zeit. (p > 0,05) *Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
Der Anteil der PI- und FITC-PNA-negativen (plasma- und
akrosomenmembranintakter) Spermien (Abb. 30) wurde nur von der Lagerungsdauer
beeinflusst. Die Ergebnisse lagen während der gesamten Versuchsdauer in einem
engen Bereich von 92,1 ± 1,8 % bis 89,0 ± 1,7 %.
Bei den bei 32 °C gehaltenen Proben wurden weniger Unterschiede deutlich als bei
den bei 21 °C gehaltenen Proben. Die für 30 min bei 32 °C gehaltenen Proben
enthielten am Verdünnungstag (d 0) signifikant mehr membranintakte Samenzellen
als nach 3 Tagen Lagerung, wobei die mittlere Differenz mit 2 % niedrig war. Am Tag
der Verdünnung (d 0) waren signifikant mehr membranintakte Spermien bei den für
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3
Mem
bra
nin
takte
Sperm
ien in
%
Lagerungsdauer
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
Kontrolle*

Ergebnisse
88
60 min gehaltenen Proben als nach 1 oder 3 Tagen Lagerung. Auch hier war die
mittlere Differenz mit 1,7 % an d 1 und 2 % nach 3 Tagen Lagerung gering.
Proben, die einer Haltetemperatur von 21 °C ausgesetzt wurden, wiesen bei allen
Haltezeiten signifikant höhere Ergebnisse am Tag der Verdünnung auf, verglichen
mit den Ergebnissen nach 1 oder 3 Tagen Lagerungsdauer.
Nach 30 min Haltezeit wurden am Tag der Verdünnung (d 0) 91,5 ± 1,3 %
membranintakte Samenzellen gemessen, an d 1 ergab die Messung einen Wert von
90,3 ± 0,8 %. Ähnliche Zahlen zeigten sich nach 60 min Haltezeit, mit 91,6 ± 1,0 %
(d 0) und 90,5 ± 0,5 % (d 3). Die Werte der für 3 h gehaltenen Proben sanken von
91,6 ± 1,3 % (d 0) auf 90,2 ± 1,0 % (d 1). Über alle 3 Lagerungstage wurden
Veränderungen bei den für 6 h gehaltenen Proben gemessen: Der Wert 90,5 ± 1,4 %
(d 0) verringerte sich auf 90,3 ± 0,7 % (d 1) und 89,4 ± 0,9 % nach 72 h Lagerung.
4.5 Experiment 4 b: Auswirkungen von Haltezeiten des
ausverdünnten Samens nach einstufig isothermer Verdünnung
4.5.1 Temperaturverlauf während der Samenverarbeitung
Nach der einstufig isothermen Verdünnung (in 100 ml Ebersamenflaschen) lag die
Temperatur der Proben bei 31,6 °C. Die Kontrollprobe wurde 90 min bei 21 °C
gehalten. In den ersten 30 min betrug die Abkühlrate 0,13 ± 0,04 °C/ min; sie sank
dann auf 0,08 ± 0,05 °C/ min (31 -60 min) und schließlich auf 0,05 ± 0,04 °C/ min
(61 – 90 min). Bei der Umlagerung in den 17 °C Klimaschrank betrug die
Probentemperatur 23,8 °C.
Nach der Ausverdünnung wurden die anderen ausverdünnten Proben bei 21 °C bzw.
32 °C für 30 min, 60 min, 3 h bzw. 6 h gehalten. Anschließend kühlten sie weitere 90
min bei 21 °C ab. Die Proben zeigten gleiche Abkühlraten wie die Kontrollproben.
Die für 30 min bei 21 °C gehaltenen Proben zeigten nach der Haltezeit noch eine
Temperatur von 27,4 °C, nach weiteren 90 min bei 21 °C sank diese auf 22,7 °C.
Nach 60 min Halten ergab die Temperaturmessung 25,3 °C, nach den
anschließenden 90 min verringerte sich diese auf 22,1 °C. Sowohl die für 3 h und für

Ergebnisse
89
6 h gehaltenen Proben waren nach der Haltezeit mit 21,6 °C vollständig auf die
Umgebungstemperatur abgekühlt. Die Lagerungstemperatur wurde nach 245 min
(Kontrolle), 270 min (30 min Haltezeit), 292 min (60 min Haltezeit), 439 min (3 h
Haltezeit) und 625 min (6 h Haltezeit) erreicht.
Der Temperaturverlauf ist in Abb. 31 dargestellt.
Abb. 31: Temperaturverlauf der bei Experiment 4b einstufig isotherm verdünnten und anschließend bei 21 °C gehaltenen Proben. Mittelwerte von je 3 Samenproben. Diese wurden einstufig isotherm mit Beltsville Thawing Solution (BTS, 21 °C) ausverdünnt und in 100 ml Ebersamenflaschen abgefüllt. Die ausverdünnten Proben wurden bei 32 °C bzw. 21 °C gehalten (für 30 min, 60 min, 3 h und 6 h), und danach für weitere 90 min bei 21 °C gehalten und anschließend bei 17 °C gelagert. . Dargestellt sind der Temperaturverlauf der bei 21 °C gehaltenen Proben und die Kontrollproben. Die Klammern beschreiben den Verlauf der roten bzw. blauen Kurve. Die rote Kurve entspricht den Kontrollproben, die ohne Haltezeiten direkt weiterverarbeitet wurden. Die blaue Kurve entspricht den Proben, die nach der Ausverdünnung 3 h bei 21 °C gehalten wurde. Um die Grafik übersichtlicher zu gestalten wurde, auf die Darstellung der Haltezeiten bei 32 °C verzichtet. *Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
10
15
20
25
30
35
0 60 120 180 240 300 360 420 480 540 600
Tem
pera
tur
in °
C
Minuten
Kontrolle* 30 min Haltezeit 60 min Haltezeit
3 h Haltezeit 6 h Haltezeit
Ausverdünnt 90 min bei 21 °C
weitere Abkühlung und Lagerung bei 17 °C
Ausverdünnter Samen 3h Haltezeit bei 21 °C
90 min bei 21 °C
weitere Abkühlung und Lagerung bei 17 °C

Ergebnisse
90
4.5.2 CASA-Motilitätsparameter
Abb. 32: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen). Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. Klammer: Unterschiede zwischen Lagerungstagen für eine Verarbeitungsvariante. (p < 0,05; n=6) *Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
Die Haltezeit hatte auf die Motilität der Spermien keinen Einfluss.
Nur die Parameter Gesamtmotilität, progressive Motilität, straightness, linearity und
wobble wurden von der Haltetemperatur beeinflusst (p < 0,05). Bei der Gesamt- und
progressiven Motilität konnten im Paarvergleich allerdings keine Unterschiede
festgestellt werden. Veränderungen bei straightness und linearity konnten nur am
Tag der Verdünnung bei den für 6 h gehaltenen Proben gemessen werden. Der
Parameter wobble zeigte diese bei den für 60 min gehaltenen Proben, ebenfalls an
d 0 (Tabelle 11).
Alle Motilitätsparameter, mit Ausnahme der ALH wurden durch die Lagerungsdauer
beeinflusst (p < 0,05). Bei der Gesamtmotilität (Abb. 32) konnten Veränderungen nur
bei der Kontrolle bzw. den bei 32 °C gehaltenen Proben ermittelt werden. Die
Kontrolle zeigte signifikante Unterschiede zwischen d 0 mit 93,4 ± 1,6 % und d 1 mit
89,9 ± 2,7 % beweglichen Spermien. Die für 60 min bei 32 °C gehaltenen Proben
0
10
20
30
40
50
60
70
80
90
100
d1 d2 d3
Mo
tilit
ät in
%
Lagerungsdauer
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
*

Ergebnisse
91
zeigten eine Gesamtmotilität von 91,5 ± 3,2 % an d 0. Dieser Wert sank auf
86,4 ± 10,2 % nach 3 Tagen Lagerung. Die für 3 h gehaltenen Proben reduzierten
sich von 92,0 ± 3,7 % (d 1) auf 89,3 ± 3,9 % bewegliche Spermien an d 3.
4.5.3 Membranintegrität
Abb. 33: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW * SD) in Abhängigkeit von der der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen). Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. Klammer: Unterschiede zwischen Lagerungstagen für eine Verarbeitungsvariante. (p < 0,05) *Kontrolle = 0 min Haltezeit, direkte Messung und Abkühlung, Verarbeitung nach ZDS-Empfehlung
(STÄHR et al., 2009)
Der Anteil der PI- und FITC-PNA-negativen (plasma- und akrosommembranintakter)
Spermien wurde sowohl von der Haltetemperatur als auch von der Lagerungsdauer
beeinflusst (p < 0,05). Bezüglich der Haltetemperatur konnten aber keine
signifikanten Veränderungen ermittelt werden (Abb. 33).
0
10
20
30
40
50
60
70
80
90
100
d0 d1 d3
Mem
bra
nin
takt
e Sp
erm
ien
in %
Lagerungsdauer
Kontrolle
32°C-30min
32°C-60min
32°C-3h
32°C- 6h
21°C-30min
21°C-60min
21°C-3h
21°C-6h
*

Ergebnisse
92
Während der Lagerungsdauer sank der Anteil der membranintakten Spermien im
Mittel nicht unter 90 %. Die Werte in der Kontrolle verringerten sich von 92,7 ± 1,7 %
an d 0 auf 91,7 ± 2,1 % an d 1.
Weitere Veränderungen wurden hauptsächlich bei den bei 32 °C gehaltenen Proben
festgestellt. Die für 60 min bei 32 °C gehaltenen Proben enthielten am Tag der
Verdünnung 92,6 ± 1,6 % membranintakte Spermien. Dieser Wert sank auf
91,2 ± 1,0 % (d 1) und 91,0 ± 2,5 % (d 3). Signifikante Veränderungen zeigte auch
die für 6 h gehaltene Probe mit 93,1 ± 0,9 % an d 0 und 90,9 ± 1,5 %
membranintakter Spermien nach 3 Tagen Lagerung.
Bei den bei 21 °C gehaltenen Proben zeigten sich signifikante Unterschiede der bei
3 h gehaltenen Proben von 93,3 ± 2,0 % (d 0) auf 91,8 ± 2,5 % (d 3).
4.6 Experiment 5: Auswirkungen einer beschleunigten Abkühlung
des ausverdünnten Samens nach einstufig isothermer Verdünnung
4.6.1 Temperaturverlauf während der Samenverarbeitung
Im Anschluss an die isotherme Ausverdünnung in 100 ml Ebersamenflaschen wurde
ein Teil der Proben gemäß ZDS-Empfehlungen zunächst 90 min bei 21 °C gehalten
und anschließend bei 17 °C gelagert (Kontrolle). Der andere Teil der Proben wurde
direkt nach der Ausverdünnung ohne weitere Haltezeit direkt bei 17 °C gelagert.
Die bei 21 °C gehaltenen Proben sanken während der ersten 90 min von 31,6 °C auf
23,7 °C. Der Temperaturrückgang pro Minute belief sich hier auf 0,13 ± 0,04 °C/ min
(0 – 30 min), 0,08 ± 0,05 °C/ min (31 – 60 min) und 0,05 ± 0,04 °C/ min (61 –
90 min).
In derselben Zeit verringerte sich die Temperatur der direkt bei 17 °C gehaltenen
Proben von 31,1 °C auf 20,6 °C. Die Abkühlraten betrugen 0,18 ± 0,05 °C/ min (0 –
30 min), 0,11 ± 0,05 °C/ min (31 - 60 min) und 0,07 ± 0,06 °C/ min (61 -90 min).
Die Lagerungstemperatur erreichten die Proben beider Abkühlungsverfahren
beinahe zeitgleich nach 246 min (Haltezeit bei 21 °C) und nach 252 min (direkte
Abkühlung bei 17 °C).
Der Temperaturverlauf ist in Abb. 34 dargestellt.

Ergebnisse
93
Abb. 34: Temperaturverlauf der bei Experiment 5 durchgeführten beschleunigten Abkühlung (32 °C – 17 °C) und der Abkühlung gemäß ZDS-Empfehlungen (32 °C – 21 °C – 17°C) der verdünnten Samenproben in 100 ml Ebersamenflaschen. Mittelwerte von je 3 Samenproben. Die Samenproben einstufig isotherm mit 32 °C warmer Beltsville Thawing Solution (BTS) auf die Endkonzentration 20 x 10
6 verdünnt. Anschließend kühlte ein Teil der Proben nach ZDS-Empfehlungen 90 min bei 21 °C
ab (blaue Kurve) und wurden bei 17 °C gelagert. Der andere Teil kühlte direkt nach der Verdünnung bei 17 °C ab (rote Kurve).
10
15
20
25
30
35
0 60 120 180 240
Tem
pera
tur
in °
C
Minuten
32°C - 21°C - 17°C 32°C - 17°C
direkte Lagerung bei 17 °C
90 min bei 21°C Lagerung bei 17°C

Ergebnisse
94
4.6.2 Casa-Motilitätsparameter
Abb. 35: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit der Abkühlraten und Lagerungsdauer; n=6 Ejakulate. Blaue Säulen: Abkühlung laut ZDS Empfehlungen; Nach Verdünnung mit 32 °C 90 min Abkühlung bei 21 °C, anschließend weitere Abkühlung und Lagerung bei 17 °C. Rote Säulen: Proben mit schneller Abkühlrate (Verdünnung bei 32 °C, direkte Abkühlung bei 17 °C)
Die Gesamtmotilität (Abb. 35) sowie die progressive Motilität wurden weder durch die
Art der Abkühlung noch durch die Lagerungsdauer beeinflusst. Die Werte bewegten
sich bei beiden Varianten in einem sehr engen Bereich.
Straightness (STR), linearity (LIN), wobble (WOB) und aplitude of lateral head-
displacement (ALH) wurden von der Lagerungsdauer beeinflusst (Tabelle 12).
0
10
20
30
40
50
60
70
80
90
100
d1 d3 d6
mo
tile
Sp
erm
ien
in %
Lagerungsdauer
Abkühlung 32 -21 - 17 Abkühlung 32 - 17

Ergebnisse
95
4.6.3 Membranintegrität
Abb. 36: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW + SD) in Abhängigkeit der Abkühlrate und Lagerungsdauer. Blaue Säulen: Abkühlung laut ZDS Empfehlungen; Nach Verdünnung mit 32 °C 90 min Abkühlung bei 21 °C, anschließend Lagerung bei 17 °C. Rote Säulen: Proben mit schneller Abkühlrate (nach Verdünnung bei 32 °C direkte Abkühlung bei 17 °C); n=6 Ejakulate. a, b: Unterschiede zwischen Lagerungstagen (d1 – d6) innerhalb derselben Abkühlgruppe (p < 0,05)
Der Anteil der PI- und FITC-PNA-negativen (plasma- und
akrosomenmembranintakter) Spermien wurde nur von der Lagerungsdauer
beeinflusst (p < 0,05).
Bei den Proben welche für 90 min bei 21 °C gehalten wurden reduzierte sich der
Anteil membranintakter Spermien von 86,8 ± 3,5 % nach 24 h auf 84,3 ± 3,5 % nach
72 h und auf 83,0 ± 2,7 % nach 144 h Lagerungsdauer.
Die direkt bei 17 °C abgekühlten Proben zeigten ähnliche Werte. An d 1 wurden
86,2 ± 3,3 % gemessen, dies erniedrigte sich auf 83,8 ± 2,9 % (d 3) und 81,3 ± 2,5 %
nach 6 Tagen Lagerung.
0
10
20
30
40
50
60
70
80
90
100
d1 d3 d6
Mem
bra
nin
takt
e Sp
erm
ien
in %
Lagerungsdauer
Abkühlung 32 -21 - 17 Abkühlung 32 - 17
a a b b b c

Ergebnisse
96
4.7 Experiment 6: Temperaturverlauf in der Prozesskette der
Samenverarbeitung auf einer Besamungsstation und Vergleich mit
Laborstandards
4.7.1 Temperaturverlauf während der Samenverarbeitung
Der Temperaturverlauf der einstufig isothermen Verdünnung (Laborstandard) wurde
in Kapitel 4.5.1 beschrieben.
In der Besamungsstation wurden die Ejakulate nach Ankunft im Labor mit auf 32 °C
temperierten BTS vorverdünnt und während der mikroskopischen Untersuchung und
vor der Abfüllung im Mittel 28 min bei 21 °C gehalten und kühlten dabei auf 29,5 °C
ab. Die anschließende Ausverdünnung wurde mit einem auf 26 °C angewärmten
Verdünner durchgeführt, danach erfolgte die Abfüllung in die 80 ml Tuben. Nach der
Abfüllung war die Temperatur der Tuben auf 24,5 °C abgekühlt. Für die nächsten
2,5 h wurden 40 Tuben in einer Plastikbox bei 21 °C gehalten. Die außen liegenden
Tuben kühlten in dieser Zeit auf 22,4 °C mit einer durchschnittlichen Abkühlrate von
0,02 ± 0,08 °C/ min ab. Die inneren Tuben erreichten 23,2 °C mit einer Rate von
0,01 ± 0,04 °C/ min. Anschließend wurden die Boxen in einen auf 17 °C temperierten
Klimaschrank verbracht. Die nach Laborstandard verarbeiteten und einzeln
abgekühlten Proben erreichten nach 3 h bei 17 °C die Lagerungstemperatur mit einer
Abkühlrate von 0,025 °C/ min. Im selben Zeitraum erreichten die außen liegenden
Tuben in der Box 20,7 °C mit einer Abkühlrate von 0,014 °C/ min und die innen
liegenden Tuben 19,4 °C mit einer Abkühlrate 0,012 °C/ min.
Am Ende des Versuchs, 7 h nach Beginn der Temperaturmessung, betrug die
Temperatur der außen liegenden Tuben 19,4 °C, die der innen liegenden Tuben
18,5 °C.
Für die Simulation der Verarbeitung der Station wurden die Proben nach der
isothermen Vorverdünnung im Verhältnis 1:1 für 30 min bei 21 °C gehalten. Die
Temperatur der Ausverdünnung und weiteren Haltetemperaturen wurde wie oben
beschrieben durchgeführt. Das Halten der abgefüllten Proben erfolgte in einer
Plastikbox mit 40 auf 24 °C vortemperierten Tuben.
Der Temperaturverlauf ist in Abb. 37 dargestellt.
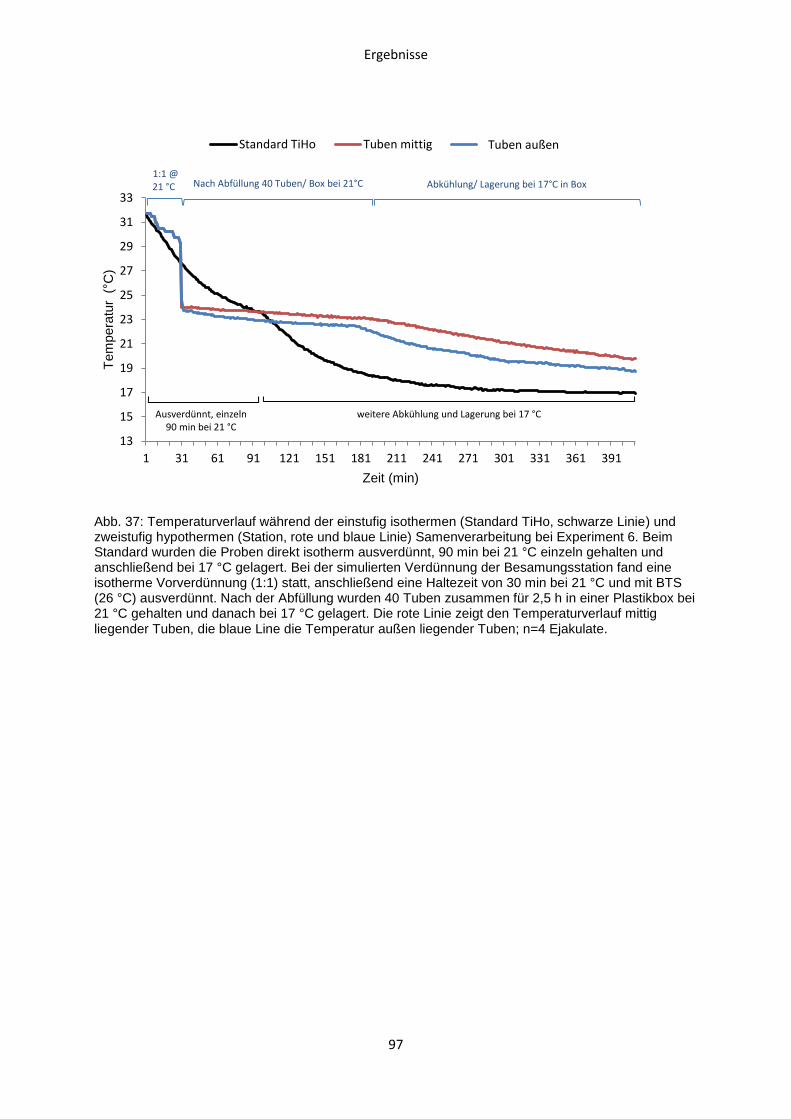
Ergebnisse
97
Abb. 37: Temperaturverlauf während der einstufig isothermen (Standard TiHo, schwarze Linie) und zweistufig hypothermen (Station, rote und blaue Linie) Samenverarbeitung bei Experiment 6. Beim Standard wurden die Proben direkt isotherm ausverdünnt, 90 min bei 21 °C einzeln gehalten und anschließend bei 17 °C gelagert. Bei der simulierten Verdünnung der Besamungsstation fand eine isotherme Vorverdünnung (1:1) statt, anschließend eine Haltezeit von 30 min bei 21 °C und mit BTS (26 °C) ausverdünnt. Nach der Abfüllung wurden 40 Tuben zusammen für 2,5 h in einer Plastikbox bei 21 °C gehalten und danach bei 17 °C gelagert. Die rote Linie zeigt den Temperaturverlauf mittig liegender Tuben, die blaue Line die Temperatur außen liegender Tuben; n=4 Ejakulate.
13
15
17
19
21
23
25
27
29
31
33
1 31 61 91 121 151 181 211 241 271 301 331 361 391
Tem
pera
tur
(°C
)
Zeit (min)
Standard TiHo Tuben mittig Tuben vorne
1:1 @ 21 °C Nach Abfüllung 40 Tuben/ Box bei 21°C Abkühlung/ Lagerung bei 17°C in Box
Ausverdünnt, einzeln 90 min bei 21 °C
weitere Abkühlung und Lagerung bei 17 °C
Tuben außen

Ergebnisse
98
4.7.2 CASA-Motilitätsparameter
Abb. 38: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Verarbeitungsart und Lagerungsdauer; n=6 Ejakulate. Standard (blaue Säule): einstufig isotherme Verdünnung (32°C), Abfüllung in 100ml Flaschen, Haltezeit 90 min bei 21°C, Lagerung bei 17°C. Station (rote Säule): zweistufige Verdünnung (32 °C, 26 °C), Abfüllung in 100 ml Flaschen, Positionierung mittig im Bündel von 40 auf 26°C temperierten Besamungstuben, Haltezeit 2,5h bei 21°C, Lagerung bei 17 °C.
Bei der Gesamtmotilität (Abb. 38) konnten keine Unterschiede festgestellt werden.
Die progressive Motilität (Tabelle 15) zeigte Unterschiede nach 72 h und 144 h
Lagerungsdauer. Die Werte der nach dem Standard verdünnten Proben waren mit
81,3 ± 7,3 % (d 3) signifikant geringer als die der Proben, die nach der Methode der
Besamungsstation verdünnt wurden. Diese zeigten noch 85,3 ± 6,0 %
vorwärtsbewegliche Spermien. An d 6 war mit 73,3 ± 17,3 % (Standard) und
76,8 ± 18,2 % (Station) ein signifikanter Unterschied zu sehen.
0
10
20
30
40
50
60
70
80
90
100
d 1 d 3 d 6
motile
Sperm
ien in %
Lagerungsdauer
TiHo
Station
Standard

Ergebnisse
99
4.7.3 Membranintegrität
Abb. 39: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW + SD) in Abhängigkeit von der Verarbeitungsart und Lagerungsdauer; n=6 Ejakulate. Standard (blaue Säule): einstufig isotherme Verdünnung (32°C), Abfüllung in 100ml Flaschen, Haltezeit 90 min bei 21°C, Lagerung bei 17°C. Station (rote Säule): zweistufige Verdünnung (32 °C, 26 °C), Abfüllung in 100 ml Flaschen, Positionierung mittig im Bündel von 40 auf 26°C temperierten Besamungstuben, Haltezeit 2,5h bei 21°C, Lagerung bei 17 °C.
Die Werte der membranintakten Spermien bewegten sich bei beiden Varianten in
einem sehr engen Bereich. Signifikante Unterschiede konnten nicht ermittelt werden.
0
10
20
30
40
50
60
70
80
90
100
d 1 d 2 d 3
Mem
bra
nin
takte
Sperm
ien in %
Lagerungsdauer
TiHo
Station
Standard

Ergebnisse
100
4.7.4 Spezifische Reaktivität gegenüber Bikarbonat (Calcium-Influx)
Abb. 40: Spezifische Reaktivität auf Bicarbonat berechnet anhand der Propidiumiodid-negativen (PI-neg.) und Fluo-3-negativen (Fluo-3-neg) Spermienpopulation nach 60 min Inkubationsdauer; n=6 Ejakulate. Darstellung der Werte (MW + SD) in Abhängigkeit von der Verarbeitungsart und Lagerungsdauer. Standard (blaue Säule): einstufig isotherme Verdünnung (32°C), Abfüllung in 100ml Flaschen, Haltezeit 90 min bei 21°C, Lagerung bei 17°C. Station (rote Säule): zweistufige Verdünnung (32 °C, 26 °C), Abfüllung in 100 ml Flaschen, Positionierung mittig im Bündel von 40 auf 26°C temperierten Besamungstuben, Haltezeit 2,5h bei 21°C, Lagerung bei 17 °C. a, b: signifikante Unterschiede (p < 0,05)
Nach 24 h Lagerung war die spezifische Reaktivität auf den Kapazitationsstimulus
Bikarbonat bei der Standard-Verarbeitung mit 31,7 ± 7,4 % signifikant niedriger als
bei der Verarbeitungsart der Station mit 40,3 ± 6,5 %.
0
10
20
30
40
50
60
70
80
90
100
spez.
Reaktivität, %
Sperm
ien
d 1
Lagerungsdauer
Standard
Station
b
a

Diskussion
101
5 Diskussion
Aufgrund ihrer Membranzusammensetzung sind Eberspermien besonders
empfindlich gegenüber Abkühlung. Die Flüssigkonservierung ist deshalb noch immer
mit Schwierigkeiten verbunden. Die initialen Schritte der Samenverarbeitung sind
dabei maßgeblich für die Qualität des konservierten Samens.
Ziel der vorliegenden Studie war es, den Einfluss von Haltezeiten zwischen 30 min
und 6 h bei 21 °C sowie 32 °C während der Frühphase der Konservierung auf die
Qualität gelagerten Eberspermas unter Anwendung gängiger Praxisverfahren zu
untersuchen. Dabei wurden neben spermatologischen Parametern, wie der Motilität
und Membranintegrität, mögliche Einflüsse auf den Energiestoffwechsel und das
bakterielle Wachstum betrachtet. Zusätzlich sollten praxisübliche
Verarbeitungsmethoden überprüft und Temperaturprofile während der
Verarbeitungskette erhoben werden.
Bei der zweistufig isothermen Verdünnung mit einem auf 32 °C temperierten
Verdünnter verläuft, erwartungsgemäß und wie die Temperaturmessungen
bestätigen, die initiale Abkühlung der Samenproben langsamer, als bei der zweistufig
hypothermen Verdünnung. Bei letzterer wurde die erste Verdünnung ebenfalls mit
32 °C warmem Verdünner und die zweite Verdünnung mit auf 21 °C temperierten
Verdünner durchgeführt. Es ist seit langem bekannt, dass langsames Abkühlen die
Resistenz der Eberspermien gegen Kälteschock erhöht (PURSEL et al., 1972; 1973).
Die deshalb aufgestellte Hypothese, dass Samenproben, die zweistufig isotherm
verdünnt wurden, eine bessere Qualität während der Lagerung aufweisen als
hypotherm verdünnte Proben, konnte vor allem nach längerer Lagerungsdauer,
sowie bei höheren Verdünnungsgraden von 10 x 106 Spermien/ ml bestätigt werden
(Exp. 1). Die Gesamtmotilität war bei den hypotherm verdünnten Proben der
vorliegenden Studie mit ca. 5 % signifikant niedriger als bei den isotherm verdünnten
Proben. Eine mögliche Ursache für die erhöhte Empfindlichkeit ist, dass bei der
hohen Verdünnung der Samenproben auf 10 Millionen Spermien/ ml ein geringerer
Anteil von Seminalplasma in den Proben vorhanden ist. Es ist bekannt, dass

Diskussion
102
Seminalplasma einen stabilisierenden Effekt auf die Spermienmembran von
Eberspermien hat, der sich vor allem beim Abkühlen der Samenzellen zeigt (LEAHY
u. GADELLA, 2011). Signifikante Unterschiede zwischen hypothermer und
isothermer Verdünnung und der positive Einfluss der isothermen Verarbeitung
konnten in einem Feldversuch belegt werden (SCHULZE et al., 2013). Hier konnten
bei der isothermen Variante höhere mitochondriale Aktivität und höhere Akrosomen-
und Membranintegrität der Samenzellen ermittelt werden. Weiterhin erwiesen sich
die isotherm verdünnten Samenzellen zudem besser in einem Thermoresistenztest
als die hypotherm verdünnten Spermien, was auf eine hohe Stressresistenz des
isotherm verdünnten Samens schließen lässt.
Bei der vorliegenden Studie zeigten sich tendenziell nach sechs Tagen Lagerung
auch bei den Proben mit 20 x 106 Spermien/ ml Veränderungen der Gesamtmotilität
und der progressiven Motilität bei den hypotherm verdünnten Proben; die Werte
sanken um 9 % im Vergleich zu den isotherm verdünnten Proben. Ähnliche
Ergebnisse wurden bei der Membranintegrität deutlich. Bei den höher ausverdünnten
Proben deuten sich die Unterschiede an allen Lagerungstagen an.
Da zurzeit noch eine Spermienkonzentration von ca. 20 Millionen Spermien/ ml als
weltweiter Standard gilt, wurden die folgenden Experimente mit dieser Konzentration
durchgeführt. Es gibt allerdings eine Tendenz zur Reduktion der Spermienzahl pro
Besamungsportion. Da dadurch die Empfindlichkeit der Spermien erhöht werden
kann, muss dabei auf das Temperaturregime noch genauer geachtet werden. Neben
der Art der Verdünnung sind Haltezeiten eine weitere Möglichkeit die
Abkühlgeschwindigkeit zu reduzieren und die Adaptionszeit für die Zellen zu
verlängern. Die Hypothese, dass Haltezeiten bis zu 60 min bei 32 °C, im Gegensatz
zu Haltezeiten bei 21 °C, die Qualität des gelagerten Samens verbessern, konnte im
Experiment 2 mit Einschränkung bestätigt werden.
Während der gesamten Versuchsdauer wurde hoch standardisiert und mit Sperma
auf hohem Qualitätsniveau gearbeitet. Dies wurde vor allem nach längerer
Lagerungsdauer deutlich, denn an Tag 6 waren bei allen Samenproben noch über
80 % der Spermien motil. Unter anderen, weniger optimalen Voraussetzungen, wie

Diskussion
103
sie in der Praxis vorherrschen können, wie beispielsweise eberindividuelle
Kälteschocktoleranz, saisonal herabgesetzte Spermaqualität und höhere
Verdünnungsgrade bei hochkonzentriertem Nativsperma sind größere Abweichungen
der Messergebnisse zu erwarten. Die Haltetemperatur beeinflusste die Gesamt- und
progressive Motilität zwischen d 0 und d 3 (n=12), allerdings konnten nur marginale
signifikante Unterschiede gemessen werden.
Bei der Analyse der Subpopulationen weiterer kinematischer Parameter zeigte sich,
dass bei einigen Parametern Unterschiede des Bewegungsmusters zwischen den
frisch verdünnten Proben an d 0 und den gelagerten Proben auftraten. Vor allem die
für 6 h bei 21 °C gehaltenen Proben wichen an d 0 vom Kurvenverlauf der anderen
Proben ab. Am deutlichsten wurde dies bei den Parametern velocity curved line
(VCL), wobble (WOB), linearity (LIN) und straightness (STR). Bereits nach 24 h
Lagerungsdauer zeigten die Kurven einen einheitlicheren Verlauf. Die Ergebnisse
deuten auf einen so genannten „Verdünnungsschock“ hin, der durch eine initiale
Erhöhung der Zellaktivität charakterisiert ist, was zu Motilitätsverlust und
Membranschäden führen kann. Es gibt keine eindeutigen Erklärungen, aber es wird
vermutet, dass dies durch den plötzlichen Milieuwechsel hervorgerufen wird
(WATSON, 1995; JOHNSON et al., 2000). Der temporäre Einfluss der Verdünnung
an d 0 scheint sich vor allem bei den für 6 h bei 21 °C gehaltenen Proben zu zeigen.
Weiterhin wurden alle Proben nach der Haltezeit vor der Motilitätsmessung auf 38 °C
erwärmt. Die bei 21 °C gehaltenen Proben waren, im Gegensatz zu den anderen in
dieser Analyse berücksichtigten Proben, nach 6 h bereits auf 21 °C abgekühlt; die
erneute Erwärmung stellt einen zusätzlichen Einfluss auf die Samenzellen dar, der
aber offensichtlich ebenfalls temporär zu sein scheint.
Die Hypothese, dass sich Haltezeiten, bei 21 °C und vor allem bei 32 °C, von mehr
als 60 min negativ auf den Energiestoffwechsel auswirken, konnte nicht bestätigt
werden (Exp. 2). Der Adenylat-Gehalt der Spermien wurde weder unmittelbar nach
der Haltezeit noch nach Lagerung bis zu 144 h beeinflusst. Die vorliegende Studie
zeigte ebenfalls, dass Eberspermien über einen sehr effizienten Energiestoffwechsel
verfügen, der von Haltezeiten bis zu 6 h, Haltetemperatur bis zu 32 °C und der

Diskussion
104
Lagerung bei 17 °C über sechs Tage nicht beeinflusst wird. In der Literatur gibt es
unterschiedliche Angaben bezüglich der Stabilität des ATP-Gehalts während der
Lagerung. GOGOL et al. (2009) berichteten von einer Verringerung des ATP-Gehalts
während der Lagerung nach vier und sieben Tagen bei 15 °C. LONG und GUTHRIE
(2006) dagegen zeigten stabile Werte über fünf Tage Lagerungsdauer bei 17 °C. Es
ist allerdings schwierig, die Ergebnisse der ATP-Analysen unterschiedlicher Studien
zu vergleichen, da unterschiedliche Methoden angewendet wurden.
Die Energieladung (EC) zeigte sich während der Versuche der vorliegenden Studie
als sehr stabil; während der gesamten Versuchsdauer wurde dieser Parameter nicht
von der Haltezeit, -temperatur oder Lagerungsdauer beeinflusst und der Wert von 0,6
wurde nicht unterschritten. Dies bestätigt die Aussage von CHULAVATNATOL et al.
(1977), die bei humanen Samenzellen zeigten, dass die Energieladung auch unter
Stressbedingungen stabil bleibt. Die Beobachtung von KAMP et al. (2003), dass
eine Haltezeit von 60 min bei 35 °C einen Abfall der Energieladung auf 0,3
verursachte, konnte in den eigenen Versuchen nicht bestätigt werden. Die
Energieladung sank wie oben erwähnt auch nach einer Haltezeit 6 h bei 32 °C nicht
unter 0,6. Vermutlich ist dieser Unterschied durch die Verfügbarkeit von Sauerstoff
für die Samenzellen erklärbar. Die Haltezeiten erfolgten in der vorliegenden Studie
unter aeroben Bedingungen, bei KAMP et al. (2003) dagegen fanden diese unter
anaeroben Bedingungen statt. Vor allem bei höheren Temperaturen scheint die
Sauerstoffzufuhr essentiell für den Energiehaushalt der Samenzellen zu sein, da bei
der anaeroben Lagerung der Samenproben bei 15 – 17 °C Werte um 0,6 gemessen
werden konnten.
Um die mitochondriale ATP-Produktion aufrechtzuerhalten, ist ein elektrochemischer
Protonengradient innerhalb der Mitochondrien notwendig. Dieses sogenannte
mitochondriale Membranpotential (MMP) entsteht bei der Elektronenkopplung
innerhalb der oxidativen Phosphorylierung (CRAMER u. KNAFF, 1990).
Durchflusszytometrisch konnten lebende Zellen mit hohem MMP von denen mit
niedrigem MMP, also mit weniger bis nicht aktiven Mitochondrien mit Hilfe des
Farbstoffs JC-1 unterschieden werden (COSSARIZZA et al., 1993). Das MMP wurde

Diskussion
105
in den eigenen Versuchen weder von der Haltezeit noch von der Haltetemperatur
beeinflusst.
Die Qualität der Samenproben hängt nicht nur von der Vitalität der Samenzellen ab,
sondern ist auch abhängig vom mikrobiellen Status, der vor allem durch das Halten
bei höheren Temperaturen beeinflusst werden könnte. Durch das natürliche
Vorkommen von Bakterien im männlichen Genitaltrakt und möglichen Keimeintrag
aus der Umwelt ist es unvermeidlich, dass Ejakulate bakteriell belastet sind.
Bakteriospermie kann zu Reduktion der Motilität, Membranschäden und vermehrter
Agglutination der Zellen führen. Der negative Einfluss der Bakterien ist abhängig von
der Keimart und -konzentration und kann die Haltbarkeit der Portionen beeinflussen
(ALTHOUSE u. LU, 2005).
Da sich die Wachstumsrate von Bakterien durch Inkubation bei Temperaturen nahe
der Körpertemperatur rasant erhöhen kann (ALTHOUSE et al., 2008), war eine
Hypothese, dass sich eine Inkubation bei 32 °C von im Verhältnis 1:1 verdünnten
Ejakulaten negativ auf die mikrobielle Qualität der Samenproben auswirkt, zumal bei
einer Verdünnung in diesem Verhältnis nur eine geringe Konzentration des
Antibiotikums im Verdünner vorliegt (Exp. 2). Diese Annahme konnte jedoch nicht
bestätigt werden. Im frischen Ejakulat befanden sich im Mittel erwartungsgemäß ca.
5 x 104 KbE/ ml. Nach 60 min Haltezeit war der Keimgehalt bei 32 °C und 21 °C im
Vergleich zur Kontrolle unverändert und sogar nach 6 h Haltezeit blieben die Werte
bei beiden Temperaturen im geringgradigen Bereich von unter 1 x 103 KbE/ ml. In
den ausverdünnten Proben wurde nach drei Tagen Lagerungsdauer weder bei den
Kontrollproben noch bei den für 60 min gehaltenen Proben (beider Temperaturen)
Keimwachstum festgestellt. Die Werte der für 6 h gehaltenen Probe wiesen einen
geringgradigen Keimgehalt von 138 KbE/ ml (32 °C) und 50 KbE/ ml (21 °C) auf.
Berücksichtigt werden muss, dass das Keimspektrum hinsichtlich
Wachstumstemperaturoptimum und Antibiotika-Empfindlichkeit variieren kann. Bei
resistenten Bakterien muss davon ausgegangen werden, dass sich eine Haltezeit,
vor allem bei 32 °C, negativ auf die mikrobielle Qualität auswirkt. Resistente Keime
stellen aber ohnehin in der Ebersamenverarbeitung und –lagerung ein Problem dar;

Diskussion
106
ein vermehrtes Wachstum ist auch ohne Haltezeiten zu erwarten (ALTHOUSE et al.,
2008).
Die Hypothese, dass die Ausverdünnung mit einem im Vergleich zur Samenprobe
hyperthermen Verdünner (32 °C) einen negativen Einfluss auf die Samenqualität hat
konnte nicht bestätigt werden (Exp. 3). Die Temperaturerhöhungen von Proben, die
bereits 28 – 21 °C abgekühlt waren, scheinen keine großen Veränderungen der
Membranen zu verursachen. Lediglich Proben, die bereits vollständig auf 21 °C
abgekühlt waren, zeigten bei langer Lagerung (144 h) leicht verringerte Werte der
membranintakten Spermien, was auf eine ausgeprägte Toleranz der Spermien
gegenüber höheren Temperaturen hinweist.
Die Hypothese, dass Haltezeiten von 30 min, 60 min, 3 h und 6 h bei 21 °C und
32 °C auf die bereits ausverdünnten Samenproben einen positiven Einfluss haben,
konnte ebenfalls nicht bestätigt werden (Exp. 4). Die Haltezeiten hatten keinen
Einfluss auf die standardspermatologischen Parameter. Ein Einfluss der
Haltetemperatur auf Motilitätsparameter konnte bei den Proben, die nach einstufig
isothermer (Exp. 4 b) oder zweistufig hypothermer (Exp. 4 a) Ausverdünnung
gehalten wurden nur tendenziell gezeigt werden.
PURSEL et al. (1973 a) stellten fest, dass die Proben, die vor den Haltezeiten
ausverdünnt wurden, eine höhere Kälteschockempfindlichkeit aufwiesen, als Proben,
die erst nach den Haltezeiten ausverdünnt wurden. Die Autoren schlossen daraus,
dass durch die Verdünnung die Kälteempfindlichkeit begünstigt werden kann und mit
der Ausbildung der Kälteschockresistenz interferiert. In den Versuchen der
vorliegenden Studie konnte dieser Unterschied nicht bestätigt werden. Ein direkter
Vergleich zu den Versuchen von PURSEL et al. (1973) ist aber schwer zu ziehen, da
die Versuchsbedingungen von denen der vorliegenden Studie abwichen. Der größte
Unterschied ist, dass die Autoren den Kälteschock bei 5 °C bei einem Aliquot von nur
1 ml durchführten. Damit induzierten sie drastischere Effekte, die sich in einem
drastischen Motilitätsverlust widerspiegeln (max. Motilität: 55 %). In der vorliegenden
Studie galt es jedoch, den Einfluss subtiler Temperaturabsenkungen, wie sie unter

Diskussion
107
Praxisbedingungen gegeben sind, mit einem erweiterten Methodenspektrum zu
untersuchen. Es wurden bei keiner Probe weniger als 80 % bewegliche Spermien
gemessen; die Motilität ist demnach ein robuster Parameter unter den vorliegenden
Versuchsbedingungen.
Die Hypothese, dass eine schnelle Abkühlrate einstufig isotherm verdünnter
Samenproben während der Frühphase der Konservierung einen negativen Einfluss
auf die Qualität der gelagerten Samenproben hat, konnte tendenziell bestätigt
werden (Exp. 5). Die ermittelten Werte lagen auch bei diesem Versuch auf einem
sehr hohen Niveau, so dass sich entgegen früheren Versuchen keine signifikanten
Unterschiede zeigten. Die schnelle Abkühlrate, die durch die direkt an die
Ausverdünnung anschließende Haltezeit bei 17 °C hervorgerufen wurde, verringerte
die Motilität nach 144 h Lagerung um 6 % im Vergleich zur Messung nach 72 h. Im
gleichen Zeitraum verringerte sich bei den langsam abgekühlten Proben die Motilität
nur um 3 %. Zahlenmäßig deutlicher wurde der Unterschied bei der progressiven
Motilität, die in diesem Zeitraum durch die schnelle Abkühlung um 13 % abnahm.
Ähnliche Tendenzen zeigten sich in der Membranintegrität der Samenzellen.
Dass bei Lagerungstemperaturen unter 17 °C der Einfluss von Haltezeiten deutlicher
wird, zeigten PURSEL et al. (1973 b). Es ist außerdem anzunehmen, dass
beispielsweise durch eberindividuelle Unterschiede und weniger standardisierte
Bedingungen bei Temperatur- und Zeitmanagement größere Unterschiede messbar
sind.
In einem Feldversuch konnte gezeigt werden, dass bei Besamungsstationen, die die
zweistufig hypotherme Verdünnung anwenden, die Temperatur des zweiten
Verdünners zwischen 21 °C und 27 °C variierte (SCHULZE et al., 2013). Bei der in
der vorliegenden Studie beteiligten Besamungsstation wurde die zweite Verdünnung
mit einem auf 26 °C temperierten Verdünner durchgeführt. Eine Aussage darüber, ob
die hypotherme Verdünnung in der Praxis zur Qualitätsminderung der
Samenportionen führt, konnte deshalb im eigenen Praxisversuch (Exp. 6) nur
eingeschränkt getroffen werden. Da das vorverdünnte Sperma zu diesem Zeitpunkt

Diskussion
108
eine Temperatur von 29,5 °C aufwies, entsprach das Vorgehen nicht einer mit dem
eigenen Laborstandard vergleichbaren hypothermen Verdünnung. Ein weiterer
Unterschied bestand darin, dass die Tuben nicht wie im Labor einzeln mit Abstand
zueinander, sondern alle Tuben eines Ebers (20 – 60 Stück) in einer Plastikbox
liegend abkühlten. Im Vergleich zum Temperaturverlauf im Labor wird deutlich, dass
die Samenproben in der Station deutlich langsamer abgekühlt wurden. Vor allem die
Temperatur der Tuben, die sich in der Mitte der Box befanden, differierte zeitweise
bei gleicher Versuchsdauer bis zu 5 °C im Vergleich zur einstufig isothermen
Verdünnung im Labor. Die außen liegenden Tuben kühlten etwas schneller ab, aber
auch hier wurde während der Versuchsdauer von ca. 7 h die Lagerungstemperatur
nicht erreicht.
Bei dem im Labor nachgestellten Vergleich waren die Membranintegrität und die
Gesamtmotilität bei der langsameren Abkühlung in der Praxis tendenziell besser als
beim Laborstandard, bei der progressiven Motilität konnten nach drei Tagen
Lagerung sogar signifikante bessere Ergebnisse bei der Praxisvariante gemessen
werden. Zudem war die spezifische Reaktivität der Spermien auf den
Kapazitatationstimulus Bicarbonat, die wie in der Besamungsstation verarbeitet
wurden, nach 24 h Lagerung um 9 % höher als bei der Standardverarbeitung des
Labors. Insgesamt kann also gesagt werden, dass die langsamere Abkühlung der
gebündelten Samenportionen unter Praxisbedingungen vorteilhafter ist für die
Samenqualität als der Laborstandard.
Bei den vorliegenden Experimenten wurde akribisch auf die Einhaltung eines
standardisierten Temperaturverlaufs geachtet. Temperaturprofile von Eberejakulaten
während der Samenverarbeitung sind in der Literatur bisher nur sehr spärlich
beschrieben. Ein Augenmerk dieser Arbeit lag deshalb darauf, den Verlauf der
Abkühlung jeder Verarbeitungsvariante im Labor zu dokumentieren. Weiterhin wurde
die Temperaturkurve der Standardverarbeitungsvariante unter Laborbedingungen mit
dem Temperaturverlauf der Verarbeitung in einer Besamungsstation unter
Praxisbedingungen verglichen. Ein wesentlicher Unterschied zwischen der
Spermaverarbeitung unter Standardbedingungen im Labor und in

Diskussion
109
Besamungsstationen liegt in der Aufbereitung der Anzahl von Besamungsportionen
pro Ejakulat und dem anschließender Versand.
Wie bereits erwähnt, ist ein Argument für die zweistufig hypotherme Verdünnung in
der Praxis, dass der schnellere Temperaturabfall, der durch den zweiten, um ca.
10 °C kälteren Verdünner hervorgerufen wird, dazu führt, dass die
Lagerungstemperatur von 17 °C schneller erreicht wird. Dadurch soll der Versand
zeitnah zur Verarbeitung bei konstanter Temperatur ermöglicht werden und das
Risiko von bakteriellem Wachstum gesenkt werden. Zusätzlich werden durch die
niedrigere Temperatur Energiekosten eingespart. Außerdem scheint es die
Erwartungshaltung einiger Landwirte zu geben, dass frische Samenproben bei
Ankunft auf dem Betrieb auf 17 °C abgekühlt sein müssen und wärmere
Probentemperaturen einen Qualitätsmangel bedeuten können.
Die Temperaturmessungen des Laborstandards ergaben allerdings, dass trotz des
schnellen Abfalls bei der hypothermen Verdünnung eine Temperatur von 20 °C nicht
vorzeitig, sondern zeitgleich mit der isothermen Verdünnung nach 150 min erreicht
wird (Exp. 1). Folglich wurde die Lagerungstemperatur ebenfalls nach der gleichen
Zeitspanne erreicht. Erklärbar ist dies, da nach der hypothermen Verdünnung die
Temperatur der Proben (23 °C) relativ nahe der Umgebungstemperatur (21 °C) ist.
Dadurch verringert sich deren Abkühlrate und die Zeitspanne bis zum Erreichen der
Temperatur verlängert sich.
Das gleiche Bild zeigte sich sogar bei den Proben, die direkt nach der isothermen
Verdünnung bei 17 °C, statt zunächst bei 21 °C, abgekühlt wurden (Exp. 5). Sie
erreichten zeitgleich mit den Proben, die zunächst 90 min bei 21 °C und erst dann
bei 17 °C gelagert wurden, die Lagerungstemperatur von 17 °C nach ca. 210 min.
Schlussfolgernd sind Haltezeiten bis zu 60 min des 1:1 vorverdünnten als auch des
ausverdünnten Samens, zwischen 21 °C und 32 °C, vorteilhaft für die Samenqualität.
Temperaturen von 32 °C beeinträchtigen die Samenqualität nicht. Das Halten des
1:1 verdünnten Samens bis zu 6 h bei 32 °C hatte dabei keinen Einfluss auf den
Energiestoffwechsel der Spermien während der Lagerung. Allerdings könnten

Diskussion
110
Haltezeiten über 60 min je nach Keimbelastung und Bakterienart der Proben kritisch
für den Keimgehalt während der Lagerung sein.
Entgegen bisheriger Annahmen, führt hypotherme Verdünnung von Eberspermien,
mit auf 21 °C temperierten Verdünner, weder zum schnellen Erreichen der
Lagerungstemperatur, geringerer Keimzahl noch zu höherem Energiegehalt der
Spermien. Da nachgewiesen werden konnte, dass isotherme Verdünnung und ein
langsames Abkühlen zu verbesserter Qualität der Besamungsportionen führt, sollte
die Erwartungshaltung der Landwirte dementsprechend korrigiert werden. Der
Nachteil, dass das Erwärmen des Verdünners für die isotherme Verdünnung die
Energie- und damit Produktionskosten erhöht, könnte durch die bessere
Produktqualität und Kundenzufriedenheit ausgeglichen werden.

Zusammenfassung
111
6 Zusammenfassung
Ulrike Wallner (2015):
Einfluss von Haltezeiten während der Frühphase der Konservierung auf den
Energiemetabolismus und die Qualität von Eberspermien
Ziel der vorliegenden Studie war es, den Einfluss von Haltezeiten zwischen 30 min
und 6 h bei 21 °C sowie 32 °C während der Frühphase der Konservierung auf die
Qualität gelagerten Eberspermas unter Anwendung gängiger Praxisverfahren zu
untersuchen. Dabei wurden neben spermatologischen Parametern, wie der Motilität
und Membranintegrität, mögliche Einflüsse auf den Energiestoffwechsel und das
bakterielle Wachstum betrachtet. Zusätzlich sollten die praxisüblichen
Verarbeitungsmethoden überprüft und Temperaturprofile während der
Verarbeitungskette erhoben werden.
Es wurden sechs Experimente durchgeführt. Bei allen Experimenten wurden die
Motilität der Spermien mittels CASA und der Anteil membranintakter Spermien
mittels Durchflusszytometrie bestimmt. In Experiment 2 wurde zudem mit dem
Fluoreszenzfarbstoff JC-1 durchflusszytometrisch das Mitochondrienmembran-
potential ermittelt, sowie der Nukleotidgehalt (ATP, ADP, AMP) und die
Energieladung der Samenzellen gemessen. Ein weiteres Augenmerk wurde darauf
gelegt, den genauen Verlauf der Abkühlung jeder Verarbeitungsvariante im Labor zu
dokumentieren.
Im ersten Experiment wurden zunächst die zweistufig isotherme Verdünnung mit der
zweistufig hypothermen Verdünnung bei zwei Verdünnungsgraden,
10 x 106 Spermien/ ml und 20 x 106 Spermien/ ml verglichen. Während der Lagerung
bei 17 °C wiesen die isotherm verdünnten Proben eine bessere Qualität auf. Dies
zeigte sich vor allem beim höheren Verdünnungsgrad.
In den Experimenten 2 bis 4 wurden daraufhin die Auswirkungen von Haltezeiten für
30 min, 60 min, 3 h und 6 h bei 32 °C und 21 °C bei verschiedenen
Verarbeitungsbedingungen geprüft. Die Haltezeiten beeinträchtigten dabei die
Spermienqualität, den Adenylat-Gehalt sowie das Mitochondrienmembranpotential
nicht. Dies deutet darauf hin, dass Eberspermien über einen sehr effizienten

Zusammenfassung
112
Energiestoffwechsel verfügen. Das Keimwachstum wurde durch die Haltezeiten bis
zu 60 min des 1:1 verdünnten Samens nicht negativ beeinflusst.
Im fünften Experiment wurden die Auswirkungen einer beschleunigten Abkühlrate
des ausverdünnten Samens nach isothermer Verdünnung untersucht. Die schnelle
Abkühlrate bei einer direkten Abkühlung bei 17 °C wirkte sich tendenziell negativ auf
die Motilität aus. Allerdings wurden hier aufgrund des hohen spermatologischen
Niveaus nur geringe Schwankungen der Werte ermittelt.
Im abschließenden Experiment wurde im ersten Schritt der Temperaturverlauf der
Prozesskette der hypothermen Samenverarbeitung auf einer Besamungsstation mit
dem Verlauf des isotherm durchgeführten Laborstandards verglichen. Die
Samenportionen in der Besamungsstation kühlten aufgrund der höheren Temperatur
der Verdünner und der Abkühlung in Bündeln von ca. 40 – 60 Tuben langsamer ab
als die des Laborstandards. Im zweiten Schritt wurden die beiden
Verarbeitungsverfahren im Hinblick auf die Spermienqualität verglichen, wobei die
Motilität und Membranintegrität sich bei der langsamen Abkühlung der
Besamungsstation als tendenziell besser erwiesen.
Die Temperaturmessungen der Verdünnungsverfahren ergaben bei dem Vergleich
der zweistufig hypothermen mit der zweistufig isothermen Verdünnung, dass
entgegen den Erwartungen durch den hypothermen Verdünner, eine schnellere
Abkühlung auf die Lagerungstemperatur nicht erreicht wird. Bei beiden Methoden
wurde diese nach der gleichen Zeitspanne erreicht.
Schlussfolgernd sind Haltezeiten bis zu 60 min bei 32 °C und 21 °C sowohl des 1:1
vorverdünnten als auch des ausverdünnten Samens vorteilhaft für die Samenqualität;
verlängerte Haltezeiten führen nicht zum Energiedefizit der konservierten Spermien.
Allerdings könnten Haltezeiten über 60 min je nach Keimbelastung und Bakterienart
kritisch für den Keimgehalt während der Lagerung sein. Es konnte nachgewiesen
werden, dass eine isotherme Verdünnung und ein langsames Abkühlen zu einer
verbesserten Qualität der Besamungsportionen führt. Das derzeit in der Praxis noch
häufig durchgeführte zweistufig isotherme Verfahren ist aus spermatologischer Sicht
nicht empfehlenswert.

Summary
113
7 Summary
Ulrike Wallner (2015):
Effect of holding times during early stage of preservation on energy
metabolism and quality of boar spermatozoa
The aim of the present study was to test the effect of holding times between 30 min
and 6 h at 21 °C as well as 32 °C during the early stage of semen preservation on
semen quality during storage. Spermatology parameters such as motility and
membrane integrity as wells as possible influences on energy metabolism and
bacterial growth were examined. Additionally, current routine procedures in artificial
insemination (AI) laboratories were reviewed and temperature curves during semen
processing were determined.
In total, six experiments were performed. In all experiments the sperm motility was
determined by CASA and the proportion of membrane intact sperm was analyzed
using flow cytometry. In the second experiment, additionally the mitochondrial
membrane potential was determined by flow cytometry using the fluorescent dye
JC-1. Moreover, the content of nucleotides (ATP, ADP and AMP) and the energy
charge of the sperm cells were measured. Special attention was paid to document
the exact cooling curves for each variation of semen processing in all laboratory
experiments.
In the first experiment the two-step isothermic dilution was compared with the two-
step hypothermic dilution using two dilution rates, namely 10 x 106 sperm/ ml and
20 x 106 sperm/ ml. As a result, isothermic diluted samples showed a higher semen
quality during storage at 17 °C, especially at higher dilution level.
In Experiments 2 to 4 the effects of holding times for 30 min, 60 min, 3 h and 6 h at
32 °C and 21 °C were examined using different processing conditions.
Holding times neither had influence on semen quality nor on adenylate content and
mitochondrial membrane potential. This suggests that boar sperm energy metabolism
is very efficient. Bacterial growth was not negatively affected by holding times of the
1:1 pre-diluted semen up to 60 min.

Summary
114
In the fifth experiment, the effect of an accelerated cooling rate after isothermic
dilution semen was studied. Rapid cooling rates to the storage temperature at17 °C
tend to show adverse effects on motility. However, due to the high sperm quality level
only minor variations between the experimental groups were found.
In the sixth experiment the temperature profile of hypothermically processed semen
in an AI center was compared with the profile of the isothermically processed
laboratory standard. Due to the higher temperature of the diluent and the packing of
40 to 60 semen tubes in bundles, semen doses in the AI-center cooled down slower
than the single tube samples used in the laboratory standard. In the second step of
this experiment, the two methods were compared in terms of semen quality. The
results showed that semen processed with a slower cooling rate as in the AI-center
tend to have better motility parameters and membrane integrity compared to the
laboratory standard.
Contrary to expectations, the comparison of the temperature profiles of two-step
isothermic and two-step hypothermic dilution showed that hypothermic diluted
samples did not reach the final storage temperature earlier. Thus, in both dilution
methods, the storage temperature of 17°C was reached after the same period of
time.
In conclusion, holding times up to 60 min at 32 °C and 21 °C of prediluted semen and
semen diluted to final concentration are beneficial for semen quality; prolonged
holding times do not impair energy metabolism of stored semen. However,
depending on germ load and the bacterial species, prolonged holding times up to 6 h
may lead to moderately increased bacterial growth during storage. It is demonstrated
that isothermic dilution and retarded cooling results in higher sperm quality. As a
consequence, the in AI practice commonly used two-step isothermic method is no
longer recommended.

Literaturverzeichnis
115
8 Literaturverzeichnis
ALTHOUSE, G., M. WILSON, C. KUSTER, u. M. PARSLEY (1998): Characterization of lower temperature storage limitations of fresh-extended porcine semen. Theriogenology, 50, 535–543 ALTHOUSE, G.C. u. K.G. LU (2005): Bacteriospermia in extended porcine semen. Theriogenology, 63, 573–84 ALTHOUSE, G.C., M.S. PIERDON u. K.G. LU (2008): Thermotemporal dynamics of contaminant bacteria and antimicrobials in extended porcine semen. Theriogenology, 70, 1317–1323 ARMSTRONG, J.S., M. RAJASEKARAN, W. CHAMULITRAT, P. GATTI, W.J. HELLSTROM, u. S.C. SIKKA (1999): Characterization of reactive oxygen species induced effects on human spermatozoa movement and energy metabolism. Free Rad. Biol. Med., 26, 869-880 ATKINSON, D.E. (1968): The energy charge of the adenylate pool as a regulatory parameter. Interaction with feedback modifiers. Biochemistry, 7, 4030–4034 ATKINSON, D.E. u. G.M. WALTON (1967): Communications : Adenosine Triphosphate Conservation in Metabolic Regulation: Rat liver citrate cleavage enzyme. J. Biol. Chem., 242, 3239-3241 BOHNENSACK, R. u. W. HALANGK (1986): Control of respiration and of motility in ejaculated bull spermatozoa. Biochim. Biophys. Acta, 850, 72–79 BUTLER, W.J. u. T.K. ROBERTS (1975): Effects of some phosphatidyl compounds on boar spermatozoa following cold shock or slow cooling. J. Reprod. Fertil., 43, 183–187 CANVIN, A.T. u. M.M BUHR (1989): Effect of temperature on the fluidity of boar sperm membranes. J. Reprod. Fertil., 85, 533–540

Literaturverzeichnis
116
CASAS, I. u. G.C. ALTHOUSE (2013): The protective effect of a 17°C holding time on boar sperm plasma membrane fluidity after exposure to 5°C. Cryobiology, 1, 69-75 CEROLINI, S., A. MALDJIAN, P. SURAI, u. R. NOBLE (2000): Viability, susceptibility to peroxidation and fatty acid composition of boar semen during liquid storage. Anim. Reprod. Sci., 58, 99-111 CHAPMAN, A.G., L. FALL, E. ATKINSON, u. D.E. ATKINSON (1971): Adenylate energy charge in escherichia coli during growth and starvation. J. Bacteriol., 108, 1072–1086 CHULAVATNATOL, M. u. A. HAESUNGCHARERN (1977): Stabilistation of adenylate energy charge and its relation to human sperm motility. J. Biolog. Chem., 22, 8088-8091 COSSARIZZA, A., M., BACCARANI-CONTRI, G. KALASHNIKOVA, C. FRANCESCHI, (1993): A new method for the cytofluorimetric analysis of mitochondrial membrane potential using the j-aggregate forming lipophilic cation 5,5’-6,6'-tetrachloro-1,1',3,3' tetraethylbenzimidazolcarbocyanine iodide (JC-1). Biochemical and biophysical research communications, 197, 40-45 CRAMER W. u. D. KNAFF (Hrsg.) (1990): Energy transduction in biological membranes. A textbook of bioenergetics. Springer-Verlag New York, Inc DARIN-BENNETT, A. u. I.G. WHITE (1977): Influence of the cholesterol content of mammalian spermatozoa on susceptibility to cold-shock. Cryobiology, 14, 466–470 DE LEEUW, F.E., B. COLENBRANDER, u. A.J. VERKLEIJ (1991): The role membrane damage plays in cold shock and freezing injury. Reprod. Domest. Anim., Suppl.1, 95 – 104 DE LEEUW, F.E., H.C. CHEN, B. COLENBRANDER, u. A.J. VERKLEIJ (1990): Cold-induced ultrastructural changes in bull and boar sperm plasma membranes. Cryobiology, 27, 171–183 DROBNIS, E.Z., L.M. CROWE, T. BERGER, T.J. ANCHORDOGUY, u. J.W. OVERSTREET, u. J.H. CROWE (1993): Cold shock damage is due to lipid phase transitions in cell membranes: A demonstration using sperm as a model. J. Exp. Zool., 265, 432–437

Literaturverzeichnis
117
DZIEKONSKA, A., L. FRASER u. J. STRZEZEK (2009): Effect of different storage temperatures on the metabolic activity of spermatozoa following liquid storage of boar semen. J. Anim. Feed. Sci., 18, 638-649 ERIKSSON, B.M., J.M. VAZQUEZ, E.A. MARTINEZ, J. ROCA, X. LUCAS, u. H.R. MARTINEZ (2001): Effects of holding time during cooling and of type of package on plasma membrane integrity, motility and in vitro oocyte penetration ability of frozen-thawed boar spermatozoa. Theriogenology, 55, 1593–1605 FAWCETT, D.W. (1975): The Mammalian Spermatozoon. Dev. Biol., 7, 394–436 FORD, S.R. u. F.R. LEACH (1998): Bioluminescent assay of the adenylate energy charge. Methods Mol. Biol., 102, 69–81 FORD, W.C.L. (2006): Glycolysis and sperm motility: Does a spoonful of sugar help the flagellum go round? Hum. Reprod. Update, 12, 269–274 GOGOL, P., B. SZCZESNIAK-FABIANCZYK u. A. WIERZCHOS-HILCZER (2009): The photon emission, ATP level and motility of boar spermatozoa during liquid storage. Reprod. Biol., 9, 39-49 GUTHRIE, H.D. u. G.R. WELCH (2005): Impact of storage prior to cryopreservation on plasma membrane function and fertility of boar sperm. Theriogenology, 63, 396–410. GUTHRIE, H.D., G.R. WELCH u. J.A. LONG (2008): Mitochondrial function and reactive oxygen species action in relation to boar motility. Theriogenology, 70, 1209-1215 HENNING, H., A.M. PETRUNKINA, R.A.P. HARRISON u. D. WABERSKI (2012): Bivalent response to long-term storage in liquid-preserved boar semen: a flow cytometric analysis. Cytometry Part A, 81 A, 576-587 JOHNSON, L.A., K.F. WEITZE, P. FISER u. W.M. MAXWELL (2000): Storage of boar semen. Anim. Reprod. Sci., 62, 143–72

Literaturverzeichnis
118
KAMP, G., G. BÜSSELMANN, N. JONES, B. WIESNER u. J. LAUTERWEIN (2003): Energy metabolism and intracellular pH in boar spermatozoa. Reproduction, 126, 517-525 KUMARESAN, A., G. KADIRVEL, K.M BUJARBARUAH, R.K.BARDOLOI, A. DAS, S.KUMAR, u. S. NASKAR (2009): Preservation of boar semen at 18 °C induces lipid peroxidation and apoptosis like changes in spermatozoa. Anim. Reprod. Sci., 110, 162-171 LADBROOKE, B.D. u. D. CHAPMAN (1969): Thermal analysis of lipids, proteins and biological membranes. A review and summary of some recent studies. Chem. Phys. Lipids, 3, 304–56 LEAHY, T. u. B.M. GADELLA (2011): Sperm surface changes and physiological consequences induced by sperm handling and storage. Reproduction, 142, 759–778 LONG, J. A. u. H.D. GUTHRIE (2006): Validation of a rapid, large-scale assay to quantify ATP concentration in spermatozoa. Theriogenology, 65, 1620–1630 LONG, J. u. H. GUTHRIE (2006): Validation of a rapid, large-scale assay to quantify ATP concentration in spermatozoa. Theriogenology, 65, 1620-1630 LÓPEZ RODRÍGUEZ A., T. RIJSSELAERE, P. VYT, A. VAN SOOM, u. D. MAES (2012): Effect of dilution temperature on boar semen quality. Reprod. Dom. Anim., 47, e63–e66 MARIN, S., K. CHIANG, S. BASSILIAN, W.N.P. LEE, L.G. BOROS, J.M. FERNÁNDEZ-NOVELL, J.J. CENTELLES, A. MEDRANO, J.E. RODRIGUEZ-GIL, u. M. CASCANTE (2003): Metabolic strategy of boar spermatozoa revealed by a metabolomic characterization. FEBS Lett., 554, 342–346 MIKI, K., (2007): Energy metabolism and sperm function. in: E.R.S. ROLDAN u. M. GOMENDIO (Hrsg.): Spermatology, Proceedings of the 10th international Symposium on Spermatology, Madrid. Verlag Nottingham University Press, Nottingham, UK S. 309–325

Literaturverzeichnis
119
MISRO, M.M. u. T. RAMYA, (2012): Fuel/ Energy Sources of Spermatozoa, In: S.J. PAREKATTIL u. A. AGARWAL (Hrsg.): Male fertility Verlag Springer, Heidelberg S. 209-223 NIKOLOPOULOU, M., D.A. SOUCEK, u. J.C. Vary (1985): Changes in the lipid content of boar sperm plasma membranes during epididymal maturation. Biochim. Biophys. Acta, 815, 486–498 PARKS, J. u. D. LYNCH (1992): Lipid composition and thermotropic phase behavior of boar, bull, stallion, and rooster sperm Membranes Cryobiology, 266, 255–266 PETRUNKINA, A.M., G. VOLKER, K.F. WEITZE, M. BEYERBACH, E. TÖPFER-PETERSEN, u. D. WABERSKI, (2005): Detection of cooling-induced membrane changes in the response of boar sperm to capacitating conditions. Theriogenology, 63, 2278–2299 PURSEL, V.G., L.A. JOHNSON, u. G.B. RAMPACEK (1972): Acrosome morphology of boar spermatozoa incubated before cold shock. J. Anim. Sci., 34, 278–283 PURSEL, V.G., L.A. JOHNSON, u. L.L. SCHULMAN (1973 a): Effect of dilution, seminal plasma and incubation period on cold shock susceptibility of boar spermatozoa. J. Anim. Sci., 37, 528–531 PURSEL, V.G., L.L. SCHULMAN, u. L.A. JOHNSON (1973 b): Effect of holding time on storage of boar spermatozoa at 5 °C. J. Anim. Sci., 37, 785 – 789 RIESENBECK, A. (2011): Review on international trade with boar semen. Reprod. Domest. Anim., 46, 1-3 RODRIGUEZ-GIL, J.E. (2013): Biological aspects of the mature boar spermatozoa In: S. BONET, I. CASAS, W.V. HOLT, M. YESTE (Hrsg.): Boar Reproduction Verlag Springer, Heidelberg S. 49-64

Literaturverzeichnis
120
SCHMID, S., H. HENNING, A.M. PETRUNKINA, K.F. WEITZE, u. D. WABERSKI (2014): Response to capacitating stimuli indicates extender-related differences in boar sperm function J. Anim. Sci., 91, 5018–5025 SCHMID, S., H. HENNING, H. OLDENHOF, W.F. WOLKERS, A.M. PETRUNKINA, u. D. WABERSKI (2013): The specific response to capacitating stimuli is a sensitive indicator of chilling injury in hypothermically stored boar spermatozoa. Andrology, 1, 376 – 386 SCHULZE, M., H. HENNING, K. RÜDIGER, U. WALLNER, u. D. WABERSKI (2013): Temperature management during semen processing: Impact on boar sperm quality under laboratory and field conditions. Theriogenology, 9, 990-998 STÄHR, B, L. ROTHE u. D. WABERSKI (2009): Empfehlungen zur Gewinnung, Aufbereitung, Lagerung und Transport von Ebersperma - Handbuch für Besamungsstationen. Institut für Fortpflanzung landwirtschaftlicher Nutztiere Schönow e. V. / Tierärztliche Hochschule Hannover, Schönow/Hannover TAMULI M.K., u. P.F. WATSON (1994): Cold resistance of live boar spermatozoa during incubation after ejaculation. Vet. Rec., 135, 160-162 TRAVIS, A.J., J.A. FOSTER, N.A. ROSENBAUM, P.E. VISCONTI, G.L. GERTON, G.S. KOPF, u. S.B. MOSS (1998): Targeting of a germ cell-specific type 1 hexokinase lacking a porin-binding domain to the mitochondria as well as to the head and fibrous sheath of murine spermatozoa. Mol. Biol. Cell, 9, 263–276 TURNER, R.M. (2003): Tales from the tail: what do we really know about sperm motility? J. Androl., 24, 790–803 WABERSKI, D. (2009): Critical steps from semen collection to insemination. In: Proceedings of the Annua Meeting of the EU-AI-Vets, Ghent, Belgium. S. 66–69 WABERSKI, D. K.F. WEITZE, C. LIETMANN, W. L. ZURLAGE, F.P. BORTOLOZZO, T. WILLMEN u. R. PETZOLD (1994): The initial fertilizing-capacity of long-term-stored liquid boar semen following preovulatory and postovulatory insemination. Theriogenology, 41, 1367-1377

Literaturverzeichnis
121
WATERHOUSE, K.E., P.M DE ANGELIS, T. HAUGAN, H. PAULENZ, P.O. HOFMO, u. W. FARSTAD (2004): Effects of in vitro storage time and semen-extender on membrane quality of boar sperm assessed by flow cytometry. Reproduction, 62, 1638–1651 WATERHOUSE, K.E., P.O. HOFMO, A. TVERDAL, u. R.R. MILLER (2006): Within and between breed differences in freezing tolerance and plasma membrane fatty acid composition of boar sperm. Reproduction, 131, 887–894 WATSON, P.F. (1996): Cooling of spermatozoa and fertilizing capacity. In: D. RATH, L. A. JOHNSON, u. K. F. WEITZE (Hrsg.): Boar Semen Preservation III, Proc 3rd in. conf. Boar Semen Preservaton, Mariensee, Deutschland Reprod. Domest. Anim. 31, S. 135–140 WHITE, I.G. (1993): Lipids and calcium uptake of sperm in relation to cold shock and preservation: a review. Reprod. Fertil. Dev., 5, 639–658 WIEBE, W.J. u. BANCROFT, K. (1975): Use of the adenylate energy charge ratio to measure growth state of natural microbial communities. Proc. Natl. Acad. Sci. U. S. A., 72, 2112–2115 YI, Y.J., Z.H. LI, E.S. KIM, E.S. SONG, H.B. KIM, P.Q. CONG, J.M LEE u. C.S. PARK (2008): Comparison of motility, acrosome, viability and ATP of boar sperm with or without cold shock resistance in liquid semen at 17 °C and 4 °C, and frozen-thawed semen. Asian-Aust. J. Anim. Sci., 21, 190-197

Anhang
122
9 Anhang
9.1 Tabellen und Grafiken
Tabelle 1: CASA Motilitätsparameter (MW ± SD) von Experiment 1 in Abhängigkeit der Verdünnungsart, des Verdünnungsgrades und der Lagerungsdauer (n = 6 Eber)
Parameter Verdün-
nungsgrad Verdün-nungsart
Lagerungsdauer
d 0 d1 d 3 d 6
Gesamt-motilität
[%]
20 x 106/ml
isotherm hypotherm
92,8 ± 3,1a
91,1 ± 2,2
a
89,9 ± 5,6a
86,4 ± 5,1b
86,2 ± 6,9a,*
77,1 ± 12,1b,*
68,4 ± 18,4b
10 x 106/ml
isotherm hypotherm
90,5 ± 2,8a
89,3 ± 4,5
a,#
84,9 ± 4,57a,#
81,2 ± 6,8
a,b
76,5 ± 10,3a,b,
* 61,9 ± 16,0
b,*
60,6 ± 15,0b
Progressive Motilität
[%]
20 x 106/ml
isotherm hypotherm
86,5 ± 4,6a
83,9 ± 3,9
a
81,9 ± 10,3a
72,3 ± 5,6b
71,9 ± 13,8a
62,4 ± 18,2b,*
53,5 ± 24,1b
10 x 106/ml
isotherm hypotherm
84,2 ± 7,5a
78,8 ± 9,1
a
75,7 ± 3,2a
62,7 ± 11,2b
58,5 ± 15,5a,b
45,8 ± 19,9
b,*
40,3 ± 17,0b
VAP [µm/s]
20 x 106/ml
isotherm hypotherm
79,6 ± 14,0a
67,6 ± 8,7
b
66,6 ± 11,4a
55,2 ± 9,6b
56,1 ± 16,4a,b
54,5 ± 18,6a,b
50,5 ± 14,4b
10 x 106/ml
isotherm hypotherm
73,3 ± 13,7a
64,8 ± 14,0
a,b
67,3 ± 11,2a
48,1 ± 16,0b,c
44,6 ± 7,4b
39,6 ± 10,8c
42,2 ± 8,7b
VCL [µm/s]
20 x 106/ml
isotherm hypotherm
117,0 ± 21,6
110,9 ±1 7,9
108,3 ± 20,7
87,2 ± 15,3
88,8 ± 31,4
83,4 ± 30,6
78,9 ± 24,4
10 x 106/ml
isotherm hypotherm
109,8 ± 23,6
109,4 ± 23,8
116,3 ± 21,7 75,4 ± 25,1
72,5 ± 8,6 61,2 ± 14,5
69,5 ± 15,6
VSL [µm/s]
20 x 106/ml
isotherm hypotherm
64,6 ± 12,5a
49,9 ± 4,3
b
49,5 ± 7,5
44,1 ± 8,3b
43,8 ± 11,4
44,2 ± 14,5b
40,6 ± 11,4
10 x 106/ml
isotherm hypotherm
59,8 ± 12,4a
47,4 ± 9,6
b
48,6 ± 6,8a
39,1 ± 14,2b
35,4 ± 7,9a,b
32,1 ± 10,3
b
32,6 ± 6,6b
STR
20 x 106/ml
isotherm hypotherm
0,81 ± 0,03
0,74 ± 0,04
0,74 ± 0,02
0,79 ± 0,02
0,79 ± 0,06
0,8 ± 0,05
0,8 ± 0,05
10 x 106/ml
isotherm hypotherm
0,81 ± 0,04a
0,73 ± 0,03
b
0,72 ± 0,03 0,8 ± 0,05
a,b
0,78 ± 0,06 0,8 ± 0,06
a,b
0,77 ± 0,05
LIN
20 x 106/ml
isotherm hypotherm
0,55 ± 0,04a
0,45 ± 0,04
b
0,45 ± 0,02
0,5 ± 0,04a
0,5 ± 0,07
0,53 ± 0,07a,b
0,52 ± 0,08
10 x 106/ml
isotherm hypotherm
0,54 ± 0,05a
0,43 ± 0,03
b
0,42 ± 0,04 0,51 ± 0,07
a,b
0,48 ± 0,07 0,51 ± 0,08
a,b
0,47 ± 0,06
WOB
20 x 106/ml
isotherm hypotherm
0,68 ± 0,03a
0,61 ± 0,03
b
0,61 ± 0,02
0,63 ± 0,04a
0,64 ± 0,05
0,65 ± 0,04a,b
0,64 ± 0,06
10 x 106/ml
isotherm hypotherm
0,67 ± 0,03a
0,59 ± 0,02
b
0,58 ± 0,03 0,64 ± 0,06
a,b
0,61 ± 0,05 0,64 ± 0,05
a,b
0,61 ± 0,05
ALH [µm]
20 x 106/ml
isotherm hypotherm
2,43 ± 0,44a,b,
*
2,53 ± 0,51
a
2,40 ± 0,48
2,15 ± 0,30a,b,
*
2,20 ± 0,86
2,00 ± 0,58b
1,99 ± 0,77
10 x 106/ml
isotherm hypotherm
2,11 ± 0,42a,b,
*
2,30 ± 0,44
a
2,46 ± 0,48a
1,76 ± 0,33b,c,
*
1,85 ± 0,35b
1,65 ± 0,28c
1,95 ± 0,47a,b
BCF [Hz]
20 x 106/ml
isotherm hypotherm
39,9 ± 2,4a
38,0 ± 2,0
a,b
37,7 ± 3,6a
34,0 ± 4,8b
32,3 ± 5,4b
31,1 ± 6,7b
28,9 ± 7,2b
10 x 106/ml
isotherm hypotherm
40,3 ± 3,3a
37,3 ± 5,5
a,b
38,1 ± 3,0a
29,8 ± 8,3b,c
28,6 ± 4,5b
24,0 ± 6,2c
25,3 ± 4,4b
a-c: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
*: gleiche Symbole zeigen signifikante Unterschiede zwischen Verdünnungsgraden bei gleicher Verdünnungsart (p < 0,05)
#: gleiche Symbole zeigen signifikante Unterschiede zwischen Verdünnungsarten bei gleichem Verdünnungsgrad (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency
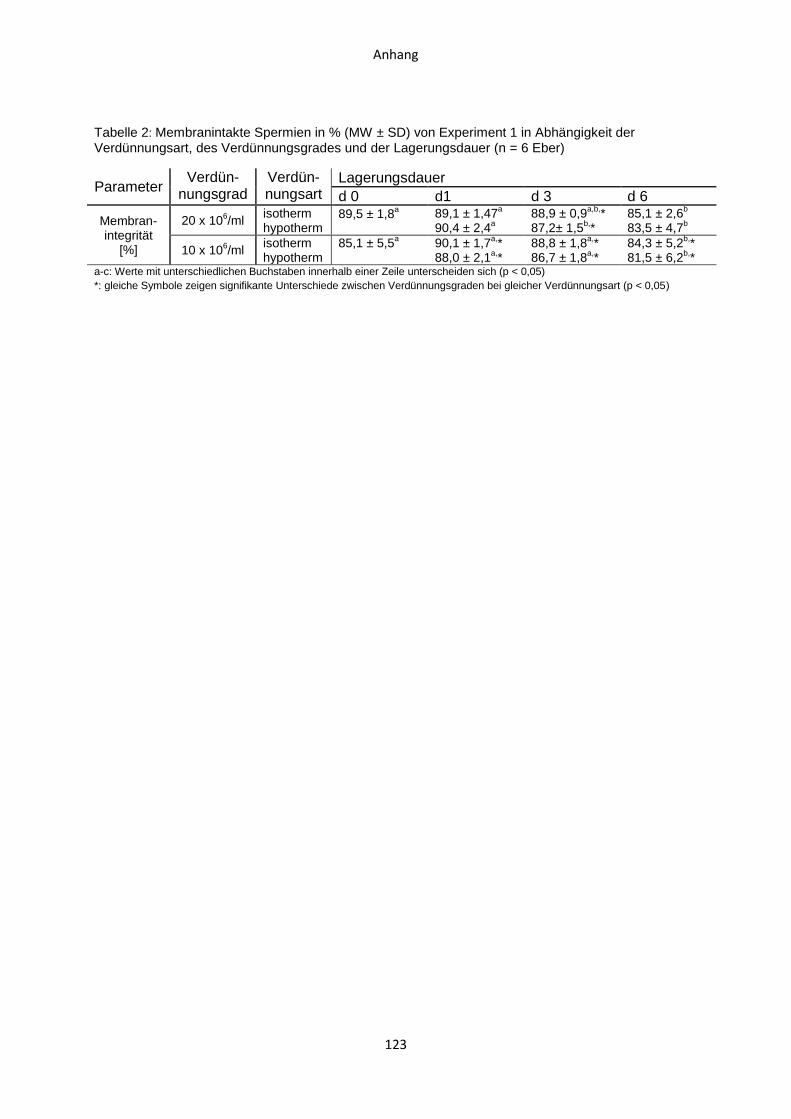
Anhang
123
Tabelle 2: Membranintakte Spermien in % (MW ± SD) von Experiment 1 in Abhängigkeit der Verdünnungsart, des Verdünnungsgrades und der Lagerungsdauer (n = 6 Eber)
Parameter Verdün-
nungsgrad Verdün-nungsart
Lagerungsdauer
d 0 d1 d 3 d 6
Membran-integrität
[%]
20 x 106/ml
isotherm hypotherm
89,5 ± 1,8a 89,1 ± 1,47
a
90,4 ± 2,4a
88,9 ± 0,9a,b,
* 87,2± 1,5
b,*
85,1 ± 2,6b
83,5 ± 4,7b
10 x 106/ml
isotherm hypotherm
85,1 ± 5,5a
90,1 ± 1,7a,*
88,0 ± 2,1a,*
88,8 ± 1,8a,*
86,7 ± 1,8a,*
84,3 ± 5,2b,*
81,5 ± 6,2b,*
a-c: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
*: gleiche Symbole zeigen signifikante Unterschiede zwischen Verdünnungsgraden bei gleicher Verdünnungsart (p < 0,05)

Anhang
124
Tabelle 3: CASA Motilitätsparameter (MW ± SD) von Experiment 2 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber)
Parameter Haltezeit Haltetem-peratur
Lagerungsdauer
d0 d 1 d 3 d 6*
Gesamt-motilität
[%]
K 32 °C 90,1 ± 2,9 90,4 ± 3,6 88,8 ± 5,0 88,3 ± 1,8
30 min 32 °C 91,1 ± 2,3 90,5 ± 3,1
# 89,0 ± 5,4 87,7 ± 4,2
21 °C 91,1 ± 3,5 89,7 ± 4,2#
88,3 ± 6,6 88,7 ± 1,6
60 min 32 °C 90,8 ± 2,8 91,1 ± 3,4 89,1 ± 6,1 88,4 ± 3,6
21 °C 90,8 ± 3,4 89,4 ± 5,5 87,8 ± 6,8 88,8 ± 3,4
3 h 32 °C 91,3 ± 3,8 89,7 ± 3,7 89,1 ± 5,0 88,2 ± 3,6
21 °C 90,5 ± 3,5 90,9 ± 3,8 87,4 ± 9,4 88,6 ± 2,0
6 h 32 °C 91,9 ± 2,7 89,9 ± 7,2
# 90,0 ± 4,3 88,8 ± 2,9
21 °C 90,7 ± 3,6 90,1 ± 3,5#
88,5 ± 9,5 89,1 ± 2,3
Progressive Motilität
[%]
K 32 °C 82,3 ± 5,9 81,2 ± 3,9 77,8 ± 5,8 77,1 ± 4,1
30 min 32 °C 81,9 ± 5,4
# 81,1 ± 6,4 79,7 ± 7,1
# 76,1 ± 6,0
21 °C 82,5 ± 6,6#
80,4 ± 5,3 78,7 ± 7,7#
78,2 ± 5,1
60 min 32 °C 82,6 ± 4,9 80,7 ± 3,9 80,3 ± 7,9 79,2 ± 4,6
21 °C 82,7 ± 6,4 80,3 ± 7,1 78,0 ± 9,6 77,3 ± 6,8
3 h 32 °C 83,2 ± 6,5 79,5 ± 6,8 79,8 ± 7,7 78,8 ± 5,4
21 °C 81,7 ± 5,8 82,4 ± 5,8 78,2 ± 11,6 78,1 ± 5,9
6 h 32 °C 84,0 ± 4,9 81,3 ± 7,9
# 81,5 ± 5,5 78,6 ± 5,1
21 °C 82,1 ± 5,5 79,6 ± 7,7#
79,1 ± 10,9 78,6 ± 2,0
VAP [µm/s]
K 32 °C 73,2 ± 11,7
65,2 ± 9,9
64,8 ± 12,1
63,3 ± 11,1
30 min 32 °C 71,1 ± 10,3
a,# 65,4 ± 9,3
a,b 65,0 ± 12,0
a,b,# 61,9 ± 11,4
b
21 °C 71,9 ± 7,8# 66,5 ± 11,9 62,0 ± 118
# 63,8 ± 15,4
60 min 32 °C 71,7 ± 8,6 64,7 ± 10,1 63,7 ± 11,7 69,1 ± 10,3
21 °C 71,7 ± 9 63,9 ± 12,1 62,9 ± 13,8 63,9 ± 12,9
3 h 32 °C 71,8 ± 7,7 65,6 ± 11,4 64,7 ± 10,3 67,0 ± 14,6
21 °C 68,7 ± 8,7 65,8 ± 12,0 64,6 ± 11,4 69,2 ± 13,7
6 h 32 °C 74,9 ± 8,7 65,6 ± 9,9 67,0 ± 11,6 69,1 ± 11,5
21 °C 70,2 ± 7,9 66,6 ± 10,8 63,3 ± 10,0 68,3 ± 12,4
VCL [µm/s]
K 32 °C 108,2 ± 21,9
108,9 ± 24,8
104,5 ± 24,6
107,1 ± 30,8
30 min 32 °C 108,3 ± 2,8 113,4 ± 26,5 106,3 ± 7,0 105,8 ± 28,5
21 °C 109,0 ± 18,8 115,6 ± 32,6 100,5 ± 26,3 108,2 ± 38,9
60 min 32 °C 107,7 ± 20,5
a 109,9 ± 26,3
a,b 104,3 ± 26,1
a,b 118,4 ± 28,0
b
21 °C 110,0 ± 22,0 110,3 ± 31,0 102,4 ± 28,0 107,9 ± 33,6
3 h 32 °C 107,6 ± 17,1 112,4 ± 29,3 106,3 ± 23,0 113,4 ± 34,7
21 °C 111,0 ± 24,3 114,8 ± 31,0 104,8 ± 26,6 121,4 ± 34,0
6 h 32 °C 114,5 ± 18,1 115,6 ± 30,0 110,4 ± 24,9 121,0 ± 30,6
21 °C 117,1 ± 21,9 118,3 ± 28,9 103,7 ± 23,9 117,4 ± 33,1
VSL [µm/s]
K 32 °C 59,2 ± 9,7
46,3 ± 4,63 49,0 ± 7,7
47,2 ± 3,3
30 min 32 °C 56,7 ± 7,7
a,# 46,7 ± 4,1
a,b 48,9 ± 6,6
a,b,# 45,8 ± 4,6
b
21 °C 57,4 ± 6,0# 46,6 ± 5,1 47,5 ± 6,9
# 46,9 ± 3,7
60 min 32 °C 57,3 ± 5,8
a,# 46,3 ± 4,5
a 48,3 ± 6,7
a,b 49,8 ± 2,8
b
21 °C 56,0 ± 5,9# 45,5 ± 4,6 47,9 ± 8,8 47,0 ± 5,2
3 h 32 °C 58,6 ± 6,3
a 47,0 ± 4,8
a,b 49,4 ± 5,8
a,b,# 48,2 ± 5,5
b
21 °C 51,1 ± 4,4 45,4 ± 5,8 49,1 ± 6,8# 48,9 ± 5,0
6 h 32 °C 60,9 ± 7,6
a 47,6 ± 4,6
a,b,# 51,2 ± 7,5
a,b 49,8 ± 3,4
b
21 °C 50,4 ± 5,9 46,4 ± 6,6# 48,0 ± 6,1 49,4 ± 3,3

Anhang
125
STR
K 32 °C 0,80 ± 0,03
0,72 ± 0,09
0,76 ± 0,07
0,75 ± 0,08
30 min
32 °C 0,79 ± 0,04 0,72 ± 0,08 0,76 ± 0,07 0,75 ± 0,07
21 °C 0,79 ± 0,04 0,71 ± 0,09 0,77 ± 0,06 0,75 ± 0,11
60 min
32 °C 0,80 ± 0,04 0,72 ± 0,07 0,76 ± 0,06 0,72 ± 0,08
21 °C 0,78 ± 0,05 0,72 ± 0,07 0,76 ± 0,06 0,74 ± 0,08
3 h
32 °C 0,81 ± 0,04 0,72 ± 0,07 0,76 ± 0,05 0,73 ± 0,08
21 °C 0,75 ± 0,07 0,69 ± 0,08 0,76 ± 0,06 0,71 ± 0,10
6 h 32 °C 0,81 ± 0,04 0,73 ± 0,07 0,76 ± 0,05 0,73 ± 0,08
21 °C 0,72 ± 0,08 0,70 ± 0,09 0,76 ± 0,07 0,73 ± 0,10
LIN
K 32 °C 0,55 ± 0,06 0,43 ± 0,09 0,48 ± 0,08 0,46 ± 0,10
30 min
32 °C 0,53 ± 0,07 0,43 ± 0,08 0,47 ± 0,08 0,45 ± 0,08
21 °C 0,53 ± 0,07 0,42 ± 0,09 0,48 ± 0,08 0,46 ± 0,11
60 min
32 °C 0,54 ± 0,07 0,43 ± 0,08 0,47 ± 0,07 0,43 ± 0,08
21 °C 0,52 ± 0,07 0,43 ± 0,08 0,48 ± 0,07 0,46 ± 0,09
3 h
32 °C 0,55 ± 0,06 0,43 ± 0,08 0,47 ± 0,07 0,44 ± 0,09
21 °C 0,47 ± 0,08 0,41 ± 0,09 0,48 ± 0,08 0,42 ± 0,09
6 h 32 °C 0,53 ± 0,07 0,43 ± 0,08 0,47 ± 0,06 0,42 ± 0,08
21 °C 0,44 ± 0,08 0,41 ± 0,09 0,47 ± 0,08 0,44 ± 0,10
WOB
K 32 °C 0,68 ± 0,05 0,60 ± 0,07 0,62 ± 0,05 0,60 ± 0,07
30 min
32 °C 0,66 ± 0,06 0,59 ± 0,06 0,62 ± 0,05 0,59 ± 0,06
21 °C 0,66 ± 0,06 0,59 ± 0,07 0,62 ± 0,06 0,60 ± 0,07
60 min
32 °C 0,67 ± 0,06 0,59 ± 0,05 0,62 ± 0,05 0,59 ± 0,06
21 °C 0,66 ± 0,05 0,59 ± 0,06 0,62 ± 0,06 0,60 ± 0,06
3 h
32 °C 0,67 ± 0,05 0,59 ± 0,05 0,61 ± 0,05 0,60 ± 0,06
21 °C 0,63 ± 0,07 0,58 ± 0,06 0,62 ± 0,06 0,58 ± 0,05
6 h 32 °C 0,66 ± 0,06 0,59 ± 0,06 0,61 ± 0,05 0,58 ± 0,05
21 °C 0,60 ± 0,06 0,58 ± 0,06 0,62 ± 0,06 0,59 ± 0,06
ALH [µm]
K 32 °C 2,42 ± 0,61
2,54 ± 0,63
2,33 ± 0,55
2,47 ± 0,69
30 min 32 °C 2,29 ± 0,50 2,51 ± 0,64 2,33 ± 0,60 2,43 ± 0,63
21 °C 2,27 ± 0,44 2,54 ± 0,67 2,26 ± 0,57 2,50 ± 0,85
60 min 32 °C 2,29 ± 0,47 2,49 ± 0,62 2,31 ± 0,58 2,59 ± 0,64
21 °C 2,30 ± 0,49 2,46 ± 0,70 2,26 ± 0,57 2,48 ± 0,79
3 h 32 °C 2,29 ± 0,41 2,50 ± 0,61 2,34 ± 0,52 2,64 ± 0,74
21 °C 2,40 ± 0,56 2,61 ± 0,66 2,29 ± 0,59 2,77 ± 0,79
6 h 32 °C 2,41 ± 0,40 2,48 ± 0,62 2,37 ± 0,55 2,71 ± 0,74
21 °C 2,50 ± 0,49a 2,64 ± 0,62
a,b 2,32 ± 0,60
a,b 2,77 ± 0,78
b
BCF [Hz]
K 32 °C 39,2 ± 2,5 36,9 ± 2,5 36,8 ± 4,0 35,6 ± 2,4
30 min 32 °C 39,4 ± 2,8 37,7 ± 1,8 36,9 ± 3,6
# 35,1 ± 3,8
21 °C 40,1 ± 2,4 37,5 ± 2,1 35,6 ± 4,0#
35,4 ± 2,4
60 min 32 °C 39,5 ± 2,5 36,8 ± 1,5 36,4 ± 3,8 37,1 ± 2,4
21 °C 39,4 ± 2,5 36,2 ± 3,1 35,7 ± 4,5 35,8 ± 3,4
3 h 32 °C 40,2 ± 2,1 37,3 ± 2,6 37,2 ± 3,4 36,4 ± 2,8
21 °C 39,1 ± 2,1 37,1 ± 3,0 36,9 ± 3,7 36,7 ± 3,9
6 h 32 °C 40,1 ± 2,0 37,0 ± 2,0 37,6 ± 3,7 36,6 ± 2,4
21 °C 39,6 ± 2,3 38,0 ± 2,9 36,2 ± 2,0 36,3 ± 3,1
K= Kontrolle; *: d 6: n = 6 Eber;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
#: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltetemperaturen bei gleicher Lagerungsdauer (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency

Anhang
126
Tabelle 4: Ergebnisse der durchflusszytometrischen Messungen in % (MW ± SD) von Experiment 2 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber)
Parameter Haltezeit Haltetem-peratur
Lagerungsdauer
d0 d 1 d 3 d 61
Membran-integrität
[%]
K 32 °C 89,6 ± 2,9 88,5 ± 3,3 90,0 ± 2,6 85,9 ± 3,6
30 min 32 °C 90,0 ± 2,4 89,3 ± 3,9* 90,0 ± 2,6 85,5 ± 5,0
21 °C 90,2 ± 2,2 89,4 ± 2,9** 89,8 ± 2,3 84,8 ± 5,2
60 min 32 °C 89,8 ± 2,9 89,0 ± 3,1* 89,9 ± 2,4*
,** 84,9 ± 4,5
21 °C 90,3 ± 2,6 89,5 ± 3,1*** 90,1 ± 2,4 86,4 ± 4,4
3 h 32 °C 90,1 ± 2,7 89,0 ± 3,6 89,2 ± 2,7* 85,5 ± 4,2
21 °C 90,8 ± 3,0 89,1 ± 3,1**;*** 89,9 ± 2,4 85,4 ± 4,3
6 h 32 °C 89,5 ± 2,4 88,3 ± 4,6 88,9 ± 2,6** 85,0 ± 4,0
21 °C 90,5 ± 3,2 89,6 ± 2,8 90,3 ± 2,1 86,9 ± 5,0
Mito-chondrien-membran-potential
[%]
K 32 °C 79,8 ± 6,9 86,4 ± 12,4 87,4 ± 9,8 78,3 ± 14,1
30 min 32 °C 77,4 ± 10,9 88,1 ± 10,6 87,2 ± 7,6 76,6 ± 18,2 21 °C 83,6 ± 7,0 83,5 ± 20,2 82,3 ± 17,7 80,6 ± 12,9
60 min 32 °C 76,2 ± 15,9 85,9 ± 13,9 83,4 ± 9,3 77,3 ± 20,8
21 °C 82,1 ± 8,8 82,6 ± 20,0 80,8 ± 11,9 77,0 ± 18,4
3 h 32 °C 87,3 ± 5,2 82,8 ± 15,8 82,6 ± 11,1 77,6 ± 17,5 21 °C 83,1 ± 6,9 82,1 ± 19,8 76,7 ± 17,8 73,4 ± 20,1
6 h 32 °C 86,9 ± 4,4 82,7 ± 19,8 83,3 ± 8,4 80,2 ± 12,3 21 °C 80,8 ± 5,6 80,5 ± 21,1 78,6 ± 14,5 74,4 ± 16,4
Mittlere Fluo-
reszenz-intensität
aller Spermien
[AU]
K 32 °C 14,1 ± 5,1 14 ± 4,8 13,5 ± 7,2 10,9 ± 5,1
30 min 32 °C 16,2 ± 3,3 16,5 ± 7,1 13,7 ± 5,3 12,3 ± 6 21 °C 19,9 ± 8,1 16,4 ± 6,6 12,6 ± 6,0 11,2 ± 4,6
60 min 32 °C 13,8 ± 7,8 15,9 ± 8,1 11,6 ± 4,4 13,8 ± 6,5 21 °C 13,6 ± 4,0 15,8 ± 7,2 11,9 ± 5,6 11,9 ± 5,1
3 h 32 °C 15,5 ± 5,8 14,9 ± 8,9 11,6 ± 4,7 11,2 ± 5,8 21 °C 13,7 ± 6,0 15,2 ± 7,9 11,4 ± 4,2 10,5 ± 5,3
6 h 32 °C 14,0 ± 4,1 15,9 ± 8,4 11,8 ± 5,2 11,6 ± 5 21 °C 12,9 ± 3,9 15,0 ± 8,8 11,3 ± 5,0 11,8 ± 5,2
K= Kontrolle; 1: d 6: n = 6 Eber;
*-***: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltezeiten bei gleicher Lagerungsdauer (p < 0,05)

Anhang
127
Tabelle 5: Ergebnisse der Analyse des Energiestoffwechsels (MW + SD) von Experiment 2 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber)
Parameter Haltezeit Haltetem-peratur
Lagerungsdauer
d0 d 1 d 3 d 61
ATP [pmol/ 50000
Spermien]
nativ 298 ± 78 K 32 °C 336 ± 115 257 ± 127 260 ± 59 272 ± 77
30 min 32 °C 296 ± 101 237 ± 103 239 ± 72 226 ± 73
21 °C 298 ± 142a
252 ± 93b
250 ± 51a,b
240 ± 68b
60 min 32 °C 270 ± 90 223 ± 105 272 ± 60 202 ± 68
21 °C 306 ± 94 231 ± 60 239 ± 75 254 ± 66
3 h 32 °C 313 ± 85 232 ± 93 232 ± 62 251 ± 64
21 °C 278 ± 73 261 ± 88 232 ± 71 249 ± 71
6 h 32 °C 282 ± 103 250 ± 99 238 ± 61 259 ± 62
21 °C 277 ± 70 245 ± 50 236 ± 55 237 ± 62
ADP
[pmol]
nativ 104 ± 21 K 32 °C 98 ± 32 88 ± 38 125 ± 46 81 ± 40
30 min 32 °C 80 ± 51 85 ± 20 65 ± 26 66 ± 20
21 °C 90 ± 30 80 ± 19 74 ± 39 80 ± 31
60 min 32 °C 82 ± 20 78 ± 21 69 ± 27 52 ± 32
21 °C 88 ± 27 67 ± 14 73 ± 40 91 ± 42
3 h 32 °C 89 ± 38 79 ± 22 61 ± 26 70 ± 25
21 °C 86 ± 27 80 ± 31 72 ± 44 78 ± 29
6 h 32 °C 80 ± 29 70 ± 30 58 ± 38 59 ± 23
21 °C 84 ± 36 84 ± 25 69 ± 31 78 ± 36
AMP [pmol]
nativ 81 ± 56 K 32 °C 86 ± 58 103 ± 54 51 ± 58 86 ± 37
30 min 32 °C 75 ± 49 92 ± 35 104 ± 56 73 ± 44
21 °C 74 ± 43 105 ± 52 99 ± 61 91 ± 26
60 min 32 °C 93 ± 53 76 ± 49 117 ± 32 70 ± 39
21 °C 88 ± 74 90 ± 49 94 ± 52 75 ± 28
3 h 32 °C 99 ± 70 90 ± 65 88 ± 39 74 ± 27
21 °C 72 ± 71 83 ± 36 65 ± 35 80 ± 39
6 h 32 °C 74 ± 52 75 ± 53 89 ± 46 84 ± 36
21 °C 59 ± 53 78 ± 73 51 ± 39 81 ± 24
Energie-ladung
nativ 0,68 ± 0,12 K 32 °C 0,69 ± 0,11 0,61 ± 0,14 0,77 ± 0,18 0,66 ± 0,09
30 min 32 °C 0,70 ± 0,10 0,62 ± 0,12 0,62 ± 0,14 0,66 ± 0,08
21 °C 0,68 ± 0,10 0,62 ± 0,11 0,65 ± 0,12 0,67 ± 0,11
60 min 32 °C 0,67 ± 0,12 0,65 ± 0,13 0,62 ± 0,08 0,69 ± 0,10
21 °C 0,67 ± 0,13 0,64 ± 0,12 0,65 ± 0,12 0,66 ± 0,10
3 h 32 °C 0,70 ± 0,14 0,66 ± 0,21 0,64 ± 0,09 0,67 ± 0,09
21 °C 0,72 ± 0,13 0,66 ± 0,10 0,69 ± 0,11 0,67 ± 0,07
6 h 32 °C 0,70 ± 0,11 0,70 ± 0,18 0,66 ± 0,08 0,66 ± 0,10
21 °C 0,72 ± 0,12 0,68 ± 0,16 0,75 ± 0,16 0,67 ± 0,08 K= Kontrolle;
1: d 6: n = 6 Eber;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)

Anhang
128
Tabelle 6: CASA Motilitätsparameter (MW ± SD) von Experiment 3 in Abhängigkeit der Haltezeit, der Verdünnertemperatur und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Verdünner-temperatur
Lagerungsdauer
d0 d 1 d 3
Gesamt-motilität
[%]
K 32 °C 93,2 ± 2,0
a 91,8 ± 2,9
a 91,4 ± 4,0
a
30 min 32 °C 92,0 ± 3,1* 89,7 ± 4,2
89,0 ± 4,8
21 °C 92,0 ± 2,7a,b
92,8 ± 3,0a
89,5 ± 5,2b
60 min 32 °C 91,4 ± 2,6 90,6 ± 4,6 90,3 ± 5,8
21 °C 91,4 ± 3,1
91,4 ± 3,5
88,3 ± 6,4
3 h 32 °C 93,3 ± 2,3* 91,5 ± 2,9
90,6 ± 4,7
21 °C 92,7 ± 2,3
89,9 ± 6,7
86,7 ± 10,8
6 h 32 °C 93,2 ± 2,7
90,0 ± 5,2
89,8 ± 5,1
21 °C 93,0 ± 2,2a 86,2 ± 14,6
a,b 84,3 ± 15,4
b
Progressive Motilität
[%]
K 32 °C 87,1 ± 3,79 83,77 ± 3,82 83,57 ± 6,41
30 min 32 °C 85,6 ± 4,0
a 81,2 ± 7,2
a,b 80,3 ± 6,5
b
21 °C 85,2 ± 4,2a 85,2 ± 6,0
a,b 78,6 ± 10,8
b
60 min 32 °C 84,9 ± 3,7 83,6 ± 6,4 80,5 ± 7,5
21 °C 84,7 ± 4,7 83,4 ± 5,6 77,4 ± 8,4
3 h 32 °C 86,7 ± 4,2 84,9 ± 5,0 81,2 ± 6,1
21 °C 85,5 ± 3,4a 83,9 ± 8,2
a 76,6 ± 13,3
b
6 h 32 °C 86,4 ± 4,4 82,8 ± 7,7 81,8 ± 7,6
21 °C 85,9 ± 4,7a 78,4 ± 14,8
a 72,4 ± 18,0
b
VAP [µm/s]
K 32 °C 74,5 ± 8,1
a 65,4 ± 13,2
a,b 59,5 ± 7,6
b
30 min 32 °C 74,2 ± 6,9
a 66,3 ± 11,6
b 62,6 ± 7,1
b
21 °C 72,1 ± 4,36a 64,5 ± 14,7
a 56,9 ± 3,3
b
60 min 32 °C 72,1 ± 8,1
a 65,55 ± 12,56
a,b 59,51 ± 7,04
b
21 °C 69,8 ± 6,1a 61,95 ± 13,93
a,b 56,98 ± 9,1
b
3 h 32 °C 66,9 ± 7,0 67,7 ± 13,2 63,97 ± 11,55
21 °C 68,7 ± 4,4 66,56 ± 11,91 61,5 ± 14,41
6 h 32 °C 67,7 ± 8,9 66,52 ± 14,2 63,97 ± 11,55
#
21 °C 65,9 ± 6,0 63,85 ± 11,23 60,35 ± 9,97#
VCL [µm/s]
K 32 °C 107,9 ± 14,7
a 106,0 ± 23,6
a,b 91,3 ± 15,7
b
30 min 32 °C 107,1 ± 12,9
a 107,1 ± 23,0
a,b 98,2 ± 15,5
b
21 °C 103,7 ± 9,3a,b
103,5 ± 25,1a 89,5 ± 26,4
b
60 min 32 °C 104,5 ± 14,8
a,b 104,4 ± 23,4
a 92,4 ± 17,6
b
21 °C 100,6 ± 10,0 100,5 ± 27,4 89,3 ± 21,3
3 h 32 °C 103,2 ± 16,6 108,2 ± 24,8 101,0 ± 21,8
21 °C 106,9 ± 12,9 108,2 ± 23,4 97,3 ± 28,4
6 h 32 °C 104,9 ± 20,5 107,3 ± 26,8 105,5 ± 26,8
21 °C 105,6 ± 17,0a,b
106,4 ± 23,8a 99,0 ± 23,7
b
VSL [µm/s]
K 32 °C 61,3 ± 6,6
a 49,0 ± 7,6
b 48,16 ± 5,53
b
30 min 32 °C 61,1 ± 5,4
a 49,0 ± 6,6
b 49,6 ± 4,9
b
21 °C 59,8 ± 2,9a 49,6 ± 9,8
b 45,1 ± 8,8
c
60 min 32 °C 59,4 ± 5,2
a 50,0 ± 7,7
a,b 47,4 ± 4,3
b
21 °C 58,0 ± 4,9a 47,0 ± 8,2
b 45,4 ± 5,5
b
3 h 32 °C 52,4 ± 3,4 50,8 ± 8,4 49,8 ± 7,7
21 °C 53,6 ± 3,0 49,5 ± 7,3 47,9 ± 8,8
6 h 32 °C 51,1 ± 3,7 50,0 ± 8,0 50,1 ± 4,8
#
21 °C 48,9 ± 3,4 47,1 ± 5,8 46,2 ± 4,9#

Anhang
129
STR
K 32 °C 0,82 ± 0,03a 0,75 ± 0,04
b 0,81 ± 0,03
a
30 min
32 °C 0,82 ± 0,03a,
* 0,74 ± 0,03b 0,79 ± 0,04
a
21 °C 0,82 ± 0,02a,
*** 0,77 ± 0,03b 0,79 ± 0,04
a,b
60 min
32 °C 0,82 ± 0,03a,
** 0,76 ± 0,04b 0,8 ± 0,03
a,b
21 °C 0,83 ± 0,03a,
**** 0,76 ± 0,05b 0,8 ± 0,05
a,b,*
3 h
32 °C 0,78 ± 0,05*,** 0,75 ± 0,04 0,78 ± 0,04
21 °C 0,78 ± 0,05***,**** 0,74 ± 0,04 0,78 ± 0,05
6 h 32 °C 0,76 ± 0,08 0,76 ± 0,04 0,76 ± 0,07
21 °C 0,74 ± 0,08**** 0,74 ± 0,06 0,77 ± 0,06*
LIN
K 32 °C 0,57 ± 0,04a 0,47 ± 0,05
b 0,53 ± 0,06
a
30 min
32 °C 0,57 ± 0,05a,
*,** 0,46 ± 0,05
b 0,51 ± 0,06
c
21 °C 0,57 ± 0,03a,
**** 0,48 ± 0,04b 0,51 ± 0,07
a,b
60 min
32 °C 0,57 ± 0,04a,
*** 0,48 ± 0,05b,
* 0,52 ± 0,07a,b
21 °C 0,57 ± 0,04a,
***** 0,48 ± 0,06b 0,52 ± 0,09
a,b
3 h
32 °C 0,51 ± 0,06*,*** 0,47 ± 0,05 0,50 ± 0,07
21 °C 0,50 ± 0,06**** 0,46 ± 0,06 0,50 ± 0,07
6 h 32 °C 0,50 ± 0,08**
,*** 0,47 ± 0,05 0,49 ± 0,09
21 °C 0,47 ± 0,09****,***** 0,45 ± 0,07* 0,48 ± 0,08
WOB
K 32 °C 0,69 ± 0,03
a,* 0,62 ± 0,03
b 0,65 ± 0,04
a,b
30 min
32 °C 0,69 ± 0,04a,
** 0,62 ± 0,03b 0,64 ± 0,04
b
21 °C 0,69 ± 0,02a,
**** 0,62 ± 0,03b 0,64 ± 0,06
a,b
60 min
32 °C 0,69 ± 0,03a,
*** 0,63 ± 0,03b 0,65 ± 0,05
a,b
21 °C 0,69 ± 0,03a,
***** 0,62 ± 0,04b 0,65 ± 0,07
a,b,*
3 h
32 °C 0,65 ± 0,04*** 0,63 ± 0,04 0,63 ± 0,05
21 °C 0,65 ± 0,04****,***** 0,62 ± 0,05 0,64 ± 0,05
6 h 32 °C 0,65 ± 0,04
a,*
,**
,*** 0,62 ± 0,04
b 0,64 ± 0,06
a,b
21 °C 0,62 ± 0,06***** 0,60 ± 0,05 0,62 ± 0,06*
ALH [µm]
K 32 °C 2,29 ± 0,41
a,b 2,37 ± 0,63
a 2,07 ± 0,48
b,*
30 min 32 °C 2,25 ± 0,31 2,36 ± 0,64 2,24 ± 0,50*
21 °C 2,21 ± 0,31a,b,
** 2,32 ± 0,60a 2,12 ± 0,63
b
60 min 32 °C 2,17 ± 0,39* 2,32 ± 0,56 2,15 ± 0,53**
21 °C 2,09 ± 0,33a,
**,*** 2,37 ± 0,68
a 2,10 ± 0,62
a
3 h 32 °C 2,31 ± 0,42* 2,41 ± 0,63 2,29 ± 0,61**
21 °C 2,29 ± 0,34*** 2,43 ± 0,59* 2,22 ± 0,68
6 h 32 °C 2,31 ± 0,50* 2,44 ± 0,70 2,43 ± 0,75
21 °C 2,37 ± 0,43*** 2,42 ± 0,63* 2,39 ± 0,68
BCF [Hz]
K 32 °C 40,0 ± 0,9
a 38,1 ± 2,1
a,b 36,0 ± 2,7
b
30 min 32 °C 40,0 ± 1,3
a 37,3 ± 3,1
a,b 37,0 ± 2,5
b
21 °C 39,7 ± 1,0a 37,6 ± 4,4
a 34,0 ± 4,8
b
60 min 32 °C 40,2 ± 0,8
a 37,7 ± 2,9
b 35,2 ± 2,5
b
21 °C 39,9 ± 0,8a 35,7 ± 4,3
b 33,7 ± 4,5
c
3 h 32 °C 39,0 ± 2,1 37,8 ± 3,0 36,7 ± 3,2
21 °C 39,8 ± 1,4a 38,0 ± 3,0
a,b 35,4 ± 5,0
b
6 h 32 °C 38,9 ± 2,5
a 37,2 ± 2,9
b 36,2 ± 2,3
b
21 °C 38,9 ± 1,0a 36,6 ± 1,8
b 34,4 ± 2,6
c
K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
#: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltetemperaturen bei gleicher Lagerungsdauer (p < 0,05)
* -
*****: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltezeiten bei gleicher Lagerungsdauer (p < 0,05) VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency

Anhang
130
Tabelle 7: Membranintakte Spermien in % (MW ± SD) von Experiment 3 in Abhängigkeit der Haltezeit, der Verdünnertemperatur und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Verdünner-temperatur
Lagerungsdauer
d0 d 1 d 3
Membran-intakte
Spermien [%]
K 32 °C 91,4 ± 2,3
91,1 ± 1,2
90,9 ± 1,7
30 min 32 °C 91,4 ± 2,0
91,2 ± 2,5
90,6 ± 1,9
21 °C 92,7 ± 2,3a
92,4 ± 2,0a,b
90,6 ± 1,6b
60 min 32 °C 91,8 ± 2,0 91,2 ± 2,3
# 90,8 ± 2,2
21 °C 92,9 ± 1,9a
92,0 ± 2,1b,#
90,8 ± 2,1b
3 h 32 °C 91,6 ± 2,3 90,9 ± 1,9
90,1 ± 1,5
21 °C 92,4 ± 1,6a
91,3 ± 1,9a,b
90,7 ± 0,7b
6 h 32 °C 91,4 ± 2,4
91,1 ± 2,1
91,0 ± 2,1
21 °C 90,8 ± 3,8 90,8 ± 1,8 89,8 ± 1,9 K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05);
#: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltetemperaturen bei gleicher Lagerungsdauer (p < 0,05)
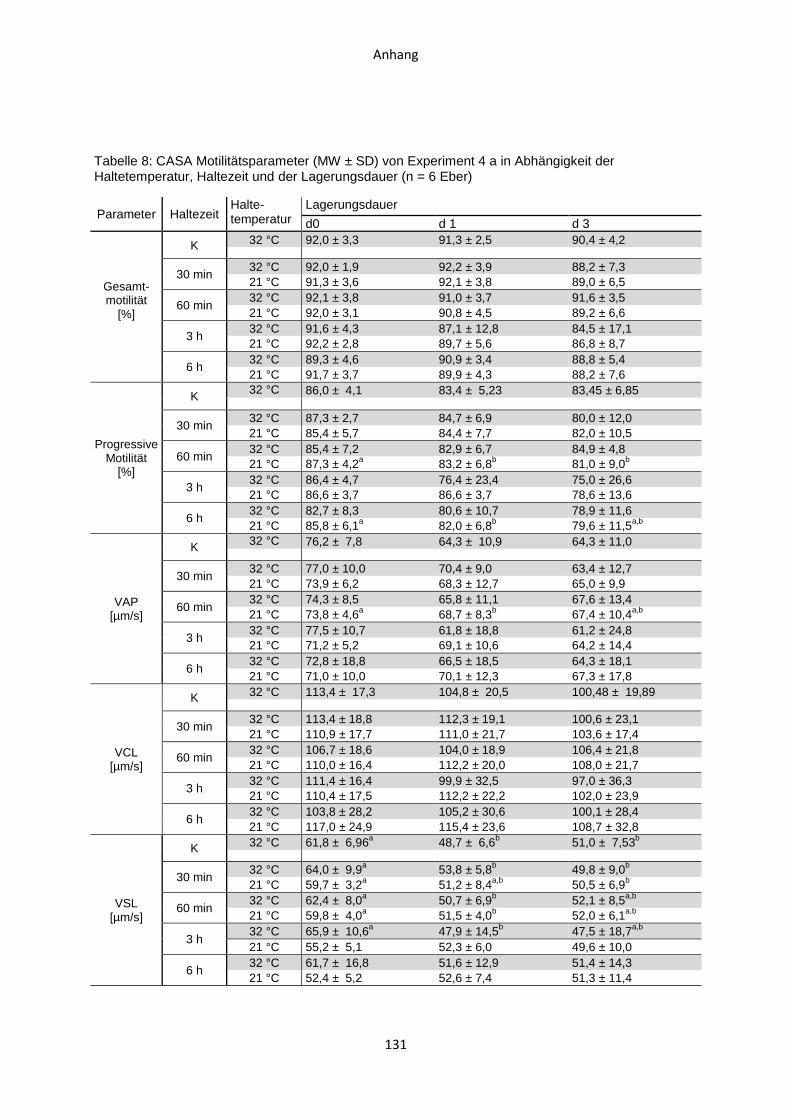
Anhang
131
Tabelle 8: CASA Motilitätsparameter (MW ± SD) von Experiment 4 a in Abhängigkeit der Haltetemperatur, Haltezeit und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Halte-temperatur
Lagerungsdauer
d0 d 1 d 3
Gesamt-motilität
[%]
K 32 °C 92,0 ± 3,3 91,3 ± 2,5 90,4 ± 4,2
30 min 32 °C 92,0 ± 1,9 92,2 ± 3,9 88,2 ± 7,3
21 °C 91,3 ± 3,6 92,1 ± 3,8 89,0 ± 6,5
60 min 32 °C 92,1 ± 3,8 91,0 ± 3,7 91,6 ± 3,5
21 °C 92,0 ± 3,1 90,8 ± 4,5 89,2 ± 6,6
3 h 32 °C 91,6 ± 4,3 87,1 ± 12,8 84,5 ± 17,1
21 °C 92,2 ± 2,8 89,7 ± 5,6 86,8 ± 8,7
6 h 32 °C 89,3 ± 4,6 90,9 ± 3,4 88,8 ± 5,4
21 °C 91,7 ± 3,7 89,9 ± 4,3 88,2 ± 7,6
Progressive Motilität
[%]
K 32 °C 86,0 ± 4,1 83,4 ± 5,23 83,45 ± 6,85
30 min 32 °C 87,3 ± 2,7 84,7 ± 6,9 80,0 ± 12,0
21 °C 85,4 ± 5,7 84,4 ± 7,7 82,0 ± 10,5
60 min 32 °C 85,4 ± 7,2 82,9 ± 6,7 84,9 ± 4,8
21 °C 87,3 ± 4,2a 83,2 ± 6,8
b 81,0 ± 9,0
b
3 h 32 °C 86,4 ± 4,7 76,4 ± 23,4 75,0 ± 26,6
21 °C 86,6 ± 3,7 86,6 ± 3,7 78,6 ± 13,6
6 h 32 °C 82,7 ± 8,3 80,6 ± 10,7 78,9 ± 11,6
21 °C 85,8 ± 6,1a 82,0 ± 6,8
b 79,6 ± 11,5
a,b
VAP [µm/s]
K 32 °C 76,2 ± 7,8 64,3 ± 10,9 64,3 ± 11,0
30 min 32 °C 77,0 ± 10,0 70,4 ± 9,0 63,4 ± 12,7
21 °C 73,9 ± 6,2 68,3 ± 12,7 65,0 ± 9,9
60 min 32 °C 74,3 ± 8,5 65,8 ± 11,1 67,6 ± 13,4
21 °C 73,8 ± 4,6a 68,7 ± 8,3
b 67,4 ± 10,4
a,b
3 h 32 °C 77,5 ± 10,7 61,8 ± 18,8 61,2 ± 24,8
21 °C 71,2 ± 5,2 69,1 ± 10,6 64,2 ± 14,4
6 h 32 °C 72,8 ± 18,8 66,5 ± 18,5 64,3 ± 18,1
21 °C 71,0 ± 10,0 70,1 ± 12,3 67,3 ± 17,8
VCL [µm/s]
K 32 °C 113,4 ± 17,3 104,8 ± 20,5 100,48 ± 19,89
30 min 32 °C 113,4 ± 18,8 112,3 ± 19,1 100,6 ± 23,1
21 °C 110,9 ± 17,7 111,0 ± 21,7 103,6 ± 17,4
60 min 32 °C 106,7 ± 18,6 104,0 ± 18,9 106,4 ± 21,8
21 °C 110,0 ± 16,4 112,2 ± 20,0 108,0 ± 21,7
3 h 32 °C 111,4 ± 16,4 99,9 ± 32,5 97,0 ± 36,3
21 °C 110,4 ± 17,5 112,2 ± 22,2 102,0 ± 23,9
6 h 32 °C 103,8 ± 28,2 105,2 ± 30,6 100,1 ± 28,4
21 °C 117,0 ± 24,9 115,4 ± 23,6 108,7 ± 32,8
VSL [µm/s]
K 32 °C 61,8 ± 6,96
a 48,7 ± 6,6
b 51,0 ± 7,53
b
30 min 32 °C 64,0 ± 9,9
a 53,8 ± 5,8
b 49,8 ± 9,0
b
21 °C 59,7 ± 3,2a 51,2 ± 8,4
a,b 50,5 ± 6,9
b
60 min 32 °C 62,4 ± 8,0
a 50,7 ± 6,9
b 52,1 ± 8,5
a,b
21 °C 59,8 ± 4,0a 51,5 ± 4,0
b 52,0 ± 6,1
a,b
3 h 32 °C 65,9 ± 10,6
a 47,9 ± 14,5
b 47,5 ± 18,7
a,b
21 °C 55,2 ± 5,1 52,3 ± 6,0 49,6 ± 10,0
6 h 32 °C 61,7 ± 16,8 51,6 ± 12,9 51,4 ± 14,3
21 °C 52,4 ± 5,2 52,6 ± 7,4 51,3 ± 11,4

Anhang
132
STR
K 32 °C 0,81 ± 0,05a 0,76 ± 0,05
b 0,79 ± 0,04
a
30 min
32 °C 0,83 ± 0,04a 0,76 ± 0,05
b 0,78 ± 0,04
a,b
21 °C 0,80 ± 0,05a 0,75 ± 0,05
b 0,77 ± 0,04
a,b
60 min
32 °C 0,84 ± 0,05a,#
0,77 ± 0,04b 0,77 ± 0,03
a,b
21 °C 0,80 ± 0,06a,#
0,75 ± 0,05b 0,77 ± 0,05
a,b
3 h
32 °C 0,84 ± 0,04a,##
0,77 ± 0,03b 0,78 ± 0,03
b
21 °C 0,77 ± 0,06##
0,76 ± 0,05 0,77 ± 0,03
6 h 32 °C 0,84 ± 0,03
a,### 0,78 ± 0,05
b,# 0,80 ± 0,02
a,b
21 °C 0,74 ± 0,06###
0,75 ± 0,04# 0,77 ± 0,04
LIN
K 32 °C 0,55 ± 0,08a 0,47 ± 0,06
b 0,51 ± 0,05
a
30 min
32 °C 0,57 ± 0,07a 0,48 ± 0,07
b 0,50 ± 0,06
b
21 °C 0,54 ± 0,08a 0,46 ± 0,06
b 0,49 ± 0,05
b
60 min
32 °C 0,59 ± 0,09a,#
0,49 ± 0,06b 0,49 ± 0,04
a,b
21 °C 0,54 ± 0,09a,#
0,46 ± 0,06b 0,49 ± 0,07
b
3 h
32 °C 0,59 ± 0,09a,##
0,48 ± 0,06b 0,48 ± 0,05
a,b
21 °C 0,51 ± 0,09##
0,47 ± 0,07 0,49 ± 0,05
6 h 32 °C 0,60 ± 0,07
a,### 0,50 ± 0,07
b,# 0,51 ± 0,05
a,b
21 °C 0,46 ± 0,08###
0,46 ± 0,06# 0,48 ± 0,00
WOB
K 32 °C 0,67 ± 0,06
a 0,61 ± 0,04
b 0,64 ± 0,04
a
30 min
32 °C 0,68 ± 0,06a 0,63 ± 0,05
b 0,63 ± 0,04
a,b
21 °C 0,67 ± 0,06a 0,61 ± 0,03
b 0,63 ± 0,03
b
60 min
32 °C 0,70 ± 0,07a 0,63 ± 0,05
b 0,63 ± 0,04
b
21 °C 0,67 ± 0,07a 0,62 ± 0,04
b 0,63 ± 0,05
a,b
3 h
32 °C 0,70 ± 0,08a,#
0,62 ± 0,05b 0,62 ± 0,05
a,b
21 °C 0,65 ± 0,06# 0,62 ± 0,05 0,63 ± 0,04
6 h 32 °C 0,71 ± 0,07
a,## 0,64 ± 0,06
b 0,64 ± 0,05
a,b
21 °C 0,61 ± 0,06##
0,61 ± 0,06 0,63 ± 0,05
ALH [µm]
K 32 °C 2,32 ± 0,47 2,34 ± 0,45 2,27 ± 0,46
30 min 32 °C 2,31 ± 0,36 2,43 ± 0,41 2,29 ± 0,45
21 °C 2,29 ± 0,44 2,40 ± 0,49 2,34 ± 0,45
60 min 32 °C 2,19 ± 0,49 2,23 ± 0,39
# 2,28 ± 0,38
21 °C 2,27 ± 0,47a 2,44 ± 0,49
b,# 2,35 ± 0,49
a,b
3 h 32 °C 2,28 ± 0,44 2,24 ± 0,66 2,15 ± 0,59
21 °C 2,31 ± 0,48 2,46 ± 0,52 2,30 ± 0,43
6 h 32 °C 2,11 ± 0,53
# 2,24 ± 0,58 2,16 ± 0,48
21 °C 2,47 ± 0,6# 2,47 ± 0,56 2,39 ± 0,65
BCF [Hz]
K 32 °C 40,6 ± 0,64 37,5 ± 3,6 37,1 ± 3,4
30 min 32 °C 40,4 ± 1,2
a 39,3 ± 2,9
a,b 36,2 ± 4,9
b
21 °C 40,1 ± 2,4a 38,7 ± 3,6
a,b 37,0 ± 3,0
b
60 min 32 °C 39,2 ± 2,8 38,2 ± 3,5 37,7 ± 4,8
21 °C 40,3 ± 1,3 39,2 ± 2,4 37,7 ± 3,6
3 h 32 °C 39,8 ± 1,5 35,5 ± 9,7 34,0 ± 12,0
21 °C 39,7 ± 1,2 38,6 ± 2,6 36,2 ± 5,5
6 h 32 °C 38,1 ± 5,2 37,3 ± 6,4 36,1 ± 6,6
21 °C 39,9 ± 2,9a 39,2 ± 3,4
a,b 36,7 ± 5,9
b
K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
# - ###: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltetemperaturen bei gleicher Lagerungsdauer (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency
Tabelle 9: Membranintakte Spermien in % (MW ± SD) von Experiment 4 a in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber)
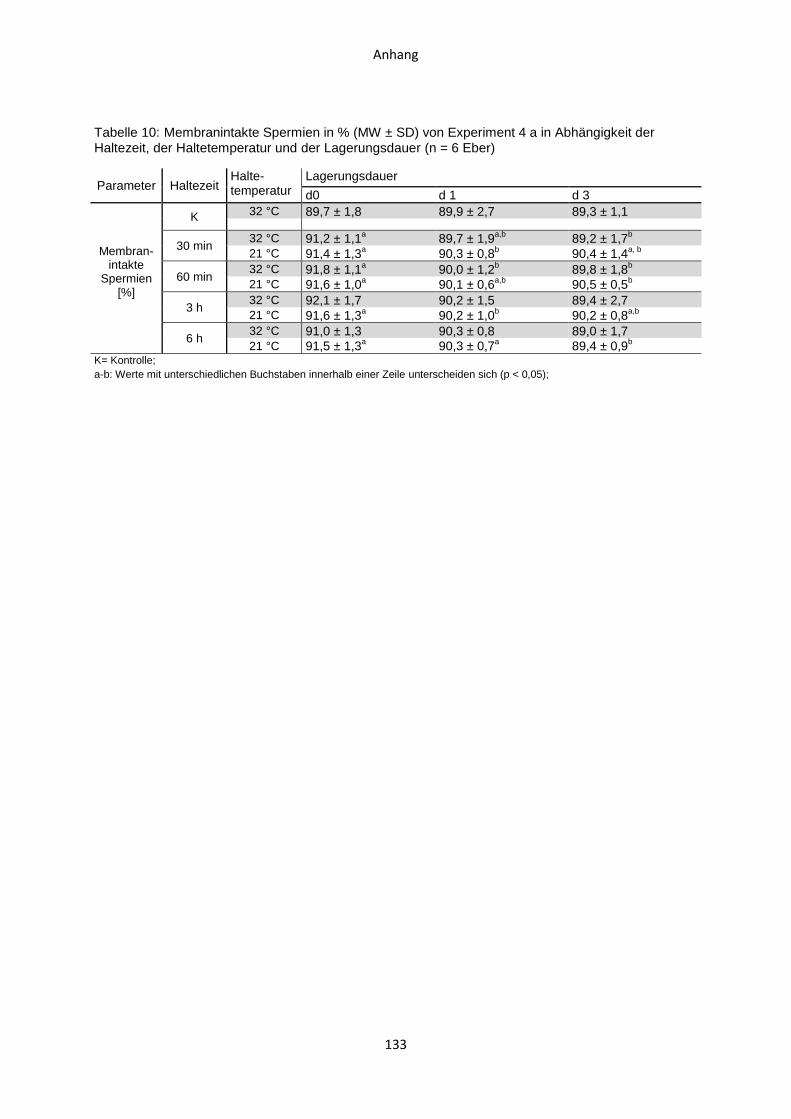
Anhang
133
Tabelle 10: Membranintakte Spermien in % (MW ± SD) von Experiment 4 a in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Halte-temperatur
Lagerungsdauer
d0 d 1 d 3
Membran-intakte
Spermien [%]
K 32 °C 89,7 ± 1,8 89,9 ± 2,7 89,3 ± 1,1
30 min 32 °C 91,2 ± 1,1
a 89,7 ± 1,9
a,b 89,2 ± 1,7
b
21 °C 91,4 ± 1,3a
90,3 ± 0,8b
90,4 ± 1,4a, b
60 min 32 °C 91,8 ± 1,1
a 90,0 ± 1,2
b 89,8 ± 1,8
b
21 °C 91,6 ± 1,0a
90,1 ± 0,6a,b
90,5 ± 0,5b
3 h 32 °C 92,1 ± 1,7 90,2 ± 1,5 89,4 ± 2,7
21 °C 91,6 ± 1,3a
90,2 ± 1,0b
90,2 ± 0,8a,b
6 h 32 °C 91,0 ± 1,3 90,3 ± 0,8 89,0 ± 1,7
21 °C 91,5 ± 1,3a
90,3 ± 0,7a
89,4 ± 0,9b
K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05);
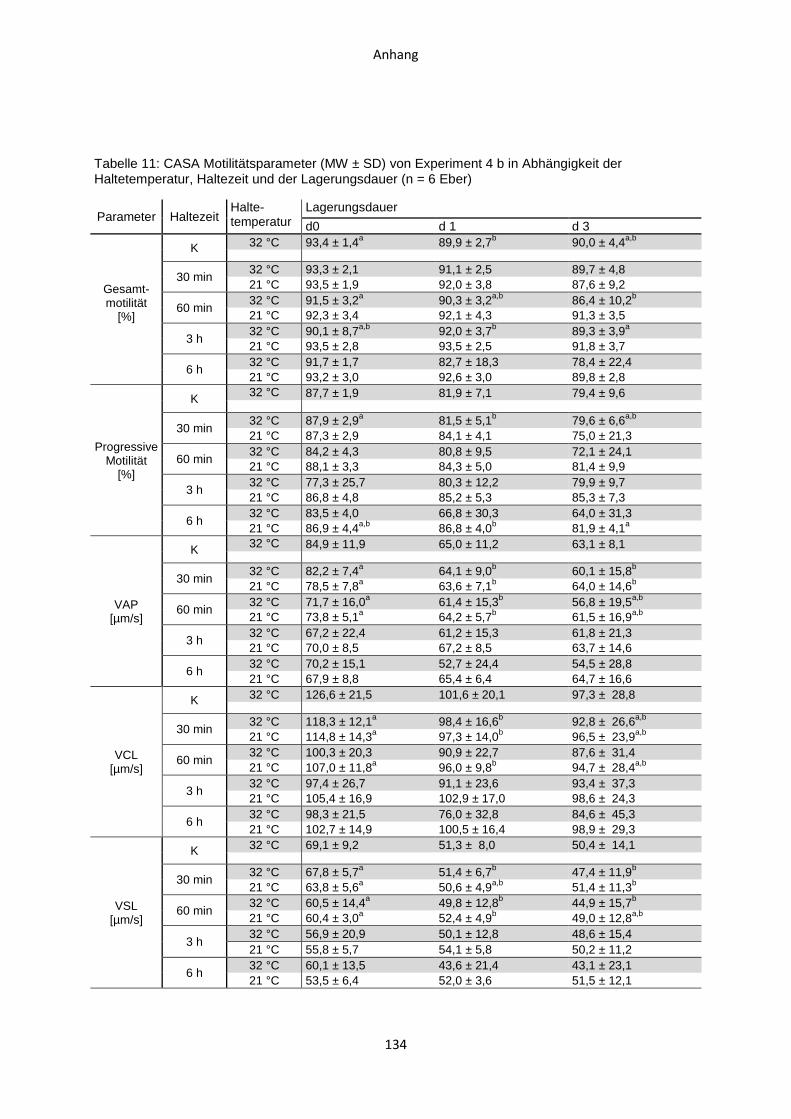
Anhang
134
Tabelle 11: CASA Motilitätsparameter (MW ± SD) von Experiment 4 b in Abhängigkeit der Haltetemperatur, Haltezeit und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Halte-temperatur
Lagerungsdauer
d0 d 1 d 3
Gesamt-motilität
[%]
K 32 °C 93,4 ± 1,4a 89,9 ± 2,7
b 90,0 ± 4,4
a,b
30 min 32 °C 93,3 ± 2,1 91,1 ± 2,5 89,7 ± 4,8
21 °C 93,5 ± 1,9 92,0 ± 3,8 87,6 ± 9,2
60 min 32 °C 91,5 ± 3,2
a 90,3 ± 3,2
a,b 86,4 ± 10,2
b
21 °C 92,3 ± 3,4 92,1 ± 4,3 91,3 ± 3,5
3 h 32 °C 90,1 ± 8,7
a,b 92,0 ± 3,7
b 89,3 ± 3,9
a
21 °C 93,5 ± 2,8 93,5 ± 2,5 91,8 ± 3,7
6 h 32 °C 91,7 ± 1,7 82,7 ± 18,3 78,4 ± 22,4
21 °C 93,2 ± 3,0 92,6 ± 3,0 89,8 ± 2,8
Progressive Motilität
[%]
K 32 °C 87,7 ± 1,9 81,9 ± 7,1 79,4 ± 9,6
30 min 32 °C 87,9 ± 2,9
a 81,5 ± 5,1
b 79,6 ± 6,6
a,b
21 °C 87,3 ± 2,9 84,1 ± 4,1 75,0 ± 21,3
60 min 32 °C 84,2 ± 4,3 80,8 ± 9,5 72,1 ± 24,1
21 °C 88,1 ± 3,3 84,3 ± 5,0 81,4 ± 9,9
3 h 32 °C 77,3 ± 25,7 80,3 ± 12,2 79,9 ± 9,7
21 °C 86,8 ± 4,8 85,2 ± 5,3 85,3 ± 7,3
6 h 32 °C 83,5 ± 4,0 66,8 ± 30,3 64,0 ± 31,3
21 °C 86,9 ± 4,4a,b
86,8 ± 4,0b 81,9 ± 4,1
a
VAP [µm/s]
K 32 °C 84,9 ± 11,9 65,0 ± 11,2 63,1 ± 8,1
30 min 32 °C 82,2 ± 7,4
a 64,1 ± 9,0
b 60,1 ± 15,8
b
21 °C 78,5 ± 7,8a 63,6 ± 7,1
b 64,0 ± 14,6
b
60 min 32 °C 71,7 ± 16,0
a 61,4 ± 15,3
b 56,8 ± 19,5
a,b
21 °C 73,8 ± 5,1a 64,2 ± 5,7
b 61,5 ± 16,9
a,b
3 h 32 °C 67,2 ± 22,4 61,2 ± 15,3 61,8 ± 21,3
21 °C 70,0 ± 8,5 67,2 ± 8,5 63,7 ± 14,6
6 h 32 °C 70,2 ± 15,1 52,7 ± 24,4 54,5 ± 28,8
21 °C 67,9 ± 8,8 65,4 ± 6,4 64,7 ± 16,6
VCL [µm/s]
K 32 °C 126,6 ± 21,5 101,6 ± 20,1 97,3 ± 28,8
30 min 32 °C 118,3 ± 12,1
a 98,4 ± 16,6
b 92,8 ± 26,6
a,b
21 °C 114,8 ± 14,3a 97,3 ± 14,0
b 96,5 ± 23,9
a,b
60 min 32 °C 100,3 ± 20,3 90,9 ± 22,7 87,6 ± 31,4
21 °C 107,0 ± 11,8a 96,0 ± 9,8
b 94,7 ± 28,4
a,b
3 h 32 °C 97,4 ± 26,7 91,1 ± 23,6 93,4 ± 37,3
21 °C 105,4 ± 16,9 102,9 ± 17,0 98,6 ± 24,3
6 h 32 °C 98,3 ± 21,5 76,0 ± 32,8 84,6 ± 45,3
21 °C 102,7 ± 14,9 100,5 ± 16,4 98,9 ± 29,3
VSL [µm/s]
K 32 °C 69,1 ± 9,2 51,3 ± 8,0 50,4 ± 14,1
30 min 32 °C 67,8 ± 5,7
a 51,4 ± 6,7
b 47,4 ± 11,9
b
21 °C 63,8 ± 5,6a 50,6 ± 4,9
a,b 51,4 ± 11,3
b
60 min 32 °C 60,5 ± 14,4
a 49,8 ± 12,8
b 44,9 ± 15,7
b
21 °C 60,4 ± 3,0a 52,4 ± 4,9
b 49,0 ± 12,8
a,b
3 h 32 °C 56,9 ± 20,9 50,1 ± 12,8 48,6 ± 15,4
21 °C 55,8 ± 5,7 54,1 ± 5,8 50,2 ± 11,2
6 h 32 °C 60,1 ± 13,5 43,6 ± 21,4 43,1 ± 23,1
21 °C 53,5 ± 6,4 52,0 ± 3,6 51,5 ± 12,1

Anhang
135
STR
K 32 °C 0,81 ± 0,03 0,79 ± 0,03 0,80 ± 0,02
30 min
32 °C 0,82 ± 0,03 0,80 ± 0,04 0,79 ± 0,02
21 °C 0,81 ± 0,04 0,79 ± 0,04 0,80 ± 0,02
60 min
32 °C 0,84 ± 0,03a 0,81 ± 0,03
b 0,78 ± 0,03
b
21 °C 0,81 ± 0,04 0,81 ± 0,02 0,79 ± 0,02
3 h
32 °C 0,83 ± 0,06 0,82 ± 0,03 0,79 ± 0,03
21 °C 0,80 ± 0,05 0,80 ± 0,02 0,78 ± 0,02
6 h 32 °C 0,85 ± 0,02
a,b,# 0,81 ± 0,04
a 0,78 ± 0,03
b
21 °C 0,79 ± 0,03# 0,79 ± 0,04 0,80 ± 0,04
LIN
K 32 °C 0,55 ± 0,05 0,51 ± 0,05 0,51 ± 0,02
30 min
32 °C 0,57 ± 0,06 0,52 ± 0,06 0,51 ± 0,03
21 °C 0,56 ± 0,07 0,52 ± 0,05 0,53 ± 0,03
60 min
32 °C 0,60 ± 0,07a 0,54 ± 0,05
b 0,51 ± 0,05
b
21 °C 0,57 ± 0,06 0,54 ± 0,0 0,52 ± 0,03
3 h
32 °C 0,56 ± 0,11 0,55 ± 0,05 0,53 ± 0,05
21 °C 0,53 ± 0,08 0,53 ± 0,04 0,51 ± 0,03
6 h 32 °C 0,61 ± 0,06
a,# 0,55 ± 0,07
a,b 0,50 ± 0,07
b
21 °C 0,52 ± 0,06# 0,52 ± 0,07 0,53 ± 0,06
WOB
K 32 °C 0,67 ± 0,04 0,64 ± 0,04 0,64 ± 0,02
30 min
32 °C 0,69 ± 0,05a 0,65 ± 0,04
b 0,65 ± 0,02
a,b
21 °C 0,68 ± 0,05 0,65 ± 0,03 0,66 ± 0,02
60 min
32 °C 0,71 ± 0,05a,#
0,67 ± 0,04b 0,65 ± 0,04
b
21 °C 0,69 ± 0,05# 0,67 ± 0,04 0,65 ± 0,02
3 h
32 °C 0,67 ± 0,09 0,67 ± 0,04 0,67 ± 0,05
21 °C 0,66 ± 0,06 0,65 ± 0,03 0,65 ± 0,02
6 h 32 °C 0,71 ± 0,05 0,68 ± 0,05 0,64 ± 0,06
21 °C 0,66 ± 0,05 0,66 ± 0,05 0,66 ± 0,05
ALH [µm]
K 32 °C 2,75 ± 0,66 2,18 ± 0,44 2,2 ± 0,55
30 min 32 °C 2,55 ± 0,62 2,22 ± 0,42 2,19 ± 0,50
21 °C 2,52 ± 0,58 2,18 ± 0,33 2,18 ± 0,48
60 min 32 °C 2,18 ± 0,57 2,00 ± 0,38 2,11 ± 0,65
21 °C 2,32 ± 0,45 2,18 ± 0,34 2,20 ± 0,59
3 h 32 °C 2,14 ± 0,40 2,01 ± 0,47 2,07 ± 0,68
21 °C 2,21 ± 0,32 2,27 ± 0,40 2,24 ± 0,49
6 h 32 °C 2,12 ± 0,47 1,79 ± 0,43 1,98 ± 0,79
21 °C 2,17 ± 0,30 2,29 ± 0,45 2,19 ± 0,63
BCF [Hz]
K 32 °C 39,8 ± 3,1 37,9 ± 3,9 35,9 ± 6,2
30 min 32 °C 40,4 ± 1,4
a 37,0 ± 2,9
a,b 34,3 ± 4,8
b
21 °C 40,0 ± 1,3 36,7 ± 2,9 36,2 ± 5,5
60 min 32 °C 37,8 ± 4,5 34,9 ± ,2 32,8 ± 8,9
21 °C 39,9 ± 1,2 36,9 ± 2,0 35,1 ± 6,3
3 h 32 °C 35,9 ± 10,0 35,0 ± 7,3 34,1 ± 8,1
21 °C 39,0 ± 2,8 38,1 ± 3,1 36,9 ± 5,8
6 h 32 °C 37,2 ± 4,1 29,2 ± 12,6 29,6 ± 13,9
21 °C 39,0 ± 3,2 37,6 ± 2,5 35,7 ± 5,6 K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
#: gleiche Symbole zeigen signifikante Unterschiede zwischen Haltetemperaturen bei gleicher Lagerungsdauer (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN =
linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency
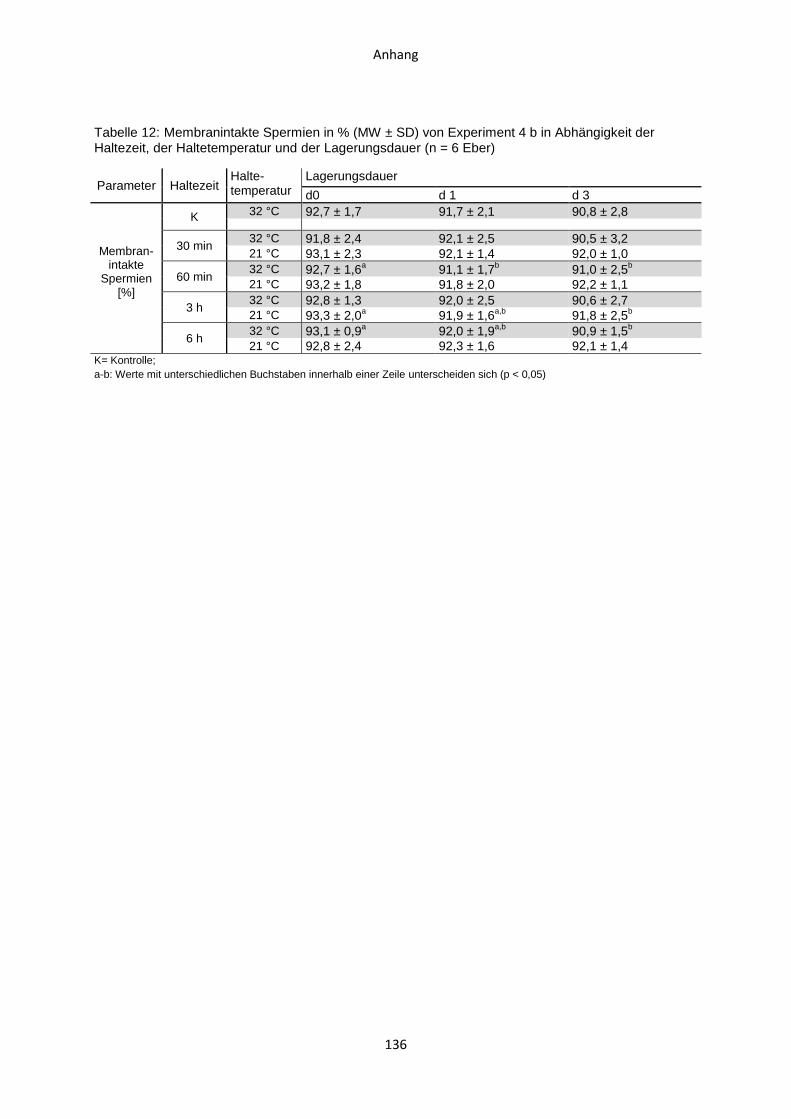
Anhang
136
Tabelle 12: Membranintakte Spermien in % (MW ± SD) von Experiment 4 b in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber)
Parameter Haltezeit Halte-temperatur
Lagerungsdauer
d0 d 1 d 3
Membran-intakte
Spermien [%]
K 32 °C 92,7 ± 1,7 91,7 ± 2,1 90,8 ± 2,8
30 min 32 °C 91,8 ± 2,4 92,1 ± 2,5 90,5 ± 3,2
21 °C 93,1 ± 2,3 92,1 ± 1,4 92,0 ± 1,0
60 min 32 °C 92,7 ± 1,6
a 91,1 ± 1,7
b 91,0 ± 2,5
b
21 °C 93,2 ± 1,8 91,8 ± 2,0 92,2 ± 1,1
3 h 32 °C 92,8 ± 1,3 92,0 ± 2,5 90,6 ± 2,7
21 °C 93,3 ± 2,0a
91,9 ± 1,6a,b
91,8 ± 2,5b
6 h 32 °C 93,1 ± 0,9
a 92,0 ± 1,9
a,b 90,9 ± 1,5
b
21 °C 92,8 ± 2,4 92,3 ± 1,6 92,1 ± 1,4 K= Kontrolle;
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)

Anhang
137
Tabelle 13: CASA Motilitätsparameter (MW ± SD) von Experiment 5 in Abhängigkeit vom Abkühlregime und der Lagerungsdauer (n = 6 Eber)
Parameter Abkühlregime Lagerungsdauer
d1 d 3 d 6
Gesamt-motilität [%]
32°C–21°C–17°C
32°C–17°C
85,8 ± 4,7
84,8 ± 2,0
87,9 ± 3,6
86,5 ± 2,0
84,7 ± 4,5
80,5 ± 5,1
Progressive Motilität [%]
32°C–21°C–17°C
32°C–17°C
75,8 ± 7,5
74,1 ± 7,3
78,0 ± 6,0
77,22 ± 3,8
73,3 ± 9,3
64,23 ± 9,3
VAP [µm/s]
32°C–21°C–17°C
32°C–17°C
64,9 ± 6,0
62,6 ± 10,7
61,3 ± 8,4
63,2 ± 12,0
64,6 ± 5,1
54,1 ± 12,9
VCL [µm/s]
32°C–21°C–17°C
32°C–17°C
108,5 ± 19,5
104,7 ± 28,0
98,7 ± 16,5
103,6 ± 27,4
98,5 ± 11,5
85,9 ± 24,8
VSL [µm/s]
32°C–21°C–17°C
32°C–17°C
47,1 ± 3,4
45,5 ± 4,0
45,6 ± 6,1
46,7 ± 4,3
51,6 ± 5,4
41,2 ± 7,9
STR 32°C–21°C–17°C
32°C–17°C
0,73 ± 0,09a
0,74 ± 0,09
0,74 ± 0,06a,b
0,75 ± 0,09
0,79 ± 0,07b
0,77 ± 0,08
LIN
32°C–21°C–17°C
32°C–17°C
0,44 ± 0,09a
0,45 ± 0,1
0,46 ± 0,06a,b
0,47 ± 0,09
0,53 ± 0,09b
0,49 ± 0,09
WOB
32°C–21°C–17°C
32°C–17°C
0,60 ± 0,05a
0,61 ± 0,06
0,62 ± 0,04a,b
0,61 ± 0,05
0,66 ± 0,06b
0,64 ± 0,05
ALH [µm]
32°C–21°C–17°C
32°C–17°C
2,62 ± 0,48a
2,52 ± 0,61a,b
2,39 ± 0,48a,b
2,48 ± 0,57a
2,32 ± 0,39a
2,09 ± 0,53b
BCF [Hz]
32°C–21°C–17°C
32°C–17°C
36,8 ± 1,3
35,1 ± 2,2
34,4 ± 3,3
35,2 ± 2,7
35,5 ± 1,9
30,7 ± 4,6
K= Kontrolle
a-b: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency
Tabelle 14: Membranintakte Spermien in % (MW ± SD) von Experiment 5 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber)
Parameter Abkühlregime Lagerungsdauer
d1 d 3 d 6
Membran-intakte
Spermien [%]
32°C–21°C–17°C
32°C–17°C
86,80 ± 3,45a
86,15 ± 3,31a
84,30 ± 3,45b
83,78 ± 2,91b
82,97 ± 2,73b
81,28 ± 2,48c
K= Kontrolle;
a-c: Werte mit unterschiedlichen Buchstaben innerhalb einer Zeile unterscheiden sich (p < 0,05)

Anhang
138
Tabelle 15: CASA Motilitätsparameter (MW ± SD) von Experiment 6 in Abhängigkeit von der Art der
Verarbeitung und der Lagerungsdauer (n = 6 Eber)
Parameter Verarbeitung Lagerungsdauer
d 1 d 3 d 6
Gesamt-motilität [%]
Standard
Station
88,8 ± 2,7
90,3 ± 2,6
87,9 ± 3,0
90,6 ± 1,9
82,6 ± 7,0
85,6 ± 5,6
Progressive Motilität [%]
Standard
Station
83,7 ± 4,5
86,3 ± 4,0
81,3 ± 7,3#
85,3 ± 6,0#
73,3 ± 17,3#
76,8 ± 18,2#
VAP [µm/s] isotherm
hypotherm
70,4 ± 16,3
69,2 ± 12,6
66,8 ± 14,5
69,1 ± 12,5
64,6 ± 22,7
63,5 ± 21,1
VCL [µm/s] isotherm
hypotherm
103,7 ± 29,8
99,8 ± 23,5
93,1 ± 22,3
98,4 ± 18,3
89,5 ± 30,0
88,4 ± 29,5
VSL [µm/s] isotherm
hypotherm
54,5 ± 9,0
55,3 ± 7,6
54,5 ± 9,8
56,2 ± 8,6
53,3 ± 18,6
52,1 ± 17,6
STR isotherm
hypotherm
0,78 ± 0,10
0,8 ± 0,07
0,82 ± 0,06
0,81 ± 0,04
0,82 ± 0,04
0,81 ± 0,04
LIN
isotherm
hypotherm
0,54 ± 0,11
0,57 ± 0,11
0,59 ± 0,08
0,57 ± 0,06
0,59 ± 0,07
0,59 ± 0,07
WOB
isotherm
hypotherm
0,69 ± 0,06
0,7 ± 0,08
0,72 ± 0,05
0,7 ± 0,04
0,71 ± 0,05
0,71 ± 0,04
ALH [µm] isotherm
hypotherm
2,25 ± 0,68
2,11 ± 0,52
1,99 ± 0,47
2,07 ± 0,45
1,94 ± 0,5
1,93 ± 0,47
BCF [Hz] isotherm
hypotherm
37,4 ± 4,8
37,2 ± 4,6
36,1 ± 4,2
37,7 ± 3,3
33,8 ± 7,5
33,8 ± 8,1
#: gleiche Symbole zeigen signifikante Unterschiede zwischen der Art Verarbeitung bei gleicher Lagerungsdauer (p < 0,05)
VAP = velocity average path; VCL = velocity curved line; VSL = velocity straight-line; STR = straightness (VSL/VAP); LIN = linearity (VAP/VCL); WOB = wobble (VAP/VCL); ALH = amplitude of lateral head-displacement; BCF = beat cross frequency
Tabelle 16: Ergebnisse der durchflusszytometrischen Messungen in % (MW ± SD) von Experiment 6 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber)
Parameter Verarbeitung Lagerungsdauer
d 1 d 3 d 6
Membran-integrität[%]
Standard
Station 92,3 ± 1,8 91,9 ± 1,6
92,1 ± 1,9 92,0 ± 1,7
90,7 ± 2,8 91,4 ± 1,5
Spezifische Reaktivität gg. Bicarbonat [%]
Standard
Station 31,7 ± 7,4
#
40,3 ± 6,5#
#: gleiche Symbole zeigen signifikante Unterschiede zwischen der Art Verarbeitung bei gleicher Lagerungsdauer (p < 0,05)

Anhang
139
Abb. 41: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VAP (velocity average path; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
Ante
il der
Sperm
ien p
ro K
lasse in %
Geschwindigkeit in µm/ s
VAP
K d0
21-6 d0
32-6 d0
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
Ante
il der
Sperm
ien p
ro K
lasse in %
Geschwindigkeit in µm/ s
VAP
Kontrolle
32-6
21-6
Kontrolle*-d1
32°C-6h-d1
21°C-6h-d1
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
A
B

Anhang
140
Abb. 42: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VSL (velocity straight line; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
20%
Sper
mie
nan
teil
pro
Kla
sse
in %
Geschwindigkeitsklassen in µm/s
VSL
K d0
21-6 d0
32-6 d0
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
20%
Sper
mie
nan
teil
pro
Kla
sse
in %
Geschwindigkeitsklassen in µm/s
VSL
K d1
21-6 d1
32-6 d1
B
A
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1
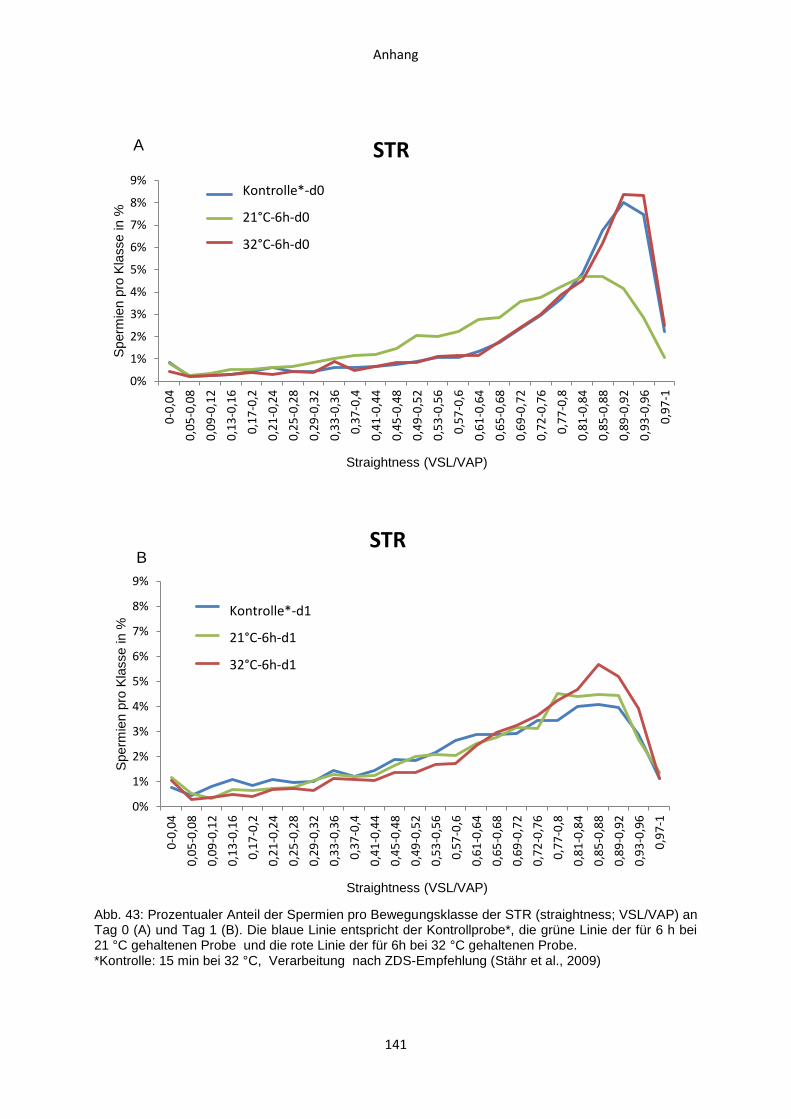
Anhang
141
Abb. 43: Prozentualer Anteil der Spermien pro Bewegungsklasse der STR (straightness; VSL/VAP) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%0
-0,0
4
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sperm
ien p
ro K
lasse in %
Straightness (VSL/VAP)
STR
K d0
21-6 d0
32-6 d0
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
0-0
,04
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sperm
ien p
ro K
lasse in %
Straightness (VSL/VAP)
STR
K d1
21-6 d1
32-6 d1
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
A
B

Anhang
142
Abb. 44: Prozentualer Anteil der Spermien pro Bewegungsklasse der LIN (linearity; VSL/VCL) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
1%
2%
3%
4%
5%
0-0
,04
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sper
mie
nan
teil
pro
Kla
sse
in %
Linearität (VSL/VCL)
LIN K d0
21-6 d0
32-6 d0
0%
1%
2%
3%
4%
5%
0-0
,04
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sper
mie
nan
teil
pro
Kla
sse
in %
Linearität (VSL/VCL)
LIN 21-6 d1
32-6
K-d1
21°C-6h-d1
32°C-6h-d1
Kontrolle*-d1
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
A
B

Anhang
143
Abb. 45: Prozentualer Anteil der Spermien pro Bewegungsklasse der WOB (wobble; VAP/VCL) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
1%
2%
3%
4%
5%
6%
7%
8%0
-0,0
4
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sperm
ien p
ro K
lasse in %
Wobble (VAP/VCL)
WOB
K d0
21-6 d0
32-6 d0
0%
1%
2%
3%
4%
5%
6%
7%
8%
0-0
,04
0,0
5-0
,08
0,0
9-0
,12
0,1
3-0
,16
0,1
7-0
,2
0,2
1-0
,24
0,2
5-0
,28
0,2
9-0
,32
0,3
3-0
,36
0,3
7-0
,4
0,4
1-0
,44
0,4
5-0
,48
0,4
9-0
,52
0,5
3-0
,56
0,5
7-0
,6
0,6
1-0
,64
0,6
5-0
,68
0,6
9-0
,72
0,7
2-0
,76
0,7
7-0
,8
0,8
1-0
,84
0,8
5-0
,88
0,8
9-0
,92
0,9
3-0
,96
0,9
7-1
Sperm
ien p
ro K
lasse in %
Wobble (VAP/VCL)
WOB
K d1
21-6 d1
32-6 d1
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1
A
B
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0

Anhang
144
Abb. 46: Prozentualer Anteil der Spermien pro Bewegungsklasse der ALH (amplitude of lateral head displacement; µm) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al.,2009)
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
20%
0-0,5 0,5-1 1-1,5 1,5-2 2-2,5 2,5-3 3-3,5 3,5-4 4-4,5 4,5-5 5-5,5 5,5-6
Sperm
ien p
ro K
lasse in %
amplitude of lateral head displacement (µm)
ALH
K d0
21-6 d0
32-6 d0
0%
2%
4%
6%
8%
10%
12%
14%
16%
18%
20%
0-0,5 0,5-1 1-1,5 1,5-2 2-2,5 2,5-3 3-3,5 3,5-4 4-4,5 4,5-5 5-5,5
Sperm
ien p
ro K
lasse in %
amplitude of lateral head displacement (µm)
ALH
K d1
21-6 d1
32-6 d1
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
A
B

Anhang
145
Abb. 47: Prozentualer Anteil der Spermien pro Bewegungsklasse der BCF (beat cross frequency; Hz) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C gehaltenen Probe. *Kontrolle: 15 min bei 32 °C, Verarbeitung nach ZDS-Empfehlung (Stähr et al., 2009)
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
0-4 6,1-8 10,1-12 14,1-16 18,1-20 22,1-24 26,1-28 30,1-32 34,1-36 38,1-40 42,1-44 46,1-48
Sperm
ien p
ro K
lasse in %
beat cross frequency (Hz)
BCF K d0
21-6 d0
32-6 d0
0%
1%
2%
3%
4%
5%
6%
7%
8%
9%
0-4 6,1-8 10,1-12 14,1-16 18,1-20 22,1-24 26,1-28 30,1-32 34,1-36 38,1-40 42,1-44 46,1-48
Sperm
ien p
ro K
lasse in %
beat cross frequency (Hz)
BCF K d1
21-6 d1
32-6 d1
Kontrolle*-d1
21°C-6h-d1
32°C-6h-d1
Kontrolle*-d0
21°C-6h-d0
32°C-6h-d0
A
B

Anhang
146
9.2 Chemikalien
Substanz Bezugsquelle Artikelnummer
A23187 (Calcimycin) Alexis Chemicals, Lausen (CH) 52665-69-7
Adenylatkinase Sigma Aldrich, Steinheim M 3003
ATP-Bioluminescent-Assay-Kit Sigma Aldrich, Steinheim FLAA
BSA, Fraction V SERVA Electrophoresis GmbH, Heidelberg
1193-03
Calciumchlorid Merck, Darmstadt 102393
Dimethylsulfoxid (DMSO) Roth, Karlsruhe 4720.4
DNA Control UV Partec GmbH, Münster 05-4018
FITC-PNA Axorra, Lörrach C-FL-1071-M01
Fluo-3 AM ultra pure grade Biomol, Hamburg ABD-21011
Formaldehyd 37% Roth, Karlsruhe 4980.2
HEPES PUFFERAN® Roth, Karlsruhe HN78
Hoechst 33342 Invitrogen™/ Life Technologies Carlsbad (USA)
H-1399
JC-1 iodide Enzo Lifescience, Lörrach ALX-620-053-M005
Kaliumchlorid AppliChem GmbH, Darmstadt A3582
Kaliumhydroxid AppliChem GmbH, Darmstadt A3871
Magnesiumsufat Sigma Aldrich, Schnelldorf M5921
Magnesiumchlorid Roth, Karlsruhe 2189
Natriumchlorid Roth, Karlsruhe 3957
Phosphatase Inhibitor Cocktail 2 Sigma Aldrich, Steinheim P5725

Anhang
147
Phosphoenolpyruvat Sigma Aldrich, Steinheim P7022
Propidiumiodid (PI) Sigma Aldrich, Schnelldorf P-4170-100MG
Pyruvatkinase Sigma Aldrich, Steinheim P1506
Tricin Sigma Aldrich, Schnelldorf T 0377
Tri-Natriumcitrat-Dihydrat Roth, Karlsruhe 3580.3
α-D(+)-Glucose-Monohydrat Roth, Karlsruhe 6780
9.3 Rezepte
9.3.1 Formolcitrat
9,86 mmol tri-Natriumcitrat-Dihydrat
ad 100 ml aqua dest
4 ml der Lösung verwerfen, durch 4 ml 37 %-iges Formalin ersetzen und vermischen.
Die Aufbewahrung der Lösung erfolgte aliquotiert (300 µl) bei 4 °C
9.3.2 HBS (HEPES®- buffered saline)
137,0 mmol NaCl
20,0 mmol HEPES®
10,0 mmol α-D(+)-Glucose-Monohydrat
2,5 mmol KOH
ad 1000 ml aqua dest
Osmolarität: 300 mOsmol/kg
pH 7,4 bei Gebrauchstemperatur von 38 °C (Titration mit 1 M NaOH)
Die Aufbewahrung erfolgte für höchstens 24 h in einem verschlossenen Glaskolben
im Kühlschrank

Anhang
148
9.3.3 Tyrode basierte Medien
9.3.3.1 Herstellung der Stammlösungen
NaCl 0,5 M
KCl 0,5 M
MgSO4 * 7H2O 0,1 M
Glucose * H2O 0,5 M
NaHCO3 0,3 M
CaCl2 0,5 M
KH2PO4 0,1 M
Hepes 0,2 M (pH 7,55 bei 20 °C mit NaOH (1 M) eingestellt)
Gentamycinsulfat 20 mg/ ml
9.3.3.2 Herstellung der Tyr BikCa-Vormischung
Je 96,0 mmol NaCl-Stammlösung
3,1 mmol KCl-Stammlösung
0,4 mmol MgSO4 * 7H2O-Stammlösung
5,0 mmol Glucose * H2O-Stammlösung
15,0 mmol NaHCO3-Stammlösung
2,0 mmol CaCl2-Stammlösung
0,3 mmol KH2PO4-Stammlösung
20,0 mmol Hepes-Stammlösung
wurden in eine Eberflasche pipettiert und bei -20 °C bis zum Gebrauch gelagert. Am
Versuchstag wurde die Vormischung aufgetaut und wie unter … beschrieben
komplettiert.

Anhang
149
9.3.3.3 Herstellung der TyrCa-Vormischung
Je 111,0 mmol NaCl-Stammlösung
3,1 mmol KCl-Stammlösung
0,4 mmol MgSO4 * 7H2O-Stammlösung
5,0 mmol Glucose * H2O-Stammlösung
2,0 mmol CaCl2-Stammlösung
0,3 mmol KH2PO4-Stammlösung
20,0 mmol Hepes-Stammlösung
wurden in eine Eberflasche pipettiert und bei -20 °C bis zum Gebrauch gelagert. Am
Versuchstag wurde die Vormischung aufgetaut und wie unter … beschrieben
komplettiert.
9.3.3.4 Komplettierung der Medien
Tyr BikCa - bzw. TyrCa –Vormischung
21,7 mmol Na-Lactat
1,0 mmol Na-Pyruvat
0,5 ml Gentamycin-Stammlösung
0,002 g Phenolrot
ad 100 ml Aqua dest
Die Osmolarität der Lösungen lag bei 300 ± 5 mOsmol/ kg. Der pH des
Kapazitationsmediums wurde auf 7,6 (+20 °C) eingestellt, der pH-Wert des nicht
kapazitierenden Mediums lag bei 7,55 (+20 °C). Nach Sterilfiltration wurden die
beiden Medien mit jeweils 3 mg/ ml BSA versetzt.

Anhang
150
9.4 Geräte und Verbrauchsmaterialien
96 well Mikrotiterplatte
white wall, clear bottom
Fa. Biohit, Rosbach
CO2-Inkubator MCO-17AC Sanyo, Bad Nenndorf
Deckgläser 18 x 18 mm Interessengemeinschaft der
Laborfachhändler (IDL) GmbH & Co KG,
Nidderau
Durchflusszytometer Dako Galaxy Partec GmbH, Münster
Ebersamenflaschen mit Verschluss Fa. Minitüb GmbH & Co KG, Tiefenbach
Einmalhandschuhe
Manuplast
Nobaglove Latex
Nobaglove Nitril
Vinyl 2000 PF
Landgraf Laborsysteme, Langenhagen
NOBA Verbandmittel Danz GmbH & Co
KG, Wetter
Meditrade, Kiefersfelden
Einmalspritzen, steril 20 ml Fa. Braun, Melsungen
Filter; PES Membran; 0,22 µm
Porendurchmesser
Milipore, Irland
Gefrierpunktosmometer, Osmomat 030 Gonotec, Berlin
Glas-Filtrationsgerät mit Glasfilterplatte Interessenfachgemeinschaft der
Laborfachhändler (IDL) GmbH & Co KG,
Nidderau
Heizplatten
HT 50
HT 200
Fa. Minitüb GmbH & Co KG, Tiefenbach
Klimaschränke mit Digitalregler Fa. Minitüb GmbH & Co KG, Tiefenbach

Anhang
151
Laborwaagen
PT 12
ALC-80
Sartorius AG, Göttingen
Laborzentrifuge
Universal 30 RF
Sepatech Biofuge 13
Megafuge 2.0 R
Fa.Hettich, Tuttlingen
Fa. Heraeus, Hanau
Fa. Heraeus, Hanau
Magnetrührer, MR3001K Heidolph, Schwabach
Messgefäße Osmomat 030 Gonotec, Berlin
Mikromec multisens Fa. Minitüb GmbH & Co KG, Tiefenbach
NaCl-Röhrchen Sarstedt, Nürnbrecht
Objektträger 76 x 26 mm Gerhard Menzel GmbH, Braunschweig
Parafilm Landgraf Laborsysteme, Langenhagen
Phantom für Eber Fa. Minitüb GmbH & Co KG, Tiefenbach
Phasenkontrastmikroskope
Anxiostar Plus
Zeiss 4713849
Zeiss, Jena
pH-Meter, Multiplex 3000/ pmx WTW, Weilheim
Pipette, elektronisch
Biohit eLine E 1000
Fa. Biohit, Rosbach
Pipettenspitzen Nerbe plus, Winsen/ Luhe
Plattenphotometer
Tecan GENios pro plate reader
Tecan Group Ltd., Männedorf, Schweiz

Anhang
152
Reagenzröhrchen 3,5 ml mit
Eindrückstopfen
Sarstedt, Nürnbrecht
Reaktionsgefäße 1,5 ml
schwarz und transparent
Landgraf Laborsysteme, Langenhagen
Samenauffangbecher Fa. Minitüb GmbH & Co KG, Tiefenbach
Samenauffangbeutel, US Bag™ Fa. Minitüb GmbH & Co KG, Tiefenbach
Spritzenansatzfilter, PES-Membran,
0,22 µm Porendurchmesser, steril
Carl Roth GmbH & Co KG, Karlsruhe
Thermomixer 5436 Fa. Eppendorf, Hamburg
Variable Einkanalpipetten Eppendorf, Hamburg
Vortex, Reax top Heidolph, Schwabach
Wärmeschränke Memmert, Schwabach
Wasserbäder
Typ 1013
WNB 745
GFL, Burgwedel
Memmert, Schwabach
Wasserstrahlpumpe Landgraf Laborsysteme, Langenhagen
Zählkammer „Leja“
20 micron, 4-chamber-slide
Leja Products B.V., Nieuw-Vennep,
Niederlande
Zählkammer nach Thoma, „neu“ Fa. Jürgens, Hannover

Abbildungsverzeichnis
153
10 Abbildungsverzeichnis
Abb. 1: Übersicht aller durchgeführten Experimente .............................................................17
Abb. 2: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 1. Vergleich von
hypothermer und isothermer Verdünnung mit Haltezeiten bei zwei Haltetemperaturen (21 °C/
32 °C) und zwei Verdünnungsstufen (10 x 106 Spermien/ ml; 20 x 106 Spermien/ ml). .........23
Abb. 3: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 2. Vergleich von
Haltezeiten bei verschiedenen Haltetemperaturen (21 °C/ 32 °C) bei zweistufig isothermer
Verdünnung. .........................................................................................................................26
Abb. 4: Temperatur- und Zeitregime der Samenverarbeitung in Experiment 3. Vergleich
zweistufig isothermer und zweistufig hyperthermer Verdünnung nach Haltezeiten bei 21 °C.
.............................................................................................................................................28
Abb. 5: Temperatur- und Zeitregime der zweistufig hypothermen Samenverarbeitung in
Experiment 4 a. Vergleich von Haltezeiten bei verschiedenen Haltetemperaturen (21 °C/
32 °C) des ausverdünnten Samens nach zweistufig hypothermer Verdünnung. ...................30
Abb. 6: Temperatur- und Zeitregime der einstufig isothermen Samenverarbeitung in
Experiment 4 b. Vergleich von Haltezeiten bei verschiedenen Haltetemperaturen
(21 °C/ 32 °C) des ausverdünnten Samens nach einstufig isothermer Verdünnung. ............31
Abb. 7: Temperatur- und Zeitregime der einstufig isothermen Samenverarbeitung in
Experiment 5. Vergleich von einer beschleunigten Abkühlrate ohne Haltezeit zur
Standardabkühlung mit 90 min Haltezeit nach einstufig isothermer Verdünnung. .................32
Abb. 8: Prozesskette der Samenverarbeitung in einer Besamungsstation. Die grauen
Flächen stellen Zeitpunkte für mögliche Verzögerungen in der Verarbeitung dar, aus denen
sich Haltezeiten unterschiedlicher Länge ergeben. ...............................................................35
Abb. 9: Vergleich von Temperatur- und Zeitregime der Samenverarbeitung in der
Besamungsstation mit Laborstandard in Experiment 6. ........................................................37
Abb. 10: Beispiel einer Standardkurve des ATP-Assays ......................................................55
Abb. 11: Temperaturverlauf während der zweistufig hypothermen (blaue Linie) und
isothermen (rote Linie) Samenverarbeitung bei Experiment 1. Mittelwerte von je 3
Samenproben. Die Samenproben wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville
Thawing Solution (BTS, 32 °C) verdünnt. Für die zweistufig isotherme Verdünnung wurde
eine Hälfte des vorverdünnten Samen für ca. 15 min bei 32 °C (Wasserbad) gehalten um
dann mit BTS (32 °C) auf die Endkonzentration in Ebersamenflaschen verdünnt zu werden.
Abkühlung der Proben 90 min bei 21 °C anschließend weitere Abkühlung und Lagerung in
einen auf 17 °C temperierten Klimaschrank verbracht. Für zweistufig hypotherme
Verarbeitung wurde nach der Vorverdünnung die andere Hälfte der Probe für 20 min bei

Abbildungsverzeichnis
154
21 °C gehalten. Die weitere Verdünnung erfolgte mit auf 21 °C temperierter BTS, wodurch
der schnelle Abfall der Probentemperatur zu erklären ist. Die weitere Verarbeitung entsprach
den zweistufig isotherm verarbeiteten Proben. .....................................................................60
Abb. 12: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der Art der
zweistufigen Verdünnung (isotherm vs. hypotherm), Verdünnungsgrad (20 x 106 und
10 x 106 Spermien/ ml) und Lagerungsdauer (d 1, d 3, d 6) bei 17 °C; n = 6 Ejakulate. ........61
Abb. 13: Prozentualer Anteil membranintakter (PI und FITC-PNA negativer) Spermien (MW +
SD) in Abhängigkeit von der Art der zweistufigen Verdünnung (isotherm vs. hypotherm),
Verdünnungsgrad (20 x 106 und 10 x 106 Spermien/ ml) und Lagerungsdauer (d 1, d3, d 6)
bei 17 °C; n = 6 Ejakulate. ....................................................................................................63
Abb. 14: Temperaturverlauf der bei Experiment 2 bei 21 °C gehaltenen Proben. Mittelwerte
von je 3 Samenproben. Diese wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville
Thawing Solution (BTS, 32°C) verdünnt und zweistufig isotherm weiterverarbeitet. Je 200ml
des vorverdünnten Samen wurden bei 32 °C bzw. 21 °C gehalten (für 30 min, 60 min, 3 h
und 6 h), danach ausverdünnt (in 100 ml Ebersamenflaschen), für weitere 90 min bei 21 °C
gehalten und anschließend bei 17 °C gelagert. ....................................................................66
Abb. 15: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der
Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die
gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von
32 °C und blaue Säulen der Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen
Haltezeiten (30 min – 6 h); ...................................................................................................67
Abb. 16: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VCL (velocity
curvilinear; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der
Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der
für 6h bei 32 °C gehaltenen Probe. ......................................................................................69
Abb. 17: Prozentualer Anteil membranintakter (PI und FITC-PNA negative) Spermien (MW +
SD) in Abhängigkeit von der Haltetemperatur, Haltezeit und der Lagerungsdauer des
vorverdünnten Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den
Proben mit Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C. Die
Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=12 Ejakulate für d0 – d3;
n=6 Ejakulate für d6). ...........................................................................................................70
Abb. 18: Prozentualer Anteil der PI-negativen (Plasmamembran-intakten) Spermien mit
mehrheitlich hohem Mitochondrienmembranpotential (MMP) in Abhängigkeit von der
Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die
gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit Haltetemperatur von
32 °C und blaue Säulen der Haltetemperatur von 21 °C. Die Farbabstufungen:
unterschiedlichen Haltezeiten; n=6 Ejakulate. ......................................................................71

Abbildungsverzeichnis
155
Abb. 19: Mittlere Fluoreszenzintensität (orange Jagg) aller Spermien (MW + SD) in
Abhängigkeit von der Lagerungsdauer, Haltetemperatur und der Haltezeit des vorverdünnten
Samens. Die gestreifte Säule entspricht der Kontrolle*, rote Säulen den Proben mit
Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C.
Farbabstufungen: unterschiedlichen Haltezeiten;n=6 Ejakulate. ...........................................72
Abb. 20: ATP – Gehalt der Spermien in pmol/ 50000 Spemien (MW + SD) in Abhängigkeit
von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Die
grüne Säule entspricht dem ATP – Gehalt der unbehandelten Samenprobe, die gestreifte
Säule der Kontrolle*. Rote Säulen entsprechen den Proben mit einer Haltetemperatur von
32 °C und blaue Säulen der Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen
Haltezeiten (30 min – 6 h); n=6 Ejakulate. ............................................................................73
Abb. 21: Energieladung (EC) der Spermien in Abhängigkeit von der Lagerungsdauer, Halte-
temperatur und der Haltezeit des vorverdünnten Samens. Die grüne Säule entspricht der EC
der unbehandelten Samenproben, die gestreifte Säule der Kontrollen*. Rote Säulen
entsprechen den Proben mit einer Haltetemperatur von 32 °C und die blauen Säulen Proben
mit einer Haltetemperatur von 21 °C. Farbabstufungen: unterschiedlichen Haltezeiten
(30 min – 6 h); n=6 Ejakulate. ...............................................................................................74
Abb. 22: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW + SD) in Abhängigkeit
von der Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens (n =
4). .........................................................................................................................................75
Abb. 23: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW + SD) der Proben an
Tag 0 bei 32 °C (A) und 21 °C Haltetemperatur (B) in Abhängigkeit der Eber und der
Haltezeit des vorverdünnten Samens. Rote Säulen entsprechen den Proben mit einer
Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C (je 60 min und 6
h); n=4 Ejakulate ..................................................................................................................77
Abb. 24: Gesamtkeimzahlen (GKZ) der Samenproben in KbE/ ml (MW * SD) der Proben an
Tag 3 bei 32 °C (A) und 21 °C Haltetemperatur (B) in Abhängigkeit der Eber und der
Haltezeit des vorverdünnten Samens. Rote Säulen entsprechen den Proben mit einer
Haltetemperatur von 32 °C und blaue Säulen der Haltetemperatur von 21 °C (je 60 min und 6
h); n=4 Ejakulate. .................................................................................................................78
Abb. 25: Temperaturverlauf der bei Experiment 3 bei 21 °C gehaltenen und anschließend
hypertherm verdünnten Proben. Mittelwerte von je 3 Samenproben. Diese wurden im
Verhältnis 1:1 (200ml: 200 ml) mit Beltsville Thawing Solution (BTS, 32 °C) verdünnt und
zweistufig hypertherm bzw. isotherm weiterverarbeitet. Beide vorverdünnte Proben wurden
bei 21 °C gehalten (für 30 min, 60 min 3 h und 6 h), danach hypertherm (BTS, 32 °C) bzw.
isotherm ausverdünnt, für weitere 90 min bei 21 °C gehalten und anschließend bei 17 °C
gelagert. ...............................................................................................................................80
Abb. 26: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der
Verdünnertemperatur im zweiten Verdünnungsschritt, der Haltezeit des vorverdünnten

Abbildungsverzeichnis
156
Samens und der Lagerungsdauer. Dargestellt sind die Kontrolle* (gestreifte Säulen),
hypertherm ausverdünnte Proben (rote Säulen) und isotherm ausverdünnte Proben (blaue
Säulen). Farbabstufungen: unterschiedliche Haltezeiten; n=6 Ejakulate. ..............................81
Abb. 27: Prozentualer Anteil membranintakter (PI und FITC-PNA negative) Spermien (MW +
SD) in Abhängigkeit von der Verdünnertemperatur im zweiten Verdünnungsschritt, der
Haltezeit des vorverdünnten Samens und der Lagerungsdauer; n=6 Ejakulate ....................83
Abb. 28: Temperaturverlauf der bei Experiment 4a zweistufig hypotherm verdünnten und
anschließend bei 32 °C gehaltenen Proben. Mittelwerte von je 3 Samenproben. Diese
wurden im Verhältnis 1:1 (200 ml: 200 ml) mit Beltsville Thawing Solution (BTS 32 °C)
verdünnt 20 min bei 21 °C gehalten und schließlich mit BTS (21 °C) hypotherm ausverdünnt.
Die ausverdünnten Proben (in 100 ml Ebersamenflaschen) wurden bei 32 °C bzw. 21 °C
gehalten (für 30 min, 60 min, 3 h und 6 h), und danach für weitere 90 min bei 21 °C gehalten
und anschließend bei 17 °C gelagert. ...................................................................................85
Abb. 29: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der
Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Dargestellt
sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote Säulen), und
Proben mit Haltetemperatur 21 °C (rote Säulen). Farbabstufungen: unterschiedlichen
Haltezeiten (30 min – 6 h); n=6 Ejakulate. ............................................................................86
Abb. 30: Prozentualer Anteil membranintakter (PI und FITC-PNA negativer) Spermien (MW +
SD) in Abhängigkeit von der der Haltetemperatur, Haltezeit und der Lagerungsdauer des
vorverdünnten Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit
Haltetemperatur 32 °C (rote Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen). 87
Abb. 31: Temperaturverlauf der bei Experiment 4b einstufig isotherm verdünnten und
anschließend bei 21 °C gehaltenen Proben. Mittelwerte von je 3 Samenproben. Diese
wurden einstufig isotherm mit Beltsville Thawing Solution (BTS, 21 °C) ausverdünnt und in
100 ml Ebersamenflaschen abgefüllt. Die ausverdünnten Proben wurden bei 32 °C bzw. 21
°C gehalten (für 30 min, 60 min, 3 h und 6 h), und danach für weitere 90 min bei 21 °C
gehalten und anschließend bei 17 °C gelagert. . ..................................................................89
Abb. 32: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der
Haltetemperatur, Haltezeit und der Lagerungsdauer des vorverdünnten Samens. Dargestellt
sind die Kontrolle* (gestreifte Säulen), Proben mit Haltetemperatur 32 °C (rote Säulen), und
Proben mit Haltetemperatur 21 °C (rote Säulen). Farbabstufungen: unterschiedlichen
Haltezeiten (30 min – 6 h); n=6 Ejakulate. ............................................................................90
Abb. 33: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW
* SD) in Abhängigkeit von der der Haltetemperatur, Haltezeit und der Lagerungsdauer des
vorverdünnten Samens. Dargestellt sind die Kontrolle* (gestreifte Säulen), Proben mit
Haltetemperatur 32 °C (rote Säulen), und Proben mit Haltetemperatur 21 °C (rote Säulen).
Farbabstufungen: unterschiedlichen Haltezeiten (30 min – 6 h); n=6 Ejakulate. ...................91

Abbildungsverzeichnis
157
Abb. 34: Temperaturverlauf der bei Experiment 5 durchgeführten beschleunigten Abkühlung
(32 °C – 17 °C) und der Abkühlung gemäß ZDS-Empfehlungen (32 °C – 21 °C – 17°C) der
verdünnten Samenproben in 100 ml Ebersamenflaschen. Mittelwerte von je 3 Samenproben.
Die Samenproben einstufig isotherm mit 32 °C warmer Beltsville Thawing Solution (BTS) auf
die Endkonzentration 20 x 106 verdünnt. Anschließend kühlte ein Teil der Proben nach ZDS-
Empfehlungen 90 min bei 21 °C ab (blaue Kurve) und wurden bei 17 °C gelagert. Der andere
Teil kühlte direkt nach der Verdünnung bei 17 °C ab (rote Kurve). .......................................93
Abb. 35: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit der Abkühlraten
und Lagerungsdauer; n=6 Ejakulate. Blaue Säulen: Abkühlung laut ZDS Empfehlungen;
Nach Verdünnung mit 32 °C 90 min Abkühlung bei 21 °C, anschließend weitere Abkühlung
und Lagerung bei 17 °C. .......................................................................................................94
Abb. 36: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW
+ SD) in Abhängigkeit der Abkühlrate und Lagerungsdauer. Blaue Säulen: Abkühlung laut
ZDS Empfehlungen; Nach Verdünnung mit 32 °C 90 min Abkühlung bei 21 °C, anschließend
Lagerung bei 17 °C. .............................................................................................................95
Abb. 37: Temperaturverlauf während der einstufig isothermen (Standard TiHo, schwarze
Linie) und zweistufig hypothermen (Station, rote und blaue Linie) Samenverarbeitung bei
Experiment 6. Beim Standard wurden die Proben direkt isotherm ausverdünnt, 90 min bei 21
°C einzeln gehalten und anschließend bei 17 °C gelagert. Bei der simulierten Verdünnung
der Besamungsstation fand eine isotherme Vorverdünnung (1:1) statt, anschließend eine
Haltezeit von 30 min bei 21 °C und mit BTS (26 °C) ausverdünnt. Nach der Abfüllung wurden
40 Tuben zusammen für 2,5 h in einer Plastikbox bei 21 °C gehalten und danach bei 17 °C
gelagert. Die rote Linie zeigt den Temperaturverlauf mittig liegender Tuben, die blaue Line
die Temperatur außen liegender Tuben; n=4 Ejakulate. .......................................................97
Abb. 38: Prozentualer Anteil motiler Spermien (MW + SD) in Abhängigkeit von der
Verarbeitungsart und Lagerungsdauer; n=6 Ejakulate. .........................................................98
Abb. 39: Prozentualer Anteil membranintakter (PI- und FITC-PNA negativer) Spermien (MW
+ SD) in Abhängigkeit von der Verarbeitungsart und Lagerungsdauer; n=6 Ejakulate. .........99
Abb. 40: Spezifische Reaktivität auf Bicarbonat berechnet anhand der Propidiumiodid-
negativen (PI-neg.) und Fluo-3-negativen (Fluo-3-neg) Spermienpopulation nach 60 min
Inkubationsdauer; n=6 Ejakulate. Darstellung der Werte (MW + SD) in Abhängigkeit von der
Verarbeitungsart und Lagerungsdauer. .............................................................................. 100
Abb. 41: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VAP (velocity
average path; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der
Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der
für 6h bei 32 °C gehaltenen Probe. .................................................................................... 139
Abb. 42: Prozentualer Anteil der Spermien pro Geschwindigkeitsklasse der VSL (velocity
straight line; in µm/ s) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der

Abbildungsverzeichnis
158
Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der
für 6h bei 32 °C gehaltenen Probe. .................................................................................... 140
Abb. 43: Prozentualer Anteil der Spermien pro Bewegungsklasse der STR (straightness;
VSL/VAP) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die
grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C
gehaltenen Probe. .............................................................................................................. 141
Abb. 44: Prozentualer Anteil der Spermien pro Bewegungsklasse der LIN (linearity;
VSL/VCL) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die
grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C
gehaltenen Probe. .............................................................................................................. 142
Abb. 45: Prozentualer Anteil der Spermien pro Bewegungsklasse der WOB (wobble;
VAP/VCL) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die
grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C
gehaltenen Probe. .............................................................................................................. 143
Abb. 46: Prozentualer Anteil der Spermien pro Bewegungsklasse der ALH (amplitude of
lateral head displacement; µm) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der
Kontrollprobe*, die grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der
für 6h bei 32 °C gehaltenen Probe. .................................................................................... 144
Abb. 47: Prozentualer Anteil der Spermien pro Bewegungsklasse der BCF (beat cross
frequency; Hz) an Tag 0 (A) und Tag 1 (B). Die blaue Linie entspricht der Kontrollprobe*, die
grüne Linie der für 6 h bei 21 °C gehaltenen Probe und die rote Linie der für 6h bei 32 °C
gehaltenen Probe. .............................................................................................................. 145

Tabellenverzeichnis
159
11 Tabellenverzeichnis
Tabelle 1: CASA Motilitätsparameter (MW ± SD) von Experiment 1 in Abhängigkeit der
Verdünnungsart, des Verdünnungsgrades und der Lagerungsdauer (n = 6 Eber) .............. 122
Tabelle 2: Membranintakte Spermien in % (MW ± SD) von Experiment 1 in Abhängigkeit
der Verdünnungsart, des Verdünnungsgrades und der Lagerungsdauer (n = 6 Eber) ........ 123
Tabelle 3: CASA Motilitätsparameter (MW ± SD) von Experiment 2 in Abhängigkeit der
Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber) ............................... 124
Tabelle 4: Ergebnisse der durchflusszytometrischen Messungen in % (MW ± SD) von
Experiment 2 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n
= 12 Eber) .......................................................................................................................... 126
Tabelle 5: Ergebnisse der Analyse des Energiestoffwechsels (MW + SD) von Experiment 2 in
Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 12 Eber) ... 127
Tabelle 6: CASA Motilitätsparameter (MW ± SD) von Experiment 3 in Abhängigkeit der
Haltezeit, der Verdünnertemperatur und der Lagerungsdauer (n = 6 Eber) ........................ 128
Tabelle 7: Membranintakte Spermien in % (MW ± SD) von Experiment 3 in Abhängigkeit der
Haltezeit, der Verdünnertemperatur und der Lagerungsdauer (n = 6 Eber) ........................ 130
Tabelle 8: CASA Motilitätsparameter (MW ± SD) von Experiment 4 a in Abhängigkeit der
Haltetemperatur, Haltezeit und der Lagerungsdauer (n = 6 Eber) ....................................... 131
Tabelle 9: Membranintakte Spermien in % (MW ± SD) von Experiment 4 a in Abhängigkeit
der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber) .......................... 132
Tabelle 10: Membranintakte Spermien in % (MW ± SD) von Experiment 4 a in Abhängigkeit
der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber) .......................... 133
Tabelle 11: CASA Motilitätsparameter (MW ± SD) von Experiment 4 b in Abhängigkeit der
Haltetemperatur, Haltezeit und der Lagerungsdauer (n = 6 Eber) ....................................... 134
Tabelle 12: Membranintakte Spermien in % (MW ± SD) von Experiment 4 b in Abhängigkeit
der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber) .......................... 136
Tabelle 13: CASA Motilitätsparameter (MW ± SD) von Experiment 5 in Abhängigkeit vom
Abkühlregime und der Lagerungsdauer (n = 6 Eber) .......................................................... 137
Tabelle 14: Membranintakte Spermien in % (MW ± SD) von Experiment 5 in Abhängigkeit
der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n = 6 Eber) .......................... 137
Tabelle 15: CASA Motilitätsparameter (MW ± SD) von Experiment 6 in Abhängigkeit von der
Art der Verarbeitung und der Lagerungsdauer (n = 6 Eber) ................................................ 138

Tabellenverzeichnis
160
Tabelle 16: Ergebnisse der durchflusszytometrischen Messungen in % (MW ± SD) von
Experiment 6 in Abhängigkeit der Haltezeit, der Haltetemperatur und der Lagerungsdauer (n
= 12 Eber) .......................................................................................................................... 138

Danksagung
161
Danksagung
Zuerst möchte ich mich herzlich bei Frau Prof. Dagmar Waberski für die Überlassung
des interessanten Themas und die stets gewährte Unterstützung und Motivation
bedanken, sowie für die Möglichkeit des eigenständigen wissenschaftlichen
Arbeitens.
Der Firma Minitüb und dem Förderverein Biotechnologieforschung (FBF) danke ich
für die finanzielle Unterstützung der Arbeit.
Ein besonderer Dank geht an Herrn Dr. Heiko Henning für die Begleitung der
Dissertation vor Ort und aus der Ferne mit vielen Hilfestellungen sowie fachlichen
Anregungen.
Ich möchte auch den Mitarbeitern der Reproduktionsmedizinischen Einheit, vor allem
Anja, Xuyên und Quynh, allen Tierpflegern und meinen Mitdoktoranden, allen voran
Sabine und Annett, für ihre Hilfe danken.
Ein weiterer Dank geht an Frau Dr. Marion Piechotta aus der Klinik für Rinder für die
Bereitstellung der Räumlichkeiten und Geräte für die Nukleotid-Analyse.
Außerdem danke ich Frau Dr. Anja Riesenbeck und ihrem Team für die
Unterstützung auf der Besamungsstation.
Speziell möchte ich Thomas Willer danken. Er hat mich mit viel Geduld unterstützt,
mich mit seiner ruhigen Art immer wieder auf den Boden der Tatsaschen zurück
gebracht, und ist immer da, wenn ich ihn brauche. Außerdem haben unsere Ausflüge
für unvergessliche Erlebnisse und gelungene Abwechslung gesorgt.
Ganz herzlich bedanke ich mich bei meiner Familie, meiner Schwester Eva mit ihrer
Familie und ganz besonders bei meinen Eltern für ihre Geduld und Unterstützung in
jeglicher Hinsicht während des Studiums, der Promotion und darüber hinaus. Danke!







