
Recombinant Protein Synthesis in Chinese HamsterOvary Cells Using a Vaccinia Virus/Bacteriophage T7Hybrid Expression System*
(Received for publication, February 13, 1996, and in revised form, April 24, 1996)
Anna Ramsey-Ewing and Bernard Moss‡
From the Laboratory of Viral Diseases, NIAID, National Institutes of Health, Bethesda, Maryland 20892
The vaccinia virus/bacteriophage T7 expression sys-tem was adapted to Chinese hamster ovary (CHO) cells.Vaccinia virus undergoes abortive infection in CHOcells, which is characterized by a sharp reduction inprotein synthesis at the stage of viral intermediate geneexpression. We determined that expression of a T7 pro-moter-regulated chloramphenicol acetyltransferasegene was at least 20 times more efficient in permissiveBS-C-1 than in CHO cells. The encephalomyocarditis vi-rus 5*-untranslated region, which confers cap-indepen-dent translatability to mRNA, stimulated recombinantprotein synthesis by 10-fold in both cell lines, maintain-ing the advantage of the BS-C-1 cells over CHO cells.Since the cowpox virus hr gene overcomes vaccinia vi-rus host range restriction in CHO cells, we constructeda recombinant virus that carries an intact hr gene inaddition to the T7 RNA polymerase gene. With this virus,synthesis of T7 RNA polymerase was enhanced and pro-duction of a recombinant protein occurred in CHO cellsat the level observed in permissive cell lines. Extensionof the vaccinia virus/bacteriophage T7 expression sys-tem to CHO cells should be of wide interest, as thesecells have advantages for preparation of recombinantproteins in research and biotechnology.
Recombinant expression systems may use regulatory ele-ments derived from phylogenetically distinct lineages. A suc-cessful mammalian expression system that exploits the bacte-riophage T7 RNA polymerase to transcribe T7 promoter-regulated target genes was described using vaccinia virus(VV)1 as the vector (1). In this system, recombinant VV carriesan integrated T7 polymerase gene regulated by a viral pro-moter. Originally, the T7 promoter-regulated template wascarried on either a plasmid that was transfected into infectedcells or on a second recombinant virus that was coinfected withthe T7 polymerase-expressing virus (2, 3). The low translat-ability of the largely uncapped mRNAs was overcome by initi-ating T7 transcripts with the leader sequence of encephalomyo-carditis virus (EMCV) (4). More recently, the VV/T7 hybridsystem was rendered inducible by inclusion of the Escherichiacoli lac operator/repressor system (5, 6), thereby allowing the
construction of recombinant viruses containing both the T7RNA polymerase and the T7 promoter-regulated gene. Becausevaccinia virus infects a broad range of cells in tissue culture,this system has been used to express recombinant proteins in avariety of cells.One notable exception to the usable cell lines is the Chinese
hamster ovary (CHO) cell line. Despite the fact that CHO cellsare one of a few cell lines approved for production of recombi-nant proteins for use in clinical trials, the VV/T7 technologyhas not yet been adapted to these cells. One major obstacle hasbeen the restriction in growth and gene expression of VV inCHO cells (7). Insertion of the CHO hr gene of cowpox virus(CPV) into the genome of VV, however, enables VV to produc-tively infect CHO cells (8). Replication of VV on CHO cells isblocked at the stage of viral intermediate protein synthesis (9).This defect is overcome by the CHO hr gene (9), which alsodelays the onset of apoptosis in VV-infected CHO cells (10). Weconstructed a new recombinant VV containing both the CHO hrand the bacteriophage T7 RNA polymerase genes, which allowscomparable levels of expression in CHO and fully permissivecell lines. Expression was further enhanced by incorporatingthe EMCV untranslated leader sequence.
MATERIALS AND METHODS
Cells and Viruses—BS-C-1 (kidney, African green monkey) and CV-1(kidney, African green monkey) cells were grown in minimum essentialmedium supplemented with 5% fetal calf serum. HeLa S3 monolayercultures were maintained in Dulbecco’s modified Eagle’s medium sup-plemented with 10% fetal calf serum. CHO cells were grown in mono-layer culture in Ham’s F-12 medium supplemented with 5% fetal calfserum. In suspension culture, CHO cells were maintained in an equalmixture of Ham’s F-12 and CHO-S-SFM II (Life Technologies, Inc.) orUltraCHO (BioWhittaker) media. VV strain WR and recombinantsthereof were propagated as described (11).Construction of Recombinant Viruses—Recombinant viruses vTF7–3,
vT7EMCAT, and vT7CAT have been described elsewhere (2, 3) and aredepicted in Fig. 1. To construct the vT7CP recombinant virus, a 2.3-kilobase pair EcoRI/PstI fragment containing the entire open readingframe of the CPV hr gene CP77 was excised from pEA36 (8) and ligatedto EcoRI/PstI cleaved pUC19. The resultant plasmid, pRECP77 (9), wasused as a donor in homologous recombination with vTF7–3 to createvT7CP. In addition to the T7 polymerase gene regulated by a viralearly/late promoter (P7.5) at the thymidine kinase locus, recombinantvirus vT7CP has an intact copy of the CP77 open reading frame, insteadof the disrupted WR copy (12), at the corresponding region ofHindIII C.Analysis of Viral Protein Synthesis—To examine viral protein syn-
thesis, we seeded 5 3 105 permissive or nonpermissive cells in mini-mum essential medium with 5% fetal calf serum. After 16–24 h, thecells were coinfected with recombinant viruses at a total multiplicity of30 plaque-forming units (pfu)/cell, 15 pfu/cell for each virus. 10–20 minbefore each labeling period, cells were washed twice and incubated inminimum essential medium with 5% fetal calf serum without methio-nine. The cells were then incubated in the presence of 75 mCi of [35S]me-thionine in 250 ml of methionine-free minimum essential medium with5% fetal calf serum for 30 min. The labeling medium was removed andthe cells were washed twice with ice-cold phosphate-buffered saline(PBS) and then incubated at 37 °C for 3–5 min with hypotonic lysis
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.‡ To whom correspondence should be addressed: Bldg. 4, Rm. 229,
NIAID, National Institutes of Health, Bethesda, MD 20892. Tel: 301-496-9869; Fax: 301-480-1147; E-mail: [email protected].
1 The abbreviations used are: VV, vaccinia virus; CPV, cowpox virus,EMCV, encephalomyocarditis virus; CHO, Chinese hamster ovary; hr,host range; hpi, h postinfection; PBS, phosphate-buffered saline; SDS,sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; pfu,plaque-forming unit; CAT, chloramphenicol acetyltransferase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 271, No. 28, Issue of July 12, pp. 16962–16966, 1996Printed in U.S.A.
16962
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from
buffer (20 mM Tris-HCl (pH 8.0), 10 mM NaCl, and 0.5% Nonidet P-40).The lysates were collected and centrifuged for 2 min at 12,000 3 g tosediment nuclei. The supernatants containing 35S-labeled polypeptideswere stored at 220 °C. A portion of each sample was mixed with anappropriate volume of 3 3 or 5 3 sodium dodecyl sulfate/b-mercapto-ethanol (SDS/2-mercaptoethanol, 59 to 39, Inc.) sample buffer and boiledfor 5 min. The proteins were resolved by polyacrylamide gel electro-phoresis (PAGE) in 10, 12, or 15% gels.Immunoprecipitation—Cells were infected with 15 pfu/cell of each
recombinant virus, labeled with [35S]methionine as described above,and harvested at various times. Lysates were prepared in isotonic lysisbuffer (50 mM Tris-HCl (pH 8.0), 150 mM NaCl, 0.02% sodium azide,0.1% SDS, 100 mg/ml phenylmethylsulfonyl fluoride, and 1% NonidetP-40), centrifuged to remove nuclei, and then incubated with polyclonalantiserum to CAT at a 1:500 dilution at 4 °C overnight. An equalvolume of 20% (w/v) protein A-Sepharose beads in PBS was added andincubation continued at room temperature for an additional 2–3 h or at4 °C overnight. Immune complexes were washed twice in Triton buffer(300 mM NaCl, 50 mM Tris-HCl (pH 7.4), and 0.1% Triton X-100),denatured and solubilized in Laemmli buffer, and then resolved bySDS-PAGE in 15% gels.Western Blot Analysis—For analysis of steady-state levels of pro-
teins, Western blot analysis was performed. Infected cells (1 3 106)were incubated with 0.5 ml of lysis buffer (20 mM Tris-HCl (pH 7.0),0.5% Triton X-100 in PBS) for 5 min at 37 °C. Lysates were centrifugedat 14,000 3 g for 5 min and the supernatants stored at 220 °C. Aportion was mixed with 5 3 SDS/2-mercaptoethanol sample buffer andboiled for 5 min, and proteins were resolved by SDS-PAGE in 10% gels.Proteins were electrophoretically transferred to nitrocellulose mem-branes and incubated with antisera (anti-CAT 1:500 and anti-T7 po-lymerase 1:1000) overnight at 4 °C and then with 125I-protein A over-night at 4 °C.Preparation and Analysis of Infected Cell RNA—RNA was isolated
from cells that had been infected with two recombinant viruses at amultiplicity of 15 pfu/cell for each. At various times after infection, thecells were washed twice in ice-cold PBS, and then they were lysed andthe RNA was extracted by the guanidinium thiocyanate/phenol/chloro-form extraction method (13), using RNAzol B, as described by themanufacturer (Tel-Test).To quantitate the steady-state levels of RNAs, samples of infected
cell RNA were applied to a nylon membrane (Schleicher & Schullnytran) using a Hoefer 24-well slot blot apparatus. Membranes werehybridized with a 32P-labeled DNA probe complementary to CATmRNA.Analysis of Reporter Gene Expression—Cells were infected with re-
combinant viruses at a multiplicity of 30 pfu/cell. At various times afterinfection, the cells were washed twice with PBS and incubated for 15min at 37 °C in 1 3 reporter lysis buffer (Promega). The lysate wascentrifuged and the supernatant was retained for further analysis.Protein content of each lysate was determined colorimetrically usingthe Pierce Coomassie Blue protein reagent. Either equal volumes of
lysates or equal amounts of protein were used in CAT assays as de-scribed by the manufacturer (Promega). Standard curves of both pro-tein content and enzyme activity were prepared for quantitative anal-ysis of assay results.
RESULTS
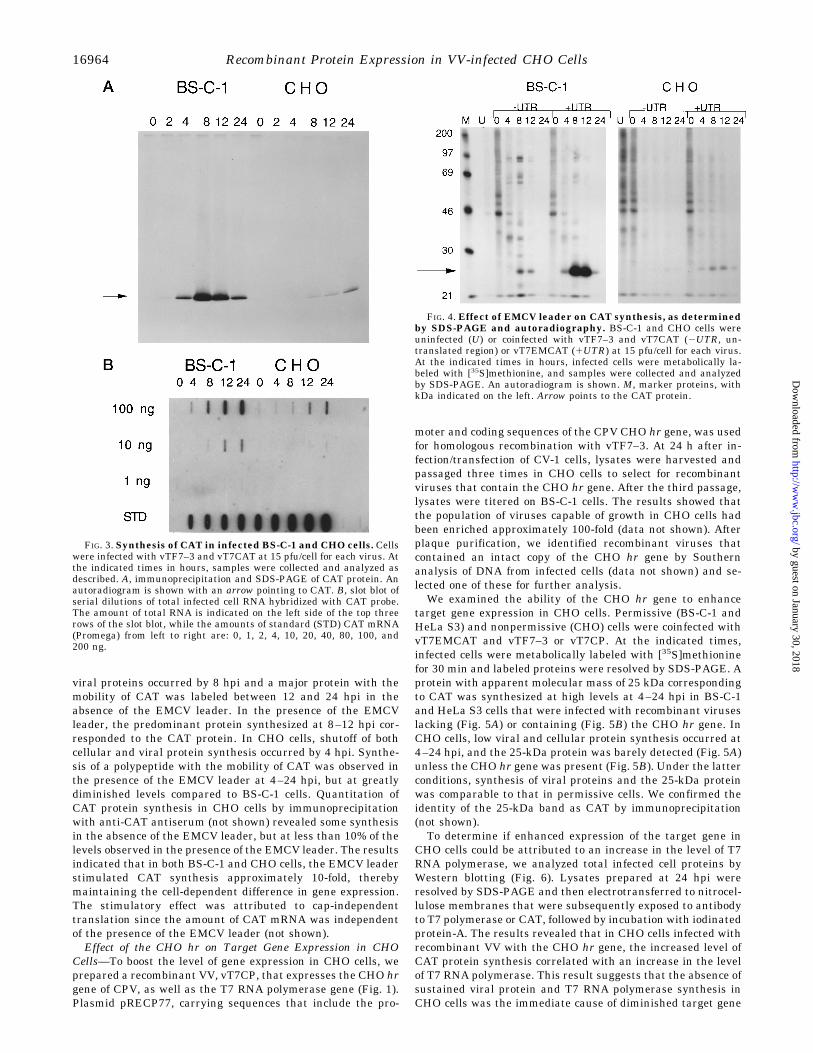
Recombinant Protein Synthesis in CHO Cells—We measuredthe impact of host range restriction of VV in CHO cells onexpression of a target gene using the VV/T7 system. Permissive(BS-C-1) or nonpermissive (CHO) cells were coinfected with 15pfu/cell of vTF7–3 (3), which encodes the T7 RNA polymerasegene regulated by the vaccinia virus early/late promoter (P7.5)and with vT7CAT, which has a T7 promoter-regulated CATgene. In BS-C-1 cells, enzymatically active CAT accumulatedrapidly between 4 and 24 h postinfection (hpi). In contrast, lowamounts of CAT accumulated between 12 and 24 hpi in CHOcells (Fig. 2). To monitor CAT protein synthesis, we pulselabeled infected cells with [35S]methionine and incubated ali-quots of the lysates with polyclonal antiserum directed againstthe CAT protein. Immune complexes were dissociated withSDS and electrophoretically resolved (Fig. 3A). The resultsconfirmed both the delayed and the low rate of CAT proteinsynthesis in CHO cells. Thus, there was at least 10-fold moreCAT protein synthesis in BS-C-1 than CHO cells.To determine if the differences in CAT expression reflected
the steady-state levels of CAT mRNA, we performed slot blotanalysis on total RNA from infected BS-C-1 and CHO cells witha CAT-specific DNA probe (Fig. 3B). Densitometric analysis ofthe autoradiogram revealed a 4–7-fold difference in the steady-state levels of CATmRNA in BS-C-1 and CHO cells. This resultsuggests that the differences in the levels of CAT protein syn-thesis and enzymatic activity in the two cell lines could be dueto the difference in the levels of CAT mRNA as well as in theirtranslation.Effect of Cap-independent Translation on Target Gene Ex-
pression—Previous studies (14) had shown that the mRNAssynthesized by T7 RNA polymerase are largely uncapped. Thetranslation of the uncapped transcripts was enhanced by incor-porating the EMCV leader at the 59-end of the mRNA (4).Therefore, we examined the ability of the EMCV leader tostimulate target gene expression in CHO cells. Total proteinsynthesis (Fig. 4) was analyzed in cells coinfected with vTF7–3and vT7CAT or vTF7–3 and vT7EMCAT, which contains acDNA copy of the EMCV leader preceding the CAT open read-ing frame. In BS-C-1 cells, decreased cellular protein synthesiswas observed at 4 h and had virtually ceased by 12 hpi, asindicated by labeling of the 43-kDa actin protein. Synthesis of
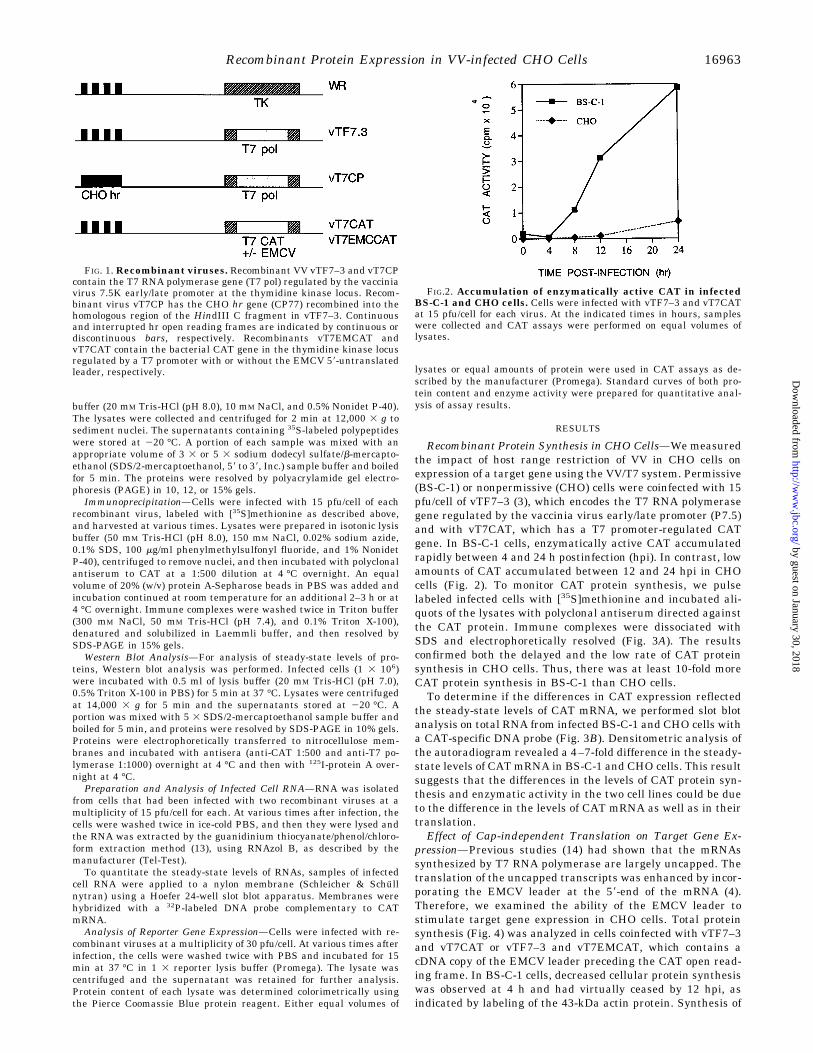
FIG. 1. Recombinant viruses. Recombinant VV vTF7–3 and vT7CPcontain the T7 RNA polymerase gene (T7 pol) regulated by the vacciniavirus 7.5K early/late promoter at the thymidine kinase locus. Recom-binant virus vT7CP has the CHO hr gene (CP77) recombined into thehomologous region of the HindIII C fragment in vTF7–3. Continuousand interrupted hr open reading frames are indicated by continuous ordiscontinuous bars, respectively. Recombinants vT7EMCAT andvT7CAT contain the bacterial CAT gene in the thymidine kinase locusregulated by a T7 promoter with or without the EMCV 59-untranslatedleader, respectively.
FIG.2. Accumulation of enzymatically active CAT in infectedBS-C-1 and CHO cells. Cells were infected with vTF7–3 and vT7CATat 15 pfu/cell for each virus. At the indicated times in hours, sampleswere collected and CAT assays were performed on equal volumes oflysates.
Recombinant Protein Expression in VV-infected CHO Cells 16963
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from
viral proteins occurred by 8 hpi and a major protein with themobility of CAT was labeled between 12 and 24 hpi in theabsence of the EMCV leader. In the presence of the EMCVleader, the predominant protein synthesized at 8–12 hpi cor-responded to the CAT protein. In CHO cells, shutoff of bothcellular and viral protein synthesis occurred by 4 hpi. Synthe-sis of a polypeptide with the mobility of CAT was observed inthe presence of the EMCV leader at 4–24 hpi, but at greatlydiminished levels compared to BS-C-1 cells. Quantitation ofCAT protein synthesis in CHO cells by immunoprecipitationwith anti-CAT antiserum (not shown) revealed some synthesisin the absence of the EMCV leader, but at less than 10% of thelevels observed in the presence of the EMCV leader. The resultsindicated that in both BS-C-1 and CHO cells, the EMCV leaderstimulated CAT synthesis approximately 10-fold, therebymaintaining the cell-dependent difference in gene expression.The stimulatory effect was attributed to cap-independenttranslation since the amount of CAT mRNA was independentof the presence of the EMCV leader (not shown).Effect of the CHO hr on Target Gene Expression in CHO
Cells—To boost the level of gene expression in CHO cells, weprepared a recombinant VV, vT7CP, that expresses the CHO hrgene of CPV, as well as the T7 RNA polymerase gene (Fig. 1).Plasmid pRECP77, carrying sequences that include the pro-
moter and coding sequences of the CPV CHO hr gene, was usedfor homologous recombination with vTF7–3. At 24 h after in-fection/transfection of CV-1 cells, lysates were harvested andpassaged three times in CHO cells to select for recombinantviruses that contain the CHO hr gene. After the third passage,lysates were titered on BS-C-1 cells. The results showed thatthe population of viruses capable of growth in CHO cells hadbeen enriched approximately 100-fold (data not shown). Afterplaque purification, we identified recombinant viruses thatcontained an intact copy of the CHO hr gene by Southernanalysis of DNA from infected cells (data not shown) and se-lected one of these for further analysis.We examined the ability of the CHO hr gene to enhance
target gene expression in CHO cells. Permissive (BS-C-1 andHeLa S3) and nonpermissive (CHO) cells were coinfected withvT7EMCAT and vTF7–3 or vT7CP. At the indicated times,infected cells were metabolically labeled with [35S]methioninefor 30 min and labeled proteins were resolved by SDS-PAGE. Aprotein with apparent molecular mass of 25 kDa correspondingto CAT was synthesized at high levels at 4–24 hpi in BS-C-1and HeLa S3 cells that were infected with recombinant viruseslacking (Fig. 5A) or containing (Fig. 5B) the CHO hr gene. InCHO cells, low viral and cellular protein synthesis occurred at4–24 hpi, and the 25-kDa protein was barely detected (Fig. 5A)unless the CHO hr gene was present (Fig. 5B). Under the latterconditions, synthesis of viral proteins and the 25-kDa proteinwas comparable to that in permissive cells. We confirmed theidentity of the 25-kDa band as CAT by immunoprecipitation(not shown).To determine if enhanced expression of the target gene in
CHO cells could be attributed to an increase in the level of T7RNA polymerase, we analyzed total infected cell proteins byWestern blotting (Fig. 6). Lysates prepared at 24 hpi wereresolved by SDS-PAGE and then electrotransferred to nitrocel-lulose membranes that were subsequently exposed to antibodyto T7 polymerase or CAT, followed by incubation with iodinatedprotein-A. The results revealed that in CHO cells infected withrecombinant VV with the CHO hr gene, the increased level ofCAT protein synthesis correlated with an increase in the levelof T7 RNA polymerase. This result suggests that the absence ofsustained viral protein and T7 RNA polymerase synthesis inCHO cells was the immediate cause of diminished target gene
FIG. 3. Synthesis of CAT in infected BS-C-1 and CHO cells. Cellswere infected with vTF7–3 and vT7CAT at 15 pfu/cell for each virus. Atthe indicated times in hours, samples were collected and analyzed asdescribed. A, immunoprecipitation and SDS-PAGE of CAT protein. Anautoradiogram is shown with an arrow pointing to CAT. B, slot blot ofserial dilutions of total infected cell RNA hybridized with CAT probe.The amount of total RNA is indicated on the left side of the top threerows of the slot blot, while the amounts of standard (STD) CAT mRNA(Promega) from left to right are: 0, 1, 2, 4, 10, 20, 40, 80, 100, and200 ng.
FIG. 4. Effect of EMCV leader on CAT synthesis, as determinedby SDS-PAGE and autoradiography. BS-C-1 and CHO cells wereuninfected (U) or coinfected with vTF7–3 and vT7CAT (2UTR, un-translated region) or vT7EMCAT (1UTR) at 15 pfu/cell for each virus.At the indicated times in hours, infected cells were metabolically la-beled with [35S]methionine, and samples were collected and analyzedby SDS-PAGE. An autoradiogram is shown. M, marker proteins, withkDa indicated on the left. Arrow points to the CAT protein.
Recombinant Protein Expression in VV-infected CHO Cells16964
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from

expression.Quantitation of CAT Protein Synthesized in CHO Cells Using
the Vaccinia Virus/T7 System—To quantitate the amount ofCAT protein synthesized in the vaccinia virus/T7 systemadapted to CHO cells and compare it with that obtained inother cell types, we performed CAT assays on samples fromthree cell types grown in monolayer culture (Fig. 7) and quan-titative Western analysis on samples from CHO cells grown insuspension culture (not shown). When the CHO hr gene waspresent, the amounts of CAT activity recovered per cell at 24hpi were comparable in BS-C-1, HeLa S3, and CHO cells. Weadapted CHO cells to growth in suspension culture in either oftwo media and coinfected with vT7CP and vT7EMCAT. Quan-titation of the results revealed that by 24 hpi, 2–5 mg of CATprotein were produced per 108 suspension CHO cells.
DISCUSSION
The present studies were initiated to learn more about themechanism of vaccinia virus host restriction in CHO cells andto adapt the widely used vaccinia virus/T7 hybrid expressionsystem to these cells. Vaccinia virus gene expression is regu-
lated by a cascade mechanism: early, intermediate, and latemRNAs are synthesized in succession (15). In CHO cells, how-ever, only early and intermediate mRNAs are synthesized (9).Translation of intermediate mRNAs is severely inhibited, pre-venting the synthesis of late stage transcription factors andsubsequent transcription of late genes. Since the translationaldefect pertains to mRNAs transcribed from both authenticintermediate stage genes and reporter genes regulated by in-
FIG. 5. Effect of CHO hr gene on CAT synthesis, as determinedby SDS-PAGE and autoradiography. BS-C-1, HeLa S3, and CHOcells were coinfected with vT7CAT and vTF7–3 (panel A) or vT7CP(panel B) at 15 pfu/cell for each virus. At the indicated times in hours,infected cells were metabolically labeled with [35S]methionine and totalprotein synthesis was determined by SDS-PAGE and autoradiography.Arrow points to CAT.
FIG. 6. Effect of CHO hr gene on T7 RNA polymerase and CATsynthesis, as determined by Western blot analysis. BS-C-1, HeLaS3, and CHO cells were coinfected with vT7CAT and vTF7–3 (-CP) orvT7CP (1CP) at 15 pfu/cell for each virus. Samples harvested at 24 hpiwere analyzed by Western blotting with antisera directed against T7RNA polymerase (T7 pol) or CAT. M, marker; B, BS-C-1; C, CHO; H,HeLa S3. Arrows point to T7 polymerase and CAT in the upper andlower panels, respectively.
FIG. 7. Quantitation of CAT gene expression. BS-C-1, HeLa S3,and CHO cells were coinfected with vT7CAT and vTF7–3 (2CP) orvT7CP (1CP) at 15 pfu/cell for each virus. At 24 hpi, lysates werecollected and CAT assays were performed. Values shown are normal-ized for the number of cells contributing to each lysate.
Recombinant Protein Expression in VV-infected CHO Cells 16965
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from

termediate promoters, sequence-specific features of intermedi-ate mRNAs are probably not involved. Instead, translation maybe generally blocked at the time of intermediate gene transcrip-tion. We were curious to determine whether we could overcomethe expression block by using T7 RNA polymerase to transcribereporter genes and by using the leader sequence of EMCV toachieve cap-independent translation. In recombinant VVvTF7–3, the T7 RNA polymerase gene is regulated by an early/late promoter; therefore, synthesis of some T7 RNA polymeraseshould occur in CHO cells, followed by transcription of the T7promoter-regulated reporter gene. We found that T7-regulatedreporter gene expression was delayed and severely reduced inCHO cells relative to that in permissive BS-C-1 cells. Moreover,the EMCV leader stimulated expression to a similar extent inCHO and BS-C-1 cells, suggesting that the translational blockwas not related to a specific cap-dependent step in translation.Further analysis indicated the presence of low amounts of T7transcripts in CHO cells relative to BS-C-1 cells, regardless ofwhether the EMCV leader was present.An alternative approach was taken to adapt the VV/T7 hy-
brid expression system to CHO cells. Spehner et al. (8) hadoriginally shown, and we confirmed (9), that the CP77 hr geneof CPV allows VV to replicate in CHO cells. Sequence studieshad shown that the open reading frame of the VV homolog ofCP77 was interrupted in the WR strain of VV (12). We there-fore used homologous recombination to repair the VV hr gene ofa recombinant VV containing the T7 RNA polymerase gene.The resulting recombinant vT7CP replicated in CHO cells andproduced larger amounts of viral proteins and T7 RNA polym-erase. Importantly, vT7CP could be used as an efficient expres-sion vector either with transfected plasmids containing a T7promoter-regulated gene (data not shown) or by co-infectingCHO cells with a second VV containing an integrated copy of aT7 promoter-regulated gene. As the CP77 gene is dominant,the second recombinant VV need not have an intact copy. Thatis a great convenience, since recombinant VV previously con-structed for use in permissive cells can be used with vT7CP inCHO cells. With this system, we estimated that approximately2–5 mg of CAT protein was produced in 108 CHO cells in 24 h.This value could probably be doubled by adapting the inducibleVV/T7 system (6) to CHO cells by inserting the CP77 gene intothe vector vT7lacOI.Extension of the VV/T7 expression system to CHO cells was
important for several reasons. First, CHO cells are frequentlyused for overexpression of proteins (16–20) and can be grown in
large scale suspension or hollow fiber reactor cultures in de-fined serum-free medium. CHO cells are among the very fewcontinuous cells lines approved for expression of recombinantproteins for use in humans. For example, recombinant DNase I,for treatment of cystic fibrosis, has been produced in CHO cells(21). Furthermore, because of the genetic stability of CHO cells,mutants lines deficient in metabolic steps such as glycosylation(22) have been made. In addition, both monochromosomal andpolychromosomal human/CHO somatic cell hybrids are avail-able (Coriell Institute for Medical Research). In our laboratory,we have used glycosylation-deficient CHO cells to study theprocessing of the HIV-1 envelope protein synthesized with thenewly modified VVCP/T7system.2
REFERENCES
1. Moss, B., Elroy-Stein, O., Mizukami, T., Alexander, W. A., and Fuerst, T. R.(1990) Nature 348, 91–92
2. Fuerst, T. R., Niles, E. G., Studier, F. W., and Moss, B. (1986) Proc. Natl. Acad.Sci. U. S. A. 83, 8122–8126
3. Fuerst, T. R., Earl, P. L., and Moss, B. (1987) Mol. Cell. Biol. 7, 2538–25444. Elroy-Stein, O., Fuerst, T., and Moss, B. (1989) Proc. Natl. Acad. Sci. U. S. A.
86, 6126–61305. Alexander, W. A., Moss, B., and Fuerst, T. R. (1992) J. Virol. 66, 2934–29426. Ward, G. A., Stover, C. K., Moss, B., and Fuerst, T. R. (1995) Proc. Natl. Acad.
Sci. U. S. A. 92, 6773–67777. Drillien, R., Spehner, D., and Kirn, A. (1978) J. Virol. 28, 843–8508. Spehner, D., Gillard, S., Drillien, R., and Kirn, A. (1988) J. Virol. 62,
1297–13049. Ramsey-Ewing, A. L., and Moss, B. (1995) Virology 206, 984–99310. Ink, B. S., Gilbert, C. S., and Evan, G. I. (1995) J. Virol. 69, 661–66811. Earl, P. L., and Moss, B. (1991) in Current Protocols in Molecular Biology
(Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G.,Smith, J. A., and Struhl, K., eds) pp. 16.17.1–16.17.16, Greene PublishingAssociates and Wiley Interscience, New York
12. Kotwal, G. J., and Moss, B. (1988) Virology 167, 524–53713. Chomczynski, P., and Sacchi, N. (1987) Anal. Biochem. 162, 156–15914. Fuerst, T. R., and Moss, B. (1989) J. Mol. Biol. 206, 333–34815. Moss, B. (1990) Annu. Rev. Biochem. 59, 661–68816. Collen, D., Stassen, J. M., Marafino, B. J., Jr., Builder, S., DeCock, F., Ogez, J.,
Tajiri, D., Pennica, D., Bennett, W. F., Salwa, J., and Hoyng, C. F. (1984) J.Pharmacol. Exp. Ther. 231, 146–152
17. Egrie, J. C., Brown, J., Lai, P., and Fu-Kuen, L. (1985) in ExperimentalApproaches for the Study of Hemoglobin Switching, pp. 339–350, Alan R.Liss, Inc., New York
18. Patzer, E. J., Nakamura, G. R., Hershberg, R. D., Gregory, T. J., Crowley, C.,Levinson, A. D., and Eichberg, J. W. (1986) Bio/Technology 4, 630–636
19. Scahill, S. J., Devos, R., Van der Heyden, J., and Fiers, W. (1983) Proc. Natl.Acad. Sci. U. S. A. 80, 4654–4658
20. Zettmeibl, G., Ragg, H., and Karges, H. E. (1987) BioTechniques 5, 720–72521. Hubbard, R. C., McElvaney, N. G., Birrer, P., Shak, S., Robinson, W. W.,
Jolley, C., Wu, M., Chernick, M. S., and Crystal, R. G. (1992) N. Engl.J. Med. 326, 812–815
22. Elbein, D. (1991) Trends Biotechnol. 9, 346–352
2 V. Karacostas, personal communication.
Recombinant Protein Expression in VV-infected CHO Cells16966
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from

Anna Ramsey-Ewing and Bernard MossVirus/Bacteriophage T7 Hybrid Expression System
Recombinant Protein Synthesis in Chinese Hamster Ovary Cells Using a Vaccinia
doi: 10.1074/jbc.271.28.169621996, 271:16962-16966.J. Biol. Chem.
http://www.jbc.org/content/271/28/16962Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/271/28/16962.full.html#ref-list-1
This article cites 20 references, 10 of which can be accessed free at
by guest on January 30, 2018http://w
ww
.jbc.org/D
ownloaded from







